
Human Amnion Epithelial Cell Transplantation Abrogates Lung Fibrosis and Augments Repair
9
Human Amnion Epithelial Cell Transplantation Abrogates Lung Fibrosis and Augments Repair Yuben Moodley 1 , Sivagami Ilancheran 2 , Chrishan Samuel 3 , Vijesh Vaghjiani 4 , Daniel Atienza 1 , Elizabeth D. Williams 5 , Graham Jenkin 1 , Euan Wallace 2 , Alan Trounson 1 *, and Ursula Manuelpillai 2† 1 Monash Immunology and Stem Cell Laboratories, School of Biomedical Sciences, 2 Centre for Women’s Health Research, Monash Institute of Medical Research (MIMR), 3 Howard Florey Research Institute, 4 Centre for Reproduction and Development, MIMR, and 5 Centre for Cancer Research, MIMR, Monash University, Melbourne, Australia Rationale: Chronic lung disease characterized by loss of lung tissue, inflammation, and fibrosis represents a major global health burden. Cellular therapies that could restore pneumocytes and reduce in- flammation and fibrosis would be a major advance in management. Objectives: To determine whether human amnion epithelial cells (hAECs), isolated from term placenta and having stem cell–like and antiinflammatory properties, could adopt an alveolar epithelial phenotype and repair a murine model of bleomycin-induced lung injury. Methods: Primary hAECs were cultured in small airway growth medium to determine whether the cells could adopt an alveolar epithelial phenotype. Undifferentiated primary hAECs were also injected parenterally into SCID mice after bleomycin-induced lung injury and analyzed for production of surfactant protein (SP)-A, SP-B, SP-C, and SP-D. Mouse lungs were also analyzed for inflammation and collagen deposition. Measurements and Main Results: hAECs grown in small airway growth medium developed an alveolar epithelial phenotype with lamellar body formation, production of SPs A–D, and SP-D secretion. Al- though hAECs injected into mice lacked SPs, hAECs recovered from mouse lungs 2 weeks post-transplantation produced SPs. hAECs remained engrafted over the 4-week test period. hAEC administra- tion reduced inflammation in association with decreased monocyte chemoattractant protein-1, tumor necrosis factor-a, IL-1 and -6, and profibrotic transforming growth factor-b in mouse lungs. In addi- tion, lung collagen content was significantly reduced by hAEC treatment as a possible consequence of increased degradation by matrix metalloproteinase-2 and down-regulation of the tissue in- hibitors of matrix metalloproteinase-1 and 2. Conclusions: hAECs offer promise as a cellular therapy for alveolar restitution and to reduce lung inflammation and fibrosis. Keywords: amnion epithelial cells; fetal membranes; lung fibrosis; bleomycin Lung fibrosis is the central pathologic end point of many diseases characterized by progressive inflammation, cell death, and disor- dered repair. Such conditions as acute respiratory distress syn- drome, with a mortality rate that can reach up to 40% and idiopathic pulmonary fibrosis represent a major burden to global health (1). Although immunosuppressive agents are frequently used in the therapeutic armamentarium, there are no effective pharmacologic agents that impact on the morbidity and mortality associated with acute respiratory distress syndrome and idio- pathic pulmonary fibrosis. Studies have shown that allogeneic murine stem cell transplantation reduced inflammation and fibrosis after lung injury (2–4). However, investigations examin- ing human stem cells in treating lung fibrosis are limited. Human bone marrow–derived stem cells, umbilical cord blood mesen- chymal stem cells (uMSCs), and amniotic fluid stem cells (AFSCs) expressed some markers of alveolar epithelium after injection into mice, albeit at very low rates of engraftment in the lungs. However, there are no data on their ability effectively to treat disease models (5–7). Human placental stem cells (PSCs) and MSCs obtained from Wharton jelly of the umbilical cord have been shown to reduce inflammation and fibrosis in bleomycin- injured mice (8, 9). The bleomycin-induced model of lung injury is a well-characterized model of initial inflammation and subse- quent fibrosis. This model has an overlap with acute respiratory distress syndrome and idiopathic pulmonary fibrosis (10). Nota- bly, however, PSCs and Wharton jelly MSCs did not differentiate into lung epithelium in bleomycin-treated mice. Recent studies have shown that human amnion epithelial cells (hAECs), an abundant cell type, isolated from amnion membranes of term- delivered placenta display features of embryonic and multipotent stem cells (11, 12). The amnion and chorion form part of the AT A GLANCE COMMENTARY Scientific Knowledge on the Subject Cellular therapies may become future therapeutic option for lung disease. Both murine and human stem cells have been shown to express some markers of lung phenotype, whereas predominantly murine stem cells reduce fibrosis in animal models of lung injury. What This Study Adds to the Field We use human stem-like cells isolated from a plentiful tissue source to demonstrate that human amnion epithelial cells differentiate into lung epithelium in addition to reducing inflammation and abrogating fibrosis post–lung injury. Furthermore, this study demonstrates functionality of the differentiated stem cells and makes use of novel antibodies to isolate human cells from a mouse lung suspension. (Received in original form January 5, 2010; accepted in final form June 2, 2010) * Present address: California Institute for Regenerative Medicine, San Francisco, California † Present address: Centre for Reproduction and Development, MIMR, Monash University, Melbourne, Australia Supported by NH&MRC project grant 491145 and a Monash University Collaborative Grant; NH&MRC Biomedical Career Development Award 519539 (E.D.W.); NHFA/NH&MRC RD Wright Fellowship (C.S.); and NH&MRC grant 436836 (U.M.). Correspondence and requests for reprints should be addressed to Yuben Moodley, M.D., Ph.D., F.R.A.C.P. School of Medicine and Pharmacology, University of Western Australia, Level 4, Medical Research Foundation Building, 50 Murray Street, Perth, Western Australia 6000, Australia. E-mail: yuben.moodley@uwa. edu.au This article has an online supplement, which is accessible from this issue’s table of contents at www.atsjournals.org Am J Respir Crit Care Med Vol 182. pp 643–651, 2010 Originally Published in Press as DOI: 10.1164/rccm.201001-0014OC on June 3, 2010 Internet address: www.atsjournals.org
Transcript of Human Amnion Epithelial Cell Transplantation Abrogates Lung Fibrosis and Augments Repair

Human Amnion Epithelial Cell TransplantationAbrogates Lung Fibrosis and Augments Repair
Yuben Moodley1, Sivagami Ilancheran2, Chrishan Samuel3, Vijesh Vaghjiani4, Daniel Atienza1,Elizabeth D. Williams5, Graham Jenkin1, Euan Wallace2, Alan Trounson1*, and Ursula Manuelpillai2†
1Monash Immunology and Stem Cell Laboratories, School of Biomedical Sciences, 2Centre for Women’s Health Research, Monash Institute of
Medical Research (MIMR), 3Howard Florey Research Institute, 4Centre for Reproduction and Development, MIMR, and 5Centre for Cancer Research,
MIMR, Monash University, Melbourne, Australia
Rationale: Chronic lung disease characterized by loss of lung tissue,inflammation, and fibrosis represents a major global health burden.Cellular therapies that could restore pneumocytes and reduce in-flammation and fibrosis would be a major advance in management.Objectives: To determine whether human amnion epithelial cells(hAECs), isolated from term placenta and having stem cell–like andantiinflammatory properties, could adopt an alveolar epithelialphenotype and repair a murine model of bleomycin-induced lunginjury.Methods: Primary hAECs were cultured in small airway growthmedium to determine whether the cells could adopt an alveolarepithelial phenotype. Undifferentiated primary hAECs were alsoinjected parenterally into SCID mice after bleomycin-induced lunginjuryandanalyzed forproductionof surfactantprotein(SP)-A,SP-B,SP-C, and SP-D. Mouse lungs were also analyzed for inflammationand collagen deposition.Measurements and Main Results: hAECs grown in small airway growthmedium developed an alveolar epithelial phenotype with lamellarbody formation, production of SPs A–D, and SP-D secretion. Al-though hAECs injected into mice lacked SPs, hAECs recovered frommouse lungs 2 weeks post-transplantation produced SPs. hAECsremained engrafted over the 4-week test period. hAEC administra-tion reduced inflammation in association with decreased monocytechemoattractant protein-1, tumor necrosis factor-a, IL-1 and -6, andprofibrotic transforming growth factor-b in mouse lungs. In addi-tion, lung collagen content was significantly reduced by hAECtreatment as a possible consequence of increased degradation bymatrix metalloproteinase-2 and down-regulation of the tissue in-hibitors of matrix metalloproteinase-1 and 2.Conclusions: hAECs offer promise as a cellular therapy for alveolarrestitution and to reduce lung inflammation and fibrosis.
Keywords: amnion epithelial cells; fetal membranes; lung fibrosis;
bleomycin
Lung fibrosis is the central pathologic end point of many diseasescharacterized by progressive inflammation, cell death, and disor-dered repair. Such conditions as acute respiratory distress syn-drome, with a mortality rate that can reach up to 40% andidiopathic pulmonary fibrosis represent a major burden to globalhealth (1). Although immunosuppressive agents are frequentlyused in the therapeutic armamentarium, there are no effectivepharmacologic agents that impact on the morbidity and mortalityassociated with acute respiratory distress syndrome and idio-pathic pulmonary fibrosis. Studies have shown that allogeneicmurine stem cell transplantation reduced inflammation andfibrosis after lung injury (2–4). However, investigations examin-ing human stem cells in treating lung fibrosis are limited. Humanbone marrow–derived stem cells, umbilical cord blood mesen-chymal stem cells (uMSCs), and amniotic fluid stem cells(AFSCs) expressed some markers of alveolar epithelium afterinjection into mice, albeit at very low rates of engraftment in thelungs. However, there are no data on their ability effectively totreat disease models (5–7). Human placental stem cells (PSCs)and MSCs obtained from Wharton jelly of the umbilical cord havebeen shown to reduce inflammation and fibrosis in bleomycin-injured mice (8, 9). The bleomycin-induced model of lung injury isa well-characterized model of initial inflammation and subse-quent fibrosis. This model has an overlap with acute respiratorydistress syndrome and idiopathic pulmonary fibrosis (10). Nota-bly, however, PSCs and Wharton jelly MSCs did not differentiateinto lung epithelium in bleomycin-treated mice. Recent studieshave shown that human amnion epithelial cells (hAECs), anabundant cell type, isolated from amnion membranes of term-delivered placenta display features of embryonic and multipotentstem cells (11, 12). The amnion and chorion form part of the
AT A GLANCE COMMENTARY
Scientific Knowledge on the Subject
Cellular therapies may become future therapeutic optionfor lung disease. Both murine and human stem cells havebeen shown to express some markers of lung phenotype,whereas predominantly murine stem cells reduce fibrosis inanimal models of lung injury.
What This Study Adds to the Field
We use human stem-like cells isolated from a plentifultissue source to demonstrate that human amnion epithelialcells differentiate into lung epithelium in addition toreducing inflammation and abrogating fibrosis post–lunginjury. Furthermore, this study demonstrates functionalityof the differentiated stem cells and makes use of novelantibodies to isolate human cells from a mouse lungsuspension.
(Received in original form January 5, 2010; accepted in final form June 2, 2010)
* Present address: California Institute for Regenerative Medicine, San Francisco,
California
† Present address: Centre for Reproduction and Development, MIMR, Monash
University, Melbourne, Australia
Supported by NH&MRC project grant 491145 and a Monash University
Collaborative Grant; NH&MRC Biomedical Career Development Award 519539
(E.D.W.); NHFA/NH&MRC RD Wright Fellowship (C.S.); and NH&MRC grant
436836 (U.M.).
Correspondence and requests for reprints should be addressed to Yuben
Moodley, M.D., Ph.D., F.R.A.C.P. School of Medicine and Pharmacology, University
of Western Australia, Level 4, Medical Research Foundation Building, 50 Murray
Street, Perth, Western Australia 6000, Australia. E-mail: yuben.moodley@uwa.
edu.au
This article has an online supplement, which is accessible from this issue’s table of
contents at www.atsjournals.org
Am J Respir Crit Care Med Vol 182. pp 643–651, 2010
Originally Published in Press as DOI: 10.1164/rccm.201001-0014OC on June 3, 2010
Internet address: www.atsjournals.org

embryonically derived inner fetal membranes of the placentawith the maternally derived decidua comprising the outer layer.The amniotic epithelium arises from pluripotent embryonicepiblast cells about 8 days after fertilization in the human,whereas the chorion forms from the placenta several weeks laterin gestation. Because the epiblast is also the origin of the threeembryonic germ layers, it has been suggested that the amnioticepithelium could harbor stem cells even at term pregnancy (13).Although we and others have differentiated hAECs from termpregnancy into lineages derived from all three germ layers (11,12), it is unknown whether they fulfill all the characteristics ofstem cells. For example, clonal expansion is limited and stem cellproperties of clonally expanded hAECs have not been deter-mined. Furthermore, hAECs have morphologic features ofdifferentiated epithelial cells unlike quiescent cells. hAECs arealso known to have low immunogenicity and have been success-fully transplanted into allogeneic recipients (14). Furthermore,hAECs and amnion membranes are widely used in ophthalmol-ogy and dermal applications for their antiinflammatory proper-ties (14). Although these features suggest that hAECs may beattractive for potential cellular therapies, little is known abouttissue-specific differentiation in vivo and if their antiinflamma-tory effects can be exploited after transplantation. We studied thecapacity of hAECs to differentiate into alveolar epithelium andreduce inflammation and fibrosis in bleomycin-injured mice.
MATERIALS AND METHODS
Human and Animal Research Ethics Committees of Monash MedicalCentre and Monash University approved the study.
hAEC Isolation and Culture
Amnion was obtained from normal term pregnancies delivered bycaesarean section. hAECs were isolated, as described previously, andanalyzed by flow cytometry for cytokeratins 7, 8/18 (12); batchesgreater than 99% purity as determined by the cytokeratins wereused (12). For characterization, CD73, CD105, CD117 (c-kit),CD90, CD166, CD31, CD34, CD45, HLA-A-B-C, HLA-DP-DQ-DR,aquaporin-5, and caveolin were analyzed by fluorescence-activated cellsorter. Oct-4, Sox-2, SSEA-4, and GCTM2 were assessed by immuno-histochemistry (12). Nkx2.1, occludin, and mucin mRNA expressionwas analyzed by reverse transcriptase–polymerase chain reaction(qRT-PCR) (n 5 6).
To induce differentiation, primary hAECs were cultured in smallairway growth medium (SAGM; Clonetics, San Diego, CA) or controlsmaintained in Dulbecco’s modified Eagle medium (DMEM)/F12(Gibco, Grand Island, NY) plus 10% FCS (n 5 6) for 4 weeks andassessed for lamellar body formation (electron microscopy) and surfac-tant protein (SP) production (immunohistochemistry and ELISA).
hAEC Injection into Mice
Bleomycin-sulfate (0.15 mg in 20 ml saline; Sigma-Aldrich, St. Louis,MO) was administered intranasally under weak anesthesia (isoflurane,Baxter Healthcare, Toongabbie, New South Wales, Australia) to inducepulmonary fibrosis in 8-week-old SCID mice. To determine the optimalcell dose, 0.5, 1, and 2 3 106 hAECs were injected into the tail vein 24hours after bleomycin and culled 2 weeks later (n 5 4 mice per dose).Lungs analyzed for human DNA by PCR showed mice given 1 3 106
hAECs had the highest human DNA content; hence, this dose wassubsequently used (15). hAECs were localized in murine lungs byimmunohistochemistry for the human specific inner mitochondrial mem-brane protein (IMM; 1:100, Chemicon, Boronia, Victoria, Australia).
Differentiation of hAECs in murine lungs was assessed by injectinghAECs obtained from pregnancies of male babies into female micegiven bleomycin and culled 2 weeks later (n 5 8 mice). Lungs weredigested in type I collagenase (3 mg/ml; Invitrogen) for 45 minutes anderythrocytes lysed. hAECs were recovered from lung cell suspensionsby fluorescence-activated cell sorter using antihuman CD29 antibody.Cytospins were prepared from flow– sorted cells and analyzed for XY/
XX chromosomes by fluorescence in situ hybridization (FISH) and SPsA–D by immunohistochemistry.
To assess hAEC effects on inflammation and fibrosis, the controlgroups were as follows: healthy (intranasal saline, 20 ml); saline (tailvein, 100 ml); healthy mice 1 hAECs; bleomycin alone; bleomycin 1
fibroblasts; and bleomycin 1 cytotrophoblasts. Experimental groupscomprised bleomycin 1 hAECs (24 hours postbleomycin) and bleo-mycin 1 hAECs (2 weeks postbleomycin) (n 5 8 mice per group).Animals were killed 2 and 4 weeks later. Inflammation and fibrosis wasdetermined by the Ashcroft score on lung histology (16, 17). CytokinemRNA expression in lungs was measured by RT-qPCR and protein bya cytometric bead array (BD Biosciences, San Jose, CA) or ELISAs(R&D Systems, Minneapolis, MN). Lung collagen content was mea-sured by the hydroxyproline assay, matrix metalloproteinase (MMPs)by gelatin zymography, and tissue inhibitor of metalloproteinases(TIMPs) by RT-qPCR and reverse zymography (18, 19). METHODS aredescribed in greater detail in the online supplement.
Statistics
Data are expressed as the mean 6 SEM and analyzed by one-wayanalysis of variance and the Bonferroni post hoc test. P , 0.05 wasconsidered to be significant. GraphPad Prism software (v5.03; La Jolla,CA) was used for data analysis.
RESULTS
hAECs Produce Surfactant Proteins In Vitro
hAECs isolated from term amnion expressed some epithelial(cytokeratin-7, 8/18), pluripotent (Oct-4, Sox-2, SSEA-4,GCTM2), and MSC markers (CD73, CD117, CD166, CD90).HLA-A, -B, and -C was low, whereas HLA-DP, HLA-DQ, HLA-DR, endothelial (CD31, CD34), and hematopoietic (CD45)lineage markers were absent. The primary hAECs also expressedlung-associated markers Nkx2.1, mucin, occludin, aquaporin-5,and caveolin-1 (Table 1). Noting the expression of Nkx 2.1, anearly lung lineage specification marker in primary hAECs, wecultured the cells in small airway growth medium (SAGM),a medium that induces differentiation of embryonic stem cellsinto type II pneumocytes (20). hAECs grown in SAGM producedSP-A (62 6 6% of cells); SP-B (73 6 8%); pro–SP-C (78 6 8%);and SP-D (48 6 6%). Cultures maintained in DMEM/F12 did not
TABLE 1. CHARACTERIZATION OF PRIMARY HUMAN AMNIONEPITHELIAL CELLS
Marker Percentage (n 5 4–7)
Cytokeratin 7* 98.7 6 0.7
Cytokeratin 8/18* 98.4 6 1.1
Oct-4† 4.8 6 1.2
Sox-2† 6.4 6 1.8
SSEA-4† 98.6 6 0.6
GCTM2† 97.4 6 1.3
c-kit (CD117) * 6.7 6 1.3
CD73* 95.3 6 1.2
CD166* 91.9 6 4.2
CD90* 22.5 6 3.8
CD105* Not detected
HLA-A, -B, -C* 2.73 6 0.3
HLA-DP, -DQ, -DR* Not detected
CD45* Not detected
CD31* Not detected
CD34* Not detected
Nkx 2.1‡ Present
Mucin‡ Present
Occludin‡ Present
Caveolin-1* 91.1 6 2.1
Values are expressed as means 6 SEM.
* Flow analyses.† Immunocytochemistry.‡ Real-time polymerase chain reaction.
644 AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE VOL 182 2010

produce SPs (Figure 1A). We also examined if hAECs cultured inSAGM secreted SP-D, which would suggest functionality. After 4weeks in SAGM, hAECs were stimulated with dexamethasoneand SP-D measured in the supernatant by ELISA. SP-D wassecreted by hAECs grown in SAGM alone, whereas dexameth-asone stimulation increased SP-D by a further 30% (Figure 1A).Cells grown in DMEM/F12 did not secrete SP-D. Furthermore,ultrastructural examination revealed that hAECs grown inSAGM for 2–4 weeks had a distinct cell population withcytoplasmic organelles containing surfactant-associated lamellarbodies typical of type II pneumocytes. These features were absentin hAECs cultured in DMEM/F12 (Figure 1B).
hAECs Localize to Injured Lung
hAECs injected into the tail vein 24 hours after bleomycin-induced lung injury was shown to locate to the lung by differentassays. Lung tissue retrieved 2 weeks post-hAEC injection andanalyzed for human Alu repeat sequences by DNA-PCR (15)confirmed the presence of hAECs within the lung. Furthermore,hAECs were identified in the lung by immunostaining forhuman IMM. The specificity of the IMM antibody was demon-strated by immunostaining of human amniotic epithelium andhAECs but not mouse lung tissue (Figure 1C). IMM-positivehAECs were identified in fibrotic regions of injured lungs 4weeks post–cell injection (10 6 0.7 hAECs per 100 lung cells)(Figure 1D). IMM-positive cells were also present in alveolarregions (6 6 0.4 hAECs per 100 lung cells) (Figure 1D). hAECswere absent in lung tissue from healthy mice injected with cells.There was no evidence of tumor formation in lung tissueexamined at 4 weeks post-hAEC injection.
hAECs Demonstrate a Type II Pneumocyte Phenotype In Vivo
To determine whether hAECs adopted characteristics of type IIpneumocytes in mice, antihuman CD29 antibody was used to
recover human cells from mouse lung cell suspensions by flowcytometry. The specificity of the CD29 antibody was demon-strated by positively stained human amniotic epithelium andisolated hAECs, but not mouse lung tissue (Figure 2A). At 2weeks post-hAEC injection, CD29-positive cells constituted 5.3 6
0.9% of the total lung suspension (Figure 2A). To verify that theCD29-positive cells were indeed human, we injected hAECsobtained from pregnancies with male babies into female SCIDmice and analyzed the recovered cells by FISH for the XY chro-mosomes to identify human cells and XX chromosomes repre-senting mouse cells. Greater than 98% of flow sorted cells werehuman by this criterion (Figure 2B). Notably, primary hAECsthat were injected did not produce pro–SP-C, SP-A, SP-B, orSP-D (Figure 2C). The antibodies against the SPs showed specificcytoplasmic staining of human lung tissue, whereas SP-B and pro–SP-C also showed nonspecific nuclear staining of mouse lung cells(Figure 2C). The CD29-positive cells recovered from bleomycin-injured lungs by fluorescence-activated cell sorter were positivefor SP-A (65 6 5%), SP-B (72 6 7%), pro–SP-C (79 6 8%), andSP-D (70 6 8%) by immunohistochemistry (Figure 2D).
hAECs Reduce Inflammation in the Injured Lung
Bleomycin instillation caused a significant inflammatory andfibrotic infiltration of the lung that was attenuated by hAECinjection (Figure 3A). Ashcroft scores of inflammation andfibrosis on histology showed that bleomycin-induced pneumo-nitis was significantly reduced at 2 and 4 weeks by hAECadministration (Figure 3B) (P , 0.03; Student t test). Theinflammatory response to bleomycin was accompanied by in-creased mRNA expression of IL-1, IL-2, IL-6, IFN-g, tumornecrosis factor (TNF)-a, monocyte inflammatory protein, andtransforming growth factor (TGF)-b compared with controls(P , 0.001 for all cytokines). The expression of these cytokineswas significantly attenuated in mice receiving hAECs (P , 0.01
Figure 1. Human am-
nion epithelial cells
(hAECs) acquire an alve-
olar epithelial cell phe-notype. (A) Primary
hAECs cultured in small
airway growth medium
(SAGM) for 4 weeks pro-duce prosurfactant pro-
tein (SP)-C compared
with controls in DMEM/
F12. Similar results (notshown) were obtained
for SP-A, SP-B, and SP-
D. hAECs grown inSAGM secreted SP-D
when stimulated by
dexamethasone treat-
ment. (B) Transmissionelectron microscopy of
hAECs grown in SAGM
showed lamellar body–
like organelles (arrows);these were absent in
controls. (C) hAECs lin-
ing term amnion andisolated hAECs show
strong immunostaining
for human inner mito-
chondrial membrane (IMM) protein. (D) Lung tissue sections from bleomycin-injured mice and isotype controls (not shown) did not stain forIMM. Lung tissue sections from mice given bleomycin 1 hAECs, 4 weeks after cell injection, showed IMM1 cells in fibrotic and alveolar areas (brown
staining, original magnification 350, 3100, and 3200). Scale bars 5 50 mm.
Moodley, Ilancheran, Samuel, et al.: Amnion Epithelial Cells Augment Lung Repair 645

for all cytokines). IL-10 mRNA was significantly increased inmice given hAECs compared with bleomycin (P , 0.01) (Figure4A). The expression of human specific cytokines in lungs ofhAEC-injected mice was also investigated. hAECs expressedmRNA for IL-1, IL-10, TNF-a, TGF-b, migration inhibitionfactor, and IFN-g. We detected low levels of human specificcytokine mRNA for IL-1, IL-10, TNF-a, and TGF-b in lungs ofmice culled two weeks after hAEC transplantation using real-time PCR. However, mRNA for migration inhibition factor andIFN-g were not detectable. Gel electrophoresis showing thePCR products generated is shown in Figure 4B.
We also examined changes in cytokine protein levels inmouse lung tissue lysates; IL-1, IL-6, monocyte chemoattractantprotein-1, and TGF-b were reduced significantly in mice treatedwith hAECs compared with animals given bleomycin alone(P , 0.05 for all cytokines) (Figure 4C).
hAECs Reduce Fibrosis in the Injured Lung
We investigated whether hAECs reduced fibrosis by assessingcollagen deposition using the hydroxyproline assay. Collagen was
elevated 4 weeks after bleomycin instillation (P , 0.05 versushealthy controls), but was significantly reduced in the cohortreceiving hAECs (P , 0.05 versus bleomycin). Notably, injectionof human lung fibroblast cells or placental cytotrophoblast cellsinto bleomycin-injured mice did not lower lung collagen content.Furthermore, hAECs did not influence collagen levels in lungs ofhealthy mice. The effect of hAECs on well-established injury wasalso investigated by administering the cells 2 weeks after bleo-mycin instillation. hAECs significantly reduced collagen in thiscohort (P , 0.05 vs. bleomycin alone) (Figure 5A).
hAEC Treatment Alters Protease Levels in the Injured Lung
Gelatin zymography demonstrated that active MMP-2 wassignificantly elevated in bleomycin-injured mice compared withhealthy controls (P , 0.05), which is in keeping with previousstudies (21). Notably, hAEC injection into bleomycin-exposedmice resulted in a significant increase in latent and active MMP-2 compared with bleomycin alone or healthy controls (P , 0.05for all comparisons). Active MMP-2 was not elevated in healthycontrols treated with hAECs or bleomycin-injured mice given
Figure 2. Analysis of CD29-positive cells from mouse lung cell suspensions. (A) Human amnion epithelial cells (hAECs) lining term amnion and
isolated hAECs show strong immunostaining for CD29 (diaminobenzidine-DAB staining). Haemotoxylin stained lung tissue sections frombleomycin-injured mice and isotype controls (not shown) did not stain for CD29. The CD291 population was easily identified and separated by
flow cytometry 2 weeks after hAEC injection and comprised 5.3 6 0.9% of the total lung cell suspension. Magnification 100x. (B) hAECs from male
placenta (XY) were injected into female (XX) SCID mice. Fluorescence in situ hybridization analysis for XY chromosomes (Poseidon RAB9B)confirmed that flow-sorted CD291 cells were human and that the CD292 population were murine (XX) cells. (C) Human lung alveolar epithelial cells
stained positively for surfactant protein (SP)-A and pro–SP-C (DAB). Mouse lung cells showed no cross reactivity for SP-A. Nonspecific nuclear
staining was seen for pro–SP-C. Primary hAECs that were injected lacked staining for surfactant proteins. Cells and tissue sections counterstained
with haemotoxylin. (D) Immunohistochemistry showed that greater than 65% of flow-sorted CD291 cells recovered from murine lungs producedSP-A, SP-B, pro–SP-C, and SP-D (DAB) Magnification 100x (A-C and 50x in D). Magnification 100x (A-C and 50x in D). Scale bars 5 20 mM in cell
spreads and 50 mM in tissue sections.
646 AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE VOL 182 2010

human fibroblasts (Figure 5B). Analyses of TIMP mRNAexpression showed a down-regulation of TIMP-1, TIMP-3,and TIMP-4 in mice treated with hAECs (P , 0.05). Reversezymography demonstrated that TIMP-1 and TIMP-2 proteinswere up-regulated after bleomycin and significantly reduced byhAEC treatment (P , 0.05) (Figure 5C).
DISCUSSION
This study described several novel findings that extend the fieldof stem cell therapy for lung disease. We have used a novelsource of cells (hAECs) to treat lung injury. Furthermore, toour knowledge, this is the first study to describe human cellsthat not only differentiate into an alveolar epithelial phenotypebut also rescue an injury model of lung fibrosis. Significantly,hAECs were not only positive for lung epithelial markers afterboth in vitro and in vivo differentiation assays, but alsodisplayed functionality in the secretion of SP-D after stimula-tion with corticosteroids.
Preliminary characterization studies of hAECs generatedfrom our laboratory by Ilancheran and coworkers (12) revealedthat the primary cells express a number of pluripotent andmultipotent stem cell markers, are clonogenic, and express lowlevels of HLA Class IA and IIB antigens. Furthermore, hAECsdifferentiated into lineages derived from all three germ layersin vitro (12). Based on this study, we hypothesized that hAECsmay be an effective modality of cellular therapy; we thereforeexplored their reparative and differentiation properties ina model of lung injury.
As part of the focus on lung differentiation and repair, wefound that hAECs express thyroid transcription factor or Nkx 2.1mRNA, which is among the earliest lineage specification markersof the developing lung. Nkx. 2.1 is a transcription factor critical forbranching lung morphogenesis and the formation of type IIpneumocytes (22). Primary hAECs also expressed epithelialmRNAs occludin, mucin, caveolin-1, and aquaporin-5, but lackedthe more lung-specific proSP-C.
Upon noting the expression of Nkx 2.1 in primary hAECs,we cultured these cells in SAGM, which has been shown toinduce differentiation of uMSC and embryonic stem cells into
type II pneumocytes (20, 23). SAGM contains hydrocortisone,human epidermal growth factor, and retinoic acid, which arefactors required for lung development (24). Primary hAECsgrown in SAGM produced SP-A, SP-B, pro–SP-C, and SP-D,whereas control cells cultured in DMEM/F12 did not expressSPs. SP-A, SP-B, and SP-C are synthesized in the endoplasmicreticulum of type II pneumocytes and form lamellar bodies,whereas SP-D is synthesized through a different pathway (25).Collectively, the SPs contribute to lowering the surface tensionbetween the air–liquid interfaces in the lung. SP-D and SP-Aalso play a part in the innate defense system of the lung (25).The production of all SPs is suggestive of hAEC differentiationto a phenotype of type II pneumocytes. Furthermore, ultrastruc-tural examination revealed that cells grown in SAGM for 2 to 4weeks contained a distinct cell population with cytoplasmicorganelles containing lamellar bodies typical of type II pneumo-cytes. These features were absent in hAECs cultured in DMEM/F12. We also examined whether hAECs cultured in SAGMsecreted SP-D, which would suggest functionality. After 4 weeksof culture in SAGM, hAECs were stimulated with dexametha-sone and SP-D measured in the supernatant. Corticosteroids areused to accelerate lung maturation in preterm infants by aug-menting surfactant production and secretion (26). Notably, SP-Dproduction was increased in hAECs after stimulation with cor-ticosteroids, suggesting a phenotype that is functionally similarto type II pneumocytes. We noted other markers of type IIpneumocytes including CD44v6 (27), CD208 (28), and gp600(29). However, these markers share epitopes with other celltypes and tissue and are not as specific as SPs for type IIpneumocytes.
Primary hAECs that lacked SP production were also injectedinto bleomycin-treated SCID mice and identified by IMMimmunostaining in fibrotic and alveolar regions 4 weeks post–cell transplantation without evidence of tumor formation. Fur-thermore, the hAECs were recovered from lung suspensions byflow cytometry using CD29. FISH analysis of both human andmouse chromosomes verified that the flow–sorted CD29-positivecells were human. Notably, the flow–sorted cells produced all SPsdemonstrating that hAECs were capable of differentiating intotype II pneumocytes in vivo. Taken together, these data support
Figure 3. Human amnion ep-
ithelial cells (hAECs) reduce
inflammation of the lung afterbleomycin injury. (A) Bleomy-
cin induced an inflammatory
infiltrate at 2 weeks and an
inflammatory and fibroblasticinfiltrate of the lung at 4
weeks. The lung injury was
attenuated by hAEC injection
at both time-points. Scalebars 5 50 mM. (B) The Ash-
croft score confirmed reduced
inflammation and fibrosis after
hAEC injection (*P , 0.03).
Moodley, Ilancheran, Samuel, et al.: Amnion Epithelial Cells Augment Lung Repair 647
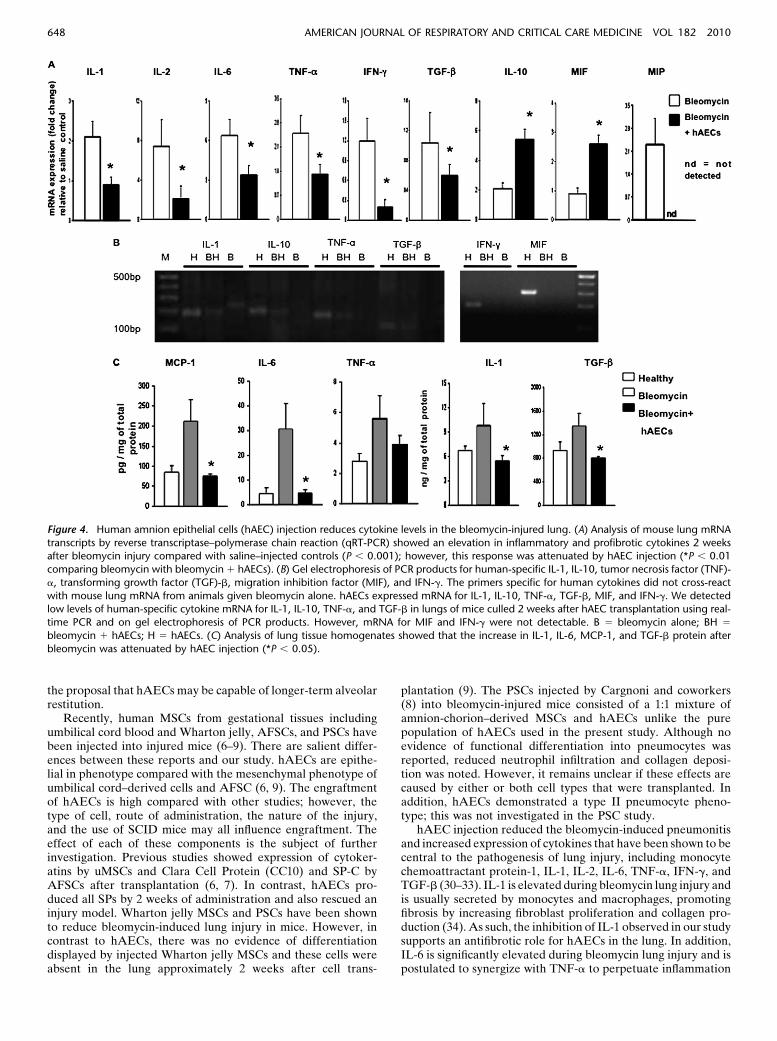
the proposal that hAECs may be capable of longer-term alveolarrestitution.
Recently, human MSCs from gestational tissues includingumbilical cord blood and Wharton jelly, AFSCs, and PSCs havebeen injected into injured mice (6–9). There are salient differ-ences between these reports and our study. hAECs are epithe-lial in phenotype compared with the mesenchymal phenotype ofumbilical cord–derived cells and AFSC (6, 9). The engraftmentof hAECs is high compared with other studies; however, thetype of cell, route of administration, the nature of the injury,and the use of SCID mice may all influence engraftment. Theeffect of each of these components is the subject of furtherinvestigation. Previous studies showed expression of cytoker-atins by uMSCs and Clara Cell Protein (CC10) and SP-C byAFSCs after transplantation (6, 7). In contrast, hAECs pro-duced all SPs by 2 weeks of administration and also rescued aninjury model. Wharton jelly MSCs and PSCs have been shownto reduce bleomycin-induced lung injury in mice. However, incontrast to hAECs, there was no evidence of differentiationdisplayed by injected Wharton jelly MSCs and these cells wereabsent in the lung approximately 2 weeks after cell trans-
plantation (9). The PSCs injected by Cargnoni and coworkers(8) into bleomycin-injured mice consisted of a 1:1 mixture ofamnion-chorion–derived MSCs and hAECs unlike the purepopulation of hAECs used in the present study. Although noevidence of functional differentiation into pneumocytes wasreported, reduced neutrophil infiltration and collagen deposi-tion was noted. However, it remains unclear if these effects arecaused by either or both cell types that were transplanted. Inaddition, hAECs demonstrated a type II pneumocyte pheno-type; this was not investigated in the PSC study.
hAEC injection reduced the bleomycin-induced pneumonitisand increased expression of cytokines that have been shown to becentral to the pathogenesis of lung injury, including monocytechemoattractant protein-1, IL-1, IL-2, IL-6, TNF-a, IFN-g, andTGF-b (30–33). IL-1 is elevated during bleomycin lung injury andis usually secreted by monocytes and macrophages, promotingfibrosis by increasing fibroblast proliferation and collagen pro-duction (34). As such, the inhibition of IL-1 observed in our studysupports an antifibrotic role for hAECs in the lung. In addition,IL-6 is significantly elevated during bleomycin lung injury and ispostulated to synergize with TNF-a to perpetuate inflammation
Figure 4. Human amnion epithelial cells (hAEC) injection reduces cytokine levels in the bleomycin-injured lung. (A) Analysis of mouse lung mRNA
transcripts by reverse transcriptase–polymerase chain reaction (qRT-PCR) showed an elevation in inflammatory and profibrotic cytokines 2 weeksafter bleomycin injury compared with saline–injected controls (P , 0.001); however, this response was attenuated by hAEC injection (*P , 0.01
comparing bleomycin with bleomycin 1 hAECs). (B) Gel electrophoresis of PCR products for human-specific IL-1, IL-10, tumor necrosis factor (TNF)-
a, transforming growth factor (TGF)-b, migration inhibition factor (MIF), and IFN-g. The primers specific for human cytokines did not cross-react
with mouse lung mRNA from animals given bleomycin alone. hAECs expressed mRNA for IL-1, IL-10, TNF-a, TGF-b, MIF, and IFN-g. We detectedlow levels of human-specific cytokine mRNA for IL-1, IL-10, TNF-a, and TGF-b in lungs of mice culled 2 weeks after hAEC transplantation using real-
time PCR and on gel electrophoresis of PCR products. However, mRNA for MIF and IFN-g were not detectable. B 5 bleomycin alone; BH 5
bleomycin 1 hAECs; H 5 hAECs. (C) Analysis of lung tissue homogenates showed that the increase in IL-1, IL-6, MCP-1, and TGF-b protein after
bleomycin was attenuated by hAEC injection (*P , 0.05).
648 AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE VOL 182 2010

and fibrosis (31). Therefore, the reduction in IL-6 after hAECinjection may serve to reduce further the development of fibrosis.TGF-b is a central cytokine in the pathogenesis of lung fibrosis(35). By Day 14 after bleomycin administration, TGF-b isincreased in macrophages, epithelial cells, fibroblasts, and myofi-broblasts (33). Khalil and coworkers (33) conclusively showedthat the amelioration of bleomycin fibrosis is dependent on theinhibition of TGF-b. As such, the significant down-regulation ofTGF-b also supports the antifibrotic role of hAECs. In addition,hAECs reduce fibrosis when injected after early (24 hourspostbleomycin) and during established injury (2 weeks postbleo-mycin), suggesting that hAEC therapy may be useful in lungdiseases with established fibrosis and tissue remodeling. Thedetection, albeit low levels, of human specific mRNA for IL-I,IL-10, TNF-a, and TGF-b suggests that hAECs engraft in thelung, remain viable, and are capable of synthesizing mRNA.However, despite the presence of human mRNA, there is a re-duction in protein levels for IL-1, TNF-a, and TGF-b after hAECinjection. Furthermore, it is unlikely that the detected low levelsof human IL-10 mRNA contributes significantly to the increase inIL-10 mRNA in the mouse lung. Therefore, we propose that theoverall antiinflammatory effect of hAECs is to modify theresponse of the mouse lung to injury.
MMPs and their endogenous inhibitors (TIMPs) are primarilyresponsible for the degradation of extracellular matrix proteins,such as collagen. Consistent with our findings TIMP concentra-tions increase before collagen deposition in the bleomycin mousemodel at 14–28 days after injury (36). Protein analysis of lungtissue lysates demonstrated an hAEC-induced increase in MMP-2 and a reduction in TIMP-1 and TIMP-2 protein, which otherstudies suggest would augment lung repair (21). In contrast, Ortizand colleagues (37) demonstrated down-regulation of MMP-2when using murine bone marrow MSCs to reduce the fibroticresponse to bleomycin injury. This difference may be caused bythe source of cells and the mouse strains (C57/Bl6 versus SCID)used.
The mechanisms delineating the salutary effects of stem cellson models of lung injury are largely unknown. However, studiesin MSCs have found that several different molecules may playa part in augmenting repair. MSCs reduced lung fibrosis byinhibition of IL-1 receptor antagonist (IL-1Ra) after bleomycin-induced lung injury (2). Furthermore, MSCs reduced endotoxin-induced lung injury by decreasing inflammation and cytokinescaused by the actions of angiopoeitin-1 (3, 38). In addition,hepatocyte growth factor reduced elastase-induced lung injuryby mobilizing endothelial progenitor cells to the lung (39). In an
Figure 5. Human amnion epithelial cells (hAECs) reduce lung fibrosis after bleomycin injury and alter matrix metalloproteinase (MMPs) and tissue
inhibitor of metalloproteinases (TIMPs). (A) hAEC injection resulted in a significant fall in collagen content compared with cellular controls placentaltrophoblasts and lung fibroblasts. hAECs injected into healthy mice did not induce a fibrotic response. hAECs also reduced well-developed fibrosis.
*P , 0.05 compared with saline controls; 1 P , 0.05 compared with bleomycin alone. (B) Active MMP-2 increased significantly in mice receiving
bleomycin and hAECs (*P , 0.05 versus bleomycin alone). (C) Compared with controls, TIMP mRNA expression is up-regulated in bleomycin-
injured mice (P , 0.001). However, hAEC treatment led to a reduction in TIMP-1, -3, and -4 mRNA expression (*P , 0.05). Reverse zymography oflung homogenates show that the increase in TIMP-1 and TIMP-2 after bleomycin instillation is reduced after hAEC injection (*P , 0.05).
Moodley, Ilancheran, Samuel, et al.: Amnion Epithelial Cells Augment Lung Repair 649

elegant study, Lee and coworkers (40) found that keratinocytegrowth factor was central to increasing alveolar fluid clearanceand reducing capillary leak after injection of human MSCs in anex vivo human lung model of LPS-induced lung injury. Althoughwe have not identified a specific molecule by which hAECsmediated lung repair, we suggest these cells act through severalmechanisms to reduce inflammation and fibrosis. The adoption ofa lung phenotype in vivo by hAECs would assist in restitutingalveolar epithelium, thereby reducing collagen deposition (41).In addition, the reduction in inflammation and cytokine expres-sion may be important mechanisms that limit damage to the lungand subsequent scarring. Furthermore, the elevation in MMPswith concurrent inhibition of TIMPs would constitute a prode-gradative environment for the breakdown of deposited collagen.
To date, there are no safe and effective therapeutic agents totreat chronic fibrotic lung diseases. The use of hAECs for thetreatment of lung disorders has distinct advantages because theyare highly abundant, obtained noninvasively, and have beenparentally administered into humans without adverse effects(14). In addition, there are no ethical controversies associatedwith the clinical use of these cells.
Author Disclosure: Y.M. has a pending patent PCT/AU2008000753. S.I. does nothave a financial relationship with a commercial entity that has an interest in thesubject of this manuscript. C.S. does not have a financial relationship witha commercial entity that has an interest in the subject of this manuscript. V.V.does not have a financial relationship with a commercial entity that has aninterest in the subject of this manuscript. D.A. does not have a financialrelationship with a commercial entity that has an interest in the subject of thismanuscript. E.D.W. received grant support from AstraZeneca for collaborativecontact research and from the NHMRC (more than $100,001). G.J. does not havea financial relationship with a commercial entity that has an interest in the subjectof this manuscript. E.W. does not have a financial relationship with a commercialentity that has an interest in the subject of this manuscript. A.T. does not havea financial relationship with a commercial entity that has an interest in the subjectof this manuscript. U.M. has a temporary patent.
Acknowledgment: The authors thank James Ngui for flow cytometry, Mark Malinand Jinhua Li for tail vein injections, Dhanya Sreedharan for DNA-PCR, GregoryAllen for FISH analysis, and A/Prof Phillip Bardin for providing human lungfibroblast cells.
References
1. Ware LB, Matthay MA. The acute respiratory distress syndrome. NEngl J Med 2000;342:1334–1349.
2. Ortiz LA, Dutreil M, Fattman C, Pandey AC, Torres G, Go K, PhinneyDG. Interleukin 1 receptor antagonist mediates the antiinflammatoryand antifibrotic effect of mesenchymal stem cells during lung injury.Proc Natl Acad Sci U S A 2007;104:11002–11007.
3. Mei SH, McCarter SD, Deng Y, Parker CH, Liles WC, Stewart DJ.Prevention of LPS-induced acute lung injury in mice by mesenchymalstem cells overexpressing angiopoietin 1. PLoS Med 2007;4:e269.
4. Baber SR, Deng W, Master RG, Bunnell BA, Taylor BK, Murthy SN,Hyman AL, Kadowitz PJ. Intratracheal mesenchymal stem celladministration attenuates monocrotaline-induced pulmonary hyper-tension and endothelial dysfunction. Am J Physiol Heart Circ Physiol2007;292:H1120–H1128.
5. Albera C, Polak JM, Janes S, Griffiths MJ, Alison MR, Wright NA,Navaratnarasah S, Poulsom R, Jeffery R, Fisher C, et al. Repopula-tion of human pulmonary epithelium by bone marrow cells: a potentialmeans to promote repair. Tissue Eng 2005;11:1115–1121.
6. Sueblinvong V, Loi R, Eisenhauer PL, Bernstein IM, Suratt BT, SpeesJL, Weiss DJ. Derivation of lung epithelium from human cord blood-derived mesenchymal stem cells. Am J Respir Crit Care Med 2008;177:701–711.
7. Carraro G, Perin L, Sedrakyan S, Giuliani S, Tiozzo C, Lee J, TurcatelG, De Langhe SP, Driscoll B, Bellusci S, et al. Human amniotic fluidstem cells can integrate and differentiate into epithelial lung lineages.Stem Cells 2008;26:2902–2911.
8. Cargnoni A, Gibelli L, Tosini A, Signoroni PB, Nassuato C, Arienti D,Lombardi G, Albertini A, Wengler GS, Parolini O. Transplantationof allogeneic and xenogeneic placenta-derived cells reduces bleomycin-induced lung fibrosis. Cell Transplant 2009;18:405–422.
9. Moodley Y, Atienza D, Manuelpillai U, Samuel CS, Tchongue J,Ilancheran S, Boyd R, Trounson A. Human umbilical cord mesen-
chymal stem cells reduce fibrosis of bleomycin-induced lung injury.Am J Pathol 2009;175:303–313.
10. Moore BB, Hogaboam CM. Murine models of pulmonary fibrosis. Am J
Physiol Lung Cell Mol Physiol 2008;294:L152–L160.11. Miki T, Lehmann T, Cai H, Stolz DB, Strom SC. Stem cell character-
istics of amniotic epithelial cells. Stem Cells 2005;23:1549–1559.12. Ilancheran S, Michalska A, Peh G, Wallace EM, Pera M, Manuel-
pillai U. Stem cells derived from human fetal membranes displaymulti-lineage differentiation potential. Biol Reprod 2007;77:577–588.
13. Sakuragawa N, Thangavel R, Mizuguchi M, Hirasawa M, Kamo I.
Expression of markers for both neuronal and glial cells in humanamniotic epithelial cells. Neurosci Lett 1996;209:9–12.
14. Ilancheran S, Moodley Y, Manuelpillai U. Human fetal membranes:
a source of stem cells for tissue regeneration and repair? Placenta2009;30:2–10.
15. Chaffer CL, Brennan JP, Slavin JL, Blick T, Thompson EW, Williams
ED. Mesenchymal-to-epithelial transition facilitates bladder cancermetastasis: role of fibroblast growth factor receptor-2. Cancer Res2006;66:11271–11278.
16. Ashcroft T, Simpson JM, Timbrell V. Simple method of estimating
severity of pulmonary fibrosis on a numerical scale. J Clin Pathol1988;41:467–470.
17. Szapiel SV, Elson NA, Fulmer JD, Hunninghake GW, Crystal RG.
Bleomycin-induced interstitial pulmonary disease in the nude, athy-mic mouse. Am Rev Respir Dis 1979;120:893–899.
18. Jeyabalan A, Kerchner LJ, Fisher MC, McGuane JT, Doty KD, Conrad
KP. Matrix metalloproteinase-2 activity, protein, mRNA, and tissueinhibitors in small arteries from pregnant and relaxin-treated non-pregnant rats. J Appl Physiol 2006;100:1955–1963.
19. Hewitson TD, Mookerjee I, Masterson R, Zhao C, Tregear GW, Becker
GJ, Samuel CS. Endogenous relaxin is a naturally occurring modu-lator of experimental renal tubulointerstitial fibrosis. Endocrinology2007;148:660–669.
20. Ali NN, Edgar AJ, Samadikuchaksaraei A, Timson CM, Romanska HM,
Polak JM, Bishop AE. Derivation of type II alveolar epithelial cellsfrom murine embryonic stem cells. Tissue Eng 2002;8:541–550.
21. Yaguchi T, Fukuda Y, Ishizaki M, Yamanaka N. Immunohistochemical
and gelatin zymography studies for matrix metalloproteinases inbleomycin-induced pulmonary fibrosis. Pathol Int 1998;48:954–963.
22. Minoo P, Hu L, Xing Y, Zhu NL, Chen H, Li M, Borok Z, Li C. Physical
and functional interactions between homeodomain NKX2.1 andwinged helix/forkhead FOXA1 in lung epithelial cells. Mol Cell Biol2007;27:2155–2165.
23. Berger MJ, Adams SD, Tigges BM, Sprague SL, Wang XJ, Collins DP,
McKenna DH. Differentiation of umbilical cord blood-derived multi-lineage progenitor cells into respiratory epithelial cells. Cytotherapy2006;8:480–487.
24. Warburton D, Zhao J, Berberich MA, Bernfield M. Molecular embry-
ology of the lung: then, now, and in the future. Am J Physiol 1999;276:L697–L704.
25. Baritussio A, Bellina L, Carraro R, Rossi A, Enzi G, Magoon MW,
Mussini I. Heterogeneity of alveolar surfactant in the rabbit: compo-sition, morphology, and labelling of subfractions isolated by centri-fugation of lung lavage. Eur J Clin Invest 1984;14:24–29.
26. Brownfoot FC, Crowther CA, Middleton P. Different corticosteroids
and regimens for accelerating fetal lung maturation for women at riskof preterm birth. Cochrane Database Syst Rev 2008;4:CD006764.
27. Kasper M, Gunthert U, Dall P, Kayser K, Schuh D, Haroske G, Muller
M. Distinct expression patterns of CD44 isoforms during human lungdevelopment and in pulmonary fibrosis. Am J Respir Cell Mol Biol1995;13:648–656.
28. Salaun B, de Saint-Vis B, Pacheco N, Pacheco Y, Riesler A, Isaac S,
Leroux C, Clair-Moninot V, Pin JJ, Griffith J, et al. CD208/dendriticcell-lysosomal associated membrane protein is a marker of normaland transformed type II pneumocytes. Am J Pathol 2004;164:861–871.
29. Makker SP, Kanalas JJ, Tio FO, Kotas RV. Definition of an immuno-
logic marker for type II pneumocytes. J Immunol 1989;142:2264–2269.30. Segel MJ, Izbicki G, Cohen PY, Or R, Christensen TG, Wallach-Dayan
SB, Breuer R. Role of interferon-gamma in the evolution of murinebleomycin lung fibrosis. Am J Physiol Lung Cell Mol Physiol 2003;285:L1255–L1262.
31. Smith RE, Strieter RM, Phan SH, Lukacs N, Kunkel SL. TNF and IL-6
mediate MIP-1alpha expression in bleomycin-induced lung injury.J Leukoc Biol 1998;64:528–536.
650 AMERICAN JOURNAL OF RESPIRATORY AND CRITICAL CARE MEDICINE VOL 182 2010

32. Kradin RL, Sakamoto H, Jain F, Zhao LH, Hymowitz G, Preffer F. IL-10 inhibits inflammation but does not affect fibrosis in the pulmonaryresponse to bleomycin. Exp Mol Pathol 2004;76:205–211.
33. Khalil N, Parekh TV, O’Connor RN, Gold LI. Differential expression oftransforming growth factor-beta type I and II receptors by pulmonarycells in bleomycin-induced lung injury: correlation with repair andfibrosis. Exp Lung Res 2002;28:233–250.
34. Gurujeyalakshmi G, Wang Y, Giri SN. Taurine and niacin block lunginjury and fibrosis by down-regulating bleomycin-induced activationof transcription nuclear factor-kappaB in mice. J Pharmacol Exp Ther2000;293:82–90.
35. Khalil N, Bereznay O, Sporn M, Greenberg AH. Macrophage pro-duction of transforming growth factor beta and fibroblast collagensynthesis in chronic pulmonary inflammation. J Exp Med 1989;170:727–737.
36. Oggionni T, Morbini P, Inghilleri S, Palladini G, Tozzi R, Vitulo P,Fenoglio C, Perlini S, Pozzi E. Time course of matrix metallopro-teases and tissue inhibitors in bleomycin-induced pulmonary fibrosis.Eur J Histochem 2006;50:317–325.
37. Ortiz LA, Gambelli F, McBride C, Gaupp D, Baddoo M, Kaminski N,Phinney DG. Mesenchymal stem cell engraftment in lung is enhancedin response to bleomycin exposure and ameliorates its fibrotic effects.Proc Natl Acad Sci USA 2003;100:8407–8411.
38. Xu J, Qu J, Cao L, Sai Y, Chen C, He L, Yu L. Mesenchymal stem cell-based angiopoietin-1 gene therapy for acute lung injury induced bylipopolysaccharide in mice. J Pathol 2008;214:472–481.
39. Ishizawa K, Kubo H, Yamada M, Kobayashi S, Suzuki T, Mizuno S,Nakamura T, Sasaki H. Hepatocyte growth factor induces angiogen-esis in injured lungs through mobilizing endothelial progenitor cells.Biochem Biophys Res Commun 2004;324:276–280.
40. Lee JW, Fang X, Gupta N, Serikov V, Matthay MA. Allogeneic humanmesenchymal stem cells for treatment of E. coli endotoxin-inducedacute lung injury in the ex vivo perfused human lung. Proc Natl AcadSci U S A 2009;106:16357–16362.
41. Sisson TH, Mendez M, Choi K, Subbotina N, Courey A, Cunningham A,Dave A, Engelhardt JF, Liu X, White ES, et al. Targeted injury oftype II alveolar epithelial cells induces pulmonary fibrosis. Am JRespir Crit Care Med 2010;181:254–263.
Moodley, Ilancheran, Samuel, et al.: Amnion Epithelial Cells Augment Lung Repair 651




















