
Identification of the protein Zibra, its genomic organization, regulation, and expression in breast...
12
Identification of the protein Zibra, its genomic organization, regulation, and expression in breast cancer cells H. Garrett R. Thompson, Joseph W. Harris, Limin Lin, and James P. Brody * Department of Biomedical Engineering, University of California-Irvine, Irvine, CA 92697-2715, USA Received 29 October 2003, revised version received 14 January 2004 Abstract The mRNA that encodes zibra (zinc, in-between-ring finger, ubiquitin-associated domain), previously known as hypothetical protein FLJ10111, or RNF31 is expressed in several distinct cancers. Little is known about the genomic organization, expression, or regulation of zibra. Using RNA ligase-mediated rapid amplification of cDNA ends (RLM-RACE), we cloned the full-length zibra cDNA from a transformed breast cell line. We identified a novel exon, the 5V untranslated region including the +1 start site, and three alternatively spliced zibra transcripts. The zibra protein contains three zinc ring-finger motifs, an ubiquitin-associated domain, and an in-between-ring-finger domain, characteristic of ubiquitin ligases. We obtained an antibody to zibra and confirmed the presence of translated zibra protein for the first time. Promoter studies localized a core element responsible for basal activity to a 14-bp region in the 5V untranslated region. Although there are numerous consensus Ets factor binding sites in the zibra promoter, we found no affect on promoter activity from Ets-1, PDEF, or PEA-3/E1A-F. Treatment of cells with the proteasome inhibitor I (PSI) decreased zibra protein to an undetectable level after 8 h. Zibra remained undetectable even after 32 h, while mRNA levels remained essentially unchanged. In conclusion, zibra is a translationally regulated putative ubiquitin ligase that is frequently overexpressed in different forms of cancer. D 2004 Elsevier Inc. All rights reserved. Keywords: Breast cancer; Cell lines; E3; FLJ10111; Gene regulation; Hypothetical protein; Proteasome; PSI; RNF31; Zibra Introduction Several unifying principles are shared by most, if not all, cancers. Genetic changes lead to abnormal regulation of physiological processes, which in turn lead to uncon- trolled proliferation of the cancer, bypassing mechanisms that govern normal cells. These abnormally regulated physiological processes have been described as the hall- marks of cancer. They include evasion of apoptosis, angiogenesis, tissue invasion and metastasis, unlimited replication, and independence from both positive and negative growth signaling [1]. One strategy for identifying the molecular basis for these hallmarks of cancer is to search for common molecular signatures among dissimilar cancers. We identified a module of genes that are expressed in several distinct types of cancers using publicly available large-scale gene expression data [2]. Zibra, previously known as the hypothetical protein FLJ10111, or Ring-finger protein 31 (RNF31) is one member of this module. Other members include several breast cancer and proteasome- related gene products [2]: PSMD7, a 19S proteasome lid non-ATPase subunit; SQSTM1, encoding the ubiquitin- binding protein p62 overexpressed in breast tumors [3]; and PDEF, an Ets family transcription factor whose mRNA is overexpressed in breast cancer [2–4]. Zibra was first identified by the Nedo Full-length cDNA Sequencing Project in Japan. Although several similar mes- sages for the gene have been found, most compiled from individual expressed sequence tags (ESTs) [5,6], no protein product has yet been identified. Transcripts of zibra mRNA are increased in some breast cancer cell lines compared to untransformed cells [2] (this report) and represent a high percentage of transcripts in various cancer tissues, including oligodendroglioma, and bronchioalveolar, papillary serous, borderline ovarian, and breast carcinomas based on EST and Unigene normalized expression data (http://genome-www3. Stanford.edu/cgi-bin/SMD/source/sourceResult?option= Number&criteria=NM _ 017999&choice=Gene). 0014-4827/$ - see front matter D 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.yexcr.2004.01.019 * Corresponding author. Department of Biomedical Engineering, University of California-Irvine, 204 Rockwell Engineering Center, Irvine, CA 92697-2715. Fax: +1-949-824-9968. E-mail address: [email protected] (J.P. Brody). www.elsevier.com/locate/yexcr Experimental Cell Research 295 (2004) 448 – 459
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Identification of the protein Zibra, its genomic organization, regulation, and expression in breast...

www.elsevier.com/locate/yexcr
Experimental Cell Research 295 (2004) 448–459
Identification of the protein Zibra, its genomic organization,
regulation, and expression in breast cancer cells
H. Garrett R. Thompson, Joseph W. Harris, Limin Lin, and James P. Brody*
Department of Biomedical Engineering, University of California-Irvine, Irvine, CA 92697-2715, USA
Received 29 October 2003, revised version received 14 January 2004
Abstract
The mRNA that encodes zibra (zinc, in-between-ring finger, ubiquitin-associated domain), previously known as hypothetical protein
FLJ10111, or RNF31 is expressed in several distinct cancers. Little is known about the genomic organization, expression, or regulation of
zibra. Using RNA ligase-mediated rapid amplification of cDNA ends (RLM-RACE), we cloned the full-length zibra cDNA from a
transformed breast cell line. We identified a novel exon, the 5V untranslated region including the +1 start site, and three alternatively spliced
zibra transcripts. The zibra protein contains three zinc ring-finger motifs, an ubiquitin-associated domain, and an in-between-ring-finger
domain, characteristic of ubiquitin ligases. We obtained an antibody to zibra and confirmed the presence of translated zibra protein for the
first time. Promoter studies localized a core element responsible for basal activity to a 14-bp region in the 5V untranslated region. Although
there are numerous consensus Ets factor binding sites in the zibra promoter, we found no affect on promoter activity from Ets-1, PDEF, or
PEA-3/E1A-F. Treatment of cells with the proteasome inhibitor I (PSI) decreased zibra protein to an undetectable level after 8 h. Zibra
remained undetectable even after 32 h, while mRNA levels remained essentially unchanged. In conclusion, zibra is a translationally regulated
putative ubiquitin ligase that is frequently overexpressed in different forms of cancer.
D 2004 Elsevier Inc. All rights reserved.
Keywords: Breast cancer; Cell lines; E3; FLJ10111; Gene regulation; Hypothetical protein; Proteasome; PSI; RNF31; Zibra
Introduction
Several unifying principles are shared by most, if not
all, cancers. Genetic changes lead to abnormal regulation
of physiological processes, which in turn lead to uncon-
trolled proliferation of the cancer, bypassing mechanisms
that govern normal cells. These abnormally regulated
physiological processes have been described as the hall-
marks of cancer. They include evasion of apoptosis,
angiogenesis, tissue invasion and metastasis, unlimited
replication, and independence from both positive and
negative growth signaling [1]. One strategy for identifying
the molecular basis for these hallmarks of cancer is to
search for common molecular signatures among dissimilar
cancers.
We identified a module of genes that are expressed in
several distinct types of cancers using publicly available
0014-4827/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.yexcr.2004.01.019
* Corresponding author. Department of Biomedical Engineering,
University of California-Irvine, 204 Rockwell Engineering Center, Irvine,
CA 92697-2715. Fax: +1-949-824-9968.
E-mail address: [email protected] (J.P. Brody).
large-scale gene expression data [2]. Zibra, previously
known as the hypothetical protein FLJ10111, or Ring-finger
protein 31 (RNF31) is one member of this module. Other
members include several breast cancer and proteasome-
related gene products [2]: PSMD7, a 19S proteasome lid
non-ATPase subunit; SQSTM1, encoding the ubiquitin-
binding protein p62 overexpressed in breast tumors [3];
and PDEF, an Ets family transcription factor whose mRNA
is overexpressed in breast cancer [2–4].
Zibra was first identified by the Nedo Full-length cDNA
Sequencing Project in Japan. Although several similar mes-
sages for the gene have been found, most compiled from
individual expressed sequence tags (ESTs) [5,6], no protein
product has yet been identified. Transcripts of zibra mRNA
are increased in some breast cancer cell lines compared to
untransformed cells [2] (this report) and represent a high
percentage of transcripts in various cancer tissues, including
oligodendroglioma, and bronchioalveolar, papillary serous,
borderline ovarian, and breast carcinomas based on EST and
Unigene normalized expression data (http://genome-www3.
Stanford.edu/cgi-bin/SMD/source/sourceResult?option=
Number&criteria=NM_017999&choice=Gene).

H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459 449
Several lines of evidence hint that the zibra protein is
involved in the ubiquitin–proteasome pathway. Zibra is
located in a tightly packed region of chromosome 14q11.2
that contains other proteasome-related genes including
PA28a and PA28h [5], which bind the 20S proteasome
and enhance the generation of major MHC class I-binding
peptides [5,7]. Transcription of these genes as well as the
ISGF3 gene located immediately downstream of zibra is
induced by interferon g [5], although zibra itself is not
regulated by interferon g [5] (this report). The products of
other genes in this region are not well characterized with the
notable exception of NEDD8, an ubiquitin-like molecule.
Furthermore, zibra codes for both the in-between-ring finger
(IBR) motif, common among E2 and E3 enzymes in the
ubiquitination of targeted substrates, and the ubiquitin-
associated (UBA) domain, responsible for binding multi-
ubiquitin chains noncovalently.
The RING finger is a zinc-binding protein domain that
was initially characterized as a protein interaction domain
[8]. More recently, it has become clear that a large number
of proteins containing RING finger motifs function as E3
ligases, with the RING finger motif itself serving to recruit
specific E2s [9]. In addition, the RING finger has been
demonstrated to be necessary and sufficient to specifically
catalyze the ubiquitin ligase reaction [10].
Methods and materials
Cell culture
Normal breast epithelial cells MCF10-2A, MCF10-2F,
MCF12A, and MCF12F were cultured in 1:1 DMEM/
Ham’s F12 medium supplemented with 20 ng/ml EGF,
100 ng/ml cholera toxin, 0.01 mg/ml bovine insulin, and
500 ng/ml hydrocortisone. These four cell lines were
cultured with 5% horse serum, and in the case of MCF10-
2F and MCF12F, the serum was chelexed before use. The
transformed cell lines MCF-7, HCC1806, HCC1428,
DU4475, and BT-20 were cultured with 10% FBS supple-
mented EMEM (MCF-7 and BT-20) or RPMI 1640
(HCC1806, HCC1428, and DU4475). Cells were routinely
cultured in monolayer in 10 cm2 culture dishes in the
recommended media until <80% confluent and passaged
at 1:4 dilution two to three times per week. NIH3T3 cells
were cultured in DMEM with 10% CS. All cell lines were
purchased from ATCC (Manassas, VA).
Proteasome inhibitor I (PSI) (Calbiochem, San Diego,
CA) at a final concentration of 10 AM or 0.075% DMSO
(v/v) vehicle was added to cultured HCC1806 cells for the
times indicated, and the cells were harvested for RNA or
protein collection. INFg (Biosource International, Camarillo,
CA) was added to cultured HCC1806 cells at a final
concentration of 200 Units/ml for 2–72 h before harvesting
RNA. 17, h-estradiol (Sigma, St. Louis, MO) at a final
concentration of 10�7 M was added to HCC1806 cells
cultured in phenol red-free DMEM (Invitrogen, Carlsbad,
CA) with 10% charcoal stripped FBS (Gemini Bioproducts,
Woodland, CA) for 24 h. Media were changed to serum-free
overnight for NIH3T3 cells transfected with the zibra pro-
moter after 48 h, and then treated for 8 h with either 10 ng/ml
IL-1 or 20 ng/ml TNF-a in serum-free DMEM.
Recovery of mRNA from cultured cells
Total RNA was harvested from cells solubilized in TRI
Reagent (Molecular Research Center, Cincinnati, OH)
according to manufacturer’s instructions. RNA extraction,
precipitation, and solubilization were performed as de-
scribed [2]. A measure of 250 Ag total RNA was used for
recovery of polyA mRNA from each cell line cultured
using a spin-column format (Qiagen, Valencia, CA).
Message RNA was quantitated by fluorescence using
the RiboGreen RNA Quantitation Reagent (Molecular
Probes, Eugene, OR) assayed in triplicate using the
Fluoroskan Accent FL combination luminometer/fluorom-
eter (LabSystems, Finland).
RT-PCR
Reverse transcription was performed using the Thermo-
Script RT-PCR system according to manufacturer’s instruc-
tions (Invitrogen). In the case where cDNA was prepared
by RLM-RACE (Ambion, Austin, TX), 2 Al of 50 AMrandom decamers was substituted in place of oligo (dT).
RLM-RACE PCR
5V RLM-RACE PCR was performed according to
Ambion’s recommendations and general PCR conditions.
The forward primers used were specific to the 5V RACEadapter that was ligated to the RNA before the RT-PCR
reaction. Both Taq Polymerase (Promega, Madison, WI) and
Pfu Turbo polymerase (Stratagene, La Jolla, CA) were
utilized for the 5V RLM RACE PCR reactions. The first
round of PCR used ‘‘outer primers’’; the second round used
‘‘inner primers’’ (Table 1) to specifically amplify the first
reaction. 3V RLM-RACE did not involve a second round
reaction as the inside primer designed with the cloning site
worked well in a single round.
Cloning Ets factor genes
Genes of interest were cloned in the usual way either
using prostate cDNA as a template (Clontech, Palo Alto,
CA) or cDNA prepared in house from various cell lines as
described [2]. Primers were designed from sequences avail-
able through the NCBI database (Table 2). Genes were
cloned using Pfu Turbo DNA polymerase (Stratagene).
Subsequently, the genes were cloned into pcDNA3.1 (Invi-
trogen) for mammalian expression. Usable amounts of the
constructs were made by Maxi prep (Qiagen).

Table 1
RLM-RACE primers
Primer name Sequence AK000973
(1804 bp)
BC012077
(3496 bp)
AY256461
(zibra: 3211 bp)
Tm (jC)
5V OR-RACE1 5V-TGAGCGTTGCAGCGGACAAG-3V 384 354 No binding 79
5V IR-RACE1 5V-CGCAAGCTT-AAACGCCTGCCCGGAAT-3V 286 256 No binding 75
5V OR-RACE2 5V-ATCTGGTTGCTCCTCTGTGTAG-3V No binding 559 219 69
5V IR-RACE2 5V-CGCAAGCTT-CTTGACACCACGCCAGTA-3V No binding 466 126 67
5V OR-RACE3 5V-TCTCCTGCAGCAGTGACAGT-3V 530 2174 1898 71
5V IR-RACE3 5V-CGCAAGCTT-CTGACCAGGCTCTGCTTGT-3V 459 2103 1827 70
3V OF 5V-CTCGAGACTGCCTCTTCTACCT-3V 1278 2949 2673 70
3V IF 5V-CGCAAGCTT-CGGCTGCCGAGTGATAGA-3V 1399 3070 2794 72
Zibra-specific primers and their relative binding sites on GenBank sequences. OR (outside reverse), IR (inside reverse), OF (outside forward), IF (inside
forward) are indicated for 5V or 3V RLM-RACE primers. RACE1, RACE2, and RACE3 correspond to graphical representation in Fig. 1. HindIII restriction site
is bolded.
H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459450
Cloning of the Zibra promoter region
Genomic DNA was extracted, precipitated, and quanti-
tated as previously described from HCC1806 [3].
HCC1806 genomic DNA (100 ng) was used as a template
for cloning the 2.1 kb zibra promoter and enhancer region.
We targeted approximately 2.3 kb upstream of the putative
+1 transcriptional start site based on sequences available in
NCBI databases (BC017376, AK000973, and BC012077).
Nested PCR was performed to obtain the specific product
using the outside forward primer 5V-CCTGAGTGCCT-CACCTCATC-3V and the outside reverse primer 5V-GTCACTCAGACCACGGTCAC-3V. The 2nd round used
the forward primer 5V-GGGGTACC-GAGGGAAGT-
CAAGTCAGCCACAT-3V and the reverse primer 5V-GTGGTACC-CAGGGTTAGCCGAGAACAGGAA-3V,where the bolded sequences are KpnI restriction sites. The
zibra sense and antisense constructs were subsequently
cloned into the KpnI site of pGL3basic vector (Promega),
screened for direction, and sequenced. All zibra promoter
constructs were made from these parent clones using the
primers in Table 3.
Transfections
Transfections of plasmid DNA were as previously de-
scribed [3], with the following additions and modifications.
Table 2
Ets factor cloning primers
Name Sequence
PDEF-F 5V-CCCAAGCTT-ACCAGGCAGCTAACAGACACA-3VPDEF-R 5V-CGCGGATCC-ATGCCAGGTGTGGTGCAGAAT-3VPEA3-F 5V-CTTAAGCTT-CTCCTGGGAGCAGGTC-3VPEA3-R 5V-GTGGATCC-GCTGGGGGCTAGTAAGA-3VEts1-F 5V-GTTAAGCTT-CCATCCCGTACGTCCC-3VEts1-R 5V-ACGGTACC-TCACTCGTCGGCATCTG-3V
Gene-specific primers for PDEF, PEA3/E1A-F, and Ets1 are shown. Bolded seq
primers were generated from publicly available sequence data of the GenBank ac
NIH3T3 cells were plated in 12-well culture plates at a
density of 80,000 cells/well. After 24 h, cells were trans-
fected with appropriate DNA constructs using the Effectene
Transfection Reagent (Qiagen) in triplicate transfections for
48 h. Control transfections included empty vectors for both
the pGL3-basic and pcDNA3.1(+). In total, 0.5 Ag empty
vector, wild-type zibra promoter, and various deletion
constructs were cotransfected with 0.2 Ag PDEF construct
or the empty pcDNA3.1(+) vector. In some cases, 0.2 AgpSV40 hgal (Promega) was cotransfected. Values normal-
ized to hgal activity used the FluoReporter LacZ/Galacto-
sidase Quantitation kit (Molecular Probes), according to the
manufacturer’s instructions, measured in triplicate. An NF-
nB reporter construct (Stratagene) containing five tandem
NF-nB consensus sequences to drive luciferase expression
was used as a positive control to demonstrate cell response
to TNF-a and IL-1 as well as inducible NF-nB activity.
Where specified, samples were also normalized to total
protein content.
Generation of peptide antibody
Rabbits were immunized with zibra C-terminal peptide
(C)EEIPLGQSIPRRRK conjugated with KLH through the
N-terminus cysteine residue and emulsified in Freund’s
adjuvant at 3-week intervals. The antiserum was harvested
after the third booster (Zymed Labs, South San Francisco,
Binding
position
Restriction
enzyme
Tm (jC) GenBank
accession no.
371 HindIII 74 NM_012391
1813 C BamHI 78 NM_012391
7 HindIII 65 NM_001986
1533 C BamHI 64 NM_001986
249 HindIII 69 NM_005238
1604 C KpnI 68 NM_005238
uences are restriction sites listed. ‘‘C’’ denotes complement strand. These
cession number shown.

Table 3
Zibra promoter primers
Primer sequence (restriction enzyme sites are in bold) Binding position Restriction enzyme
5V-GGGGTACC-GAGGGAAGTCAAGTCAGCCACAT-3V �1457 KpnI
5V-GTGGTACC-CAGGGTTAGCCGAGAACAGGAA-3V +640 C KpnI
5V-GGCGATATC-CTGTGGCAAGAATCTGTAGAG-3V �1147 EcoRV
5VGCGGATATC-ATGGCAGAACGAAGGTTAGG-3V �958 EcoRV
5V-CCGGATATC-TGGAAGCTCCTGCTCACT-3V �527 EcoRV
5V-GCGGATATC-GGCCATGCTGCTTCAGT-3V �311 EcoRV
5V-GGCGATATC-AAGGCTCGACCGCAAGTA-3V +77 EcoRV
5V-GGCGATATC-ACCGGACTGGTCCAGCTA-3V +321 EcoRV
5V-GTCGATATC-GCGCCTGCGGCTAT-3V +375 EcoRV
5V-GTCGATATCAGAGCGCTTGACTGAGG-3V +426 EcoRV
5V-GTCGATATC-TGACACCGCTCCAACCT-3V +540 EcoRV
5V-GGTGATATC-GCCCGTTCCTCCGAAA-3V +571 EcoRV
5V-GGTGATATC-TCCGAAATTGGGTCGCAGTC-3V +580 EcoRV
5V-GTCGATATC-CTTCCTGTTCTCGGCTAAC-3V +618 EcoRV
5V-AGGTTGGAGCGGTGTCA-3V +556 C None
5V-CCGGATATC-TCTGGCACCTGCACTTG-3V 4801 C pGL3basic EcoRV
5V-GTCGCTAGATCTCTGGTCTC-3V �296 None
5V-AGCCAGTTGCCACTAGGA-3 �140 None
5V-CAAGTAGCGGACGGAGA-3V +89 None
5VATGGCAGAACGAAGGTTAGG-3V �958 None
5V-CCGCTCGAG-TCTTCTAAGAGCTGCTAGTCCT-3V +178 C XhoI
5V-CCGCTCGAG-ATGGCAGAACGAAGGTTA-3V �958 XhoI
5V-TCTTCTAAGAGCTGCTAGTCC-3V +178 C None
Promoter-specific primers for the various promoter constructs seen in Fig. 2 are shown. Bolded sequences are restriction sites listed. ‘‘C’’ denotes
complement strand.
H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459 451
CA). This peptide was used as competitive inhibitor to
demonstrate antibody specificity.
SDS-PAGE and Western blots
PBS-washed cells were lysed on ice in RIPA buffer (50
mM Tris pH 8.0, 150 mM NaCl, 1.0% NP-40, 0.5%
sodium deoxycholate, 0.1% SDS, 0.1 mM sodium ortho-
vanadate) supplemented with protease inhibitor cocktail
(Sigma). Protein concentration was determined by addition
of Bradford dye modified for use with detergents (BioRad,
Hercules, CA) [11]. Protein (15 or 40 Ag) was subjected to
SDS-PAGE and transferred to nitrocellulose. Western blot
analyses were carried out in the usual way using appro-
priate primary and horseradish peroxidase (HRP)-conju-
gated secondary antibodies. Two primary anti-zibra
antibodies were used. ‘‘Large’’ zibra was generated from
the same peptide with the first isoleucine of ‘‘small’’ zibra
replaced by valine, as predicted by the zibra mRNA
sequence (Abcam, Cambridge, UK). Anti-large zibra was
diluted 1:250 and anti-small zibra was diluted 1:2000.
HRP-conjugated donkey anti-rabbit, donkey anti-goat, and
sheep anti-mouse IgG secondary antibodies were pur-
chased from Abcam and diluted 1:2500, 1:3000, and
1:1500, respectively.
Specific competitive peptide inhibition was carried out
using 200 Ag unconjugated peptide preincubated with the
primary antibody for 2 h before addition to the membrane.
Noncompetitive control peptide RPPGFSPFR (Peptide
Institute, Inc. Osaka, Japan) was incubated under identical
conditions.
Results
Analysis of zibra’s location on chromosome 14q11.2
shows three previously sequenced full-length transcripts
from cDNA libraries. GenBank accession number
BC017376 and AK00973 list 1975- and 1804-bp (‘‘short’’)
sequences cloned from ovarian adenocarcinoma and the
head of a human 10-week embryo, respectively. Both nearly
perfectly overlap the 3496-bp (‘‘long’’) sequence BC012077
cloned from human placenta choriocarcinoma (Fig. 1), but
when translated results in the deletion of 560 and 10 amino
acids. The 10 amino acid deletion would result in the
clipping of the IBR domain, which would presumably affect
the function of the molecule where it is expressed. Another
notable exception is a single nucleotide difference near the
3V end which allows a conservative amino acid substitution
from isoleucine to valine. Other sequences are available
through GenBank, but are comprised of overlapping
expressed sequence tags (ESTs) and the composite sequen-
ces may or may not represent true transcripts.
Identification of zibra mRNA structure
To determine the +1 transcription start site of the zibra
gene, we performed 5V RLM-RACE on HCC1806-trans-

Fig. 1. Alternative transcripts identified by 5V RLM-RACE. Genomic alignment of three alternative zibra transcripts identified by RLM-RACE on chromosome
14q11.2. Genomic numbering is noted as base position. Adjacent regions to the +1 were sequenced. RLM-RACE2 was amplified by PCR using a reverse
primer generated from 3V RLM-RACE data, subsequently cloned, and sequenced. The full-length sequence is shown (zibra). Two other fully sequenced
transcripts, AK000973 and BC012077, are shown for reference. Alignment was performed using the UCSC Genome Browser located at http://
www.genome.ucsc.edu/ using the April 2003 freeze [42,43]. The location of a predicted CpG island is indicated. The hollow arrow indicates a novel 21-bp
upstream start site. The filled arrow indicated a novel 64-bp region in the full-length transcript previously unreported. Zibra sequence data have been deposited
in GenBank under accession number AY256461. Full 5V sequence data for these transcripts are provided with the exon splice sites bold and underlined.
H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459452
formed breast epithelial cells in which mature 5V-cappedmRNA is selectively reverse transcribed, amplified, and
sequenced. Using a reverse primer that was expected to
bind both the long and the short zibra sequences, we
identified a novel 21-bp sequence with the +1 site approx-
imately 650 bp upstream of the previous clones (‘‘RLM-
Fig. 2. Identification of the zibra domain structure. Analysis of the deduced ami
ubiquitin associated domain (UBA), zinc finger (ZF_RING_1, zf-RanBP, ZF_RA
(IBR) domain. Each domain type is represented separately for clarity due to the h
refers to a database entry for the domain, accessible at http://www.ebi.ac.uk/interpro
method. Entries beginning with PS, PF, and SM refer to ProSmart, ProFam, or S
protein sequence used for this analysis is the sequence from our sequenced full
Q86VI2.
RACE1,’’ Fig. 1). Using a reverse primer that was
expected only to bind the long zibra sequence, we identi-
fied a second distinct transcript with the same +1 site
(‘‘RLM-RACE2,’’ Fig. 1). Unexpectedly, when we used a
reverse primer that bound midway in the gene, an addi-
tional +1 start site was identified (‘‘RLM-RACE3’’, Fig. 1),
no acid sequence revealed the presence of several domains, including the
NBP2_1, ZF_RANBP2_2, and ZnF_RBZ), and an in-between-ring finger
omology of zibra with several zinc finger domain types. The Interpro entry
/ISpy?mode=single&ac=Q86VI2. Method accession refers to the predictive
MART, respectively. Vertical bars denote 10 amino acid blocks. The zibra
-length zibra, GenBank accession number AAP12522, TrEMBL accession

H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459 453
with the immediate downstream sequence identical to the
long zibra sequence.
The finding of the RLM-RACE3 transcription start site
midway in the zibra gene raises the possibility that this is an
artifact. We are unable to rule this possibility out as
Northern blot analysis is not sensitive enough to detect the
relatively low level of gene expression, and this sequence
cannot be differentiated from other transcripts by real-time
PCR as it is entirely contained within the zibra sequence and
is not unique. However, the RLM-RACE method is
designed to exclusively amplify the mature message that
has been 5V-capped, and we were unable to detect any
product in multiple negative control RLM-RACE reactions.
The DNA corresponding to RLM-RACE2 was cloned,
sequenced, and submitted as GenBank accession number
AY256461 (‘‘zibra,’’ Fig. 1). In addition to the novel +1
start site (hollow arrow, Fig. 1), an additional 64-bp tran-
Fig. 3. Zibra mRNA and protein expression levels in normal and transformed
HCC1806, and MCF-7) breast epithelial cells were used for these studies. (A) Zibr
lines were measured by real-time PCR. Samples were assayed three times in dup
SEM. (B) Zibra protein expression of the same cell lines. Identical duplicate blots
show antibody specificity. Molecular weight markers are indicated. Arrow shows
scribed sequence previously unreported is identified (filled
arrow, Fig. 1). The 3V polyA tail of the mRNA was also
determined by 3V RLM-RACE with only one identified end
sequence.
Because the RLM-RACE data demonstrate the presence
of at least two different transcripts with the novel 21 bp
upstream region, we expected multiple PCR products using
a forward primer specific to the novel 21 bp sequence and a
reverse primer designed from the 3V RLM-RACE data.
However, the full-length zibra sequence was the only
detectable transcript. Similarly, primers designed to amplify
both the long and short zibra sequences (BC012077
andAK000973) generated only the long product from
HCC1806 total cDNA. Using these same primers, the long
zibra product could be amplified by means of cDNA from
several other cultured breast epithelial cell lines, including
HCC1428, MCF-7, MCF10-2A, and MCF10-2F (data not
breast epithelial cells. Normal (MCF10-2A) and transformed (HCC1428,
a mRNA levels from one normal and three transformed breast epithelial cell
licate. Data are represented as a percentage of total mRNA. Error bars are
assayed, without or with competitive peptide (cp), indicated by � or +, to
the specific immunoreactive band.

Fig. 4. Genomic sequence of zibra promoter–enhancer. The 1.5-kb
promoter–enhancer construct is presented with putative transcription factor
binding sites identified. The newly identified 21-bp 5V UTR sequence is
bold. Asterisks at some Sp1 sites denote that sequence is identical to an ER
binding site, although estradiol had no effect on promoter activity. Positions
of various promoter transcripts are identified with their appropriate number
for reference. The +1 start sites of the two other FLJ10111 clones (Fig. 1)
are represented with down arrowheads, and the up arrowhead indicates the
beginning of a putative CpG island. The transcription start site of PSME2 is
also identified.
H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459454
shown). Furthermore, a forward primer designed to span the
21 bp exon and the unique exon 2 of RLM-RACE1 poorly
amplified cDNA weakly and nonspecifically from each of
these cell lines, and thus only the sequence of the reported
transcript could be determined.
Zibra protein sequence similarities and expression analysis
The deduced translated protein product of the zibra
transcript is 921 amino acids with a predicted molecular
weight of 102 kDa. The overall structure of the protein is
presented in Fig. 2. Because the zinc finger regions of zibra,
between amino acids 147 and 282, match several different
zinc finger classes, each class is listed individually. A
composite of the overall domain structure is indicated
directly below.
Although zibra mRNA is not detectable by Northern blot
(data not shown), we could quantitatively measure tran-
script levels by real-time PCR. Because we previously
found zibra mRNA to be generally increased in transformed
breast epithelial cells [2] and six of the top 10 zibra-
expressing tissue types are carcinomas based on Unigene
and EST expression information (http://genome-www3.
Stanford.edu/cgi-bin/SMD/source/sourceResult?option=
Number&criteria=NM_017999&choice=Gene), we mea-
sured relative RNA expression in normal and transformed
breast epithelial cell lines. In addition, we analyzed zibra
protein expression by Western blot. To demonstrate antibody
specificity, a competitive peptide generated against the small
zibra sequence was preincubated with the primary antibody
before assay. One immunospecific band is identified at
approximately 145 kDa (Fig. 3B). Curiously, zibra protein
was undetectable in MCF-7 cells which express the highest
level of zibra mRNA.
Zibra migrates at a molecular weight greater than pre-
dicted based on the sum of amino acid residues. Our
sequence data showed that the expressed mRNA transcript
contains a single nucleotide difference allowing for the
conservative amino acid change. Thus, the antibody would
have higher affinity for the short form, and also for the
protein product predicted from the transcript that was the
source of the cDNA sequenced from HCC1806 cells. While
it is possible that we could have observed the short form
(AK000973, predicted 57.6 kDa), we did not observe this
band. Instead, the only specific band (e.g., that could be
competed off with a specific peptide) was the observed band
at approximately 145 kDa. While migrating slower than
expected in the polyacrylamide gel, the presence of the
larger MW protein is in accordance with the full-length
transcript that we were able to sequence from these cells.
The retarded migration by polyacrylamide gel electro-
phoresis could be explained by several possibilities, includ-
ing overall hydrophobicity, posttranslational modifications,
of which there are several high-probability amino acid sites
including those for phosphorylation [12], O-Gln [13], and
sumoylation based on the tetrapeptide motif B-K-x-D/E (B
H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459 455
is a hydrophobic residue, K is the lysine conjugated to
SUMO, x is any amino acid, and D or E is an acidic
residue), and propensity of these amino acids to maintain
similar hydrophobicity.
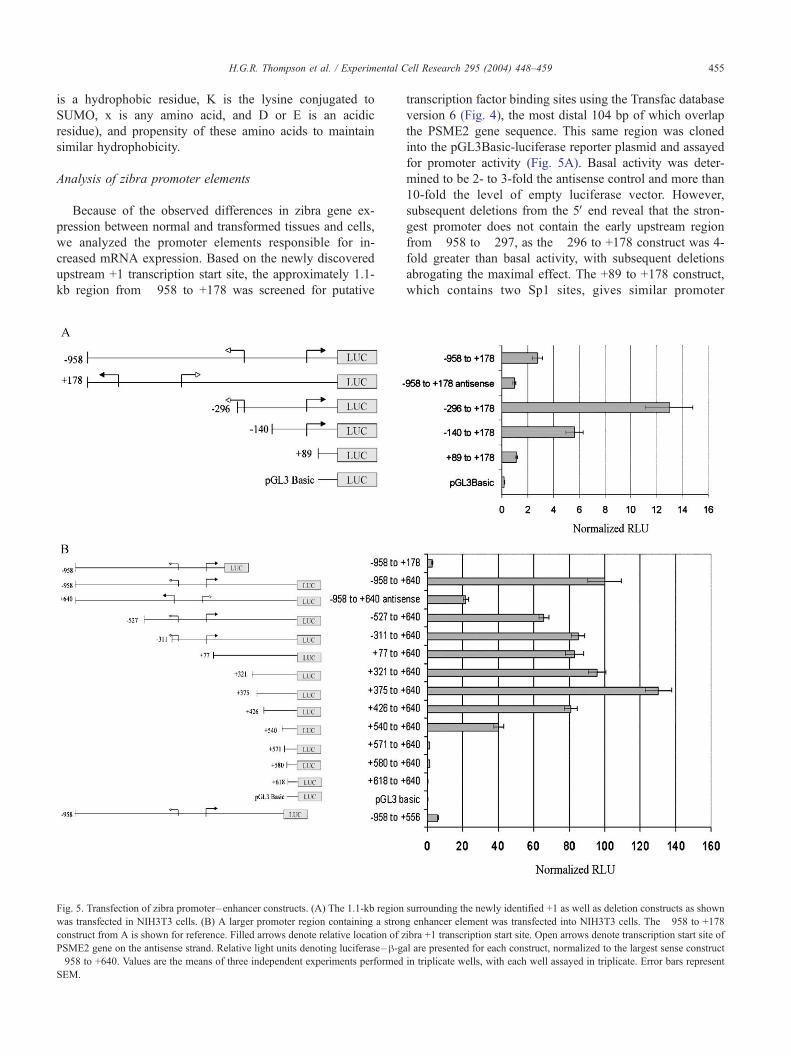
Analysis of zibra promoter elements
Because of the observed differences in zibra gene ex-
pression between normal and transformed tissues and cells,
we analyzed the promoter elements responsible for in-
creased mRNA expression. Based on the newly discovered
upstream +1 transcription start site, the approximately 1.1-
kb region from �958 to +178 was screened for putative
Fig. 5. Transfection of zibra promoter–enhancer constructs. (A) The 1.1-kb region
was transfected in NIH3T3 cells. (B) A larger promoter region containing a stron
construct from A is shown for reference. Filled arrows denote relative location of z
PSME2 gene on the antisense strand. Relative light units denoting luciferase–h-ga�958 to +640. Values are the means of three independent experiments performed
SEM.
transcription factor binding sites using the Transfac database
version 6 (Fig. 4), the most distal 104 bp of which overlap
the PSME2 gene sequence. This same region was cloned
into the pGL3Basic-luciferase reporter plasmid and assayed
for promoter activity (Fig. 5A). Basal activity was deter-
mined to be 2- to 3-fold the antisense control and more than
10-fold the level of empty luciferase vector. However,
subsequent deletions from the 5V end reveal that the stron-
gest promoter does not contain the early upstream region
from �958 to �297, as the �296 to +178 construct was 4-
fold greater than basal activity, with subsequent deletions
abrogating the maximal effect. The +89 to +178 construct,
which contains two Sp1 sites, gives similar promoter
surrounding the newly identified +1 as well as deletion constructs as shown
g enhancer element was transfected into NIH3T3 cells. The �958 to +178
ibra +1 transcription start site. Open arrows denote transcription start site of
l are presented for each construct, normalized to the largest sense construct
in triplicate wells, with each well assayed in triplicate. Error bars represent

H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459456
activity as the much larger �958 to +178 promoter, suggest-
ing that these Sp1 sites contribute to the basal promoter
activity. Deletion of the �296 to �140 region eliminates
more than half of the maximal promoter activity. Analysis of
this region reveals the presence of two consensus sequences
for TFIID/TBP/Sp1 (Fig. 5A).
Next, we studied a larger approximately 1.5 kb promoter-
luciferase construct using the same 5V end with an additional
462 bp downstream that encompassed the first base of the
previously reported short zibra sequence. The largest pro-
moter, �958 to +640, drove luciferase expression approx-
imately 40-fold higher than the �958 to +178 construct
(Fig. 5B), suggesting regulatory elements between +178 and
+640. Because of the proximal location of zibra to PSME2
(encoding PA28h) in a head-to-head configuration less than
1 kb away, the antisense promoter construct was assayed for
activity. While this construct possessed significant promoter
activity, it is interesting to note that deletion of the final 84
bp of the sense construct abolishes activity to levels lower
than antisense. These data suggest that the genomic region
between PSME2 and zibra confers some activity in both
orientations.
Subsequent deletions of the large promoter construct
from the 5V end reveal essentially full promoter activity
with the +426 to +640 construct. This is in accordance with
previous finding that most promoter activity is located
between +178 and +640, as seen when the �958 to +178
and �958 to +640 constructs are compared. Deletion of the
next 114 bp from the 5V end dramatically reduces promoter
activity by half, and deletion of the next 31 bp nearly
abolishes all activity. We see a similar knockdown of
promoter activity with the nearly full-length �958 to +556
construct. Taken together, these data provide evidence of a
core element of basal promoter activity that lies between
+557 and +570. The only identified transcriptional ele-
ments, based on the TRANSFAC database V6.0, that may
be responsible for this activity is a TFIID/TBP/Sp1 consen-
sus site located +566 to +575 (Fig. 4). The location of this
Fig. 6. Transfection of zibra promoter–enhancer constructs and treatment with TN
as a deletion construct eliminating a strong consensus NF-nB binding site was tra
positive control. After transfection, cells were treated with TNF-a or IL-1 for 8 h a
the means of three independent experiments performed in triplicate wells, with e
sequence 48 bp upstream of the +1 reported for the short
zibra sequence and 78 bp upstream of the +1 reported for
the long zibra sequence is suggestive that this element may
play an important role in the basal regulation of these
transcripts.
Because PDEF and zibra mRNA are coordinately in-
creased in transformed breast epithelial cells in culture [2],
and several Ets factor family members shown to be upregu-
lated in cancer [14–17] are potent regulators of promoter
activity [18–21], cotransfection experiments with the zibra
promoter and three Ets factors were performed. Initially, it
appeared as though Ets-1, PDEF, and PEA3 all dramatically
stimulated the zibra promoter, in order of increasing poten-
cy, as measured by luciferase activity. However, it was
determined that each of these transcription factors affected
h-gal expression via the SV40 enhancer. When luciferase
activity was divided by total protein, no promoter regulation
was observed alone or in combination (data not shown). In
addition, several consensus NF-nB and estrogen receptor
binding sites are present in the upstream 0.5-kb region (Fig.
4). Therefore, we assessed the possibility that NF-nB or
estrogen regulates zibra expression. While addition of TNF-
a or IL-1 increased NF-nB activity as measured by the NF-
nB reporter construct, neither cytokine affected the zibra
promoter (Fig. 6) or protein concentration (data not shown).
In addition, neither 200 U/ml INFa˜nor 10�7 M estradiol
had any effect.
Previous studies have assessed the role of proteasome
inhibitors on cellular death in several different cell types
[22–25]. An appropriate inhibitor and dose was determined
with cell proliferation assays [3]. Previous findings demon-
strate that PSI treatment of cultured HCC1806 cells, as well
as other normal and transformed breast epithelial cells,
results in a rapid increase of ubiquitinated proteins and a
time-dependent increase of p62 protein and stabilization of
p62 mRNA. The ability of p62 to bind ubiquitin through its
UBA domain, its coexpression with zibra mRNA, and its
ability to bind several signaling proteins suggest that p62
F-a and IL-1. The 1.1-kb region surrounding the newly identified +1 as well
nsfected in NIH3T3 cells. A NF-nB reporter construct was transfected as a
nd promoter activity measured by luciferase–h-gal. Relative light units areach well assayed in triplicate. Error bars represent SEM.

Fig. 7. Effect of PSI on zibra mRNA and protein expression. HCC1806
cells were cultured in the presence of 10 AM PSI for 8, 16, or 32 h.
Untreated and 32-h DMSO-treated cells served as controls. (A) Total cell
lysate was assayed for zibra protein by Western blot. h-actin is included as aloading control. (B) mRNA levels were measured by real-time PCR. Values
are the means of three independent experiments performed in duplicate and
represented as a percentage of total mRNA. Error bars represent SEM.
H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459 457
may play a regulatory role in the ubiquitin-proteasome
pathway. To assess the effects of PSI on zibra expression,
HCC1806 cells were treated with 10 AM PSI for 0, 8, 16, or
32 h. A 32 h DMSO vehicle treatment was included as a
control. A measure of 40 Ag total cell lysate, extracted in
RIPA buffer, was subjected to Western blot analysis for
zibra and h-actin and visualized by chemiluminescence. PSI
dramatically decreased zibra protein expression after 8 h and
was still undetectable after 32 h (Fig. 7A). To show equal
loading of cell lysate, h-actin is included as a control.
To determine whether the decrease of zibra protein in
PSI-treated cells was because of decreased mRNA, mRNA
expression level of zibra from HCC1806 cells treated with
10 AM PSI was measured by real-time PCR over 32 h (Fig.
7B). DMSO-treated vehicle values are shown for reference.
The addition of PSI to cultured HCC1806 cell had essen-
tially no effect on mRNA levels of the zibra transcript at any
time point. Cells treated with DMSO for 32 h served as
control.
Discussion
Because both the long and short submitted zibra sequen-
ces were obtained from cDNA libraries, it is possible that
these sequences are not full-length or mature transcripts.
Thus, we determined both the 5V and 3V end by RLM-RACE.
As such, we identified a novel 21-bp transcribed region
upstream of the previously reported sequence as well as an
additional 64-bp region at the end of exon 3 (filled arrow,
Fig. 1). In an effort to confirm the presence of the short zibra
sequence, we designed a reverse RLM-RACE primer at the
5V end of exon 11. While, we could not verify the presence
of the short transcript, we identified an additional transcript
(‘‘RLM-RACE3’’, Fig. 1) that begins within exon 9 of zibra,
but contains all of exon 10, as in the full-length zibra
sequence. Interestingly, a recent paper predicted and vali-
dated 845 significantly tumor-associated alternative splic-
ings, including FLJ10111 (zibra) with a 23-bp insertion at
position 1026 of the NM_017999 (short zibra) sequence
[26]. However, we did not observe this insertion.
Recent analysis of publicly available mRNA expression
data revealed a module of genes that are coexpressed in
several tissue types, including breast cancer [2]. Member
genes of this module include the Ets factor transcription
factor PDEF, SQSTM1/p62, PSMD7, and zibra. Because
EST and UniGene data suggest that zibra mRNA is over-
expressed in several cancers and our previous findings in
breast cell lines, we analyzed zibra mRNA and protein
expression in breast cancer cells. In this study, we demon-
strate that while zibra mRNA expression may in fact be a
marker of breast cancer, zibra protein levels may not parallel
mRNA levels. For instance, we observe that MCF-7 cells in
which zibra mRNA is approximately 3-fold greater express
less protein than other cell lines studied. Of the members of
the module of coexpressed genes in breast cancer, zibra is
not alone in this regard. PDEF mRNA is expressed up to
three orders of magnitude greater in breast cancer cells
compared to normal cells. However, no studies to date have
identified PDEF protein in breast cancer, and one study
finds that PDEF protein is reduced in human invasive breast
cancer and is absent in invasive breast cancer cell lines [27].
The apparent discrepancy between zibra mRNA and
protein levels suggests several possibilities. The 5V end of
the transcript contains a putative CpG island, and the
methylation status of this region may confer epigenetic
control on the expression of the gene. In cancer cell lines,
we noted increased expression of the zibra gene, which may
suggest decreased methylation. In addition, capped mRNA
can also be methylated, although the effects on translation
are not yet clear. Alternatively, feedback mechanisms, the
simplest of which is that lack of protein, either by reduced
translation or increased degradation, may signal an increase
in zibra transcription. This is not likely the case, as addition
of proteasome inhibitor rapidly reduces zibra protein con-
centration within the cell without any change in mRNA.
Even after 32 h, the protein is decreased, with only minor
changes on mRNA. Another more likely scenario supported
by our data is that zibra protein concentration is tightly
regulated, and mRNA expression is a function of the
individual cell type. Transformed breast epithelial cells
may, in general, express more zibra mRNA due to altered
RNA stability or transcription mechanisms.
Thus, in an effort to determine the functional regulation
of zibra gene expression, we cloned the promoter encom-

H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459458
passing both the previously reported transcription start site
and the newly identified +1 site. The �958 to +178
construct, based on the new start site, has approximately
3-fold promoter activity compared to its antisense. Remov-
ing 662 bp from the 5V end increases basal promoter activity
another 4-fold (Fig. 5A). However, adding 462 bp to the 3Vend increased promoter activity greater than 30-fold (Fig.
5B). Thus, most basal promoter activity lies between +178
and +640.
While analysis of the promoter identified a Kozak
consensus sequence at +39 (Fig. 4), identical activities are
observed between the �311 construct and the +77 construct
which does not contain this sequence (Fig. 5B). Therefore,
we conclude that this site has little or no effect on the
transcription of the reporter gene. Various deletion con-
structs found that half of the maximal activity is regulated
between +426 and +540, and the remaining between +540
and +571. Strikingly, the �958 to +556 construct had
minimal activity (compared to �958 to +640), indicating
that nearly all basal promoter activity lies between +557 and
+570.
This 14-bp sequence of DNA contains a short region
predicted to be homologous to only TFIID, TATA Binding
Protein (TBP), Sp1, and estrogen receptor (denoted by the
asterisk on Sp1 in Fig. 4) binding sites. Sp1 is a basal
promoter transcription factor and Sp1 activation of TATA-
less promoters, such as this one, requires interaction with
TFIID [28] and multiple Sp1 sites are synergistic and
required for maximal stimulation [29]. In TATA-less pro-
moters, Sp1 also interacts with other proteins other than
TFIID to stimulate transcription, including E2 [30,31], NF-
nB [32], Oct-1 [33], SREBP-1 [34,35], and the Ets factors
Ets-1 [36,37] and PU.1 [38].
To determine the mechanism of zibra gene regulation, we
first looked at Ets factors because of the large number of Ets
factor consensus sequences in the promoter, synergistic
activation with Sp1, and the predicted and confirmed coex-
pression of zibra with PDEF. We cotransfected PDEF, Ets-1,
and PEA-3/E1A-F with various zibra promoter constructs
either alone or in combination. Each of these factors is
overexpressed in cancer models and directly implicated in
tumorigenesis. Surprisingly, we observed no changes in
promoter activity with the addition of these transcription
factors.
We next assessed the effect of estrogen, as the zibra
promoter contains a large number of estrogen receptor
binding sites including one in the critical short basal region,
and many breast cancer genes are dysregulated by estrogens.
We treated promoter-transfected cells with physiologically
relevant concentrations of 17-h-estradiol but observed no
changes in promoter activity. Lastly, because NF-nB has
been shown to regulate genes involved in cancers, its link to
Sp1, its observed synergistic transcriptional activation with
Ets-1 [39], and the finding that biological morphogens, such
as TNF-a, upregulate the expression of Ets-1 transcription
factor [40,41], we treated cells with either TNF-a or IL-1 to
increase endogenous NF-nB activity, again with no effect on
promoter activity or zibra protein expression.
Zibra maps to a region of chromosome 14 in a locus
rich in genes involved in the structure and function of the
proteasome. Because the amino acid motifs are consistent
with E3 ubiquitin ligases in the proteasome degradation
pathway and our previous finding that zibra is coex-
pressed with other genes likely involved in the ubiqui-
tin-proteasome pathway, we assessed whether chemically
inhibiting the proteasome would exert a regulatory effect
on gene expression. We found that while there was
essentially no change in mRNA levels, protein levels
dropped by 8 h and were still undetectable after 32 h.
These data taken together suggest that the zibra protein is
regulated posttranscriptionally. This is in accordance with
our previous and current findings that zibra mRNA levels
are increased in transformed breast epithelial cells com-
pared to normal cells, but protein levels are essentially the
same or lower.
In conclusion, zibra mRNA is expressed at high levels in
several distinct cancers, but its protein expression appears
to be uncorrelated to mRNA levels. It has at least three
alternative transcripts. Its domain structure, location on
chromosome 14, and responsiveness to proteasome inhibi-
tion suggest that one cellular function is as an ubiquitin
ligase. Zibra’s promoter contains a small 14-bp region that
is important for basal transcription, and protein levels can
be regulated posttranscriptionally.
Acknowledgment
This work was supported by the National Institutes of
Health Human Genome Research Institute grant number
1K22HG00047-01.
References
[1] D. Hanahan, R.A. Weinberg, The hallmarks of cancer, Cell 100
(2000) 57–70.
[2] H.G.R. Thompson, J.W. Harris, B.J. Wold, S.R. Quake, J.P. Brody,
Identification and confirmation of a module of coexpressed genes,
Genome Res. 12 (2002) 1517–1522.
[3] H.G.R. Thompson, J.W. Harris, B.J. Wold, F. Lin, J.P. Brody, p62
overexpression in breast tumors and regulation by prostate-derived
Ets factor in breast cancer cells, Oncogene 22 (2003) 2322–2333.
[4] A. Ghadersohi, A.K. Sood, Prostate epithelium-derived Ets transcrip-
tion factor mRNA is overexpressed in human breast tumors and is a
candidate breast tumor marker and a breast tumor antigen, Clin. Can-
cer Res. 7 (2001) 2731–2738.
[5] M. Yawata, S. Murata, K. Tanaka, Y. Ishigatsubo, M. Kasahara, Nu-
cleotide sequence analysis of the approximately 35-kb segment con-
taining interferon-gamma-inducible mouse proteasome activator
genes, Immunogenetics 53 (2001) 119–129.
[6] J. Kent, C. Sugnet, T. Furey, D. Haussler, M. Schwartz, A. Hinrichs,
D. Karolchik, H. Trumbower, UCSC Genome Browser, The Regents
of the University of California, Santa Cruz, CA, 2001.

H.G.R. Thompson et al. / Experimental Cell Research 295 (2004) 448–459 459
[7] C.P. Ma, C.A. Slaughter, G.N. DeMartino, Identification, purification,
and characterization of a protein activator (PA28) of the 20 S protea-
some (macropain), J. Biol. Chem. 267 (1992) 10515–10523.
[8] K.L. Borden, RING fingers and B-boxes: zinc-binding protein–pro-
tein interaction domains, Biochem. Cell Biol. 76 (1998) 351–358.
[9] C.M. Pickart, Mechanisms underlying ubiquitination, Annu. Rev.
Biochem. 70 (2001) 503–533.
[10] T. Araki, J. Milbrandt, ZNRF proteins constitute a family of presyn-
aptic E3 ubiquitin ligases, J. Neurosci. 23 (2003) 9385–9394.
[11] M.M. Bradford, A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding, Anal. Biochem. 72 (1976) 248–254.
[12] N. Blom, S. Gammeltoft, S. Brunak, Sequence and structure-based
prediction of eukaryotic protein phosphorylation sites, J. Mol. Biol.
294 (1999) 1351–1362.
[13] J.E. Hansen, O. Lund, N. Tolstrup, A.A. Gooley, K.L. Williams, S.
Brunak, NetOglyc: prediction of mucin type O-glycosylation sites
based on sequence context and surface accessibility, Glycoconjugate
J. 15 (1998) 115–130.
[14] D.K. Watson, M.J. McWilliams-Smith, M.F. Nunn, P.H. Duesberg,
S.J. O’Brien, T.S. Papas, The ets sequence from the transforming gene
of avian erythroblastosis virus, E26, has unique domains on human
chromosomes 11 and 21: both loci are transcriptionally active, Proc.
Natl. Acad. Sci. U. S. A. 82 (1985) 7294–7298.
[15] M.S. Trimble, J.H. Xin, C.T. Guy, W.J. Muller, J.A. Hassell, PEA3 is
overexpressed in mouse metastatic mammary adenocarcinomas, On-
cogene 8 (1993) 3037–3042.
[16] C.C. Benz, R.C. O’Hagan, B. Richter, G.K. Scott, C.H. Chang, X.
Xiong, K. Chew, B.M. Ljung, S. Edgerton, A. Thor, J.A. Hassell,
HER2/Neu and the Ets transcription activator PEA3 are coordinate-
ly upregulated in human breast cancer, Oncogene 15 (1997)
1513–1525.
[17] I. Afrikanova, E. Yeh, D. Bartos, S.S. Watowich, G.D. Longmore,
Oncogene cooperativity in Friend erythroleukemia: erythropoietin re-
ceptor activation by the env gene of SFFV leads to transcriptional
upregulation of PU.1, independent of SFFV proviral insertion, Onco-
gene 21 (2002) 1272–1284.
[18] Y. de Launoit, A. Chotteau-Lelievre, C. Beaudoin, L. Coutte, S.
Netzer, C. Brenner, I. Huvent, J.L. Baert, The PEA3 group of ETS-
related transcription factors. Role in breast cancer metastasis, Adv.
Exp. Med. Biol. 480 (2000) 107–116.
[19] F. Higashino, K. Yoshida, T. Noumi, M. Seiki, K. Fujinaga, Ets-
related protein E1A-F can activate three different matrix metallopro-
teinase gene promoters, Oncogene 10 (1995) 1461–1463.
[20] Z. Su, Y. Shi, P.B. Fisher, Cooperation between AP1 and PEA3 sites
within the progression elevated gene-3 (PEG-3) promoter regulate
basal and differential expression of PEG-3 during progression of
the oncogenic phenotype in transformed rat embryo cells, Oncogene
19 (2000) 3411–3421.
[21] X. Xing, S.C. Wang, W. Xia, Y. Zou, R. Shao, K.Y. Kwong, Z. Yu, S.
Zhang, S. Miller, L. Huang, M.C. Hung, The ets protein PEA3 sup-
presses HER-2/neu overexpression and inhibits tumorigenesis, Nat.
Med. 6 (2000) 189–195.
[22] B. An, R.H. Goldfarb, R. Siman, Q.P. Dou, Novel dipeptidyl protea-
some inhibitors overcome Bcl-2 protective function and selectively
accumulate the cyclin-dependent kinase inhibitor p27 and induce ap-
optosis in transformed, but not normal, human fibroblasts, Cell Death
Differ. 5 (1998) 1062–1075.
[23] L.A. Pasquini, M. Besio Moreno, A.M. Adamo, J.M. Pasquini, E.F.
Soto, Lactacystin, a specific inhibitor of the proteasome, induces
apoptosis and activates caspase-3 in cultured cerebellar granule cells,
J. Neurosci. Res. 59 (2000) 601–611.
[24] X.M. Fan, B.C. Wong, W.P. Wang, X.M. Zhou, C.H. Cho, S.T. Yuen,
S.Y. Leung, M.C. Lin, H.F. Kung, S.K. Lam, Inhibition of proteasome
function induced apoptosis in gastric cancer, Int. J. Cancer 93 (2001)
481–488.
[25] E. Kuusisto, T. Suuronen, A. Salminen, Ubiquitin-binding protein
p62 expression is induced during apoptosis and proteasomal inhibi-
tion in neuronal cells, Biochem. Biophys. Res. Commun. 280 (2001)
223–228.
[26] Z. Wang, H.S. Lo, H. Yang, S. Gere, Y. Hu, K.H. Buetow, M.P. Lee,
Computational analysis and experimental validation of tumor-associ-
ated alternative RNA splicing in human cancer, Cancer Res. 63
(2003) 655–657.
[27] R.J. Feldman, V.I. Sementchenko, M. Gayed, M.M. Fraig, D.K.
Watson, Pdef expression in human breast cancer is correlated with
invasive potential and altered gene expression, Cancer Res. 63
(2003) 4626–4631.
[28] K.H. Emami, T.W. Burke, S.T. Smale, Sp1 activation of a TATA-less
promoter requires a species-specific interaction involving transcrip-
tion factor IID, Nucleic Acids Res. 26 (1998) 839–846.
[29] X. Chen, J.C. Azizkhan, D.C. Lee, The binding of transcription factor
Sp1 to multiple sites is required for maximal expression from the rat
transforming growth factor alpha promoter, Oncogene 7 (1992)
1805–1815.
[30] R. Li, J.D. Knight, S.P. Jackson, R. Tjian, M.R. Botchan, Direct
interaction between Sp1 and the BPV enhancer E2 protein mediates
synergistic activation of transcription, Cell 65 (1991) 493–505.
[31] J.D. Knight, R. Li, M. Botchan, The activation domain of the bovine
papillomavirus E2 protein mediates association of DNA-bound
dimers to form DNA loops, Proc. Natl. Acad. Sci. U. S. A. 88
(1991) 3204–3208.
[32] B. Majello, P. De Luca, G. Hagen, G. Suske, L. Lania, Different
members of the Sp1 multigene family exert opposite transcriptional
regulation of the long terminal repeat of HIV-1, Nucleic Acids Res. 22
(1994) 4914–4921.
[33] A.C. Strom, M. Forsberg, P. Lillhager, G. Westin, The transcription
factors Sp1 and Oct-1 interact physically to regulate human U2
snRNA gene expression, Nucleic Acids Res. 24 (1996) 1981–1986.
[34] L. Yieh, H.B. Sanchez, T.F. Osborne, Domains of transcription factor
Sp1 required for synergistic activation with sterol regulatory element
binding protein 1 of low density lipoprotein receptor promoter, Proc.
Natl. Acad. Sci. U. S. A. 92 (1995) 6102–6106.
[35] H.B. Sanchez, L. Yieh, T.F. Osborne, Cooperation by sterol regulatory
element-binding protein and Sp1 in sterol regulation of low density
lipoprotein receptor gene, J. Biol. Chem. 270 (1995) 1161–1169.
[36] A. Gegonne, R. Bosselut, R.A. Bailly, J. Ghysdael, Synergistic acti-
vation of the HTLV1 LTR Ets-responsive region by transcription
factors Ets1 and Sp1, EMBO J. 12 (1993) 1169–1178.
[37] J. Dittmer, A. Gegonne, S.D. Gitlin, J. Ghysdael, J.N. Brady, Regu-
lation of parathyroid hormone-related protein (PTHrP) gene expres-
sion. Sp1 binds through an inverted CACCC motif and regulates
promoter activity in cooperation with Ets1, J. Biol. Chem. 269
(1994) 21428–21434.
[38] B. Nelsen, G. Tian, B. Erman, J. Gregoire, R. Maki, B. Graves, R.
Sen, Regulation of lymphoid-specific immunoglobulin mu heavy
chain gene enhancer by ETS-domain proteins, Science 261 (1993)
82–86.
[39] R. Li, H. Pei, D.K. Watson, Regulation of Ets function by protein–
protein interactions, Oncogene 19 (2000) 6514–6523.
[40] Y. Chen, H.S. Lillehoj, C.H. Hsu, S.L. Carpenter, S.J. Lamont, Func-
tional characterization of a chicken major histocompatibility complex
class II B gene promoter, Immunogenetics 45 (1997) 242–248.
[41] F. Gilles, M.B. Raes, D. Stehelin, B. Vandenbunder, V. Fafeur, The
c-ets-1 proto-oncogene is a new early-response gene differentially
regulated by cytokines and growth factors in human fibroblasts,
Exp. Cell Res. 222 (1996) 370–378.
[42] W.J. Kent, C.W. Sugnet, T.S. Furey, K.M. Roskin, T.H. Pringle, A.M.
Haussler, D. Haussler, The human genome browser at UCSC, Ge-
nome Res. 12 (2002) 996–1006.
[43] W.J. Kent, BLAT-the BLAST-like alignment tool, Genome Res. 12
(2002) 656–664.




















