Immunochemical assessment of deoxynivalenol tissue distribution following oral exposure in the mouse
14
Immunochemical Assessment of Deoxynivalenol Tissue Distribution Following Oral Exposure in the Mouse James J. Pestka *,†,§,1 , Zahidul Islam * , and Chidozie J. Amuzie §,¶ *Department of Food Science and Human Nutrition, Michigan State University, East Lansing, Michigan 48824 ¶Comparative Medicine and Integrative Biology Program, Michigan State University, East Lansing, Michigan 48824 †Department of Microbiology and Molecular Genetics, Michigan State University, East Lansing, Michigan 48824 §Center for Integrative Toxicology, Michigan State University, East Lansing, Michigan 48824 Abstract Deoxynivalenol (DON or vomitoxin) is a trichothecene mycotoxin commonly found in cereal grains that adversely affects growth and immune function in experimental animals. A competitive enzyme- liked immunosorbent assay (ELISA) was used to monitor the kinetics of distribution and clearance of DON in tissues of young adult B6C3F1 male mice that were orally administered 25 mg/kg bw of the toxin. DON was detectable from 5 min to 24 h in plasma, liver, spleen and brain and from 5 min to 8 h in heart and kidney. The highest DON plasma concentrations were observed within 5 to 15 min (12 μg/ml) after dosing. There was rapid clearance following two-compartment kinetics (t 1/2 α=20.4 min, t 1/2 β=11.8 h) with 5 and 2% maximum plasma DON concentrations remaining after 8 and 24 h, respectively. DON distribution and clearance kinetics in other tissues were similar to that of plasma. At 5 min, DON concentrations in μg/g were 19.5±1.9 in liver, 7.6±0.5 in kidney, 7.3±0.8 in spleen, 6.8±0.9 in heart and 0.8±0.1 in the brain. DON recoveries in tissues by ELISA were comparable to a previous study that employed 3 H-DON and 25 mg/kg bw DON dose. The ELISA was further applicable to the detection of DON in plasma of mice exposed to the toxin via diet. This approach provides a simple strategy that can be used to answer relevant questions in rodents of how dose, species, age, gender, genetic background and route/duration of exposure impact DON uptake and clearance. Introduction The trichothecene mycotoxins are group of more than 200 structurally related sesquiterpenoid metabolites produced by foodborne and environmental fungi that are characterized by the tetracyclic 12,13-epoxytrichothec-9-ene ring system (Grove, 2007). Toxicological effects associated with trichothecene mycotoxin poisoning in humans and animals include anorexia, gastroenteritis, emesis and hematological disorders (Pestka and Casale, 1990). The immune system is extremely sensitive to trichothecenes (Pestka et al., 2004). Exposure to low 1To whom correspondence should be addressed at 234 G.M. Trout Building, Michigan State University, East Lansing, MI 48824-1224. Fax: 517-353-8963. E-mail: E-mail: [email protected]. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Toxicol Lett. Author manuscript; available in PMC 2009 June 15. Published in final edited form as: Toxicol Lett. 2008 May 5; 178(2): 83–87. doi:10.1016/j.toxlet.2008.02.005. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Immunochemical assessment of deoxynivalenol tissue distribution following oral exposure in the mouse

Immunochemical Assessment of Deoxynivalenol TissueDistribution Following Oral Exposure in the Mouse
James J. Pestka*,†,§,1, Zahidul Islam*, and Chidozie J. Amuzie§,¶
*Department of Food Science and Human Nutrition, Michigan State University, East Lansing, Michigan48824
¶Comparative Medicine and Integrative Biology Program, Michigan State University, East Lansing,Michigan 48824
†Department of Microbiology and Molecular Genetics, Michigan State University, East Lansing, Michigan48824
§Center for Integrative Toxicology, Michigan State University, East Lansing, Michigan 48824
AbstractDeoxynivalenol (DON or vomitoxin) is a trichothecene mycotoxin commonly found in cereal grainsthat adversely affects growth and immune function in experimental animals. A competitive enzyme-liked immunosorbent assay (ELISA) was used to monitor the kinetics of distribution and clearanceof DON in tissues of young adult B6C3F1 male mice that were orally administered 25 mg/kg bw ofthe toxin. DON was detectable from 5 min to 24 h in plasma, liver, spleen and brain and from 5 minto 8 h in heart and kidney. The highest DON plasma concentrations were observed within 5 to 15min (12 μg/ml) after dosing. There was rapid clearance following two-compartment kinetics(t1/2α=20.4 min, t1/2β=11.8 h) with 5 and 2% maximum plasma DON concentrations remaining after8 and 24 h, respectively. DON distribution and clearance kinetics in other tissues were similar to thatof plasma. At 5 min, DON concentrations in μg/g were 19.5±1.9 in liver, 7.6±0.5 in kidney, 7.3±0.8in spleen, 6.8±0.9 in heart and 0.8±0.1 in the brain. DON recoveries in tissues by ELISA werecomparable to a previous study that employed 3H-DON and 25 mg/kg bw DON dose. The ELISAwas further applicable to the detection of DON in plasma of mice exposed to the toxin via diet. Thisapproach provides a simple strategy that can be used to answer relevant questions in rodents of howdose, species, age, gender, genetic background and route/duration of exposure impact DON uptakeand clearance.
IntroductionThe trichothecene mycotoxins are group of more than 200 structurally related sesquiterpenoidmetabolites produced by foodborne and environmental fungi that are characterized by thetetracyclic 12,13-epoxytrichothec-9-ene ring system (Grove, 2007). Toxicological effectsassociated with trichothecene mycotoxin poisoning in humans and animals include anorexia,gastroenteritis, emesis and hematological disorders (Pestka and Casale, 1990). The immunesystem is extremely sensitive to trichothecenes (Pestka et al., 2004). Exposure to low
1To whom correspondence should be addressed at 234 G.M. Trout Building, Michigan State University, East Lansing, MI 48824-1224.Fax: 517-353-8963. E-mail: E-mail: [email protected]'s Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptToxicol Lett. Author manuscript; available in PMC 2009 June 15.
Published in final edited form as:Toxicol Lett. 2008 May 5; 178(2): 83–87. doi:10.1016/j.toxlet.2008.02.005.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

trichothecene doses induces rapid, transient upregulation of proinflammatory cytokinescausing immune stimulation, whereas high doses of trichothecenes cause apoptosis inlymphoid tissues resulting in immunosuppression.
Deoxynivalenol (DON, vomitoxin) is a trichothecene commonly found in wheat, barley andcorn that have been infected by the mold Fusarium graminearum (Placinta et al., 1999;Schothorst and van Egmond, 2004). The presence of ppm levels of DON in foods is of majorhuman health concern worldwide (Pestka and Smolinski, 2005). All animal species evaluatedto date are susceptible to DON according to the rank order of swine>mice>rats>poultry≈ruminants (Rotter et al., 1996). Differences in metabolism, absorption, distribution, andelimination of DON among animal species have been suggested to account for this differentialsensitivity (Pestka, 2007).
In addition to species sensitivity, critical questions still remain as to how age, gender, geneticbackground and route of exposure affect absorption distribution, metabolism and excretion ofDON. Radiolabeled DON has been employed to investigate the toxicokinetics and tissuedistribution of this mycotoxin in several animal species in several studies (Azcona-Olivera etal., 1995)(Rotter et al., 1996) (Lake et al., 1987; Lun et al., 1989; Meky et al., 2003; Preluskyet al., 1986; Prelusky et al., 1988). Limitations of such radioisotope approaches include (1)challenges associated with preparing these compounds with high specific activity, (2)unavailability of radiolabeled DON from commercial sources, (3) difficulties applying theiruse in large animal studies and (4) general safety issues related to handling and disposal ofthese materials. While conventional chemical analyses have also been employed to track DONabsorption and distribution in tissues (Prelusky et al., 1985; Prelusky et al., 1990), thesemethods lack sensitivity, require large amounts of tissue and involve extensive sample clean-up.
Mycotoxin immunoassays are rapid, extremely sensitive, do not require extensive clean-upand typically do not require evaporation or concentration (Pestka,1988). Our laboratory wasthe first to describe DON-specific antibodies and an ELISA for this mycotoxin (Casale et al.,1988). Immunochemical assays are now widely employed for the analysis of DON in cerealgrains (Schneider et al., 2004). ELISA might be similarly applicable to detecting DON intissues of experimental animals exposed to this and other trichothecenes. The purpose of thisstudy was to apply the ELISA to monitoring the kinetics of DON distribution and clearance inmouse plasma and tissues following a single oral exposure to the toxin. The results indicatedthat the ELISA was comparable to a previously described radioisotope approach for monitoringDON toxicokinetics, with both showing DON to be quickly distributed throughout the bodyand removed during sequential rapid clearance and slower terminal elimination phases.
Materials and MethodsChemicals
All reagents were of reagent-grade quality or better and purchased from Sigma Chemical Co.(St. Louis, MO) unless otherwise described. For acute oral exposure studies DON waspurchased from Sigma and used. For the feeding study, DON was produced in Fusariumgraminearum R6576 cultures and purified by silica gel chromatography (Clifford et al.,2003). Purity of DON was verified by a single HPLC peak at 224 nm. Concentrated toxinsolutions were handled in a fume hood. Labware that was contaminated with mycotoxin wasdetoxified by soaking for >1 h in 100 mL/L sodium hypochlorite.
Pestka et al. Page 2
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

AnimalsMale B6C3F1 (C57B1/6J×C3H/HeJ) mice (7 wk) were obtained from Charles River (Portage,MI). Animal handling was conducted in accordance with recommendations established by theNational Institutes of Health and were approved by the Michigan State University InstitutionalAnimal Care and Use Committee. Mice were kept in environmentally protected transparentpolypropylene cages with stainless steel wire tops and filter covers and acclimated for 1 wkprior to onset of different experimental treatments. Environmental conditions included 23–25°C, relative humidity of 45–55%, and a 12-h light:dark cycle. Mice were 3 housed per cage andprovided food and water ad libitum.
Experimental designFor the acute exposure study, food and water were withdrawn from cages 1 h prior to DONadministration. DON was administered at 25 mg/kg bw by oral gavage in 250 μl of endotoxin-free water. Food and water were restored immediately after gavaging. Ungavaged animals wereused as controls. At 5, 15, 30 min and 1, 2, 4, 8, 24 h, mice were anesthetized and bloodcollected. Mice were then euthanized and liver, kidney, heart, spleen and brain removed.Plasma was isolated from blood and stored at -20°C. Organs were frozen at -80°C, pulverizedusing a mortar and pestle and then mixed with phosphate-buffered saline (PBS) at a 1:5 ratio.The tissue extract was centrifuged for 10 min at 14,000 × g. The resultant supernatant fractionwas heated to 100°C for 5 min to inactivate enzymes and precipitate proteins. The heated extractwas centrifuged for 10 min at 14,000 × g and the supernatant fraction subsequently used withappropriate dilution for ELISA.
For the subchronic feeding study, purified DON was added at 0, 2, 5, 10 and 20 mg/kg ofpowdered high fat AIN-93 G Purified Rodent Diet 101847 (Dyets Inc, Bethlehem, PA) asdetailed previously (Pestka et al., 1989). Experimental diets were placed in special containersto minimize spillage. Cages were kept in class II ventilated cabinets for the duration of theexperiment. Mice were fed for 4 wk and blood collected from the saphenous vein as describedpreviously (Hem et al., 1998).
Measurement of DON by competitive direct ELISADON was analyzed using a commercial DON ELISA (Veratox for DON, Neogen Corp.,Lansing, MI) which is based on a previously described direct competitive ELISA (Casale etal., 1988). The monoclonal antibody used in this assay binds to DON and 3-ADON but not toother trichothecenes. Since this assay was designed to analyze DON in grain and grain products,the manufacturer's protocol was adapted for use on plasma and organ tissue samples bypreparing separate DON (Sigma) standards (4 to 1,000 ng/ml) in 10% (v/v) human plasma(Innovative Research Inc., Southfield, MI) or 1% bovine serum albumin (w/v) in PBS whichfunctioned to prevent non-specific binding. Samples or standards (100 μl each) were combinedwith 100 μl of DON horseradish peroxidase conjugate solution in mixing wells provided bythe manufacturer. A 100 μl aliquot of this mixture was transferred to antibody-coated wellsand incubated for 5 min at 25°C. Wells were washed, 100 μl of K-blue Max TMB substrate(Neogen, Lansing, MI) added and the wells incubated for an additional 5 min at 25°C. Thereaction was stopped with 100 μl of 2N sulfuric acid and absorbance read on a MolecularDevices ELISA reader (Menlo Park, CA) using 450 nm filter. DON concentrations werequantified from a standard curve using Molecular Devices Softmax software. DON recoveriesof 90 percent or higher were observed in preliminary studies either in which heated liverextracts were spiked directly with the toxin at 12.5 to 250 ng/g tissue and analyzed or in whichliver homogenates were spiked with the toxin at 10 to 250 ng/g tissue, heated and supernatantsanalyzed. Since this ELISA might detect DON as well as some of its metabolites, data werereported as DON equivalents per ml plasma or per g organ tissue.
Pestka et al. Page 3
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Pharmacokinetic analysisA two-compartment open model (Shargel et al., 2004) was employed to calculate toxicokineticparameters. DON concentrations in plasma and tissue were fitted to biexponential expressionto calculate clearance rates (Li et al., 1997).
StatisticsData were analyzed using Sigma Stat for Windows (Jandel Scientific, San Rafael, CA). AKruskal-Wallis One Way Analysis of Variance was performed. Data sets were significantlydifferent when p<0.05.
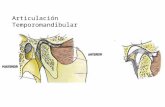
ResultsDON was rapidly absorbed into plasma following oral exposure to a 25 mg/kg bw dose. Thehighest concentrations, 12.1 and 11.5 μg/ml, were detectable at 5 and 15 min, respectively (Fig.1). There was rapid clearance following two-compartment kinetics with half-lives ofdistribution (t1/2α) and clearance (t1/2β) being 20.4 min and 11.8 h, respectively.
DON distribution and clearance kinetics in liver (Fig. 2A) and kidney (Fig. 2B) similarlyfollowed a two-compartment kinetic pattern. As observed for plasma, concentrations of DONpeaked at 5 to 15 min in liver (19.6 and 12.1 μg/g, respectively) (Fig. 2A) and kidney (7.6 and9.0 μg/g, respectively) (Fig. 2B). The t1/2α and t1/2β in liver were 22 min and 19.0 h,respectively, whereas the t1/2α and t1/2β for kidney were 47 min and 20.9 h, respectively.
As seen in spleen and liver, the above tissues, the highest DON concentrations in spleen wereobserved at 5 min (7.3 μg/g) and 15 min (7.9 μg/g (Fig. 3A). The t1/2α and t1/2β were 29 minand 9.0 h, respectively. The highest DON concentrations were observed in the heart at 5 min(6.7 μg/g) and 15 min (6.8 μg/g) with t1/2α and t1/2β being 41 min and 12.3 h, respectively (Fig.3B).
Compared to other tissues, DON entered the brain much more slowly and peaked at muchlower concentrations (0.7-1.0 μg/g), lasting from 5 min to 2 h (Fig. 4).
The effects of subchronic dietary exposure to DON on plasma concentrations of this mycotoxinwere also assessed in a 4 wk feeding study. Dose-dependent increases in plasma DON wereobserved in mice fed the toxin at 2, 5, 10 and 20 mg/kg diet with concentrations ranging from20 to 100 ng/ml (Fig.5). All four of these doses reduced weight gain (data not shown) as hasbeen reported previously for the B6C3F1 mouse (Forsell et al., 1986)
DiscussionThe results presented here suggested that DON uptake and distribution in the mouse could bemonitored by ELISA. DON was rapidly taken up into tissues after exposure and maximumconcentrations followed the rank order liver> plasma> kidney> spleen> heart> brain. DONwas cleared initially at a rapid rate in all tissues except for brain and this was followed by arelatively slower rate of clearance. The data presented here were highly consistent with aprevious [3H]-DON study conducted in the mouse by our laboratory (Azcona-Olivera et al.,1995). That prior study also reported two compartment kinetics with an initial rapid clearancerate and slower terminal elimination in plasma following exposure to DON doses of 5 mg/kgbw (t1/2α = 21.6 min and t1/2β =7.6 h, respectively) and 25 mg/kg (t1/2α = 33.6 min and t1/2β= 88.9 h, respectively). When DON uptake and clearance in plasma, liver and kidney observedhere during the first 4 h were graphically compared to the aforementioned study at the 25 mg/kg bw dose, the results were found to be remarkably similar (Fig. 6). The ELISA was thuscomparable to the radiolabel method for monitoring DON tissue concentrations and
Pestka et al. Page 4
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

toxicokinetics in the mouse. Key advantages of this immunoassay approach are that (1) it wasvery sensitive and specific, (2) it employed a facile sample preparation protocol that did notrequire extensive clean-up and (3) it did not require evaporation or concentration.
The acute dose employed here, 25 mg/kg bw, is equivalent to 1/3 the LD50 previously reportedfor B6C3F1 female mice (Forsell et al., 1987). Although there were no obvious signs of toxicityat this dose, DON disposition and clearance kinetics seen here are highly consistent with thetemporal patterns for biomarkers of stress-related responses DON rapidly induces theexpression of the proinflammatory cytokines (IL-1β, IL-6, TNF-α) that potentially contributeto induction of anorexia and multiple immunologic effects (Azcona-Olivera et al., 1995; Zhouet al., 1997). Cytokine upregulation is mediated by mitogen-activated protein kinase (MAPK)-driven activation of transcription factors and enhancement of mRNA stability (Pestka et al.,2004). In mice, acute oral DON exposure (25 mg/kg bw), sequentially induces in the spleen:(1) phosphorylation of JNK 1/2, ERK 1/2 and p38 (15-30 min), (2) transcription factoractivation (1-2 h), and (3) cytokine mRNA expression (1-4 h) (Zhou et al., 2003a). Thus, thetime required to reach the highest DON concentrations in serum and tissues (5 to 15 min)correlates well with rapid stress-mediated gene expression.
It should be noted that in the subchronic feeding study, plasma DON levels increased in dose-dependent fashion with increasing concentrations of the toxin in diet. The DON concentrationrange of 2 to 20 mg/kg diet utilized here encompasses low and high levels that have previouslybeen observed in human food products (Abouzied et al., 1991) and cause reduced weight gainin mice (Forsell et al.,1986). The DON concentrations found in plasma are also consistent withthose having effects in leukocyte culture studies. For example, in cloned macrophages, DONat 100 ng/ml has previously been shown to rapidly induce MAPK kinase activation as well ascytokine gene expression (Zhou et al., 2003b; Zhou et al., 2005). Furthermore, we haveobserved that DON induces p38 activation in cultured human peripheral blood mononuclearcell cultures at concentrations as low as 25 ng/ml (Islam et al., 2006). Further investigation iswarranted on how prolonged chronic consumption impacts plasma and tissue DONconcentrations of how these effects might affect both kinase signaling and cytokine geneexpression in immunocompetent cells.
DON has also been reported to have neuroendocrine effects (Rotter et al., 1996). It wastherefore notable that this mycotoxin was detected in brain, albeit at peak concentrations thatwere approximately one-tenth those found in other organs. In a study of fate and distributionof the radiolabeled T-2 toxin, in guinea pigs, it was similarly observed that concentrations ofthis trichothecene in brain represented 10 to 25 percent that of other organs (Pace et al.,1985). Thus, while the blood brain barrier might impede uptake of trichothecenes, theyapparently can still enter brain. Interestingly, DON's anorectic and emetic responses have beensuggested to be mediated by the serotoninergic system based on increased levels of serotoninor its metabolites in DON-treated animals as well as the capacity of serotonin receptorantagonists to prevent DON-induced emesis (Prelusky et al., 1992; Prelusky, 1993; Preluskyand Trenholm, 1993). Clarification is needed on whether the neuroendocrine effects of DONresult from direct interaction with the brain or, rather, are secondary effects of proinflammatorycytokines or other mediators evoked by the toxin in other tissues.
Numerous investigations have been conducted in several animal species to elucidate DON'stissue targets, mechanisms of action and threshold doses for adverse effects. These have beenapplied to establish tolerable daily intakes (TDIs) and regulatory limits in specific foods(Canady, 2001; Pieters et al., 2002). The potential for chemicals to differentially affect humansubpopulations is an important consideration when determining TDIs and establishingregulatory standards (Falk-Filipsson et al., 2007). One way that subpopulations can differ inis their ability to distribute and clear a toxin, which will ultimately impact its concentration in
Pestka et al. Page 5
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

a target organ and the magnitude of its pathophysiologic effects. The results presented heresuggest that ELISA could be used to analyze for DON in animal plasma and tissues withoutextensive clean-up and that, in the mouse, the toxin was taken up into tissues throughout thebody within a very short time. This approach provides a simple strategy that can be used toanswer relevant questions of how dose, species, age, gender, genetic background and route/duration of exposure impact DON uptake and clearance in rodents.
AcknowledgmentsThis work was supported in part by the USDA, under a cooperative project with U.S. Wheat and Barley Scab Initiative.Any findings, opinions, conclusions or recommendations expressed in this publication are those of the authors and donot necessarily reflect the views of USDA. This project was also funded in part by Public Health Service Grant ES3358 (JJP) from the National Institute for Environmental Health Sciences. We would like to thank Dr. PavlinaYordanova, Sarah Godbehere and Mary Rosner for technical assistance.
ReferencesAbouzied MM, Azcona JI, Braselton WE, Pestka JJ. Immunochemical assessment of mycotoxins in 1989
grain foods: evidence for deoxynivalenol (vomitoxin) contamination. Appl Environ Microbiol1991;57:672–677. [PubMed: 1828138]
Azcona-Olivera JI, Ouyang Y, Murtha J, Chu FS, Pestka JJ. Induction of cytokine mRNAs in mice afteroral exposure to the trichothecene vomitoxin (deoxynivalenol): relationship to toxin distribution andprotein synthesis inhibition. Toxicol Appl Pharmacol 1995;133:109–120. [PubMed: 7597700]
Canady RA, Coker RD, Egan SK, Krska R, Kuiper-Goodman T, Olsen M, Pestka J, Resnik S, SchlatterJ. Deoxynivalenol. WHO/IPCS Safety evaluation of certain mycotoxins in food 2001;47:419–555.WHO Food Addit. Ser.
Casale WL, Pestka JJ, Hart LP. Enzyme-linked immunosorbent-assay employing monoclonal-antibodyspecific for deoxynivalenol (vomitoxin) and several analogs. J Agric Food Chem 1988;36:663–668.
Clifford LJ, Jia Q, Pestka JJ. An improved method for the purification of the trichothecene deoxynivalenol(vomitoxin) from Fusarium graminearum culture. J Agric Food Chem 2003;51:521–523. [PubMed:12517120]
Falk-Filipsson A, Hanberg A, Victorin K, Warholm M, Wallen M. Assessment factors--applications inhealth risk assessment of chemicals. Environ Res 2007;104:108–127. [PubMed: 17166493]
Forsell JH, Witt MF, Tai JH, Jensen R, Pestka JJ. Effects of 8-week exposure of the B6C3F1 mouse todietary deoxynivalenol (vomitoxin) and zearalenone. Food Chem Toxicol 1986;24:213–219.[PubMed: 2937701]
Forsell JH, Jensen R, Tai JH, Witt MF, Lin WS, Pestka JJ. Comparison of acute toxicities ofdeoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1 mouse. Food Chem Toxicol1987;25:155–162. [PubMed: 3557238]
Grove JF. The trichothecenes and their biosynthesis. Fortschr Chem Org Naturst 2007;88:63–130.[PubMed: 17302179]
Hem A, Smith AJ, Solberg P. Saphenous vein puncture for blood sampling of the mouse, rat, hamster,gerbil, guinea pig, ferret and mink. Lab Anim 1998;32:364–368. [PubMed: 9807749]
Islam Z, Gray JS, Pestka JJ. p38 Mitogen-activated protein kinase mediates IL-8 induction by theribotoxin deoxynivalenol in human monocytes. Toxicol Appl Pharmacol 2006;213:235–244.[PubMed: 16364386]
Lake BG, Phillips JC, Walters DG, Bayley DL, Cook MW, Thomas LV, Gilbert J, Startin JR, BaldwinNC, Bycroft BW. Studies on the metabolism of deoxynivalenol in the rat. Food Chem Toxicol1987;25:589–592. [PubMed: 3623350]
Li S, Marquardt RR, Frohlich AA, Vitti TG, Crow G. Pharmacokinetics of ochratoxin A and itsmetabolites in rats. Toxicol Appl Pharmacol 1997;145:82–90. [PubMed: 9221827]
Lun AK, Moran ET Jr, Young LG, McMillan EG. Absorption and elimination of an oral dose of 3H-deoxynivalenol in colostomized and intact chickens. Bull Environ Contam Toxicol 1989;42:919–925. [PubMed: 2743027]
Pestka et al. Page 6
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Meky FA, Turner PC, Ashcroft AE, Miller JD, Qiao YL, Roth MJ, Wild CP. Development of a urinarybiomarker of human exposure to deoxynivalenol. Food Chem Toxicol 2003;41:265–273. [PubMed:12480302]
Pace JG, Watts MR, Burrows EP, Dinterman RE, Matson C, Hauer EC, Wannemacher RW Jr. Fate anddistribution of 3H-labeled T-2 mycotoxin in guinea pigs. Toxicol Appl Pharmacol 1985;80:377–385.[PubMed: 4035695]
Pestka JJ. Enhanced surveillance of foodborne mycotoxins by immunochemical assay. J Assoc Off AnalytChem 1988;71:1075–1081. [PubMed: 3071526]
Pestka JJ. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim Feed Sci Technol2007;137:283–298.
Pestka JJ, Smolinski AT. Deoxynivalenol: toxicology and potential effects on humans. J Toxicol EnvironHealth B Crit Rev 2005;8:39–69. [PubMed: 15762554]
Pestka JJ, Zhou HR, Moon Y, Chung YJ. Cellular and molecular mechanisms for immune modulationby deoxynivalenol and other trichothecenes: unraveling a paradox. Toxicol Lett 2004;153:61–73.[PubMed: 15342082]
Pestka, JJ.; Casale, WL. Naturally occurring fungal toxins. In: Hriaga, JO.; Simmons, MS., editors. FoodContamination from Environmental Sources. John Wiley & Sons; New York: 1990. p. 613-638.
Pestka JJ, Moorman MA, Warner RL. Dysregulation of IgA production and IgA nephropathy inducedby the trichothecene vomitoxin. Food Chem Toxicol 1989;27:361–368. [PubMed: 2676788]
Pieters MN, Freijer J, Baars BJ, Fiolet DC, van KJ, Slob W. Risk assessment of deoxynivalenol in food:concentration limits, exposure and effects. Adv Exp Med Biol 2002;504:235–248. [PubMed:11924598]
Placinta CM, DMello JPF, Macdonald AMC. A review of worldwide contamination of cereal grains andanimal feed with Fusarium mycotoxins. Anim Feed Sci Technol 1999;78:21–37.
Prelusky DB. The effect of low-level deoxynivalenol on neurotransmitter levels measured in pig cerebralspinal fluid. J Environ Sci Health B 1993;28:731–761. [PubMed: 7903675]
Prelusky DB, Trenholm HL. The efficacy of various classes of anti-emetics in preventing deoxynivalenol-induced vomiting in swine. Nat Toxins 1993;1:296–302. [PubMed: 8167950]
Prelusky DB, Yeung JM, Thompson BK, Trenholm HL. Effect of deoxynivalenol on neurotransmittersin discrete regions of swine brain. Arch Environ Contam Toxicol 1992;22:36–40. [PubMed:1372799]
Prelusky DB, Hartin KE, Trenholm HL. Distribution of deoxynivalenol in cerebral spinal fluid followingadministration to swine and sheep. J Environ Sci Health B 1990;25:395–413. [PubMed: 2144863]
Prelusky DB, Hartin KE, Trenholm HL, Miller JD. Pharmacokinetic fate of 14C-labeled deoxynivalenolin swine. Fundam Appl Toxicol 1988;10:276–286. [PubMed: 3356314]
Prelusky DB, Hamilton RM, Trenholm HL, Miller JD. Tissue distribution and excretion of radioactivityfollowing administration of 14C-labeled deoxynivalenol to White Leghorn hens. Fundam ApplToxicol 1986;7:635–645. [PubMed: 3803758]
Prelusky DB, Veira DM, Trenholm HL. Plasma pharmacokinetics of the mycotoxin deoxynivalenolfollowing oral and intravenous administration to sheep. J Environ Sci Health B 1985;20:603–624.[PubMed: 4093544]
Rotter BA, Prelusky DB, Pestka JJ. Toxicology of deoxynivalenol, vomitoxin. J Toxicol Environ Health1996;48:1–34. [PubMed: 8637056]
Schneider E, Curtui V, Seidler C, Dietrich R, Usleber E, Martlbauer E. Rapid methods for deoxynivalenoland other trichothecenes. Toxicol Lett 2004;153:113–121. [PubMed: 15342088]
Schothorst RC, van Egmond HP. Report from SCOOP task 3.2.10 “collection of occurrence data ofFusarium toxins in food and assessment of dietary intake by the population of EU member states”.Subtask: trichothecenes. Toxicol Lett 2004;153:133–143. [PubMed: 15342090]
Shargel, L.; Wu-Pong, S.; Yu, ABC. Applied Biopharmaceutics and Pharmacokinetics. McGraw-HillMedical; New York, NY: 2004.
Zhou HR, Jia Q, Pestka JJ. Ribotoxic stress response to the trichothecene deoxynivalenol in themacrophage involves the SRC family kinase Hck. Toxicol Sci 2005;85:916–926. [PubMed:15772366]
Pestka et al. Page 7
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Zhou HR, Islam Z, Pestka JJ. Rapid, sequential activation of mitogen-activated protein kinases andtranscription factors precedes proinflammatory cytokine mRNA expression in spleens of miceexposed to the trichothecene vomitoxin. Toxicol Sci 2003a;72:130–142. [PubMed: 12604842]
Zhou HR, Lau AS, Pestka JJ. Role of double-stranded RNA-activated protein kinase R (PKR) indeoxynivalenol-induced ribotoxic stress response. Toxicol Sci 2003b;74:335–344. [PubMed:12773753]
Zhou HR, Yan D, Pestka JJ. Differential cytokine mRNA expression in mice after oral exposure to thetrichothecene vomitoxin, deoxynivalenol: dose response and time course. Toxicol Appl Pharmacol1997;144:294–305. [PubMed: 9194413]
Pestka et al. Page 8
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
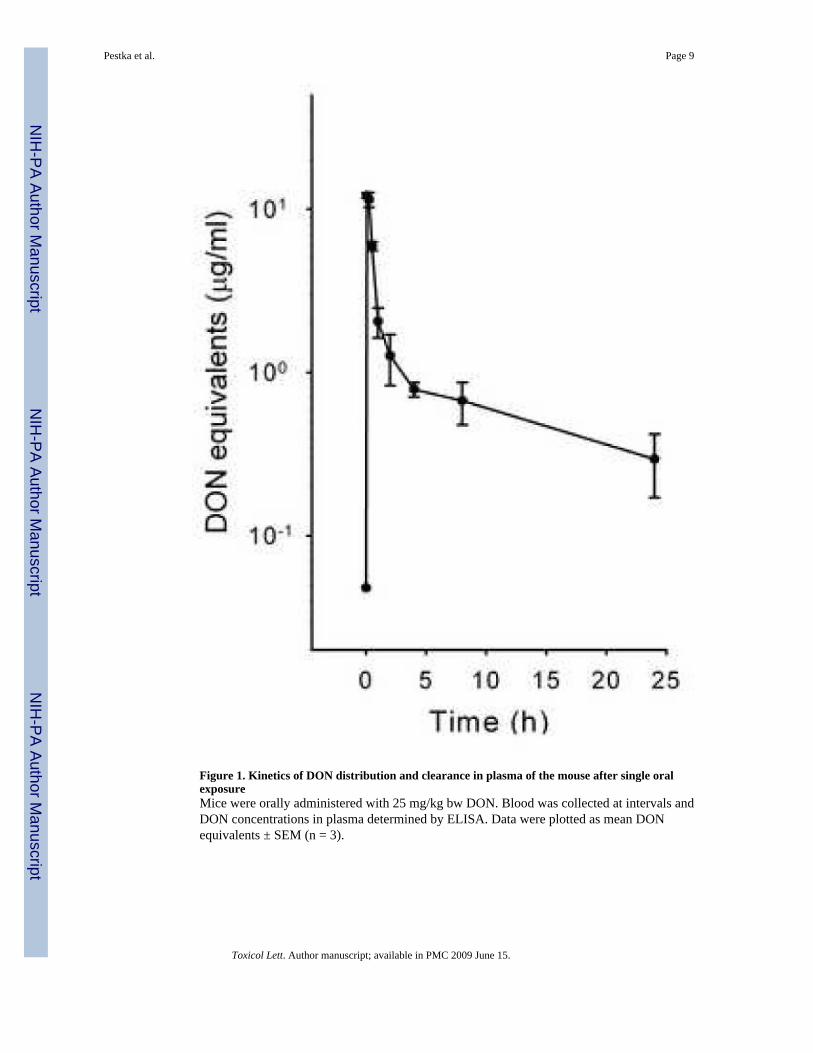
Figure 1. Kinetics of DON distribution and clearance in plasma of the mouse after single oralexposureMice were orally administered with 25 mg/kg bw DON. Blood was collected at intervals andDON concentrations in plasma determined by ELISA. Data were plotted as mean DONequivalents ± SEM (n = 3).
Pestka et al. Page 9
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
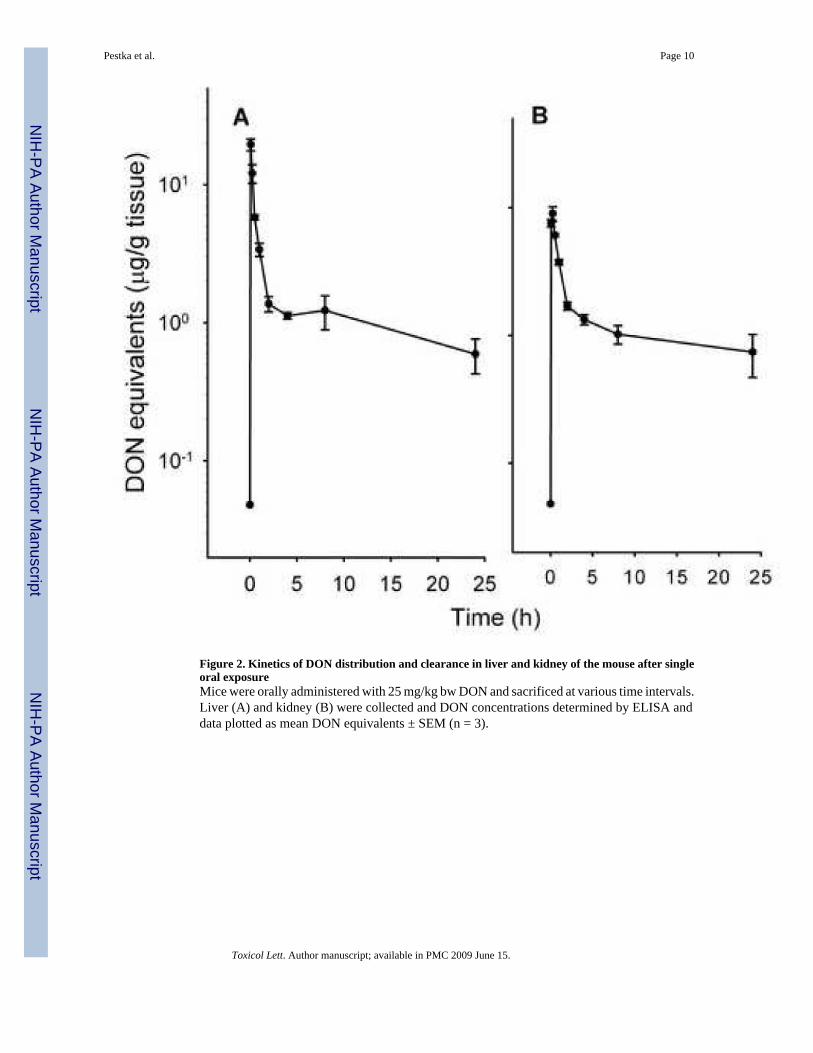
Figure 2. Kinetics of DON distribution and clearance in liver and kidney of the mouse after singleoral exposureMice were orally administered with 25 mg/kg bw DON and sacrificed at various time intervals.Liver (A) and kidney (B) were collected and DON concentrations determined by ELISA anddata plotted as mean DON equivalents ± SEM (n = 3).
Pestka et al. Page 10
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Kinetics of DON distribution and clearance in spleen and heart of the mouse after singleoral exposureMice were orally administered with 25 mg/kg bw DON. Spleen (A) and heart (B) were collectedand DON concentrations determined by ELISA and data plotted as mean DON equivalents ±SEM (n = 3).
Pestka et al. Page 11
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
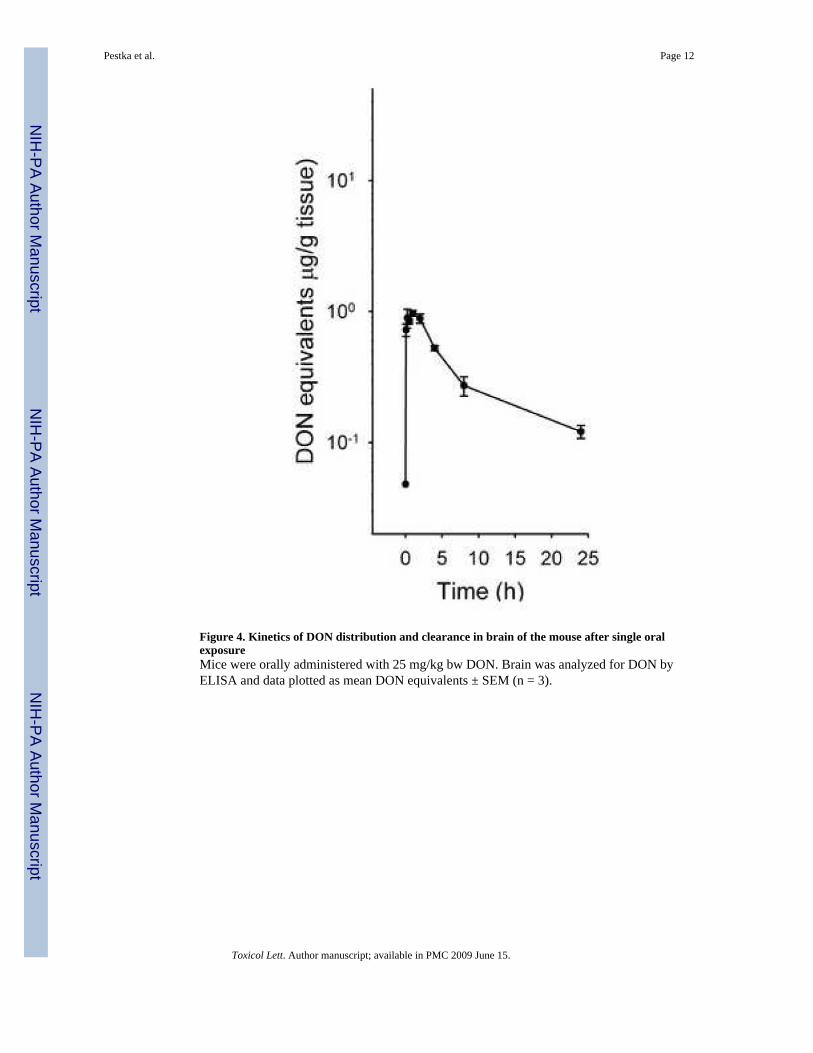
Figure 4. Kinetics of DON distribution and clearance in brain of the mouse after single oralexposureMice were orally administered with 25 mg/kg bw DON. Brain was analyzed for DON byELISA and data plotted as mean DON equivalents ± SEM (n = 3).
Pestka et al. Page 12
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. Dose-dependent effects of short-term DON consumption on DON plasma concentrationsMice were fed various concentrations of DON for 4 wk and blood collected. Plasma wasanalyzed for DON by ELISA. Data plotted as mean DON equivalents ± SEM (n = 3).
Pestka et al. Page 13
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6. Estimations by ELISA and radiolabel methods of DON distribution in plasma, liver andkidney after a single oral exposure are comparableData, determined as described in Figs. 1 and 2, were arithmetically plotted as mean DONequivalents (solid circles) and compared to data from a similar previous study (Azcona-Oliveraet al. 1995) conducted with [3H]-DON and employing the identical DON dose and mouse strain(open circles).
Pestka et al. Page 14
Toxicol Lett. Author manuscript; available in PMC 2009 June 15.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript