
Immune responses against human papillomavirus (HPV) type
10
Journal of General Virology (1999), 80, 399–408. Printed in Great Britain ................................................................................................................................................................................................................................................................................... Immune responses against human papillomavirus (HPV) type 16 virus-like particles in a cohort study of women with cervical intraepithelial neoplasia I. Differential T-helper and IgG responses in relation to HPV infection and disease outcome Tanja D. de Gruijl, 1 Hetty J. Bontkes, 1 Jan M. M. Walboomers, 1 Pierre Coursaget, 3 Marij J. Stukart, 1 Catherine Dupuy, 3 Esther Kueter, 1 Rene ! H. M. Verheijen, 2 Theo J. M. Helmerhorst, 2 † Margaret F. Duggan-Keen, 4 Peter L. Stern, 4 Chris J. L. M. Meijer 1 and Rik J. Scheper 1 1, 2 Departments of Pathology 1 and Obstetrics and Gynaecology 2 , Free University Hospital, PO Box 7057, 1007 MB Amsterdam, The Netherlands 3 Laboratoire d’Immunologie des Maladies Infectieuses, Faculte ! de Pharmacie, Tours, France 4 Department of Immunology, Paterson Institute for Cancer Research, Christie Hospital NHS Trust, Manchester, UK T-helper (Th) cell-dependent IL-2 production and plasma IgG responses to virus-like particles consisting of the human papillomavirus type 16 (HPV-16) major capsid protein L1 (L1-VLP) were determined in patients with cytological evidence of cervical intraepithelial neoplasia (CIN) participating in a non-intervention prospective cohort study. IgG responses were associated with HPV-16 persistence and high-grade CIN lesions, while high frequencies of Th responses were observed in patients with both virus clearance and virus persistence, irrespective of CIN grade. The IgG response was found in conjunction with an IL-2 response to L1-VLP in 87 % of the patients. Recognition of the HPV-16 L1 Th epitope (amino acids 311–335) was found to be more closely associated than recognition of L1-VLP as a whole to HPV exposure and CIN development. Among the HPV-16 M patients included in this study, those showing a Th response to amino acids 311–335 were more likely to carry the HLA DRB1*11/DQB1*0301 haplotype, while those showing an IgG response to L1-VLP were more likely to carry DRB1*0101/DQB1*0501. However, neither cell- mediated nor humoral immune responses against HPV-16 L1 appear to be sufficient for the natural control of HPV infection and CIN development. Introduction Mucosal, oncogenic human papillomaviruses (HPV), of which HPV-16 is the most prevalent, play an important role in the development of cancer of the cervix uteri and its precursor lesions (IARC, 1995). Failure of the host immune system to control HPV infections may contribute to virus persistence and the concomitant development and progression of premalignant cervical lesions (Frazer, 1996 ; Tindle, 1996). Author for correspondence : Jan Walboomers. Fax ›31 20 4442964. e-mail jmm.walboomers!azvu.nl † Present address : Dept of Gynaecology, University Hospital Rotterdam (Dijkzigt), Erasmus University, Rotterdam, The Netherlands. The possibility of immunological intervention in cervical neoplasia has been widely discussed (Tindle, 1996) and protocols for prophylactic and}or therapeutic immunization against HPV-derived proteins are in development (Borysiewicz et al., 1996 ; Tindle, 1996). For prophylactic vaccination, the use of the HPV-16 capsid proteins, particularly in the form of virus-like particles (VLP) comprising either L1 or L1 in combination with L2, to induce neutralizing antibodies has been proposed (Schiller & Roden, 1995 ; Frazer, 1996 ; Tindle, 1996 ; Kirnbauer et al., 1993; Le Cann et al., 1994). However, the presence of IgG antibody responses against HPV-16 capsid proteins in humans does not appear to correlate with the prevention or clearance of HPV-induced cervical lesions (Carter et al., 1996 ; Wikstro $ m et al., 1995 ; Chua et al., 1996 ; de Gruijl 0001-5973 # 1999 SGM DJJ
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Immune responses against human papillomavirus (HPV) type

Journal of General Virology (1999), 80, 399–408. Printed in Great Britain. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Immune responses against human papillomavirus (HPV) type16 virus-like particles in a cohort study of women with cervicalintraepithelial neoplasia
I. Differential T-helper and IgG responses in relation to HPVinfection and disease outcome
Tanja D. de Gruijl,1 Hetty J. Bontkes,1 Jan M. M. Walboomers,1 Pierre Coursaget,3 Marij J. Stukart,1
Catherine Dupuy,3 Esther Kueter,1 Rene! H. M. Verheijen,2 Theo J. M. Helmerhorst,2†Margaret F. Duggan-Keen,4 Peter L. Stern,4 Chris J. L. M. Meijer1 and Rik J. Scheper1
1, 2 Departments of Pathology1 and Obstetrics and Gynaecology2, Free University Hospital, PO Box 7057, 1007 MB Amsterdam,The Netherlands3 Laboratoire d’Immunologie des Maladies Infectieuses, Faculte! de Pharmacie, Tours, France4 Department of Immunology, Paterson Institute for Cancer Research, Christie Hospital NHS Trust, Manchester, UK
T-helper (Th) cell-dependent IL-2 production and plasma IgG responses to virus-like particlesconsisting of the human papillomavirus type 16 (HPV-16) major capsid protein L1 (L1-VLP) weredetermined in patients with cytological evidence of cervical intraepithelial neoplasia (CIN)participating in a non-intervention prospective cohort study. IgG responses were associated withHPV-16 persistence and high-grade CIN lesions, while high frequencies of Th responses wereobserved in patients with both virus clearance and virus persistence, irrespective of CIN grade. TheIgG response was found in conjunction with an IL-2 response to L1-VLP in 87% of the patients.Recognition of the HPV-16 L1 Th epitope (amino acids 311–335) was found to be more closelyassociated than recognition of L1-VLP as a whole to HPV exposure and CIN development. Amongthe HPV-16M patients included in this study, those showing a Th response to amino acids 311–335were more likely to carry the HLA DRB1*11/DQB1*0301 haplotype, while those showing an IgGresponse to L1-VLP were more likely to carry DRB1*0101/DQB1*0501. However, neither cell-mediated nor humoral immune responses against HPV-16 L1 appear to be sufficient for the naturalcontrol of HPV infection and CIN development.
IntroductionMucosal, oncogenic human papillomaviruses (HPV), of
which HPV-16 is the most prevalent, play an important role inthe development of cancer of the cervix uteri and its precursorlesions (IARC, 1995). Failure of the host immune system tocontrol HPV infections may contribute to virus persistence andthe concomitant development and progression of premalignantcervical lesions (Frazer, 1996 ; Tindle, 1996).
Author for correspondence: Jan Walboomers.
Fax 31 20 4442964. e-mail jmm.walboomers!azvu.nl
† Present address: Dept of Gynaecology, University Hospital
Rotterdam (Dijkzigt), Erasmus University, Rotterdam, The Netherlands.
The possibility of immunological intervention in cervicalneoplasia has been widely discussed (Tindle, 1996) andprotocols for prophylactic and}or therapeutic immunizationagainst HPV-derived proteins are in development (Borysiewiczet al., 1996 ; Tindle, 1996). For prophylactic vaccination, the useof the HPV-16 capsid proteins, particularly in the form ofvirus-like particles (VLP) comprising either L1 or L1 incombination with L2, to induce neutralizing antibodies hasbeen proposed (Schiller & Roden, 1995 ; Frazer, 1996 ; Tindle,1996 ; Kirnbauer et al., 1993 ; Le Cann et al., 1994). However,the presence of IgG antibody responses against HPV-16 capsidproteins in humans does not appear to correlate with theprevention or clearance ofHPV-induced cervical lesions (Carteret al., 1996 ; Wikstro$ m et al., 1995 ; Chua et al., 1996 ; de Gruijl
0001-5973 # 1999 SGM DJJ

T. D. de Gruijl and othersT. D. de Gruijl and others
et al., 1997) and can be correlated with increased risk of thedevelopment of cervical cancer (Nonnenmacher et al., 1995 ;Lehtinen et al., 1996).
Patients with cervical intraepithelial neoplasia (CIN) showevidence of T-helper (Th) cell responses to HPV-16 L1-derivedpeptide sequences in a proliferation assay (Shepherd et al.,1997) and delayed-type hypersensitivity reactions after cu-taneous injection of a recombinant HPV-16 L1 fusion protein(Ho$ pfl et al., 1991). How these responses influence HPVinfection and disease outcome is unknown. This study wasperformed to determine the relationship between cellular (Thcell-dependent IL-2 release) and humoral (plasma IgG re-activity) immune responses against HPV-16 L1 VLPs (L1-VLP)and the possible influence of these responses on the clearanceof virus infection and the behaviour of premalignant cervicallesions in a non-intervention follow-up study of patients withCIN.
Methods+ Controls, patients and study design. Sixteen umbilical cordblood samples were included in the study as immunologically naivenegative controls (UC controls). Nine HPV-16+ women attending ageneral city hospital in Amsterdam and participating in a follow-up studyof HPV-positive women, with normal cervical cytology (Pap 1 or Pap 2)and no history of previous cervical lesions, were included in the study ascontrols for disease status (mean age 36±6 years ; SD 9±6).
The study of women with cervical dysplasia described in this paperwas nested within a larger prospective non-intervention cohort study ofDutch women with CIN that was designed and conducted to study therelationship between HPV infection patterns and the natural diseasecourse of premalignant cervical lesions (Remmink et al., 1995). Womenwith cytologically determined mild to moderate cervical dyskaryosiswere referred to the gynaecological outpatient clinic of the FreeUniversity Hospital in Amsterdam and, after giving written, informedconsent, were enrolled in the study. The study design was approved bythe ethics committee of the hospital.
Clinical follow-up consisted of cytological and colposcopical exami-nations that were performed every 3–4 months. Biopsies were not taken
Table 1. Characteristics of patient groups from the CIN cohort study at the time of cross-sectional analysis at the end of follow-up
Patients were allocated to one of four groups, as described in the text. Patient ages and the period of clinicalfollow-up are given as means ³SD. Disease outcomes, as histological diagnoses of biopsies taken at the endof follow up, were allocated to one of four classes as outlined in the text, and are given as numbers in eachclass with percentages of the total in that group in parentheses.
Disease outcome (%)Age Clinical follow-up
Patient group (n) (years) (months) CIN 0 CIN I CIN II CIN III
HPV− (14) 38±3³8±1 33±6³11±1 5 (36) 8 (57) 1 (7) 0 (0)HPV non-16 (13) 32±0³5±8 34±0³8±7 5 (38) 5 (38) 1 (8) 2 (15)HPV-16 C (15) 35±7³8±3 46±9³13±5 6 (40) 8 (53) 1 (7) 0 (0)HPV-16 P (21) 35±0³9±1 34±3³12±7 2 (10) 5 (24) 1 (5) 13 (62)
during the follow-up period, to avoid interference with the natural courseof the disease. Results from the cytomorphological tests were classifiedaccording to a modified Papanicolaou system, as employed in theNetherlands (Remmink et al., 1995) : Pap 1, no cytomorphologicalabnormalities ; Pap 2, inflammation ; Pap 3a, mild to moderatedyskaryosis ; Pap 3b, severe dyskaryosis ; Pap 4, carcinoma in situ ; Pap 5,(microinvasive) carcinoma. Colposcopy was employed to indicate thepredicted grade of dysplasia and to determine the extent of the observedlesions in cervical quadrants. At the end of follow-up, biopsies were takenand a histological diagnosis was made (CIN 0, normal or metaplasticepithelium; CIN I, mild dysplasia ; CIN II, moderate dysplasia ; CIN III,severe dysplasia or carcinoma in situ). A detailed description of the tightclinical surveillance of the patients and the evaluation of clinical follow-up was reported previously (Remmink et al., 1995).
Every 3–6 months, 40 ml heparinized peripheral blood was drawn.Blood collection started 10–56 months (varying for each patient) afterenrolment in the study (de Gruijl et al., 1997, 1998). Blood samples from187 patients, who consented to one or more blood donations, werecollected. From these patients, 63 women were selected to be tested in across-sectional analysis at the end of follow-up (i.e. at the time whenbiopsies were taken) on the basis of HPV infection patterns and torepresent the following groups : patients who were negative for HPVduring the entire follow-up period and for at least 12 months (n¯ 14,HPV−) ; patients with a current or past infection of HPV types other thanHPV-16 (n¯ 13, HPV non-16 : HPV-18, -31, -33, -51, -52, -58 or -66) ;patients with cleared HPV-16 infections (n¯ 15, HPV-16 C; mean timesince the patients were last determined to be positive for HPV-16 DNA,28±5 months ; range 12–65 months) ; and patients with persistent HPV-16infections (n¯ 21, HPV-16 P, positive for HPV-16 DNA in 4–11consecutive PCR tests over a period of 13–59 months). Mean ages,clinical follow-up times and histologically determined disease outcomefor all the CIN patient groups are presented in Table 1.
The CIN patients tested in the cross-sectional analysis (n¯ 63) had amean clinical follow-up period of 37±1 months (range 13–68). Of thesewomen, five who were HPV−, 12 from the HPV-16 C group and 13 fromthe HPV-16 P group were longitudinally tested at two to four separatetime-points over a preceding period of 9–31 months (mean 17±7 months ;SD 6±0). At the start of the follow-up period, the median time since HPV-16 DNA was last detected among the patients with cleared infections was9 months (range ®3 to 34 months).
Plasma samples were available from 54 of the 63 CIN patients and ofall nine HPV-16+ women with normal cervical cytology. These samples
EAA

Th and IgG responses to HPV-16 L1-VLPTh and IgG responses to HPV-16 L1-VLP
were tested for both IgG and IL-2 reactivity to HPV-16 L1-VLP. Of thepatients tested in follow-up, six of 13 HPV-16 P women, six of 12 HPV-16 C women and four of five HPV− women could be tested in parallel forboth IL-2 and IgG responses.
+ HPV genotyping. HPV detection and genotyping were performedon cellular material from cervical smears as described elsewhere (de RodaHusman et al., 1994, 1995). Samples were tested for the following genitalHPV types : 6, 11, 13, 16, 18, 30–33, 35, 39, 40, 42–45, 51, 52, 54–56, 58,59 and 61 (de Roda Husman et al., 1994 ; Jacobs et al., 1995).
+ HLA-DRB1 and -DQB1 typing. DRB1 and DQB1 typing wasperformed on peripheral blood mononuclear cells (PBMC) as previouslydescribed by using the second British Society of Histocompatibility andImmunogenetics (BSHI) class II oligotyping kit (Duggan-Keen et al.,1996).
+ Synthetic peptides. Two 15-mer peptides with a five amino acidoverlap were synthesized. These peptides were derived from the HPV-16L1 sequence and were previously found to contain Th cell epitopes (P1,amino acids 311–325, NLASSNYFPTPSGSM; P2, amino acids 321–335,PSGSMVTSDAQIFNK) (Shepherd et al., 1997). The peptides weresynthesized with a Millipore 9050 peptide synthesizer employing theFmoc method, dissolved in DMSO to a stock concentration of 10 mMand further diluted in culture medium.
+ HPV-16 L1-VLP. Self-assembled HPV-16 L1-VLP were expressedin baculovirus-infected Spodoptera frugiperda (Sf)-21 insect cells, isolatedand purified by sucrose gradient and caesium chloride gradientcentrifugation, as described elsewhere (Le Cann et al., 1994). Gradientfractions were tested for the presence and density of VLP by electronmicroscopy and ELISA (Le Cann et al., 1994). For use in T cell in vitrocultures, the VLP fraction was further purified by a second round ofsucrose and caesium chloride centrifugation, as previously reported(Dupuy et al., 1997). The same fractions derived from Sf-21 cells infectedwith wild-type (wt) baculoviruses were collected and used as controlpreparations in the T cell IL-2 production assay (Dupuy et al., 1997). TheVLP were suspended in PBS and used in ELISA and T cell cultures.
+ Isolation of PBMC. PBMC were isolated from 40 ml heparinizedblood by density centrifugation with Lymphoprep (Nycomed) andcryopreserved as previously described (de Gruijl et al., 1996).
+ T cell culture and stimulation. PBMC were seeded in round-bottomed, 96-well culture plates (Nunclon Delta, Intermed) at 2¬10&
cells per well in Iscove’s modified Dulbecco’s medium containing 10%human pooled serum (CLB, Amsterdam), 50 U}ml penicillin–streptomycin, 1±6 mM -glutamine and 0±01 mM β-mercaptoethanol.Cells were stimulated by the addition of HPV-16 L1-VLP, syntheticpeptides or, as a positive control, phytohaemagglutinin (PHA) (Murex)and cultured for 7 days at 37 °C in an incubator with a humidified 5%CO
#atmosphere. The anti-CD25 monoclonal antibody TB30 (hybridoma
supernatant at a final dilution of 1 :25 ; a kind gift of Dr R. van Lier, CLB,Amsterdam) was added to all wells to prevent IL-2 consumption. Thestimulatory VLP concentration used (1±5 µg}ml) was found to be optimalin titration experiments. The L1 peptides were added to the cultures at aconcentration of 10 µM, as previously described (Shepherd et al., 1997).PHA was used at 30 µg}ml. PBMC cultured in plain culture mediumserved as a negative control. All culture conditions were carried out intriplicate or quadruplicate wells. After 7 days the culture supernatantswere harvested, pooled for each test condition and stored at ®20 °Cuntil further use.
To check whether the IL-2 detected was derived from CD4+ or CD8+
T cells, cultures were incubated under the conditions described aboveafter CD4+ or CD8+ cells had been depleted. PBMC were first incubatedconsecutively with anti-CD4 or anti-CD8 mouse monoclonal antibodies
(Becton Dickinson) and goat anti-mouse IgG antibodies conjugated toMACS magnetic microbeads (Miltenyi Biotec). CD4+ or CD8+ cells werethen removed with the MiniMACS magnetic microbead separationsystem (Miltenyi Biotec), according to the manufacturer’s instructions. Tocheck the CD4+}CD8+ cell depletion, CD4 and CD8 FACS analyses wereperformed after double staining with fluorescein isothiocyanate (FITC)-and phycoerythrin-conjugated antibodies (Simulset, Becton Dickinson).In addition, PBMC were cultured with L1-VLP in the presence of the HLAclass II-blocking monoclonal antibody CR3}43 (anti-DP}DQ}DR, Dako)at 10 µg}ml. The isotype-matched MOPC 21 monoclonal antibody wasused at the same concentration as a negative control. All antibodies weredialysed against PBS before use in order to remove NaN
$.
+ IL-2 bioassay. Production of IL-2 in culture supernatants wasmeasured in a bioassay with the IL-2-dependent cell line HT2 (Ertl et al.,1989). HT2 cells (1¬10% per well) in Iscove’s modified Dulbecco’smedium supplemented with 50 U}ml penicillin–streptomycin, 1±6 mM -glutamine, 0±01 mM β-mercaptoethanol and 10% FCS were cultured for24 h with PBMC culture supernatants used at final dilutions of 1 :2, 1 :4and 1 :8. Triplicate wells were set up for each test condition andsupernatant dilution. During the last 4 h, the cells were incubated with[$H]thymidine (0±4 µCi per well) (Amersham). The cells were harvestedonto fibreglass filters and [$H]thymidine incorporation was determinedwith a flat-bed liquid scintillation counter (Wallac). IL-2 titration curveswere included in each assay (100, 50, 25, 12, 6, 3, 1±5, 0±75, 0±375 and0 IU}ml IL-2) (Cetus). Counts in the L1 test wells never exceeded thelinear range of the titration curves (usually between 12±5 and0±375 IU}ml). Samples were considered positive when the mean HT2proliferation (c.p.m.) in the test wells exceeded proliferation in themedium control wells by a factor of two (HT2 stimulation index, SI
HT#" 2) for at least two of the tested culture supernatant dilutions, with adifference of at least 500 c.p.m. between the means of the triplicate setsof test and medium control wells.
+ VLP ELISA. HPV-16 L1-VLP-specific plasma IgG reactivity wasdetermined by using 250 ng per well HPV-16 L1-VLP and 1 :10-dilutedplasma samples in an ELISA test system, as previously described forL1}L2-containing VLP (de Gruijl et al., 1997). Duplicate tests wereperformed for all plasma samples.
IgG reactivities were expressed as the mean absorbance of theduplicate tests at 492 nm. To compensate for inter-assay variability,absorbance values were adjusted in relation to the absorbances of a set ofinternal control plasma samples (with both positive and negative IgGreactivities), which were included in each plate. The background reactivityfound in the negative control wells coated with PBS and blocked withPBS–Tween–newborn calf serum was subtracted from the correspondingabsorbances obtained for the test wells of each plasma sample. A cut-offlevel, above which absorbances were considered positive, was based onthe distribution of absorbances found in the HPV-negative CIN patientsincluded in this study. This cut-off value was obtained by adding 2 SD tothe mean absorbance (cut-off absorbance¯ 0±443).
+ Statistical analysis. Frequencies of positive responses between thetest groups were compared using 2¬2 table analysis and Fisher’s exacttest. Comparisons between sets of SI
HT#were carried out using the
Mann–Whitney U test. All tests were two-tailed and differences wereconsidered significant when P! 0±05.
ResultsIL-2 production in response to HPV-16 L1-VLP
Th responses were measured by the use of a sensitive IL-2bioassay, which was described previously (de Gruijl et al.,
EAB
T. D. de Gruijl and othersT. D. de Gruijl and others
12
9
6
3
0
SI H
T2
Undepleted
(a)
2× 4× 8×Culture supernatant dilution
12
9
6
3
0
12
9
6
3
0
CD4 depleted CD8 depleted
2× 4× 8× 2× 4× 8×
12
9
6
3
0
12
9
6
3
0
iso antiDR/DP/DQ
2× 4× 8× 2× 4× 8×Culture supernatant dilution
SI H
T2
(b)
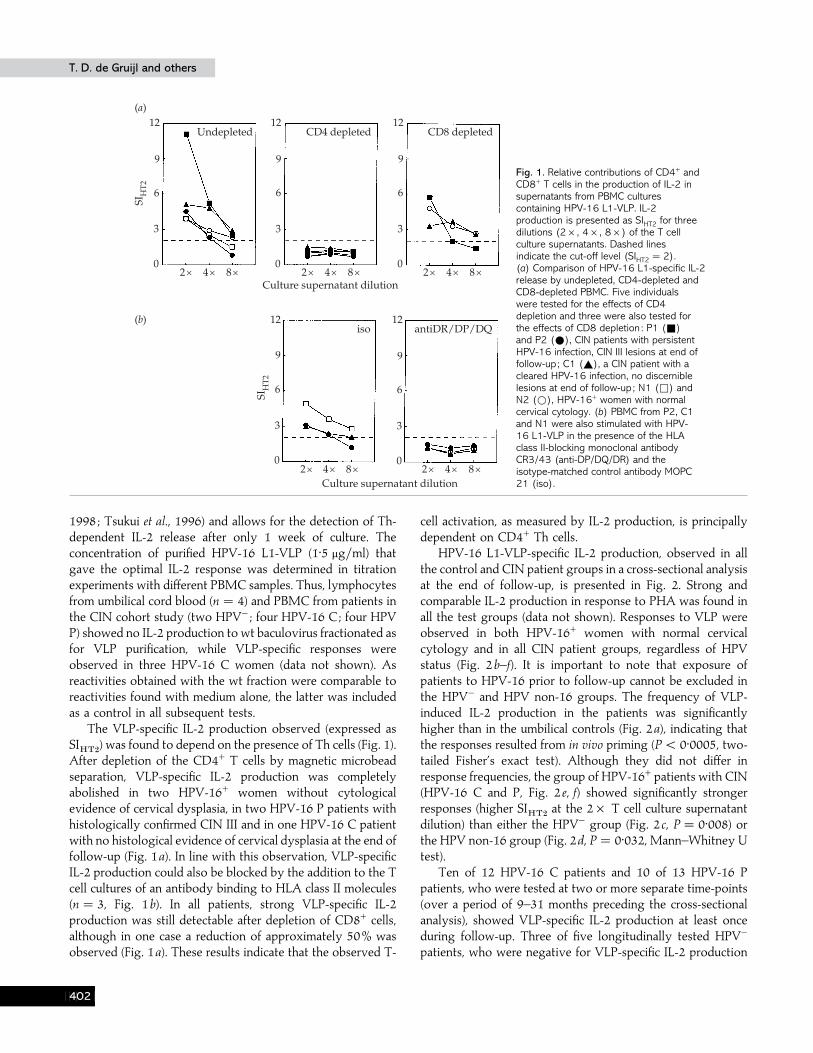
Fig. 1. Relative contributions of CD4+ andCD8+ T cells in the production of IL-2 insupernatants from PBMC culturescontaining HPV-16 L1-VLP. IL-2production is presented as SIHT2 for threedilutions (2¬, 4¬, 8¬) of the T cellculture supernatants. Dashed linesindicate the cut-off level (SIHT2 ¯2).(a) Comparison of HPV-16 L1-specific IL-2release by undepleted, CD4-depleted andCD8-depleted PBMC. Five individualswere tested for the effects of CD4depletion and three were also tested forthe effects of CD8 depletion : P1 (+)and P2 (E), CIN patients with persistentHPV-16 infection, CIN III lesions at end offollow-up; C1 (_), a CIN patient with acleared HPV-16 infection, no discerniblelesions at end of follow-up; N1 (*) andN2 (D), HPV-16+ women with normalcervical cytology. (b) PBMC from P2, C1and N1 were also stimulated with HPV-16 L1-VLP in the presence of the HLAclass II-blocking monoclonal antibodyCR3/43 (anti-DP/DQ/DR) and theisotype-matched control antibody MOPC21 (iso).
1998 ; Tsukui et al., 1996) and allows for the detection of Th-dependent IL-2 release after only 1 week of culture. Theconcentration of purified HPV-16 L1-VLP (1±5 µg}ml) thatgave the optimal IL-2 response was determined in titrationexperiments with different PBMC samples. Thus, lymphocytesfrom umbilical cord blood (n¯ 4) and PBMC from patients inthe CIN cohort study (two HPV− ; four HPV-16 C; four HPVP) showed no IL-2 production to wt baculovirus fractionated asfor VLP purification, while VLP-specific responses wereobserved in three HPV-16 C women (data not shown). Asreactivities obtained with the wt fraction were comparable toreactivities found with medium alone, the latter was includedas a control in all subsequent tests.
The VLP-specific IL-2 production observed (expressed asSI
HT#) was found to depend on the presence of Th cells (Fig. 1).
After depletion of the CD4+ T cells by magnetic microbeadseparation, VLP-specific IL-2 production was completelyabolished in two HPV-16+ women without cytologicalevidence of cervical dysplasia, in two HPV-16 P patients withhistologically confirmed CIN III and in one HPV-16 C patientwith no histological evidence of cervical dysplasia at the end offollow-up (Fig. 1a). In line with this observation, VLP-specificIL-2 production could also be blocked by the addition to the Tcell cultures of an antibody binding to HLA class II molecules(n¯ 3, Fig. 1b). In all patients, strong VLP-specific IL-2production was still detectable after depletion of CD8+ cells,although in one case a reduction of approximately 50% wasobserved (Fig. 1a). These results indicate that the observed T-
cell activation, as measured by IL-2 production, is principallydependent on CD4+ Th cells.
HPV-16 L1-VLP-specific IL-2 production, observed in allthe control and CIN patient groups in a cross-sectional analysisat the end of follow-up, is presented in Fig. 2. Strong andcomparable IL-2 production in response to PHA was found inall the test groups (data not shown). Responses to VLP wereobserved in both HPV-16+ women with normal cervicalcytology and in all CIN patient groups, regardless of HPVstatus (Fig. 2b–f). It is important to note that exposure ofpatients to HPV-16 prior to follow-up cannot be excluded inthe HPV− and HPV non-16 groups. The frequency of VLP-induced IL-2 production in the patients was significantlyhigher than in the umbilical controls (Fig. 2a), indicating thatthe responses resulted from in vivo priming (P! 0±0005, two-tailed Fisher’s exact test). Although they did not differ inresponse frequencies, the group of HPV-16+ patients with CIN(HPV-16 C and P, Fig. 2 e, f) showed significantly strongerresponses (higher SI
HT#at the 2¬ T cell culture supernatant
dilution) than either the HPV− group (Fig. 2 c, P¯ 0±008) orthe HPV non-16 group (Fig. 2d, P¯ 0±032, Mann–Whitney Utest).
Ten of 12 HPV-16 C patients and 10 of 13 HPV-16 Ppatients, who were tested at two or more separate time-points(over a period of 9–31 months preceding the cross-sectionalanalysis), showed VLP-specific IL-2 production at least onceduring follow-up. Three of five longitudinally tested HPV−
patients, who were negative for VLP-specific IL-2 production
EAC

Th and IgG responses to HPV-16 L1-VLPTh and IgG responses to HPV-16 L1-VLP
15
10
5
0
0/16 8/9 5/14
(a) UC controls (b) Normal, HPV-16+
(c) CIN, HPV–
SI H
T2
2× 4× 8× 2× 4× 8× 2× 4× 8×
15
10
5
0
5/13 8/15 12/21
(d) CIN, HPV non-16
(e) CIN, HPV-16 C
( f ) CIN, HPV-16 P
SI H
T2
2× 4× 8× 2× 4× 8× 2× 4× 8×Culture supernatant dilution
Culture supernatant dilution
Fig. 2. IL-2 responses to HPV-16 L1-VLP among patients and controls inthe cross-sectional analysis. IL-2 production is presented as SIHT2 for threedilutions (2¬, 4¬, 8¬) of the T cell culture supernatants. The dashedline indicates the cut-off level (SIHT2 ¯2). The proportion of patientsgiving a positive response is indicated for each of the test groups.(a) Umbilical cord PBMC controls, n¯16; (b) HPV-16+ women withnormal cervical cytology, n¯9; (c) HPV− patients from the CIN cohort,n¯14; (d) HPV non-16 patients from the CIN cohort, n¯13; (e)HPV-16 C patients from the CIN cohort, n¯15; (f) HPV-16 P patientsfrom the CIN cohort, n¯21.
in the cross-sectional analysis, showed positive IL-2 responsesat earlier time-points, possibly indicating the presence of HPVinfections prior to follow-up. IL-2 follow-up data from thepositively responding patients with either HPV-16 clearanceor persistence did not reveal consistently increasing ordecreasing IL-2 production over time in either of the twogroups, and no consistent trends arose in relation to cervicaldisease development. This is illustrated in Fig. 3 : increasingand decreasing VLP-specific IL-2 production was observedduring follow-up, both in HPV-16 C patients with resolvedCIN lesions (Fig. 3a, b) and in HPV-16 P patients with high-grade CIN III lesions (Fig. 3 c, d).
IgG responses to HPV-16 L1-VLP in relation to IL-2reactivity
Plasma samples were available to determine L1-VLP-specific IgG reactivity of 54 of 63 CIN patients tested and all
HPV-16+ women with normal cervical cytology. IgGresponses in the same group of patients against VLP containingboth L1 and L2 were reported previously (de Gruijl et al.,1997). For comparison with Th cell activity, IgG responses inthis study were measured against the same VLP preparation(containing only L1) as was used in the IL-2 release experi-ments. A good correlation was found between these IgGreactivities and IgG reactivities against L1}L2-VLP (de Gruijl etal., 1997) previously determined in the same plasma samples(data not shown: linear regression analysis ; r¯ 0±782, P!0±0001, two-tailed). By combining the cross-sectional andlongitudinal analyses, L1-VLP IgG and IL-2 reactivity datawere available from one to four time-points, depending on thepatient. A patient was considered to respond positively whenshe showed a positive response at least once during the follow-up. The resulting IgG response frequencies in relation to IL-2response frequencies from the same patients are shown inTable 2.
In contrast to the Th responses, IgG responses to L1-VLPwere restricted to patients known to have been exposed toHPV-16 and were significantly more frequent among patientswith persistent infections than in patients with cleared HPV-16infections (Table 2).
When the HPV-16+ patients from the CIN cohort studywere stratified according to disease status at the end of follow-up, no clear associations were found with Th IL-2 responserates (eight of 13 patients with CIN III compared to 12 of 23patients with CIN I}II or resolved lesions), whereas positiveIgG responses to L1-VLP were significantly more frequentamong HPV-16+ women with CIN III (seven of 11) thanamong HPV-16+ women with lower grade or resolved lesionsat the end of follow-up (two of 17 ; P¯ 0±01, two-tailedFisher’s exact test).
Of the 63 women tested for both IL-2 production and IgGreactivity in response to L1-VLP, 28 showed only IL-2production, 13 showed both IL-2 production and IgG reactivityand two showed only IgG reactivity (Table 2).
Th IL-2 responses to the L1-derived peptides P1 andP2: relation with IL-2 and IgG response to L1-VLP andHLA-DR/-DQ genotype
Whereas HPV-16 L1-VLP-specific IgG responses showedtype specificity, as indicated by an association with knownexposure to HPV-16, Th responses did not. The relevantspecificity of the Th response may be restricted to a particularepitope. This was further investigated by using peptidescovering L1 amino acids 311–335, which have previously beenshown to elicit an HPV-16-specific proliferative response inpatients with CIN (Shepherd et al., 1997). The overall responserates to the L1-derived 15-mer peptides P1 (amino acids311–325) and P2 (amino acids 321–335) were lower than thosefound for L1-VLP (Table 3). The responses appeared tocorrelate with HPV exposure but it could not be concludedthat they were HPV-16-specific (16 of 40 HPV+ versus one of
EAD

T. D. de Gruijl and othersT. D. de Gruijl and others
SI H
T2
15
10
5
02× 4× 8× 2× 4× 8× 2× 4× 8×
Culture supernatant dilution
52 months 58 months 63 months
(b)S
I HT
2
30
20
10
02× 4× 8× 2× 4× 8× 2× 4× 8×
Culture supernatant dilution
37 months 48 months 53 months
HPV-16 C, CIN 0(a)
SI H
T2
30
20
10
02× 4× 8× 2× 4× 8× 2× 4× 8×
Culture supernatant dilution
22 months 35 months 43 months
HPV-16 P, CIN III(c)
SI H
T2
15
10
5
02× 4× 8× 2× 4× 8× 2× 4× 8×
Culture supernatant dilution
34 months 42 months 51 months
(d )
Fig. 3. Follow-up of Th cell-dependent IL-2 production in four CIN patients. IL-2 production is presented as SIHT2 for threedilutions (2¬, 4¬, 8¬) of the T cell culture supernatants. The dashed line indicates the cut-off level (SIHT2 ¯2). The time oftesting is indicated in months since the start of clinical follow-up; the last indicated time-point is at the end of follow-up (i.e.when biopsies were taken). (a)–(b) CIN patients with a cleared HPV-16 infection and resolved lesions at the end of follow-up.IL-2 follow-up started 5 months (a) and 20 months (b) after HPV-16 DNA was last detected. (c)–(d) CIN patients with apersistent HPV-16 infection and CIN III lesions at the end of follow-up.
Table 2. IL-2 and IgG response frequencies to HPV-16 L1-VLP among patients andcontrols : combined results from the cross-sectional and longitudinal analyses
The number of patients from different HPV-status groups (as described in Methods) giving IL-2 and IgGresponses to VLP is shown as no. positive}no. tested, with the percentage of each group shown inparentheses.
Disease state HPV status IL-2+ (%) IgG+ (%) IL-2+ and IgG+ (%)
Normal HPV-16+ 8}9 (88±9) 5}9 (55±6) 4}9 (44±4)CIN HPV− 8}13 (61±5) 1}13 (7±7) 1}13 (7±7)
HPV-16 PC 20}28 (71±4) 9}28 (32±1)* 8}28 (28±6)HPV-16 P 12}16 (75±0) 8}16 (50±0)† 7}16 (43±8)HPV-16 C 8}12 (66±7) 1}12 (8±3) 1}12 (8±3)HPV non-16 5}13 (38±5) 0}13 (0) 0}13 (0)
* HPV-16 PC versus HPV non-16, P¯ 0±038, two-tailed Fisher’s exact test.† HPV-16 P versus HPV-16 C, P¯ 0±039, two-tailed Fisher’s exact test.
12 HPV−, P¯ 0±076; 11 of 29 HPV-16+ versus five of 11 HPVnon-16+, P¯ 0±733; two-tailed Fisher’s exact tests).
Of the 19 subjects with positive responses to amino acids311–335, 15 also showed IL-2 responsiveness to L1-VLP.
There was no absolute association between IL-2 respon-siveness to P1 or P2 and L1-VLP-specific IgG reactivity, but allIgG-positive patients also showed IL-2 responses either to L1-VLP or to P1 or P2.
EAE

Th and IgG responses to HPV-16 L1-VLPTh and IgG responses to HPV-16 L1-VLP
Table 3. IL-2 response frequencies to the HPV-16 L1-derived peptides P1 and P2among patients and controls
The number of patients in each group showing an IL-2 response to peptides P1 and}or P2 is shown as no.positive}no. tested, with the percentage within the group shown in parentheses.
Disease state HPV status P1 (%) P2 (%) P1 or P2 (%)
Normal HPV-16+ 1}7 (14±3) 1}7 (14±3) 1}7 (14±3)CIN HPV− 0}12 (0) 1}12 (8±3) 1}12 (8±3)
HPV-16 PC 7}29 (24±1) 8}29 (27±6) 11}29 (37±9)*HPV-16 P 4}18 (22±2) 6}18 (33±3) 7}18 (38±9)HPV-16 C 3}11 (27±3) 2}11 (18±2) 4}11 (36±4)HPV non-16 4}11 (36±4) 3}11 (27±3) 5}11 (45±5)
UC controls – 0}7 (0) 0}7 (0) 0}7 (0)
* HPV-16 PC versus HPV−, P¯ 0±073, two-tailed Fisher’s exact test.
Of the 36 HPV-16+ CIN patients included in this study, 35were tissue-typed for their HLA DR and DQ alleles (HPV-16C, n¯ 15 ; HPV-16 P, n¯ 20). In these patients, recognitionof the immunogenic region covering amino acids 311–335 wasfound to be associated with the HLA DRB1*11}DQB1*0301haplotype (P¯ 0±05). This association appears to depend onthe DRB1*11 allele, as eight of 11 patients that responded toP1}P2 carried this allele as compared to four of 17 that did notrespond (P¯ 0±019), while no significant difference in fre-quency was found for the DQB1*0301 allele between thesetwo groups.
Further examination of the Th IL-2 and IgG responsivenessin the HPV-16+ patient group in relation to the patient tissuetype revealed an association between the HLA DRB1*0101}DQB1*0501 haplotype and the presence of IgG responses toHPV-16 L1-VLP (six of nine giving an IgG response versusone of 19 that did not respond, P¯ 0±0014). Again, thisassociation was found to depend more on the DR allele (P¯0±0014) than on the DQ allele, which showed a slightly weakerassociation (P¯ 0±0068).
DiscussionThis study has investigated whether Th and}or IgG
responses to HPV-16 L1-VLP are correlated with clearance ofthe virus and associated CIN lesions. Differential patterns ofTh and IgG responsiveness were observed in terms of HPVinfection and CIN development.
Th-dependent, HPV-16 L1-VLP-specific IL-2 responseswere significantly stronger in patients with abnormal cervicalcytology and known HPV-16 infections compared with HPV−
patients or HPV non-16 patients. This, together with the lackof responses in umbilical cord lymphocytes, is consistent withthe detection of memory responses. Responses detected inHPV− and HPV non-16 patients may be due to exposure toHPV-16 prior to follow-up or by cross-reactivity with other
HPV types. The possibility of prior HPV exposure in HPV−
patients was supported by IL-2 responses observed early onduring follow-up, but found to decline over time, in three outof five longitudinally tested HPV− patients.
Th IL-2 responses did not correlate with clinical outcome,defined by either virus clearance or persistence or CIN grade.In contrast, IgG responses to L1-VLP were associated withpersistent HPV-16 infection and the development of high-grade CIN. Thirteen of 15 patients producing anti-VLP IgGalso showed Th IL-2 release. This is consistent with Th activitypreceding and required for the generation of L1-specific IgGresponses.
Using the data from this study and other previouslyreported results, we can now attempt to reconstruct thetemporal relationship between HPV-16 infection, the gen-eration of immune responses to the viral capsid proteins andthe development of cervical neoplasia.
A high frequency of Th cell-dependent IL-2 responses to L1was found among women with HPV-16 infections but normalcervical cytology (eight of nine or 88±9%). This is consistentwith L1-specific T-cell responses occurring upon virus ex-posure and preceding the development of any lesions. Inaddition, five of these nine women also showed positive IgGresponses. However, as these were prevalent HPV-16 in-fections, there was no way of knowing how long these womenhad been infected by HPV-16 or within what time-period thesubsequent T-cell activation and seroconversion occurred.Other longitudinal studies have shown that upon primaryinfection, HPV-16 VLP IgG seroconversion can occur within6–8 months, preceding any overt lesions (Wikstro$ m et al.,1995 ; Carter et al., 1996). As the generation of IgG responsesdepends on Th activity, it is to be expected that some womenwill show Th responses without IgG seropositivity.
Two previous studies showed that cytologically normalwomen who harboured persistent HPV-16 infections and whoseroconverted were more likely to develop dysplastic cervical
EAF

T. D. de Gruijl and othersT. D. de Gruijl and others
lesions (Carter et al., 1996 ; Chua et al., 1996). Thus, serum IgGantibodies to L1 do not necessarily contribute to the clearanceof HPV at an early stage of infection. Indeed, seropositivity toVLP is predictive for the development of CIN III and cervicalcarcinoma (Nonnenmacher et al., 1995 ; Lehtinen et al., 1996 ; deGruijl et al., 1997).
In the longitudinal analysis, there were no consistent trendsin Th IL-2 activity among patients with either virus clearanceor persistence. This contrasts with IL-2 responses to the HPV-16 E7 oncoprotein, which show a significant decline after HPVclearance but increase in patients with persistent HPVinfections and CIN III lesions (de Gruijl et al., 1998). Transientexposure to HPV virions may be sufficient to trigger memoryTh cells specifically reactive to capsid proteins, influencing thedetection of IL-2 release during follow-up but independent ofovert disease or HPV DNA positivity.
The longitudinal analysis revealed VLP-specific IL-2responses in a majority of patients with CIN, independent ofHPV status and disease course. These results suggest that theT-cell activity detected does not necessarily influence diseaseoutcome or virus clearance.
In addition, we have studied HPV-16 specificity in Thresponsiveness in relation to IgG production. To this end, IL-2 release induced by peptides P1 and P2 covering L1 aminoacids 311–335 was examined. This response showed con-siderable overlap with responses to L1-VLP but no significantcorrelation with IgG production, consistent with a recentreport (Luxton et al., 1997). A strong association among HPV-16+ patients between IL-2 responsiveness to amino acids311–335 and the HLA DRB1*11 allele suggests that thisepitope may be specifically recognized by DR11-restricted Thcells. However, responses to L1 amino acids 311–335 wereshown to be more frequent in HPV+ patients with CIN than inHPV− patients with resolved or low-grade lesions or in HPV-16+ women with normal cervical cytology, suggesting thatrecognition of this region does not protect from the de-velopment of premalignant cervical lesions. Indeed, the highestresponse rates reported by Shepherd et al. (1997) were inpatients with CIN III.
The IgG response to HPV-16 L1-VLP showed a strongassociation with the DRB1*0101}DQB1*0501 haplotype.Although the small number of subjects included in this studyand the high polymorphism of the HLA locus (for which theprobabilities listed in the text were not corrected) necessitatescaution, it is an interesting observation that particular HLAclass II alleles may be involved in the generation of IgGresponses. This is in line with recent studies showing thatdifferent peptide}MHC II combinations can give rise toqualitatively different Th responses (Th1 or Th2), in turnleading to the preferential generation of either cell-mediated orhumoral immune responses (Chaturvedi et al., 1995).
It is apparent that neither the Th IL-2 nor the IgG responsesmeasured in this study are sufficient to deliver natural controlof HPV infection and CIN development. This may be due to
the expression of HPV capsid proteins in the top layers of thesquamous epithelium of the cervix. This relatively remotelocalization may hamper transport of L1 by migratingprofessional antigen-presenting cells (APC) to draining lymphnodes, which has been proposed as a crucial step in thegeneration of virus-specific immune responses (Zinkernagel,1996). This in turn would account for the observed lagbetween HPV-16 infection and the generation of VLP-specificantibody responses at generally low serum titres (Carter et al.,1996). However, prophylactic vaccination with VLP mayinduce immediate and vigorous immune responses and highserum titres of neutralizing IgG antibodies. Such responseshave proved to be protective against subsequent experimentalchallenge with papillomaviruses and to prevent the devel-opment of papillomavirus-induced lesions in various animalmodels (Breitburd et al., 1995 ; Suzich et al., 1995 ; Kirnbauer etal., 1996).
In terms of natural immunity against HPV capsids, localIgA responses in the cervix, with the capacity to neutralizevirions at the actual site of infection, may be more relevantthan peripherally recirculating T cells and serum IgG levels(Wang et al., 1996). Therefore, in the following paper, we havedetermined IgA reactivity to HPV-16 VLP in cervical mucussamples of women with CIN, to relate these responses to HPVand disease status over time (Bontkes et al., 1999). Identificationof the most effective type of immunity will also haveimplications for immune intervention protocols, as the methodof vaccine administration and routing will depend on the typeof immune response to be targeted.
Dr Peter L. Stern was Visiting Professor of the Pasman Chair forInternationalization of the Oncology Research Institute of the VrijeUniversteit, Amsterdam. This study was supported by grants from thePrevention Fund (28-1502.2) and the University Stimulation Fund (USF)of the Free University (T.D.G., H. J.B., J.M.M.W., M. J.S., R.H.M.V.,Th. J.M.H., C. J.L.M.M. and R. J.S.) and received financial support fromthe Cancer Research Campaign (M.F.D.-K. and P.L.S.). We thank AdamStevens, Paul Sinnott and Philip Dyer for HLA typing of the CIN patients(North West Regional Tissue Typing Laboratory, St Mary’s Hospital,Manchester, UK), Adrie van den Muysenberg, Rene! Pol and all ourcolleagues at the Laboratory of Clinical Immunology of the FreeUniversity Hospital in Amsterdam for excellent technical assistance, andAns Remmink, Rence Rozendaal and Elle Risse for assisting in theevaluation of the clinical data.
ReferencesBontkes, H. J., de Gruijl, T. D., Walboomers, J. M. M., Schiller, J. T.,Dillner, J., Helmerhorst, T. J. M., Verheijen, R. H. M., Scheper, R. J. &Meijer, C. J. L. M. (1999). Immune responses against human papillo-mavirus (HPV) type 16 virus-like particles in a cohort study of womenwith cervical intraepithelial neoplasia. II. Systemic but not local IgAresponses correlate with clearance of HPV-16. Journal of General Virology80, 409–417.
Borysiewicz, L. K., Fiander, A., Nimako, M., Man, S., Wilkinson, G. W.G., Westmoreland, D., Evans, A. S., Adams, M., Stacey, S. N., Boursnell,M. E. G., Rutherford, E., Hickling, J. K. & Inglis, S. C. (1996). A
EAG

Th and IgG responses to HPV-16 L1-VLPTh and IgG responses to HPV-16 L1-VLP
recombinant vaccinia virus encoding human papillomavirus types 16 and18, E6 and E7 proteins as immunotherapy for cervical cancer. Lancet 347,1523–1527.
Breitburd, F., Kirnbauer, R., Hubbert, N. L., Nonnenmacher, B., Trin-Dinh-Desmarquet, C., Orth, C., Schiller, J. T. & Lowy, D. R. (1995).Immunization with viruslike particles from cottontail rabbit papilloma-virus (CRPV) can protect against experimental CRPV infection. Journal ofVirology 69, 3959–3963.
Carter, J. J., Koutsky, L. A., Wipf, G. C., Christensen, N. D., Lee, S.-K.,Kuypers, J., Kiviat, N. & Galloway, D. A. (1996). The natural history ofhuman papillomavirus type 16 capsid antibodies among a cohort ofuniversity women. Journal of Infectious Diseases 174, 927–936.
Chaturvedi, P., Yu, Q., Southwood, S., Sette, A. & Singh, B. (1995).Peptide analogs with different affinities for MHC alter the cytokineprofile of helper T cells. International Immunology 8, 745–755.
Chua, K. L., Wiklund, L., Lenner, P., Angstrom, T., Hallmans, G.,Bergman, F., Sapp, M., Schiller, J., Wadell, G., Hjerpe, A. & Dillner, J.(1996). A prospective study on the risk of cervical intra-epithelialneoplasia among healthy subjects with serum antibodies to HPVcompared with HPV DNA in cervical smears. International Journal ofCancer 68, 54–59.
de Gruijl, T. D., Bontkes, H. J., Stukart, M. J., Walboomers, J. M. M.,Remmink, A. J., Verheijen, R. H. M., Helmerhorst, T. J. M., Meijer, C. J.L. M. & Scheper, R. J. (1996). T cell proliferative responses againsthuman papillomavirus type 16 E7 oncoprotein are most prominent incervical intraepithelial neoplasia patients with a persistent viral infection.Journal of General Virology 77, 2183–2191.
de Gruijl, T. D., Bontkes, H. J., Walboomers, J. M. M., Schiller, J. T.,Stukart, M. J., Groot, B. S., Chabaud, M. M. R., Remmink, A. J.,Verheijen, R. H. M., Helmerhorst, Th. J. M., Meijer, C. J. L. M. &Scheper, R. J. (1997). Immunoglobulin G responses against humanpapillomavirus type 16 virus-like particles in a prospective nonin-tervention cohort study of women with cervical intraepithelial neoplasia.Journal of the National Cancer Institute 89, 630–638.
de Gruijl, T. D., Bontkes, H. J., Walboomers, J. M. M., Stukart, M. J.,Doekhie, F. S., Remmink, A. J., Helmerhorst, Th. J. M., Verheijen,R. H. M., Duggan-Keen, M. F., Stern, P. L., Meijer, C. J. L. M. &Scheper, R. J. (1998). Differential T helper cell responses to humanpapillomavirus type 16 E7 related to viral clearance or persistence inpatients with cervical neoplasia : a longitudinal study. Cancer Research 58,1700–1706.
de Roda Husman, A. M., Walboomers, J. M. M., Meijer, C. J. L. M.,Risse, E. K. J., Schipper, M. E. I., Helmerhorst, T. J. M., Bleker, O. P.,Delius, H., van den Brule, A. J. C. & Snijders, P. J. F. (1994). Analysisof cytomorphologically abnormal cervical scrapes for the presence of 27mucosotropic human papillomavirus genotypes, using polymerase chainreaction. International Journal of Cancer 56, 802–806.
de Roda Husman, A.-M., Walboomers, J. M. M., van den Brule, A. J. C.,Meijer, C. J. L. M. & Snijders, P. J. F. (1995). The use of general primersGP5 and GP6 elongated at their 3’ ends with adjacent highly conservedsequences improves human papillomavirus detection by PCR. Journal ofGeneral Virology 76, 1057–1062.
Duggan-Keen, M. F., Keating, P. J., Stevens, F. R. A., Sinnott, P.,Snijders, P. J. F., Walboomers, J. M. M., Davidson, S., Hunter, R. D.,Dyer, P. A. & Stern, P. L. (1996). Immunogenetic factors in HPV-associated cervical cancer : influence on disease progression. EuropeanJournal of Immunogenetics 23, 275–284.
Dupuy, C., Buzoni-Gatel, D., Touze, A., Le Cann, P., Bout, D. &Coursaget, P. (1997). Cell mediated immunity induced in mice by HPV-16 L1 virus-like particles. Microbial Pathogenesis 22, 219–225.
Ertl, H. C. J., Dietzschold, B., Gore, M., Otvos, L., Larson, J. K.,
Wunner, W. H. & Koprowski, H. (1989). Induction of rabies virus-specific T-helper cells by synthetic peptides that carry dominant T-helpercell epitopes of the viral ribonucleoprotein. Journal of Virology 63,2885–2892.
Frazer, I. H. (1996). Immunology of papillomavirus infection. CurrentOpinions in Immunology 8, 484–491.
Ho$ pfl, R., Sandbichler, M., Sepp, N., Heim, K., Mu$ ller-Holzner, E.,Wartusch, B., Dapunt, O., Jochmus-Kudielka, I., ter Meulen, J.,Gissmann, L. & Fritsch, P. (1991). Skin test for HPV type 16 proteinsin cervical intraepithelial neoplasia. Lancet i, 373–374.
IARC (1995). Monographs on the Evaluation of the Carcinogenic Risks toHumans, vol. 64, The Human Papillomavirus, Lyon : International Agencyfor Research on Cancer.
Jacobs, M. V., de Roda Husman, A. M., van den Brule, A. J. C., Snijders,P. J. F., Meijer, C. J. L. M. & Walboomers, J. M. M. (1995). Group-specific differentiation between high- and low-risk human papillomavirusgenotypes by using the general primer-mediated PCR and two cocktailsof oligonucleotide probes. Journal of Clinical Microbiology 33, 901–905.
Kirnbauer, R., Taub, J., Greenstone, H., Roden, R., Du$ rst, M., Gissmann,L., Lowy, D. R. & Schiller, J. T. (1993). Efficient self-assembly of humanpapillomavirus type 16 L1 and L1–L2 into virus-like particles. Journal ofVirology 67, 6929–6936.
Kirnbauer, R., Chandrachud, L. M., O’Neil, B. W., Wagner, E. R.,Grindlay, G. J., Armstrong, A., McGarvie, G. M., Schiller, J. T., Lowy,D. R. & Campo, M. S. (1996). Virus-like particles of bovine papilloma-virus type 4 in prophylactic and therapeutic immunization. Virology 219,37–44.
Le Cann, P., Coursaget, P., Iochmann, S. & Touze, A. (1994). Self-assembly of human papillomavirus type 16 capsids by expression of theL1 protein in insect cells. FEMS Microbiology Letters 117, 269–274.
Lehtinen, M., Dillner, J., Knekt, P., Luostarinen, T., Aromaa, A.,Kirnbauer, R., Koskela, P., Paavonen, J., Peto, R., Schiller, J. T. &Hakama, M. (1996). Serologically diagnosed infection with humanpapillomavirus type 16 and risk for subsequent development of cervicalcarcinoma : nested case-control study. British Medical Journal 312,537–539.
Luxton, J. C., Rose, R. C., Coletart, T., Wilson, P. & Shepherd, P. S.(1997). Serological and T-helper cell responses to human papillomavirustype 16 L1 in women with cervical dysplasia or cervical carcinoma and inhealthy controls. Journal of General Virology 78, 917–923.
Nonnenmacher, B., Hubbert, N. L., Kirnbauer, R., Shah, K. V., Munoz,N., Bosch, F. X., Desanjose, S., Viscidi, R., Lowy, D. R. & Schiller, J. T.(1995). Serologic response to human papillomavirus type 16 (HPV-16)virus-like particles in HPV-16 DNA-positive invasive cervical cancer andcervical intraepithelial neoplasia grade III patients and controls fromColombia and Spain. Journal of Infectious Diseases 172, 19–24.
Remmink, A. J., Walboomers, J. M. M., Helmerhorst, T. J. M.,Voorhorst, F. J., Rozendaal, L., Risse, E. K. J., Meijer, C. J. L. M. &Kenemans, P. (1995). The presence of persistent high-risk HPVgenotypes in dysplastic cervical lesions is associated with progressivedisease : natural history up to 36 months. International Journal of Cancer61, 306–311.
Schiller, J. T. & Roden, R. B. S. (1995). Papillomavirus-like particles.Papillomavirus Report 6, 121–128.
Shepherd, P. S., Rowe, A. J., Cridland, J. C., Coletart, T., Wilson, P. &Luxton, J. C. (1997). Proliferative T cell responses to human papilloma-virus type 16 L1 peptides in patients with cervical dysplasia. Journal ofGeneral Virology 77, 593–602.
Suzich, J. A., Ghim, S.-J., Palmer-Hill, F. J., White, W. I., Tamura, J. K.,Bell, J. A., Newsome, J. A., Jenson, A. B. & Schlegel, R. (1995).
EAH

T. D. de Gruijl and othersT. D. de Gruijl and others
Systemic immunization with papillomavirus L1 protein completelyprevents the development of viral mucosal papillomas. Proceedings of theNational Academy of Sciences, USA 92, 11553–11557.
Tindle, R. W. (1996). Human papillomavirus vaccines for cervical cancer.Current Opinions in Immunology 8, 643–650.
Tsukui, T., Hildesheim, A., Schiffman, M. H., Luci, J., III, Contois, D.,Lawler, P., Rush, B. B., Lorincz, A. T., Corrigan, A., Burk, R. D., Qu, W.,Marshall, M. A., Mann, D., Carrington, M., Clerici, M., Shearer, G. M.,Carbone, D. P., Scott, D. R., Houghten, R. A. & Berzofsky, J. A. (1996).Interleukin 2 production in vitro by peripheral lymphocytes in responseto human papillomavirus-derived peptides : correlation with cervicalpathology. Cancer Research 56, 3967–3974.
Wang, Z., Hansson, B. G., Forslund, O., Dillner, L., Sapp, M., Schiller,J. T., Bjerre, B. & Dillner, J. (1996). Cervical mucus antibodies againsthuman papillomavirus type 16, 18, and 33 capsids in relation to presenceof viral DNA. Journal of Clinical Microbiology 34, 3056–3062.
Wikstro$ m, A., van Doornum, G. J. J., Quint, W. G. V., Schiller, J. T. &Dillner, J. (1995). Identification of human papillomavirus sero-conversions. Journal of General Virology 76, 529–539.
Zinkernagel, R. M. (1996). Immunology taught by viruses. Science 271,173–178.
Received 23 September 1998; Accepted 30 September 1998
EAI




















