The Gut Microbiota and Developmental Programming of the Testis in Mice
Upload
independentCategory
view
2download
0
IDENTIFICATION OF A DEVELOPMENTALLY-REGULATEDPATHWAY OF MEMBRANE RETRIEVAL IN NEURONAL GROWTHCONES
Dario Bonanomi1,§,*, Eugenio F. Fornasiero1,*, Gregorio Valdez2, Simon Halegoua2, FabioBenfenati3,4, Andrea Menegon1, and Flavia Valtorta1,41S.Raffaele Scientific Institute/Vita-Salute University and IIT Unit of Molecular Neuroscience,Milano, Italy2Dept. of Neurobiology and Behavior, State University of New York at Stony Brook, Stony Brook,NY3Dept. of Experimental Medicine, University of Genova and Dept. of Neuroscience and BrainTechnologies, IIT Central Laboratories, Genova4Istituto Nazionale di Neuroscienze
AbstractDuring axon navigation and upon target recognition the growth cone plasma membrane is constantlyreconfigured as a result of changes in cytoskeletal and membrane dynamics. The identity andregulation of the membrane pathway(s) participating in remodeling of the growth cone surface remainelusive. Here, we identify a constitutive, high capacity plasma membrane recycling activity in theaxonal growth cones which is mediated by a novel bulk endocytic pathway mechanistically relatedto macropinocytosis. This pathway, involving large compartments distributed at sites of intense actin-based membrane ruffling, requires phosphatidylinositol 3-kinase activity, the small GTPase Rac1and the pinocytic chaperone Pincher. At early developmental stages, the synaptic vesicle and classicalendosomal recycling pathways do not participate in the rapid retrieval of the growth cone plasmamembrane. At later stages, during the onset of synaptogenesis, an intrinsic program of maturationleads to downregulation of basal bulk endocytosis and the emergence of depolarization-inducedsynaptic vesicle exo-endocytosis. We propose that the control of bulk membrane retrieval contributesto the homeostatic regulation of the axonal plasma membrane and growth cone remodeling duringaxonal outgrowth. In addition, we suggest that the downregulation of bulk endocytosis duringsynaptogenesis might contribute to the preservation of synaptic vesicle specificity.
Keywordsendocytosis; axolemma; synaptic vesicle; trafficking; fluorescence microscopy
Address correspondence to: Flavia Valtorta, DIBIT 3A3, San Raffaele Scientific Institute, via Olgettina 58, 20132 Milan, Italy; Tel.:39-022643-4826; Fax: 39-022643-4813; E-mail: [email protected].§Present address: Gene Expression Laboratory, The Salk Institute for Biological Studies, La Jolla, CA*These two authors contributed equally to the work
NIH Public AccessAuthor ManuscriptJ Cell Sci. Author manuscript; available in PMC 2009 August 25.
Published in final edited form as:J Cell Sci. 2008 November 15; 121(Pt 22): 3757–3769. doi:10.1242/jcs.033803.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

INTRODUCTIONIn the nervous system, axons are guided to their specific targets by highly motile growth cones,present at their distal tip, which exhibit a remarkable organization. Operationally, they includea peripheral (P) domain composed of actin-rich lamellipodia and filopodia, a central (C) domaincontaining microtubules and various organelles, and a transitional (T) domain, at the interfacebetween the P and C domains, characterized by intense F-actin remodeling and membraneruffling (Dailey and, 1989 Bridgman; Forscher and Smith, 1988; Schaefer et al.,2002). Duringaxon navigation and synapse formation growth cones respond to environmental cues withdrastic rearrangements of their membrane and cytoskeletal components (Dent and Gertler,2003).
The growth cone is recognized as the major site of axonal membrane addition and recycling(Bray, 1970; Sinclair et al., 1988; Parton et al., 1992; Craig et al, 1995; Zakharenko and Popov,2000); however, the control of its membrane dynamics during axon growth and retraction isstill poorly understood. Earlier ultrastructural analyses reported in the C domain thelocalization of a plethora of organelles, still largely undefined in terms of identity (Yamada etal., 1971; Pfenninger and Maylié-Pfenninger, 1981; Tsui et al., 1983; Cheng and Reese,1985). Some of these organelles, including clusters of synaptic vesicle (SV) precursors(Fletcher et al., 1991), are exocytic, subjected to regulatory mechanisms similar to thoseoperative at mature synapses (Bonanomi et al., 2005). These precursors contribute to theformation of the presynaptic pool of vesicles at nascent synaptic contacts (Chow and Poo,1985; Matteoli et al., 1992; Ahmari et al., 2000; for review, see Bonanomi et al 2006) but haveno role in neurite outgrowth and in the expansion of the growth cone plasma membrane(Lockerbie et al., 1991; Leoni et al., 1999; Verhage et al., 2000).
Endocytosis in the growth cone has been shown to take place by at least two processes, oneconstitutive and one evoked, carried out by distinct populations of vesicles (Diefenbach et al.,1999). Interestingly, these processes do not operate independently, but coordinately withexocytosis and in strict interaction with cytoskeletal dynamics. As a consequence, endocytosisis enhanced during growth cone collapse induced by repulsive cues (Fournier et al., 2000;Jurney et al., 2002), but is also required for axon outgrowth (Kim and Wu, 1987; Torre et al.,1994; Mundigl et al., 1998; Albertinazzi et al., 2003). Counterintuitively, membrane retrievalat the newly formed growth cone is a main process associated with vigorous extension of axonsafter axotomy (Ashery et al., 1996). Rapid, bidirectional changes of surface membrane dotherefore take place at growth cones during axon navigation. By combining imaging ofendocytic tracers with interference and overexpression approaches we have investigated thenature and properties of the endocytic processes taking place at the growth cones of developinghippocampal neurons in culture.
MATERIALS AND METHODSMaterials
The following primary antibodies were used: mouse anti-synaptobrevin 2, rabbit anti-APP andrabbit anti-syntaxin13 (Synaptic Systems, Göttingen, Germany); mouse anti Tau-1(MAB3420) (Millipore corporation, Billerica, MA); mouse anti clathrin heavy chain (cloneX22) (ABR-Affinity BioReagents, Golden, CO); mouse anti-hemagglutinin (HA) (RocheDiagnostics, Indianapolis, IN); rabbit anti-panTrk (C-14) (Santa Cruz Biotechnology, Inc.,Santa. Cruz, CA); mouse anti-FLAG M5 (Sigma-Aldrich, St. Louis, MO); rabbit anti L1 (agift of F. Rathjen, Max-Delbrueck-Centrum fuer Molekulare Medizin, Berlin, Germany),rabbit anti Rabankyrin-5 (a gift of M. Zerial, MPI Molecular Cell Biology and genetics,Dresden, Germany); rabbit anti-β1 integrin (a gift from K. Rubin, Uppsala University, Uppsala,Sweden). TRITC and FITC-conjugated secondary antibodies were from Jackson
Bonanomi et al. Page 2
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ImmunoResearch (West Grove, PA). FITC-conjugated phalloidin, filipin III, Lucifer yellowCH, brefeldin A (BFA), LY294002, methyl-β-cyclodextrin (MβCD) were from Sigma-Aldrich(St. Louis, MO). FM4-64, yellow-green (505/515) carboxylate-modified FluoSpheres® beads,transferrin-Alexa488 Fluor, cholera toxin-Alexa488 Fluor were from Invitrogen-MolecularProbes (Carlsbad, CA). Cytochalasin D, was from Calbiochem (La Jolla, CA). All chemicalswere diluted in Kreb’s-Ringer-HEPES (KRH) (in mM: 150 NaCl, 5 KCl, 1.2 MgSO4, 1.2KH2PO4, 2 CaCl2, 10 glucose, and 10 HEPES, pH 7.4) at the indicated concentrations. Fordepletion of extracellular Ca2+, KRH was supplemented with 2 mM EGTA and CaCl2 wassubstituted with 2 mM MgCl2.
Plasmids and viral vectorsThe pFLAG-N17-Rac1A plasmid coding for the dominant negative form of the avian Rac1AGTPase, and the pFLAG-V12-Rac1A plasmid coding for the constitutively active form of theRac GTPases were a gift of Dr. Ivan de Curtis (DIBIT-San Raffaele Scientific Institute, Milan,Italy) (Albertinazzi et al., 1998). The pEYFP-N3 vector was from BD Biosciences Clontech,(Palo Alto, CA). Lentiviruses expressing ECFP-VAMP2 have been previously described(Bonanomi et al., 2005). Defective recombinant adenoviruses driving the simultaneousexpression of GFP and either HA-Pincher or HA-PincherG68E were made using the Ad-Easysystem (Valdez,et al, 2005). Adenoviruses expressing GFP have been produced at the AAVVector Unit (AVU) of the International Centre for Genetic Engineering and Biotechnology(ICGEB) Trieste, Italy as previously described (Zentilin et al., 2001).
RNA interferenceThe clathrin heavy chain Stealth (Invitrogen, Carlsbad, CA) efficiently knock-down clathrinin mouse cells (Lampugnani et al., 2006). Stealth were designed with the BLOCK-iT™ RNAiDesigner (Invitrogen, Carlsbad, CA), starting from Mus musculus clathrin, heavy polypeptideNM_001003908. The starting nucleotide is 1,284 bases downstream of the start codon (5’-3’sequence GAAGAACUCUUUGCCCGGAAAUUUA, antisenseUAAAUUUCCGGGCAAAGA-GUUCUUC). As a control, a Stealth siRNA-negative controlduplex oligonucleotide with a C/G content equivalent to the positive oligonucleotide was used(5’-3’ sequence GAAUCAUUCCGUGC-CAAGUAGAUUA, antisenseUAAUCUACUUGGCACGGAAUGAUUC). After dissection and trypsinization, mouseneurons were nucleofected as described below and cultured for 48 h before the experiments.Efficient silencing was verified by retrospective immunofluorescence.
Cell culture and Transduction of neuronsPrimary neuronal cultures were prepared from the hippocampi of either Sprague-Dawley E18rat embryos or E17 C57BL/6J mouse embryos (Charles River Italica, Calco, Italy) aspreviously described (Banker and Cowan, 1977). For infection, neurons were placed the dayafter plating in a clean dish containing glia-conditioned medium (MEM supplemented with1% N2 supplement (Invitrogen), 2 mM glutamine (Biowhittaker Inc., Biggs Ford Rd.Walkersville, MD), 0.1% ovalbumin, 1 mM sodium pyruvate (Sigma-Aldrich), and 4 mMglucose) and incubated for 10–15 h at 37° C in a 5% CO2 humidified atmosphere in the presenceof viral supernatant. After transduction, neurons were returned to the original dishes andmaintained in culture in glia-conditioned medium. In some experiments, rat neurons (~1.5 ×106 cells) were nucleofected in suspension immediately after harvesting with 3 µg of DNAusing the Basic Nucleofector Kit for primary neurons (Amaxa Biosystems, Cologne, Germany)(O-03 program). For RNAi experiments mouse neurons (~3 × 106 cells) were nucleofectedwith 3 µg of oligonucleotides together with 0.5 µg pEYFP-N3 plasmid using the O-05 program.After electroporation, neurons were diluted and plated at the standard low density.
Bonanomi et al. Page 3
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Cell labeling protocolsFor FM4-64 uptake experiments, neurons were incubated with FM4-64 (10 µM) diluted ineither KRH (basal medium) or KRH containing 55 mM KCl (depolarizing solution) for 1 minat room temperature (RT), rinsed three times by complete medium substitution with KRH overa course of 2 min and live imaged. When indicated, FM4-64 uptake was carried out at 37° C.Since fixation does not result in noticeable loss of FM4-64 fluorescence (Diefenbach et al.,1999), in several experiments neurons were fixed after loading with FM4-64 in order to obtainlarge numbers of growth cones for analysis. Fixation was performed for 15 min at RT with 4%paraformaldehyde, 4% sucrose in 120 mM sodium phosphate buffer (pH 7.4) supplementedwith 2 mM EGTA. In some cases, fixed neurons were processed for immunofluorescence aspreviously described (Menegon et al., 2002). As FM4-64 is lost upon cell permeabilization, acircle was inscribed on the bottom coverslip using a diamond-tip scribing objective (Zeiss,Oberkochen, Germany) and was used as a reference to relocate previously imaged growthcones. For loading of fluorescent polystyrene beads, neurons were incubated with 1 µl of anaqueous suspension containing 2% solids diluted in 1 ml KRH.
Videomicroscopy and Image AnalysisSpecimens were viewed with an Axiovert 135 inverted microscope (Zeiss, Oberkochen,Germany) equipped with epifluorescence optics. Images were recorded with a C4742-98ORCA II cooled charge-coupled device camera (Hamamatsu Photonics, Hamamatsu City,Japan) and processed using Image Pro Plus 4.5 (Media Cybernetics, Silver Spring, MD) andAdobe Photoshop 6.0 (Adobe System, San Jose, CA). For image deconvolution, Z-stacks ofoptical sections taken with an Olympus IX70 with DeltaVision RT Deconvolution System(Olympus, Hamburg, Germany) were analyzed by the WoRx Deconvolve software (AppliedPrecision, Issaquah, WA). In order to quantify the FM4-64 signal in individual growth cones,images were acquired at constant parameters of illumination and gain, and the specificfluorescence intensity, calculated by subtracting a fixed fluorescence background level fromthe total pixel intensity, was measured within the whole growth cone area determined by eitherdifferential interference contrast (DIC) or GFP/YFP fluorescence images. One-way ANOVAfollowed by either Dunnetts's or Tukey's multiple comparison tests were used and p values<0.05 were considered significant.
RESULTSThe axonal growth cone is a site of intense constitutive plasma membrane endocytosis
To study constitutive membrane recycling in the growth cone, we used rat hippocampal neuronsat 2–3 days in vitro (DIV) (stage 3; Dotti et al., 1988) displaying an unequivocally identifiableaxon tipped with a growth cone. Neurons incubated in basal medium (KRH) were exposed tothe styryl dye FM4-64, which binds reversibly to the outer surface of the plasma membraneand is internalized in endocytic vesicles (Betz and Bewick, 1992; Betz et al, 1996) (Fig. 1A).After brief (1 min) incubation, intense fluorescence appeared in 50–70% of growth cones,associated with large structures, often exhibiting a vacuolar organization, clustered at the distaledge of the C domain. In contrast, FM4-64 staining was undetectable in both the lamellar Pdomain of the growth cone and the axon shaft (Fig. 1A). The axonal specificity of this bulkendocytosis was confirmed by retrospective staining of the neurons with the markerdephospho-Tau1 (Fig 1B). Interestingly, at each axonal branching bulk endocytic accumulationof the dye was typically maintained in only one of the growth cones (Fig. 1C). Intense basalendocytosis in the C domain was also observed in neurons exposed for 1 min to either themembrane-impermeable fluid-phase fluorescent dye lucifer yellow or green fluorescentpolystyrene beads, 20 nm in diameter (Fig. 1A). A similar pattern of endocytosis was observedafter a 1 min-application of large fluid-phase markers, namely fluorescent beads, 200 nm indiameter, or 40 kDa dextran, which have restricted access to small vesicles (Fig. 1A and data
Bonanomi et al. Page 4
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

not shown). In growth cones labeled with the 20 nm beads and subsequently exposed toFM4-64, the two tracers displayed largely overlapping patterns of internalization. It should benoted that in dendritic cells dextran and small latex beads (< 100 nm) have been shown to bepreferentially internalized via macropinocytosis (Reece et al., 2001; Falcone et al., 2006; seeDiscussion). Consistently, we observed a partial degree of colocalization with rabankyrin-5, aRab5 effector that modulates macropinocytosis in non-neuronal cells (Fig. 3D) (Schnatwinkelet al., 2004).
The C domain of the growth cone is characterized by high density of organelles, includingendosomes and SVs. In order to investigate the possible involvement of these vesicles in bulkendocytosis we carried out retrospective staining of FM4-64-loaded growth cones with the SVmarker synaptobrevin 2/vesicle-associated membrane protein 2 (VAMP2) or the endosomalmarker syntaxin13. In either case, virtually no colocalization with the compartments ofconstitutive plasma membrane uptake was observed (Fig. 1D).
The fast, efficient uptake of 200 nm-diameter beads was suggestive of a clathrin-independentendocytic pathway (Rejman et al. 2004). To test this possibility, we performed an RNAi-mediated clathrin heavy chain (CHC) knock-down (Lampugnani et al. 2006, Granseth et al.2006) in cells later exposed to FM4-64 uptake (Fig. 2A,B). Effective down regulation of CHC,as assessed by retrospective immunofluorescence of growth cones, did not affect FM4-64uptake, indicating that the bulk plasma membrane retrieval was indeed clathrin-independent.Consistently, Alexa488-conjugated transferrin, a marker of receptor-mediated, clathrin-dependent endocytosis, was internalized into endosomes in the cell body but not in thecompartments of bulk FM4-64 uptake in the growth cone (Fig. 2C).
Constitutive FM4-64 uptake was prevented by incubation of neurons at 4° C, a treatment whichattenuates endocytic processes. In contrast, it was apparently unaffected by prolonged (30 min)incubation of neurons in Ca2+-free KRH, a maneuver leading to depletion of intracellularCa2+ stores (Cohen and Fields, 2006), and by a 1 h incubation in the presence of BFA, an ADP-ribosylation factor-1 inhibitor that prevents traffic from the Golgi complex (Lippincott-Schwartz et al., 1989; Miller et al.,1992) and blocks axon growth (Jareb and Banker, 1997)(Fig. 3A,C). FM4-64 internalized for 1 min at rest was progressively released during 30 minwashing (Fig. 3C and Fig. 5). In some instances, we obtained evidence for retrograde transportof dye-containing organelles along the axon (Supplementary Fig. 1). We conclude that, at earlydevelopmental stages, a rapid constitutive process of bulk plasma membrane internalization,independent of the SV and coated vesicle recycling, is associated with the axonal growth cone.
Constitutive plasma membrane endocytosis takes place at the sites of membrane rufflingand depends on F-actin, cholesterol and phosphatidylinositol-3 kinase
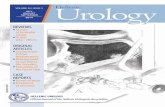
Time-lapse DIC imaging of growth cones of stage 3 neurons exposed to FM4-64 in KRH for1 min revealed a tight correlation between the sites of bulk constitutive endocytosis and thesites of plasma membrane ruffling, which is particularly intense in the T domain (see alsoForscher et al., 1992; Schaefer et al., 2002) (Fig. 3B).
In the T zone, the compartments of bulk FM4-64 uptake overlapped with F-actin rich rufflinghotspots (Fig. 3E). F-actin disruption by cytochalasin D, which impairs the addition of actinmonomers to filaments, significantly reduced basal FM4-64 uptake, indicating that actinpolymerization is required for constitutive plasma membrane retrieval in the growth cone (Fig.3A,C).
Phosphatidylinositol (PI) 3-kinase is localized at the tip of newly specified axons of stage 3neurons (Shi et al., 2003). In other cell types this enzyme has been implicated in high-volume
Bonanomi et al. Page 5
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

endocytosis (Lindmo and Stenmark, 2006). Application of the selective PI 3-kinase inhibitor,LY294002, significantly reduced constitutive FM4-64 uptake (Fig. 3A,C).
Staining of neurons with filipin, a fluorescent marker of cholesterol, revealed an enrichmentof this lipid in the C domain, partially overlapping with the compartments of bulk FM4-64internalization (Fig. 3F). Acute (3 min) cholesterol extraction with MβCD drastically affectedFM4-64 uptake in the growth cones (Fig. 3A,C).
F-actin disruption, PI 3-kinase inhibition and cholesterol depletion also inhibited theconstitutive internalization of 20 nm beads in the growth cone. Bead uptake was recovered 30min after MβCD washout, in parallel to the normalization of cholesterol levels (SupplementaryFig. 2).
Selectivity of bulk endocytosisWe investigated whether at the growth cone bulk endocytosis mediated unselectiveinternalization of plasma membrane proteins. Growth cones loaded with FM4-64 during a 1min-incubation in resting solution were immediately fixed and retrospectively stained for Trk,L1, β1 integrin or APP, four proteins that undergo recycling at the axonal plasma membrane(Yamazaki et al., 1995; Condic and Letourneau, 1997; Kamiguchi and Lemmon, 2000; Chenet al., 2005) (Fig 4). None of these membrane proteins colocalized with the FM4-64-positivecompartments.
Next, we monitored over time the internalization of FM4-64 applied to the same growth conetogether with another marker, Alexa488-conjugated cholera toxin B subunit (CTB), whichbinds the lipid raft component ganglioside GM1 (Fig. 5). In other cell types CTB is internalizedvia clathrin-coated pits, caveolae (Torgersen et al., 2001) or clathrin-independent pathways(Kirkham et al., 2005). After a 1 min-incubation, when FM4-64 was already internalized inlarge compartments at the T domain of the growth cone, CTB was still associated with theplasma membrane. During the following 30 min, fluorescent CTB was redistributed intopuncta, most probably corresponding to endocytic vesicles enriched at the T domain. At notime points did the pattern of CTB overlap with the FM4-64-positive compartments. Theseresults document the existence, at the growth cone, of one (or more) sorting processes whichexclude most specific plasma membrane components from the bulk endocytic organelles.
Rac1 controls bulk plasma membrane retrieval in the growth coneRac1 has been implicated in the stimulation of membrane ruffling and bulk fluid-phase uptake(i.e., macropinocytosis) in non-neuronal cells (Symons and Rusk, 2003). Thus, we reasonedthat Rac1 might participate in the process of constitutive plasma membrane internalization inthe growth cone. Neurons coexpressing soluble YFP and FLAG-tagged either constitutivelyactive or dominant negative versions of Rac1 (Rac1-N17 and Rac1-V17, respectively) wereexposed to FM4-64 in basal medium. The expression of Rac1 mutants in YFP-positive cellswas confirmed by retrospective staining with anti- FLAG antibody. Remarkably, expressionof dominant-negative Rac1 dramatically impaired bulk FM4-64 uptake in growth cones. Incontrast, the constitutively active Rac1 form did not affect FM4-64 internalization comparedto control growth cones expressing only soluble YFP (Fig. 6 and Supplementary Fig. 3). Ofnote, dye loading into smaller endocytic organelles was not prevented by Rac1-N17 expression.Rac1-N17 expression led to a reduction in growth cone size, which could result from acombined action of this protein on the actin cytoskeleton and membrane trafficking (see alsoKuhn et al., 1998; Ruchhoeft et al., 1999; Woo and Gomez, 2006).
Bonanomi et al. Page 6
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Constitutive bulk plasma membrane endocytosis is regulated by PincherPincher is the first regulator of a recently described process of macroendocytosis that underliesligandinduced internalization and signaling of neurotrophin receptors (Shao et al., 2002;Valdez et al., 2005; Valdez et al., 2007). The involvement of Pincher in constitutive bulk plasmamembrane retrieval in the growth cone was studied by infecting neurons with adenovirusesindependently driving expression of GFP and HA-tagged either wild-type or dominant-negative Pincher (Valdez et al., 2005). In a first set of experiments aimed at determining thelocalization of Pincher in the growth cone, we expressed the exogenous proteins at low levelsby reducing both virus concentration and infection times. Infected neurons were exposed for1 min to FM4-64 in resting medium and stained with anti-HA antibody to detect either HA-Pincher or HA-Pincher G68E (Fig. 7A). Pincher showed a partial colocalization with thecompartments of FM4-64 uptake. Similar results were obtained when constitutive endocytosiswas assayed in HA-Pincher-expressing growth cones by internalization of fluorescent beads(Fig. 7B). In contrast, Pincher G68E displayed a diffuse distribution throughout the growthcone, likely resulting from its irreversible binding to the plasma membrane (Shao et al.,2002; Fig. 7A). Next, we increased the expression levels of Pincher or Pincher G68E. In growthcones overexpressing either wild-type or the G68E Pincher mutant the constitutive FM4-64uptake was prevented compared to either uninfected neurons used as an internal control (Fig.7C) or neurons infected with adenoviruses expressing only GFP (Fig. 7D,E). As observed withRac1-N17, smaller endocytic organelles were still loaded with FM4-64 upon overexpressionof wild-type/G68E Pincher (Fig. 7D), pointing to the coexistence of at least two mechanisticallydistinct pathways of constitutive endocytosis in the growth cones.
Developmental control of plasma membrane endocytosis in the axonal growth coneTo investigate the development-related changes in the membrane recycling properties atgrowth cones, we took advantage of the stereotyped and well-characterized sequence of eventsfollowed by hippocampal neurons differentiating in culture (Dotti et al., 1988). The effects ongrowth cone endocytosis of a depolarizing solution (55 mM KCl) were first investigated atearly stages of neuronal differentiation (3 DIV). Growth cones were loaded for 1 min withFM4-64 in basal medium and subsequently exposed for an additional min to a high K+-containing solution devoid of FM4-64 prior to fixation and staining for VAMP2 (Fig. 8A).K+ application neither elicited unloading of the dye nor promoted membrane transfer from thecompartments of bulk endocytosis to the SV recycling pathway, as indicated by the lack ofFM4-64 in the VAMP2-positive SVs. In a second set of experiments, neurons infected withlentiviruses expressing ECFP-tagged VAMP2 (ECFP-VAMP2) were stimulated with highK+ for 1 min in the presence of FM4-64. The intensity and pattern of FM4-64 fluorescenceafter depolarization was remarkably similar to the pattern observed at this stage afterconstitutive uptake of the dye, being associated with large structures at the distal area of the Cdomain (Fig. 3Cand Fig. 8B, compare with Fig. 1). Importantly, also in this case no ECFP-VAMP2-positive SVs were labeled by the dye. These results indicate that at earlydevelopmental stages both bulk plasma membrane endocytosis and SV recycling areinsensitive to depolarizing stimuli.
Growth cones of ECFP-VAMP2-expressing neurons at 4–5 DIV (stage 4, characterized byprominent dendrite outgrowth; Dotti et al., 1988) were incubated in KRH containing FM4-64for 1 min. After a 15 min interval in basal medium (to unload the internalized dye) followedby photobleaching of the residual fluorescence, the neurons were exposed for 1 min to a highK+-containing solution containing FM4-64. The same growth cones where, under restingconditions, uptake FM4-64 occurred via bulk plasma membrane internalization separate fromSV recycling, after application of the depolarizing stimulus displayed a punctate patternselectively associated with endocytosis of VAMP2-positive SVs (Fig. 9A). Remarkably, bulkendocytosis ceased in growth cones during depolarizing stimuli which induced SV recycling.
Bonanomi et al. Page 7
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Neurons at 7 DIV display a complex dendritic arbor and a long axon with various branches(stage 5: full maturation; Dotti et al., 1988). Typically, the growth cone associated with themain axonal process has already encountered several potential targets, and synaptic contactshave started to be established. Moreover, at this developmental stage the distinctivemorphological compartmentalization and dynamics of growth cones are less obvious (ourunpublished observations). Upon 1 min exposure of 7 DIV neurons to KRH containingFM4-64, the axonal growth cone was largely devoid of FM4-64 labeling, whereas the dye wasinternalized throughout the soma and neuronal arborization in punctuate endocytic structuresnot overlapping with synaptic terminals (Fig. 9B and data not shown). When neurons at 7 DIVwere exposed to FM4-64 during a 1 min depolarization with high K+, the dye was loaded inECFP-VAMP2-positive SVs at both growth cones and synapses (Fig. 9C and data not shown).
The extent of the developmental suppression of bulk membrane retrieval was measured inneurons at 3, 5 and 7 DIV exposed to FM4-64 in basal medium (1 min) and subsequentlydepolarized to exclude any contribution of activity-dependent endocytic pathways. Thesestudies showed that bulk constitutive plasma membrane retrieval associated with growth conesis progressively down-regulated during neuronal differentiation (Fig. 9D and Fig. 10). Inaddition, SVs, which appear to be reluctant to undergo both spontaneous and evoked exo-endocytosis at early developmental stages, become competent for depolarization-inducedrecycling at later stages.
DISCUSSIONIn this study we identify a novel process of bulk plasma membrane retrieval as the mainpathway of constitutive endocytosis in growth cones at early stages of neuronal differentiation.Neither the classical endosomal system nor SVs participate in high-volume endocytosis of thegrowth cone plasmalemma, which involves large compartments associated with sites of intenseactin-based membrane ruffling. Our results provide an explanation for the previousidentification of the growth cone as the major site of axonal endocytosis (Sinclair et al.,1988; Parton et al., 1992; Zakharenko and Popov, 2000; Pfenninger and Maylié-Pfenninger,1981; Cheng and Reese, 1987). In addition, we have shown that basal endocytosis at the growthcone is subjected to developmental control, being down regulated at the onset ofsynaptogenesis, concomitantly with the appearance of depolarization-induced SV recycling.
Constitutive membrane retrieval in the growth cone appears to be mechanistically related tomacropinocytosis, a form of high-volume, clathrin-independent endocytosis described invarious cell types and particularly well-characterized in macrophages and dendritic cells(Swanson and Watts, 1995; Johannes and Lamaze, 2002; Kirkham and Parton, 2005).Macropinosomes are generated when membrane protrusions that form during actin-basedruffling fuse back with the plasma membrane trapping fluid-phase material. Strikingsimilarities between bulk membrane retrieval in the growth cone and macropinocytosis includethe association with membrane ruffling, the involvement of pleiomorphic endocytic structuresof large size, the dependence on actin dynamics, the requirement of PI3-kinase activity, thesensitivity to cholesterol depletion, and the involvement of the small GTPase Rac1 (Swansonand Watts, 1995; Johannes and Lamaze, 2002; Kirkham and Parton, 2005; Lindmo andStenmark, 2006). In the growth cone, the role of PI3-kinase and Rac1 in bulk plasma membraneretrieval is plausibly linked to the remodeling of the subcortical actin cytoskeleton, controlledby PI3-kinase via PI(3,4,5)P3 effectors, including the small GTPases Rac1, Cdc42 and Arf6(Govek et al., 2005; Lindmo and Stenmark, 2006). Yet, the compartments of bulk endocytosisin the growth cone do not overlap with the distribution of GM1, which is enriched inmacropinosomes in non-neuronal cells (Watarai et al, 2001), and show only partialcolocalization with rabankyrin-5 (Schnatwinkel et al., 2004). Of note, various integral proteinsof the growth cone plasma membrane are excluded from bulk endocytosis, in analogy with the
Bonanomi et al. Page 8
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

sorting process that accompanies macropinosome formation (Mercanti et al., 2006). Sorting isconceivably required to preserve the specific composition of both the plasma membrane andintracellular compartments.
Our results extend the role of the chaperone Pincher to the control of constitutive membraneretrieval at the growth cone. Pincher mediates ligand-stimulated, clathrin-independentendocytosis of neurotrophin receptors through macroendosomes generated at sites of plasmamembrane ruffling (Shao et al., 2002; Valdez et al., 2005 and 2007). Overexpression of eitherwild-type or dominant-negative Pincher interfered with bulk FM4-64 uptake. It is possible thatthe effect of high doses of wild-type Pincher reflects the exhaustion of basal endocytic activityfollowing an early phase of accelerated membrane retrieval that might have escaped detectionin our assay.
During development, an intrinsic program of maturation appears to contribute to changing theendocytic properties of growth cones (Fig. 10). Remarkably, depolarization of mature (stage4) growth cones induces selective internalization of the dye in SVs but not in the largecompartments labeled at rest. This situation is reminiscent of the spatial and temporaldissociation between bulk compensatory endocytosis and SV recycling previously reported atthe ribbon synapse of retinal bipolar cells (Holt et al., 2003). While the switch in the sensitivityof growth cones to depolarization is likely to reflect developmental changes in the Ca2+-sensingapparatus (Pravettoni et al., 2000;Menegon et al., 2002), dissociation of basal bulk endocytosisfrom depolarization-induced SV endocytosis could represent a way to uncouple majorrearrangements of the plasma membrane from ongoing SV recycling, thus preserving themolecular identity of the SV pool. Indeed, both bulk membrane endocytosis and SV recyclingtake place preferentially in the T zone of growth cones (Bonanomi et al., 2005).
The mechanism(s) targeted to occlude bulk membrane retrieval in mature neurons is (are)unknown. One attractive possibility is that during synaptogenesis spontaneous network activitygenerates ‘maturation’ signals that locally repress the PI3-kinase/Rac1/F-actin machinery inthe growth cone. This control may be exerted at different levels, for instance involving guanine-nucleotide exchange factors (GEFs) and/or GTPase activating proteins (GAPs), which tuneRac1 activity. A similar working model might explain the effect of depolarization on isolatedstage 4 growth cones. At this stage growth cones, which appear to have already acquired themolecular apparatus to sense developmental signals and repress bulk uptake, still lack actualmaturation inputs from the forming network. Interestingly, constitutive macropinocytosis,which is down regulated in mature dendritic cells that have initiated the processing of anacquired antigen, can be restored upon expression of active forms of Rac and Cdc42 (Garrettet al., 2000). In an alternative model, during neuronal differentiation bulk membrane retrievalmight be replaced by selective retrieval directed by factors, such as the neurotrophins, via asimilar Rac- and Pincher-dependent pathway (Valdez et al., 2007).
What is the role of constitutive bulk membrane retrieval in the growth cone? The absence ofspecific molecular markers and regulators hampers selective experimental manipulations ofthis endocytic pathway. The most parsimonious view is that the continuous uptake of plasmamembrane via a high-volume/high-capacity pathway counterbalances the addition of newmembrane at the axon tip, required for axon outgrowth, keeping the growth cone in a highlydynamic state necessary for navigation and response to attractive/repulsive cues. However,FM4-64 uptake is not affected by prolonged incubation of neurons in the presence of BFA,which arrests trafficking of Golgi-derived vesicles and axon elongation but not growth conemotility (Craig et al., 1995; Jareb and Banker, 1997). Hence, bulk constitutive endocytosisappears to reflect a local recycling activity of the growth cone, which can occur independentlyof the supply of new membrane from the cell body, contributing to homeostasis of the axonalplasma membrane.
Bonanomi et al. Page 9
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

The bulk endocytic process described in this study might provide the growth cone with areservoir of ready-to-use membrane to be added to the cell surface to rapidly reconfigure andupdate the plasma membrane make-up during pathfinding and establishment of contacts withtargets. In addition, it is possible that such bulk endocytosis enables the growth cone to sampleextracellular proteins in a non-specific fashion to be retrogradely transported to the cell body(von Bartheld, 2004). As growth cones undergo constant morphological remodeling duringguidance and target recognition (Mason and Erskine, 2000), and the axonal sensor apparatusneeds continuous renewal by insertion and removal of proteins to adjust its sensitivity toenvironmental cues (Vogt et al., 1996; van Horck et al., 2004), the new membrane retrievalpathway described here may power a highly efficient membrane remodeling mechanism thatcan be essential during the early stages of neural development.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
ACKNOWLEDGMENTSWe wish to thank Drs. Ivan de Curtis and Jacopo Meldolesi (S. Raffaele Scientific Institute/Vita-Salute University,Milano, Italy) for critical reading of the manuscript. We also thank Dr. Lorena Zentilin and the Telethon AVU CoreFacility for the generous gift of the GFP-adeno-associated virus and Dr. Ivan de Curtis for the Rac1 constructs. Thiswork was supported by grants from the Italian Ministry of University (Cofin 2005 and 2006 to F.V. and F.B.,respectively), from the CARIPLO Foundation and the Mariani Foundation for Infantile Neurology (to F.V. and F.B).The financial support of Telethon-Italy (grant n. GGP05134 to F.B and F.V.) is gratefully acknowledged.
REFERENCESAhmari SE, Buchanan J, Smith SJ. Assembly of presynaptic active zones from cytoplasmic transport
packets. Nat. Neurosci 2000;3:445–451. [PubMed: 10769383]Albertinazzi C, Gilardelli D, Paris S, Longhi R, de Curtis I. Overexpression of a neural-specific rho family
GTPase, cRac1B, selectively induces enhanced neuritogenesis and neurite branching in primaryneurons. J. Cell Biol 1998;142:815–825. [PubMed: 9700168]
Albertinazzi C, Za L, Paris S, de Curtis I. ADP-ribosylation factor 6 and a functional PIX/p95-APP1complex are required for Rac1B-mediated neurite outgrowth. Mol. Biol. Cell 2003;14:1295–1307.[PubMed: 12686588]
Ashery U, Penner R, Spira ME. Acceleration of membrane recycling by axotomy of cultured aplysianeurons. Neuron 1996;16:641–651. [PubMed: 8785061]
Banker GA, Cowan WM. Rat hippocampal neurons in dispersed cell culture. Brain Res 1977;126:397–342. [PubMed: 861729]
Betz WJ, Bewick GS. Optical analysis of synaptic vesicle recycling at the frog neuromuscular junction.Science 1992;255:200–203. [PubMed: 1553547]
Betz WJ, Mao F, Smith CB. Imaging exocytosis and endocytosis. Curr. Opin. Neurobiol 1996;6:365–371. [PubMed: 8794083]
Bonanomi D, Benfenati F, Valtorta F. Protein sorting in the synaptic vesicle life cycle. Prog. Neurobiol.2006
Bonanomi D, Menegon A, Miccio A, Ferrari G, Corradi A, Kao HT, Benfenati F, Valtorta F.Phosphorylation of synapsin I by cAMP-dependent protein kinase controls synaptic vesicle dynamicsin developing neurons. J. Neurosci 2005;25:7299–7308. [PubMed: 16093379]
Bray D. Surface movements during the growth of single explanted neurons. Proc. Natl. Acad. Sci. U. S.A 1970;65:905–910. [PubMed: 5266160]
Chen ZY, Ieraci A, Tanowitz M, Lee FS. A novel endocytic recycling signal distinguishes biologicalresponses of trk neurotrophin receptors. Mol. Biol. Cell 2005;16:5761–5772. [PubMed: 16207814]
Cheng TP, Reese TS. Polarized compartmentalization of organelles in growth cones from developingoptic tectum. J. Cell Biol 1985;101:1473–1480. [PubMed: 3930511]
Bonanomi et al. Page 10
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Cheng TP, Reese TS. Recycling of plasmalemma in chick tectal growth cones. J. Neurosci 1987;7:1752–1759. [PubMed: 3598645]
Chow I, Poo MM. Release of acetylcholine from embryonic neurons upon contact with muscle cell. J.Neurosci 1985;5:1076–1082. [PubMed: 3884749]
Cohen JE, Fields RD. CaMKII inactivation by extracellular ca(2+) depletion in dorsal root ganglionneurons. Cell Calcium 2006;39:445–454. [PubMed: 16519936]
Condic ML, Letourneau PC. Ligand-induced changes in integrin expression regulate neuronal adhesionand neurite outgrowth. Nature 1997;389:853–856.
Craig AM, Wyborski RJ, Banker G. Preferential addition of newly synthesized membrane protein ataxonal growth cones. Nature 1995;375:592–594. [PubMed: 7791876]
Dailey ME, Bridgman PC. Dynamics of the endoplasmic reticulum and other membranous organelles ingrowth cones of cultured neurons. J. Neurosci 1989;9:1897–1909. [PubMed: 2723756]
Dent EW, Gertler FB. Cytoskeletal dynamics and transport in growth cone motility and axon guidance.Neuron 2003;40:209–227. [PubMed: 14556705]
Diefenbach TJ, Guthrie PB, Stier H, Billups B, Kater SB. Membrane recycling in the neuronal growthcone revealed by FM1-43 labeling. J. Neurosci 1999;19:9436–9444. [PubMed: 10531447]
Dotti CG, Sullivan CA, Banker GA. The establishment of polarity by hippocampal neurons in culture. J.Neurosci 1988;8:1454–1468. [PubMed: 3282038]
Falcone S, Cocucci E, Podini P, Kirchhausen T, Clementi E, Meldolesi J. Macropinocytosis: Regulatedcoordination of endocytic and exocytic membrane traffic events. J. Cell. Sci 2006;19:4758–4769.[PubMed: 17077125]
Fletcher TL, Cameron P, De Camilli P, Banker G. The distribution of synapsin I and synaptophysin inhippocampal neurons developing in culture. J. Neurosci 1991;11:1617–1626. [PubMed: 1904480]
Forscher P, Lin CH, Thompson C. Novel form of growt h cone motility involving site-directed actinfilament assembly. Nature 1992;357:515–518. [PubMed: 1608453]
Forscher P, Smith SJ. Actions of cytochalasins on the organization of actin filaments and microtubulesin a neuronal growth cone. J. Cell Biol 1988;107:1505–1516. [PubMed: 3170637]
Fournier AE, Nakamura F, Kawamoto S, Goshima Y, Kalb RG, Strittmatter SM. Semaphorin3A enhancesendocytosis at sites of receptor-F-actin colocalization during growth cone collapse. J. Cell Biol2000;149:411–422. [PubMed: 10769032]
Garrett WS, Chen LM, Kroschewski R, Ebersold M, Turley S, Trombetta S, Galan JE, Mellman I.Developmental control of endocytosis in dendritic cells by Cdc42. Cell 2000;102:325–334. [PubMed:10975523]
Govek EE, Newey SE, Van Aelst L. The role of the rho GTPases in neuronal development. Genes Dev2005;19:1–49. [PubMed: 15630019]
Granseth B, Odermatt B, Royle SJ, Lagnado L. Clathrin-mediated endocytosis is the dominant mechanismof vesicle retrieval at hippocampal synapses. Neuron 2006;51:773–786. [PubMed: 16982422]
Holt M, Cooke A, Wu MM, Lagnado L. Bulk membrane retrieval in the synaptic terminal of retinalbipolar cells. J. Neurosci 2003;23:1329–1339. [PubMed: 12598621]
Jareb M, Banker G. Inhibition of axonal growth by brefeldin A in hippocampal neurons in culture. J.Neurosci 1997;17:8955–8963. [PubMed: 9364043]
Johannes L, Lamaze C. Clathrin-dependent or not: Is it still the question? Traffic 2002;3:443–451.[PubMed: 12047552]
Jurney WM, Gallo G, Letourneau PC, McLoon SC. Rac1-mediated endocytosis during ephrin-A2- andsemaphorin 3A-induced growth cone collapse. J. Neurosci 2002;22:6019–6028. [PubMed:12122063]
Kamiguchi H, Lemmon V. Recycling of the cell adhesion molecule L1 in axonal growth cones. J. Neurosci2000;20:3676–3686. [PubMed: 10804209]
Kim YT, Wu CF. Reversible blockage of neurite development and growth cone formation in neuronalcultures of a temperature-sensitive mutant of drosophila. J. Neurosci 1987;7:3245–3255. [PubMed:3117981]
Bonanomi et al. Page 11
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Kirkham M, Fujita A, Chadda R, Nixon SJ, Kurzchalia TV, Sharma DK, Pagano RE, Hancock JF, MayorS, Parton RG. Ultrastructural identification of uncoated caveolin-independent early endocyticvehicles. J. Cell Biol 2005;168:465–476. [PubMed: 15668297]
Kirkham M, Parton RG. Clathrin-independent endocytosis: New insights into caveolae and non-caveolarlipid raft carriers. Biochim. Biophys. Acta 2005;1745:273–286. [PubMed: 16046009]
Kuhn TB, Brown MD, Bamburg JR. Rac1-dependent actin filament organization in growth cones isnecessary for beta1-integrin-mediated advance but not for growth on poly-D-lysine. J. Neurobiol1998;37:524–540. [PubMed: 9858256]
Lampugnani MG, Orsenigo F, Gagliani MC, Tacchetti C, Dejana E. Vascular endothelial cadherincontrols VEGFR-2 internalization and signaling from intracellular compartments. J. Cell Biol2006;174:593–604. [PubMed: 16893970]
Leoni C, Menegon A, Benfenati F, Toniolo D, Pennuto M, Valtorta F. Neurite extension occurs in theabsence of regulated exocytosis in PC12 subclones. Mol. Biol. Cell 1999;10:2919–2931. [PubMed:10473636]
Lindmo K, Stenmark H. Regulation of membrane traffic by phosphoinositide 3-kinases. J. Cell. Sci2006;119:605–614. [PubMed: 16467569]
Lippincott-Schwartz J, Yuan LC, Bonifacino JS, Klausner RD. Rapid redistribution of golgi proteins intothe ER in cells treated with brefeldin A: Evidence for membrane cycling from golgi to ER. Cell1989;56:801–813. [PubMed: 2647301]
Lockerbie RO, Miller VE, Pfenninger KH. Regulated plasmalemmal expansion in nerve growth cones.J. Cell Biol 1991;112:1215–1227. [PubMed: 1999470]
Mason C, Erskine L. Growth cone form, behavior, and interactions in vivo: Retinal axon pathfinding asa model. J. Neurobiol 2000;44:260–270. [PubMed: 10934327]
Matteoli M, Coco S, Schenk U, Verderio C. Vesicle turnover in developing neurons: How to build apresynaptic terminal. Trends Cell Biol 2004;14:133–140. [PubMed: 15003622]
Menegon A, Verderio C, Leoni C, Benfenati F, Czernik AJ, Greengard P, Matteoli M, Valtorta F. Spatialand temporal regulation of Ca2+/calmodulin-dependent protein kinase II activity in developingneurons. J. Neurosci 2002;22:7016–7026. [PubMed: 12177199]
Mercanti V, Charette SJ, Bennett N, Ryckewaert JJ, Letourneur F, Cosson P. Selective membraneexclusion in phagocytic and macropinocytic cups. J. Cell. Sci 2006;119:4079–4087. [PubMed:16968738]
Miller SG, Carnell L, Moore HH. Post-golgi membrane traffic: Brefeldin A inhibits export from distalgolgi compartments to the cell surface but not recycling. J. Cell Biol 1992;118:267–283. [PubMed:1629235]
Mundigl O, Ochoa GC, David C, Slepnev VI, Kabanov A, De Camilli P. Amphiphysin I antisenseoligonucleotides inhibit neurite outgrowth in cultured hippocampal neurons. J. Neurosci 1998;18:93–103. [PubMed: 9412489]
Parton RG, Simons K, Dotti CG. Axonal and dendritic endocytic pathways in cultured neurons. J. CellBiol 1992;119:123–137. [PubMed: 1527164]
Pfenninger KH, Maylie-Pfenninger MF. Lectin labeling of sprouting neurons. II. relative movement andappearance of glycoconjugates during plasmalemmal expansion. J. Cell Biol 1981;89:547–559.[PubMed: 7251665]
Pravettoni E, Bacci A, Coco S, Forbicini P, Matteoli M, Verderio C. Different localizations and functionsof L-type and N-type calcium channels during development of hippocampal neurons. Dev. Biol2000;227:581–594. [PubMed: 11071776]
Reece JC, Vardaxis NJ, Marshall JA, Crowe SM, Cameron PU. Uptake of HIV and latex particles byfresh and cultured dendritic cells and monocytes. Immunol. Cell Biol 2001;79:255–263. [PubMed:11380679]
Rejman J, Oberle V, Zuhorn IS, Hoekstra D. Size-dependent internalization of particles via the pathwaysof clathrin- and caveolae-mediated endocytosis. Biochem. J 2004;377:159–169. [PubMed:14505488]
Ruchhoeft ML, Ohnuma S, McNeill L, Holt CE, Harris WA. The neuronal architecture of xenopus retinalganglion cells is sculpted by rho-family GTPases in vivo. J. Neurosci 1999;19:8454–8463. [PubMed:10493746]
Bonanomi et al. Page 12
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Schaefer AW, Kabir N, Forscher P. Filopodia and actin arcs guide the assembly and transport of twopopulations of microtubules with unique dynamic parameters in neuronal growth cones. J. Cell Biol2002;158:139–152. [PubMed: 12105186]
Schnatwinkel C, Christoforidis S, Lindsay MR, Uttenweiler-Joseph S, Wilm M, Parton RG, Zerial M.The Rab5 effector rabankyrin-5 regulates and coordinates different endocytic mechanisms. PLoSBiol 2004;2:E261. [PubMed: 15328530]
Shao Y, Akmentin W, Toledo-Aral JJ, Rosenbaum J, Valdez G, Cabot JB, Hilbush BS, Halegoua S.Pincher, a pinocytic chaperone for nerve growth factor/TrkA signaling endosomes. J. Cell Biol2002;157:679–691. [PubMed: 12011113]
Shi SH, Jan LY, Jan YN. Hippocampal neuronal polarity specified by spatially localized mPar3/mPar6and PI 3-kinase activity. Cell 2003;112:63–75. [PubMed: 12526794]
Sinclair GI, Baas PW, Heidemann SR. Role of microtubules in the cytoplasmic compartmentation ofneurons. II. endocytosis in the growth cone and neurite shaft. Brain Res 1988;450:60–68. [PubMed:2900046]
Swanson JA, Watts C. Macropinocytosis. Trends Cell Biol 1995;5:424–428. [PubMed: 14732047]Symons M, Rusk N. Control of vesicular trafficking by rho GTPases. Curr. Biol 2003;13:R409–R418.
[PubMed: 12747855]Torgersen ML, Skretting G, van Deurs B, Sandvig K. Internalization of cholera toxin by different
endocytic mechanisms. J. Cell. Sci 2001;114:3737–3747. [PubMed: 11707525]Torre E, McNiven MA, Urrutia R. Dynamin 1 antisense oligonucleotide treatment prevents neurite
formation in cultured hippocampal neurons. J. Biol. Chem 1994;269:32411–32417. [PubMed:7798241]
Tsui HC, Ris H, Klein WL. Ultrastructural networks in growth cones and neuritis of cultured centralnervous system neurons. Proc. Natl. Acad. Sci. U. S. A 1983;80:5779–5783. [PubMed: 6577454]
Valdez G, Akmentin W, Philippidou P, Kuruvilla R, Ginty DD, Halegoua S. Pincher-mediatedmacroendocytosis underlies retrograde signaling by neurotrophin receptors. J. Neurosci2005;25:5236–5247. [PubMed: 15917464]
Valdez G, Philippidou P, Rosenbaum J, Akmentin W, Shao Y, Halegoua S. Trk-signaling endosomes aregenerated by rac-dependent macroendocytosis. Proc. Natl. Acad. Sci. U. S. A 2007;104:12270–12275. [PubMed: 17640889]
van Horck FP, Weinl C, Holt CE. Retinal axon guidance: Novel mechanisms for steering. Curr. Opin.Neurobiol 2004;14:61–66. [PubMed: 15018939]
Verhage M, Maia AS, Plomp JJ, Brussaard AB, Heeroma JH, Vermeer H, Toonen RF, Hammer RE, vanden Berg TK, Missler M, et al. Synaptic assembly of the brain in the absence of neurotransmittersecretion. Science 2000;287:864–869. [PubMed: 10657302]
Vogt L, Giger RJ, Ziegler U, Kunz B, Buchstaller A, Hermens WTJMC, Kaplitt MG, Rosenfeld MR,Pfaff DW, Verhaagen J, et al. Continuous renewal of the axonal pathway sensor apparatus by insertionof new sensor molecules into the growth cone membrane. Curr. Biol 1996;6:1153–1158. [PubMed:8805367]
von Bartheld CS. Axonal transport and neuronal transcytosis of trophic factors, tracers, and pathogens.J. Neurobiol 2004;58:295–314. [PubMed: 14704960]
Watarai M, Derre I, Kirby J, Growney JD, Dietrich WF, Isberg RR. Legionella pneumophila isinternalized by a macropinocytotic uptake pathway controlled by the Dot/Icm system and the mouseLgn1 locus. J. Exp. Med 2001;194:1081–1096. [PubMed: 11602638]
Woo S, Gomez TM. Rac1 and RhoA promote neurite outgrowth through formation and stabilization ofgrowth cone point contacts. J. Neurosci 2006;26:1418–1428. [PubMed: 16452665]
Yamada KM, Spooner BS, Wessells NK. Ultrastructure and function of growth cones and axons ofcultured nerve cells. J. Cell Biol 1971;49:614–635. [PubMed: 4326456]
Yamazaki T, Selkoe DJ, Koo EH. Trafficking of cell surface beta-amyloid precursor protein: Retrogradeand transcytotic transport in cultured neurons. J. Cell Biol 1995;129:431–442. [PubMed: 7721945]
Zakharenko S, Popov S. Plasma membrane recycling and flow in growing neurites. Neuroscience2000;97:185–194. [PubMed: 10771350]
Bonanomi et al. Page 13
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Zentilin L, Marcello A, Giacca M. Involvement of cellular double-stranded DNA break binding proteinsin processing of the recombinant adeno-associated virus genome. J. Virol 2001;75:12279–12287.[PubMed: 11711618]
Bonanomi et al. Page 14
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Constitutive bulk membrane retrieval in growth cones is distinct from the endosomal andsynaptic vesicle recycling pathways(A) (a–b’) Basal uptake of FM4-64 (red in a’ and b’) in neurons at 2 DIV incubated with KRHcontaining the dye and imaged after a 2-min washing. (a and a’) A broad region of intenseFM4-64 staining is associated with the growth cone (arrowhead) while few dispersed punctaappear in the cell body (asterisk). Note the lack of FM4-64 internalization along the axon shaft.(b and b’) High magnification of a growth cone showing internalization of FM4-64 in largeorganelles at the interface between the central and peripheral domains (T domain). Note thevacuolar organization of the FM4-64 positive compartment in b. (c–e) Constitutive endocytosisin growth cones probed by exposure to various tracers (green) for 1 min in KRH: lucifer yellow(4 mg/ml) (c), 200 nm-beads (d), 20 nm-beads (e). FM4-64 applied during a subsequentincubation is internalized in bead-containing compartments (right panel in e; red). The overlaybetween the corresponding fluorescence and DIC images is shown in a’, b’, c–e. (B) Basaluptake of FM4-64 (red) in neurons incubated with the dye for 1 min, fixed and retrospectivelystained for the axon specific marker dephospho-Tau-1 (green). Intense FM4-64 uptake is
Bonanomi et al. Page 15
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

visible in growth cones of Tau-1-positive axons (asterisks) but not in Tau-1-negativeperspective dendrites (arrowheads). (C) Differential uptake of FM 4–64 (red) in growth conesassociated with distinct axonal branches of the same neuron. One of the growth cones(arrowhead) shows strong constitutive endocytosis while the other (asterisk) is inactive. (D)Basal uptake of FM 4–64 (red in the merge image) in a growth cone incubated with the dyefor 1 min, fixed and retrospectively labeled for VAMP2 (blue in the merge image) and syntaxin13 (green in the merge image). The overlay between the fluorescence and DIC images is shownin the merge image. The bulk of FM4–64 fluorescence colocalizes with neither VAMP2 norsyntaxin 13, although occasional puncta of overlap are observed in more proximal regions(arrowheads).Bar, 13.5 µm in a, a’; 5µm in b, b’; 25 µm in B; 10 µm in C. 10 µm in c, d, e and D.
Bonanomi et al. Page 16
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Inhibition of clathrin-mediated endocytosis does not impair bulk plasma membraneretrieval(A) Depletion of the clathrin heavy chain (CHC) in mouse hippocampal neurons by RNAi.Representative images of neurons cotransfected with YFP and either control Stealth (upperpanels) or CHC Stealth (bottom panels). After basal FM4–64 uptake (red), 3 DIV mouseneurons were fixed and retrospectively immunostained to assess clathrin down-regulation(green). (B) Quantification of the effect of CHC RNAi on bulk plasma membrane endocytosis.A five-fold reduction of CHC immunoreactivity (green bars; mean ± SEM; n=20 growth conesper treatment) in individual growth cones is observed following RNAi. FM4–64 bulkendocytosis (red bars) is unchanged (* p < 0.001, Student’s t test, treated vs. control). (C)Neuron exposed to Alexa488-conjugated transferrin (green in the merge image) for 1 h andsubsequently to FM4–64 (red in the merge im age) for 1 min in KRH. Transferrin, primarilyinternalized in the cell body (asterisk), does not overlap with FM4–64 endocytosed in thegrowth cone (arrowhead).Bar, 10 µm in A; 25 µm in C.
Bonanomi et al. Page 17
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
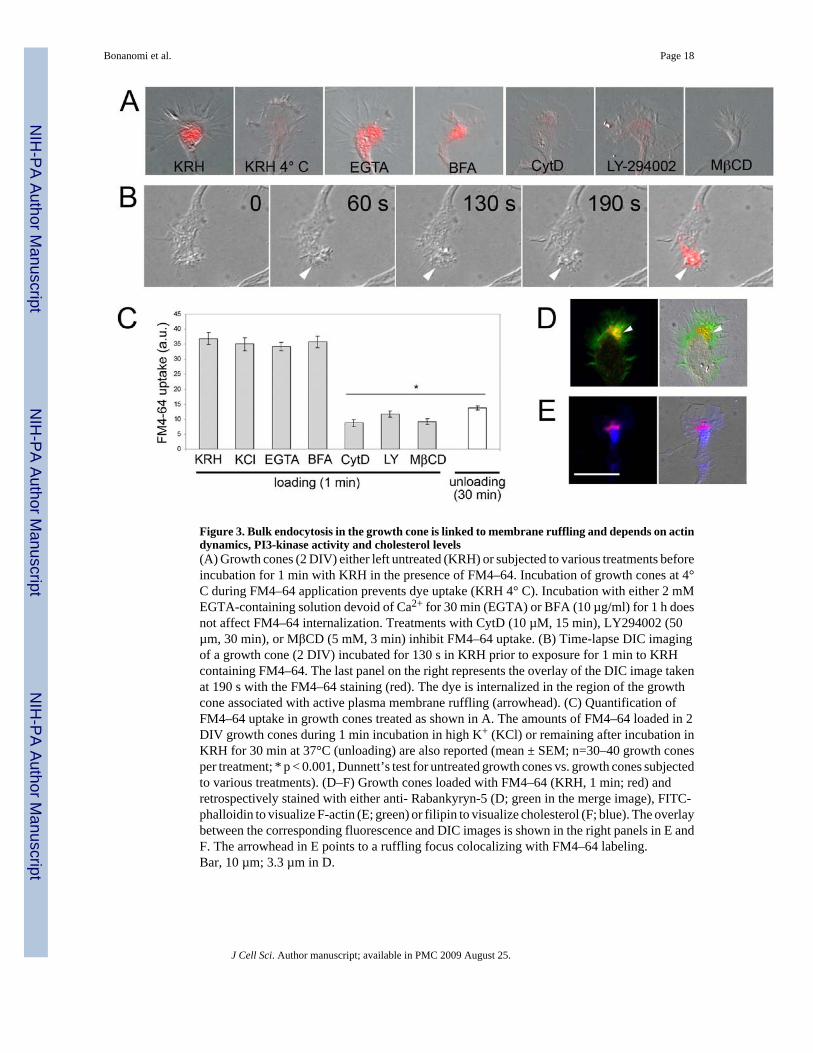
Figure 3. Bulk endocytosis in the growth cone is linked to membrane ruffling and depends on actindynamics, PI3-kinase activity and cholesterol levels(A) Growth cones (2 DIV) either left untreated (KRH) or subjected to various treatments beforeincubation for 1 min with KRH in the presence of FM4–64. Incubation of growth cones at 4°C during FM4–64 application prevents dye uptake (KRH 4° C). Incubation with either 2 mMEGTA-containing solution devoid of Ca2+ for 30 min (EGTA) or BFA (10 µg/ml) for 1 h doesnot affect FM4–64 internalization. Treatments with CytD (10 µM, 15 min), LY294002 (50µm, 30 min), or MβCD (5 mM, 3 min) inhibit FM4–64 uptake. (B) Time-lapse DIC imagingof a growth cone (2 DIV) incubated for 130 s in KRH prior to exposure for 1 min to KRHcontaining FM4–64. The last panel on the right represents the overlay of the DIC image takenat 190 s with the FM4–64 staining (red). The dye is internalized in the region of the growthcone associated with active plasma membrane ruffling (arrowhead). (C) Quantification ofFM4–64 uptake in growth cones treated as shown in A. The amounts of FM4–64 loaded in 2DIV growth cones during 1 min incubation in high K+ (KCl) or remaining after incubation inKRH for 30 min at 37°C (unloading) are also reported (mean ± SEM; n=30–40 growth conesper treatment; * p < 0.001, Dunnett’s test for untreated growth cones vs. growth cones subjectedto various treatments). (D–F) Growth cones loaded with FM4–64 (KRH, 1 min; red) andretrospectively stained with either anti- Rabankyryn-5 (D; green in the merge image), FITC-phalloidin to visualize F-actin (E; green) or filipin to visualize cholesterol (F; blue). The overlaybetween the corresponding fluorescence and DIC images is shown in the right panels in E andF. The arrowhead in E points to a ruffling focus colocalizing with FM4–64 labeling.Bar, 10 µm; 3.3 µm in D.
Bonanomi et al. Page 18
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
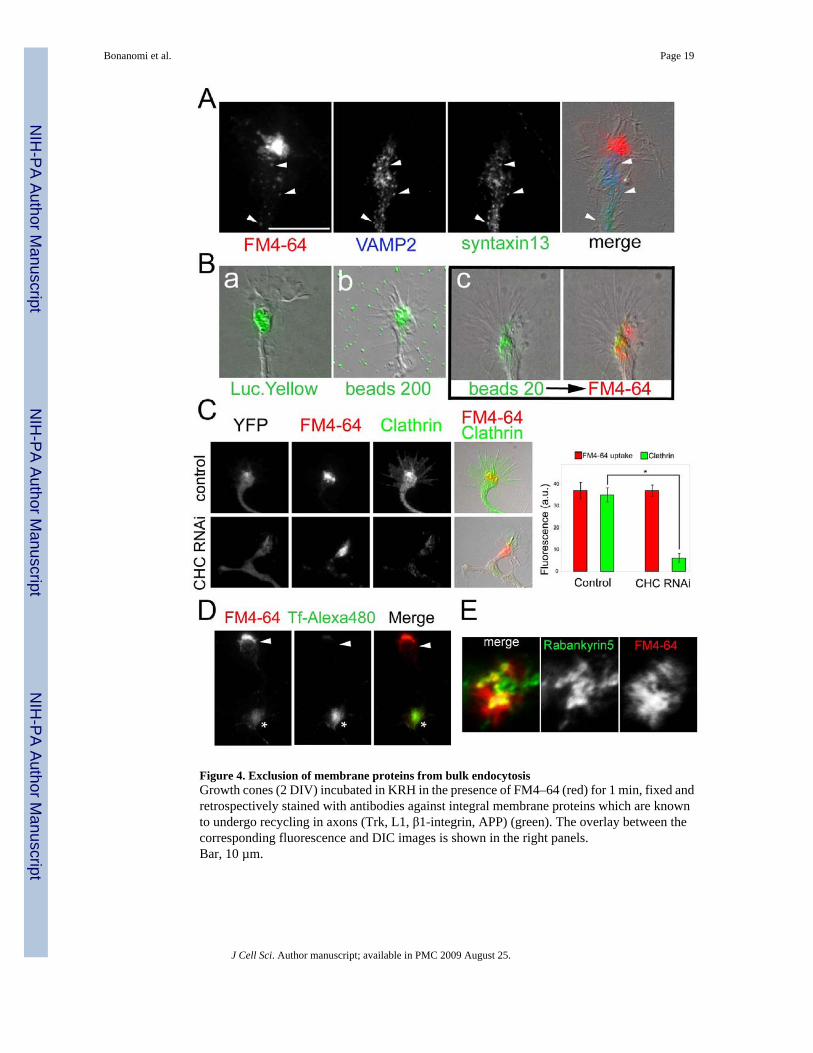
Figure 4. Exclusion of membrane proteins from bulk endocytosisGrowth cones (2 DIV) incubated in KRH in the presence of FM4–64 (red) for 1 min, fixed andretrospectively stained with antibodies against integral membrane proteins which are knownto undergo recycling in axons (Trk, L1, β1-integrin, APP) (green). The overlay between thecorresponding fluorescence and DIC images is shown in the right panels.Bar, 10 µm.
Bonanomi et al. Page 19
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Figure 5. The pathway of bulk membrane retrieval is distinct from the pathway of cholera toxininternalizationGrowth cone (2 DIV) exposed simultaneously to FM4–64 (red in the merge images) andAlexa488-conjugated cholera toxin subunit B (CTB; green in the merge images) for 1 min inKRH, washed and imaged during the following 30 min at 37° C. Immediately after loading(t=0) FM4–64 is internalized in large compartments while CTB is still associated with theplasma membrane and is subsequently endocytosed in puncta not overlapping with FM4-64labeling.Bar, 10 µm.
Bonanomi et al. Page 20
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
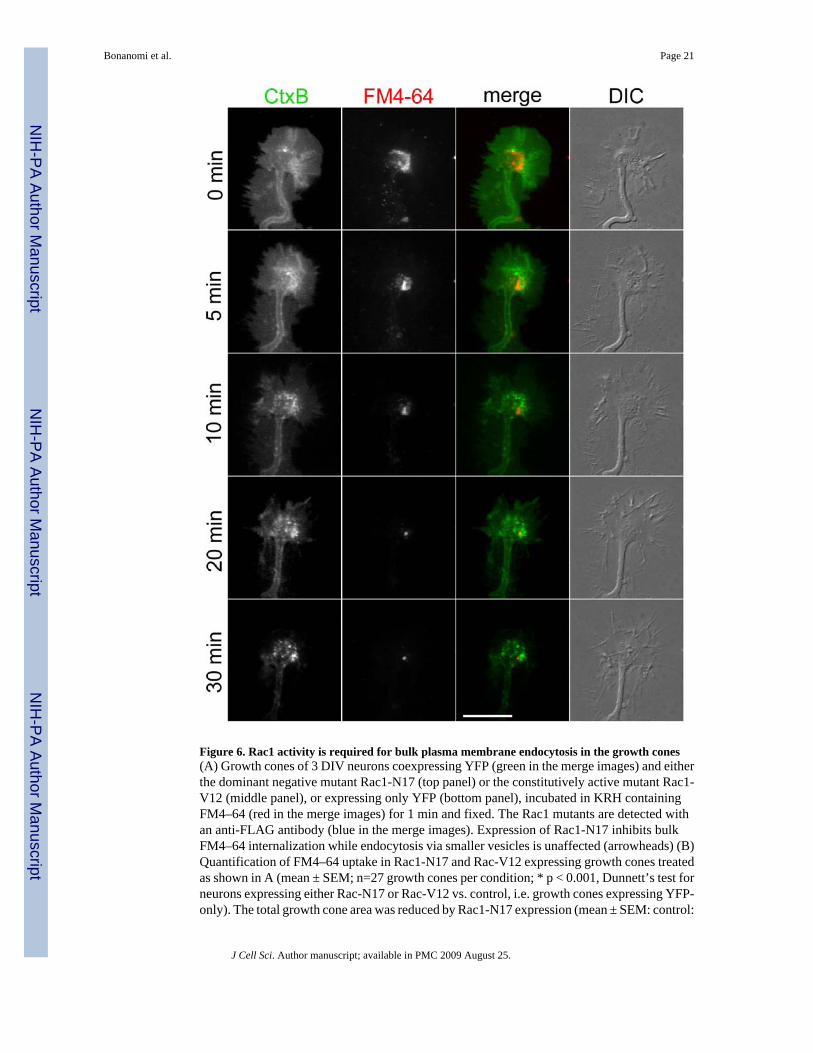
Figure 6. Rac1 activity is required for bulk plasma membrane endocytosis in the growth cones(A) Growth cones of 3 DIV neurons coexpressing YFP (green in the merge images) and eitherthe dominant negative mutant Rac1-N17 (top panel) or the constitutively active mutant Rac1-V12 (middle panel), or expressing only YFP (bottom panel), incubated in KRH containingFM4–64 (red in the merge images) for 1 min and fixed. The Rac1 mutants are detected withan anti-FLAG antibody (blue in the merge images). Expression of Rac1-N17 inhibits bulkFM4–64 internalization while endocytosis via smaller vesicles is unaffected (arrowheads) (B)Quantification of FM4–64 uptake in Rac1-N17 and Rac-V12 expressing growth cones treatedas shown in A (mean ± SEM; n=27 growth cones per condition; * p < 0.001, Dunnett’s test forneurons expressing either Rac-N17 or Rac-V12 vs. control, i.e. growth cones expressing YFP-only). The total growth cone area was reduced by Rac1-N17 expression (mean ± SEM: control:
Bonanomi et al. Page 21
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

44 ± 2.5 µm2; Rac1-N17: 35 ± 2.6 µm2; p<0.05, Dunnett’s test vs. control), but not by Rac1-V17 expression (mean ± SEM: 42 ± 2.7 µm2).Bar, 10 µm.
Bonanomi et al. Page 22
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 7. The marker of neuronal macroendocytosis Pincher regulates constitutive plasmamembrane retrieval at the growth comeGrowth cones of neurons at 3 DIV infected with adenoviruses driving the simultaneousexpression of GFP and either HA-Pincher or HA-Pincher G68E. (A) Growth cones expressinglow levels of HA-Pincher or HA-Pincher G68E stained with anti-HA antibody (green) afterbasal FM4–64 (red) uptake (1 min) to reveal the distribution of the exogenous proteins. Theoverlay between the fluorescence and DIC images is shown in the right panels (B) Growthcone expressing HA-Pincher at low levels exposed to 20 nm beads (red in the merge image)for 1 min in KRH, fixed and stained with anti-HA antibody (green in the merge image). Theimages are maximal projections of deconvoluted Z-stacks. Pincher overlaps with thecompartments of bulk endocytosis. (C and D) Basal FM4–64 uptake (1 min) in growth conesoverexpressing GFP and either HA-Pincher or HA-Pincher G68E, or infected withadenoviruses expressing only GFP (control). C shows the overlay between the fluorescenceand DIC images of a HA-Pincher/GFP expressing growth cone (arrowhead) and of anuninfected growth cone (asterisk) located in the same field of view. Overexpression of eitherPincher or the dominant negative mutant Pincher G68E inhibits constitutive bulk plasmamembrane uptake, while endocytosis via smaller vesicles is unaffected (arrowheads in D). (E)Quantification of FM4–64 uptake in HA-Pincher or HA-Pincher G68E expressing growth
Bonanomi et al. Page 23
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

cones (mean ± SEM; n=50 growth cones per condition; * p < 0.001, Dunnett’s test vs. control,growth cones expressing only GFP). Expression of either mutants does not affect the totalgrowth cone area (p<0.05, Dunnett’s test vs. control).Bar, 10 µm in A, C, D; 3.3 µm in B.
Bonanomi et al. Page 24
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
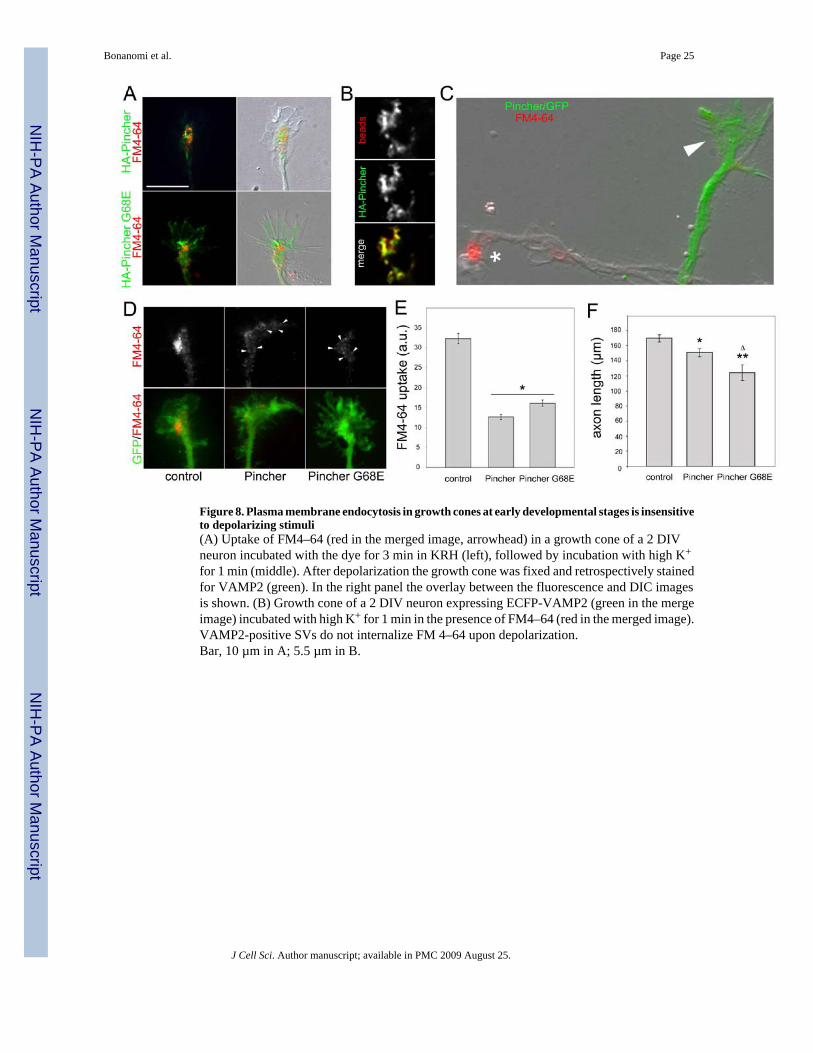
Figure 8. Plasma membrane endocytosis in growth cones at early developmental stages is insensitiveto depolarizing stimuli(A) Uptake of FM4–64 (red in the merged image, arrowhead) in a growth cone of a 2 DIVneuron incubated with the dye for 3 min in KRH (left), followed by incubation with high K+
for 1 min (middle). After depolarization the growth cone was fixed and retrospectively stainedfor VAMP2 (green). In the right panel the overlay between the fluorescence and DIC imagesis shown. (B) Growth cone of a 2 DIV neuron expressing ECFP-VAMP2 (green in the mergeimage) incubated with high K+ for 1 min in the presence of FM4–64 (red in the merged image).VAMP2-positive SVs do not internalize FM 4–64 upon depolarization.Bar, 10 µm in A; 5.5 µm in B.
Bonanomi et al. Page 25
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 9. Developmental control of endocytic activities in the growth cone(A) Growth cone (DIC, left) of neuron at 5 DIV expressing ECFP-VAMP2 (green) subjectedto two 15 min-spaced applications of FM4–64 (red) for 1 min in either KRH (first application,middle) or high K+ (second application, right). The depolarizing step induces FM4–64 loadingin VAMP2-positive SVs (arrowheads) with a pattern markedly different from that ofconstitutive endocytic compartments. (B) Neurons at 7 DIV incubated with FM4–64 for 1 minin KRH. At this stage, constitutive FM4–64 uptake is not associated with growth cones(asterisk), whereas is present in punctate endocytic compartments along the neurite network.(C) Growth cone of a 7 DIV neuron expressing ECFP-VAMP2 (green) incubated with highK+ for 1 min in the presence of FM4–64 (red). Arrowheads point to VAMP2-positive SVs thatinternalized the dye upon depolarization. (D) Quantification of basal FM4–64 uptake in growthcones of 3, 5, and 7 DIV neurons incubated with FM4–64/KRH for 1 min, rapidly washed andincubated in high K+ for 1 min. Bulk endocytosis is suppressed at later developmental stages(mean ± SEM; n=30–40 growth cones per treatment). At all time points, FM4–64 uptake wassignificantly different (p < 0.001, Tukey’s test).Bar, 7 µm in A and C; 12.5 µm in B, left; 7.5 µm in B, right.
Bonanomi et al. Page 26
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 10. Developmental changes in the endocytic properties of the growth coneTop: At early stages of differentiation (3 DIV), constitutive high-capacity plasma membranerecycling occurs via a bulk endocytic pathway mediated by large compartments (red) in the Tdomain of the growth cone. SV precursors (white circles), which at these stages are reluctantto both constitutive and evoked recycling, do not participate in the bulk retrieval process.Middle: As neuronal differentiation proceeds (4–5 DIV), the intensity of the bulk recyclingprocess decreases (pink), concomitantly with the emergence of activity-dependent SVrecycling (green circles) in both the growth cone and nascent synaptic structures. Bottom: Bulkplasma membrane retrieval ceases at later stages of neuronal differentiation (7 DIV), whenfrequent interactions of the growth cone with targets result in the establishment of synaptic
Bonanomi et al. Page 27
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

contacts associated with robust activity-dependent SV recycling. Exo-endocytosis of SVs ismaintained also in the growth cone. The dissociation of bulk endocytosis from the SV recyclingpathway at the onset of synaptogenesis may preserve the molecular identity of the formingpool of SVs and the stability of nascent synaptic structures.
Bonanomi et al. Page 28
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
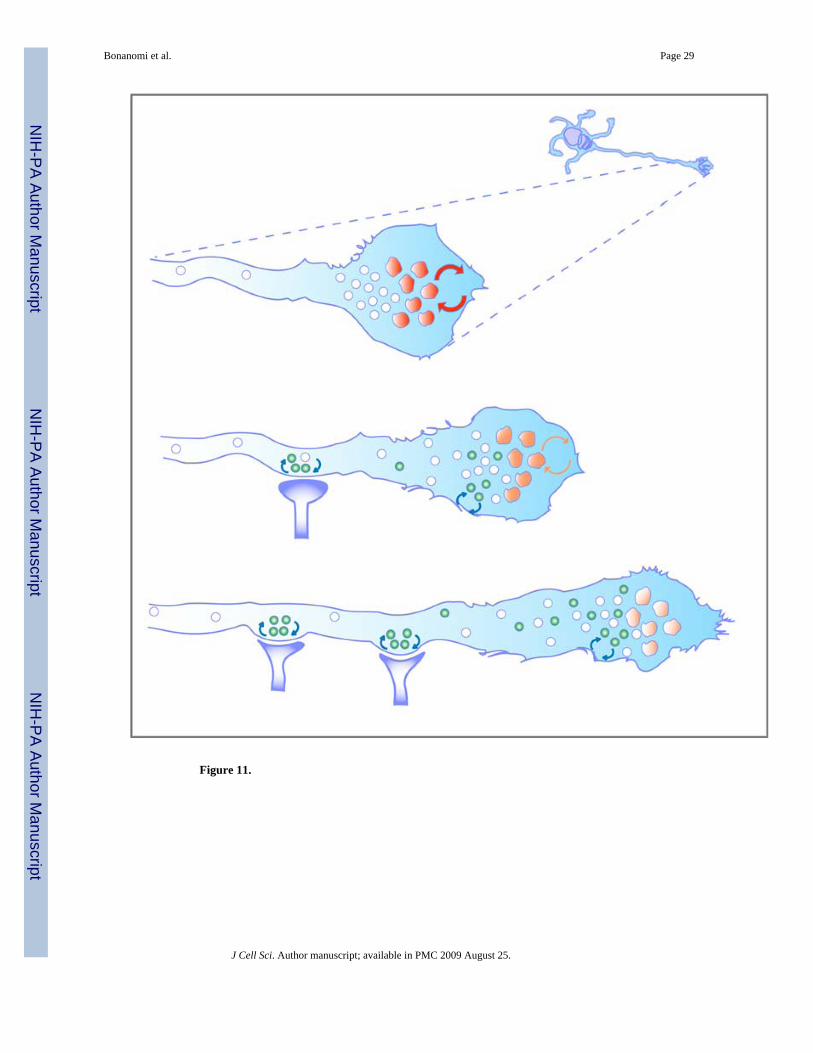
Figure 11.
Bonanomi et al. Page 29
J Cell Sci. Author manuscript; available in PMC 2009 August 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Copyright © 2022 FDOKUMEN