
How do global and local geometries shape exploratory behavior in rats?
9
Available online at www.sciencedirect.com Behavioural Brain Research 187 (2008) 334–342 Research report How do global and local geometries shape exploratory behavior in rats? Osnat Yaski, David Eilam ∗ Department of Zoology, Tel-Aviv University, Ramat Aviv 69978, Israel Received 4 July 2007; received in revised form 18 September 2007; accepted 21 September 2007 Available online 29 September 2007 Abstract The present study examines how rats process object information in relation to other objects as well as to the global shape of an enclosure. Rats were introduced into either a round or a square arena with various arrays of freestanding portable corners (objects) that differed in spacing, orientation, and number. We found that the time spent at the objects was substantially higher in the round compared with the square arena. Rats in the square arena distributed their time evenly between arena perimeter and objects; however, they visited the objects more frequently than the perimeter. Rats tested in either round or square arena favored staying at the interior of the object regardless of its orientation in the arena. Finally, only geometric changes in the object array affected the rats’ level of activity. These findings demonstrate that the rats were able to detect the geometry of the enclosure, the array, and the objects. We suggest that the context of the global enclosure geometry affects the meaning of landmarks (objects) for the observer, which in turn results in a different distribution of activity. Specifically, an object’s local geometry served mainly in the context of a safe place in the round arena, as manifested in extended duration of stay at the objects, however in the square arena served mainly as a place of interest, as manifested in the frequent visits, but without their staying there. Thus, the geometries of the enclosure, the object array, and the discrete objects, together shape exploratory behavior in rats. © 2007 Elsevier B.V. All rights reserved. Keywords: Exploration; Geometric module; Landmark; Array; Open field; Home base; Spatial cognition; Spatial representation 1. Introduction The notion that numerous vertebrate species use the geometry of the surrounding environment to orient in space [3] assumes the existence of a ‘geometric module’ in animals’ spatial represen- tation. According to this notion, animals first encode geometric properties whereas non-geometric information is secondary and subordinate in this processing mode [3]. A theoretical mech- anism for how geometry is encoded and used is the ‘global matching process’, in which a principal axis of space is encoded without encoding the entire shape [14]. This mechanism was further refined with the suggestion that specific axes, for exam- ple, symmetry axes and medial axes, might be encoded [4,5]. The global matching process of encoding internal coordinates in space constrains the use of featural cues and prevents over- load of spatial information [6,14]. Such geometric processing of environmental information has been further supported by neu- rological evidence: e.g., hippocampal place cells of rats tested in ∗ Corresponding author. Tel.: +972 3 6406471; fax: +972 3 6406988. E-mail address: [email protected] (D. Eilam). two different environments, a round and a square arenas, were shown to be sensitive to the environment’s geometry [24]. Prominent landmarks or key locations in the environment can be important for animals in organizing their locomotor activity [7,8,18,21]. An example for a behavior organized in reference to a key location is the formation of a ‘home base’ in rodents, which is the place where they stay for relatively long periods and from which they set out on round trips in the environment [12]. When a shelter is available, the home base is located at it [7,10,31], but without a shelter rodents tend to establish the home base close to a salient landmark [7,8,21]. In a square empty open field, the home base is typically established at one of the arena corners [1,10–12], probably because these are the most distinct environmental features and therefore easy to recognize. Arena corners may also be perceived as a relatively protected area, a notion supported by the finding that the home base is where rats tend to groom and crouch [12]. In examining which are the physical properties of corners that shape behavior, rats were challenged with additional freestanding portable corners placed inside the arena [37]. The routes of these rats converged to the additional corner placed against the arena wall, testifying about the prominence of the added corner against the background 0166-4328/$ – see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.bbr.2007.09.027
Transcript of How do global and local geometries shape exploratory behavior in rats?

A
RoitFtlmso©
K
1
oetpsamwfpTiler
0d
Available online at www.sciencedirect.com
Behavioural Brain Research 187 (2008) 334–342
Research report
How do global and local geometries shape exploratory behavior in rats?
Osnat Yaski, David Eilam ∗Department of Zoology, Tel-Aviv University, Ramat Aviv 69978, Israel
Received 4 July 2007; received in revised form 18 September 2007; accepted 21 September 2007Available online 29 September 2007
bstract
The present study examines how rats process object information in relation to other objects as well as to the global shape of an enclosure.ats were introduced into either a round or a square arena with various arrays of freestanding portable corners (objects) that differed in spacing,rientation, and number. We found that the time spent at the objects was substantially higher in the round compared with the square arena. Ratsn the square arena distributed their time evenly between arena perimeter and objects; however, they visited the objects more frequently thanhe perimeter. Rats tested in either round or square arena favored staying at the interior of the object regardless of its orientation in the arena.inally, only geometric changes in the object array affected the rats’ level of activity. These findings demonstrate that the rats were able to detect
he geometry of the enclosure, the array, and the objects. We suggest that the context of the global enclosure geometry affects the meaning ofandmarks (objects) for the observer, which in turn results in a different distribution of activity. Specifically, an object’s local geometry served
ainly in the context of a safe place in the round arena, as manifested in extended duration of stay at the objects, however in the square arenaerved mainly as a place of interest, as manifested in the frequent visits, but without their staying there. Thus, the geometries of the enclosure, thebject array, and the discrete objects, together shape exploratory behavior in rats.
2007 Elsevier B.V. All rights reserved.
e ba
ts
b[twa[ihoadA
eywords: Exploration; Geometric module; Landmark; Array; Open field; Hom
. Introduction
The notion that numerous vertebrate species use the geometryf the surrounding environment to orient in space [3] assumes thexistence of a ‘geometric module’ in animals’ spatial represen-ation. According to this notion, animals first encode geometricroperties whereas non-geometric information is secondary andubordinate in this processing mode [3]. A theoretical mech-nism for how geometry is encoded and used is the ‘globalatching process’, in which a principal axis of space is encodedithout encoding the entire shape [14]. This mechanism was
urther refined with the suggestion that specific axes, for exam-le, symmetry axes and medial axes, might be encoded [4,5].he global matching process of encoding internal coordinates
n space constrains the use of featural cues and prevents over-
oad of spatial information [6,14]. Such geometric processing ofnvironmental information has been further supported by neu-ological evidence: e.g., hippocampal place cells of rats tested in∗ Corresponding author. Tel.: +972 3 6406471; fax: +972 3 6406988.E-mail address: [email protected] (D. Eilam).
awawpta
166-4328/$ – see front matter © 2007 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2007.09.027
se; Spatial cognition; Spatial representation
wo different environments, a round and a square arenas, werehown to be sensitive to the environment’s geometry [24].
Prominent landmarks or key locations in the environment cane important for animals in organizing their locomotor activity7,8,18,21]. An example for a behavior organized in referenceo a key location is the formation of a ‘home base’ in rodents,hich is the place where they stay for relatively long periods
nd from which they set out on round trips in the environment12]. When a shelter is available, the home base is located att [7,10,31], but without a shelter rodents tend to establish theome base close to a salient landmark [7,8,21]. In a square emptypen field, the home base is typically established at one of therena corners [1,10–12], probably because these are the mostistinct environmental features and therefore easy to recognize.rena corners may also be perceived as a relatively protected
rea, a notion supported by the finding that the home base ishere rats tend to groom and crouch [12]. In examining which
re the physical properties of corners that shape behavior, rats
ere challenged with additional freestanding portable cornerslaced inside the arena [37]. The routes of these rats convergedo the additional corner placed against the arena wall, testifyingbout the prominence of the added corner against the background
Brain
oewmbf
meeaaotpwvisrte
2
2
w((pc7a(
2
(rtimtiItGoap
2
(owef
aabr
(oTapt
2
tfiiadaicc4eodo
‘
12
3
4
56
2
wtovt
3
3t
aHr
O. Yaski, D. Eilam / Behavioural
f the arena enclosure. That is, it was suggested that the ratsncoded the geometry of the enclosure with the corners andalls perceived as one entity and not as a set of discrete land-arks. The distinctness of the added corner was thus due to it
eing ‘pasted’ on a geometric module that served as a featurelessrame [37].
Natural environments are likely to provide arrays of land-arks whereas experimental set-ups usually comprise an
nclosure. Therefore, one might expect that a mechanism forncoding geometry will not be restricted to enclosures [6,29]nd that rats are able to use geometry inferred from an objectrray [2]. Nevertheless, recognition of an object either as a partf an array or as an independent object may also depend onhe context or the observer’s view [22]. In light of this, theresent study examined how object information is processedith respect to other objects and to the global information pro-ided by the shape of the enclosure. Specifically, we placed ratsn either a round or a square arena with a geometric array of free-tanding corners (objects), and subsequently changed the localelationships among the objects in order to study the encoding ofhe geometry of the individual objects, the array, and the overallnclosure.
. Materials and methods
.1. Animals
Adult male Long Evans hooded rats (n = 17; 3–5 months old;eight > 200 g) were housed in groups of 3–4 individuals in metal cages
60 cm × 40 cm × 25 cm) with sawdust bedding, in an air-conditioned room24 ± 2 ◦C) under 12/12 light/dark cycle (lights on 8:00–20:00). Standard ratellets and water were provided ad lib. Rats were individually marked byolored ink on the tail and were each handled for 15 min/day during the
days before testing. At all times, animals were maintained and treatedccording to the institutional guidelines for animal care and use in researchpermit L-06-015).
.2. Apparatus
Testing was carried out in two arenas: (1) a square open field200 cm × 200 cm) enclosed with 50 cm-high opaque Plexiglas walls; and (2) aound open field (200 cm diameter) enclosed with 50 cm-high opaque flexiblein wall. Both arenas were placed in an air-conditioned room (22 ◦C), centeredn the same location and thus providing the same distal room panorama (e.g.
onitor, closet and air-conditioner) and same blue PVC floor. During testing,he room was illuminated by three 300 W light bulbs directed to the white ceil-ng, providing diffused illumination of the arena. A video camera (Ikegami B/WCD-47E) was placed 2.5 m above the center of the arena, providing a top viewhat was recorded onto a computer by means of a video grabber (VideoHomerabBeeX) as MPEG1 digital file. According to the specific array setting, fourr three objects were placed in the open field. Each of the objects was shaped ascorner made of two orthogonal (20 cm wide and 50 cm high) opaque Plexiglaslates.
.3. Procedure
Rats were randomly assigned to a group that was tested either in the square
n = 8), or the round arena (n = 9). In each arena, rats were randomly exposednce to each of the five arrays (Fig. 1A) over 5 consecutive days. The five arraysere based on changing the spacing, the position and the number of objects inither the square or round arena, as follows: array 1 – empty arena; array 2 –our objects facing the arena center, arrayed in a square and distanced 66 cm
spt(
Research 187 (2008) 334–342 335
part; array 3 – four adjacent objects facing the arena center, arrayed in a squarend spaced 26 cm apart; array 4 – same as array 2, but the objects were rotatedy 180◦ to face the arena walls; array 5 – as in array 4, but with top-right objectemoved, thus forming a triangle.
At the beginning of each test session, a rat was placed at a fixed ‘start point’marked by asterisk in Fig. 1A), facing the center of the arena. The behaviorf the rat was then recorded for 15 min, after which it was returned to its cage.he arena was cloth-wiped with detergent and adjusted to the next randomlyssigned array. All rats were tested over five consecutive days during the lighthase with each animal randomly tested once a day at a different time in ordero reduce biological cyclic rhythm effect.
.3.1. Data acquisition and analysisVideo files were analyzed using ‘Ethovision’ (by Noldus, NL), which tracks
he progression of the rat in the arena, providing the time and location of the ratve times per second. For analysis, arenas were divided so that each division
n the round arena comprised the same percentage of the area of the squarerena, enabling parallel comparison between the arenas. A perimeter area wasefined as a strip section (13 and 14 cm along the arena walls in round and squarerena, respectively; Fig. 1B). The perimeter was further divided into eight zonesdentical in area, one of which, the start zone (marked by asterisk in Fig. 1B) wasentered at the fixed start point. Object zones comprised each one of the portableorners, and were defined as a square area (40 cm × 40 cm in round arena and5 cm × 45 cm in square arena; Fig. 1B). The total area of four object zonesqualed the area of the perimeter, enabling a comparison between activity atbjects and activity at arena perimeter. Each object zone was further subdividediagonally into interior object sub-zone (the inside of the corner) and exteriorbject sub-zone (Fig. 1B).
The following parameters were extracted from ‘Ethovision’ and exported toMicrosoft Excel’ program for further analyses:
. Travel distance: overall metric distance (m) that a rat traveled in the arena.
. Travel speed: average velocity (m/s) of a rat, measured during motion seg-ments.
. Number of stops: incidence of stops in the arena or in defined arena area. Astop was defined as the absence of forward movement of the rat for at least1 s or longer.
. Duration of stops: total duration of a stop in the arena or in defined arenaarea.
. Time spent: total duration of stay (s) in defined arena area.
. Number of visits: the incidence of entering a defined arena area.
.4. Statistics
Data of the test groups were compared by repeated-measure ANOVA, inhich the between-group factor was arena shape (‘square’ or ‘round’ arena) and
he within-group factor was the different arrays. In several comparisons, onef the following additional within-group factors was also included: (1) interiorersus exterior object sub-zones; (2) perimeter area versus object zones. ANOVAests were followed by Tukey HSD test or Dunnett test.
. Results
.1. Overall activity in the arena, at the perimeter, and athe start zone
Arena shape and the various object arrays did not affect over-ll activity in the arena (Table 1, third and fourth columns, top).owever, they affected activity at the perimeter area, with the
ats moving mainly along the arena walls in both round and
quare empty arenas (Fig. 2, array 1). But, when objects wereresent, rats were attracted to them, moving from one object tohe next, along the walls, and between the objects and wallsFig. 2, arrays 2–5). The rats spend significantly more time
336 O. Yaski, D. Eilam / Behavioural Brain Research 187 (2008) 334–342
Fig. 1. (A) Rats were tested in either a round (top) or square (bottom) arena in five arrays (columns). An asterisk at the bottom of each arena-sketch representsthe ‘start point’ at which rats were introduced into the arena at the beginning of each test. In array 1, rats were tested in an empty arena. In array 2, four portablecorners (objects) were distanced from each other in a square array with the interior of each corner facing the arena center. In array 3, the four portable corners werespaced more closely than in array 2, with each corner interior facing the arena center. In array 4, the portable corners in the square array were each rotated so thatthe interior faced the arena walls. In array 5, the top right portable corner was removed, changing the object array from square to triangular. Rats in each groupwere tested only once in each array in a random order. (B). The division of arenas is depicted by the grey dashed line in round (top) and square (bottom) arena. Ineach arena, a strip along the walls was defined as the perimeter area. This was further divided into eight identical perimeter zones, with the start zone marked by anasterisk, indicating where rats were placed at the beginning of each test. A square around each of the objects (portable corners) was defined as the object zone andw ct zone ed the
awtppc
ia
pzcas
as relocated according to the location of the object in each array setting. Objexterior object sub-zone. Each defined area in round and square arenas compris
nd paid more stops at the perimeter of the square comparedith the round arena (Table 1, third column, top two rows in
he center). Time spent and incidence of stops and visits at theerimeter were higher in empty arenas (round or square) com-ared with arenas with added objects (Table 1, fourth column,
enter).Arena shape and the various object arrays also affected activ-ty at the start zone, which constitutes a part of the perimeterrea. In addition, the start zone had a polarizing effect on other
ttbc
es were further subdivided by a diagonal line into interior object sub-zone andsame percentage of arena area in order to allow parallel comparisons.
erimeter zones in terms of time spent and visits paid to theseones. Rats spent a longer duration at the start zone of the roundompared with the square arena (Table 1, third column, top rowt the bottom). Object arrays had a significant effect on the timepent and the number of stops and visits at the start zone, with all
hese three parameters being higher in the round compared withhe square arena (Table 1, fourth column, bottom). Interactionsetween arena shape and object arrays were mostly insignifi-ant, except for the time spent at the start zone (Table 1, fifth
O. Yaski, D. Eilam / Behavioural Brain Research 187 (2008) 334–342 337
Table 1Activity in the arena, at the perimeter, and in the start zone
Round arena(mean ± S.E.M.)
Square arena(mean ± S.E.M.)
Effect ofarena shape(F1,15; p)
Effect ofobject arrays(F4,60; p)
Interactionarena × arrays(F4,60; p)
All over the arenaTraveled distance (m) 96.98 ± 8.80 108.91 ± 11.79 0.97; 0.339 1.19; 0.323 0.52; 0.723Travel speed (m/s) 11.16 ± 0.99 12.40 ± 1.38 0.77; 0.394 1.58; 0.192 0.57; 0.682Number of stops 96.56 ± 8.40 100.45 ± 10.50 0.15; 0.706 6.11; <0.001a 0.03; 0.999Duration of stops (s) 641.40 ± 30.84 607.60 ± 36.60 0.83; 0.376 1.43; 0.235 0.36; 0.833
At the perimeterTime spent (s) 329.68 ± 50.27 465.66 ± 55.78 5.00; 0.041 48.51; <0.0001b 1.65; 0.174Number of stops 32.69 ± 3.79 41.43 ± 4.31 8.46; 0.011 27.29; <0.0001b 0.32; 0.863Number of visits 37.89 ± 4.71 36.93 ± 4.40 0.04; 0.839 3.18; 0.019b 1.52; 0.208
At the start zoneTime spent (s) 129.23 ± 35.20 57.00 ± 9.81 16.76; <0.001 5.79; <0.001 3.59; 0.011Number of stops 11.62 ± 1.99 8.65 ± 1.49 3.52; 0.080 8.25; <0.0001 1.71; 0.159Number of visits 17.73 ± 2.93 14.40 ± 1.95 1.63; 0.221 6.40; <0.0003 1.24; 0.305
p-values below α = 0.05 are depicted in boldface numerals.a Tukey HSD analysis revealed this difference to be between arrays 3 and 4 (Fig. 1A).b Tukey HSD analysis revealed these differences to be between empty arenas compared with all other arrays with objects (Fig. 1A).
Fig. 2. Routes of progression of a representative rat in the round (top) and square (bottom) arenas and in each array (columns) for the entire 15 min session. In thee d squac arenas
cswRctrtevb
3
wb(a
mpty arena (array 1), routes were mainly along the arena walls in both round anonnecting them, and extending along the arena walls and between objects andimilar in both round and square arenas.
olumn). In the round arena, duration and number of visits at thetart zone compared with the other perimeter zones decreasedith the increase of distance from the start zone (Fig. 3, top).ats in the square arena spent the longest duration at an arenaorner adjacent to the start zone (Fig. 3, bottom). The preferenceo this specific arena corner was probably dictated by the distaloom panorama and the close proximity of this arena corner to
he start point. Time at other perimeter zones was biased towardxtended stay at other arena corners. However, the number ofisits to the start zone and its adjacent corner were similar (Fig. 3,ottom).tutt
re arenas. With objects (arrays 2–5), routes converged mainly upon the objects,walls. This locomotor pattern was maintained in all object arrays and seemed
.2. Activity at object zones
Arena shape affected the cumulative time spent at the objects,hich was higher in the round compared with the square arena,ut did not affect the number of visits and number of stopsTable 2, third column). In contrast, the different arrays did notffect the total time spent at the array, but significantly affected
he number of visits and stops to object zones (Table 2, fourth col-mn). The total time spent at the array was preserved even whenhe number of objects was reduced from four to three, implyinghat rats compensated for the absence of one object by extending
338 O. Yaski, D. Eilam / Behavioural Brain Research 187 (2008) 334–342
Fig. 3. Time spent (mean ± S.E.M.; black columns) and visits (mean ± S.E.M.;white columns) in each of the eight perimeter zones in round (top) and square(bottom) arenas. The location of each zone is sketched at the bottom of eachset of columns in both graphs. As shown, time spent and visits were highestin the start zone in the round arena, and declined in the other zones accordingto their distance from the start. In the square arena most time was spent at acorner adjacent to the start zone, whereas the number of visits to that corner wassimilar to that of the start zone. Time spent at other perimeter zones was biasedtoward extended duration of stay at the other arena corners. A two-way ANOVAwith repeated measures of time spent revealed a significant within-group fac-tor (F7,56 = 33.41; p < 0.0001), which in a subsequent Dunnett comparison wasfound to differ between the start zone and other perimeter zones. The interactionbetween arena shape and zones was also significant (F7,56 = 15.78; p < 0.0001),as was the difference in the square arena between the start zone and the adjacentarena corner. A two-way ANOVA with repeated measures for the number of vis-its revealed a significant within-group factor (F7,56 = 167.14; p < 0.0001), whichin a subsequent Dunnett comparison was found to be between the start zone andthe other perimeter zones. The interaction between arena shape and zones wasalso significant (F7,56 = 8.45; p < 0.005) as was the difference between the startzone and all other perimeter zones in both the round and square arenas, withthe exception that in the square arena there was no difference in time spent andvisits between the start zone and the adjacent arena corner.
Table 3Time spent at the interior and exterior object sub-zones
Time spent (s) Round arena Square arena
Interior sub-zone 345.74 ± 41.36 220.46 ± 28.55Exterior sub-zone 205.73 ± 22.77 177.26 ± 26.73
Factor F(d.f.) p
Arena shape 6.54(1,15) 0.022Object arrays 0.72(3,45) 0.542Arena × arrays 0.66(3,45) 0.580Sub-zones 41.15(1,15) <0.0001Sub-zones × arena 11.49(1,15) 0.004a
Arrays × sub-zones 12.87(3,45) <0.0001b
Arrays × sub-zones × arena 0.55(3,45) 0.650
p-values below α = 0.05 are depicted in boldface numerals.a Tukey HSD analysis revealed a significant difference between time spent at
the interior and exterior sub-zones in the round arena but not in the square arena.
a
to5otttp
3
mvinra5o
3
t
Table 2Activity at object zones
Round arena(mean ± S.E.M.)
Square arena(mean ± S.E.M.)
Effect(F1,15;
Objects zonesTime spent (s) 551.53 ± 50.52 397.73 ± 49.64 6.54; 0Number of visits 81.25 ± 8.65 63.56 ± 9.38 2.75; 0Number of stops 59.42 ± 5.76 56.13 ± 7.17 0.19; 0
p-values below α = 0.05 are depicted in boldface numerals.a Tukey HSD analysis revealed a significant difference between array 3 and 4 (Fig.b Tukey HSD analysis revealed the difference to be between arrays 4 and 5 (Fig. 1A
b Tukey HSD analysis revealed the difference between sub-zones to be inrrays 3–5 (Fig. 1A).
heir stay at the remaining three. Specifically, average durationf stay at an object in the three-object array (Fig. 1A; array) was longer than at four-object arrays (151.6 ± 16.7 s at eachf the three objects, 120.3 ± 12.5 s at each of the four objects;wo-way ANOVA with repeated measures, within-group fac-or, F3,45 = 9.93; p < 0.0001; significant difference between thehree- and four-object arrays in a subsequent Tukey HSD com-arison).
.3. Activity at the interior and exterior object sub-zones
As described in Section 2, objects were freestanding cornersade of two perpendicular plates. Each object zone was subdi-
ided into an interior and an exterior zone (Fig. 1B). As shownn Fig. 4, time spent at the interior of these corners was sig-ificantly longer than that spent at the exterior (Table 3). Theesult was consistent even when the objects were rotated (Fig. 4,rrays 4 and 5) or when one object was removed (Fig. 4, array) implying that the rats had a general preference for the interiorf the objects, regardless of their orientation or number.
.4. Activity at perimeter area compared with object zones
As described above, rats spent a greater amount of time athe perimeter of the square compared with the round arena.
of arena shapep)
Effect of object arrays(F3,45; p)
Interaction arena × arrays(F3,45; p)
.022 0.72; 0.543 0.66; 0.580
.118 14.06; <0.0001a 1.46; 0.238
.668 3.39; 0.026b 0.85; 0.474
1A) and a significant difference between array 5 and all other arrays (Fig. 1A).).

O. Yaski, D. Eilam / Behavioural Brain
Fig. 4. Time spent (mean ± S.E.M.) in the interior (black columns) and exterior(white columns) sub-zones of the portable corner (object), in round (top) andsquare (bottom) arenas. A sketch of object array is depicted at the bottom ofeach set of columns. As shown, time spent at the interior was longer than that atthe exterior, and this difference was greater in the round arena. The differenceb(
AceatmT
4
b[evsahaotevoeIfopris‘Stp
4
tby it. The term was coined two decades ago, when rats were
TA
A
PO
F
AOAAAAA
p
etween interior and exterior sub-zone was preserved when corners were rotatedarrays 4 and 5).
ccordingly, they spent more time at the objects in the roundompared with the square arena (Fig. 5 and Table 4) at thexpense of duration at the perimeter, whereas rats in the squarerena distributed their time evenly between objects and perime-
er. However, in both round and square arenas, visits wereore frequent to the objects than to the perimeter (Fig. 5 andable 4).
sog
able 4ctivity at the perimeter compared with object zones
rea Time spent (s)
Round arena Square aren
erimeter 255.50 ± 49.75 411.44 ± 5bject zones 545.74 ± 50.62 392.13 ± 4
actor F(d.f.)
rena shape 0.01(1,15)
bject arrays 1.67(3,45)
rena × arrays 0.89(3,45)
rea 4.70(1,15)
rea × arena 6.14(1,15)
rrays × area 0.83(3,45)
rrays × area × arena 0.72(3,45)
-values below α = 0.05 are depicted in boldface numerals.a Tukey HSD analysis revealed a significant difference between arrays 5 and arraysb Tukey HSD analysis revealed a significant difference between time spent in perim
Research 187 (2008) 334–342 339
. Discussion
The spatial organization of locomotor behavior is affectedy the physical and geometric properties of the environment26]. In order to reveal the effect of local and global geom-try, rats were tested in a round or a square open field witharious arrays of objects. We found that in both a round and aquare empty arena, rats stayed mainly at the perimeter, near therena walls. When objects were placed in either type of arena,owever, the rats traded activity at the perimeter for activityt the objects, regardless of object array. The time spent at thebjects was substantially higher in the round compared withhe square arena. Rats in the square arena distributed their timevenly between perimeter and objects; however their numerousisits to the objects indicate that these had become focal pointsf greater interest than the arena corners. No apparent differ-nce in activity was observed among the different object arrays.n terms of this local geometry of the corner-shaped objects, ratsavored staying in the interior regardless of the orientation of thebject within the arena, thus indicating a preference for the morerotected object region. Finally, in the round arena the rats wereepeatedly attracted to visit and stay at the start zone, whereasn the square arena they stayed more at an arena corner near thetart zone. In the following discussion, we suggest that the ratspaste’ the local geometry of objects onto the global geometry.pecifically, the global geometric shape of the enclosure and
hat of the object array is detected by the rats and affects theirerception of distinctness and meaning of the objects.
.1. Detecting the global enclosure geometry
The term ‘geometric module’ refers to the ability of animalso encode the global geometry of the environment and orient
hown to be able to find a hidden goal in reference to the shapef the environment [3]. Since then, evidence of encoding theeometry of enclosures was found in young chickens [34,35],
Number of visits
a Round arena Square arena
9.49 36.69 ± 4.02 34.88 ± 4.099.02 86.33 ± 9.26 67.76 ± 9.89
p F(d.f.) p
0.903 1.77(1,15) 0.2030.187 5.29(3,45) 0.003a
0.453 2.03(3,45) 0.1230.047 102.22(1,15) <0.00010.026b 4.80(1,15) 0.0450.482 34.38(3,45) <0.00010.543 0.99(3,45) 0.405
2 and 3 (Fig. 1A).eter and object zones in the round arena but not in the square arena.
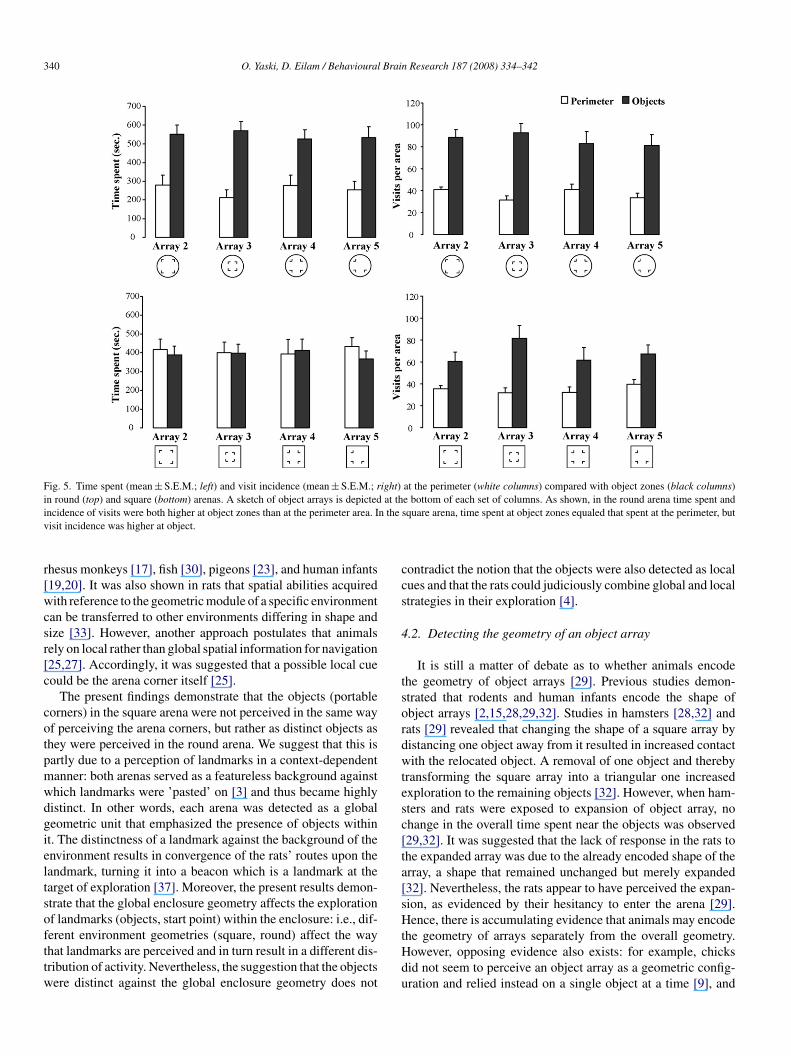
340 O. Yaski, D. Eilam / Behavioural Brain Research 187 (2008) 334–342
Fig. 5. Time spent (mean ± S.E.M.; left) and visit incidence (mean ± S.E.M.; right) at the perimeter (white columns) compared with object zones (black columns)in round (top) and square (bottom) arenas. A sketch of object arrays is depicted at the bottom of each set of columns. As shown, in the round arena time spent andi n thev
r[wcsr[c
cotpmwdgieltsofttw
ccs
4
tsordwtesc[ta[sH
ncidence of visits were both higher at object zones than at the perimeter area. Iisit incidence was higher at object.
hesus monkeys [17], fish [30], pigeons [23], and human infants19,20]. It was also shown in rats that spatial abilities acquiredith reference to the geometric module of a specific environment
an be transferred to other environments differing in shape andize [33]. However, another approach postulates that animalsely on local rather than global spatial information for navigation25,27]. Accordingly, it was suggested that a possible local cueould be the arena corner itself [25].
The present findings demonstrate that the objects (portableorners) in the square arena were not perceived in the same wayf perceiving the arena corners, but rather as distinct objects ashey were perceived in the round arena. We suggest that this isartly due to a perception of landmarks in a context-dependentanner: both arenas served as a featureless background againsthich landmarks were ’pasted’ on [3] and thus became highlyistinct. In other words, each arena was detected as a globaleometric unit that emphasized the presence of objects withint. The distinctness of a landmark against the background of thenvironment results in convergence of the rats’ routes upon theandmark, turning it into a beacon which is a landmark at thearget of exploration [37]. Moreover, the present results demon-trate that the global enclosure geometry affects the explorationf landmarks (objects, start point) within the enclosure: i.e., dif-
erent environment geometries (square, round) affect the wayhat landmarks are perceived and in turn result in a different dis-ribution of activity. Nevertheless, the suggestion that the objectsere distinct against the global enclosure geometry does nottHdu
square arena, time spent at object zones equaled that spent at the perimeter, but
ontradict the notion that the objects were also detected as localues and that the rats could judiciously combine global and localtrategies in their exploration [4].
.2. Detecting the geometry of an object array
It is still a matter of debate as to whether animals encodehe geometry of object arrays [29]. Previous studies demon-trated that rodents and human infants encode the shape ofbject arrays [2,15,28,29,32]. Studies in hamsters [28,32] andats [29] revealed that changing the shape of a square array byistancing one object away from it resulted in increased contactith the relocated object. A removal of one object and thereby
ransforming the square array into a triangular one increasedxploration to the remaining objects [32]. However, when ham-ters and rats were exposed to expansion of object array, nohange in the overall time spent near the objects was observed29,32]. It was suggested that the lack of response in the rats tohe expanded array was due to the already encoded shape of therray, a shape that remained unchanged but merely expanded32]. Nevertheless, the rats appear to have perceived the expan-ion, as evidenced by their hesitancy to enter the arena [29].ence, there is accumulating evidence that animals may encode
he geometry of arrays separately from the overall geometry.owever, opposing evidence also exists: for example, chicksid not seem to perceive an object array as a geometric config-ration and relied instead on a single object at a time [9], and

Brain
co
ti4rcptamhfhniif
tttcdttrtTospars
4
tadiotec
cttapfaf
dlsagbbt
A
dNb
R
[
[
[
[
[
[
O. Yaski, D. Eilam / Behavioural
hildren failed to use the geometric configuration of objects inrder to reorient themselves [16].
The present findings are consistent with earlier ones regardinghe detection of array geometry [28,29,32]. Positional changesn object proximity (arrays 2 and 3) and connectedness (array) preserved the geometry of the array and did not affect theats’ level of activity. Indeed, even when the objects (portableorners) were rotated, the rats preserved their activity and theirreference for the interior of the corners. When the geometry ofhe array was changed from a square (four objects, arrays 2–4) totriangle (three objects, array 5), rats in the triangular array spentore time at each object than they had done in the square array;
owever, the cumulative time spent at the triangular did not differrom that spent at the square arrays. Thus, the rats appeared toave a fix time allocation for the entire array, regardless of theumber of objects that comprised it. Altogether, these findingsmply that rats detected the geometry of the array and encodedt separately, and consequently raise questions concerning theunctional value of geometry detection.
When tested in a water maze, rats performed a navigationask based on the geometric arrangement of objects (isoscelesriangle), thus using the array as a whole unit and ignoring fea-ural information provided by object identity [2]. Under theseircumstances, rats perceived the arrangement of objects as airectional cue [4], which is presumably simpler than deducinghe position of a single object relative to the distance and posi-ion of other objects within the array [22]. In the present study,ats related to the specific features of the objects regardless ofheir position or distance from the walls and from other objects.his indicates that the objects arranged in the array were rec-gnized as independent objects and did not provide directionalource of spatial information [22]. Thus, the present study com-lements the findings that both rats [2,29] and infants [15] areble to detect the geometry of object array. However, unlike theats in the water maze [2], infants [15] and rats in the presenttudy also detect objects’ identity, as discussed next.
.3. Detecting the local object geometry
In addition to the global geometry of the enclosure and ofhe array, each of the objects was structured as a portable cornernd thereby had a characteristic geometry. The rats appeared toetect this local object geometry, exhibiting a preference for thenterior of the corner and maintaining this preference regardlessf object orientation. The animals also seemed to incorporatehis local object geometry within the global geometry of thenclosure by preferring to stay longer at the objects in the roundompared with the square arena.
We suggest that the local geometry of the object (a portableorner) provided a protected area that was more apparent againsthe geometry of the round enclosure, a geometry that is a fea-ureless background compared with the more structured squarerena. Indeed, in the square enclosure, arena corners appear to be
referred safe locations [7,13,36], while the objects constituteocal points of interest [37]. In accordance, rats in the squarerena stayed for extended durations at the arena corners butrequently visited the objects.[
[
Research 187 (2008) 334–342 341
In conclusion, the context of the global geometry providesifferent meanings: the same objects and array form a safeocation in the round arena, and a source of attraction in thequare arena. Thus, the physical structure of the environment,s manifested in the interface of the global enclosure and arrayeometry together with the local object geometry, is perceivedy the rats and has a major share in shaping their exploratoryehavior in terms of the places at which they stay or visit andheir trajectories of progression among these places.
cknowledgments
We are grateful to Hadas Benhar for her help in testing andata acquisition, to Reut Avni for her useful comments, and toaomi Paz for editing the article. This research was supportedy grant 471/04 of The Israel Science Foundation.
eferences
[1] Avni R, Zadicario E, Eilam D. Exploration in a dark open field: a shift fromdirectional to positional progression and a proposed model of acquiringspatial information. Behav Brain Res 2006;171:313–23.
[2] Benhamou S, Poucet B. Landmark use by navigating rats (Rattus norvegi-cus): contrasting geometric and featural information. J Comp Psychol1998;112:317–22.
[3] Cheng K. A purely geometric module in the rat’s spatial representation.Cognition 1986;23:149–78.
[4] Cheng K. Reflections on geometry and navigation. Connect Sci2005;17:5–21.
[5] Cheng K, Gallistel CR. Shape parameters explain data from spatial trans-formations: comment on Pearce et al (2004) and Tommasi & Polli (2004).J Exp Psychol Anim Behav Process 2005;31:254–9.
[6] Cheng K, Newcombe NS. Is there a geometric module for spatial orienta-tion? Squaring theory and evidence. Psychon Bull Rev 2005;12:1–23.
[7] Clark BJ, Hamilton DA, Whishaw IQ. Motor activity (exploration) andformation of home bases in mice (c57bl/6) influenced by visual and tactilecues: modification of movement distribution, distance, location, and speed.Physiol Behav 2006;87:805–16.
[8] Clark BJ, Hines DJ, Hamilton DA, Whishaw IQ. Movements of explorationintact in rats with hippocampal lesions. Behav Brain Res 2005;163:91–9.
[9] Della Chiesa A, Speranza M, Tommasi L, Vallortigara G. Spatial cognitionbased on geometry and landmarks in the domestic chick (Gallus gallus).Behav Brain Res 2006;175:119–27.
10] Drai D, Kafkafi N, Benjamini Y, Elmer G, Golani I. Rats and mice sharecommon ethologically relevant parameters of exploratory behavior. BehavBrain Res 2001;125:133–40.
11] Eilam D, Dank M, Maurer R. Voles scale locomotion to the size of theopen-field by adjusting the distance between stops: a possible link to pathintegration. Behav Brain Res 2003;141:73–81.
12] Eilam D, Golani I. Home base behavior of rats (Rattus norvegicus) explor-ing a novel environment. Behav Brain Res 1989;34:199–211.
13] Ennaceur A, Michalikova S, Chazot PL. Models of anxiety: responses ofrats to novelty in an open space and an enclosed space. Behav Brain Res2006;171:26–49.
14] Gallistel CR. The organization of learning. Cambridge, MA: Brad-ford/MIT Press; 1990.
15] Garrad-Cole F, Lew AR, Bremner JG, Whitaker CJ. Use of cue configura-tion geometry for spatial orientation in human infants (Homo sapiens). JComp Psychol 2001;115:317–20.
16] Gouteux S, Spelke ES. Children’s use of geometry and landmarks to reori-ent in an open space. Cognition 2001;81:119–48.
17] Gouteux S, Thinus-Blanc C, Vauclair J. Rhesus monkeys use geometricand nongeometric information during a reorientation task. J Exp PsycholGen 2001;130:505–19.

3 Brain
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
42 O. Yaski, D. Eilam / Behavioural
18] Graham P, Fauria K, Collett TS. The influence of beacon-aiming on theroutes of wood ants. J Exp Biol 2003;206:535–41.
19] Hermer L, Spelke E. Modularity and development: the case of spatialreorientation. Cognition 1996;61:195–232.
20] Hermer L, Spelke E. A geometric process for spatial reorientation in youngchildren. Nature 1994;370:57–9.
21] Hines DJ, Whishaw IQ. Home bases formed to visual cues but not to self-movement (dead reckoning) cues in exploring hippocampectomized rats.Eur J Neurosci 2005;22:2363–75.
22] Jacobs LF, Schenk F. Unpacking the cognitive map: the parallelmap theory of hippocampal function. Psychol Rev 2003;110:285–315.
23] Kelly DM, Spetch ML. Pigeons encode relative geometry. J Exp PsycholAnim Behav Process 2001;27:417–22.
24] Lever C, Wills T, Cacucci F, Burgess N, O’Keefe J. Long-term plasticity inhippocampal place-cell representation of environmental geometry. Nature2002;416:90–4.
25] McGregor A, Jones PM, Good MA, Pearce JM. Further evidence that ratsrely on local rather than global spatial information to locate a hidden goal:reply to Cheng and Gallistel (2005). J Exp Psychol Anim Behav Process2006;32:314–21.
26] Paulus MP, Geyer MA. Environment and unconditioned motor behavior:influences of drugs and environmental geometry on behavioral organization
in rats. Psychobiology 1997;25:327–37.27] Pearce JM, Good MA, Jones PM, McGregor A. Transfer of spatial behaviorbetween different environments: implications for theories of spatial learn-ing and for the role of the hippocampus in spatial learning. J Exp PsycholAnim Behav Process 2004;30:135–47.
[
[
Research 187 (2008) 334–342
28] Poucet B, Chapuis N, Durup M, Thinus-Blanc C. A study of exploratorybehavior as an index of spatial knowledge in hamsters. Anim Learn Behav1986;14:93–100.
29] Skov-Rackette SI, Shettleworth SJ. What do rats learn about the geometryof object arrays? Tests with exploratory behavior. J Exp Psychol AnimBehav Process 2005;31:142–54.
30] Sovrano VA, Bisazza A, Vallortigara G. Modularity as a fish (Xeno-toca eiseni) views it: conjoining geometric and nongeometric informationfor spatial reorientation. J Exp Psychol Anim Behav Process 2003;29:199–210.
31] Tchernichovski O, Benjamini Y, Golani I. The dynamics of long-termexploration in the rat – part I. A phase-plane analysis of the relationshipbetween location and velocity. Biol Cybern 1998;78:423–32.
32] Thinus-Blanc C, Bouzouba L, Chaix K, Chapuis N, Durup M, Poucet B. Astudy of spatial parameters encoded during exploration in hamsters. J ExpPsychol Anim Behav Process 1987;13:418–27.
33] Tommasi L, Thinus-Blanc C. Generalization in place learning and geometryknowledge in rats. Learn Mem 2004;11:153–61.
34] Tommasi L, Vallortigara G, Zanforlin M. Young chickens learn to localizethe centre of a spatial environment. J Comp Physiol A 1997;180:567–72.
35] Vallortigara G, Zanforlin M, Pasti G. Geometric modules in animals spatialrepresentations – a test with chicks (Gallus gallus domesticus). J CompPsychol 1990;104:248–54.
36] Whishaw IQ, Gharbawie OM, Clark BJ, Lehmann H. The exploratorybehavior of rats in an open environment optimizes security. Behav BrainRes 2006;171:230–9.
37] Yaski O, Eilam D. The impact of landmark properties in shaping explorationand navigation. Anim Cogn 2007;10:415–28.




















