
Hydraulic redistribution in Eucalyptus kochii subsp. borealis with variable access to fresh...
10
ORIGINAL PAPER Hydraulic redistribution in Eucalyptus kochii subsp. borealis with variable access to fresh groundwater K. Brooksbank • D. A. White • E. J. Veneklaas • J. L. Carter Received: 22 July 2010 / Revised: 11 February 2011 / Accepted: 12 February 2011 Ó Springer-Verlag 2011 Abstract Salinity caused by land clearing is an important cause of land degradation in the Western Australian wheatbelt. Returning a proportion of the cleared land to higher water use perennial vegetation is one option for reducing or slowing the salinisation of land. Over the course of a year patterns of water use by Eucalyptus kochii subsp borealis (C. Gardner) D. Nicolle, a mallee eucalypt species, were monitored in three landscape positions with different water availability. One treatment had groundwater at 2 m, a second at 4.5 m and a third had groundwater below a silcrete hardpan thought to be impenetrable to roots. Hydraulic redistribution was observed in all land- scape positions, and rates were positively correlated with the magnitude of soil water potential gradients within the soil. High rates of hydraulic redistribution, facilitated by abundant deep water may increase tree water use by wet- ting surface soils and reducing stomatal closure. This effect may be countered by increased soil evaporation of water moved from root to soil following hydraulic redistribution; the net volumes of redistributed water though lateral roots was calculated to be the equivalent of up to 27% of transpiration. Keywords Sap flow Salinity Hydraulic redistribution Mallee eucalypt Root architecture Introduction In the low rainfall agricultural region of south Western Australia, replacement of deep-rooted native vegetation with a farming system based on annual grain crops and pastures has altered local hydrology, causing water tables to rise and mobilize salt stored in the soil profile (George et al. 1999). Salinisation and waterlogging significantly reduces agricultural productivity (George 1991) and is predicted to affect more than 25% of Western Australia’s agricultural land before a new hydrologic equilibrium is reached (McFarlane et al. 2004). Re-establishing deep-rooted perennial vegetation, such as tree crops has potential to at least partially address this environmental degradation (Stirzaker et al. 1999). It is widely recognized that commercial options are needed to facilitate tree planting on a scale sufficient to restore hydrologic balance in the annual cropping zone of Aus- tralia (Pate and Verboom 2009). One prospective com- mercial tree crop is a group of native multi-stemmed or mallee-form eucalypts collectively known as oil mallees. Since the early 1990s, over 23 million oil mallees have been planted in this cropping zone, mostly in narrow belts separated by wider alleys of annual crops and pastures (M. Kerkmans, pers comm). Integrating trees into the existing farming system distributes the hydrological bene- fits of these plantings across the landscape (Stirzaker et al. 1999). Communicated by C. Lovelock. K. Brooksbank (&) D. A. White E. J. Veneklaas School of Plant Biology, University of Western Australia, 35 Stirling Hwy, Crawley, Perth, WA 6009, Australia e-mail: [email protected] K. Brooksbank D. A. White E. J. Veneklaas J. L. Carter CRC for Future Farm Industries, University of Western Australia, 35 Stirling Hwy, Crawley, Perth, WA 6009, Australia D. A. White J. L. Carter CSIRO Sustainable Ecosystems, CSIRO Centre for Environment and Life Sciences, Private Bag 5, Wembley, WA 6913, Australia 123 Trees DOI 10.1007/s00468-011-0551-0
Transcript of Hydraulic redistribution in Eucalyptus kochii subsp. borealis with variable access to fresh...

ORIGINAL PAPER
Hydraulic redistribution in Eucalyptus kochii subsp. borealiswith variable access to fresh groundwater
K. Brooksbank • D. A. White • E. J. Veneklaas •
J. L. Carter
Received: 22 July 2010 / Revised: 11 February 2011 / Accepted: 12 February 2011
� Springer-Verlag 2011
Abstract Salinity caused by land clearing is an important
cause of land degradation in the Western Australian
wheatbelt. Returning a proportion of the cleared land to
higher water use perennial vegetation is one option for
reducing or slowing the salinisation of land. Over the
course of a year patterns of water use by Eucalyptus kochii
subsp borealis (C. Gardner) D. Nicolle, a mallee eucalypt
species, were monitored in three landscape positions with
different water availability. One treatment had groundwater
at 2 m, a second at 4.5 m and a third had groundwater
below a silcrete hardpan thought to be impenetrable to
roots. Hydraulic redistribution was observed in all land-
scape positions, and rates were positively correlated with
the magnitude of soil water potential gradients within the
soil. High rates of hydraulic redistribution, facilitated by
abundant deep water may increase tree water use by wet-
ting surface soils and reducing stomatal closure. This effect
may be countered by increased soil evaporation of water
moved from root to soil following hydraulic redistribution;
the net volumes of redistributed water though lateral roots
was calculated to be the equivalent of up to 27% of
transpiration.
Keywords Sap flow � Salinity � Hydraulic redistribution �Mallee eucalypt � Root architecture
Introduction
In the low rainfall agricultural region of south Western
Australia, replacement of deep-rooted native vegetation
with a farming system based on annual grain crops and
pastures has altered local hydrology, causing water tables
to rise and mobilize salt stored in the soil profile (George
et al. 1999). Salinisation and waterlogging significantly
reduces agricultural productivity (George 1991) and is
predicted to affect more than 25% of Western Australia’s
agricultural land before a new hydrologic equilibrium is
reached (McFarlane et al. 2004).
Re-establishing deep-rooted perennial vegetation, such
as tree crops has potential to at least partially address this
environmental degradation (Stirzaker et al. 1999). It is
widely recognized that commercial options are needed to
facilitate tree planting on a scale sufficient to restore
hydrologic balance in the annual cropping zone of Aus-
tralia (Pate and Verboom 2009). One prospective com-
mercial tree crop is a group of native multi-stemmed or
mallee-form eucalypts collectively known as oil mallees.
Since the early 1990s, over 23 million oil mallees have
been planted in this cropping zone, mostly in narrow
belts separated by wider alleys of annual crops and pastures
(M. Kerkmans, pers comm). Integrating trees into the
existing farming system distributes the hydrological bene-
fits of these plantings across the landscape (Stirzaker et al.
1999).
Communicated by C. Lovelock.
K. Brooksbank (&) � D. A. White � E. J. Veneklaas
School of Plant Biology, University of Western Australia,
35 Stirling Hwy, Crawley, Perth, WA 6009, Australia
e-mail: [email protected]
K. Brooksbank � D. A. White � E. J. Veneklaas � J. L. Carter
CRC for Future Farm Industries, University of Western
Australia, 35 Stirling Hwy, Crawley,
Perth, WA 6009, Australia
D. A. White � J. L. Carter
CSIRO Sustainable Ecosystems, CSIRO Centre for Environment
and Life Sciences, Private Bag 5, Wembley,
WA 6913, Australia
123
Trees
DOI 10.1007/s00468-011-0551-0

Recent analysis suggests that up to 80% of the landscape
in some catchments will need to be planted to reduce the
area affected by salinity (Bennett and George 2008),
meaning more strategic designs of tree planting will be
required to effect change while minimizing the area
required to be planted. Trees must be planted in locations
where their water use has the effect of reducing ground-
water recharge and where they grow quickly enough to be
an economically viable part of farming systems. This could
be achieved either by planting trees in locations where they
have access to water in addition to current rainfall or by
combining trees with active surface water management
systems. Textural discontinuities, including hardpans and
duplex soils, can cause perched water tables within reach
of the roots of trees. The impact of these water sources on
productivity and the resilience of the farming systems will
depend on the capacity of trees to access and use this water
and to convert it to biomass.
Direct groundwater use has been demonstrated for a
number of species but the amount will vary as a function of
species (White et al. 2002), soil structure (Casper et al.
2003), water quality (Costelloe et al. 2008) and depth to
groundwater (Benyon et al. 2006). The root system archi-
tecture has a significant influence on patterns of soil water
utilization by trees and ultimately canopy transpiration
(Ong et al. 2002). One way that plants can improve access
to limited soil water resources is by hydraulic redistribution
(HR) (Burgess et al. 1998) or transport of water via roots
along water potential gradients from wetter to drier parts of
the soil profile (Richards and Caldwell 1987). This usually
occurs nocturnally when transpiration is low (Hultine et al.
2003), but has also been shown to occur during the day
along water potential gradients due to variation in soil
salinity (Hao et al. 2009). Hydraulic lift is the redistribution
of water from deep soil to surface soil (Horton and Hart
1998) and is the primary focus of this paper.
The ability of a tree to relocate water from one soil
horizon to another or even one zone to another within a
particular soil horizon has been well documented in many
species around the world (Meinzer et al. 2001). A 1998
review found hydraulic redistribution had been confirmed
in 27 species of grasses, herbs, shrubs and trees (Caldwell
et al. 1998), and noted that the phenomenon was likely to
occur wherever plants experienced gradients in soil mois-
ture within their root zone. The adaptive significance of
hydraulic redistribution remains unclear but a number of
possible roles have been proposed: (1) uniform distribution
of water in the soil column after rainfall may promote
water conservation due to lower water potential and soil
conductivity (Ryel et al. 2004); (2) redistribution of water
to the surface may improve availability of nutrients
(Caldwell et al. 1998) (Scholz et al. 2006; Scholz et al.
2008) and (3) improve access to that water for existing
vegetation (Caldwell and Richards 1989; Burgess et al.
2000) and maintain turgor of surface roots (Baker and van
Bavel 1986). In environments that experience long dry
periods, HR has been shown to substantially reduce the rate
of decline of soil water potential over the dry season
(Scholz et al. 2010; Meinzer et al. 2004).
Hydration of surface soils has been identified as poten-
tially playing a role in maintaining stomatal conductance
either through altering the concentration of the hormone
abscisic acid (Wilkinson and Davies 2002), or through direct
hydrostatic signals (Caldwell and Richards 1989; Whitehead
1998). This suggests that rehydration of surface soils by
hydraulic redistribution may help keep stomata open, and
thus improve rates of growth by enabling the trees to main-
tain higher rates of photosynthesis, similar to the stomatal
response to surface irrigation noted by White et al. (1999).
The effects of the presence and depth of groundwater on root
structure and sap flow patterns are less well understood.
Species level information on hydraulic redistribution is
important for understanding ecosystem hydrology and as an
adaptation to resource scarcity (Espeleta et al. 2004).
In this study we investigate patterns of water use and
hydraulic redistribution of Eucalyptus kochii subsp. bore-
alis, an oil mallee, with variable depths to permanent
groundwater and without groundwater by measuring sap
flow in stems, tap roots and lateral roots. The root system of
Eucalyptus kochii subsp. borealis has surface lateral roots
as well as deep tap roots (unpublished data), an architecture
that is well adapted for hydraulic redistribution. We test the
hypothesis that variable access to groundwater will affect
both diurnal and seasonal patterns of water use, and attempt
to quantify net loss of water as a result of efflux following
hydraulic redistribution. The results are used to explore the
potential role of hydraulic redistribution in maintaining
water use during the dry season.
Materials and methods
Site description
All measurements were made between November 2003 and
November 2004 at the Calecono Springs farm on the
Buntine-Marchagee Rd, approximately 30 km south east of
the town of Coorow in Western Australia (30�0202800S,
116�1300000E; Fig. 1). In 1999 the farmer planted Euca-
lyptus kochii subsp. borealis (C. Gardner) D. Nicolle in
north–south oriented belts separated by approximately
110 m, primarily to reduce wind erosion. These tree belts
were comprised of two rows separated by 2 m and within
rows trees were planted every 2 m. The study commenced
when the trees were just over 4 years old. The site was also
used by Carter and White (2009) who also describe details
Trees
123
of the site and experimental infrastructure. The salient
details for this paper are also described below.
The study site has a Mediterranean climate characterized
by a hot dry season (*November to April) and cool wet
season (*May to October). The mean annual rainfall
(1899–2003) was 382 mm, more than 80% of which falls in
the wet season. Annual Priestley–Taylor potential evapo-
transpiration over the same period (Bureau of Meteorology)
was 2,340 mm with a monthly peak of more than 350 mm
in January (Fig. 2). The average annual rainfall since the
trees were planted in 1999 was 380 mm, but only 234 mm
fell in 2004 (measured on site).
Soils consisted of 4–6 m depth of pale yellow sands
consisting mainly of quartz ([80%) with the remainder
goethite and kaolinite. This was overlying a relatively
impermeable granitoid saprolite or silcrete approximately
6 m deep (George 1992). A perched groundwater system
emerged above the silcrete mid-slope and mixed with the
highly saline regional system further downslope. One study
plot was established in each of three landscape positions,
where groundwater was available at 2 m (shallow
groundwater, Shallow GW) and 4.5 m below the surface
(deep groundwater, Deep GW), and where there was no
accessible groundwater (no groundwater, No GW). At the
beginning of the study, the average heights and basal
areas of the trees were 4.31 m, 8.28 9 10-3 m2; 3.18 m,
4.79 9 10-3 m2 and 1.68 m, 1.61 9 10-3 m2 in the
Shallow DW, Deep GW and No GW landscape positions,
respectively. The three plots were approximately 500 m
apart along the same belt of trees running perpendicular to
a 3% slope. Groundwater salinities were relatively uniform
across the site ranging from about 440 mS m-1 under the
crop to 540 mS m-1 under the trees. This is moderately
saline and suitable for most tree species (Department of
Agriculture and Food Western Australia 2005).
Sap flow
Heat pulse sap flow probes (University of Western Aus-
tralia) were installed in three trees per plot using the
approach described by Burgess et al. (2000). To test our
hypothesis regarding hydraulic redistribution and access to
groundwater, sap flow was monitored in stems to estimate
tree water use and in tap and lateral roots to examine
diurnal changes in rates and directions of flow. The heat
ratio method was used to calculate sap velocity and flux
because it can measure low- and reverse-flow and is
therefore ideal for observing hydraulic redistribution in
woody roots (Burgess et al. 2001). The root system was
partially excavated close to the lignotuber using com-
pressed air to minimize mechanical damage to the root
systems. If suitable insertion sites were available, probes
were installed in two tap roots, two lateral roots and one
stem from each tree. We were unable to measure sap flow
in the larger tap roots because they occurred towards the
centre of the root system and access for instrumentation
was obscured by the smaller tap roots. Once the probes
were installed the exposed roots and probe sets were cov-
ered with an insulating blanket.
A total of 15 probe sets were installed across 3 trees in
each plot and connected to an AM16/32 multiplexer and a
CR10X datalogger (Campbell Scientific, Logan UT, USA).
Power was supplied using a sealed lead-acid battery
recharged using a solar panel. Measurements were made
every half hour from late November 2003 to mid April
Fig. 1 Location of the town of Coorow Western Australia where the
study took place
Time (month)
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
mm
0
100
200
300
400
Rain Evaporation
Fig. 2 Long-term (100 years) average rainfall and Priestley–Taylor
potential evapotranspiration (mm) at the Coorow town site (source:
Bureau of Meteorology)
Trees
123

2004 using probe sets comprising a central heater probe
and sensor probes 5 mm upstream and downstream of the
heater. The sensor probes had thermocouples 7.5 and
22.5 mm from the end of the probe allowing measurements
at two depths within each sapwood band. Due to the
increase in diameter of the wound surrounding the insertion
point with time and the effect this has on measurements,
the maximum period for reliable sap flow measurements
from the same installation is 6 months (Burgess et al.
2001). Thus, it was necessary to instrument new trees at the
half way point of this 12-month experiment. At the end of
each measurement period the trees were cut down and
measurements made under zero flow conditions for data
calibration (Burgess et al. 2001). In May 2004 the equip-
ment was installed in different trees that were measured
until November 2004 using the same protocol. For the
second set of measurements only one sensor from each
probe set was used for measurement because this allows for
differential wiring which is less prone to earthing problems
(Maherali and DeLucia 2000). Similarities in measured sap
velocities at two sapwood depths in the first set of mea-
surements (average differences were between 1 and 2%)
indicated this allowed estimation of average stem sap flux
density. The distal sensor was placed at the midpoint in the
sapwood band and any portion of the probe outside the
bark insulated with silicon.
Sap flux density (flow rate per cm2 of sapwood) was
calculated from heat pulse velocity data after taking into
account probe misalignment, wounding, wood density and
sapwood area (Burgess et al. 2001). Flow of sap towards the
extremities of the root system will be referred to as acrop-
etal, and flow towards the lignotuber will be referred to as
basipetal (Burgess et al. 2000). Average sapflow was cal-
culated for three seasonal 14-day periods. To represent
each period, sap flux density was calculated for each
point in time, and then the means combined to form
a curve for a single 24-h period. Standard errors for
these averages were typically less than 1.2 cm3 cm-2 h-1
where sap flow rates were above 5 cm3 cm-2 h-1 and less
than 0.5 cm3 cm-2 h-1 when sap flow rates were below
5 cm3 cm-2 h-1. The chosen periods were 11 Dec 2003–24
Dec 2003 (early dry season), 30 Mar 2004–13 Apr 2004
(late dry season) and 8 Aug 2004–22 Aug 2004 (wet sea-
son). Volumetric estimates of sap flow in stems and roots
from the No GW plot were calculated as the product of sap
flux density and the cross sectional area of conducting
wood. Due to the young age of the trees, no heartwood had
formed in the stems or roots so all wood was assumed to be
conducting and the sapwood cross sectional area was cal-
culated from under-bark diameters. This data was then used
for mass balance calculations to examine whether hydraulic
redistribution by the root system was resulting in efflux of
water into the surrounding soil.
Excavations of the root system for the installation of sap
flow monitoring equipment revealed the basic design to be
dimorphic with lateral roots emerging horizontally from the
lignotuber, and tap roots emerging vertically. This architec-
ture is common in trees growing in semi-arid environments
(Pate et al. 1995; Dawson and Pate 1996). Closer examination
of root systems showed that vertical or sinker roots branched
from along the lateral roots. The depth of these roots is
unknown but they provide access to deeper soil water stores at
a distance from the tree. Xylem water flow towards the
extremities of the instrumented root could have originated
from a secondary sinker distal to the sap flow monitoring point
meaning that total nocturnal acropetal flow measured in those
lateral roots may have been underestimated. Due to this
architectural complexity we did not attempt to assess origins
of water or calculate volumes of HR in the landscape positions
where groundwater was present.
Soil water status and environmental conditions
Soil water status was assessed using a neutron moisture meter
(NMM; California Pacific Neutron, Pacheco, California). To
install the access tubes, holes were drilled down to the sil-
crete, and 50 mm PVC pipe, capped at the end, was inserted
into each hole. The holes were drilled slightly smaller than
the pipe and the pipe was forced into the hole to ensure there
were no air spaces between the tube and the soil. The holes
were then backfilled and capped with kaolin–cement slurry
after (Prebble et al. 1981). Access tubes were positioned in a
transect perpendicular to the tree line and spacing was based
on the average tree height at the time of installation which
was 4 m for the Shallow GW plot, 2 m for the Deep GW plot
and 1.5 m for the No GW plot. The first tube was positioned
between the two tree rows, the second on the drip-line of the
belt, then at 1, 2, 3 and 4 tree heights from the belt. A final
access tube was positioned midway between tree belts
approximately 55 m from the plot. Measurements were taken
on individual days that coincided with the sap flow moni-
toring periods in the late dry season, the early dry season and
the wet season. Soil water content measurements were taken
at depths of 0.1 m then every 0.2 m down to 1.5 m and every
0.5 m down to 4.5 m. Neutron count ratios measured using
the probe were converted to a volumetric soil water fraction
by calibrating against measurements of soil cores collected in
summer and winter adjacent to the access tubes at the end of
the project. Neutron data was kriged in order to present a two-
dimensional profile of soil moisture.
Results
Nocturnal flow was observed in lateral and tap roots in all
landscape positions (Fig. 3). This was clearest in the No
Trees
123

GW plot when surface soil was drier than at depth in early
summer. At that time, water moved up the tap roots and out
along the lateral roots. As the dry season progressed and
the deeper soil moisture was depleted, the rate of hydraulic
redistribution progressively decreased (Fig. 4). In the wet
season, when surface soil was wetter than at depth, water
moved acropetally or down tap roots (No GW, Fig. 3).
In the Deep GW plot the average direction of nighttime
flow for all monitored tap roots in the dry season was
acropetal. However, examination of the individual traces
that made up this average showed that flow direction varied
as a function of the basal diameter of the roots measured. In the
smaller diameter tap roots, water tended to move acropetally
or downwards at night, while in the larger diameter tap roots
nocturnal flow was predominantly basipetal or upwards
(Fig. 5). While the monitored tap roots average flow direction
over the dry season in the Deep GW plot and in the late dry
season in the No GW plot appear to be downwards (Fig. 3),
this may have been biased due to the fact that sap flow was not
monitored in the largest tap roots.
Rates of hydraulic redistribution (HR) in the No GW
plot decreased over the early dry season, indicated by
Wet2 stems10 laterals
Early dryS
FD
(cm
3 cm
-2 h
r-1)
-20
-15
-10
-5
0
5
10
15
20
25
30
35 2 stems10 laterals1 tap
Late dry3 stems4 laterals
2 stems10 laterals1 tap
SF
D (
cm3 c
m-2
hr-1
)
-20
-15
-10
-5
0
5
10
15
20
25
30
35 3 stems5 laterals1 tap
3 stems5 laterals1 tap
SF
D (
cm3 c
m-2
hr-1
)
-20
-15
-10
-5
0
5
10
15
20
25
30
35
1 stem3 laterals5 taps
Time (hours)
1 stem3 laterals5 taps
00:00 06:00 12:00 18:00 00:00 00:00 06:00 12:00 18:00 00:00 00:00 06:00 12:00 18:00 00:00
1 stem3 laterals5 taps
A
B
C
Fig. 3 Mean stem, tap root and lateral root sap flux density
(cm3 cm-2 h-1) in Eucalyptus kochii subsp borealis in the Shallow
groundwater plot (a), the Deep groundwater plot (b) and the No
groundwater plot (c) across three seasons. Each trace is an average
over 14 days in each season. Number in legend represents number of
replicates
Trees
123
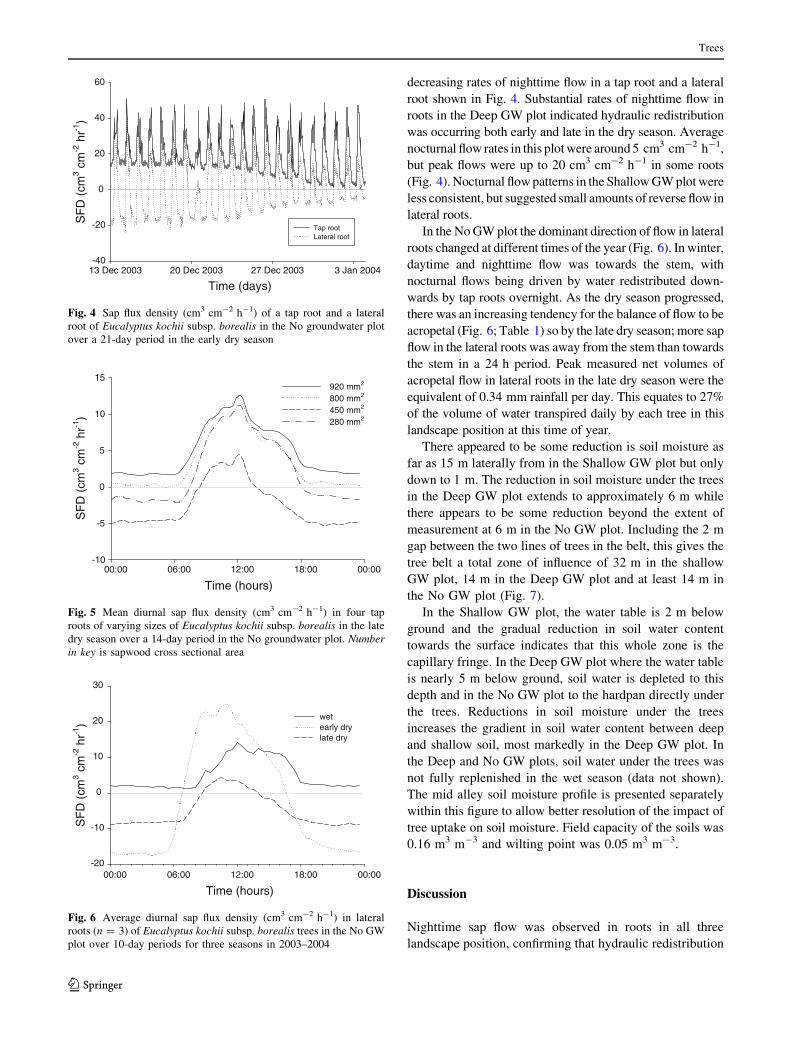
decreasing rates of nighttime flow in a tap root and a lateral
root shown in Fig. 4. Substantial rates of nighttime flow in
roots in the Deep GW plot indicated hydraulic redistribution
was occurring both early and late in the dry season. Average
nocturnal flow rates in this plot were around 5 cm3 cm-2 h-1,
but peak flows were up to 20 cm3 cm-2 h-1 in some roots
(Fig. 4). Nocturnal flow patterns in the Shallow GW plot were
less consistent, but suggested small amounts of reverse flow in
lateral roots.
In the No GW plot the dominant direction of flow in lateral
roots changed at different times of the year (Fig. 6). In winter,
daytime and nighttime flow was towards the stem, with
nocturnal flows being driven by water redistributed down-
wards by tap roots overnight. As the dry season progressed,
there was an increasing tendency for the balance of flow to be
acropetal (Fig. 6; Table 1) so by the late dry season; more sap
flow in the lateral roots was away from the stem than towards
the stem in a 24 h period. Peak measured net volumes of
acropetal flow in lateral roots in the late dry season were the
equivalent of 0.34 mm rainfall per day. This equates to 27%
of the volume of water transpired daily by each tree in this
landscape position at this time of year.
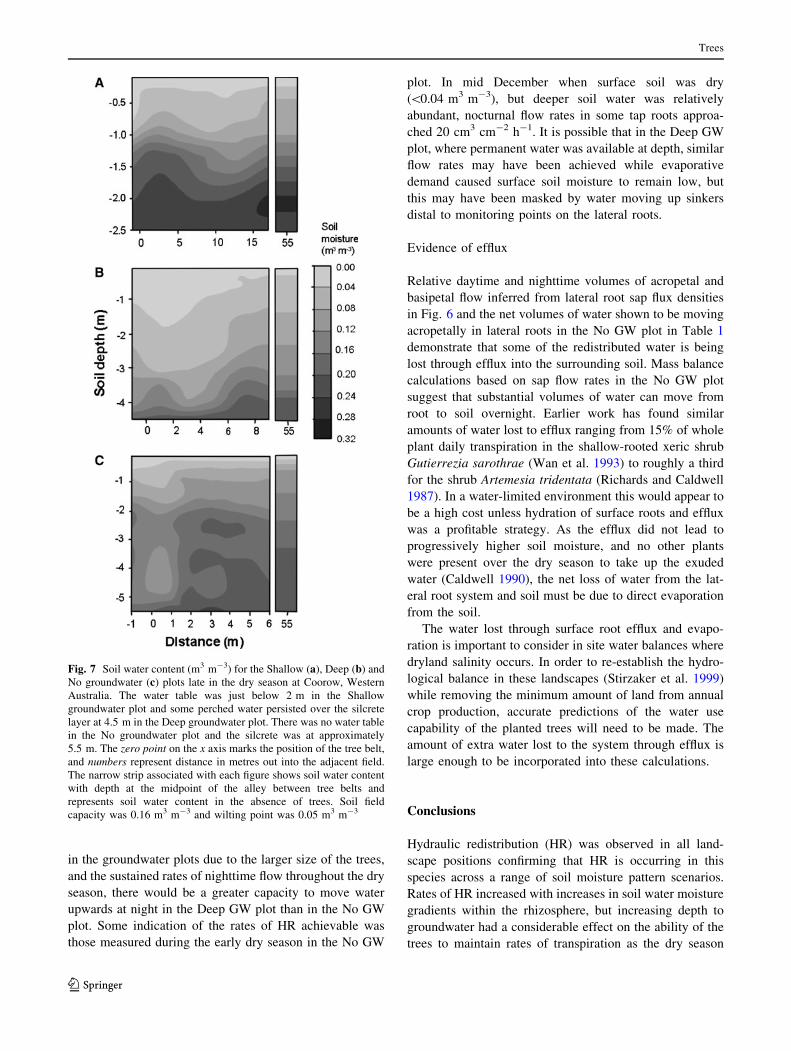
There appeared to be some reduction is soil moisture as
far as 15 m laterally from in the Shallow GW plot but only
down to 1 m. The reduction in soil moisture under the trees
in the Deep GW plot extends to approximately 6 m while
there appears to be some reduction beyond the extent of
measurement at 6 m in the No GW plot. Including the 2 m
gap between the two lines of trees in the belt, this gives the
tree belt a total zone of influence of 32 m in the shallow
GW plot, 14 m in the Deep GW plot and at least 14 m in
the No GW plot (Fig. 7).
In the Shallow GW plot, the water table is 2 m below
ground and the gradual reduction in soil water content
towards the surface indicates that this whole zone is the
capillary fringe. In the Deep GW plot where the water table
is nearly 5 m below ground, soil water is depleted to this
depth and in the No GW plot to the hardpan directly under
the trees. Reductions in soil moisture under the trees
increases the gradient in soil water content between deep
and shallow soil, most markedly in the Deep GW plot. In
the Deep and No GW plots, soil water under the trees was
not fully replenished in the wet season (data not shown).
The mid alley soil moisture profile is presented separately
within this figure to allow better resolution of the impact of
tree uptake on soil moisture. Field capacity of the soils was
0.16 m3 m-3 and wilting point was 0.05 m3 m-3.
Discussion
Nighttime sap flow was observed in roots in all three
landscape position, confirming that hydraulic redistribution
Time (days)13 Dec 2003 20 Dec 2003 27 Dec 2003 3 Jan 2004
SF
D (
cm3 c
m-2
hr-1
)
-40
-20
0
20
40
60
Tap rootLateral root
Fig. 4 Sap flux density (cm3 cm-2 h-1) of a tap root and a lateral
root of Eucalyptus kochii subsp. borealis in the No groundwater plot
over a 21-day period in the early dry season
Time (hours)00:00 06:00 12:00 18:00 00:00
SF
D (
cm3 c
m-2
hr-1
)
-10
-5
0
5
10
15920 mm2
800 mm2
450 mm2
280 mm2
Fig. 5 Mean diurnal sap flux density (cm3 cm-2 h-1) in four tap
roots of varying sizes of Eucalyptus kochii subsp. borealis in the late
dry season over a 14-day period in the No groundwater plot. Numberin key is sapwood cross sectional area
Time (hours)00:00 06:00 12:00 18:00 00:00
SF
D (
cm3 c
m-2
hr-1
)
-20
-10
0
10
20
30
wetearly drylate dry
Fig. 6 Average diurnal sap flux density (cm3 cm-2 h-1) in lateral
roots (n = 3) of Eucalyptus kochii subsp. borealis trees in the No GW
plot over 10-day periods for three seasons in 2003–2004
Trees
123

is occurring in E. kochii across a range of soil moisture
conditions. Although higher rates of hydraulic redistribu-
tion were measured in the No GW plot, complex root
architecture may have masked the extent of nocturnal flow
in the two plots where a water table was present. Increasing
depth to groundwater had a considerable effect on the
ability of the trees to maintain rates of transpiration as the
dry season progressed. There was evidence to suggest that
hydraulic redistribution results in efflux of water into sur-
face soils during the dry season which is consistent with
HR being a mechanism for maintaining stomatal conduc-
tance, thus facilitating higher rates of water use where
groundwater is present.
Patterns of HR in the No GW plot gave a clear picture of
nighttime flow driven by gradients of soil water potential.
Early in the dry season, the dry surface soil surrounding
lateral roots generated gradients in soil water potential that
provided the driving force for HR within the root system.
Extensive lateral root systems have been reported in belt
planted mallee eucalypts reaching up to 20 m from the
edge of the belt at 10 years of age (Sudmeyer and Flugge
2005; Robinson et al. 2006). Later in the dry season less
water was available to redistribute and as a result the rate
of hydraulic redistribution declined, (e.g. Fig. 4). The small
amount of daytime sapflow in lateral roots in the No GW
plot (Fig. 3 No GW) shows the extent to which trees in this
landscape position are reliant on water deeper in the
unsaturated zone at this time of year. Similar trends have
been noted in other tree species (Scholz et al. 2008).
In the Deep GW plot, tap roots and lateral roots appear
to be making an equal and consistent contribution to water
use throughout the dry season. This may be partially due to
the fact that larger amounts of redistributed water are
available via the lateral roots during the day, as well as
sinker roots emanating from laterals continuing to access
the water table throughout the year. In the presence of a
water table, the soil water potential gradients between
surface soil and deeper soil layers intensify over the dry
season providing an increasingly strong driving force for
water to be redistributed within the root system (Brooks
et al. 2002). Where a water table is present, this gradient
will reach an upper limit (saturated soil at depth, very dry
surface soil) so rates of HR will plateau as was the case in
the Deep GW plot (Fig. 3b). In the Shallow GW plot, HR
appeared to occur at a lower rate, presumably because most
of the soil profile was relatively wet providing less impetus
for nocturnal redistribution; consequently it was more
difficult to relate nocturnal flow to observed patterns of soil
stored water.
The nocturnal direction of water flow in tap roots in the
dry season also appeared to vary with root diameter; the
larger tap roots moved water up at night and smaller tap
roots moved water down at the same time in the dry season
(Fig. 5). This may be because larger diameter tap roots
penetrated deeper in the soil profile where soil water
potentials were high and the gradient for water movement
was upwards, while the smaller tap roots were in shallower,
drier soils (with more negative water potentials) and the
gradient for water movement was downwards, meaning the
same patterns were observed in these smaller tap roots as
the lateral roots. The tap roots instrumented in this study
spanned the mid-size diameters only. These results suggest
that the largest diameter roots in each tree are the dominant
source of redistributed water in the dry season and supply
water at night to both the lateral roots and shallower tap
roots. The movement of hydraulically redistributed water is
not necessarily just between the extremities of the root
system, but will occur among all soil layers where soil
water potentials differ; for example, one simulation model
developed divided the soil profile into 7 vertical layers
between which HR was occurring to varying degrees (Ryel
et al. 2002).
Although the measured rates of HR were highest in the
No GW plot, based on the relative size of the trees in each
plot, we would expect the volumes of water moved through
HR were greatest in the Deep GW plot where potential
gradients between saturated and dry soils were larger
(surface soil moisture 0.00–0.04 m3 m-3 c.f. 0.24–0.28
m3 m-3 at 4.5 m), and where larger volumes of water were
available at depth to be redistributed. Hydraulic redistri-
bution is driven along a potential gradient (Richards and
Caldwell 1987), so larger volumes are likely to be moved
when that potential gradient is larger (e.g., between satu-
rated deep soil associated with groundwater and dry sur-
face soil as found in the groundwater plots) (Meinzer et al.
2004). Given the larger sap wood area of the root systems
Table 1 Xylem sap flow in stems and lateral roots at the no groundwater (No GW) landscape position during the day (sunrise to sunset) and
night (sunset to sunrise) in different seasons
Wet Early dry Late dry
Day Night Day Night Day Night
Lateral roots 1.3 (±0.05) 0.3 (±0.01) 2.1 (±0.2) -2.7 (±0.04) 0.2 (±0.02) -1.7 (±0.02)
Stems 3.9 (±0.3) 7.4 (±0.40) 5.6 (±0.2)
Data shown are mean total daily flows in litres per day per tree (n = 3) (±SE) for 10-day periods in each of the wet season, early dry season and
late dry season over 2003–2004
Trees
123
in the groundwater plots due to the larger size of the trees,
and the sustained rates of nighttime flow throughout the dry
season, there would be a greater capacity to move water
upwards at night in the Deep GW plot than in the No GW
plot. Some indication of the rates of HR achievable was
those measured during the early dry season in the No GW
plot. In mid December when surface soil was dry
(\0.04 m3 m-3), but deeper soil water was relatively
abundant, nocturnal flow rates in some tap roots approa-
ched 20 cm3 cm-2 h-1. It is possible that in the Deep GW
plot, where permanent water was available at depth, similar
flow rates may have been achieved while evaporative
demand caused surface soil moisture to remain low, but
this may have been masked by water moving up sinkers
distal to monitoring points on the lateral roots.
Evidence of efflux
Relative daytime and nighttime volumes of acropetal and
basipetal flow inferred from lateral root sap flux densities
in Fig. 6 and the net volumes of water shown to be moving
acropetally in lateral roots in the No GW plot in Table 1
demonstrate that some of the redistributed water is being
lost through efflux into the surrounding soil. Mass balance
calculations based on sap flow rates in the No GW plot
suggest that substantial volumes of water can move from
root to soil overnight. Earlier work has found similar
amounts of water lost to efflux ranging from 15% of whole
plant daily transpiration in the shallow-rooted xeric shrub
Gutierrezia sarothrae (Wan et al. 1993) to roughly a third
for the shrub Artemesia tridentata (Richards and Caldwell
1987). In a water-limited environment this would appear to
be a high cost unless hydration of surface roots and efflux
was a profitable strategy. As the efflux did not lead to
progressively higher soil moisture, and no other plants
were present over the dry season to take up the exuded
water (Caldwell 1990), the net loss of water from the lat-
eral root system and soil must be due to direct evaporation
from the soil.
The water lost through surface root efflux and evapo-
ration is important to consider in site water balances where
dryland salinity occurs. In order to re-establish the hydro-
logical balance in these landscapes (Stirzaker et al. 1999)
while removing the minimum amount of land from annual
crop production, accurate predictions of the water use
capability of the planted trees will need to be made. The
amount of extra water lost to the system through efflux is
large enough to be incorporated into these calculations.
Conclusions
Hydraulic redistribution (HR) was observed in all land-
scape positions confirming that HR is occurring in this
species across a range of soil moisture pattern scenarios.
Rates of HR increased with increases in soil water moisture
gradients within the rhizosphere, but increasing depth to
groundwater had a considerable effect on the ability of the
trees to maintain rates of transpiration as the dry season
Fig. 7 Soil water content (m3 m-3) for the Shallow (a), Deep (b) and
No groundwater (c) plots late in the dry season at Coorow, Western
Australia. The water table was just below 2 m in the Shallow
groundwater plot and some perched water persisted over the silcrete
layer at 4.5 m in the Deep groundwater plot. There was no water table
in the No groundwater plot and the silcrete was at approximately
5.5 m. The zero point on the x axis marks the position of the tree belt,
and numbers represent distance in metres out into the adjacent field.
The narrow strip associated with each figure shows soil water content
with depth at the midpoint of the alley between tree belts and
represents soil water content in the absence of trees. Soil field
capacity was 0.16 m3 m-3 and wilting point was 0.05 m3 m-3
Trees
123

progressed. High rates of HR facilitated by abundant deep
water may appreciably increase tree water use and tree
growth through its effect on stomatal control. Substantial
amounts of water may be lost to the system through the
evaporation of water moved from root to soil following
HR, and this should be taken into account when calculating
water balances.
Acknowledgments We gratefully acknowledge financial support
for this research from the Cooperative Research Centre for Future
Farm Industries. We thank the Department of Agriculture Western
Australia (DAFWA) for providing administrative and logistical sup-
port, and Stanley Rance, Scott Walker and Shayne Micin for their
assistance in the field and laboratory. Special thanks to the Stacey
family for allowing us access their farm to conduct this work.
References
Baker JM, van Bavel CHM (1986) Resistance of plant roots to water
loss. Agron J 78:641–644
Bennett DL, George RJ (2008) Long term monitoring of groundwater
levels at 24 sites in Western Australia shows that integrated farm
forestry systems have little impact on salinity. International
Salinity Forum Conference, Adelaide, South Australia
Benyon RG, Theiveyanathan S, Doody TM (2006) Impacts of tree
plantations on groundwater in south-eastern Australia. Aust J
Bot 54:181–192
Brooks JR, Meinzer FC, Coulombe R, Gregg J (2002) Hydraulic
redistribution of soil water during summer drought in two
contrasting Pacific Northwest coniferous forests. Tree Physiol
22:1107–1117
Burgess SSO, Adams MA, Turner NC, Ong CK (1998) The
redistribution of soil water by tree root systems. Oecologia
115:306–311
Burgess SSO, Pate JS, Adams MA, Dawson TE (2000) Seasonal
water acquisition and redistribution in the Australian woody
phreatophyte, Banksia prionotes. Ann Bot 85:215–224
Burgess SSO, Adams MA, Turner NC, Beverly CR, Ong CK, Khan
AAH, Bleby TM (2001) An improved heat pulse method to
measure low and reverse rates of sap flow in woody plants. Tree
Physiol 21:589–598
Caldwell MM (1990) Water parasitism stemming from hydraulic lift:
a quantitative test in the field. Isr J Bot 39:395–402
Caldwell MM, Richards JH (1989) Hydraulic lift: water efflux from
upper roots improves effectiveness of water uptake by deep
roots. Oecologia 79:1–5
Caldwell MM, Dawson TE, Richards JH (1998) Hydraulic lift:
consequences of water efflux from the roots of plants. Oecologia
113:151–161
Carter JL, White D (2009) Plasticity in the huber value contributes to
homeostasis in leaf water relations of a mallee Eucalypt with
variation to groundwater depth. Tree Physiol 29:1407–1418
Casper BB, Schenk HJ, Jackson RB (2003) Defining a plants
belowground zone of influence. Ecology 84:2313–2321
Costelloe J, Payne E, Woodrow I, Irvine E, Western A, Leaney F
(2008) Water sources accessed by arid zone riparian trees in
highly saline environments, Australia. Oecologia 156:43–52
Dawson TE, Pate JS (1996) Seasonal water uptake and movement in root
systems of Australian phreatophytic plants of dimorphic root
morphology: a stable isotope investigation. Oecologia 107:13–20
Department of Agriculture and Food Western Australia (2005) Soil
Salinity tolerance of plants for Agriculture and revegetation
Espeleta JF, West JB, Donovan LA (2004) Species-specific patterns
of hydraulic lift in co-occuring adult trees and grasses in a
sandhill community. Oecologia 138(3):341–349
George RJ (1991) Management of sandplain seeps in the wheatbelt of
Western Australia. Agric Water Manag 19:85–104
George RJ (1992) Groundwater processes, sandplain seeps and
interactions with regional aquifer systems in South-western
Australia. J Hydrol 134:247–271
George RJ, Nulsen RA, Ferdowsian R, Raper GP (1999) Interactions
between trees and groundwaters in recharge and discharge
areas—a survey of Western Australian sites. Agric Water Manag
39:91–113
Hao GY, Jones TM, Luton C, Zhang YJ, Manzane E, Scholz FG,
Bucci SJ, Cao KF, Goldstein G (2009) Hydraulic redistribution
in dwarf Rhizophora mangle trees driven by interstitial soil water
salinity gradients: impacts on hydraulic architecture and gas
exchange. Tree Physiol 29:697–705
Horton JL, Hart SC (1998) Hydraulic lift: a potentially important
ecosystem process. Trees 13:232–235
Hultine KR, Williams DG, Burgess SSO, Keefer TO (2003)
Contrasting patterns of hydraulic redistribution in the three
desert phreatophytes. Oecologia 135:167–175
Maherali H, DeLucia EH (2000) Xylem conductivity and vulnerabil-
ity to cavitation of ponderosa pine growing in contrasting
climates. Tree Physiol 20:859–867
McFarlane DJ, George RJ, Caccetta PA (2004) The extent and
potential area of salt-affected land in Western Australia estimated
using remote sensing and digital terrain models. In: 1st national
salinity engineering conference, Perth, Western Australia
Meinzer FC, Clearwater MJ, Goldstein G (2001) Water transport in
trees: current perspectives, new insights and some controversies.
Environ Exp Bot 45:239–262
Meinzer FC, Brooks JR, Bucci S, Goldstein G, Scholz FG, Warren JM
(2004) Converging patterns of uptake and hydraulic redistribu-
tion of soil water in contrasting woody vegetation types. Tree
Physiol 24:919–928
Ong CK, Wilson J, Deans JD, Mulayta J, Raussen T, Wajja-Musekwe
N (2002) Tree-crop interactions: manipulation of water use and
root function. Agric Water Manag 53:171–186
Pate JS, Verboom WH (2009) Contemporary biogenic formation of
clay pavements by eucalypts: further support for the phytotarium
concept. Ann Bot 103:673–685
Pate JS, Jeschke WD, Aylward MJ (1995) Hydraulic architecture and
xylem structure of the dimorphic root systems of South-west
Australian species of proteaceae. J Exp Bot 46:907–915
Prebble RE, Forrest JA, Honeysett JL, Hughes MW, McIntyre DS,
Schrale G (1981) Field installation and maintenance. In: Greacen
EL (ed) Soil water assessment by the neutron method. CSIRO
Australia, Melbourne, pp 83–98
Richards JH, Caldwell MM (1987) Hydraulic lift: substantial
nocturnal water transport between soil layers by Artemisiatridentata roots. Oecologia 73:486–489
Robinson N, Harper RJ, Smettem KRJ (2006) Soil water depletion by
Eucalyptus sp. tree belts integrated into agricultural systems.
Plant Soil 286:141–155
Ryel RJ, Caldwell MM, Yoder CK, Or D, Leffler AJ (2002) Hydraulic
redistribution in a stand of Artemisia tridentata: evaluation of
benefits to transpiration assessed with a simulation model.
Oecologia 130:173–184
Ryel RJ, Leffler AJ, Peek MS, Ivans CY, Caldwell MM (2004) Water
conservation in Artemisia tridentata through redistribution of
precipitation. Oecologia 141:335–345
Scholz G, Bucci SJ, Goldstein G, Meinzer FC, Franco AC, Miralles-
Wilhelm F (2006) Removal of nutrient limitations by long-term
fertilization decreases nocturnal water loss in savanna trees. Tree
Physiol 27:551–559
Trees
123

Scholz FG, Bucci SJ, Goldstein G, Moreira MZ, Meinzer FC,
Domec J-C, Villalobos-Vega R, Franco AC, Miralles-
Wilhelm F (2008) Biophysical and life-history determinants
of hydraulic lift in neotropical savanna trees. Funct Ecol 22:773–
786
Scholz FG, Bucci SJ, Hoffmann WA, Meinzer FC, Goldstein G
(2010) Hydraulic lift in a Neotropical savanna: experimental
manipulation and model simulations. Agric For Meteorol 150:
629–639
Stirzaker RJ, Cook FJ, Knight JH (1999) Where to plant trees on
cropping land for control of dryland salinity: some appropriate
solutions. Agric Water Manag 39:115–133
Sudmeyer R, Flugge F (2005) The economics of managing tree-crop
competition in windbreak and alley systems. Aust J Exp Agric
45:1403–1414
Wan CG, Sosebee RE, McMichael BL (1993) Does hydraulic lift
exist in shallow rooted species? A quantitative examination with
a half-shrub Gutierrezia sarothrae. Plant Soil 153:11–17
White DA, Beadle CL, Sands PJ, Worledge D, Honeysett JL (1999)
Quantifying the effects of cumulative water stress on stomatal
conductance of Eucalyptus globulus and Eucalyptus nitens: a
phenomenological approach. Aust J Plant Physiol 26:17–27
White DA, Dunin FX, Turner NC, Ward BH, Galbraith JH (2002)
Water use by contour-planted belts of trees comprised of four
Eucalyptus species. Agric Water Manag 53:133–152
Whitehead D (1998) Regulation of stomatal conductance and
transpiration in forest canopies. Tree Physiol 18:633–644
Wilkinson S, Davies WJ (2002) ABA-based chemical signalling: the
co-ordination of responses to stress in plants. Plant Cell Environ
25:195–210
Trees
123




















