
Heterochronic growth of ostracods (Crustacea) from microbial deposits in the aftermath of the...
36
This article was downloaded by: [MNHN Muséum National D'Histoire Naturelle] On: 16 May 2014, At: 00:28 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Systematic Palaeontology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tjsp20 Heterochronic growth of ostracods (Crustacea) from microbial deposits in the aftermath of the end-Permian extinction Marie-Béatrice Forel a a State Key Laboratory of Geological Process and Mineral Resources, China University of Geosciences, No. 388, Lumo Road, Wuhan 430074, China Published online: 14 May 2014. To cite this article: Marie-Béatrice Forel (2014): Heterochronic growth of ostracods (Crustacea) from microbial deposits in the aftermath of the end-Permian extinction, Journal of Systematic Palaeontology, DOI: 10.1080/14772019.2014.902400 To link to this article: http://dx.doi.org/10.1080/14772019.2014.902400 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Heterochronic growth of ostracods (Crustacea) from microbial deposits in the aftermath of the...

This article was downloaded by: [MNHN Muséum National D'Histoire Naturelle]On: 16 May 2014, At: 00:28Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Systematic PalaeontologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tjsp20
Heterochronic growth of ostracods (Crustacea) frommicrobial deposits in the aftermath of the end-PermianextinctionMarie-Béatrice Forelaa State Key Laboratory of Geological Process and Mineral Resources, China University ofGeosciences, No. 388, Lumo Road, Wuhan 430074, ChinaPublished online: 14 May 2014.
To cite this article: Marie-Béatrice Forel (2014): Heterochronic growth of ostracods (Crustacea) from microbial deposits inthe aftermath of the end-Permian extinction, Journal of Systematic Palaeontology, DOI: 10.1080/14772019.2014.902400
To link to this article: http://dx.doi.org/10.1080/14772019.2014.902400
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Heterochronic growth of ostracods (Crustacea) from microbial deposits in theaftermath of the end-Permian extinction
Marie-B�eatrice Forel*
State Key Laboratory of Geological Process and Mineral Resources, China University of Geosciences, No. 388, Lumo Road,Wuhan 430074, China
(Received 14 May 2013; accepted 14 November 2013)
The Kokarkuyu Formation (Early Triassic) from the C€ur€uk Da�g section, Western Taurus, Antalya Nappes, Turkey wascarefully sampled for a taxonomic study of the ostracods. A total of 57 species belonging to 15 genera are recognized.Three species are newly described: Monoceratina hussonae sp. nov., Reviya sylvieae sp. nov. and Eukloedenellaadcapitisdolorella sp. nov. The present data are the first Neo-Tethyan illustration of ostracod survival in the aftermath ofthe end-Permian extinction in a refuge of microbial origin. Ostracods are abundant within the microbialites at the base ofthe formation and allow the reconstruction of ontogenetic series for nine species. Shape variations through ontogeny aredescribed for seven species: Bairdia? kemerensis, Praezabythocypris? ottomanensis, Liuzhinia antalyaensis, Paracyprisgaetanii, Reviya curukensis, R. sylvieae and Eukloedenella adcapitisdolorella. Paedomorphosis through deceleration andperamorphosis through acceleration are identified as secondary survival strategies following the end-Permian extinction.
http://zoobank.org/urn:lsid:zoobank.org:pub:6849AB0D-B085-47BE-950A-BC98CC50C4E3
Keywords: Crustacea; ostracods; Early Triassic; microbialites; heterochrony; C€ur€uk Da�g
Introduction
The Permian–Triassic Boundary (PTB; �252 Ma) is
marked by the most dramatic extinction of the Phanero-
zoic, when 80–96% of marine species went extinct (e.g.
Sepkoski 1984; Erwin 1993; Benton & Twitchett 2003).
According to some authors, global ocean anoxia was
widespread during the latest Permian (Changhsingian),
increasing close to the main extinction and continuing
during the Early Triassic (e.g. Bond & Wignall 2010). An
important trigger of the extinction was the eruption of the
Siberian Traps, causing global warming, ocean acidifica-
tion and possible destruction of atmospheric ozone (e.g.
Svensen et al. 2009). Subsequent enhanced weathering
and nutrient run-off might have increased the pre-existing
ocean anoxia (e.g. Payne & Clapham 2012). Marine eco-
systems saw the replacement of the Late Permian benthic
shelly communities dominating shallow marine environ-
ments by microbial communities. These microbialites
were abundant in low-latitude, shallow-marine carbonate
shelves in the central Tethys where they occupied envi-
ronments formerly populated by Late Permian reefs but
extending also into deeper waters (see Kershaw et al.
2012 for a synthesis).
Current evidence regarding oxygen levels associated
with microbial expansions is conflicting, with evidence of
both low-oxygen markers (Bond & Wignall 2010; Liao
et al. 2010; Chen et al. 2011) and abundant benthic shelly
faunas dominated by ostracods, and occasionally micro-
gastropods, micro-brachiopods, foraminifers, bivalves and
conodonts (e.g. Baud et al. 1997; Crasquin-Soleau & Ker-
shaw 2005; Groves et al. 2005; Richoz 2006; Br€uwhileret al. 2008; Ezaki et al. 2008; Forel et al. 2009; Song et al.
2009; Kaim et al. 2010; Hautmann et al. 2011; Yang et al.
2011; Forel 2012; Frisk et al. 2012; Forel et al. 2013a).
These unusual surviving faunas are explained by a two-
step oxygenation mechanism of surrounding waters by cya-
nobacterial activity (Forel et al. 2013b).
Through the end-Permian extinction (EPE), ostracods
(Crustacea) show specific extinction rates ranging from
74 to 100% (Crasquin & Forel 2013). Their recovery is
assumed to be complete in the Late Triassic when biodi-
versity returned to levels similar to those of Middle Perm-
ian faunas. Ostracod faunas described from Early Triassic
rocks are rare, so that survival mechanisms and refuge
zones at the origin of later recovery remain unclear. The
assemblages considered here from the C€ur€uk Da�g section
(Turkey) illustrate secondary survival mechanisms, based
on the modifications of the development (heterochronies),
in a refuge of microbial origin already known from sev-
eral localities from the Panthalassa and Palaeo-Tethys
Oceans (Forel 2012, 2013; Forel et al. 2013a, b).
*Corresponding author. Email: [email protected]
� The Trustees of the Natural History Museum, London 2014. All Rights Reserved.
Journal of Systematic Palaeontology, 2014
http://dx.doi.org/10.1080/14772019.2014.902400
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

Previous work on the PTB interval from C€ur€uk Da�gdocumented 38 ostracod species from 22 genera (Cras-
quin-Soleau et al. 2002, 2004a, b). To assess ostracod
populations during and after microbial build-up in the
aftermath of the EPE, the Kokarkuyu Formation of Early
Triassic age was carefully resampled. Fifty-seven species,
including three new species, belonging to 15 genera were
found and are reported in the present work. The data from
Crasquin-Soleau et al. (2002, 2004a, b) are included in
our database to provide the most complete record cur-
rently available for the ostracod biodiversity variations
and palaeo-environmental changes throughout the Kokar-
kuyu formation and more generally in the Neo-Tethys
area (Fig. 1).
Material and methods
The material described in the present work comes from
the C€ur€uk Da�g section (36� 410 32400 N, 30� 270 40.100 E),located in the Western Taurus, Antalya Nappes, Turkey
(Fig. 1). A succinct description of the lithological
succession is provided here: the reader is referred to Baud
et al. (1997, 2005), Crasquin et al. (2009) and Kershaw
et al. (2010) and references therein for further details. The
Upper Permian Pamucak Formation consists of wacke-
stones and grainstones/packstones. It is overlain by the
Kokarkuyu Formation, 40 metres thick in the C€ur€uk Da�gsection. The first 15 metres of this formation are com-
posed of cyclic microbial limestones, mostly made of stro-
matolites, thrombolites and hybrid microbialites (Kershaw
et al. 2010). The discovery of the conodonts Isarcicella
staeschei at the base of the formation and Hindeodus
parvus 55 cm higher up (Richoz 2006) indicates that
the base of the microbialites is already in the Triassic. It
is followed by 3 metres of calcilutites, overlain by
wackestones. The Upper Permian ostracod assemblages
have been described by Crasquin-Soleau et al. (2004a, b).
Samples spanning the entire exposed Kokarkuyu
Formation were collected to describe ostracod assemb-
lages from facies below, within and above the microbia-
lites (Fig. 2).
External features of carapaces are necessary to deter-
mine ostracods. Extraction of ostracods from the matrix
Figure 1. Middle Permian palaeogeographical map (modified from Crasquin-Soleau et al. 2001) and modern map showing the locationof the C€ur€uk Da�g section, Taurus, Turkey.
2 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
using acid dissolution is precluded because their carbon-
ate carapaces are enclosed in calcareous rocks. Disaggre-
gation of dehydrated hard limestone and release of
ostracod shells were performed by the hot acetolysis tech-
nique (Lethiers & Crasquin-Soleau 1988; Crasquin-Sol-
eau et al. 2005). A total of 140 samples were collected
and processed, of which 45 yielded ostracods (Fig. 3).
The productive samples are located in the lowermost Tri-
assic microbialites; only two samples yielded specimens
above the microbial crust. Because of poor preservation,
many species are left in open nomenclature. However, it
is important to figure all the material of this particular
interval (Figs 4, 6–8, 11, 12, 14, 20).
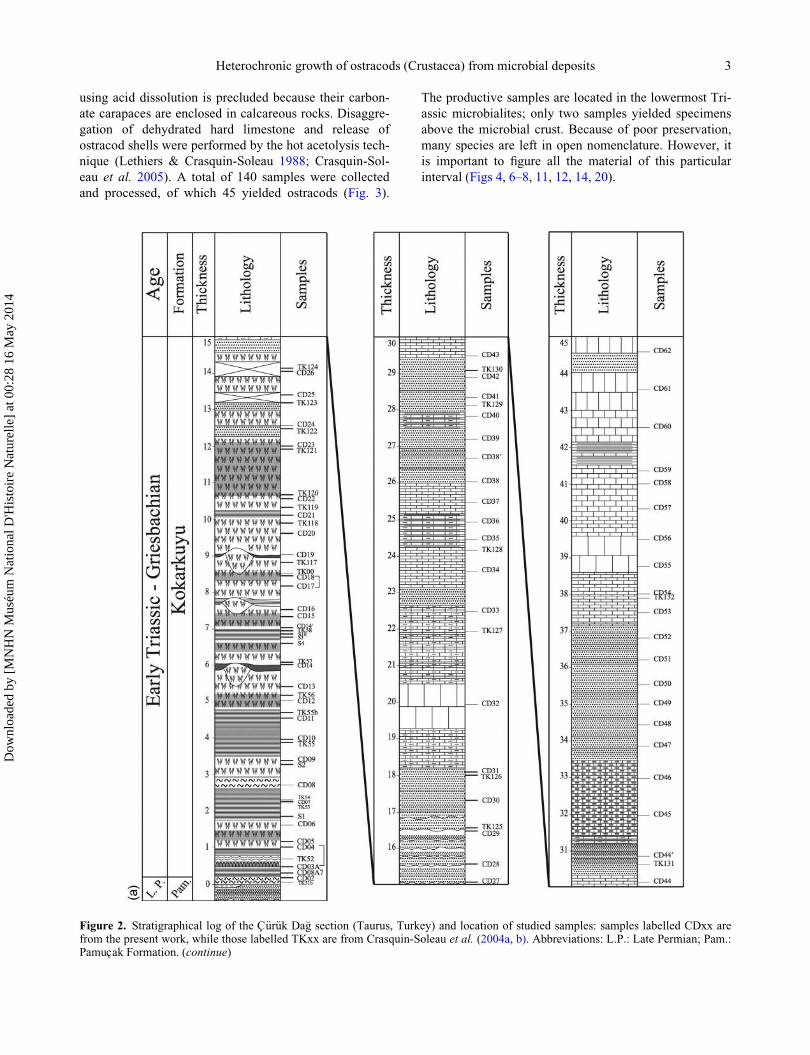
Figure 2. Stratigraphical log of the C€ur€uk Da�g section (Taurus, Turkey) and location of studied samples: samples labelled CDxx arefrom the present work, while those labelled TKxx are from Crasquin-Soleau et al. (2004a, b). Abbreviations: L.P.: Late Permian; Pam.:Pamucak Formation. (continue)
Heterochronic growth of ostracods (Crustacea) from microbial deposits 3
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

Ostracods have determinate growth: they display a step-
wise development resulting from moulting (ecdysis) and
stop moulting when they reach adult stage. Specimens are
here assigned to their corresponding instar group based on
size. The preservation makes it impossible to study hinge
structure and muscle scars. Lengths of specimens were
plotted against heights in scatter plots to distinguish size
clusters which discriminate successive juveniles and
adults. In the fossil record, the recognition of different
growth stages is difficult or impossible because of time-
averaging of specimens from different environments or
seasons (e.g. specimens of the same species tend to be
smaller in the spring than in the autumn; Morales-Ram�ırez& Jakob 2008). To overcome this difficulty, I computed
Kernel density maps (Gaussian Kernel distribution, col-
umns ¼ 100, rows ¼ 100, radius ¼ 15) using PAST soft-
ware (Hammer et al. 2001; Hammer & Harper 2005). For
each species yielding at least 15 specimens (from this
Figure 2. (Continued)
4 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
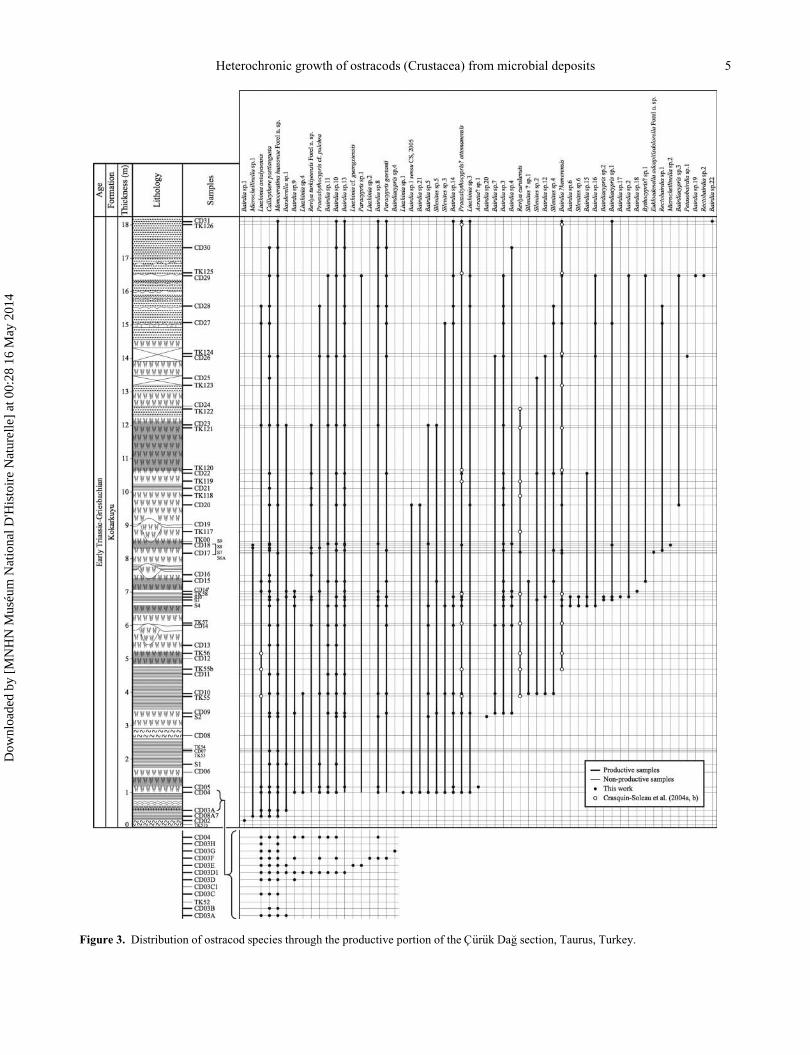
Figure 3. Distribution of ostracod species through the productive portion of the C€ur€uk Da�g section, Taurus, Turkey.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 5
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
work and the literature), it allowed the discrimination of
density patterns of individuals corresponding to different
ontogenetical stages. Successive instars of nine species
are recognized and described here. Of the 57 species iden-
tified, the Systematic palaeontology section does not
include those species left in open nomenclature.
All specimens are deposited in the collections of the
Pierre et Marie Curie University, Paris, France, under
numbers P6Mxxx.
Systematic palaeontology
For the three new species a full description is given. For
other species only the synonymy and occurrences are spec-
ified. The classification employed is that of Moore (1961),
modified after Lethiers (1981) and Horne et al. (2002).
Morphological abbreviations: RV, right valve; LV, left
valve; AB, anterior border; PB, posterior border; DB, dor-
sal border; VB, ventral border; ADB, anterodorsal border;
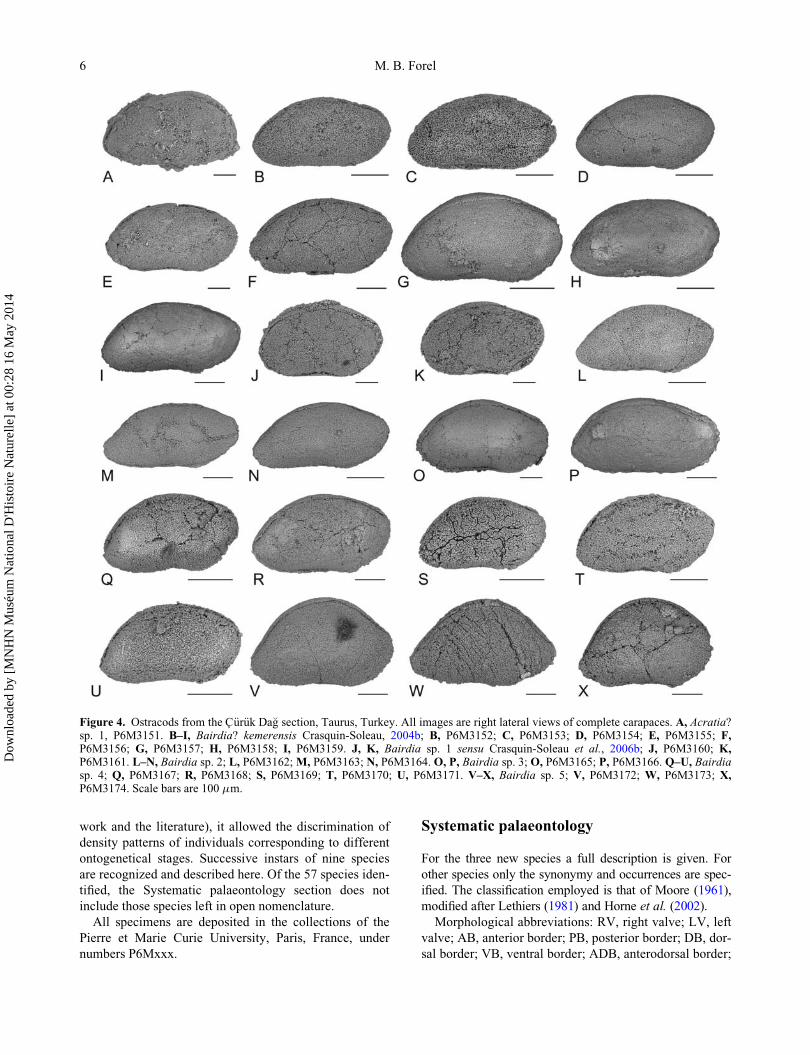
Figure 4. Ostracods from the C€ur€uk Da�g section, Taurus, Turkey. All images are right lateral views of complete carapaces. A, Acratia?sp. 1, P6M3151. B–I, Bairdia? kemerensis Crasquin-Soleau, 2004b; B, P6M3152; C, P6M3153; D, P6M3154; E, P6M3155; F,P6M3156; G, P6M3157; H, P6M3158; I, P6M3159. J, K, Bairdia sp. 1 sensu Crasquin-Soleau et al., 2006b; J, P6M3160; K,P6M3161. L–N, Bairdia sp. 2; L, P6M3162;M, P6M3163; N, P6M3164. O, P, Bairdia sp. 3; O, P6M3165; P, P6M3166. Q–U, Bairdiasp. 4; Q, P6M3167; R, P6M3168; S, P6M3169; T, P6M3170; U, P6M3171. V–X, Bairdia sp. 5; V, P6M3172; W, P6M3173; X,P6M3174. Scale bars are 100 mm.
6 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
AVB, anteroventral border; PDB, posterodorsal border;
PVB, posteroventral border; ACA, anterior cardinal angle
(for Palaeocopida only); PCA, posterior cardinal angle
(for Palaeocopida only); ADA, anterodorsal angulation;
PDA, posterodorsal angulation; AVA, anteroventral angu-
lation (angle between AVB and VB); PVA, posteroventral
angulation (PVA as angle between VB and PVB); S1,
anterior sulcus; S2, median sulcus; H, height; Hmax, max-
imal height; L, length; Lmax, maximal length.
Class Ostracoda Latreille, 1806
Subclass PodocopaM€uller, 1894Order PodocopidaM€uller, 1894Suborder Podocopina Sars, 1866
Superfamily Bairdioidea Sars, 1887
Family Bairdiidae Sars, 1887
Genus BairdiaMcCoy, 1844
Bairdia? kemerensis Crasquin-Soleau, 2004
(Fig. 4B–I)
2004b Bairdia? kemerensis Crasquin-Soleau in Crasquin-
Soleau et al.: 285, pl. 2, figs 1–5.
Occurrence. Samples S4, S3, S7, 08CD17, 20, 26, 27,
29, 31, TK55b-58, TK120, TK121, TK123–126, C€ur€ukDa�g section (Fig. 3), Western Taurus, Turkey, Kokarkuyu
Formation, Griesbachian, Induan, Early Triassic (this
work; Crasquin-Soleau et al. 2004a, b)
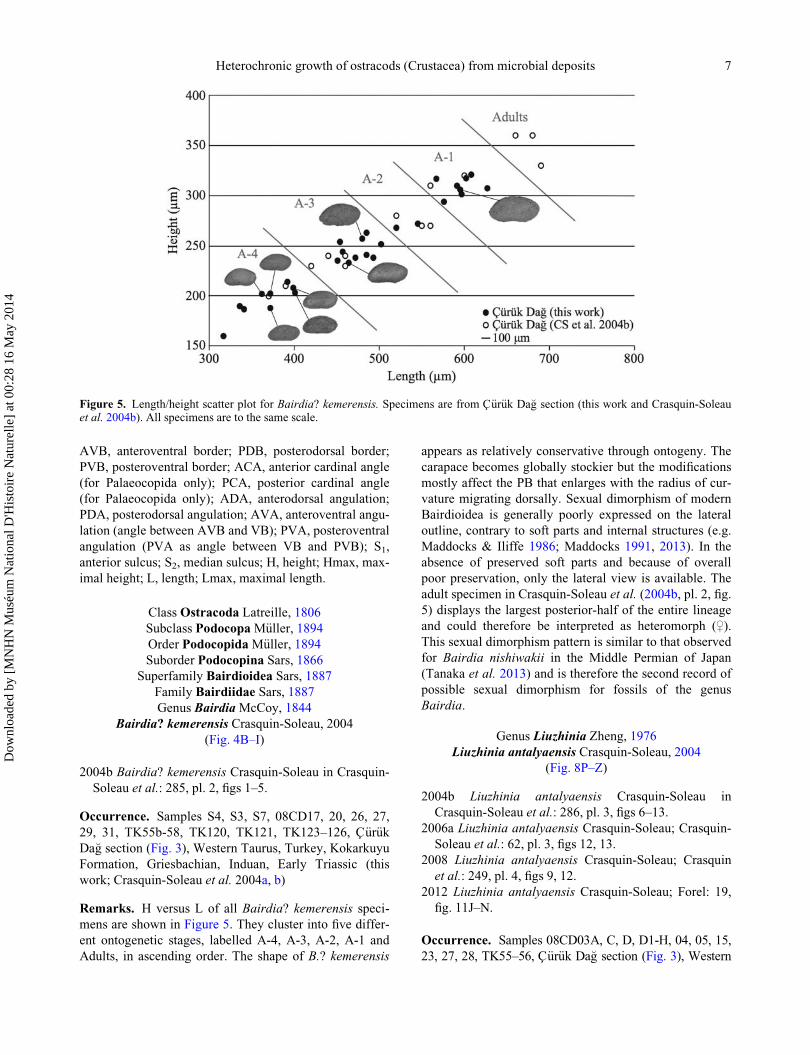
Remarks. H versus L of all Bairdia? kemerensis speci-
mens are shown in Figure 5. They cluster into five differ-
ent ontogenetic stages, labelled A-4, A-3, A-2, A-1 and
Adults, in ascending order. The shape of B.? kemerensis
appears as relatively conservative through ontogeny. The
carapace becomes globally stockier but the modifications
mostly affect the PB that enlarges with the radius of cur-
vature migrating dorsally. Sexual dimorphism of modern
Bairdioidea is generally poorly expressed on the lateral
outline, contrary to soft parts and internal structures (e.g.
Maddocks & Iliffe 1986; Maddocks 1991, 2013). In the
absence of preserved soft parts and because of overall
poor preservation, only the lateral view is available. The
adult specimen in Crasquin-Soleau et al. (2004b, pl. 2, fig.
5) displays the largest posterior-half of the entire lineage
and could therefore be interpreted as heteromorph (,).This sexual dimorphism pattern is similar to that observed
for Bairdia nishiwakii in the Middle Permian of Japan
(Tanaka et al. 2013) and is therefore the second record of
possible sexual dimorphism for fossils of the genus
Bairdia.
Genus Liuzhinia Zheng, 1976
Liuzhinia antalyaensis Crasquin-Soleau, 2004
(Fig. 8P–Z)
2004b Liuzhinia antalyaensis Crasquin-Soleau in
Crasquin-Soleau et al.: 286, pl. 3, figs 6–13.
2006a Liuzhinia antalyaensis Crasquin-Soleau; Crasquin-
Soleau et al.: 62, pl. 3, figs 12, 13.
2008 Liuzhinia antalyaensis Crasquin-Soleau; Crasquin
et al.: 249, pl. 4, figs 9, 12.
2012 Liuzhinia antalyaensis Crasquin-Soleau; Forel: 19,
fig. 11J–N.
Occurrence. Samples 08CD03A, C, D, D1-H, 04, 05, 15,
23, 27, 28, TK55–56, C€ur€uk Da�g section (Fig. 3), Western
Figure 5. Length/height scatter plot for Bairdia? kemerensis. Specimens are from C€ur€uk Da�g section (this work and Crasquin-Soleauet al. 2004b). All specimens are to the same scale.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 7
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
Figure 6. Ostracods from the C€ur€uk Da�g section, Taurus, Turkey. All images are right lateral views of complete carapaces. A–C, Bair-dia sp. 6; A, P6M3175; B, P6M3176; C, P6M3177. D, Bairdia sp. 7, P6M3178. E, F, Bairdia sp. 8; E, P6M3179; F, P6M3180. G–S,Bairdia sp. 9; G, P6M3181; H, P6M3182; I, P6M3183; J, P6M3184; K, P6M3185; L, P6M3186; M, P6M3187; N, P6M3188; O,P6M3189; P, P6M3190; Q, P6M3191; R, P6M3192; S, P6M3193. T, U, Bairdia sp. 10; T, P6M3194; U, P6M3195. V–A0, Bairdia sp.11; V, P6M3196;W, P6M3197; X, P6M3198; Y, P6M3199; Z, P6M3200; A0, P6M3201. Scale bars are 100 mm.
8 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
Taurus, Turkey, Kokarkuyu Formation, Griesbachian,
Induan, Early Triassic (this work; Crasquin-Soleau et al.,
2004b). Jinya/Waili section, Fengshan area, Guangxi Prov-
ince, South China (Crasquin-Soleau et al. 2006a). Bulla
section, Dolomites, Southern Alps, Northern Italy (Cras-
quin et al. 2008). Dajiang section, Daye Formation,
Guizhou Province, South China (Forel 2012). All occur-
rences from Griesbachian, Induan, Early Triassic.
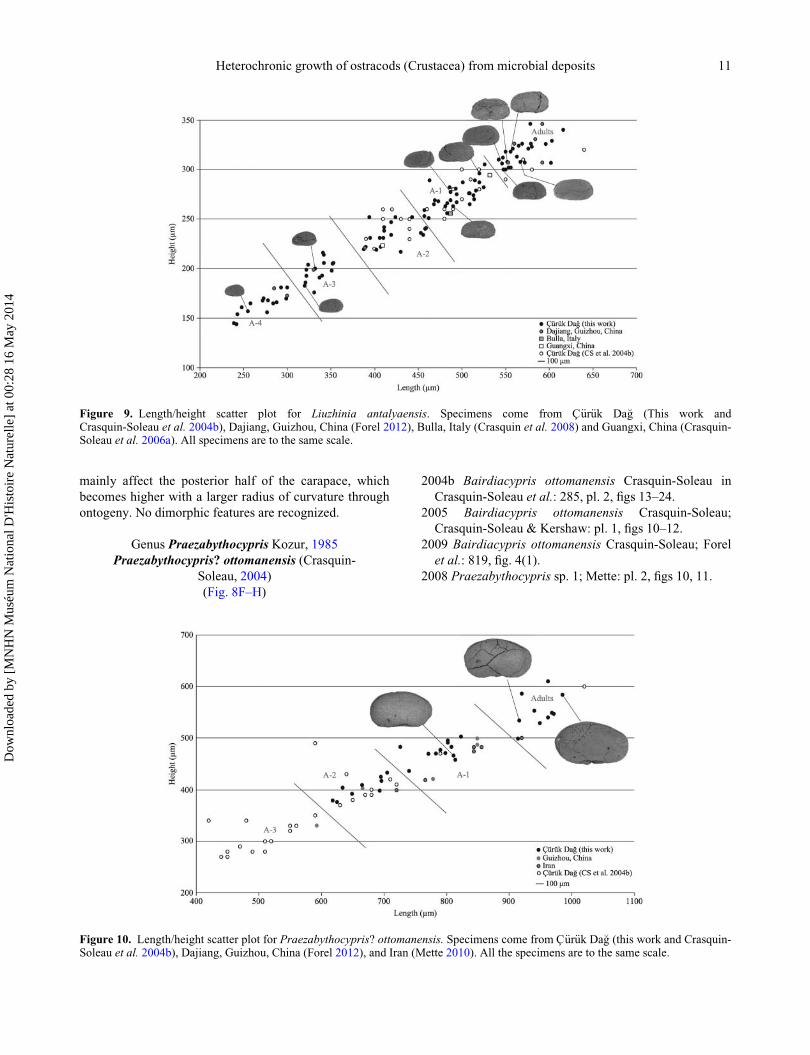
Remarks. Fig. 9 plots H versus L for all specimens of
Liuzhinia antalyaensis. Five successive ontogenetic stages
are distinguished, from A-4 to Adults. Modifications
Figure 7. Ostracods from the C€ur€uk Da�g section, Taurus, Turkey. All images are right lateral views of complete carapaces. A–D, Bair-dia sp. 12; A, P6M3202; B, P6M3203; C, P6M3204; D, P6M3205. E–G, Bairdia sp. 13; E, P6M3206; F, P6M3207; G, P6M3208. H–L, Bairdia sp. 14; H, P6M3209; I, P6M3210; J, P6M3211; K, P6M3212; L, P6M3213. M–Q, Bairdia sp. 15; M, P6M3214; N,P6M3215; O, P6M3216; P, P6M3217; Q, P6M3218. R, S, Bairdia sp. 16; R, P6M3219; S, P6M3220. T–V, Bairdia sp. 17; T,P6M3221; U, P6M3222; V, P6M3223.W, Bairdia sp. 18, P6M3224. X, Bairdia sp. 19, P6M3225. Scale bars are 100 mm.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 9
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
Figure 8. Ostracods from the C€ur€uk Da�g section, Taurus, Turkey. All images are right lateral views of complete carapaces. A, Bairdiasp. 20, P6M3226. B, Bairdia sp. 21, P6M3227. C, D, Bairdia sp. 22; C, P6M3228; D, P6M3229. E, Bairdia sp. 23, P6M3230. F–H,Praezabythocypris? ottomanensis (Crasquin-Soleau, 2004b); F, P6M3231; G, P6M3232; H, P6M3233. I, J, Bairdiacypris sp. 1; I,P6M3234; J, P6M3235. K, Bairdiacypris sp. 2, P6M3236. L, M, Bairdiacypris sp. 3 sensu Forel, 2012; L, P6M3237; M, P6M3238. N,Bairdiacypris sp. 4, P6M3239. O, Bythocypris? sp. 1, P6M3240. P–Z, Liuzhinia antalyaensis Crasquin-Soleau, 2004; P, P6M3241; Q,P6M3242; R, P6M3243; S, P6M3244; T, P6M3245; U, P6M3246; V, P6M3247; W, P6M3248; X, P6M3249; Y, P6M3250; Z,P6M3251. Scale bars are 100 mm.
10 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
mainly affect the posterior half of the carapace, which
becomes higher with a larger radius of curvature through
ontogeny. No dimorphic features are recognized.
Genus Praezabythocypris Kozur, 1985
Praezabythocypris? ottomanensis (Crasquin-
Soleau, 2004)
(Fig. 8F–H)
2004b Bairdiacypris ottomanensis Crasquin-Soleau in
Crasquin-Soleau et al.: 285, pl. 2, figs 13–24.
2005 Bairdiacypris ottomanensis Crasquin-Soleau;
Crasquin-Soleau & Kershaw: pl. 1, figs 10–12.
2009 Bairdiacypris ottomanensis Crasquin-Soleau; Forel
et al.: 819, fig. 4(1).
2008 Praezabythocypris sp. 1; Mette: pl. 2, figs 10, 11.
Figure 10. Length/height scatter plot for Praezabythocypris? ottomanensis. Specimens come from C€ur€uk Da�g (this work and Crasquin-Soleau et al. 2004b), Dajiang, Guizhou, China (Forel 2012), and Iran (Mette 2010). All the specimens are to the same scale.
Figure 9. Length/height scatter plot for Liuzhinia antalyaensis. Specimens come from C€ur€uk Da�g (This work andCrasquin-Soleau et al. 2004b), Dajiang, Guizhou, China (Forel 2012), Bulla, Italy (Crasquin et al. 2008) and Guangxi, China (Crasquin-Soleau et al. 2006a). All specimens are to the same scale.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 11
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
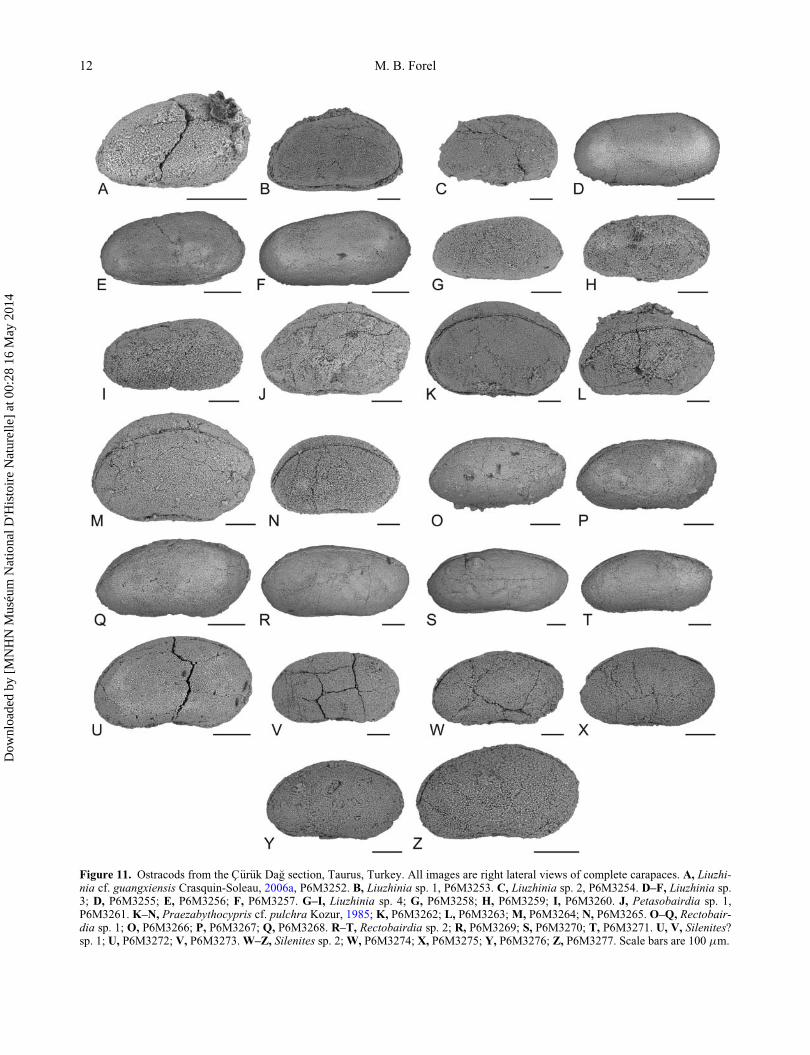
Figure 11. Ostracods from the C€ur€uk Da�g section, Taurus, Turkey. All images are right lateral views of complete carapaces. A, Liuzhi-nia cf. guangxiensis Crasquin-Soleau, 2006a, P6M3252. B, Liuzhinia sp. 1, P6M3253. C, Liuzhinia sp. 2, P6M3254. D–F, Liuzhinia sp.3; D, P6M3255; E, P6M3256; F, P6M3257. G–I, Liuzhinia sp. 4; G, P6M3258; H, P6M3259; I, P6M3260. J, Petasobairdia sp. 1,P6M3261. K–N, Praezabythocypris cf. pulchra Kozur, 1985; K, P6M3262; L, P6M3263; M, P6M3264; N, P6M3265. O–Q, Rectobair-dia sp. 1; O, P6M3266; P, P6M3267; Q, P6M3268. R–T, Rectobairdia sp. 2; R, P6M3269; S, P6M3270; T, P6M3271. U, V, Silenites?sp. 1; U, P6M3272; V, P6M3273.W–Z, Silenites sp. 2;W, P6M3274; X, P6M3275; Y, P6M3276; Z, P6M3277. Scale bars are 100 mm.
12 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
2010 Praezabythocypris? ottomanensis Crasquin-Soleau;
Mette: 28, pl. 4, figs 1–4.
non 2010 Bairdiacypris ottomanensis Crasquin-Soleau;
Liu et al.: fig. 3(1).
2012 Bairdiacypris ottomanensis Crasquin-Soleau; Forel:
13, fig. 10E–H.
Occurrence. Samples 08CD04, 09, 10, S4, S3, S10,
08CD14’, S9, 08CD22, 31, TK120, 125, 126, C€ur€uk Da�gsection (Fig. 3), Western Taurus, Turkey, Kokarkuyu For-
mation, Griesbachian, Induan, Early Triassic (this work;
Crasquin-Soleau et al. 2004a, b). Laolongdong section,
Feixianguan Formation, Griesbachian, Induan, Early
Triassic, Sichuan Province, South China (Crasquin-Soleau
& Kershaw 2005). Zal section, Ali Bashi and Zal forma-
tions, Late Permian–Early Triassic, north-west Iran
(Mette 2008, 2010). Dajiang section, Daye Formation,
Griesbachian, Induan, Early Triassic, Guizhou Province,
South China (Forel et al. 2009; Forel 2012).
Remarks. Figure 10 plots H versus L for all known Prae-
zabythocypris? ottomanensis specimens. They cluster into
four successive ontogenetic groups, in ascending order:
A-3, A-2 (Crasquin-Soleau et al. 2004b, pl. 2, fig. 13),
A-1 (Fig. 8H; Forel 2012, fig. 10E–G), and Adults
(Fig. 8F, G). Through ontogeny, the carapace becomes
more preplete and PB more rounded with a maximum
of curvature located higher. The specimen identified as
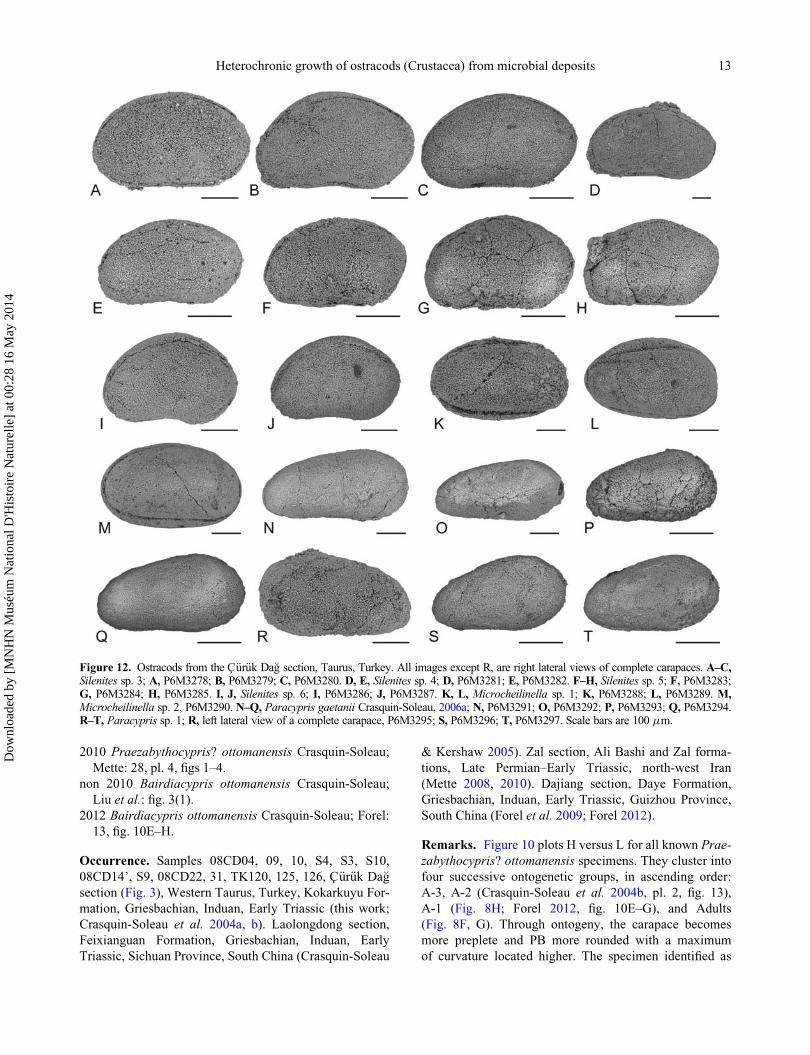
Figure 12. Ostracods from the C€ur€uk Da�g section, Taurus, Turkey. All images except R, are right lateral views of complete carapaces. A–C,Silenites sp. 3; A, P6M3278; B, P6M3279; C, P6M3280. D, E, Silenites sp. 4; D, P6M3281; E, P6M3282. F–H, Silenites sp. 5; F, P6M3283;G, P6M3284; H, P6M3285. I, J, Silenites sp. 6; I, P6M3286; J, P6M3287. K, L, Microcheilinella sp. 1; K, P6M3288; L, P6M3289. M,Microcheilinella sp. 2, P6M3290. N–Q, Paracypris gaetanii Crasquin-Soleau, 2006a; N, P6M3291; O, P6M3292; P, P6M3293; Q, P6M3294.R–T, Paracypris sp. 1; R, left lateral view of a complete carapace, P6M3295; S, P6M3296; T, P6M3297. Scale bars are 100 mm.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 13
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
Praezabythocypris? ottomanensis by Liu et al. (2010) is
excluded because it clearly displays a bairdian shape with
short DB and no angulation at dorsum.
Praezabythocypris cf. pulchra Kozur, 1985
(Figs 11K–N)
cf. 1985 Praezabythocypris pulchra Kozur: 84, taf. 17,
fig. 13.
Occurrence. Samples 08CD04, 05, S1, 08CD10, 14’, S8,
08CD26, 28, C€ur€uk Da�g section (Fig. 3), Western Taurus,
Turkey, Kokarkuyu Formation, Griesbachian, Induan,
Early Triassic (this work).
Remarks. The present species differs from Praezabytho-
cypris pulchraKozur, 1985 from the Middle and Late Perm-
ian of Hungary (Kozur 1985) by its steeper PDB, a more
inclined DB at RV, a longer and less rounded AVB, and the
posterior maximum of curvature located more ventrally.
Superfamily Cypridoidea Baird, 1845
Family Paracyprididae Sars, 1923
Genus Paracypris Sars, 1910
Paracypris gaetanii Crasquin-Soleau, 2006
(Fig. 12N–Q)
1992 Paracypris sp.; Hao: 42, pl. 1, fig. 24.
2005 Paracypris sp. sensu Hao 1999 [sic]; Crasquin-
Soleau & Kershaw: pl. I, figs 7–9.
2006a Paracypris gaetanii Crasquin-Soleau in Crasquin-
Soleau et al.: 64, pl. 4, figs 1–4.
2008 Paracypris gaetanii Crasquin-Soleau; Crasquin
et al.: 249, pl. 4, fig. 11.
2009 Paracypris gaetanii Crasquin-Soleau; Forel et al.:
819, fig. 4(5).
2011 Paracypris gaetanii Crasquin-Soleau; Forel &
Crasquin: figs 3F’–4A.
2012 Paracypris gaetanii Crasquin-Soleau; Forel: 22,
fig. 13T–X.
Occurrence. Samples 08CD03F, 09, 10, S4, S10,
08CD15, S7, 08CD22, 26–31, C€ur€uk Da�g section (Fig. 3),
Western Taurus, Turkey (this work; Crasquin-Soleau
et al. 2004a, b). Kokarkuyu Formation, Griesbachian,
Induan, Early Triassic. Feihsienkuan Formation, Griesba-
chian, Induan, Early Triassic, Zhenfeng, Guizhou Prov-
ince, South China (Hao 1992). Laolongdong section,
Feixianguan Formation, Griesbachian, Induan, Early
Triassic, Sichuan Province, South China, Induan, Early
Triassic (Crasquin-Soleau & Kershaw 2005). Jinya/Waili
section, Griesbachian, Induan, Early Triassic, Fengshan
area, Guangxi Province, China (Crasquin-Soleau et al.
2006a). Bulla section, Griesbachian, Induan, Early Trias-
sic, Dolomites, Southern Alps, Northern Italy (Crasquin
et al. 2008). Dajiang section, Wujiaping and Daye Forma-
tions, Guizhou Province, China, Late Permian–Early Tri-
assic (Forel et al. 2009; Forel 2012). Meishan section,
Figure 13. Length/height scatter plot for Paracypris gaetanii. Specimens come from C€ur€uk Da�g (this work), Dajiang, Guizhou, China(Forel 2012), Meishan, Zhejiang, China (Forel & Crasquin 2011), Bulla, Italy (Crasquin et al. 2008), Guangxi, China (Crasquin-Soleauet al. 2006a), Laolongdong, Sichuan, China (Crasquin-Soleau & Kershaw 2005), and Zhenfeng, Guizhou, China (Hao 1992). All speci-mens are at the same scale.
14 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

Griesbachian, Induan, Early Triassic, Zhejiang Province,
China (Forel & Crasquin 2011).
Remarks. Figure 13 plots H versus L for all Paracypris
gaetanii specimens. They scatter into five ontogenetic
stages, labelled A-4 to Adults. Changes through growth
stages are mainly located at the posterior-half of the cara-
pace, becoming higher and more rounded. Interestingly,
some of the larger specimens display distinct morpholo-
gies at PB: (i) tapered with a narrow radius of curvature,
maximum located relatively ventrally (e.g. Fig. 12N;
Crasquin-Soleau et al. 2006a, pl. 4, fig. 2; Hao 1992, pl. 1,
fig. 24); (ii) rounded with a larger radius of curvature,
maximum located higher (e.g. Forel 2012, fig. 13T; Cras-
quin-Soleau & Kershaw 2005, pl. 1, figs 7–9). These two
groups could respectively correspond to tecnomorphs (<)and heteromorphs (,).
Superfamily Cytheroidea Baird, 1850
Family Bythocytheridae Sars, 1928
GenusMonoceratina Roth, 1928
Monoceratina hussonae sp. nov.
(Fig. 14A–J)
Diagnosis. Subtriangular species of Monoceratina with
well-expressed S2 and faint S1; adventral alae faint; AB
and PB rimmed; PB rounded; lateral surface reticulated.
Derivation of name. Dedicated to Dr Doroth�ee Husson,
Northwestern University, Chicago, USA.
Material. Holotype: P6M3300, one complete carapace
(Fig. 14C), sample S3. Paratype: P6M3305, one complete
carapace (Fig. 14H), sample 08CD10. All from the C€ur€ukDa�g section, Western Taurus, Turkey, Kokarkuyu Forma-
tion, Griesbachian, Induan, Early Triassic. Additional
material: 152 complete carapaces and several fragments.
Occurrence. Samples CD08A7, 08CD03A-C,
08CD03D1-H, 04, 05, S1, 08CD07, S2, 08CD10, 11, 13,
14, S3, 08CD17, 20, 23, 26, 29–31, C€ur€uk Da�g section
(Fig. 3), Western Taurus, Turkey, Kokarkuyu Formation,
Griesbachian, Induan, Early Triassic.
Description. Carapace subtriangular, elongated, rela-
tively massive for the genus (H/L generally �0.5); both
valves similar in shape, size and ornamentation, no over-
lap; Hmax at or in front of anterior one-third of Lmax;
Lmax slightly above midH.
DB: long (�65–80% of Lmax), straight to slightly con-
vex; eventual presence of faint shoulders anteriorly to
PDA and posteriorly to ADA (e.g. Fig. 14F, H, J); PDA
�100�, ADA �130�.VB: long, oblique (angle with horizontal line �15�),
straight to slightly concave.
AB: broadly rounded as half-circle, maximum of curva-
ture at midH; triangular with angulation (�120–125�)located above midH at some specimens (e.g. Fig. 14F, G).
PB: tapered, rounded with a narrower radius of curva-
ture, maximum of convexity in the upper 1/3 of Hmax.
Lateral surface of the valves evenly swollen; smooth
rim along AB and PB; 2 sulci at dorsal part: S2 character-
istic of the genus well expressed in the upper-third of
Hmax, S1 poorly defined in the upper quarter of Hmax;
adventral alae short and faint, slightly pointing outward;
swollen surface faintly reticulated.
Dimensions. H ¼ 142–198 mm; L ¼ 249–342 mm; H/L
¼ 0.51–0.64 (Fig. 15).
Remarks. Until now, the only Griesbachian record of the
Monoceratina genus is an unconfirmed occurrence from
the Late Griesbachian of Pakistan (Sohn 1970). The pres-
ent species is therefore the first known occurrence of the
genus in the Early Griesbachian, directly after the end-
Permian extinction. Because of its bisulcate carapace, M.
hussonae sp. nov. differs from most species of the genus.
It is close to M.? exiqua Styk, 1972 from the Ladinian
(Middle Triassic) of Poland (Styk 1972). However, the
new species is bisulcate, ornamented and DB is not paral-
lel to VB. The new species can also be compared to M.
kozuri Styk, 1972 from the same locality (Styk 1972) but
this species is bisulcate, less elongated, and DB and VB
are not parallel. It differs from M.? buekkensis Kozur,
1985, from the Middle and Late Permian of Hungary
(Kozur 1985), in being shorter, higher, bisulcate, orna-
mented and bearing no denticulation. The new species
also differs from M.? gheorghianae Crasquin-Soleau &
Gradinaru, 1996, from the Early Anisian (Middle Triassic)
of Dobrogea in Romania (Crasquin-Soleau & Gradinaru
1996), in its stockier carapace, PB larger, S2 more
expressed and ornamentation of lateral surface. The
weakly expressed S1 is observed only rarely inMonocera-
tina species and seems to be related to the attachment of
antennulae and antennae (Schornikov 1990).
The size variations of M. hussonae sp. nov. are limited
compared with the other species described here and no
growth stages can be distinguished (Fig. 15). Several
Late Triassic species of Monoceratina are known but only
consist of single specimens with no information on ontog-
eny (e.g. Ye et al. 1977; Wei et al. 1983). The only avail-
able ontogenetic series of Monoceratina species is from
two species from the Maastrichtian (Late Cretaceous) of
Poland (Szczechura 1964): M. umbonata (Williamson,
1848) and M. laevioides similis Szczechura, 1964. The
dimensions of other species from the literature are com-
pared. A compilation of the size distribution of these
Monoceratina species clearly shows that M. hussonae is
much smaller and has more restricted H and L variations.
Monoceratina data reporting only one specimen also show
that M. hussonae is much smaller than other species
(Fig. 16). In this locality, this characteristic is shared at the
superfamilial level and will be further discussed in the
Remarks for Callicythere postiangusta Wei, 1981.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 15
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
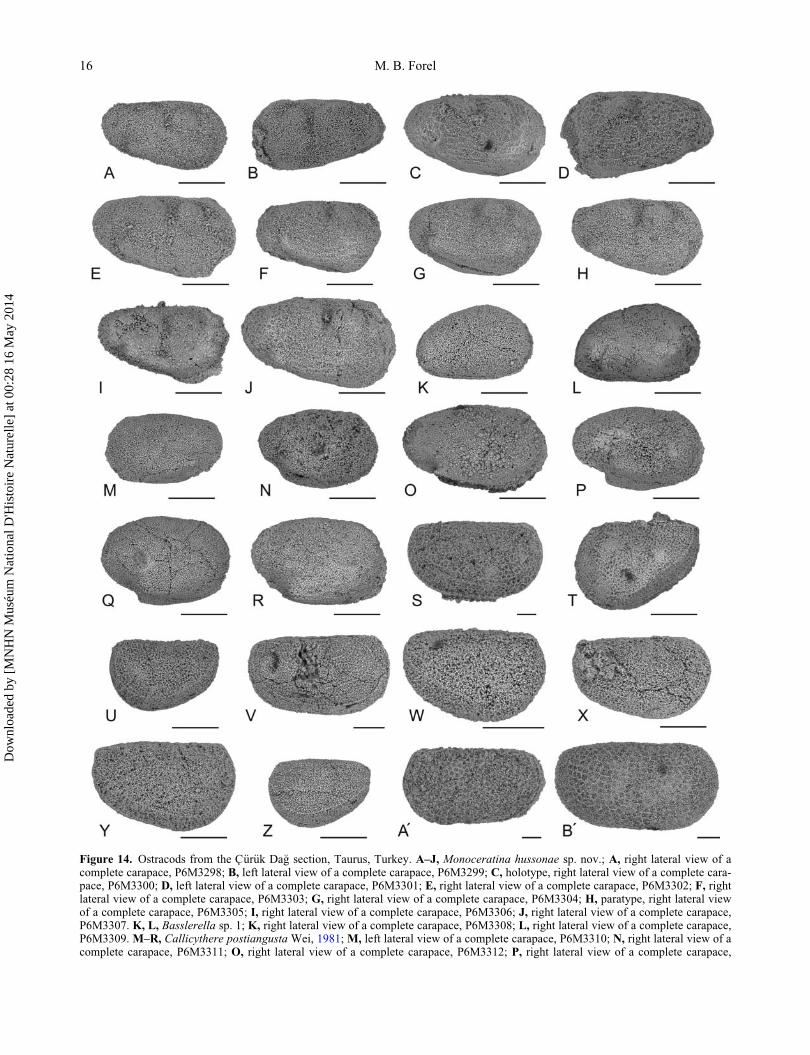
Figure 14. Ostracods from the C€ur€uk Da�g section, Taurus, Turkey. A–J, Monoceratina hussonae sp. nov.; A, right lateral view of acomplete carapace, P6M3298; B, left lateral view of a complete carapace, P6M3299; C, holotype, right lateral view of a complete cara-pace, P6M3300; D, left lateral view of a complete carapace, P6M3301; E, right lateral view of a complete carapace, P6M3302; F, rightlateral view of a complete carapace, P6M3303; G, right lateral view of a complete carapace, P6M3304; H, paratype, right lateral viewof a complete carapace, P6M3305; I, right lateral view of a complete carapace, P6M3306; J, right lateral view of a complete carapace,P6M3307. K, L, Basslerella sp. 1; K, right lateral view of a complete carapace, P6M3308; L, right lateral view of a complete carapace,P6M3309. M–R, Callicythere postiangusta Wei, 1981; M, left lateral view of a complete carapace, P6M3310; N, right lateral view of acomplete carapace, P6M3311; O, right lateral view of a complete carapace, P6M3312; P, right lateral view of a complete carapace,
16 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

Family Cytherissinellidae Kashevarova, 1958
Genus CallicythereWei, 1981
Callicythere postiangustaWei, 1981
(Fig. 14M–R)
1981 Callicythere postiangusta Wei: 504, pl. 1, figs 19–22.
2005 Callicythere postiangusta Wei; Crasquin-Soleau &
Kershaw: pl. 1, figs 1–6.
2012 Callicythere postiangusta Wei; Forel: 25, figs 14T–
W, 15A, B.
Occurrence. Samples CD08A7, 08CD03A-C, 03D-G,
04, 05, S1, 08CD07, S2, 08CD09–11, 13, 14, S4, S3, S10,
08CD14’-16, S7, S8, S9, 08CD18, 20, 21–23, 25–30,
C€ur€uk Da�g section (Fig. 3), Western Taurus, Turkey,
Kokarkuyu Formation (this work). Sichuan Province,
South China (Wei 1981). Laolondong section, Sichuan
Province, South China (Crasquin-Soleau & Kershaw
2005). Dajiang section, Daye Formation, Guizhou Prov-
ince, South China (Forel 2012). All occurrences are from
the Griesbachian, Induan, Early Triassic.
Remarks. Figure 17 plots H versus L for all specimens of
Callicythere postiangusta Wei, 1981. As observed for
Monoceratina hussonae sp. nov., the size distribution of
the present species is narrower than all other documented
species. Few species of Callicythere have been docu-
mented in the literature, making comparisons of size pat-
terns difficult. However, since the narrow size range only
affects species of the superfamily Cytheroidea, tapho-
nomic sorting can be excluded. Owing to the fact that
other Monoceratina species do not show this phenome-
non, the observed limitation of size variation is not a gen-
eral superfamilial characteristic but is restricted to the
Cytheroidea of the studied area, in space and time. As a
first approximation, I consider that this conclusion is also
valid for Callicythere. It could be the result of a specific
physiological response to an environmental stress.
Order Palaeocopida Henningsm€oen, 1953Suborder Kirkbyocopina Gr€undel, 1969
Superfamily Kirkbyoidea Ulrich & Bassler, 1906
Family Kirkbyidae Ulrich & Bassler, 1906
Genus Reviya Sohn, 1961
Reviya sylvieae sp. nov.
(Fig. 14S–Z)
Diagnosis. Species of Reviya with ridge joining cardinal
angles and parallel to free margin; surface reticulated.
Derivation of name. Dedicated to Dr Sylvie Crasquin,
CNRS, Universit�e Pierre et Marie Curie, Paris, France.
Material. Holotype: P6M3316, one complete carapace
(heteromorph; Fig. 14S), sample S8. Paratype: P6M3319,
one complete carapace (heteromorph; Fig. 14V), sample
S10. All from the C€ur€uk Da�g section, Western Taurus,
Turkey, Kokarkuyu Formation, Griesbachian, Induan,
Early Triassic (this work). Nineteen complete carapaces,
several fragments.
Occurrence. Samples 08CD03D1, 14, S3, S10, 08CD16,
17, S8, 08CD20–23, C€ur€uk Da�g section (Fig. 3), Western
Taurus, Turkey, Kokarkuyu Formation, Griesbachian,
Induan, Early Triassic (this work).
Description. Kirkbyan pit poorly defined, visible on
some broken specimens. Lateral surface of the valves: sur-
face reticulated; thin ridge parallel to free margin extend-
ing from ACA to PCB.
Heteromorphs (,; Fig. 14S, V): carapace subrectangu-
lar, slightly preplete; Lmax above midH; LV larger than
RV, slight overlap all along the free margin; DB straight
and long, �80–85% of Lmax; cardinal angles sharp,
PCA�100–110�, ACA�120�; PDB and ADB, short,
faintly convex and sharply bent ventrally; AB and PB
rounded with a smaller maximum of curvature at PB; ante-
rior maximum of convexity located around midH, posterior
maximum of convexity slightly above; VB straight, slightly
bent toward AB, �60% of Lmax; ventral angles blunt,
AVB�PVB�130�; AVB longer and more convex than
PVB; lateral surface with occasional thin ridges.
Tecnomorphs (<; Fig. 14T, U, W–Z): carapace subtrian-
gular, highly preplete with Hmax in the anterior one-third
of midL; Lmax above midH; LV larger than RV, slight
overlap all along the free margin; DB long and straight,
slightly shorter than Lmax; cardinal angles well expressed,
ACA �120–140�, PCA �90–110�; VB straight to slightly
convex, sharply bent toward AB; AB large and broadly
rounded, maximum of curvature above midL; PB narrow
finely reticulated with no accessory ridges.
Dimensions (heteromorphs and tecnomorphs). H ¼146–391 mm; L ¼ 220–708 mm; H/L ¼ 0.55–0.72
(Fig. 18).
P6M3313; Q, right lateral view of a complete carapace, P6M3314; R, right lateral view of a complete carapace, P6M3315. S–Z, Reviyasylvieae sp. nov.; S, holotype, right lateral view of a complete carapace, heteromorph, P6M3316; T, left lateral view of a complete cara-pace, tecnomorph, P6M3317; U, left lateral view of a complete carapace, tecnomorph, P6M3318; V, paratype, right lateral view of acomplete carapace, heteromorph, P6M3319; W, right lateral view of a complete carapace, tecnomorph, P6M3320; X, right lateral viewof a complete carapace, tecnomorph, P6M3321; Y, right lateral view of a complete carapace, tecnomorph, P6M3322; Z, left lateral viewof a complete carapace, tecnomorph, P6M3323. A0–B0, Reviya curukensis Crasquin-Soleau, 2004b; A0, left lateral view of a completecarapace, P6M3324; B0, right lateral view of a complete carapace, P6M3325. Scale bars are 100 mm.
3
Heterochronic growth of ostracods (Crustacea) from microbial deposits 17
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
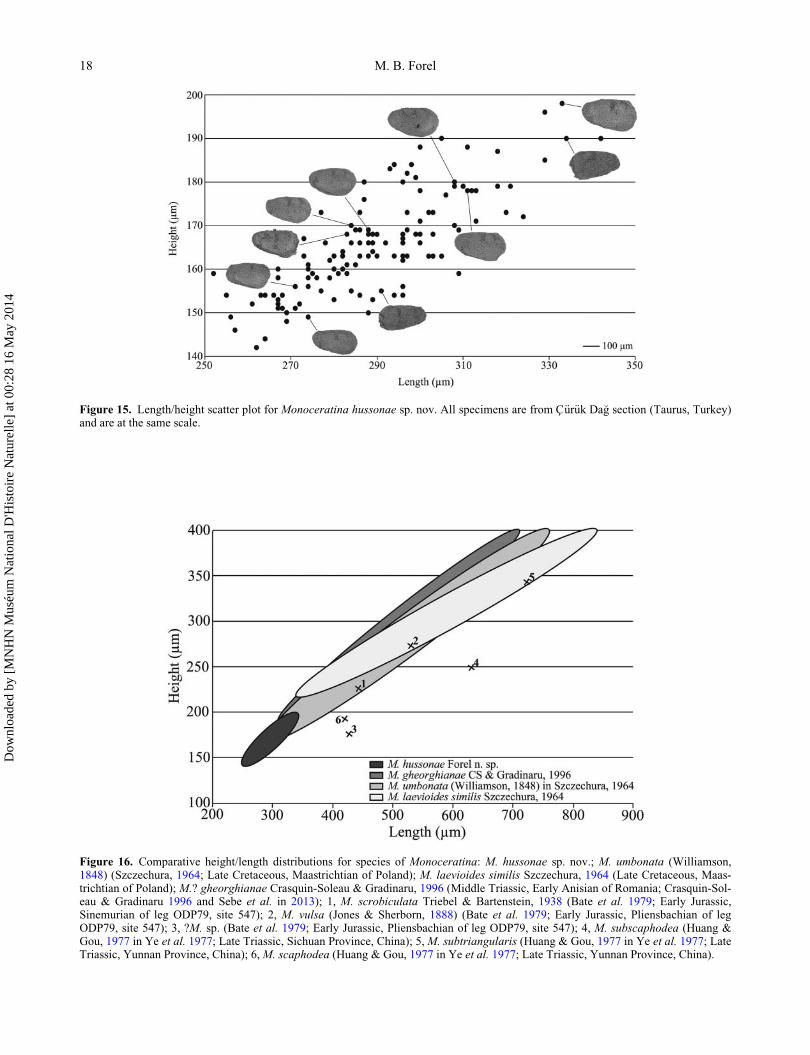
Figure 15. Length/height scatter plot for Monoceratina hussonae sp. nov. All specimens are from C€ur€uk Da�g section (Taurus, Turkey)and are at the same scale.
Figure 16. Comparative height/length distributions for species of Monoceratina: M. hussonae sp. nov.; M. umbonata (Williamson,1848) (Szczechura, 1964; Late Cretaceous, Maastrichtian of Poland); M. laevioides similis Szczechura, 1964 (Late Cretaceous, Maas-trichtian of Poland); M.? gheorghianae Crasquin-Soleau & Gradinaru, 1996 (Middle Triassic, Early Anisian of Romania; Crasquin-Sol-eau & Gradinaru 1996 and Sebe et al. in 2013); 1, M. scrobiculata Triebel & Bartenstein, 1938 (Bate et al. 1979; Early Jurassic,Sinemurian of leg ODP79, site 547); 2, M. vulsa (Jones & Sherborn, 1888) (Bate et al. 1979; Early Jurassic, Pliensbachian of legODP79, site 547); 3, ?M. sp. (Bate et al. 1979; Early Jurassic, Pliensbachian of leg ODP79, site 547); 4, M. subscaphodea (Huang &Gou, 1977 in Ye et al. 1977; Late Triassic, Sichuan Province, China); 5,M. subtriangularis (Huang & Gou, 1977 in Ye et al. 1977; LateTriassic, Yunnan Province, China); 6,M. scaphodea (Huang & Gou, 1977 in Ye et al. 1977; Late Triassic, Yunnan Province, China).
18 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
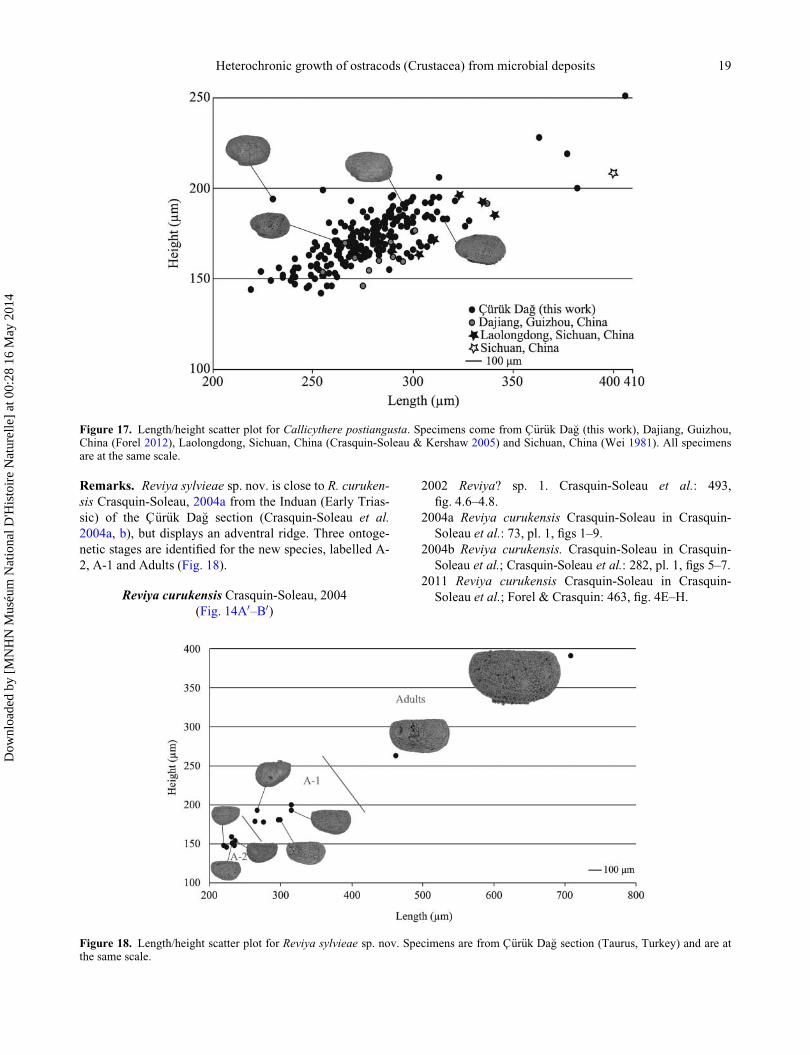
Remarks. Reviya sylvieae sp. nov. is close to R. curuken-
sis Crasquin-Soleau, 2004a from the Induan (Early Trias-
sic) of the C€ur€uk Da�g section (Crasquin-Soleau et al.
2004a, b), but displays an adventral ridge. Three ontoge-
netic stages are identified for the new species, labelled A-
2, A-1 and Adults (Fig. 18).
Reviya curukensis Crasquin-Soleau, 2004
(Fig. 14A0–B0)
2002 Reviya? sp. 1. Crasquin-Soleau et al.: 493,
fig. 4.6–4.8.
2004a Reviya curukensis Crasquin-Soleau in Crasquin-
Soleau et al.: 73, pl. 1, figs 1–9.
2004b Reviya curukensis. Crasquin-Soleau in Crasquin-
Soleau et al.; Crasquin-Soleau et al.: 282, pl. 1, figs 5–7.
2011 Reviya curukensis Crasquin-Soleau in Crasquin-
Soleau et al.; Forel & Crasquin: 463, fig. 4E–H.
Figure 17. Length/height scatter plot for Callicythere postiangusta. Specimens come from C€ur€uk Da�g (this work), Dajiang, Guizhou,China (Forel 2012), Laolongdong, Sichuan, China (Crasquin-Soleau & Kershaw 2005) and Sichuan, China (Wei 1981). All specimensare at the same scale.
Figure 18. Length/height scatter plot for Reviya sylvieae sp. nov. Specimens are from C€ur€uk Da�g section (Taurus, Turkey) and are atthe same scale.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 19
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
2013a Reviya curukensis Crasquin-Soleau in Crasquin-
Soleau et al.; Forel et al.: fig. 4U, V.
Occurrence. Samples S7, TK55–58, TK117–119, 121,
122, C€ur€uk Da�g section (Fig. 3), Western Taurus, Turkey,
Kokarkuyu Formation (this work; Crasquin-Soleau et al.
2002, 2004a, b). Meishan section, Zhejiang Province,
South China (Forel & Crasquin 2011), B�alv�any North sec-tion, Gerennav�ar Formation, B€ukk Mountains, Hungary,
Griesbachian (Forel et al. 2013a). All occurrences are
from the Griesbachian, Induan, Early Triassic.
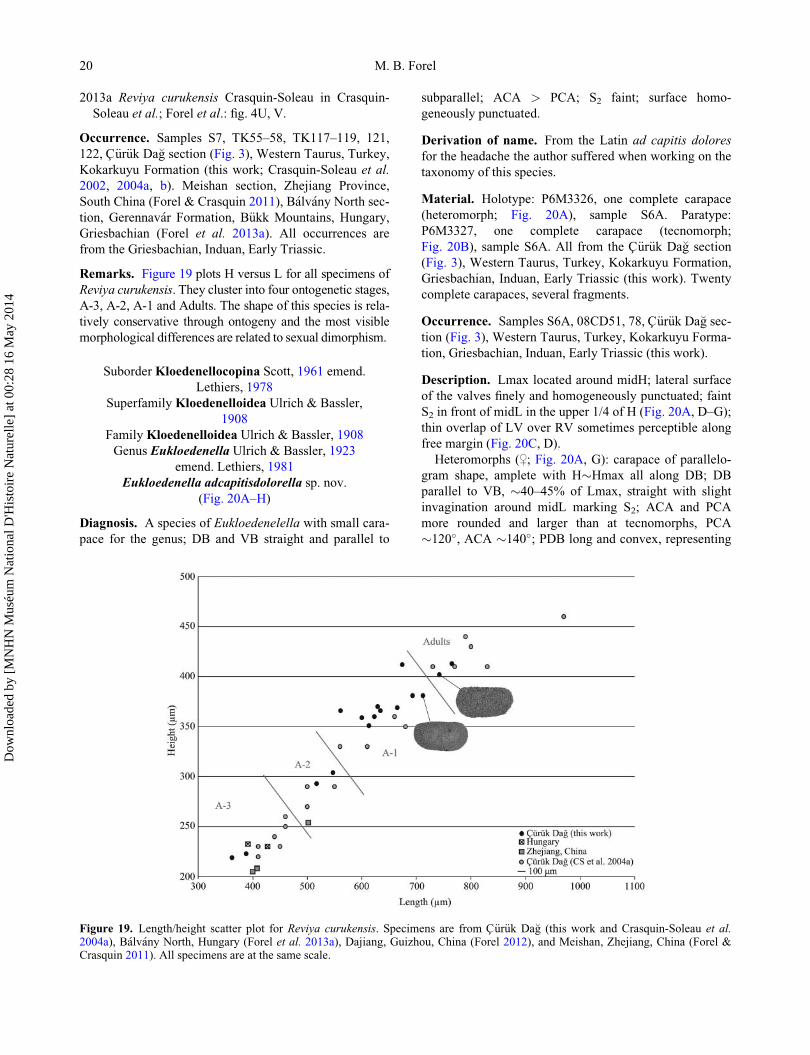
Remarks. Figure 19 plots H versus L for all specimens of
Reviya curukensis. They cluster into four ontogenetic stages,
A-3, A-2, A-1 and Adults. The shape of this species is rela-
tively conservative through ontogeny and the most visible
morphological differences are related to sexual dimorphism.
Suborder Kloedenellocopina Scott, 1961 emend.
Lethiers, 1978
Superfamily Kloedenelloidea Ulrich & Bassler,
1908
Family Kloedenelloidea Ulrich & Bassler, 1908
Genus Eukloedenella Ulrich & Bassler, 1923
emend. Lethiers, 1981
Eukloedenella adcapitisdolorella sp. nov.
(Fig. 20A–H)
Diagnosis. A species of Eukloedenelella with small cara-
pace for the genus; DB and VB straight and parallel to
subparallel; ACA > PCA; S2 faint; surface homo-
geneously punctuated.
Derivation of name. From the Latin ad capitis dolores
for the headache the author suffered when working on the
taxonomy of this species.
Material. Holotype: P6M3326, one complete carapace
(heteromorph; Fig. 20A), sample S6A. Paratype:
P6M3327, one complete carapace (tecnomorph;
Fig. 20B), sample S6A. All from the C€ur€uk Da�g section
(Fig. 3), Western Taurus, Turkey, Kokarkuyu Formation,
Griesbachian, Induan, Early Triassic (this work). Twenty
complete carapaces, several fragments.
Occurrence. Samples S6A, 08CD51, 78, C€ur€uk Da�g sec-tion (Fig. 3), Western Taurus, Turkey, Kokarkuyu Forma-
tion, Griesbachian, Induan, Early Triassic (this work).
Description. Lmax located around midH; lateral surface
of the valves finely and homogeneously punctuated; faint
S2 in front of midL in the upper 1/4 of H (Fig. 20A, D–G);
thin overlap of LV over RV sometimes perceptible along
free margin (Fig. 20C, D).
Heteromorphs (,; Fig. 20A, G): carapace of parallelo-
gram shape, amplete with H�Hmax all along DB; DB
parallel to VB, �40–45% of Lmax, straight with slight
invagination around midL marking S2; ACA and PCA
more rounded and larger than at tecnomorphs, PCA
�120�, ACA �140�; PDB long and convex, representing
Figure 19. Length/height scatter plot for Reviya curukensis. Specimens are from C€ur€uk Da�g (this work and Crasquin-Soleau et al.2004a), B�alv�any North, Hungary (Forel et al. 2013a), Dajiang, Guizhou, China (Forel 2012), and Meishan, Zhejiang, China (Forel &Crasquin 2011). All specimens are at the same scale.
20 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
� two-thirds of Hmax; PB more rounded and prominent
than at tecnomorphs, with a narrower radius of curvature,
maximum located slightly above midH; ADB strongly
convex and long, being two-thirds of Hmax; AB rounded
with a narrower radius of curvature, maximum below
midH, �40% of Hmax; VB somewhat shorter than DB,
straight to slightly concave at middle part; PVA �110–
120�, AVA �140–150�; PVB long, convex and parallel to
ADB; AVB short parallel to PDB; sexual dimorphism
strong and atypical on the entire ventral half of the cara-
pace, becoming larger and massive with rectangular out-
line, lowering of the anterior maximum of convexity
resulting in a ventrally twisted aspect of the anterior por-
tion of the carapace.
Tecnomorphs (<; Fig. 20B–F, H): carapace subovate,
slightly tapered posteriorly, preplete, Hmax around ante-
rior one-third of Lmax; DB straight to slightly convex,
long, about 70% of Lmax; ACA and PCA relatively sharp,
PCA > 90�, ACA �130�; PDB short; PB rounded with
maximum of curvature located in the upper third of
Hmax; ADB gently convex and long representing 40–
50% of Hmax; AB rounded with a large radius of convex-
ity, maximum located around midH; VB straight to
slightly convex, parallel to subparallel to DB, slightly
bent toward AB; AVA�PVA�150�; AVB and PVB
rounded; PVB narrower and longer than AVB, �65% of
Hmax.
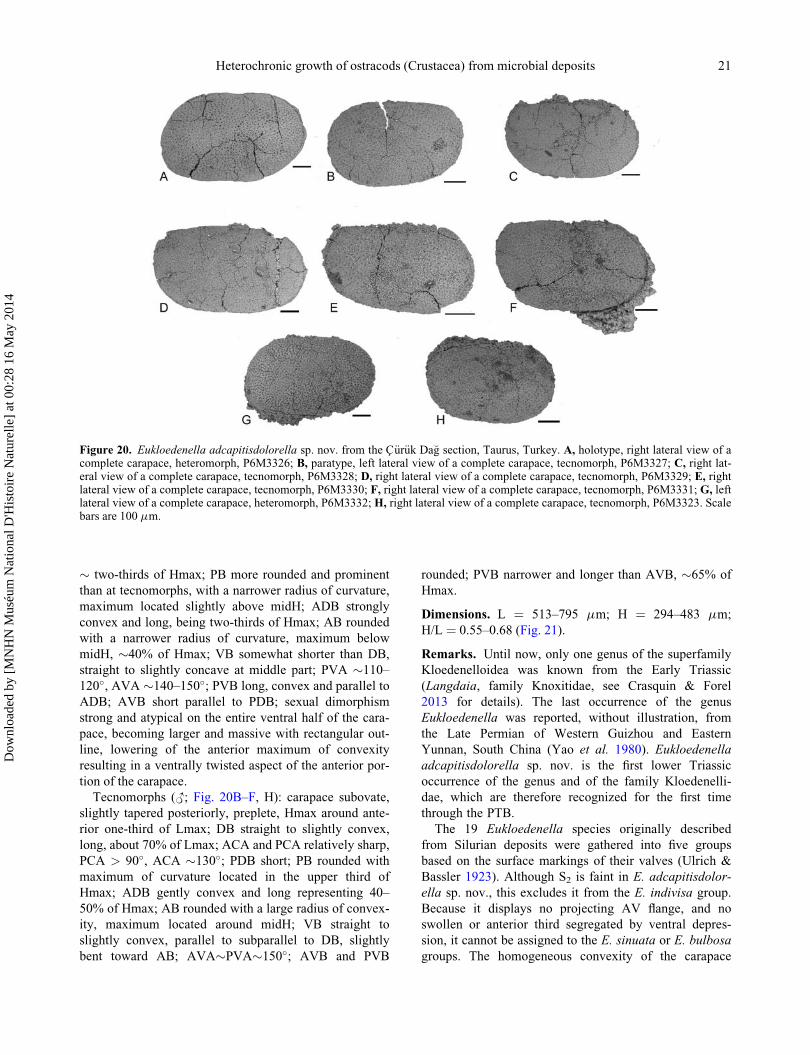
Dimensions. L ¼ 513–795 mm; H ¼ 294–483 mm;
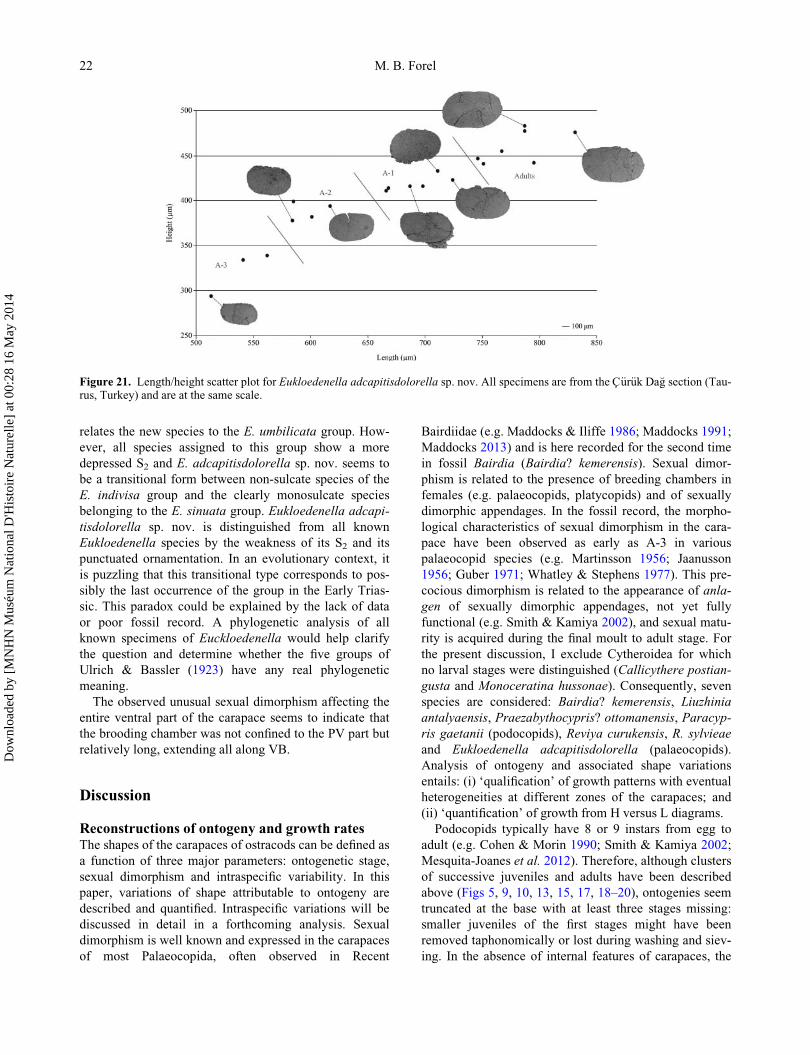
H/L ¼ 0.55–0.68 (Fig. 21).
Remarks. Until now, only one genus of the superfamily
Kloedenelloidea was known from the Early Triassic
(Langdaia, family Knoxitidae, see Crasquin & Forel
2013 for details). The last occurrence of the genus
Eukloedenella was reported, without illustration, from
the Late Permian of Western Guizhou and Eastern
Yunnan, South China (Yao et al. 1980). Eukloedenella
adcapitisdolorella sp. nov. is the first lower Triassic
occurrence of the genus and of the family Kloedenelli-
dae, which are therefore recognized for the first time
through the PTB.
The 19 Eukloedenella species originally described
from Silurian deposits were gathered into five groups
based on the surface markings of their valves (Ulrich &
Bassler 1923). Although S2 is faint in E. adcapitisdolor-
ella sp. nov., this excludes it from the E. indivisa group.
Because it displays no projecting AV flange, and no
swollen or anterior third segregated by ventral depres-
sion, it cannot be assigned to the E. sinuata or E. bulbosa
groups. The homogeneous convexity of the carapace
Figure 20. Eukloedenella adcapitisdolorella sp. nov. from the C€ur€uk Da�g section, Taurus, Turkey. A, holotype, right lateral view of acomplete carapace, heteromorph, P6M3326; B, paratype, left lateral view of a complete carapace, tecnomorph, P6M3327; C, right lat-eral view of a complete carapace, tecnomorph, P6M3328; D, right lateral view of a complete carapace, tecnomorph, P6M3329; E, rightlateral view of a complete carapace, tecnomorph, P6M3330; F, right lateral view of a complete carapace, tecnomorph, P6M3331; G, leftlateral view of a complete carapace, heteromorph, P6M3332; H, right lateral view of a complete carapace, tecnomorph, P6M3323. Scalebars are 100 mm.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 21
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
relates the new species to the E. umbilicata group. How-
ever, all species assigned to this group show a more
depressed S2 and E. adcapitisdolorella sp. nov. seems to
be a transitional form between non-sulcate species of the
E. indivisa group and the clearly monosulcate species
belonging to the E. sinuata group. Eukloedenella adcapi-
tisdolorella sp. nov. is distinguished from all known
Eukloedenella species by the weakness of its S2 and its
punctuated ornamentation. In an evolutionary context, it
is puzzling that this transitional type corresponds to pos-
sibly the last occurrence of the group in the Early Trias-
sic. This paradox could be explained by the lack of data
or poor fossil record. A phylogenetic analysis of all
known specimens of Euckloedenella would help clarify
the question and determine whether the five groups of
Ulrich & Bassler (1923) have any real phylogenetic
meaning.
The observed unusual sexual dimorphism affecting the
entire ventral part of the carapace seems to indicate that
the brooding chamber was not confined to the PV part but
relatively long, extending all along VB.
Discussion
Reconstructions of ontogeny and growth ratesThe shapes of the carapaces of ostracods can be defined as
a function of three major parameters: ontogenetic stage,
sexual dimorphism and intraspecific variability. In this
paper, variations of shape attributable to ontogeny are
described and quantified. Intraspecific variations will be
discussed in detail in a forthcoming analysis. Sexual
dimorphism is well known and expressed in the carapaces
of most Palaeocopida, often observed in Recent
Bairdiidae (e.g. Maddocks & Iliffe 1986; Maddocks 1991;
Maddocks 2013) and is here recorded for the second time
in fossil Bairdia (Bairdia? kemerensis). Sexual dimor-
phism is related to the presence of breeding chambers in
females (e.g. palaeocopids, platycopids) and of sexually
dimorphic appendages. In the fossil record, the morpho-
logical characteristics of sexual dimorphism in the cara-
pace have been observed as early as A-3 in various
palaeocopid species (e.g. Martinsson 1956; Jaanusson
1956; Guber 1971; Whatley & Stephens 1977). This pre-
cocious dimorphism is related to the appearance of anla-
gen of sexually dimorphic appendages, not yet fully
functional (e.g. Smith & Kamiya 2002), and sexual matu-
rity is acquired during the final moult to adult stage. For
the present discussion, I exclude Cytheroidea for which
no larval stages were distinguished (Callicythere postian-
gusta and Monoceratina hussonae). Consequently, seven
species are considered: Bairdia? kemerensis, Liuzhinia
antalyaensis, Praezabythocypris? ottomanensis, Paracyp-
ris gaetanii (podocopids), Reviya curukensis, R. sylvieae
and Eukloedenella adcapitisdolorella (palaeocopids).
Analysis of ontogeny and associated shape variations
entails: (i) ‘qualification’ of growth patterns with eventual
heterogeneities at different zones of the carapaces; and
(ii) ‘quantification’ of growth from H versus L diagrams.
Podocopids typically have 8 or 9 instars from egg to
adult (e.g. Cohen & Morin 1990; Smith & Kamiya 2002;
Mesquita-Joanes et al. 2012). Therefore, although clusters
of successive juveniles and adults have been described
above (Figs 5, 9, 10, 13, 15, 17, 18–20), ontogenies seem
truncated at the base with at least three stages missing:
smaller juveniles of the first stages might have been
removed taphonomically or lost during washing and siev-
ing. In the absence of internal features of carapaces, the
Figure 21. Length/height scatter plot for Eukloedenella adcapitisdolorella sp. nov. All specimens are from the C€ur€uk Da�g section (Tau-rus, Turkey) and are at the same scale.
22 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

cluster of the largest specimens of each reconstructed line-
age are considered to be adults.
Variation in shape through ontogeny. Shape variations
through ontogeny are studied by comparison of the shapes
of individuals of each growth stage in the seven selected
species (Fig. 22). Bairdia? kemerensis (Fig. 22A) shows a
relatively conservative contour. The dominant modifica-
tions are located at both extremities, with the progressive
upward migration of the posterior and anterior maxima of
convexity. This pattern results in an overall change of the
lateral outline of the carapace: from diamond-shaped in
A-4 (Lmax below Hmax) to close to rectangular in adults
(Lmax around Hmax). Liuzhinia antalyaensis and Praeza-
bythocypris? ottomanensis exhibit stronger shape modifi-
cations distributed all around the carapace. The latter
(Fig. 22B) shows enlargement of PB with the upward
migration of the maximal convexity, an increase of round-
ness at AB with a downward shift of the maximum con-
vexity, differentiation of an anteriorly inclined ADB, and
the appearance of convexity at VB. These morphological
changes mostly occur between A-3 and A-2; later stages
only showing strengthening of these characteristics. Liuz-
hinia antalyaensis (Fig. 22C) displays a relatively conser-
vative VB, becoming flatter and longer through ontogeny.
Most of the changes are located in the upper part of the
carapace, which becomes more symmetrical with wider
PB and more constant Hmax. AB becomes wider and
more rounded. Compared to Praezabythocypris? ottoma-
nensis, the observed changes are more gradual and homo-
geneously distributed amongst the successive larval
stages.
Figure 22. Mean shapes of each instar (right valves only) reconstructed for A, Bairdia? kemerensis, B, Praezabythocypris? ottomanensis,C, Liuzhinia antalyaensis, D, Paracypris gaetanii, E, Reviya sylvieae, F, R. curukensis and G, Eukloedenella adcapitisdolorella.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 23
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

The adult contour of Paracypris gaetanii has not been
represented because only left valves are available
(Fig. 22D). This species displays the most conservative
shape from A-4 to A-1: the shape of stage A-(nþ1) glob-
ally appears as an enlargement of A-n. Shape modifica-
tions mostly affect the roundness of the extremities, PB
becoming narrower and AB becoming larger. Therefore,
the dominant pattern of ontogeny for Paracypris gaetanii
is size increase, with secondary shape changes located at
both AB and PB. Most changes observed through the
ontogeny of Reviya sylvieae (Fig. 22E) affect the ventral
and posterior portions of the carapace, DB and AB being
very conservative. PB becomes larger and more rounded,
with the maximum of curvature migrating ventrally, while
VB becomes longer, straighter and parallel to DB. Reviya
curukensis displays slight modifications, with PB enlarg-
ing and VB lengthening to become parallel to DB
(Fig. 22F). The last moult from A-1 to Adult records very
different patterns between these two species: a high
increase of size for R. sylvieae but a small one for R.
curukensis.
Modifications of Eukloedenella adcapitisdolorella
(Fig. 22G) are observed mostly in PB, VB and AB. The
patterns are similar but more subtle than those described
for the species of Reviya: PB more rounded with the maxi-
mum of curvature migrating ventrally, elongation and hor-
izontalization of VB, less marked ACA, and AB
protruding anteriorly. In the three palaeocopids studied
here, DB shape is very conservative through ontogeny,
being only affected by strict elongation.
Growth rates. Brooks’s rule states that the volume of
crustaceans should double with each moult and linear
dimensions should consequently increase by 1.26 (Brooks
1886; Teissier 1960). This rule is a generalization of
growth and exceptions may provide insights into biology,
ontogeny and life cycle. Although processes accounting
for this pattern are still unknown, the growth of ostracods
follows Brooks’s rule with some amount of variability
(e.g. Hessland 1949; Sohn 1950; Kesling 1951a, b, 1952,
1953; Shaver 1953; Kesling & Takagi 1961; Kesling &
Crafts 1962). In their review, Danielopol et al. (2008)
documented values ranging from 1.21 to 1.37 for non-
marine species. Such information has not been available
previously for marine species.
The match to Brooks’s rule can be evaluated by com-
paring the observed and predicted growth rates. For the
following discussion, only specimens from the C€ur€uk Da�gsection are considered based on this work and Crasquin-
Soleau et al. (2002, 2004a, b). Lmean and Hmean for all
specimens of each ontogenetic stage of Bairdia? kemeren-
sis, Liuzhinia antalyaensis, Praezabythocypris? ottoma-
nensis, Paracypris gaetanii, Reviya sylvieae, R.
curukensis and Eukloedenella adcapitisdolorella were
determined. Growth rates of Hmean and Lmean were
calculated for each ontogenetic transition in the following
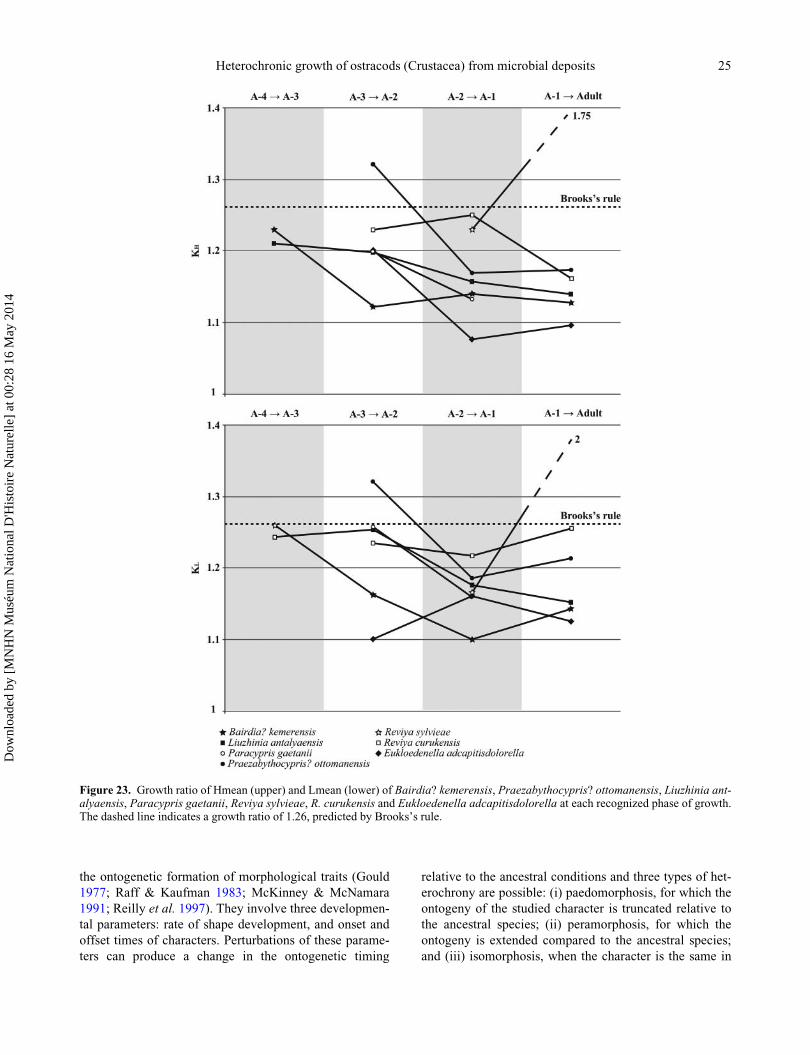
way (Fig. 23):
KH ¼ Hnþ1=Hn
KL ¼ Lnþ1=Ln;
where KH and KL are respectively Hmean and Lmean
growth rates; Hn and Ln are respectively Hmean and
Lmean at ontogenetic stage n; and Hnþ1 and Lnþ1 are
respectively Hmean and Lmean at ontogenetic stage nþ1.
For most of the species, KH and KL are generally lower
than predicted by Brooks’s rule and diminish through
ontogeny. Values range from 1.07 to 1.75 for KH, and
from 1.09 to 2 for KL. They record a decelerating and
sluggish growth, with an exception for the species Reviya
sylvieae. The slowest growth in length and height is
recorded in Bairdia? kemerensis and Eukloedenella adca-
pitisdolorella respectively. The growth of all podocopid
species displays the same decreasing pattern through
time: KH and KL decrease strongly to A-2, KH then stays
stable while KL increases before Adult stage. KH in Liuz-
hinia antalyaensis shows a constant decrease, while KL is
stable until A-2 and then decreases sharply. Although
only two transitions are considered for Paracypris gaeta-
nii, the same strong decreasing trend is recognized for
both KH and KL. All growth values for Bairdia? kemeren-
sis, Liuzhinia antalyaensis and Paracypris gaetanii are
below values predicted from Brooks’s rule. Praezabytho-
cypris? ottomanensis displays the only growth event
above Brooks’s rule, at the transition A-3 to A-2. It is then
followed by a sharp decrease and a slight increase before
Adult stage. For each ontogenetic transition in all species,
KL > KH: growth in length is greater and faster than in
height. Palaeocopid growth differs amongst the three spe-
cies under scrutiny and from podocopids. KH and KL of
the two Reviya species are below that predicted from
Brooks’s rule and anti-correlated: (i) from A-3 to A-1, KH
decreases for R. sylvieae but increases for R. curukensis,
while KL increases for R. sylvieae but decreases for R.
curukensis; (ii) from A-2 to Adult, KH increases for R. syl-
vieae but decreases for R. curukensis, while KL decreases
for R. sylvieae but increases for R. curukensis. However,
the general trend recognized for these two species is a
decrease in both KH and KL through ontogeny with values
below those predicted from Brooks’s rule. Eukloedenella
adcapitisdolorella shows a different and striking develop-
ment, with strong growth from A-2 to Adult for both KH
and KL, from values below the Brook’s rule prediction for
A-2 to A-1, to values far above it for A-1 to Adult.
Although early growth stages are missing, the observed
increase during final moulting is unusual and previously
unknown for any other ostracods.
Heterochrony. Heterochronies are generally defined as
changes in the timing and/or rates of processes underlying
24 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
the ontogenetic formation of morphological traits (Gould
1977; Raff & Kaufman 1983; McKinney & McNamara
1991; Reilly et al. 1997). They involve three developmen-
tal parameters: rate of shape development, and onset and
offset times of characters. Perturbations of these parame-
ters can produce a change in the ontogenetic timing
relative to the ancestral conditions and three types of het-
erochrony are possible: (i) paedomorphosis, for which the
ontogeny of the studied character is truncated relative to
the ancestral species; (ii) peramorphosis, for which the
ontogeny is extended compared to the ancestral species;
and (iii) isomorphosis, when the character is the same in
Figure 23. Growth ratio of Hmean (upper) and Lmean (lower) of Bairdia? kemerensis, Praezabythocypris? ottomanensis, Liuzhinia ant-alyaensis, Paracypris gaetanii, Reviya sylvieae, R. curukensis and Eukloedenella adcapitisdolorella at each recognized phase of growth.The dashed line indicates a growth ratio of 1.26, predicted by Brooks’s rule.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 25
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

the ancestor and descendant but the descendant has fol-
lowed a different ontogenetic trajectory to reach the same
shape. Paedomorphosis can be produced by deceleration
(slower rate), hypomorphosis (earlier offset time) or post-
displacement (later onset time). Peramorphosis can result
from acceleration (faster rate), hypermorphosis (later off-
set time) or pre-displacement (earlier onset time) (see
Reilly et al. 1997 for further details). Harries et al. (1996)
identified paedomorphosis as an adaptative mechanism to
survive ecological and environmental perturbations asso-
ciated with mass extinctions, with examples of progenetic
organisms (hypomorphotic according to Reilly et al.
1997). Moderate thermal fluctuations might be associated
with fast growth, while the retardation of development
was described in fluctuating regimes encompassing
extreme temperatures (Thorp & Wineriter 1981; Worner
1992). Deceleration is known for molluscs under the influ-
ence of warming that led to nutrient-depleted waters dur-
ing the Miocene (Sch€one et al. 2004), while acceleration
in fluctuating temperatures has been reported for Daphnia
(Orcutt & Porter 1983) and rotifers (Halbach 1973). Pro-
genesis (e.g. hypomorphosis) and post-displacement are
shown in Recent ostracods of the continental shelf of the
Congo and Senegal and are related to strong upwelling
and increased sedimentation rates (Bertholon 1997). Pera-
morphic and paedomorphic evolution have been impli-
cated as speciation mechanisms in Loxoconcha species
during the Pliocene–Pleistocene of the Western Pacific
(Tanaka & Ikeya 2002). Deceleration of growth rates has
been observed for several Cypridoidea (podocopids) at
low temperatures (Ganning 1971). Despite these exam-
ples, data are more abundant for freshwater ostracods, for
which it is known that salinity modulates hatching phenol-
ogy, survival and moulting, too low conductivity having a
negative effect on ostracod survival and growth (De Deck-
ker 1983; Mezquita et al. 1999). The final moulting of the
Recent Heterocypris barbara is delayed and mean size of
the adult stage decreases with lowered salinity (Rossi
et al. 2013). On the other hand, many species display
faster developmental time at higher temperatures and at
optimal salinity (Cohen & Morin 1990).
A key element in any discussion about heterochrony is
the definition of the terminal shape of the studied parame-
ter. Because the present discussion focuses on the general
ontogenetic trajectories of ostracods, the terminal shape
should be their shape at sexual maturity, i.e. at the adult
stage. The slow growth of podocopids and species of
Reviya could therefore be related to paedomorphosis by
deceleration or isomorphosis by deceleration and hyper-
morphosis, while the fast development of Eukloedenella
could result from peramorphosis by acceleration or iso-
morphosis by acceleration and hypomorphosis. Conse-
quently these patterns might result, respectively, from
paedomorphic growth by deceleration and peramorphic
growth by acceleration. As a first approximation, the
growth discrepancies between podocopids/Reviya and
Eukloedenella seem to be related to their different palae-
obiology. Deceleration of the growth of podocopids/
Reviya at A-3 or A-2 indicates that their optimal resilience
should have been reached at these stages. In contrast, the
strong acceleration of Eukloedenella growth to reach adult
stage seems to indicate that it was the most resilient for
this species.
Ostracods surviving to the EPE in association with the
PTBM (Permian-Triassic boundary microbialites) of the
C€ur€uk Da�g section are characterized by: (i) proliferation
of specimens; (ii) abundant small-sized individuals; and
(iii) developmental heterochronies for at least seven spe-
cies. Although they clearly record persistence of environ-
mental stress, how did such faunas maintain their viability
throughout the deposition of microbial deposits? To
address this problem, the way an organism acquires
energy from its environment and allocates it to different
biological processes should first be discussed. Organisms
extract resources from the environment, and use them for
three basic functions: somatic maintenance, growth and
reproduction. For a fixed amount of resource, there is a
trade-off between these functions (Roff 1992), depending
on: (i) the state of the organism, i.e. body volume
(described by size-related phenomena such as food uptake
and growth), energy reserves and ageing process; and (ii)
the environment (e.g. Lika & Kooijman 2003). Those
resources allocated to reproduction can sustain either the
production of numerous small eggs or a few large ones
(e.g. Oloffson et al. 2009). Life history theory is con-
cerned with the way individuals allocate resources to
growth and reproduction, based on the assumptions that
resources are limited and that physiological constraints act
on growth and reproduction (Roff 1992; Stearns 1992).
Ostracods do not exhibit simultaneous growth and
reproduction, except in cases of precocious sexual matu-
rity. Their reproductive effort is set to 0 until a certain age
at which growth ceases and energy is switched to repro-
duction (Perrin 1992; Bulmer 1994). As a consequence,
energy resources of eggs and juveniles are entirely dedi-
cated to maturation and conservation of the achieved
degree of maturity, while they are allocated to mainte-
nance and reproduction in adults (Lika & Kooijman
2003). There are few papers on the life history strategies
of marine ostracods but most of these organisms appear to
be iteroparous, with potentially more than one reproduc-
tive event during the adulthood of the females. It is inter-
esting to estimate how many eggs an adult female can
produce under normal and optimal conditions. To do so,
three parameters are important: the number of reproduc-
tive events per year, the number of eggs per clutch and the
lifetime of adult females. Since Bairdioidea dominate the
studied PTBM assemblages and the life cycles of
26 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

palaeocopids are relatively unknown, I will only consider
Bairdioidea. Data on longevity, breeding cycles and num-
ber of generations per year (voltinism) is scattered for
Bairdioidea, but the literature does contain some informa-
tion on other marine podocopids. Cytheroidea with sea-
sonal reproduction produce from one to four generations
per year (Horne 1983). The number of eggs per clutch is
also poorly known but is 11–50 in Cytheroidea (Cohen &
Morin 1990) and 10 to more than 20 in Cypridoidea (Mad-
docks 1990). Data for Bairdioidea are rare but Maddocks
(2013, fig. 9D, E) shows at least eight eggs of Neonesidea
tenera (Brady) in uteri, while N. oligodentata lays eggs
either singly or in groups of 7–10 (Obato 1999). Adult life-
time is known to last from 7 days to 3 months in Cytheroi-
dea, and from 1.5 to 7 months in Cypridoidea (Cohen &
Morin 1990). In the absence of precise data for Bairdioi-
dea, mean values of data observed for other podocopids
(Cypridoidea, Cytheroidea) can be used to estimate the
fecundity of adult female Bairdioidea. Considering a life-
time of 7 months, 1–4 reproductive events per year (¼ 1–2
per lifetime), 7–10 eggs per clutch, then the fecundity of a
Bairdioidea female would be 7–20 eggs in a lifetime under
optimal environmental and ecological conditions.
The observed reduction of the body size of ostracods in
the PTBM implies a reduction of the space potentially
available for eggs. To cope with this phenomenon, two
adaptations of the production of eggs are conceivable,
known as bet-hedging: (i) the same quantity of smaller
size eggs, or (ii) fewer eggs of a larger size. They have
been reported frequently and studied for all kinds of
organisms, including arthropods (Fox & Czesak 2000).
Bet-hedging strategy has been evoked to describe the
asynchronous hatching of resting eggs in populations of
freshwater Eucypris virens (Martins et al. 2008). Modifi-
cations of the production of eggs in marine ostracods
were discussed by Heip (1976), without quantification, for
the Recent Cyprideis torosa. This author considered that
in stable and predictable environments, the total number
of eggs will be reduced and that under unstable and/or del-
eterious conditions, reducing the number of eggs would be
catastrophic. According to recent observations and mod-
els, both strategies are used by arthropods, although larger
eggs seem more suitable in unstable settings (Fox &
Czesak 2000; Oloffson et al. 2009). Such information is
lacking for ostracods; however, reduction in the size of
these post-extinction ostracods seems incompatible with
larger eggs. Hence, I consider here the production of
smaller and more abundant eggs as more suitable in the
PTBM setting following the EPE.
The persistence of stable populations implies that mor-
tality and birth rates are balanced, reproductive patterns
being linked to mortality levels. Following the EPE in the
C€ur€uk Da�g section, mortality might have been higher than
normal, with two consequences: (i) higher reproductive
rates; and (ii) reduction of the time before females begin
to reproduce. Time to reproduction is further decreased
for podocopids by deceleration, resulting in a longer time
spent for the maturation of the juveniles and the shorten-
ing of adulthood duration. In these conditions, the conclu-
sion of smaller and more abundant eggs laid by PTBM
ostracods is reinforced: populations could not have been
stable with the conjunction of less abundant larger eggs
and reduced time available for reproduction. In this bet-
hedging strategy, the augmentation of the abundance of
eggs produced has never been quantified. If it is hypothe-
sized that production doubled in PTBM ostracods, the
fecundity of ostracod females should have been raised to
14–40 eggs per lifetime to maintain population viability
throughout PTBM deposition. This doubling of reproduc-
tion is fairly probable since it has been observed under
fluctuating temperatures for the Recent freshwater Heter-
ocypris virens (Rossi et al. 2013).
For freshwater ostracods, the hypothesis has been made
that more generations in a given period would allow faster
adaptation (Rossi et al. 2003, 2013; Forrest & Miller-
Rushing 2010). Together with the augmentation of the
number of eggs produced in the PTBM, increase in the
frequency of reproductive events during the adult stage is
also envisaged. Temperature, food availability and sea-
sonal time constraints regulate the duration of the life
cycle in copepods that can alternate between univoltine or
multivoltine strategies (Allan & Goulden 1980; Chen &
Folt 1996; Twombly et al. 1998; Gerten & Adrian 2002;
Winder & Schindler 2004; Adrian et al. 2009). Salinity,
temperature and day length have a great impact on freshwa-
ter ostracod reproduction, which can be reduced in low sal-
inities and doubled under fluctuating temperatures (Ganning
1971; Hagerman 1978; Otero et al. 1998; Rossi et al. 2013).
As a consequence of these factors, it is obvious that
Bairdioidea have greater adaptive potential than is tradi-
tionally considered. The faunas of the C€ur€uk Da�g section
cannot be taken as surviving in the PTBM refuge in a
straightforward and unique way. Instead they exemplify
two fundamentally different types of adaptation, poten-
tially opening new questions on the palaeobiology of
podocopids and palaeocopids. The study of ontogenetical
development of other species from other PTBM will pro-
vide additional analogues. Regarding the deceleration
observed in podocopid species, this can result in dwarf-
ism when it co-occurs with decoupled size and shape
change so that the descendant completes the same degree
and manner of shape change as the ancestor but in a nar-
rower size range (Alberch et al. 1979). In contrast, pera-
morphosis by deceleration could result in gigantism.
These phenomena could explain the overall reduction in
the size of ostracods after the EPE (Forel et al. in prog-
ress) as well as being consistent with observations that
some species become smaller and others larger in the
Heterochronic growth of ostracods (Crustacea) from microbial deposits 27
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

PTBM. It could also explain the anomalous size records
for Monoceratina hussonae and Callicythere postiangusta.
However, because no larval stage has been distinguished
for these two species, many questions are still pending: did
growth stop before terminal shape? And did they display a
very fast growth, or was it very slow? To answer these
questions, more data are required.
Ostracod assemblages
Diversity and abundance. Lower Triassic ostracods
from the C€ur€uk Da�g section are found at the base of the
Kokarkuyu Formation, which is of microbial origin, and
are very rare higher up. Species richness and abundance of
assemblages through the section can be divided into peaks
(P) and drops (D) (Fig. 24A). For clarity and because the
upper part of the section did not yield any ostracods, I
report only diversity variations up to sample CD31 (the
first 18 metres), in association with the different fabrics of
microbialites observed and described by Kershaw et al.
(2012). From the base to the top:
1. Assemblages TK51b to CD05 record increasing
richness and abundance (P1). Maximum abundance
is reached in CD05 with 1435 specimens. This inter-
val is mostly dominated by stromatolites and throm-
bolites at the top.
2. An important reduction in diversity and abundance
is observed from CD06 to CD08 (D1). This corre-
sponds to thrombolite at the base and hybrid micro-
bialites at the top.
3. Samples S2 to TK55b yielded rich and abundant
assemblages (P2), corresponding to thrombolites at
the base and hybrid microbialites at the top.
4. A diversity drop is recorded by assemblages CD12
to CD13 (D2). This event is correlated with a shift
to facies dominated by thrombolites.
5. This phase is followed by an important diversifica-
tion from CD14 to CD16 (P3). The maximum rich-
ness is recorded in S10, with 20 species.
6. Assemblages CD17 to CD25 record a long phase of
low richness and abundance (D3).
7. The interval from CD26 to CD29 records a slight
rediversification of assemblages characterized by
relatively higher richness and abundance (P4).
8. From TK125 onwards, samples are barren of ostra-
cods (D4). Only two low diversity and abundance
assemblages are found in CD51 and CD78.
Composition. The ostracods from the C€ur€uk Da�g section
belong to six superfamilies/families, the distributions of
which are shown in Figure 24B. The most common super-
family is Bairdioidea (Acratia, Bairdia, Bairdiacypris,
Bythocypris, Liuzhinia, Microcheilinella, Petasobairdia,
Praezabythocypris, Rectobairdia and Silenites). When
present, it comprises between 25% (CD03A, S1) and
100% of the assemblages (02TK51b, CD02, 02TK120,
123–126). The second most frequent family is Kirkbyidae
(Reviya), absent from specific peaks and drops (P3, D4
and P6). When present, Kirkbyidae varies between 5%
(S10) and 100% (02TK117, 122) of the recovered assemb-
lages. When present, Cytheroidea (Basslerella, Callicy-
there and Monoceratina) comprises between 6% (S4) and
100% (CD03B, 07) of the assemblages. Cypridoidea (gen-
era Macrocypris, Paracypris) are episodic components of
all diversity phases, making up between 5% (S10) and
17% of species (CD03E). Three superfamilies/families
have a unique occurrence in D3: (i) Kloedenelloidea
(genus Eukloedenella) – 50% of 08S6A, CD51, CD78
species; (ii) Cavellinidae (genus Sulcella) and (iii) Para-
parchitidae (Paraparchitidae indet.), each being 33% of
the 02TK118 assemblage.
Abundant small-sized specimens are found in some
assemblages, as in other PTBM deposits in South China
and Hungary (Forel 2012; Forel et al. 2013a). These small
specimens belong to the same genera as the larger forms:
the observed deviation of the size is not due to the pres-
ence of smaller genera per se. This aspect of ostracod fau-
nas associated with microbial deposits in the aftermath of
the EPE is the subject of a work in progress and will only
be discussed briefly below.
Palaeoenvironmental implications. In the present sec-
tion, three major units can be recognized based on both
sedimentology and composition of the ostracod faunas
(Table 1; Fig. 24B). The base of the section (up to �5 metres) is composed of stromatolites and hybrid micro-
bialites and is mostly dominated by Bairdioidea, Bythocy-
theridae and Cytherissinellidae. The middle portion of the
section (� 5 to 13.5 metres) records important facies and
faunal shifts: (i) facies shift to thrombolite deposits; and
(ii) faunal shift with the appearance and gradual increase
of Kirkbyidae. Important modifications also occur at the
top (from 13.5 to 18 metres): (i) transition from thrombo-
lites to oolites; and (ii) complete disappearance of Kirk-
byidae, together with the appearance of Cypridoidae and
the return of Cytherissinellidae. These patterns record the
forcing of environmental parameter(s) on the type of
deposits and composition of the faunas. Compared to
other PTBM (e.g. Forel 2009, 2012; Forel et al. 2013a),
faunas from the C€ur€uk Da�g section are atypical because
of the presence and abundance of Kirkbyidae, Cytherissi-
nellidae and Bythocytheridae. However, knowledge about
these last two families is still limited. Since they are key
elements of many assemblages recovered here, it would
be useful to give precise conclusions about parameters
forcing the observed patterns. A work based on the litera-
ture and new data is in progress to provide information on
Cytherissinellidae and Bythocytheridae that will allow the
28 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4
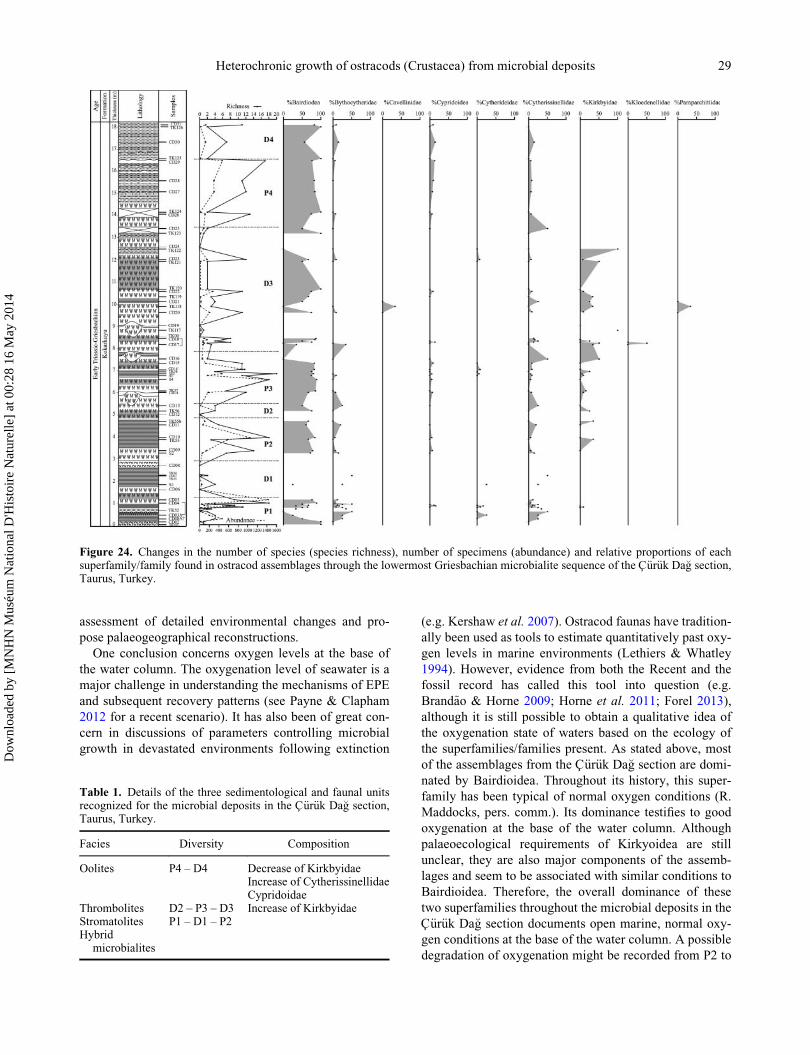
assessment of detailed environmental changes and pro-
pose palaeogeographical reconstructions.
One conclusion concerns oxygen levels at the base of
the water column. The oxygenation level of seawater is a
major challenge in understanding the mechanisms of EPE
and subsequent recovery patterns (see Payne & Clapham
2012 for a recent scenario). It has also been of great con-
cern in discussions of parameters controlling microbial
growth in devastated environments following extinction
(e.g. Kershaw et al. 2007). Ostracod faunas have tradition-
ally been used as tools to estimate quantitatively past oxy-
gen levels in marine environments (Lethiers & Whatley
1994). However, evidence from both the Recent and the
fossil record has called this tool into question (e.g.
Brand~ao & Horne 2009; Horne et al. 2011; Forel 2013),
although it is still possible to obtain a qualitative idea of
the oxygenation state of waters based on the ecology of
the superfamilies/families present. As stated above, most
of the assemblages from the C€ur€uk Da�g section are domi-
nated by Bairdioidea. Throughout its history, this super-
family has been typical of normal oxygen conditions (R.
Maddocks, pers. comm.). Its dominance testifies to good
oxygenation at the base of the water column. Although
palaeoecological requirements of Kirkyoidea are still
unclear, they are also major components of the assemb-
lages and seem to be associated with similar conditions to
Bairdioidea. Therefore, the overall dominance of these
two superfamilies throughout the microbial deposits in the
C€ur€uk Da�g section documents open marine, normal oxy-
gen conditions at the base of the water column. A possible
degradation of oxygenation might be recorded from P2 to
Table 1. Details of the three sedimentological and faunal unitsrecognized for the microbial deposits in the C€ur€uk Da�g section,Taurus, Turkey.
Facies Diversity Composition
Oolites P4 – D4 Decrease of KirkbyidaeIncrease of CytherissinellidaeCypridoidae
Thrombolites D2 – P3 – D3 Increase of KirkbyidaeStromatolites P1 – D1 – P2Hybrid
microbialites
Figure 24. Changes in the number of species (species richness), number of specimens (abundance) and relative proportions of eachsuperfamily/family found in ostracod assemblages through the lowermost Griesbachian microbialite sequence of the C€ur€uk Da�g section,Taurus, Turkey.
Heterochronic growth of ostracods (Crustacea) from microbial deposits 29
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

the top of the section, but it is still difficult to distinguish
the respective influence of oxygenation and water depth.
The growth of microbialites is not influenced by water
salinity (McNamara 2009). Despite the relative instability
of the environment recorded in the C€ur€uk Da�g section
through microbial development, the continuous presence
of stable Bairdioidea/Kirkbyoidea assemblages reveals
adaptation to environmental conditions in the area. The
impacts of the variations of salinity on ostracods are gen-
erally observed on the morphology of the valves, such as
the variation of nodation with salinity changes of Recent
Cyprideis torosa (e.g. Van Harten 1975; Boomer & Frenzel
2011). No thickening or nodation of the carapaces was
observed in the PTBM here, and the range of fluctuation of
salinity may therefore have been relatively narrow, never
reaching lethal levels for these types of ostracods. We
recently proposed a two-step oxygenation process to explain
the co-existence of low-oxygenation proxies and faunas of
relatively normal oxygenation setting (Forel et al. 2013b)
and identified microbial deposits following the EPE as ref-
uge areas. In the present section, ostracods are still present
in the 4 metres following the microbial deposits, which are
of oolitic facies. This pattern could be explained by the
non-fossilization of microbial mats within the oolites, still
providing a temporary shelter to ostracod faunas.
The heterochronies discussed above record the persis-
tence of a relatively degraded environment, in spite of the
refuge characteristics provided by the microbial ecosys-
tem. Although it is unclear whether they were the conse-
quences of environmental stress or adaptations to cope
with, they provided both individual and community abili-
ties to survive and maintain the viability of populations.
However, the nature of this stress cannot be solved in our
present state of knowledge of ostracod ecology: factors
influencing progeny size are climatic conditions, location
where eggs are laid, population densities, predation, com-
petition, starvation, dessication, oxygen stress, tempera-
ture stress, nutritional stress and environmental toxins
(Fox & Czesak 2000 and references therein).
Conclusions
1. Like in other localities yielding microbial deposits
in the earliest Griesbachian, ostracods are abundant
at the C€ur€uk Da�g section throughout microbialites
and become extinct shortly after the end of micro-
bial growth.
2. Fifty-seven species were found, amongst which
three are new: Monoceratina hussonae sp. nov.,
Reviya sylvieae sp. nov. and Eukloedenella adcapi-
tisdolorella sp. nov. Possible sexual dimorphism for
fossils of the genus Bairdia is reported for the first
time in the species Bairdia? kemerensis. This obser-
vation is the first record of the family
Kloedenellidae and the genus Eukloedenella in the
Early Triassic, showing that these taxa crossed the
Permian–Triassic boundary.
3. Heterochrony by paedomorphosis (deceleration) is
identified in six species: Bairdia? kemerensis, Liuz-
hinia antalyaensis, Praezabythocypris? ottomanen-
sis, Paracypris gaetanii, Reviya curukensis and
Reviya sylvieae. Peramorphosis (acceleration) is rec-
ognized in Eukloedenella adcapitisdolorella. These
patterns are interpreted as secondary survival strate-
gies in the aftermath of the end-Permian extinction.
4. Because of the overall dominance of Bairdioidea/
Kirkbyidae, oxygen conditions are interpreted as
normal at the base of the water column during the
growth of microbial mats at the C€ur€uk Da�g section.
Acknowledgements
This work is part of IGCP 572 ‘Restoration of marine eco-
systems following the Permian–Triassic mass extinction:
lessons for the present’ and Chinese programs NSFC
(40621002) and 111 (B08030) and was funded by Actions
transversales du museum (ATM) Biodiversit�es actuelles
et fossiles. It was undertaken at the UMR 7207 Centre de
recherche sur la pal�eobiodiversit�e et les pal�eoenvironne-ments (CR2P), Paris. I thank Prof. Dave Horne (Univer-
sity College, London, UK), Dr Gengo Tanaka (Japan
Agency for Marine-Earth Science and Technology, Japan)
and an anonymous reviewer for their comments and cor-
rections that greatly improved an earlier version of the
manuscript. I am also grateful to Prof. Horne for his valu-
able advice. My gratitude is addressed to Dr Sylvie
Crasquin (CNRS, Pierre et Marie Curie University, Paris,
France) for her advice, trust, and motherly support; it will
always be remembered. I am indebted to Prof. Steve
Kershaw (Brunel University, UK) and Dr Pierre-Yves
Collin (Burgundy University, France) for their construc-
tive remarks, and to Aymon Baud (Lausanne, Switzer-
land) and Erdal Kosun (Akdeniz University, Antalya,
Turkey) for their help in the field. I thank Martine Fordant
and Alexandre Lethiers (Pierre et Marie Curie Univer-
sity), respectively, for their help with the processing of
samples and the drawings. Finally I thank Brigitte L�etang(France) for patiently correcting my ‘frenglish’.
References
Adrian, R., O’Reilly, C. M., Zagarese, H., Baines, S. B.,Hessen, D. O., Keller, W., Livingstone, D. M., Sommar-uga, R., Straile, D., Van Donk, E., Weyhenmeyer, G. A.& Winderl, M. 2009. Lakes as sentinels of climate change.Limnology andOceanography, 54, 2283–2297.
30 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

Alberch, P., Gould, S. J., Oster, G. F. & Wake, D. B. 1979.Size and shape in ontogeny and phylogeny. Paleobiology, 5,296–317.
Allan J. D.&Goulden C.E. 1980. Some aspects of reproductivevariation among freshwater zooplankton. Pp. 388–410 in W.C. Kerfoot (ed.) Evolution and ecology of zooplankton com-munities. University Press of New England, Hanover.
Baird, W. 1845. Arrangement of the British Entomostraca, witha list of species, particularly noticing those which have asyet been discovered within the bounds of the club. Transac-tions of the Berwickshire Naturalists’ Club, 2, 145–158
Baird, W. 1850. The Natural History of the British Entomos-traca. Ray Society, London, 364 pp.
Bate, R. H., Lord, A. & Riegraf, W. 1979. Jurassic Ostracodafrom leg 79, Site 547. Pp. 703–710 in K. Hinz & E. L.Winterer (eds) Initial Reports of the Deep Sea DrillingProject. Washington.
Baud, A., Cirilli, S. & Marcoux, J. 1997. Biotic response tomass extinction: the Lowermost Triassic microbialites.Facies, 36, 238–242.
Baud, A., Richoz, S. & Marcoux, J. 2005. Calcimicrobial caprocks from the basal Triassic units: western Taurus occur-rences (SW Turkey). Comptes Rendus Palevol, 4, 569–582.
Benton, M. J. & Twitchett, R. J. 2003 How to kill (almost) alllife: the end-Permian extinction event. Trends in Ecologyand Evolution, 18(7), 358–365.
Bertholon, L. 1997. Les h�et�erochronies du d�eveloppement chezles ostracodes, indicateurs de strat�egies adaptatives. Geo-bios, 21, 277–285.
Bond, D. P. G. & Wignall, P. B. 2010. Pyrite framboid study ofmarine Permo-Triassic boundary sections: a complex anoxicevent and its relationship to contemporaneous mass extinc-tion. Geological Society of America Bulletin, 122, 1265–1279.
Boomer, I. & Frenzel, P. 2011. Possible environmental & bio-logical controls on carapace size in Cyprideis torosa (Jones,1850). Joannea – Geologie und Pal€aontologie, 11, 26–27.
Bulmer, M. 1994. Theoretical Evolutionary Ecology. SinauerAssociates, Sunderland, 352 pp.
Brand~ao, S. N. & Horne, D. J. 2009. The Platycopid Signal ofoxygen depletion in the ocean: a critical evaluation of theevidence from modern ostracod biology, ecology and depthdistribution. Palaeogeography, Palaeoclimatology, Palae-oecology, 283, 126–133.
Brooks, W. K. 1886. Report on the Stomatopoda dredged by H.M.S. “Challenger” during the years 1873–1876. Report onthe scientific results of the voyage of H.M.S. Challenger,Zoology, 16, 1–116.
Br€uwhiler, T., Brayard, A., Bucher, H. & Guodun, K. 2008.Griesbachian and Dienerian (Early Triassic) ammonoid fau-nas from Northwestern Guangxi and Southern Guizhou(South China). Palaeontology, 51, 1151–1180.
Chen, C. Y. & Folt C. L. 1996. Consequences of fall warmingfor zooplankton overwintering success. Limnology andOceanography, 41, 1077–1086.
Chen, L.,Wang, Y., Xie, S., Kershaw, S., Dong, M., Yang, H.,Liu, H.& Algeo, T. J. 2011. Molecular records of microbia-lites following the end-Permian mass extinction in Chon-gyang, Hubei Province, South China. Palaeogeography,Palaeoclimatology, Palaeoecology, 308, 151–159.
Cohen, A. C. & Morin, J. G. 1990. Patterns of reproduction inostracodes; a review. Journal of Crustacean Biology, 10,184–211.
Crasquin, S., Perri, M.C., Nicora, A. & De Wever, P. 2008.Ostracods across the Permian-Triassic boundary in Western
Tethys: the Bulla parastratotype (Southern Alps, Italy). RivistaItaliana di Palaeontologia e Stratigrafia, 114, 233–262.
Crasquin, S., Baud, A., Kershaw, S., Richoz, S., Kosun, E. &Forel, M. B. 2009. The Permian-Triassic transition in theSouthwestern Taurus Mountains (South Turkey). FieldGuide Book, IGCP 572 Annual Meeting & Field Workshopin southern Turkey, Antalya, September 2–6, 2009.
Crasquin, S. & Forel, M. B. 2013. Ostracods (Crustacea)through Permian-Triassic events. Earth-Science Reviews.doi: 10.1016/j.earscirev.2013.01.006
Crasquin-Soleau, S. & Gradinaru, E. 1996. Early Anisianostracode fauna from the Tulcea Unit (Cimmerian NorthDobrogean Orogen, Romania). Annales de Pal�eontologie,82(2), 59–116.
Crasquin-Soleau, S. & Kershaw, S. 2005. Ostracod fauna fromthe Permian-Triassic boundary interval of South China(Huaying Mountains, eastern Sichuan Province): palaeoen-vironmental significance. Palaeogeography, Palaeoclima-tology, Palaeoecology, 217, 131–141.
Crasquin-Soleau, S., Broutin, J., Besse, J. & Berthelin, M.2001. Ostracodes and paleobotany from the Middle Permianof Oman: implications on Pangea reconstruction. TerraNova, 13, 38–43.
Crasquin-Soleau, S., Richoz, S., Marcoux, J., Angiolini, L.,Nicora, A.& Baud, A. 2002. The events of the Permian-Tri-assic boundary: last survivors and/or first colonizers amongthe ostracods of the Taurides (southwestern Turkey). Comp-tes Rendus Geoscience, 334, 489–495.
Crasquin-Soleau, S., Marcoux, J., Angiolini, L. & Nicora, A.2004a. Palaeocopida (Ostracoda) across the Permian-Trias-sic events: new data from the South–Western Taurus (Tur-key). Journal of Micropalaeontology, 23, 67–76.
Crasquin-Soleau, S., Marcoux, J., Angiolini, L., Nicora, A. &Bertho, Y. 2004b. New ostracode fauna from Permian-Tri-assic boundary in Turkey (Taurus, Antalya Nappes). Micro-palaeontology, 50, 281–296.
Crasquin-Soleau, S., Vaslet, D. & Le Nindre, Y.M. 2005.Ostracods from Permian-Triassic boundary in Saudi Arabia(Khuff Formation). Palaeontology, 48, 853–868.
Crasquin-Soleau, S., Galfetti, T., Bucher, H. & Brayard, A.2006a. Early Triassic ostracods from Guangxi Province,South China. Rivista Italiana di Palaeontologia e Stratigra-fia, 112, 55–75.
Crasquin-Soleau, S., Vaslet, D. & Le Nindre, Y. M. 2006b.Ostracods of Permian-Triassic Khuff Formation, Saudi Ara-bia: paleoecology and paleobiogeography. GeoArabia, 11,55–76.
Danielopol, D. L., Baltan�as, A., Namiotko, T., Geiger, W.,Pichler, M., Reina, M. & Roidmayr, G. 2008. Developmen-tal trajectories in geographically separated populations ofnon-marine ostracods: morphometric applications for palaeoe-cological studies. Senckenbergiana lethaea, 88, 183–193.
De Deckker P. 1983. Notes on the ecology and distribution ofnonmarine ostracods in Australia. Hydrobiologia, 106, 223–234.
Erwin, D. H. 1993. The Great Paleozoic Crisis: Life and Deathin the Permian. Columbia University Press, New York,327 pp.
Ezaki, Y., Liu, J. B., Nagano, T. & Adachi, N. 2008. Geobio-logical aspects of the Earliest Triassic microbialites alongthe southern periphery of the tropical Yangtze Platform: Ini-tiation and cessation of a microbial regime. Palaios, 23,356–369.
Forel, M. B., Crasquin, S., Kershaw, S., Feng, Q. & Collin,P. Y. 2009. Ostracods (Crustacea) and water oxygenation in
Heterochronic growth of ostracods (Crustacea) from microbial deposits 31
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

earliest Triassic of South China: implications for oceanicevents of the end-Permian mass extinction. Australian Jour-nal of Earth Sciences, 56, 815–823.
Forel, M. B. & Crasquin, S. 2011. Lower Triassic ostracods(Crustacea) from Meishan section, Permian-Triassic GSSP(Zhejiang Province, South China). Journal of SystematicPalaeontology, 9, 455–466.
Forel, M. B. 2012. Ostracods (Crustacea) associated with micro-bialites across the Permian-Triassic boundary in Dajiang(Guizhou Province, South China). European Journal of Tax-onomy, 19, 1–34.
Forel, M. B. 2013. The Permian-Triassic mass extinction: ostra-cods (Crustacea) and microbialites. Comptes Rendus Geosci-ence, 345(4), 203–211.
Forel, M. B., Crasquin, S., Hips, K., Kershaw, S., Collin,P. Y. & Haas, J. 2013a. Ostracods (Crustacea) from Perm-ian-Triassic Boundary of B�alv�any North section (B€ukkMountains, Hungary). Acta Palaeontologica Polonica, 58,195–219.
Forel, M. B., Crasquin, S., Kershaw, S. & Collin, P. Y. 2013b.In the aftermath of the end-Permian extinction: the micro-bialite refuge? Terra Nova, 25, 137–143.
Forrest, J. & Miller-Rusching, A. J. 2010. Toward a syntheticunderstanding of the role of phenology in ecology and evo-lution. Philosophical Transactions of the Royal Society,Series B, 365, 3101–3112.
Fox, C. W. & Czesak, M. E. 2000. Evolutionary ecology ofprogeny size in arthropods. Annual Review of Entomology,45, 341–369.
Frisk, A., Bucher, H., Hautmann, M., Baud, A., Brosse, M. &Guodun, K. 2012. Unusually diverse mollusc assemblagesfrom the early Triassic (Griesbachian) of South China. Geolog-ical Society of America Abstracts with Program, 44(7), 166.
Ganning, B. 1971. On the ecology of Heterocypris salinus, H.incongruens and Cypridopsis aculeata (Crustacea, Ostra-coda) from Baltic brackish-water rockpools. MarineBiology, 8, 271–279.
Gerten, D. & Adrian, R. 2002. Species specific response offreshwater copepods to recent summer warming. FreshwaterBiology, 47, 2163–2173.
Gould, S. J. 1977. Ontogeny and phylogeny. Belknap Press ofHarvard University Press, Cambridge, Massachusetts, 501 pp.
Groves, J. R., Altiner, D. & Rettori, R. 2005. Extinction, sur-vival, and recovery of lagenide foraminifers in the Permian-Triassic boundary interval, central Taurides, Turkey. Pale-ontological Society Memoir, 62, 1–38.
Gr€undel, J. 1969. Neue taxonomische Einheiten der UnterklasseOstracoda (Crustacea). Neues Jahrbuch f€ur Geologie undPal€aontologie, Monatshefte, 1969, 353–361.
Guber, A. L. 1971. Problems of sexual dimorphism, populationstructure and taxonomy of the Ordovician genus Tetradella(Ostracoda). Journal of Paleontology, 45, 6–22.
Hagerman, L. 1978. The life-cycle of three species of algal-living ostracods from brackish water. Ophelia, 17,231–237.
Halbach, U. 1973. Life table data and population dynamics ofthe rotifer Brachionus calyciflorus Pallas as influenced byperiodically oscillating temperature. Pp. 217–288 in W.Wieser (ed.) Effect of temperature on ectothermic organ-isms. Springer, Heidelberg
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. 2001. PAST:Palaeontological Statistics software package for educationand data analysis. Palaeontologia Electronica, 4, 9.
Hammer, Ø. & Harper, D. A. T. 2005. Paleontological DataAnalysis. Blackwell, Oxford, 351 pp.
Hao, W. C. 1992. Lower Triassic marine ostracods from Guiz-hou. Acta Micropalaeontologica Sinica, 9, 37–44. [inChinese].
Harries, P. J., Kauffman, E. G. & Hansen, T. A. 1996. Modelsfor biotic survival following mass extinction. GeologicalSociety of London, Special Publication, 102, 41–60.
Hautmann, M., Bucher, H., Br€uhwiler, T., Goudemand, N.,Kaim, A. & N€utzel, A. 2011. An unusually diverse molluscfauna from the earliest Triassic of South China and its impli-cations for benthic recovery after the end-Permian biotic cri-sis. Geobios, 44, 71–85.
Heip, C. 1976. The life-cycle of Cyprideis torosa (Crustacea,Ostracoda). Oecologia, 24, 229–245.
Henningsmoen, G. 1953. Classification of Paleozoic Straight-Hinged Ostracods. Norsk Geologisk Tidsskrift, 31, 185–290.
Hessland, I. 1949. Investigations Lower Ordovician of the Siljandistrict. Sweden. I: lower Ordovocian ostracods of the Siljandistrict, Sweden. Bulletin of the Geological Institute ofUppsala, 33, 97–408.
Horne, D. J. 1983. Life-cycles of podocopid Ostracoda – Areview (with particular reference to marine and brackish-water species). Pp. 581–590 in R. F. Maddocks (ed.) Appli-cations of Ostracoda. Proceedings of the Eighth Interna-tional Symposium on Ostracoda. University of Houston,Geoscience, Houston, Texas.
Horne, D. J., Cohen, A. &Martens, K. 2002. Taxonomy, Mor-phology and Biology of Quaternary and living Ostracoda.Pp. 5–36 in J. A. Holmes & A. Chivas (eds) The Ostracoda:applications in Quaternary Research. Geophysical Mono-graph, American Geophysical Union, Washington DC.
Horne, D. J., Brand~ao, S. N. & Slipper, I. J. 2011. The Platy-copid Signal deciphered: responses of ostracod taxa to envi-ronmental change during the Cenomanian-TuronianBoundary Event (Late Cretaceous) in SE England. Palaeo-geography, Palaeoclimatology, Palaeoecology, 308, 304–312.
Jaanusson, V. 1956. Untersuchungen uber den oberordovizi-schen Lyckholm-Stufenkomplex in Estland. Bulletin of theGeological Institute of Uppsala, 36, 369–400.
Jones, T. R. & Sherborn, C. D. 1888. On some Ostracoda fromthe Fullers Earth Oolite and the Bradford Clay. Bath NaturalHistory and Antiquarian Field Club Proceedings, 6, 249–278.
Kaim, A., N€utzel, A., Bucher, H., Br€uwhiler, T. & Goude-mand, N. 2010. Early Triassic (Late Griesbachian) gastro-pods from South China (Shanggan, Guangxi). Swiss Journalof Geosciences, 103, 121–128.
Kashevarova, N. P. 1958. Novye vidy ostrakod verkhneperm-skikh otlozhenij (Ufimskiskh i Tatarskikh) YuzhnogoTimana i Volgo-Ural‘skoj Oblasti. Trudy Vsesoyuznogo Nef-tyanogo Nauchno-Issle-dovatel’skogo Instituta (VNIGRI),115, 302–339. [in Russian].
Kershaw, S., Li, Y., Crasquin-Soleau, S., Feng, Q., Mu, X.,Collin, P. Y., Reynolds, A. & Guo, L. 2007. Earliest Triassicmicrobialites in the South China block and other areas: con-trol on their growth and distribution. Facies, 53, 409–425.
Kershaw, S., Crasquin, S., Forel, M. B., Randon, C., Collin,P. Y., Kosun, E., Richoz, S. & Baud, A. 2010. Earliest Tri-assic microbialites in C€ur€uk Dag, southern Turkey; compo-sition, cyclicity and controls on formation. Sedimentology,58, 739–755.
Kershaw, S., Crasquin, S., Li Y., Collin, P. Y., Forel, M. B.,Mu, X., Baud, A., Wang, Y., Xie, S., Guo, L. & Maurer,F. 2012. Microbialites and global environmental changeacross the Permian-Triassic boundary: a synthesis. Geobiol-ogy, 10, 25–47.
32 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

Kesling, R. V. 1951a. The morphology of ostracod molt stages.Illinois Biological Monographs, 21(1–3), 117–126.
Kesling, R. V. 1951b. Terminology of ostracod carapaces. Con-tributions from the Museum of Paleontology University ofMichigan, 9(4), 93–171.
Kesling, R. V. 1952. Doubling in size of ostracod carapaces ineach moult stage. Journal of Paleontology, 26, 772–780.
Kesling, R. V. 1953. A slide rule for the determination of instarsin ostracod species. Contributions from the Museum of Pale-ontology, University of Michigan, 11, 97–109.
Kesling, R. V. & Takagi, R. S. 1961. Evaluation of Przibram’slaw for ostracods by use of the Zeuthen Cartesian-diverweighing technique. Contribution from the Museum of Pale-ontolog,y University of Michigan, 17, 1–58.
Kesling, R. V. & Crafts, F. C. 1962. Ontogenetic increase inArchimedean weight of the ostracod Chlamydotheca unispi-nosa (Baird). American Midland Naturalist, 68, 149–153.
Kozur, H. 1985. Neue ostracoden-Arten aus dem oberen Mittel-karbon (H€oheres Moskovian) Mittel- und Oberperm desB€ukk-Gebirges (N-Ungarn). Geologisch-Pal€aontologischeMitteilungen, 2, 1–145.
Latreille, P. A. 1806. Genera crustaceorum et insectorum: secun-dum ordinem naturalem in familias disposita, iconibus exem-plisque plurimis explicata. Tomus 1. Koenig, Paris, 301 pp.
Lethiers, F. 1978. Ostracodes du D�evonien terminal de la For-mation Big Valley, Saskatchewan et Alberta. Palaeontog-raphica, 162, 81–143.
Lethiers, F. 1981. Ostracodes du D�evonien terminal de l’Ouestdu Canada: Syst�ematique, biostratigraphie et pal�eo�ecologie.Geobios, 5, 1–234.
Lethiers, F. & Crasquin-Soleau, S. 1988. Comment extrairedes microfossiles �a tests calcitiques de roches calcairesdures. Revue de Micropal�eontologie, 31, 56–61.
Lethiers, F. & Whatley, R. 1994. The use of Ostracoda toreconstruct the oxygen levels of the Late Palaeozoic oceans.Marine Micropaleontology, 24, 57–69.
Liao, W., Wang, Y., Kershaw, S., Weng, Z. & Yang, H. 2010.Shallow-marine dysoxia across the Permian-Triassic bound-ary: evidence from pyrite framboids in the microbialite inSouth China. Sedimentary Geology, 232, 77–83.
Lika, K. & Kooijman, S. A. L. M. 2003. Life history implica-tions of allocation to growth versus reproduction in dynamicenergy budgets. Bulletin of Mathematical Biology, 65, 809–834.
Liu, H., Wang, Y. B., Yuan, A. H., Yang, H., Song, H. J. &Zhang, S. X. 2010. Ostracod fauna near the Permian-Trias-sic boundary in Chongyang, Hubei Province, and itsresponse to the mass extinction. Science in China, Earth Sci-ences, 53(6), 810–817.
Maddocks, R. F. & Iliffe, T. M. 1986. Podocopid Ostracoda ofBermuda caves. Stygologia, 2, 26–76.
Maddocks, R. F. 1990. Living and Fossil Macrocyprididae(Ostracoda). University of Kansas Paleontological Contri-butions, Monograph, 2, 1–410.
Maddocks, R. F. 1991. New Bairdiidae (Ostracoda) fromTulear, Madagascar. Journal of Micropalaeontology, 9,189–204.
Maddocks, R. F. 2013. New and poorly known species of Neon-esidea (Bairdiidae, Ostracoda, Crustacea) from French Frig-ate Shoals, the Hawaiian Islands. Zootaxa, 3608, 457–510.
Martins, M. J. F., Vandekerkhove, J. & Namiotko, T. 2008.Environmental stability and the distribution of the sexes:insights from life history experiments with the geographicparthenogen Eucypris virens (Crustacea: Ostracoda). Oikos,117, 829–836.
Martinsson, A. 1956. Neue Funde kambrischer G€ange undordovizischer Geschiebe im s€udwestlichen Finnland. Bulle-tin of the Geological Institutions of the University ofUppsala, 36, 79–105.
McCoy, F. 1844. A synopsis of the characters of the Carbonifer-ous Limestone fossils of Ireland. Dublin University Press,Dublin, 207 pp.
McKinney, M. L.&McNamara, K. J. 1991. Heterochrony: theevolution of ontogeny. Plenum, New York, 437 pp.
McNamara, K. 2009. Stromatolites. Geoscientist, 19, 16–22.Mesquita-Joanes, F., Smith, A. J.& Viehberg, F. A. 2012. The
ecology of Ostracoda across levels of biological organisationfrom individual to ecosystem: A review of recent develop-ments and future potential. Developments in Quaternary Sci-ences, 17, 15–35.
Mette, W. 2008. Upper Permian and lowermost Triassic stratig-raphy, facies and ostracods in NW Iran-implications for theP/T extinction event. Stratigraphy, 5, 205–219.
Mette, W. 2010. Ostracods from the Upper Permian and Perm-ian/Triassic boundary interval in Northwest Iran. RevistaEspa~nola de Micropaleontologia, 42(1), 11–35.
Mezquita F., Roca J. R. & Wansard G. 1999. Moulting, sur-vival and calcification: the effects of temperature and waterchemistry on an ostracod crustacean (Herpetocypris inter-media) under experimental conditions. Archiv f€ur Hydrobio-logie, 146, 219–238.
Moore, R. C. 1961. Treatise on Invertebrate Paleontology.Arthropoda 3, Crustacea, Ostracoda. Geological Society ofAmerica and University of Kansas Press, Boulder andLawrence, 442 pp.
Morales-Ram�ıres, A. & Jakob, J. 2008. Seasonal vertical dis-tribution, abundance, biomass and biometrical relationshipsof ostracods in Golfo Dulce, Pacific Coast of Costa Rica.Revista de Biolog�ıa Tropical, 56, 125–147.
M€uller, G. W. 1894. Die Ostracoden des Golfes von Neapel undder angrenzenden Meeres Abschnilte. Fauna und Flora Nea-pel, 21, 1–404.
Obato, M. 1999. Taxonomy and ecology of the genus Neonesi-dea distributed along the Japan sea coast. Unpublishedundergraduate thesis, Department of Earth Sciences, Kana-zawa University, Japan.
Oloffson, H., Ripa, J. & Jonzen, N. 2009. Bet-hedging as anevolutionary game: the trade-off between egg-size and num-ber. Proceedings of the Royal Society, Series B, 276, 2963–2969.
Orcutt, J. D. Jr & Porter, K. G. 1983. Diel migration and ther-mal fluctuation in Daphnia magna. Limnology and Ocean-ography, 28, 720–730.
Otero, M., Rossi, V., Baltan�as, A. & Menozzi, P. 1998. Effectof genotype and photoperiod on diapause strategies inEucypris virens (Jurine, 1820) (Crustacea: Ostracoda).Archiv f€ur Hydrobiologie Special Issues Advanced Limnol-ogy, 52, 229–236.
Payne, J. L. & Clapham, M. E. 2012. End-Permian MassExtinction in the Oceans: An Ancient Analog for theTwenty-First Century? Annual Review of Earth and Plane-tary Sciences, 40, 89–111.
Perrin, N. 1992. Optimal resource allocation and the marginalvalue of organs. American Naturalist, 139, 1344–1369.
Raff, R. A. & Kaufman, T. C. 1983. Embryos, genes, and evo-lution: The developmental-genetic basis of evolutionarychange. Macmillan, New York, 395 pp.
Reilly, S. M., Wiley, E. O. & Meinhardt, D. J. 1997. An inte-grative approach to heterochrony: the distinction between
Heterochronic growth of ostracods (Crustacea) from microbial deposits 33
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

interspecific and intraspecific phenomena. Biological Jour-nal of the Linnean Society, 60, 119–143.
Richoz, S. 2006. Stratigraphie et variations isotopiques du Car-bone dans le Permien sup�erieur et le Trias inf�erieur de laN�eot�ethys (Turquie, Oman et Iran). M�emoires de G�eologie(Lausanne), 46, 1–264.
Roff, D. A. 1992. The Evolution of Life Histories: Theory andAnalysis. Chapman and Hall, New York, 535 pp.
Rossi, V., Tondelli, B., Gandolfi, A., Bellavere, C.,McKenzie,K. G. & Menozzi, P. 2003. Heterocypris (Crustacea: Ostra-coda) from the Isole Pelagie (Sicily, Italy): population genet-ics. Italian Journal of Zoology, 70, 101–108.
Rossi, V., Martorella, A. & Menozzi, P. 2013. Hatching phe-nology and voltinism of Heterocypris barbara (Crustacea:Ostracoda) from Lampedusa (Sicily, Italy). Journal of Lim-nology, 72, 227–237.
Roth, R. 1928. Monoceratina: a new genus of Ostracoda fromthe Pennsylvanian of Oklahoma. Journal of Paleontology, 2,15–19.
Sars, G. O. 1866. Oversigt af marine Ostracoder. Norske Viden-skaps-Akademi, F€orhandlingar, 1865, 1–130.
Sars, G. O. 1887. Nye bidrag til kundskaben om middelhavetsinvertebrafauna: 4. Ostracods mediterranea (sydeuropaeiskeostracoder). Archiv for Mathematik og Naturvidenskab, 12,173–324.
Sars, G. O. 1910. Zoological results of the 3rd TanganyikaExpedition (1904–1905). Report on Ostracoda. Proceedingsof the Zoological Society of London, 48, 732–760.
Sars, G. O. 1922–1928. An account of the Crustacea of Norway.Vol. 9, Crustacea. Bergen Museum, 9, 1–277.
Sch€one, B. R., Oschmann, W., Tanabe, K., Dettman, D.,Fiebig, J., Houk, S. D. & Kanie, Y. 2004. Holocene seasonalenvironmental trends at Tokyo Bay, Japan, reconstructed frombivalve mollusk shells-implications for changes in the eastAsian monsoon and latitudinal shifts of the Polar Front.Quaternary Science Reviews, 23, 1137–1150.
Schornikov, E. I. 1990. Evolution and classification of Bythocy-theridae. Courier Forschungsinstitut Senckenberg, 123,291–302.
Scott, H. W. 1961. Suborder Beyrichicopina Scott n. suborder.Suborder Kloedenellocopina Scott n. suborder. Pp. Q. 111,180 in R. C. Moore (ed.) Treatise on Invertebrate Paleontol-ogy. Part Q. Arthropoda 3. Geological Society of America,and University of Kansas Press, Boulder and Kansas, 442 pp.
Sebe, O., Crasquin, S. & Gradinaru, E. 2013. Early and Mid-dle Anisian deep-water ostracods (Crustacea) from NorthDobrogea (Romania). Revue de Pal�eobiologie, 32, 509–529.
Sepkoski, J. J. Jr 1984. A kinetic model of Phanerozoic taxo-nomic diversity. III. Post-Paleozoic families and massextinctions. Paleobiology, 10, 246–267.
Shaver, R. 1953. Ontogeny and sexual dimorphism in Cyther-ella bullata. Journal of Paleontology, 27, 471–480.
Smith, R. J. & Kamiya, T. 2002. The ontogeny of Neonesideaoligodentata (Bairdioidea, Ostracoda, Crustacea). Hydrobio-logia, 489, 245–275.
Sohn, I. G. 1950. Growth series of ostracodes from the Permianof Texas. United States Geological Survey, ProfessionalPaper, 221-C, 33–43.
Sohn, I. G. 1961. Aechminella, Amphissites, Kirbyella andrelated genera. United States Geological Survey, Profes-sional Paper, 330-B, 107–160.
Sohn, I. G. 1970. Early Triassic Marine Ostracodes from theSalt Range and Surghar Range, West Pakistan. Pp. 149–206in B. Kummel & C. Teichert (eds) Stratigraphic boundaryproblems: Permian and Triassic of West Pakistan. Special
Publication, Department of Geology, University of Kansas,Lawrence.
Song, H., Tong, J., Chen, Z. Q., Yang, H. & Wang, Y. 2009.End-Permian mass extinction of foraminifers in the Nanpan-jiang basin, South China. Journal of Paleontology, 83, 718–738.
Stearns S. 1992. The evolution of life histories. Oxford Univer-sity Press, New York, 264 pp.
Styk, O. 1972. Some more important new species of foramini-fers and ostracods from the Triassic deposits of Poland.Kwartalnik Geologiczny, 16, 867–885.
Svensen, H., Planke, S, Polozov, A. G., Schmidbauer, N.,Corfu, F., Podladchikov, Y. Y. & Jamtveit, B. 2009. Sibe-rian gas venting and the end-Permian environmental crisis.Earth and Planetary Science Letters, 277, 490–500.
Szczechura, J. 1964. Monoceratina Roth (Ostracoda) from theUpper Cretaceous and Lower Paleocene of North andCentral Poland. Acta Palaeontologica Polonica, 9,357–406.
Tanaka, G. & Ikeya, N. 2002. Migration and speciation of theLoxoconcha japonica species group (Ostracoda) in EastAsia. Paleontological Research, 6, 265–284.
Tanaka, G., Ono, T., Nishimura, T. & Maeda, H. 2013. Mid-dle Permian ostracods from the Akasaka Limestone, GifuPrefecture, central Japan. Paleontological Research, 16,289–306.
Teissier, G. 1960. Reticulate growth. Pp. 541–544 in T. H.Waterman (ed.) The Physiology of Crustacea. AcademicPress, New York.
Thorp J. H. & Wineriter S. A. 1981. Stress and growthresponse of juvenile crayfish to rhythmic and arrhythmictemperature fluctuations. Archives of Environmental Con-tamination and Toxicology, 10, 69–77.
Triebel, E. & Bartenstein, H. 1938. Die Ostracoden des deut-schen Juras. Senckenbergiana, 20, 502–518.
Twombly, S., Clancy, N. & Burns, C. W. 1998. Life historyconsequences of food quality in the freshwater copepodBoeckella triarticulata. Ecology, 79, 1711–1724.
Ulrich, E. O. & Bassler, R. S. 1906. New American PaleozoicOstracoda. Notes and descriptions of the Upper Carbonifer-ous genera and species. United States National MuseumPaper, 30, 149–164.
Ulrich, E. O. & Bassler, R. S. 1908. New American PaleozoicOstracoda. preliminary revision of the Beyrichiidae, withdescriptions of new genera. Proceedings of the United StatesNational Museum, 35, 277–340.
Ulrich, E. O. & Bassler, R. S. 1923. Paleozoic Ostracoda: Theirmorphology, classification and occurrence. Maryland Geo-logical Survey, Silurian, 9, 271–391.
Van Harten, D. 1975. Size and environmental salinity in themodern euryhaline ostracod Cyprideis torosa (Jones, 1850),a biometrical study. Palaeogeography, Palaeoclimatology,Palaeoecology, 17, 35–48.
Wei, M. 1981. Early and Middle Triassic Ostracods from Sich-uan. Acta Palaeontologica Sinica, 20, 501–510. [in Chinesewith English abstract].
Wei, M., Li., Z., Jiang, Z. & Xie, L. 1983. Arthropoda. Pp. 7–253 in Chengdu Institute of Geological and Mineral Resour-ces (ed.). Palaeontological Atlas of Southwest China. Geolog-ical Publishing House, Beijing [in Chinese].
Williamson, W. C. 1848. On some of the microscopical objectsfound in the mud of Levant and other deposits; with remarkson the mode of formation of calcareous and infusorial sili-ceous rocks. Memoirs of the Literary and PhilosophicalSociety of Manchester, Series 2, 8, 1–128.
34 M. B. Forel
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4

Winder, M. & Schindler, D. E. 2004. Climatic effects on thephenology of lake processes. Global Change Biology, 10,1844–1856.
Whatley, R. C. & Stephens, M. 1977. Precocious sexualdimorphism in fossil and recent Ostracoda. Pp. 69–91 in H.L€offler & D. Danielopol (eds) Aspects of Ecology andZoogeography of Recent and Fossil Ostracoda. Junk, TheHague
Worner, S. P. 1992. Performance of phenological models undervariable temperature regimes: Consequences of the Kauf-mann or rate summation effect. Environmental Entomology,21, 689–699.
Yang, H., Chen, Z. Q., Wang, Y., Tong, J., Song, H. & Chen,J. 2011. Composition and structure of microbialite ecosys-tems following the end-Permian mass extinction in South
China. Palaeogeography, Palaeoclimatology, Palaeoecol-ogy, 308, 118–128.
Yao, Z., Xu, J., Zhang, Z. & Mo, Z. 1980. Biostratigraphy ofthe Late Permian and problems on the Permian-Triassicboundary in Western Guizhou and Eastern Yunnan. Pp. 1–69 in The coal-bearing strata and palaeontology of the LatePermian in Western Guizhou and Eastern Yunnan. SciencePress, Beijing. [in Chinese].
Ye, C. H., Gou, Y. X., Hou, Y. T. & Cao, M. Z. 1977. Meso-zoic and Cenozoic ostracode fauna from Yunnan. Pp. 153–330 in Mesozoic fossils from Yunnan, China. Volume 2,Beijing, Science Press [in Chinese].
Zheng, S. 1976. Early Mesozoic ostracodes from some localitiesin South West China. Acta Palaeontologica Sinica, 15, 77–93. [in Chinese with English abstract].
Heterochronic growth of ostracods (Crustacea) from microbial deposits 35
Dow
nloa
ded
by [
MN
HN
Mus
éum
Nat
iona
l D'H
isto
ire
Nat
urel
le]
at 0
0:28
16
May
201
4




















