
The Role of Dispositional Reappraisal in the Age-Related Positivity Effect

Expression Study on D123 Gene Product:Evidence for High Positivity in Testis
Maurizio Onisto,1 Paola Zeilante, Paolo Scannapieco, Donatella Pellati, Monica Pozza,*Cristina Caenazzo, Alessandro Negro,† and Spiridione Garbisa
Institute of Histology and Embryology, Medical School, and †Department of Biological Chemistry, University of Padua, viale G. Colombo3, 35121 Padua, Italy; and *Centre for Nervous System Physiopathology, Via Arqua 80/A, 41100 Modena, Italy
A novel 44-kDa gene product (D123) has been pro-posed as necessary for S-phase entry of the cell cycle:a point mutation resulted in a temperature-sensitivearrest in G1-phase. From human fibrosarcoma cDNAlibrary, we have isolated an identical gene and studiedits sequence and mRNA and protein expression. Com-pared with D123, three nucleotide differences withinthe human coding sequence, plus others, result in achange of two amino acids. A partial sequence similar-ity has been found with a yeast gene of unknown func-tion. The protein has several potential phosphoryla-tion sites, is highly hydrophilic, and may be highlystructured in a-helix. The mRNA is abundantly ex-pressed by a variety of normal and transformed cellsand by all tissues examined, being most highly ex-pressed in testis. Specific antibodies, raised against arhD123 polypeptide, recognize a major 42- to 44-kDamolecule in crude extract of various human cell lines.Immunohistochemistry reveals that D123 protein isnot homogeneously expressed: it is detected, often ingranular vescicles, in the cytoplasm of some epithelial,stromal, and sperm cells and in varicosities lining ner-vous fibers, while it appears to be absent in nuclei,endothelial, and smooth muscle cells. The precise linkbetween cytoplasmic occurrence of D123 and cell cycleprogression still remains to be clarified. © 1998 Academic
Press
Key Words: D123; cDNA sequence; recombinant pro-tein; tissue expression; testis.
INTRODUCTION
In the attempt to isolate and sequence an unrelatedmessenger RNA, a cDNA library of human fibrosar-coma cells was screened by PCR using an 18-mer oli-gonucleotide. Although the attempt to clone the targetmRNA failed, a novel cDNA sequence, temporarilynamed PZ32, was cloned and submitted to GenBankunder Accession Number U27112.
During completion of the experimental work, anidentical sequence was published by Okuda andKimura in Japan [1]. The sequence codes for a novelprotein D123 responsible for temperature-sensitiveG1-phase arrest in a mutant of rat fibroblast line. D123consists of 336 amino acids and is detected by specificantibodies as a 44-kDa protein in lung fibroblasts.Quantitative and qualitative changes in D123, due to asingle amino acid change, are assumed to cause thetemperature-sensitive arrest in mutant rat fibroblasts.Nevertheless, the D123 level in the mutant did notdepend significantly on whether cells were proliferat-ing or arrested at the saturation cell density. The novelprotein may be involved in the molecular mechanism ofcell-cycle control, but more molecular and biologicalinsights are required for its function to be establishedprecisely.
A sequence search reveals some similarity of D123with a number of proteins, the most significant beingthe amino acid sequence coded by L8167.23 yeast gene(36% identity), whose function has not yet been deter-mined. Our sequence differs from the original humanD123 in a number of nucleotides, resulting in twoamino acid differences. For the novel sequence, wedescribe here mRNA and protein expression and local-ization for a number of human and rat tissues.
MATERIALS AND METHODS
Cell culture. Human primary fibroblasts, HT-1080 fibrosarcoma,and SK-N-BE neuroblastoma cell lines and rat SMC (primary aorticsmooth muscle cells), IEC-6 (small intestine, ATCC), XC (Rous sar-coma virus-induced tumor), 1R (H-ras-transfected embryo fibro-blast), and L2 (yolk sac carcinoma) cells were grown in Dulbecco’smodified Eagle’s medium (DMEM, Gibco BRL, Gaithesburg, MD),with 10% fetal calf serum (FCS, Gibco BRL). IEC-6 were also sup-plemented with 0.1 U/ml of insulin. Lymphoblastoid B cell line(CEM) was kindly provided by Professor V. Bianchi (Department ofBiology, Padua) and grown in DMEM supplemented with 7% FCS.RPMI medium with 17% FCS was used for human mesangial cells;RPMI medium with 10% FCS was used for MCF7 (breast adenocar-cinoma), Jurkatt (human lymphoid T cell line), K562 (chronic my-eloid leukemia), and Raji (Burkitt lymphoma) cells. Human umbili-cal vein endothelial cell (HUVEC) line was grown in Medium 199with Earle’s salt and 10% FCS. Human HeLa (cervix epithelioidcarcinoma) and LoVo (colon adenocarcinoma) cells were grown in
1 To whom correspondence and reprint requests should be ad-dressed at Ist. Istologia ed Embriologia, viale G. Colombo 3, 35121Padova, Italy. Fax: 0039 49 8276079. E-mail: [email protected].
EXPERIMENTAL CELL RESEARCH 242, 451–459 (1998)ARTICLE NO. EX984074
0014-4827/98 $25.00451Copyright © 1998 by Academic Press
All rights of reproduction in any form reserved.

F12 medium with 10% FCS. All culture media were supplementedwith 100 U/ml penicillin, 100 mg/ml streptomycin, and 0.25 mg/mlamphotericin B. Cell cultures were maintained in 5% CO2 incubatorsat 37°C and passaged at 80% confluence.
Treatment of HT-1080 cells. Human fibrosarcoma HT-1080 cellswere grown in DMEM supplemented with 10% FCS for 24 h prior totreatment. Fresh medium containing 16 nM phorbol myristate ace-tate (PMA, Sigma, St.Louis, MO), 200 pM transforming growth fac-tor-b1 (TGF-b1, R&D System Inc., Minneapolis, MN), 1 mM all-trans-retinoic acid (Sigma), or 100 U/ml tumor necrosis factor-a(TNF-a, Sigma) was then added. The total cytoplasmic RNA wasisolated after 24 h of stimulation and Northern blot analysis wasperformed as described (see below).
Cloning and sequencing. cDNA synthesis and subsequent RACE-PCR (rapid amplification of cDNA ends) were performed using theMarathon cDNA amplification kit (Clontech Laboratories Inc., PaloAlto, CA). Briefly, first strand cDNA was synthesized starting from 1mg of HT-1080 human fibrosarcoma totRNA as template; secondstrand synthesis was performed according to described method [2]with a cocktail of Escherichia coli DNA polymerase I, RNase H, andE. coli DNA ligase. Following creation of blunt ends with T4 DNApolymerase, the double strand (ds) cDNA was ligated to the Mara-thon cDNA adaptor. This ‘‘library’’ of adaptor-ligated ds cDNA wasthen diluted 1:25 in tricine–EDTA buffer and used as template forRACE-PCR.
The PCR amplification of cDNA 39 end (39-RACE) was carried outusing the oligonucleotide GCTCCTGTGCGTGGTACC (hereafter re-ferred to as 364) as sense primer and the adaptor primer (AP-1) asantisense primer. After a 1-min incubation at 94°C, 30 cycles (94°Cfor 30 s, 55°C for 30 s, 68°C for 5 min) were run using Expand LongTemplate PCR System (Boehringer, Mannheim, Germany) in a Ce-tus-Perkin Elmer 480B DNA thermal cycler (Foster City, CA). Afteragarose gel electrophoresis separation, the major PCR product wascut and purified using Sephaglas BandPrep kit (Pharmacia, Upp-sala, Sweden) and cloned into the pCRII-TA cloning vector (Invitro-gen, San Diego, CA). The plasmid named pPZ32 was then sequencedon both strands using both dideoxy methodology [3] and the fluores-cent terminator method using the ABI 377 Sequencer (Applied Bio-systems Inc., Foster, CA).
The cDNA 59-end of the temporarily named PZ32 was obtained by59-RACE using the oligonucleotide TTCACATTCTGAGGAAGTG-GAA (rPZ32-59) as antisense primer based on the partial PZ32sequence and the adaptor primer (AP-1) as sense primer. After a1-min incubation at 94°C, PCR was performed with 30 cycles (94°Cfor 30 s, 55°C for 30 s, 68°C for 3 min) using Expand Long TemplatePCR system (Boehringer). The PCR product was then cloned into thepCRII-TA cloning vector (Invitrogen) and sequenced as describedpreviously.
Cell elutriation and FACS analysis. Exponentially growing CEMcells were separated into nine fractions by centrifugal elutriation, asdescribed by Bianchi et al. [4]. For flow cytometric analysis, 2 3 106
cells from each fraction were fixed in a solution of 80% ethanol for18 h at 4°C and stained with a solution of propidium iodide (10mg/ml) in the presence of 100 U/ml RNase A (Boehringer). The cellcycle distribution and the percentage of cells in G1, S, and G2-M,respectively, in each fraction were determined by flow-cytometricmeasurement of cellular DNA content in a FACScan flow cytometer(Becton-Dickinson, Bedford, MA), equipped with a 488-nm argon ionlaser operating at 15 mW.
RNA extraction and Northern-blot analysis. Total cytoplasmicRNA was isolated from cell lines, from elutriated CEM (lymphoblas-toid B cells), and from rat frozen tissue fragments using the Ultra-spec RNA isolation system (Biotecx Laboratories Inc., Houston, TX).Cells from elutriation were pelleted and lysed by adding 1 ml Ultra-spec RNA; cultured cells were lysed directly in 75-cm2 culture flasks(Falcon, Becton Dickinson) and the lysate was collected.
The cell lysate was transferred immediately into microfuge tubesfor subsequent chloroform RNA extraction and isopropanol precipi-
tation step. The same procedure was used for the frozen rat organfragments (heart, liver, lung, cerebellum, muscle, and testis). Frozensamples (about 100 mg) were pulverized in liquid N2 using pestle andmortar. The powder was transferred into a microfuge tube and lysedby adding 1 ml Ultraspec RNA and repeated pipetting.
Equal aliquots of total cell and tissue RNA (about 15 mg) wereapplied to a formaldehyde, 1% (w/v) agarose gel and electrophoresedbefore transfer to nylon filters Hybond-N (Amersham, Little Chal-font, UK).
In order to verify the RNA quality, the gel was stained for 10 minin an ethidium bromide solution and subsequentially destained withrepetitive washes in deionized water. The RNA capillary blottingwas performed overnight in 103 SSC buffer (1.5 M NaCl, 0.15 Msodium citrate, pH 7). The RNA was UV cross-linked to the filter andhybridized at 65°C with 107 cpm of denatured new sequence, glyc-eraldehyde-3-phosphate dehydrogenase (GAPDH), and b-actin 32P-labeled probes, using Rapid-hyb buffer (Amersham). The humanmultiple tissue Northern blot was purchased from Clontech.
32P-labeled DNA probes were prepared from cDNA of humanD123, human, and rat b-actin and human GAPDH, by the random-primed method [5] using the Klenow fragment of the DNA polymer-ase I (Boehringer).
Autoradiograms were analyzed by scanning densitometry using aHoefer GS300 densitometer (Hoefer, San Francisco, CA).
Analysis of protein sequence. Deduced amino acid compositionwas analyzed by mean of the SAPS program, developed in the groupof Karlin at Stanford University [6]. This software runs on the Webserver of the Bioinformatics Group at the Swiss Institute for Exper-imental Cancer Research (ISREC).
Hydropathicity analysis [7] was performed by ProtScale softwarerunning on the ExPASy Molecular Biology Server of Geneva Univer-sity Hospital and the University of Geneva (Geneva, Switzerland).
Protein sequence similarity search was performed using the fol-lowing software:
● BLASTP and TBLASTN [8] running on the National Center forBiotechnology Information BLAST server in Bethesda, Maryland,with the nonredundant protein and the nonredundant nucleotidedatabases, respectively;
● BLITZ [9, 10] running on the European Bioinformatics InstituteWeb server, with the Swiss-Prot database;
● FASTA [11] running on the GenQuest server, Johns HopkinsUniversity / Oak Ridge National Laboratories, with the Swiss-Protdatabase.
Occurrence within the protein sequence of particular patterns ofestablished or potential significance was verified by searching Pros-ite database [12], using ScanProsite software, running on the Ex-PASy server as above.
Preparation of antibodies to recombinant polypeptide. In order toraise antibodies against the putative protein, a plasmid containing apartial sequence was constructed.
Two oligonucleotides derived from coding region of the new humangene were used as primers for amplification by PCR, using HT-1080human fibrosarcoma cDNA as template. The forward primer con-tained a BamHI restriction site at its 59 end. The reverse primer waslocated at the 39 end of the gene and contained an EcoRI restrictionsite. The sequences of the deoxyoligonucleotide primers were a (for-ward), 59 CAGGATCCGAATAGTTCTCTGAAATGTAAAAC 39, and b(reverse), 59 CGGAATTCAGTCGTCCTCCTGCTGATTTCT 39.
After a 5-min incubation at 94°C, the following sequence wasrepeated five times: 94°C for 1 min, 45°C for 1 min, and 72°C for 3min. In the subsequent 23 cycles, the annealing temperature was55°C. The program was terminated with a 15-min incubation at72°C. The amplified sequences were analyzed on a 0.8% agarose gelto assess their purity and then cut with BamHI and EcoRI andcloned in the same restriction sites of plasmid pRSETB (Invitrogen)to give plasmid pRSETPZ32. This plasmid codes for a fusion protein
452 ONISTO ET AL.

containing the new sequence between amino acids 119 and 336 plusan NH2 tail of 34 amino acids (Fig. 2).
The plasmid pRSETPZ32 constructed in E. coli HB101 was trans-ferred in E. coli BL21 (DE3) LysE for expression of the recombinantprotein. Freshly prepared bacterial cells bearing the recombinantexpression plasmid were grown at 37°C in Luria’s broth with 100mg/ml ampicillin and 30 mg/ml chloramphenicol, until absorbance(590 nm) of the culture suspension reached 0.6 OD. Synthesis of thefusion polypeptide was induced by adding 1 mM isopropyl-b-D-thio-galactopyranoside. This inactivates the lac repressor, thereby per-mitting synthesis of T7 RNA polymerase which, in turn, transcribesDNA sequences adjacent to the T7 promoter resulting in productionof the fusion polypeptide. After 3 h, bacteria were harvested bycentrifugation and then washed with and resuspended in phosphate-buffered saline (PBS), pH 7.4. Cells were lysed by sonication (Bran-son Sonifer, power setting 6, 10 min, 4°C) in buffer A: 6 M guani-dinium, 0.1 M NaH2PO4, 10 mM Tris/HCl, 0.1% Tween 20, 10 mMb-mercaptoethanol, pH 8.0. The lysate was clarified by centrifuga-tion (18,000g, 4°C, 30 min) and the supernatant was then filteredthrough a 0.45-mm membrane (Millipore, Bedford, MA). This filtratewas then loaded onto an ion metal (Ni21) affinity chromatography(IMAC) column (0.8 3 4 cm) (Ni–NTA, Diagen, Hilden, Germany)equilibrated in buffer A. The column was washed with buffer A at pH7.5; the recombinant polypeptide was eluted by reducing buffer pH to4.5. The recovered polypeptide was exhaustively dialyzed at 4°Cagainst PBS, pH 8.3.
New Zealand White rabbits were immunized by multiple intracu-taneous microinjections of 0.2 mg of the IMAC-purified recombinantpolypeptide. Booster injections were repeated at 3-week intervals,and antisera were harvested starting 3 weeks after the third injec-tion. The antibodies were purified through an affinity column of therecombinant polypeptide immobilized to IMAC equilibrated with 50mM Tris–HCl, 10 mM NaCl, pH 7.4 [13]. Bound antibodies were theneluted using the same buffer supplemented with 3 M MgCl2, dialyzedagainst PBS, and diluted to the original serum volume.
Western blot analysis. Human cell lines harvested by 0.5% tryp-sin/0.2% EDTA and E. coli transformed and nontransformed withplasmid pPRSETPZ32 were lysed with sample buffer (50 mM Tris–HCl, pH 6.8, 2% SDS, 10% glycerol, 10% mercaptoethanol, 0.2%bromophenol blue). Conditioned medium was clarified by a 5-mincentrifugation (800g) and concentrated (35). The samples wereboiled for 3 min, electrophoresed through a 13% SDS–polyacryl-amide gel, and electrotransferred to a Hybond-C Extra nitrocellulosefilter (Amersham). Protein on the filter was detected using the affin-ity-purified antibody or the antibody blocked by the fusion proteinand the ECL Western blotting analysis system (Amersham).
Immunohistochemistry. Rats were anesthetized with ketamine(Ketalar, Parke Davis, Italy) 10 mg/kg body wt ip 1 diazepam 2mg/kg im and perfused through the ascending aorta with 100 ml (50ml at 37°C, 50 ml ice-cold) saline solution followed by about 300 mlof 4% paraformaldehyde dissolved in 0.1 M sodium phosphate buffer(pH 7.4). During the perfusion, the animals were kept in an ice-coldbath. After 25–30 min of perfusion, the tissues were removed andimmersed for 90 min in the same ice-cold fixative. The tissues werethen rinsed for 48 h in ice-cold 0.1 M Sorensen’s buffer containing 5%sucrose and cut on a Leitz cryostat (14-mm-thick sections, 220°C).Sections were collected on a glass slide coated with chrome alum andgelatin. They were then immediately processed for the immunoflu-orescence procedure.
The sections were first incubated in 0.1 M PBS at room tempera-ture for 10–30 min, followed by incubation at 4°C for 18 h in a humidatmosphere with the primary antiserum. The primary antiserumwas diluted in PBS containing 0.3% Triton X-100 (v/v). After thesections had been rinsed in PBS for 30 min (33 10 min), they wereincubated at 37°C for 30 min in humid atmosphere with fluoresceinisothiocyanate-conjugated swine anti-rabbit immunoglobulin (Dako,Denmark) containing 0.3% Triton X-100. They were then rinsed inPBS (as above) and mounted in glycerol and PBS (3:1, v/v) containing
1,4-phenylendiamine, 0.1 g/liter. Finally, they were examined in aNikon Microphot FXA microscope equipped with a 100-W mercurylamp and the B-2A filter combination. Tmax 400 film (Kodak, UK)was used for photography.
RESULTS
Isolation of the new sequence. The cDNA encodingthe new human sequence was obtained by a RACEprocedure. The major PCR product obtained by 39-RACE was about 1200 bp long (Fig. 1). After agarosegel separation it was purified and cloned into the pCRIIcloning vector, and the sequence of 1231 bp was deter-mined by manual and automatic sequencing methods.Sequence analysis revealed an ORF of 962 bp, codingfor a hypothetical protein of 320 amino acids, followedby a UTR region of 269 bp containing two putativepolyadenilation sites (Fig. 2). The following 59-RACEadded 108 bp to the 59 end of the cDNA inclusive of thefirst ATG codon and of a 16-amino-acid sequence in theN-terminus of the protein. The RACE strategy of hu-man gene cloning and nucleotide sequence of cDNA(GenBank Accession No. U27112) are shown in Fig. 2.
Sequence analysis. DNA sequence was initiallycompared with GenBank sequences. No significant ho-mologies with other genes were found by using theFASTA sequence similarity search program: the high-est score obtained indicated only extremely dissimilarsequences.
The cDNA sequence of D123 protein has been foundsuccessively by similarity search, and the human type[1] was longer but identical to that isolated in ourlaboratory. Its coding sequence differed on three nucle-otides only: 339, 1011, and 1012; the sequence analysiswas therefore repeated twice more by automatic se-quencing on both human fibrosarcoma and neuroblas-toma cDNA. In all cases, G339, G1011, and A1012 wereconfirmed (Fig. 3), thereby matching the reported ratD123 sequence. Three additional differences were de-termined at positions 1187, 1312, and 1324 of the 39-untranslated region: G, T, and A instead of A, C, and T,respectively. The thus revised complete sequence ishereafter referred to as D123.
Statistical analysis of protein sequence. The compo-sition analysis of the D123-deduced amino acid se-quence by SAPS software revealed the presence of10.4% of Asp and 7.4% of Phe, both above the averagein human proteins (above the 99 and 95% quantile,respectively); on the other hand, Ala (3.9%), Gly (3.3%),and Met (0.6%) were below the average level (all in the5% quantile). Almost one-third are charged aminoacids (60 negative, 36 positive) and global hydropath-icity analysis showed a prevalent hydrophilic pattern(graph not shown).
Northern blot analysis. To analyze the expressionof the D123 gene, total RNA from normal and tumorcell lines and from various human and rat tissues was
453NUCLEIC ACIDS, PROTEIN SYNTHESIS, AND MOLECULAR GENETICS

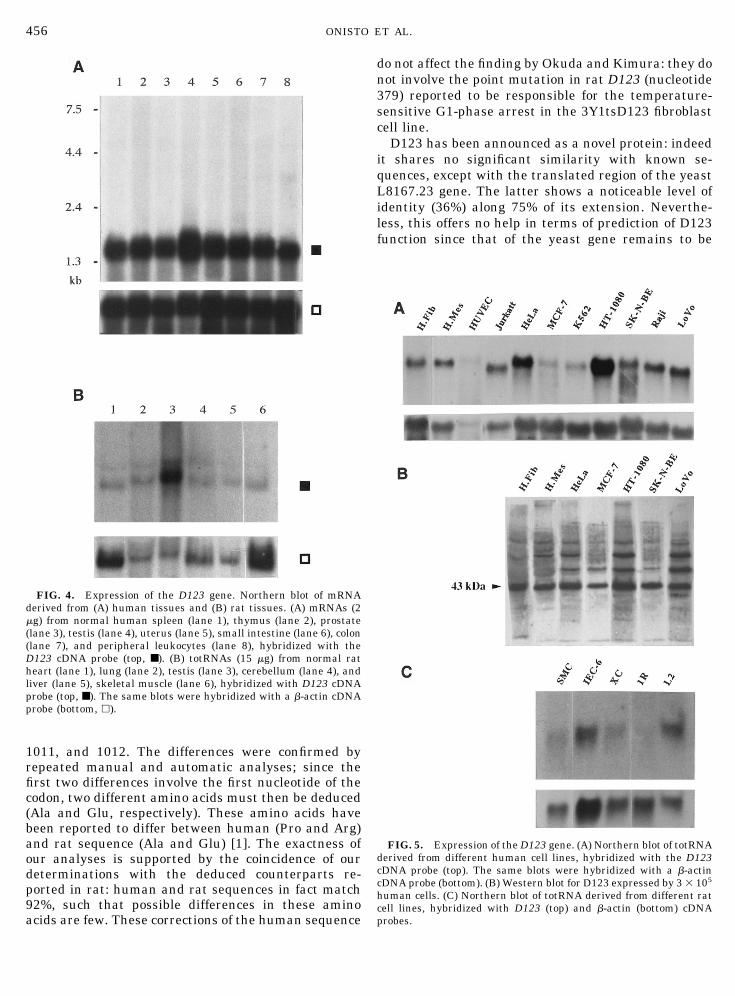
studied by Northern blotting. A major transcript ofapproximately 1.5 kb was detected in all tested humantissues—spleen, thymus, prostate, testis, ovary, smallintestine, colon, and leukocytes—with the highest ex-pression in testis (Fig. 4A). Total RNA from a differentset of rat tissues—lung, cerebellum, heart, liver, skel-etal muscle, and testis—was also analyzed: D123 genewas expressed, at various levels, by all tissues with noexceptions, and again testis showed an expression levelseveralfold higher (Fig. 4B).
Eleven human (three untransformed and eight tu-moral) cell lines were analyzed: D123 mRNA was ex-pressed by all cell types, without clear-cut differencebetween transformed and nontransformed cells, show-ing the highest expression in HT-1080 fibrosarcomaand HeLa carcinoma cells (Fig. 5A). Also in five rat(two normal and three transformed) cell lines D123mRNA was always expressed, without clear-cut differ-ence between transformed and nontransformed ones(Fig. 5C).
The mRNA expression of D123 in the HT-1080 fibro-sarcoma cells was also analyzed after treatment ofthese cells with TNF-a, retinoic acid, PMA, andTGF-b1 for 24 h. These agents were not seen to affectthe mRNA levels of D123 (data not shown).
Since the same gene product has been linked toS-phase entry [1], the expression of D123 mRNA wasanalyzed in cell populations enriched in differentphases of the cycle, obtained by elutriation of CEM
lymphoblastoid B line. Differences in the range of 10–20% in mRNA expression were recorded by Northernblotting between G1 (81% G1, 17% S, 2% G2-M), S(27% G1, 48% S, 25% G2-M), and G2-M (6% G1, 25% S,60% G2-M)-enriched fractions (Fig. 6). Further study is
FIG. 2. RACE strategy of cloning (boxed) and nucleotide anddeduced amino acid sequences of human PZ32 cDNA. The 1339-bp-long sequence is available from GenBank under Accession No.U27112. The amino acids with # and * are potential phosphorylationsites of protein kinase C and CK2 protein kinase, respectively, whileA indicates a potential site of both. Two potential glycosylation sitesare lightly shaded. Underlined nucleotides differ from those se-quenced in D123, and the two different amino acids are in heaviershadowing. The putative polyadenylation signals are double under-lined.
FIG. 1. Agarose gel electrophoresis (1%) of 39-RACE PCR prod-uct from HT-1080 human fibrosarcoma cDNA. The 18-mer 364 wasused as sense primer. Lane 1, 100-bp ladder (Pharmacia). Lane 2,PCR amplification product; the content of the major band (p) waspurified and cloned. Lane 3, b2-microglobulin PCR internal control.
454 ONISTO ET AL.

needed to determine the significance of such differ-ences.
Western blotting. As shown in Fig. 7, the rabbitpolyclonal antibodies raised against D123 polypeptidestrongly recognize on Western blot a unique band ofapproximately 36 kDa in the crude extract of trans-formed E. coli (Fig. 7, lane 4), which corresponds to themolecular weight deduced for the polypeptide ex-pressed by plasmid pRSETPZ32 (Fig. 7, lane 1). Thespecificity of the immune reaction is indicated by thelack of recognition of other antigens in E. coli crudeextract (Fig. 7, lane 5). These antibodies bind to anapproximately 44- to 42-kDa protein in cell extract(Fig. 7, lane 6) of human fibrosarcoma cells; this mo-lecular weight corresponds to that calculated by thededuced amino acid sequence of D123 (44 kDa). Tracereactivity was detectable in the conditioned medium(not shown). Addition of the recombinant fusionpolypeptide to the antiserum blocks the immune reac-tions (Fig. 7, lanes 7 and 8).
The D123 protein level was also analyzed in differenthuman cell lines. As shown in Fig. 5B, in all the linestested the antibody recognizes a main protein of ap-proximately 43 kDa, together with other products ofhigher molecular weight. The expression pattern of theantigens was variable among the different cell lines.
Immunohistochemistry. Since the protein is identi-cal in human and rat, the presence and distribution ofD123 protein was investigated by immunofluorescencein rat lung, small intestine, kidney, stomach, and testisby means of human affinity-purified antibody. The re-activity was intense in the cytoplasm of some bronchio-lar mucosal cells within irregular vescicular bodiesusually facing the luminal site and sometimes alongthe plasmalemma (Figs. 8A and 8B), while no sign ofreactivity was evident in the extracellular space. Cyto-plasm showed intense granular positivity on sparsecells of the intestinal villi and stomach submucosa(Figs. 8C and 8H), and D123-positive varicosities werelined up in nervous fibers within the intestinal mucosa(Fig. 8D). Within different cross sections of seminifer-
ous tubules, a regular front of positive granules clearlyfollowed the wavelike fashion of spermatogenesis: asshown in Fig. 8, positive cells line up either next to thebasal lamina (E) or next to the lumen of the tubules (F),with all intermediate stages (not shown). Weak stain-ing was diffuse in epithelial cells of intestine and stom-ach (Figs. 8C, 8D, and 8H) and kidney tubules (Fig.8G), while the antigen was almost undetectable, underthe same conditions, in endothelial and smooth musclecells (vessels in Fig. 8A) and renal glomeruli (Fig. 8G).Nuclei were negative in all types of cell.
Sequence similarity analysis. Alignment of theD123-deduced amino acid sequence was performed us-ing BLASTP, TBLASTN, BLITZ, and FASTA. Severalproteins showed a limited sequence identity (within a30 to 100 amino acid span), suggesting a partial simi-larity of structure [14]. A more significant similaritywas found with the yeast protein L8167.23 (PIR Acces-sion No. S48566; GenBank Accession No. U14913); nofunction has yet been assigned to this protein, whichshows 36% identity on a 271-amino-acid alignmentlength.
Potential posttranslational modifications. Two po-tential N-glycosylation sites were revealed by Scan-Prosite software: from amino acid (aa) 106 to 109(AsnTyrSerAla) and from aa 119 to 122 (AsnSerSer-Leu). In addition, D123 showed 5 potential proteinkinase C phosphorylation sites (aa 22–24, 47–49, 121–123, 191–193, and 303–305) and 12 for CK2 proteinkinase (Fig. 2).
DISCUSSION
We isolated a 1339-bp sequence containing an ORFof 1008 bp. This sequence, temporarily named PZ32and registered in GenBank in May 1995 in shorterform, was found to be identical to a human and ratcDNA, published in March 1996 and referred to asD123 [1].
Compared to D123 cDNA, our sequencing differs onthree nucleotides within the coding sequence: 339,
FIG. 3. Automatic sequence analysis of PZ32 cDNA. Differences in sequencing between PZ32 and D123 are indicated by an asterisk, andthe corresponding deduced amino acid is reported.
455NUCLEIC ACIDS, PROTEIN SYNTHESIS, AND MOLECULAR GENETICS
1011, and 1012. The differences were confirmed byrepeated manual and automatic analyses; since thefirst two differences involve the first nucleotide of thecodon, two different amino acids must then be deduced(Ala and Glu, respectively). These amino acids havebeen reported to differ between human (Pro and Arg)and rat sequence (Ala and Glu) [1]. The exactness ofour analyses is supported by the coincidence of ourdeterminations with the deduced counterparts re-ported in rat: human and rat sequences in fact match92%, such that possible differences in these aminoacids are few. These corrections of the human sequence
do not affect the finding by Okuda and Kimura: they donot involve the point mutation in rat D123 (nucleotide379) reported to be responsible for the temperature-sensitive G1-phase arrest in the 3Y1tsD123 fibroblastcell line.
D123 has been announced as a novel protein: indeedit shares no significant similarity with known se-quences, except with the translated region of the yeastL8167.23 gene. The latter shows a noticeable level ofidentity (36%) along 75% of its extension. Neverthe-less, this offers no help in terms of prediction of D123function since that of the yeast gene remains to be
FIG. 5. Expression of the D123 gene. (A) Northern blot of totRNAderived from different human cell lines, hybridized with the D123cDNA probe (top). The same blots were hybridized with a b-actincDNA probe (bottom). (B) Western blot for D123 expressed by 3 3 105
human cells. (C) Northern blot of totRNA derived from different ratcell lines, hybridized with D123 (top) and b-actin (bottom) cDNAprobes.
FIG. 4. Expression of the D123 gene. Northern blot of mRNAderived from (A) human tissues and (B) rat tissues. (A) mRNAs (2mg) from normal human spleen (lane 1), thymus (lane 2), prostate(lane 3), testis (lane 4), uterus (lane 5), small intestine (lane 6), colon(lane 7), and peripheral leukocytes (lane 8), hybridized with theD123 cDNA probe (top, ■). (B) totRNAs (15 mg) from normal ratheart (lane 1), lung (lane 2), testis (lane 3), cerebellum (lane 4), andliver (lane 5), skeletal muscle (lane 6), hybridized with D123 cDNAprobe (top, ■). The same blots were hybridized with a b-actin cDNAprobe (bottom, h).
456 ONISTO ET AL.

determined. In addition, insufficient knowledge limitsphylogenetic considerations, for the time being.
However, the D123 amino acid composition is note-worthy. Hydrophilic amino acids consistently prevailin the D123, and in some amino acids its compositionfalls outside the human proteins average: the de-duced 336-amino-acid sequence has been reported tobe rich in Asp and Leu [1]; but, while Leu turns outto be represented on the average, Asp and Phe areabove the average level in human proteins, and Ala,Gly, and Met are below. In addition, D123 shows apredominance of acidic amino acids (60 negative vs36 positive) and its calculated I.P. is 4.6. The relativeabundance of Asp together with Leu, which are in-
volved in helical structure organization, most prob-ably explains the high content of a-helix calculatedfor the secondary structure [15].
Northern blotting shows both abundant expression,comparable to that of b-actin, and wide tissue distri-bution, both in human and in rat, with expression in alltested tissues and in a variety of normal and trans-formed cell cultures. In human cultures, the protein iseasily detectable by specific antibody, in good agree-ment with their mRNA levels (Fig. 5). Products oflower Mr are recognized by the antibody, in accordancewith Okuda and Kimura, who used a different antibodypreparation [1]. However, in contrast with their re-sults, faster products are almost absent, at least ex-cluding degradation of the main protein.
Among the cells and the tissues, little difference inmRNA expression was evident except in the case oftestis, where the transcript was more abundant, byapproximately onefold in human and several times inrat, compared to the average level of the examinedtissues. Nevertheless, in testis, D123 seems almostexclusively expressed by the sperm cell lineage, whileit is not detectable in interstitial cells. Immunohisto-chemistry reveals that only some cell types expressD123: these include bronchiolar mucosal cells, somecells of the intestinal villi and stomach submucosa, andneurons in varicosities along their fibers. To explainwhy only few cell types express in vivo high levels ofD123 protein, the specific function of this novel mole-cule must be determined.
Northern blot analysis of adequately stimulated HT-1080 fibrosarcoma cells demonstrated that the D123gene is not under negative or positive transcriptionalcontrol.
FIG. 7. Detection of the D123 expressed by transformed E. coliand HT-1080 human fibrosarcoma cells. (A) Coomassie blue stainingof crude extract of E. coli transformed (lane 1) and untransformed(lane 2) with plasmid pPRSETPZ32 and of MW markers (lane 3). (B)Western blotting for D123 in crude extract of E. coli transformed(lane 4) and untransformed (lane 5) with plasmid pPRSETPZ32 andin crude extract of 106 HT-1080 cells (lane 6). No reaction wasdetected when corresponding transformed E. coli (lane 7) and HT-1080 (lane 8) samples were developed using the antibody blocked bythe fusion polypeptide.
FIG. 6. Expression of D123 gene in different cell cycle phases.FACS analysis of CEM human lymphoblastoid B cells before (A) andafter (B) elutriation. (B) Continuous line, G1-enriched fraction; dot-ted line, S-enriched fraction; bold line, G2-M-enriched fraction. (C)Northern blotting for D123 and GAPDH mRNAs in the correspond-ing fractions: compared to G1, the values of S and G2-M fractionswere 0.8 and 1.09, respectively (after normalization againstGAPDH).
457NUCLEIC ACIDS, PROTEIN SYNTHESIS, AND MOLECULAR GENETICS

FIG. 8. Immunohistochemistry for D123: rat lung (A, 3140; B, 3570), intestine (C, 3570; D 3570), testis (E, 3285; F, 3570), kidney (G,3285), and stomach (H, 3570).
458 ONISTO ET AL.

Does the D123 gene play a housekeeping like role inthe cell machinery? Though related to molecular mech-anisms of cell cycle control and found responsible fortemperature-sensitive G1-phase arrest, the new geneproduct is not found in the nucleus but in the cyto-plasm. Specific antibodies bind cytoplasmic granulesand vescicles, mainly inside the cells with secretorytraffic—intestinal mucosa, bronchial cells, and neuro-fibers—suggesting a role in intracellular trafficking orin vescicle generation / dissolution.
Lack of basic residue clusters, which—though notexclusively—act as nuclear targeting sequences [16],and lack of nuclear immune reaction reduce the likeli-hood of D123 transport into the nucleus. How thiscytoplasmic protein is involved in the control of the cellcycle is still to be determined, but an important rolecannot be excluded since control of mitosis has beenreported for a number of cytoplasmic proteins [17].However, it is worth noting that cells in G1-phase showno dramatic difference in mRNA expression for D123compared to S- and G2-M-phase, as verified in elutri-ated cells. Whether or not this gene product, recentlylinked to S-phase entry, is modulated during the cellprogression to S-phase must be investigated in moredetail.
Indeed, the presence of several potential phosphory-lation sites specific for CK2 protein kinase [18], com-bined with the relevant D123 hydrophilicity, opens thepossibility that the protein may be modulated by phos-pho/diphosphorylation and implicated in signal trans-duction.
The present report adds to knowledge on the novelgene product D123, and in particular to its cellular andtissular distribution, thereby clarifying the approachto further investigations.
This research was supported by grants from CNR-ACRO andCNR-Bilaterale, Associazione Italiana per la Ricerca sul Cancro, andMURST, Italy. We are grateful to Professor Laura Calza (Depart-ment of Physiology, Medical School of Cagliari), Professor AriannaDonella-Deana (Department of Biological Chemistry, Medical Schoolof Padua), Professor Vera Bianchi and Dr. Paola Ferraro (Depart-ment of Biology, University of Padua), Dr. Renato Zambello (Depart-ment of Clinical Medicine, Medical School of Padua), Dr. Mark Sobel(NCI, Bethesda, MD), and Dr. Antonella Peron for helpful contribu-tions and to Dr. Susan Biggin for revision of the English manuscript.
REFERENCES
1. Okuda, A., and Kimura G. (1996). An amino acid change innovel protein D123 is responsible for temperature-sensitive G1-
phase arrest in a mutant of rat fibroblast line 3Y1. Exp. CellRes. 223, 242–249.
2. Gubler, U., and Hoffman, B. J. (1983). A simple and very effi-cient method for generating cDNA libraries. Gene 25, 263–269.
3. Sanger, F., Nicklen, S., and Coulson, A. R. (1977). DNA se-quencing with chain-terminating inhibitors. Proc. Natl. Acad.Sci. USA 74, 5463–5467.
4. Bianchi, V., Borella, S., Rampazzo, C., Ferraro, P., Calderazzo,F., Chieco-Bianchi, L., Skog, S., and Reichard, P. (1997). Cellcycle-dependent metabolism of pyrimidine deoxynucleosidetriphosphates in CEM cells. J. Biol. Chem. 272(26), 16118–16124.
5. Feinberg, A. P., and Vogelstein, B. (1983). A technique forradiolabeling DNA restriction endonuclease fragments to highspecific activity. Anal. Biochem. 132, 6–13.
6. Brendel, V., Bucher, P., Nourbakhsh, I., Blaisdell, B. E., andKarlin, S. (1992). Methods and algorithms for statistical anal-ysis of protein sequences. Proc. Natl. Acad. Sci. USA 89, 2002–2006.
7. Kyte, J., and Doolittle, R. F. (1982). A simple method for dis-playing the hydropathic character of a protein. J. Mol. Biol.157, 105–132.
8. Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman,D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215,403–410.
9. Sturrock, S. S., and Collins, J. F. (1993). ‘‘MPsrch,’’ Version 1.3,Biocomputing Research Unit, University of Edinburgh, UK.
10. Smith, T. F., and Waterman, M. S. (1981). Identification ofcommon molecular subsequences. J. Mol. Biol. 147, 195–197.
11. Pearson, W. R., and Lipman, D. J. (1988). Improved tools forbiological sequence comparison. Proc. Natl. Acad. Sci. USA 85,2444–2448.
12. Bairoch A., Bucher P., and Hofmann K. (1995). The PROSITEdatabase: its status in 1995. Nucleic Acids Res. 24, 189–196.
13. Negro A., Onisto, M., Masiero, L., and Garbisa S. (1995). Syn-thesis and refolding of human TIMP-2 from E. coli, with specificactivity for MMP-2. FEBS Lett. 360, 52–56.
14. Sander, C., and Schneider, R. (1991). Database of homology-derived protein structures and the structural meaning of se-quence alignment. Proteins 9(1), 56–68.
15. Garnier, J., Osguthorpe, D. J., and Robson, B. (1978). Analysisof the accuracy and implications of simple methods for predict-ing the secondary structure of globular proteins. J. Mol. Biol.120, 97–120.
16. Wen, S.-T., Jackson, P. K., and Van Etten, R. A. (1996). Thecytostatic function of c-Abl is controlled by multiple nuclearlocalization signals and requires the p53 and Rb tumor sup-pressor gene products. EMBO J. 15, 1583–1595.
17. Maccioni, R. B., and Cambiazo V. (1995). Role of microtubule-associated proteins in the control of microtubule assembly.Physiol. Rev. 75, 835–864.
18. Meggio, F., Marin, O., and Pinna, L. A. (1994). Substrate spec-ificity of protein kinase CK2. Cell. Mol. Biol. Res. 40, 401–409.
Received September 1, 1997Revised version received February 20, 1998
459NUCLEIC ACIDS, PROTEIN SYNTHESIS, AND MOLECULAR GENETICS
Copyright © 2022 FDOKUMEN




















