
Capsular Transformation of a Multidrug‐Resistant Streptococcus pneumoniae In Vivo
Upload
independentCategory
view
2download
0
Im
MAGa
b
c
d
a
ARR2AA
KSPP
1
sgfhhpscptt
tpe
T9
0d
Vaccine 28 (2010) 3609–3616
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
mmunoprotective activities of a Streptococcus suis pilus subunit in murineodels of infection
anuela Garibaldia,1, Manuel J. Rodríguez-Ortegab,c,1, Francesca Mandanici a, Angela Cardacia,ngelina Midiri a, Salvatore Papasergia, Orazio Gambadorod, Vittorio Cavallarid,iuseppe Teti a, Concetta Beninati a,∗
The Elie Metchnikoff Department, University of Messina, Messina I-98125, ItalyDepartamento de Bioquímica y Biología Molecular, Universidad de Córdoba, Córdoba 14071 SpainInstituto Maimónides de Investigación Biomédica de Córdoba (IMIBIC), Córdoba 14071, SpainDipartimento di Patologia Umana, University of Messina, Messina I-98125, Italy
r t i c l e i n f o
rticle history:eceived 30 July 2009eceived in revised form
a b s t r a c t
Pili of gram-positive bacteria are key virulence factors and their subunits are considered excellent vaccinecandidates. Streptococcus suis is an emerging zoonotic agent that can cause epidemics of life-threateninginfections in humans, but the functional role or immunoprotective potential of its pilus components
3 November 2009ccepted 6 January 2010vailable online 14 January 2010
eywords:treptococcus suis
have not been studied yet. Using a selective proteomics approach, we have identified a surface proteinof serotype 2 S. suis showing features of an ancillary pilus subunit, as evidenced by bioinformatics anal-ysis, immunoblot and immunoelectron microscopy. Immunization with recombinant fragments of thisprotein, designated herein as PAPI-2b, markedly protected mice from systemic S. suis infection.
© 2010 Elsevier Ltd. All rights reserved.
iliroteomics
. Introduction
The gram-positive bacterium Streptococcus suis causes a range oferious swine diseases, including pneumonia, endocarditis, menin-itis and arthritis [1]. It was recently found that, after transmissionrom pigs, this pathogen can cause life-threatening disease inumans. For example in 1998 and 2005, two large outbreaks ofuman S. suis infections occurred in China, affecting hundreds ofeople and killing over fifty [2]. Therefore, this emerging zoono-is has raised public health concerns worldwide, especially inountries with intensive swine production. Based on the capsularolysaccharide, 35 serotypes have been described so far. Amonghese, serotype 2 is most commonly associated with invasive infec-ions in animals and humans worldwide [1–3].
Little is known, in comparison with other pathogenic strep-ococci, of the virulence factors of S. suis [3]. The capsularolysaccharide [4] and a serum opacity factor [5] are appar-ntly required for pathogenicity, since isogenic mutants devoid
∗ Corresponding author at: Concetta Beninati, The Elie Metchnikoff Department,orre Biologica IIp, Policlinico Universitario, Via Consolare Valeria SNC, Messina I-8125, Italy. Tel.: +39 090 221 3316; fax: +39 090 221 3312.
E-mail address: [email protected] (C. Beninati).1 These authors contributed equally.
264-410X/$ – see front matter © 2010 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2010.01.009
of either of these factors lost most of their virulence. Other pro-posed virulence factors of S. suis include suilysin (a thiol-dependenthemolysin): [6], muramidase released protein [7], extracellular fac-tor protein [8] and fibrinogen- and fibronectin-binding protein [9].In the last few years, interest in the development of protein-basedvaccines has been spurred by the increasing availability of genomicsequences from many pathogens, including S. suis [10–12]. A com-bination of putative divalent cation-binding lipoproteins [13] anda surface antigen of unknown function designated as Sao [14] wererecently reported to be immunoprotective.
In the last few years, pili were described in highly prevalentgram-positive human pathogens, including Streptococcus pyogenes[15], Streptococcus pneumoniae [16] and Streptococcus agalactiae[17]. These structures are considered as virulence factors playingkey roles in the attachment of bacteria to host surfaces, as well as inthe invasion of epithelial and endothelial barriers [18]. Moreover,pilus subunits are effective immunoprotective agents and excellentvaccine candidates for the prevention of infections by a numberof pathogens, including S. agalactiae and S. pneumoniae [16–18].Although the presence of pili was observed in S. suis as early as
1990 [19], their functional role or immunoprotective properties areunknown.Quite recently, at least 4 putative S. suis pilus islands (PIs) wereidentified in sequenced genomes and sequences obtained fromseveral clinical isolates [20–23]. However, whether these putative

3 ccine
Paccfa(2i
2
2
fcs
2
ctg(tgatwcc
2
agotppaAfufwm(ta
2
pdDUaS
2
b
antisera diluted 1:20 in blocking solution for 30 min at room tem-
610 M. Garibaldi et al. / Va
Is (two of which have a gene organization similar to those of S.galactiae PI-1 and PI-2b, respectively) actually encode for pilusomponents is presently unknown. We describe here the identifi-ation by proteomics of a surface antigen of serotype 2 S. suis witheatures of a pilus ancillary protein, as evidenced by immunoblotnd immunoelectron microscopy analysis. Moreover, this proteinreferred to herein as pilus ancillary protein of island 2b or PAPI-b) showed marked immunoprotective effects in murine S. suis
nfection models.
. Materials and methods
.1. Bacterial strains
S. suis strain 235/02, serotype 2, was originally isolated in 2002rom an infected pig in Córdoba, Spain. Species identification wasonfirmed by 16S rRNA gene sequencing [24]. Strain P1/7, also aerotype 2 S. suis, was purchased from ATCC.
.2. Proteomic analysis
We used a highly selective mass spectrometry approach, whichonsists of digesting under mild conditions the surface of live bac-eria, as previously described [25,26]. Briefly, strain 235/02 cells,rown in Todd–Hewitt broth (THB; Oxoid) to the mid-log phaseOD620 = 0.5), were incubated with trypsin or proteinase K, in ordero cleave the exposed domains of surface proteins. The peptidesenerated from “shaved” bacterial cells were recovered and thennalyzed by two-dimensional liquid chromatography coupled toandem mass spectrometry [25,26]. The mass spectra thus obtainedere used for database searching and peptide sequence identifi-
ation. Under the conditions used, no cytoplasmic contaminationould be detected.
.3. Expression of recombinant protein fragments
We expressed 3 polypeptide fragments, designated as FA1, FA3nd FA4 (Fig. 1), encoded by ORF ssu05 0473 in the sequencedenome of strain 05ZYH33 [10]. FA1 and FA3 contained eachne of the two peptides identified by tandem mass spectrome-ry (Fig. 1). Moreover, we expressed the previously described Saorotein [14] in order to use it as a positive control in immuno-rotection experiments. The corresponding DNA fragments weremplified by PCR from the genomic DNA of S. suis strain 235/02.ll recombinant protein fragments were produced as polypeptides
used to a biotinylable peptide tag (BIO) in Escherichia coli JM109,sing the PinPointTM Xa-1 Vector (Promega) according to manu-acturer’s instructions. After the induction of fusion proteins, theseere purified from the cytoplasm of bacterial cells by affinity chro-atography using the PinPointTM Xa protein purification system
Promega). Recombinant chloramphenicol acetyl transferase fusedo BIO (CAT-BIO), to be used as a negative control, was producednd purified using the same methods.
.4. Experimental animals
Four-week-old, specific pathogen-free female CD1 mice wereurchased from Charles River Italia. All of the animal experimentsescribed in the present study were conducted at the Metchnikoffepartment of the University of Messina according to the Europeannion guidelines for the handling of laboratory animals and werepproved by the relevant national authority (Istituto Superiore dianità of Italy).
.5. Production of mouse antisera
Mice were immunized by the intraperitoneal injection of recom-inant FA1-BIO, FA3-BIO or CAT-BIO (50 �g) in complete Freund’s
28 (2010) 3609–3616
adjuvant (Sigma) on day 0, and in incomplete Freund’s adjuvanton days 14 and 28. The use of complete Freund’s adjuvant in thefirst immunization was justified by our previous observations thathigh titered sera were more consistently obtained with this adju-vant, as compared to other less “inflammatory” adjuvants suchas alum. However, care was taken to minimize discomfort to theanimals by injecting a low volume of the emulsion (0.1 ml con-taining 0.05 mg of mycobacteria) and by using sterile solutionsand techniques to prepare it. Under these conditions no significantabdominal distension or complications at the injection site wereobserved throughout the experimental period. To obtain positivecontrol anti-S. suis sera, an additional group of mice was immunizedat days 0, 14 and 28 with killed bacteria (strain 235/02; 105 CFU)in incomplete Freund’s adjuvant. Bacteria were killed by treatmentwith gentamycin (400 �g/ml for 2 h at 37 ◦C), followed by extensivewashing.
2.6. Flow cytometry immunofluorescence analysis
S. suis cells, grown to the mid-log phase (OD620 = 0.5) in THB,were harvested by centrifugation, washed three times with phos-phate buffered saline (pH 7.2; PBS) and blocked for 20 min at 20 ◦Cwith PBS containing 2% fetal calf serum (PBS-FCS). Mouse antiserawere diluted 1:100 in PBS-FCS and incubated with bacterial cellsfor 40 min at 4 ◦C. Phycoerythrin-conjugated goat anti-mouse IgG(Jackson Immunoresearch), diluted 1:50 was then added to the cellsand incubated at 4 ◦C for 30 min. Bacteria were then washed andanalyzed with an LSR flow cytometer using the CellQuest software(both from BD Biosciences).
2.7. Western blot of cell-wall extracts
Bacterial cell-wall-enriched fractions were prepared asdescribed previously [15]. Briefly, bacteria grown in THB tothe mid-log phase (OD620 = 0.5) at 37 ◦C, were pelletted, washedonce in PBS, suspended in 1 ml ice-cold protoplasting buffer [40%sucrose; 0.1 M KPO4, pH 6.2; 10 mM MgCl2; EDTA-free proteaseinhibitors (Roche); 2 mg ml−1 lysozyme; 400 units of mutanolysin(Sigma)] and incubated at 37 ◦C for 3 h. After centrifuging at13,000 × g for 15 min, the supernatant (cell-wall fraction) wasseparated by 4–12% gradient gels (Bio-Rad) and transferred tonitrocellulose membranes (Bio-Rad). Immunoblots were obtainedby incubation with mouse polyclonal anti-FA3-BIO antisera(diluted 1:500 in PBS) followed by secondary antibody (alkalinephosphatase-conjugated goat anti-mouse IgG; Sigma) at a 1:5000dilution and developed with 5-bromo-4-chloro-3-indolyl phos-phate/nitro blue tetrazolium (Sigmafast BCIP/NBT tablets; Sigma)according to manufacturer’s instructions.
2.8. Immunoelectron microscopy
S. suis strain P1/7 was used in immunoelectron microscopystudies because it produced stronger signals, in preliminary flowcytometry immunofluorescence experiments, compared with the235/02 strain. Bacteria were grown on sheep blood agar plates(Oxoid), washed and resuspended in PBS to an OD620 = 0.5. Form-var carbon-coated nickel grids were floated on drops of bacterialsuspensions for 5 min, fixed in 2% paraformaldehyde for 5 min,and placed in blocking solution (PBS-FCS) for 30 min. The gridswere then floated on drops of mouse anti-FA3-BIO or anti-CAT-BIO
perature, washed, and floated for 30 min on secondary antibodyconjugated to 20 nm gold particles (diluted 1:10; BBInternational).The grids were washed with PBS, then with distilled water and air-dried. After staining with 1% uranyl acetate, grids were examinedusing a Philips EM 300 transmission electron microscope.

M. Garibaldi et al. / Vaccine 28 (2010) 3609–3616 3611
Fig. 1. (a) Deduced amino acid sequence of protein ssu05 0473 (PAPI-2b) and sequence coverage by the peptides identified by mass spectroscopy in S. suis strain 235/02,serotype 2. Covered sequences are enclosed in boxes. The arrow indicates the predicted signal peptidase cleavage site, according to SignalP v3.0 predictions. The LPXTG-cellwall anchoring motif near the C-term is underlined. The recombinant FA1, FA3 and FA4 fragments are indicated in bold. Fragments FA3 and FA4 overlap by two amino acids(highlighted in gray). (b) Structure of the putative S. suis pilus island, designated srtF cluster, in comparison with that of PI-2b from S. agalactiae. Both have the followinge subuna st.ncb4
2
m25w5dwpl1hfia
2c
bwim0
with the immunizing antigen (e.g. FA1- FA3- or FA4-BIO; 7 �g/ml).
lements: putative signal peptidase (light gray), ancillary protein (black), backbonend SAN 1575 deduced amino acid sequences, as determined by BLASTP (http://bla0–50 alignment scores.
.9. Lethal infection model
Mice were immunized as described under “production ofouse antisera” and challenged with the highly virulent strain
35/02 using a published S. suis infection model [27]. Briefly,× 108 CFUs of bacteria grown to the mid log phase (OD620 = 0.5)ere injected intraperitoneally (i.p.) in 1 ml of fresh THB usinganimals per group. This challenge was administered at 14
ays after the last immunization when the mice were 10-eek-old. Experiments were repeated twice, totaling 10 animalser group. Signs of disease (rough hair, decreased mobility,
ethargy, palsy or ataxia) and lethality were recorded daily for5 days and animals showing signs of irreversible disease wereumanely euthanized. Overwhelming S. suis infection was con-rmed as the cause of death by culturing the organs of moribundnimals.
.10. Measurement of bacterial burden after intravenoushallenge
To measure the ability of immunoprotective agents to prevent
acterial colonization of vital organs, we developed a model inhich bacterial burden is measured after the systemic admin-stration of a sublethal dose of S. suis. In initial experimentsice were injected i.v. with 2 × 107 CFUs of strain 235/02 in
.1 ml of PBS and the organs were collected at various times
it (striped pattern), class C sortase (dark grey). (c) Alignment scores for ssu05 0473i.nlm.nih.gov/Blast.cgi). Light and dark grey boxes indicate, respectively, 50–80 and
after challenge to measure CFUs using standard agar plating tech-niques. One kidney was fixed using 4% paraformaldehyde in PBS,embedded in paraffin, cut into 4 �m sections and stained withhematoxylin–eosin using standard techniques. For immunopro-tection studies, mice were immunized as described under Section2.5 and challenged as described above at 14 days after thelast immunization using 5 animals per group. Experiments wererepeated twice, totaling 10 animals per group. Mice were eutha-nized at 4 days after challenge and the organs were weighted andhomogenized for CFU counts. Two independent preliminary trialswere performed to establish the optimal bacterial dose and timepoint.
2.11. Measurement of antibody titers
The titers of specific antibodies in mouse sera (collected at14 days after the last immunization and/or at 14 days after bac-terial challenge) were measured using a previously describedindirect ELISA assay [28]. Briefly, microtiter plates were coated
Mouse sera were serially diluted with antibody buffer containing20 �g/ml of CAT-BIO to block reactivity against epitopes containedin the BIO tag or other contaminants. Plates were then developedusing phosphatase-conjugated goat anti-mouse IgG, as described[28].
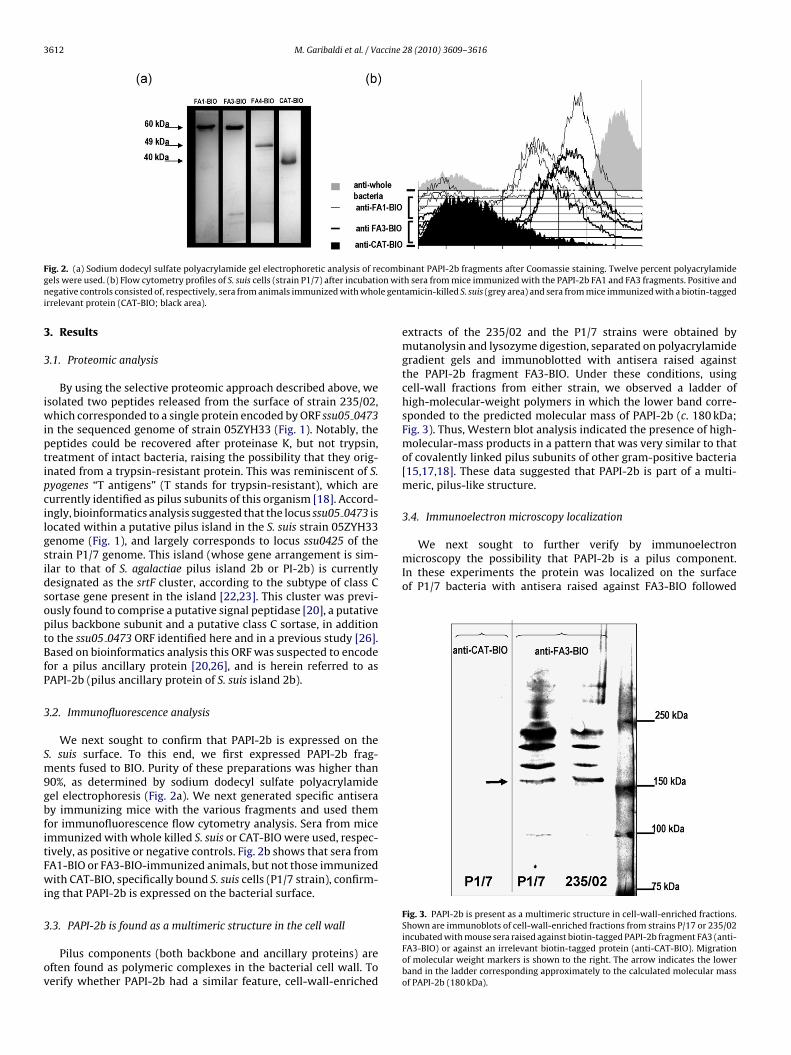
3612 M. Garibaldi et al. / Vaccine 28 (2010) 3609–3616
F combg on witn le genti
3
3
iwiptipcilgsidsoptBfP
3
Sm9gbfitFwi
3
ov
We next sought to further verify by immunoelectronmicroscopy the possibility that PAPI-2b is a pilus component.In these experiments the protein was localized on the surfaceof P1/7 bacteria with antisera raised against FA3-BIO followed
Fig. 3. PAPI-2b is present as a multimeric structure in cell-wall-enriched fractions.
ig. 2. (a) Sodium dodecyl sulfate polyacrylamide gel electrophoretic analysis of reels were used. (b) Flow cytometry profiles of S. suis cells (strain P1/7) after incubatiegative controls consisted of, respectively, sera from animals immunized with who
rrelevant protein (CAT-BIO; black area).
. Results
.1. Proteomic analysis
By using the selective proteomic approach described above, wesolated two peptides released from the surface of strain 235/02,
hich corresponded to a single protein encoded by ORF ssu05 0473n the sequenced genome of strain 05ZYH33 (Fig. 1). Notably, theeptides could be recovered after proteinase K, but not trypsin,reatment of intact bacteria, raising the possibility that they orig-nated from a trypsin-resistant protein. This was reminiscent of S.yogenes “T antigens” (T stands for trypsin-resistant), which areurrently identified as pilus subunits of this organism [18]. Accord-ngly, bioinformatics analysis suggested that the locus ssu05 0473 isocated within a putative pilus island in the S. suis strain 05ZYH33enome (Fig. 1), and largely corresponds to locus ssu0425 of thetrain P1/7 genome. This island (whose gene arrangement is sim-lar to that of S. agalactiae pilus island 2b or PI-2b) is currentlyesignated as the srtF cluster, according to the subtype of class Cortase gene present in the island [22,23]. This cluster was previ-usly found to comprise a putative signal peptidase [20], a putativeilus backbone subunit and a putative class C sortase, in additiono the ssu05 0473 ORF identified here and in a previous study [26].ased on bioinformatics analysis this ORF was suspected to encode
or a pilus ancillary protein [20,26], and is herein referred to asAPI-2b (pilus ancillary protein of S. suis island 2b).
.2. Immunofluorescence analysis
We next sought to confirm that PAPI-2b is expressed on the. suis surface. To this end, we first expressed PAPI-2b frag-ents fused to BIO. Purity of these preparations was higher than
0%, as determined by sodium dodecyl sulfate polyacrylamideel electrophoresis (Fig. 2a). We next generated specific antiseray immunizing mice with the various fragments and used themor immunofluorescence flow cytometry analysis. Sera from micemmunized with whole killed S. suis or CAT-BIO were used, respec-ively, as positive or negative controls. Fig. 2b shows that sera fromA1-BIO or FA3-BIO-immunized animals, but not those immunizedith CAT-BIO, specifically bound S. suis cells (P1/7 strain), confirm-
ng that PAPI-2b is expressed on the bacterial surface.
.3. PAPI-2b is found as a multimeric structure in the cell wall
Pilus components (both backbone and ancillary proteins) areften found as polymeric complexes in the bacterial cell wall. Toerify whether PAPI-2b had a similar feature, cell-wall-enriched
inant PAPI-2b fragments after Coomassie staining. Twelve percent polyacrylamideh sera from mice immunized with the PAPI-2b FA1 and FA3 fragments. Positive andamicin-killed S. suis (grey area) and sera from mice immunized with a biotin-tagged
extracts of the 235/02 and the P1/7 strains were obtained bymutanolysin and lysozyme digestion, separated on polyacrylamidegradient gels and immunoblotted with antisera raised againstthe PAPI-2b fragment FA3-BIO. Under these conditions, usingcell-wall fractions from either strain, we observed a ladder ofhigh-molecular-weight polymers in which the lower band corre-sponded to the predicted molecular mass of PAPI-2b (c. 180 kDa;Fig. 3). Thus, Western blot analysis indicated the presence of high-molecular-mass products in a pattern that was very similar to thatof covalently linked pilus subunits of other gram-positive bacteria[15,17,18]. These data suggested that PAPI-2b is part of a multi-meric, pilus-like structure.
3.4. Immunoelectron microscopy localization
Shown are immunoblots of cell-wall-enriched fractions from strains P/17 or 235/02incubated with mouse sera raised against biotin-tagged PAPI-2b fragment FA3 (anti-FA3-BIO) or against an irrelevant biotin-tagged protein (anti-CAT-BIO). Migrationof molecular weight markers is shown to the right. The arrow indicates the lowerband in the ladder corresponding approximately to the calculated molecular massof PAPI-2b (180 kDa).
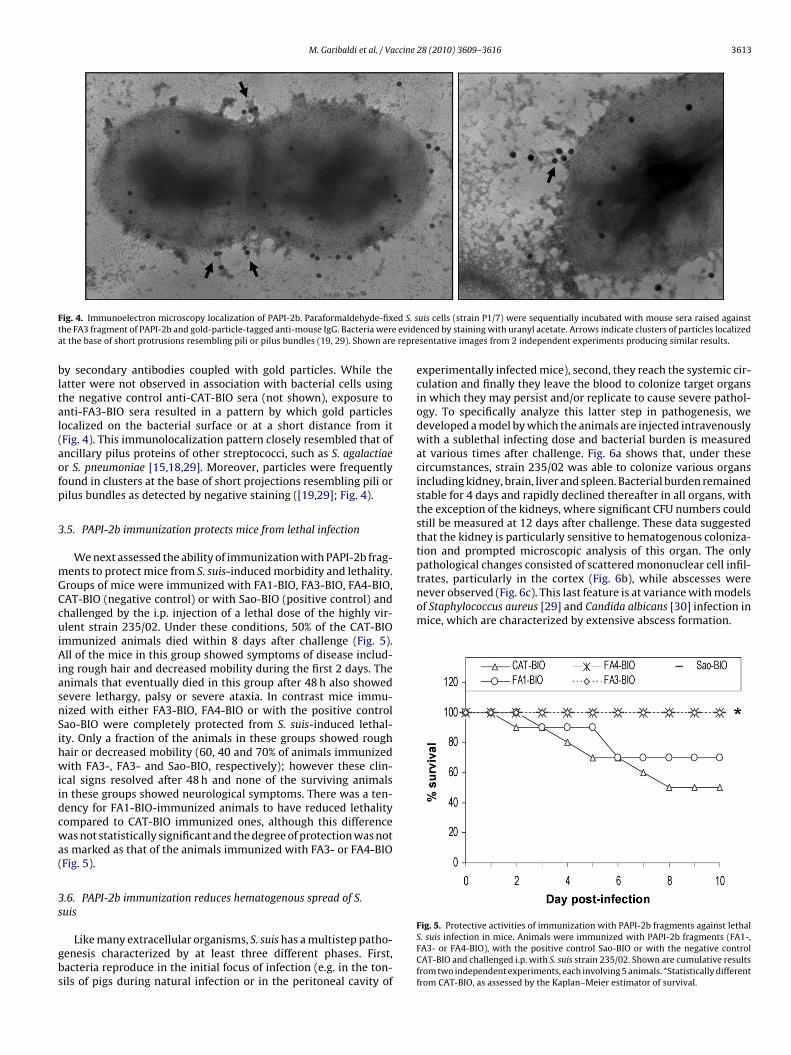
M. Garibaldi et al. / Vaccine 28 (2010) 3609–3616 3613
F ed S. st evidea repre
bltal(aofp
3
mGCcuiAiasnSihwiidcwa(
3s
gbs
trates, particularly in the cortex (Fig. 6b), while abscesses werenever observed (Fig. 6c). This last feature is at variance with modelsof Staphylococcus aureus [29] and Candida albicans [30] infection inmice, which are characterized by extensive abscess formation.
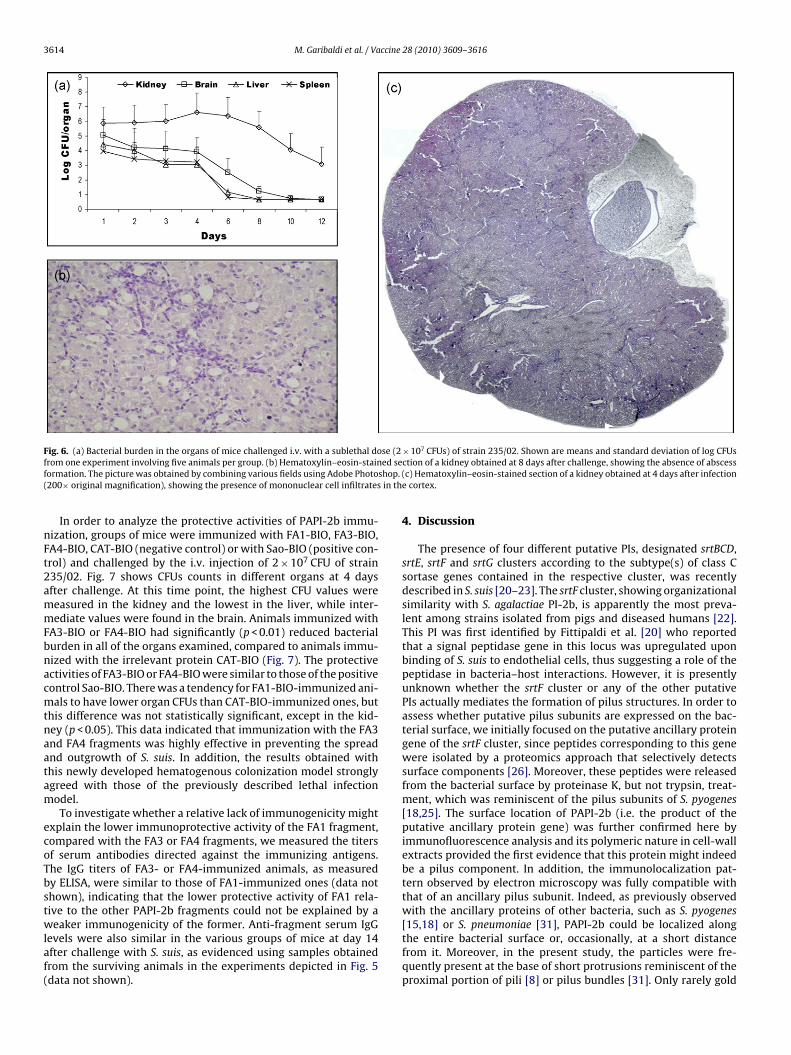
ig. 4. Immunoelectron microscopy localization of PAPI-2b. Paraformaldehyde-fixhe FA3 fragment of PAPI-2b and gold-particle-tagged anti-mouse IgG. Bacteria weret the base of short protrusions resembling pili or pilus bundles (19, 29). Shown are
y secondary antibodies coupled with gold particles. While theatter were not observed in association with bacterial cells usinghe negative control anti-CAT-BIO sera (not shown), exposure tonti-FA3-BIO sera resulted in a pattern by which gold particlesocalized on the bacterial surface or at a short distance from itFig. 4). This immunolocalization pattern closely resembled that ofncillary pilus proteins of other streptococci, such as S. agalactiaer S. pneumoniae [15,18,29]. Moreover, particles were frequentlyound in clusters at the base of short projections resembling pili orilus bundles as detected by negative staining ([19,29]; Fig. 4).
.5. PAPI-2b immunization protects mice from lethal infection
We next assessed the ability of immunization with PAPI-2b frag-ents to protect mice from S. suis-induced morbidity and lethality.roups of mice were immunized with FA1-BIO, FA3-BIO, FA4-BIO,AT-BIO (negative control) or with Sao-BIO (positive control) andhallenged by the i.p. injection of a lethal dose of the highly vir-lent strain 235/02. Under these conditions, 50% of the CAT-BIO
mmunized animals died within 8 days after challenge (Fig. 5).ll of the mice in this group showed symptoms of disease includ-
ng rough hair and decreased mobility during the first 2 days. Thenimals that eventually died in this group after 48 h also showedevere lethargy, palsy or severe ataxia. In contrast mice immu-ized with either FA3-BIO, FA4-BIO or with the positive controlao-BIO were completely protected from S. suis-induced lethal-ty. Only a fraction of the animals in these groups showed roughair or decreased mobility (60, 40 and 70% of animals immunizedith FA3-, FA3- and Sao-BIO, respectively); however these clin-
cal signs resolved after 48 h and none of the surviving animalsn these groups showed neurological symptoms. There was a ten-ency for FA1-BIO-immunized animals to have reduced lethalityompared to CAT-BIO immunized ones, although this differenceas not statistically significant and the degree of protection was not
s marked as that of the animals immunized with FA3- or FA4-BIOFig. 5).
.6. PAPI-2b immunization reduces hematogenous spread of S.uis
Like many extracellular organisms, S. suis has a multistep patho-enesis characterized by at least three different phases. First,acteria reproduce in the initial focus of infection (e.g. in the ton-ils of pigs during natural infection or in the peritoneal cavity of
uis cells (strain P1/7) were sequentially incubated with mouse sera raised againstnced by staining with uranyl acetate. Arrows indicate clusters of particles localizedsentative images from 2 independent experiments producing similar results.
experimentally infected mice), second, they reach the systemic cir-culation and finally they leave the blood to colonize target organsin which they may persist and/or replicate to cause severe pathol-ogy. To specifically analyze this latter step in pathogenesis, wedeveloped a model by which the animals are injected intravenouslywith a sublethal infecting dose and bacterial burden is measuredat various times after challenge. Fig. 6a shows that, under thesecircumstances, strain 235/02 was able to colonize various organsincluding kidney, brain, liver and spleen. Bacterial burden remainedstable for 4 days and rapidly declined thereafter in all organs, withthe exception of the kidneys, where significant CFU numbers couldstill be measured at 12 days after challenge. These data suggestedthat the kidney is particularly sensitive to hematogenous coloniza-tion and prompted microscopic analysis of this organ. The onlypathological changes consisted of scattered mononuclear cell infil-
Fig. 5. Protective activities of immunization with PAPI-2b fragments against lethalS. suis infection in mice. Animals were immunized with PAPI-2b fragments (FA1-,FA3- or FA4-BIO), with the positive control Sao-BIO or with the negative controlCAT-BIO and challenged i.p. with S. suis strain 235/02. Shown are cumulative resultsfrom two independent experiments, each involving 5 animals. *Statistically differentfrom CAT-BIO, as assessed by the Kaplan–Meier estimator of survival.
3614 M. Garibaldi et al. / Vaccine 28 (2010) 3609–3616
F se (2 7
f ed sef shop.( in th
nFt2ammFbnacmtnaatam
ecoTbstwlaf(
ig. 6. (a) Bacterial burden in the organs of mice challenged i.v. with a sublethal dorom one experiment involving five animals per group. (b) Hematoxylin–eosin-stainormation. The picture was obtained by combining various fields using Adobe Photo200× original magnification), showing the presence of mononuclear cell infiltrates
In order to analyze the protective activities of PAPI-2b immu-ization, groups of mice were immunized with FA1-BIO, FA3-BIO,A4-BIO, CAT-BIO (negative control) or with Sao-BIO (positive con-rol) and challenged by the i.v. injection of 2 × 107 CFU of strain35/02. Fig. 7 shows CFUs counts in different organs at 4 daysfter challenge. At this time point, the highest CFU values wereeasured in the kidney and the lowest in the liver, while inter-ediate values were found in the brain. Animals immunized with
A3-BIO or FA4-BIO had significantly (p < 0.01) reduced bacterialurden in all of the organs examined, compared to animals immu-ized with the irrelevant protein CAT-BIO (Fig. 7). The protectivectivities of FA3-BIO or FA4-BIO were similar to those of the positiveontrol Sao-BIO. There was a tendency for FA1-BIO-immunized ani-als to have lower organ CFUs than CAT-BIO-immunized ones, but
his difference was not statistically significant, except in the kid-ey (p < 0.05). This data indicated that immunization with the FA3nd FA4 fragments was highly effective in preventing the spreadnd outgrowth of S. suis. In addition, the results obtained withhis newly developed hematogenous colonization model stronglygreed with those of the previously described lethal infectionodel.To investigate whether a relative lack of immunogenicity might
xplain the lower immunoprotective activity of the FA1 fragment,ompared with the FA3 or FA4 fragments, we measured the titersf serum antibodies directed against the immunizing antigens.he IgG titers of FA3- or FA4-immunized animals, as measuredy ELISA, were similar to those of FA1-immunized ones (data nothown), indicating that the lower protective activity of FA1 rela-ive to the other PAPI-2b fragments could not be explained by a
eaker immunogenicity of the former. Anti-fragment serum IgGevels were also similar in the various groups of mice at day 14fter challenge with S. suis, as evidenced using samples obtainedrom the surviving animals in the experiments depicted in Fig. 5data not shown).
× 10 CFUs) of strain 235/02. Shown are means and standard deviation of log CFUsction of a kidney obtained at 8 days after challenge, showing the absence of abscess(c) Hematoxylin–eosin-stained section of a kidney obtained at 4 days after infectione cortex.
4. Discussion
The presence of four different putative PIs, designated srtBCD,srtE, srtF and srtG clusters according to the subtype(s) of class Csortase genes contained in the respective cluster, was recentlydescribed in S. suis [20–23]. The srtF cluster, showing organizationalsimilarity with S. agalactiae PI-2b, is apparently the most preva-lent among strains isolated from pigs and diseased humans [22].This PI was first identified by Fittipaldi et al. [20] who reportedthat a signal peptidase gene in this locus was upregulated uponbinding of S. suis to endothelial cells, thus suggesting a role of thepeptidase in bacteria–host interactions. However, it is presentlyunknown whether the srtF cluster or any of the other putativePIs actually mediates the formation of pilus structures. In order toassess whether putative pilus subunits are expressed on the bac-terial surface, we initially focused on the putative ancillary proteingene of the srtF cluster, since peptides corresponding to this genewere isolated by a proteomics approach that selectively detectssurface components [26]. Moreover, these peptides were releasedfrom the bacterial surface by proteinase K, but not trypsin, treat-ment, which was reminiscent of the pilus subunits of S. pyogenes[18,25]. The surface location of PAPI-2b (i.e. the product of theputative ancillary protein gene) was further confirmed here byimmunofluorescence analysis and its polymeric nature in cell-wallextracts provided the first evidence that this protein might indeedbe a pilus component. In addition, the immunolocalization pat-tern observed by electron microscopy was fully compatible withthat of an ancillary pilus subunit. Indeed, as previously observedwith the ancillary proteins of other bacteria, such as S. pyogenes
[15,18] or S. pneumoniae [31], PAPI-2b could be localized alongthe entire bacterial surface or, occasionally, at a short distancefrom it. Moreover, in the present study, the particles were fre-quently present at the base of short protrusions reminiscent of theproximal portion of pili [8] or pilus bundles [31]. Only rarely gold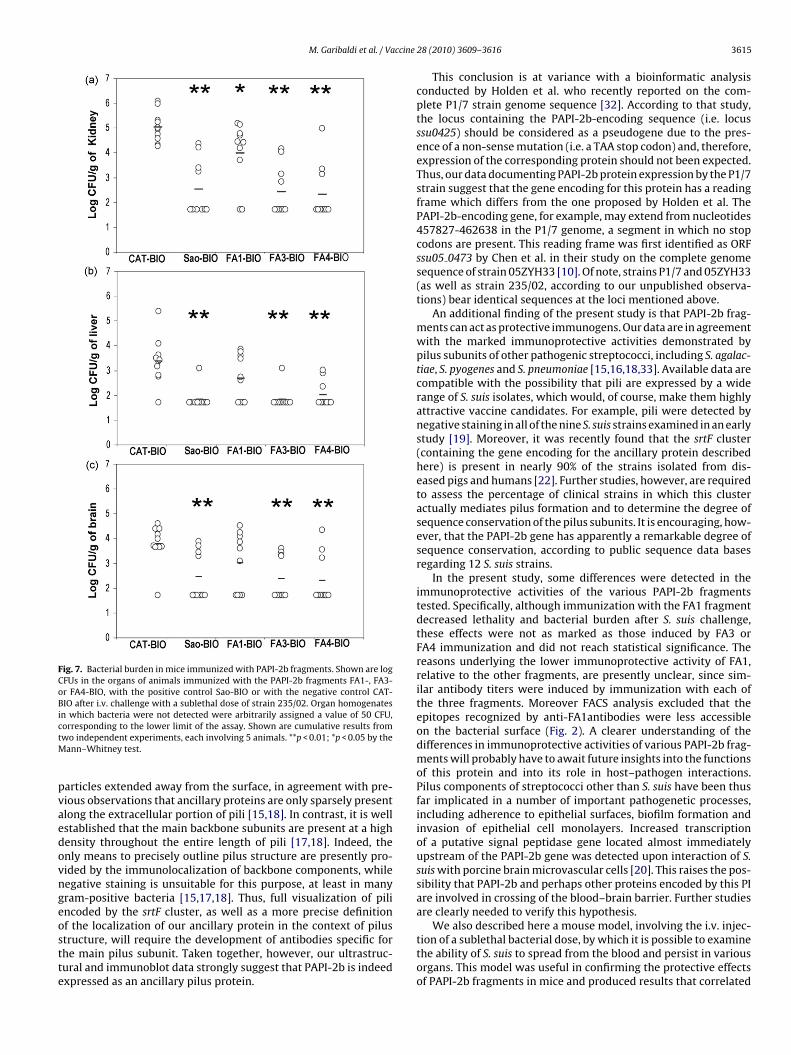
M. Garibaldi et al. / Vaccine
Fig. 7. Bacterial burden in mice immunized with PAPI-2b fragments. Shown are logCFUs in the organs of animals immunized with the PAPI-2b fragments FA1-, FA3-or FA4-BIO, with the positive control Sao-BIO or with the negative control CAT-BIO after i.v. challenge with a sublethal dose of strain 235/02. Organ homogenatesin which bacteria were not detected were arbitrarily assigned a value of 50 CFU,ctM
pvaedovngeostte
We also described here a mouse model, involving the i.v. injec-
orresponding to the lower limit of the assay. Shown are cumulative results fromwo independent experiments, each involving 5 animals. **p < 0.01; *p < 0.05 by the
ann–Whitney test.
articles extended away from the surface, in agreement with pre-ious observations that ancillary proteins are only sparsely presentlong the extracellular portion of pili [15,18]. In contrast, it is wellstablished that the main backbone subunits are present at a highensity throughout the entire length of pili [17,18]. Indeed, thenly means to precisely outline pilus structure are presently pro-ided by the immunolocalization of backbone components, whileegative staining is unsuitable for this purpose, at least in manyram-positive bacteria [15,17,18]. Thus, full visualization of pilincoded by the srtF cluster, as well as a more precise definitionf the localization of our ancillary protein in the context of pilus
tructure, will require the development of antibodies specific forhe main pilus subunit. Taken together, however, our ultrastruc-ural and immunoblot data strongly suggest that PAPI-2b is indeedxpressed as an ancillary pilus protein.28 (2010) 3609–3616 3615
This conclusion is at variance with a bioinformatic analysisconducted by Holden et al. who recently reported on the com-plete P1/7 strain genome sequence [32]. According to that study,the locus containing the PAPI-2b-encoding sequence (i.e. locusssu0425) should be considered as a pseudogene due to the pres-ence of a non-sense mutation (i.e. a TAA stop codon) and, therefore,expression of the corresponding protein should not been expected.Thus, our data documenting PAPI-2b protein expression by the P1/7strain suggest that the gene encoding for this protein has a readingframe which differs from the one proposed by Holden et al. ThePAPI-2b-encoding gene, for example, may extend from nucleotides457827-462638 in the P1/7 genome, a segment in which no stopcodons are present. This reading frame was first identified as ORFssu05 0473 by Chen et al. in their study on the complete genomesequence of strain 05ZYH33 [10]. Of note, strains P1/7 and 05ZYH33(as well as strain 235/02, according to our unpublished observa-tions) bear identical sequences at the loci mentioned above.
An additional finding of the present study is that PAPI-2b frag-ments can act as protective immunogens. Our data are in agreementwith the marked immunoprotective activities demonstrated bypilus subunits of other pathogenic streptococci, including S. agalac-tiae, S. pyogenes and S. pneumoniae [15,16,18,33]. Available data arecompatible with the possibility that pili are expressed by a widerange of S. suis isolates, which would, of course, make them highlyattractive vaccine candidates. For example, pili were detected bynegative staining in all of the nine S. suis strains examined in an earlystudy [19]. Moreover, it was recently found that the srtF cluster(containing the gene encoding for the ancillary protein describedhere) is present in nearly 90% of the strains isolated from dis-eased pigs and humans [22]. Further studies, however, are requiredto assess the percentage of clinical strains in which this clusteractually mediates pilus formation and to determine the degree ofsequence conservation of the pilus subunits. It is encouraging, how-ever, that the PAPI-2b gene has apparently a remarkable degree ofsequence conservation, according to public sequence data basesregarding 12 S. suis strains.
In the present study, some differences were detected in theimmunoprotective activities of the various PAPI-2b fragmentstested. Specifically, although immunization with the FA1 fragmentdecreased lethality and bacterial burden after S. suis challenge,these effects were not as marked as those induced by FA3 orFA4 immunization and did not reach statistical significance. Thereasons underlying the lower immunoprotective activity of FA1,relative to the other fragments, are presently unclear, since sim-ilar antibody titers were induced by immunization with each ofthe three fragments. Moreover FACS analysis excluded that theepitopes recognized by anti-FA1antibodies were less accessibleon the bacterial surface (Fig. 2). A clearer understanding of thedifferences in immunoprotective activities of various PAPI-2b frag-ments will probably have to await future insights into the functionsof this protein and into its role in host–pathogen interactions.Pilus components of streptococci other than S. suis have been thusfar implicated in a number of important pathogenetic processes,including adherence to epithelial surfaces, biofilm formation andinvasion of epithelial cell monolayers. Increased transcriptionof a putative signal peptidase gene located almost immediatelyupstream of the PAPI-2b gene was detected upon interaction of S.suis with porcine brain microvascular cells [20]. This raises the pos-sibility that PAPI-2b and perhaps other proteins encoded by this PIare involved in crossing of the blood–brain barrier. Further studiesare clearly needed to verify this hypothesis.
tion of a sublethal bacterial dose, by which it is possible to examinethe ability of S. suis to spread from the blood and persist in variousorgans. This model was useful in confirming the protective effectsof PAPI-2b fragments in mice and produced results that correlated

3 ccine
wifosbeboltlrostp
ptmt
A
PSMnER
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
616 M. Garibaldi et al. / Va
ell with those of an established immunoprotection model involv-ng lethal infection [27]. Using this newly developed model, it wasound that the kidney is particularly sensitive to hematogenous col-nization and can be used as a reliable indicator of the ability of S.uis to persist in vivo. Our data suggest that measuring bacterialurden in the kidney may be used as a rapid screening method tovaluate the immunoprotective potential of relatively large num-ers of vaccine candidates (such as those identified by proteomics)r to assess the virulence of bacterial mutants (authors’ unpub-ished observations). However, it is presently unknown whetherhis newly described murine kidney colonization model corre-ates with pig models of infection. It should be mentioned, in thisespect, that the kidney is not generally considered as a targetrgan in the course of natural S. suis infections. Therefore, furthertudies are needed to fully validate the mouse kidney coloniza-ion model described here in terms of relevance to infection inigs.
In conclusion, although delineation of the exact role of pili in theathogenesis of S. suis infection awaits ongoing genetic investiga-ions, data presented here suggest that recombinant pilus subunits
ight prove useful in the development of a protein vaccine againsthis pathogen.
cknowledgements
This research was funded by a grant from the “Ramón y Cajal”rogramme (Spanish Ministry of Education), by Project GrantAF2008-00733 (Spanish Ministry of Science and Innovation) to.J.R.-O. and by PRA 2005 and other funds from The Elie Metch-
icoff Department of the University of Messina. We thank Dr.manuela Mazzon for help with electron microscopy and Dr. Paolouggiero for help with microscopic analysis of kidneys.
eferences
[1] Lun ZR, Wang QP, Chen XG, Li AX, Zhu XQ. Streptococcus suis: an emergingzoonotic pathogen. Lancet Infect Dis 2007;7(3):201–9.
[2] Huang YT, Teng LJ, Ho SW, Hsueh PR. Streptococcus suis infection. J MicrobiolImmunol Infect 2005;38(5):306–13.
[3] Higgins R, Gottschalk M. Streptococcal diseases. In: Straw BE, D’Allaire S, Men-geling WL, Taylor DJ, editors. Diseases of swine. Ames, IA: Iowa State UniversityPress; 2005. p. 769–83.
[4] Charland N, Harel J, Kobisch M, Lacasse S, Gottschalk M. Streptococcus suisserotype 2 mutants deficient in capsular expression. Microbiology 1998;144(pt2):325–32.
[5] Baums C, Kaim GU, Fulde M, Ramachandran G, Goethe R, Valentin-WeigandP. Identification of a novel virulence determinant with serum opacificationactivity in Streptococcus suis. Infect Immun 2006;74(11):6154–62.
[6] Gottshalk M, Lacouture S, Dubreuil JS. Characterization of Streptococcus suiscapsular type 2 haemolysin. Microbiology 1995;141(pt 1):189–95.
[7] Vecht U, Arends JP, van der Molen EJ, van Leengoed LA. Differences in virulencebetween two strains of Streptococcus suis type II after experimentally inducedinfection of newborn germ-free pigs. Am J Vet Res 1989;50(7):1037–43.
[8] Vecht U, Wisselink HJ, Jellema ML, Smith HE. Identification of two pro-teins associated with virulence of Streptococcus suis type 2. Infect Immun1991;59:3156–62.
[9] de Greeff A, Buys H, Verhaar R, Dijkstra J, van Alphen L, Smith HE. Contributionof fibronectin-binding protein to pathogenesis of Streptococcus suis serotype 2.Infect Immun 2002;70(3):1319–25.
[
[
28 (2010) 3609–3616
10] Chen C, Tang J, Dong W, Wang C, Feng Y, Wang J, et al. A glimpse of streptococcaltoxic shock syndrome from comparative genomics of S. suis 2 Chinese isolates.PLoS ONE 2007;2(3):e315.
11] Geng H, Zhu L, Yuan Y, Zhang W, Li W, Wang J, et al. Identification and char-acterization of novel immunogenic proteins of Streptococcus suis serotype 2. JProteome Res 2008;7(9):4132–42.
12] Janulczyk R, Rasmussen M. Improved pattern for genome-based screeningidentifies novel cell wall-attached proteins in gram-positive bacteria. InfectImmun 2001;69(6):4019–26.
13] Garrido Aranda J, Cortés ME, Llagostera P, Barbé MJ. Analysis of the protectivecapacity of three Streptococcus suis proteins induced under divalent-cation-limited conditions. Infect Immun 2008;76(4):1590–8.
14] Li Y, Gottschalk M, Esgleas M, Lacouture S, Dubreuil JD, Willson P, et al. Immu-nization with recombinant Sao protein confers protection against Streptococcussuis infection. Clin Vaccine Immunol 2007;14(8):937–43.
15] Mora M, Bensi G, Capo S, Falugi F, Zingaretti C, Manetti AG, et al. Group AStreptococcus produce pilus-like structures containing protective antigens andLancefield T antigens. Proc Natl Acad Sci USA 2005;102(43):15641–6.
16] Gianfaldoni C, Censini S, Hilleringmann M, Maschioni M, Facciotti C, PansegrauW, et al. Streptococcus pneumoniae pilus subunits protect mice against lethalchallenge. Infect Immun 2007;75(2):1059–62.
17] Lauer P, Rinaudo CD, Soriani M, Margarit I, Maione D, Rosini R, et al. Genomeanalysis reveals pili in Group B Streptococcus. Science 2005;309(5731):105.
18] Telford JL, Barocchi MA, Margarit I, Rappuoli R, Grandi G. Pili in gram-positivepathogens. Nat Rev Microbiol 2006;4(7):509–19.
19] Jacques M, Gottschalk M, Foiry B, Higgins R. Ultrastructural study of surfacecomponents of Streptococcus suis. J Bacteriol 1990;172(6):2833–8.
20] Fittipaldi N, Gottschalk M, Vanier G, Daigle FJ, Harel J. Use of selective captureof transcribed sequences to identify genes preferentially expressed by Strepto-coccus suis upon interaction with porcine brain microvascular endothelial cells.Appl Environ Microbiol 2007;73(13):4359–64.
21] Vanier G, Fittipaldi N, Slater JD, de la Cruz Domínguez-Punaro M, Rycroft AN,Segura M, et al. New putative virulence factors of Streptococcus suis involvedin invasion of porcine brain microvascular endothelial cells. Microb Pathog2009;46(1):13–20.
22] Takamatsu D, Nishino H, Ishiji T, Ishii J, Osaki M, Fittipaldi N, et al. Geneticorganization and preferential distribution of putative pilus gene clusters inStreptococcus suis. Vet Microbiol 2009;138(1–2):132–9.
23] Wang C, Li M, Feng Y, Zheng F, Dong Y, Pan X, et al. The involvement of sortase Ain high virulence of STSS-causing Streptococcus suis serotype 2. Arch Microbiol2009;191(1):23–33.
24] Mignard S, Flandrois JP. 16S rRNA sequencing in routine bacterial identification:a 30-month experiment. J Microbiol Methods 2006;67(3):574–81.
25] Rodríguez-Ortega MJ, Norais N, Bensi G, Liberatori S, Capo S, Mora M, etal. Characterization and identification of vaccine candidate proteins throughanalysis of the group A Streptococcus surface proteome. Nat Biotechnol2006;24(2):191–7.
26] Rodríguez-Ortega MJ, Luque I, Tarradas C, Bárcena JA. Overcoming func-tion annotation errors in the Gram-positive pathogen Streptococcus suis by aproteomics-driven approach. BMC Genomics 2008;9:588.
27] Esgleas M, Dominguez-Punaro Mde L, Li Y, Harel J, Dubreuil JD, Gottschalk M.Immunization with SsEno fails to protect mice against challenge with Strepto-coccus suis serotype 2. FEMS Microbiol Lett 2009;294(1):82–8.
28] Lo Passo C, Romeo A, Pernice I, Donato P, Midiri A, Mancuso G, et al. Peptidemimics of the group B meningococcal capsule induce bactericidal and protec-tive antibodies after immunization. J Immunol 2007;178(7):4417–23.
29] Stranger-Jones YK, Bae T, Schneewind O. Vaccine assembly from surface pro-teins of Staphylococcus aureus. Proc Natl Acad Sci USA 2006;103(45):16942–7.
30] Gross O, Gewies A, Finger K, Schäfer M, Sparwasser T, Peschel C, et al. Card9controls a non-TLR signalling pathway for innate anti-fungal immunity. Nature2006;442(7103):651–6.
31] Hilleringmann M, Giusti F, Baudner BC, Masignani V, Covacci A, Rappuoli R, et al.Pneumococcal pili are composed of protofilaments exposing adhesive clustersof Rrg A. PLoS Pathog 2008;4(3):e1000026.
32] Holden MT, Hauser H, Sanders M, Ngo TH, Cherevach I, Cronin A, et al. Rapidevolution of virulence and drug resistance in the emerging zoonotic pathogenStreptococcus suis. PLoS One 2009;4(7):e6072.
33] Margarit I, Rinaudo CD, Galeotti CL, Maione D, Ghezzo C, Buttazzoni E, et al.Preventing bacterial infections with pilus-based vaccines: the group B strepto-coccus paradigm. J Infect Dis 2009;199(1):108–15.
Copyright © 2022 FDOKUMEN




















