
Diagenesis of fossil ostracods: Implications for stable isotope based palaeoenvironmental...
12
Diagenesis of fossil ostracods: Implications for stable isotope based palaeoenvironmental reconstruction Carys E. Bennett a, ⁎, Mark Williams a , Melanie J. Leng b , David J. Siveter a , Sarah J. Davies a , Hilary J. Sloane b , Ian P. Wilkinson c a Department of Geology, University of Leicester, University Road, Leicester, LE1 7RH, UK b NERC Isotope Geoscience Laboratories, British Geological Survey, Keyworth, Nottingham, NG12 5GG, UK c British Geological Survey, Keyworth, Nottingham, NG12 5GG, UK abstract article info Article history: Received 18 August 2010 Received in revised form 21 February 2011 Accepted 23 February 2011 Available online 21 March 2011 Keywords: Diagenesis Ostracod Ultrastructure Palaeoenvironment Stable isotopes Carboniferous A protocol for assessing the diagenetic alteration of ostracod carapaces for use in palaeoenvironmental reconstruction is proposed, based on comparison between fossil (Carboniferous) and recent specimens. Recent ostracods have a carapace ultrastructure of fine (b 0.5 μm diameter) crystals of primary low- magnesium calcite. The degree of recrystallisation in fossil ostracods is assessed based on the carapace ultrastructure and geochemistry. Six diagenetic stages are identified: 1, replacement of the original finely crystalline calcite carapace with neomorphic calcite; 2, framboidal and euhedral pyrite growth; 3, ferroan calcite carapace replacement and cavity (void) filling cement; 4, ferroan dolomite carapace replacement and void fills; 5, siderite growth; and 6, sphalerite and barite mineralisation. Stage 1 occurs during early shallow burial, stages 2–3 soon after, and stages 4–6 during much later burial (possibly hydrothermal) alteration. Ostracods and macrofossils exhibiting diagenetic stages 1–5 were analysed for oxygen and carbon isotope ratios. The oxygen isotope compositions clearly reflect the diagenetic stages. The most negative oxygen isotopes (δ 18 O -12.9‰) represent the precipitation of ferroan dolomite (Stage 4), and the most positive values of the neomorphic calcite (δ 18 O -0.9‰) represent Stage 1, and are within the predicted range of Carboniferous seawater (δ 18 O -4‰ to 0‰). The diagenetic protocol established here is specific to Palaeozoic fossil ostracods from carbonate and clastic sediments that were subject to shallow burial diagenesis and later hydrothermal alteration. Nevertheless, the approach could be used to assess the utility of fossil ostracods for palaeoenvironmental interpretation from a wide range of geological settings. © 2011 Elsevier B.V. All rights reserved. 1. Introduction Ostracod carapaces composed of pristine calcite have the potential to preserve geochemical and hence palaeoenvironmental information from the time of carapace formation. The ostracod carapace is composed of sub-micron crystals of low-magnesium calcite. During diagenesis the degree of recrystallisation of the carapace is linked to the lithology of the host sediment and their diagenetic history, such as burial depth and events such as volcanic intrusions. The ultrastructure of the calcified part of the carapace in Recent ostracods is well documented (Kornicker, 1969; Sylvester-Bradley and Benson, 1971; Bate and East, 1972; Okada, 1982; Sohn and Kornicker, 1988; Keyser, 1995; Yamada et al., 2005). However, only one study has compared the carapace ultrastructure of Recent specimens with fossil material (Dép che, 1982). Diagenetically altered specimens may have surfaces that are pitted, dissolved or recrystallised, although not all alteration can be identified by eye. Several techniques have been used to assess diagenesis in Recent ostracods. For example, a visual assessment of preservation under light microscopy (Dwyer et al., 2002; Keatings et al., 2002) and surface recrystallisation using scanning electron microscopy images (Mischke et al., 2008) are both routinely used for younger (Quaternary) material. If a pristine ostracod carapace is preserved, a wealth of palaeoenvironmental information can be obtained. For example, the oxygen isotope composition (coupled with other proxies) can provide information on global ice volume, salinity and temperature (Chivas et al., 1986a,b; Dwyer and Chandler, 2009). On a global scale the size of the polar ice caps significantly affects the δ 18 O composition of the oceans. For example during “icehouse” climatic periods, isotopically negative water is locked at the poles, resulting in more positive ocean δ 18 O. Glacial and interglacial episodes occurred during the late Mississippian (Lower Carboniferous) (Fielding et al., 2008). More positive oxygen isotope values measured from brachiopod calcite are interpreted as indicative of glacial conditions, in the early to mid Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161 ⁎ Corresponding author. E-mail addresses: [email protected] (C.E. Bennett), [email protected] (M. Williams), [email protected] (M.J. Leng), [email protected] (D.J. Siveter), [email protected] (S.J. Davies), [email protected] (H.J. Sloane), [email protected] (I.P. Wilkinson). 0031-0182/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2011.02.028 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Diagenesis of fossil ostracods: Implications for stable isotope based palaeoenvironmental...

Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
Diagenesis of fossil ostracods: Implications for stable isotope basedpalaeoenvironmental reconstruction
Carys E. Bennett a,⁎, Mark Williams a, Melanie J. Leng b, David J. Siveter a, Sarah J. Davies a,Hilary J. Sloane b, Ian P. Wilkinson c
a Department of Geology, University of Leicester, University Road, Leicester, LE1 7RH, UKb NERC Isotope Geoscience Laboratories, British Geological Survey, Keyworth, Nottingham, NG12 5GG, UKc British Geological Survey, Keyworth, Nottingham, NG12 5GG, UK
⁎ Corresponding author.E-mail addresses: [email protected] (C.E. Bennett), mr
[email protected] (M.J. Leng), [email protected] (D.J. [email protected] (H.J. Sloane), [email protected] (I.P. Wilkins
0031-0182/$ – see front matter © 2011 Elsevier B.V. Adoi:10.1016/j.palaeo.2011.02.028
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 August 2010Received in revised form 21 February 2011Accepted 23 February 2011Available online 21 March 2011
Keywords:DiagenesisOstracodUltrastructurePalaeoenvironmentStable isotopesCarboniferous
A protocol for assessing the diagenetic alteration of ostracod carapaces for use in palaeoenvironmentalreconstruction is proposed, based on comparison between fossil (Carboniferous) and recent specimens.Recent ostracods have a carapace ultrastructure of fine (b0.5 μm diameter) crystals of primary low-magnesium calcite. The degree of recrystallisation in fossil ostracods is assessed based on the carapaceultrastructure and geochemistry. Six diagenetic stages are identified: 1, replacement of the original finelycrystalline calcite carapace with neomorphic calcite; 2, framboidal and euhedral pyrite growth; 3, ferroancalcite carapace replacement and cavity (void) filling cement; 4, ferroan dolomite carapace replacement andvoid fills; 5, siderite growth; and 6, sphalerite and barite mineralisation. Stage 1 occurs during early shallowburial, stages 2–3 soon after, and stages 4–6 during much later burial (possibly hydrothermal) alteration.Ostracods and macrofossils exhibiting diagenetic stages 1–5 were analysed for oxygen and carbon isotoperatios. The oxygen isotope compositions clearly reflect the diagenetic stages. The most negative oxygenisotopes (δ18O −12.9‰) represent the precipitation of ferroan dolomite (Stage 4), and the most positivevalues of the neomorphic calcite (δ18O −0.9‰) represent Stage 1, and are within the predicted range ofCarboniferous seawater (δ18O−4‰ to 0‰). The diagenetic protocol established here is specific to Palaeozoicfossil ostracods from carbonate and clastic sediments that were subject to shallow burial diagenesis and laterhydrothermal alteration. Nevertheless, the approach could be used to assess the utility of fossil ostracods forpalaeoenvironmental interpretation from a wide range of geological settings.
[email protected] (M. Williams),r), [email protected] (S.J. Davies),on).
ll rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Ostracod carapaces composed of pristine calcite have the potentialto preserve geochemical and hence palaeoenvironmental informationfrom the time of carapace formation. The ostracod carapace iscomposed of sub-micron crystals of low-magnesium calcite. Duringdiagenesis the degree of recrystallisation of the carapace is linked tothe lithology of the host sediment and their diagenetic history, such asburial depth and events such as volcanic intrusions. The ultrastructureof the calcified part of the carapace in Recent ostracods is welldocumented (Kornicker, 1969; Sylvester-Bradley and Benson, 1971;Bate and East, 1972; Okada, 1982; Sohn and Kornicker, 1988; Keyser,1995; Yamada et al., 2005). However, only one study has comparedthe carapace ultrastructure of Recent specimens with fossil material(Dép che, 1982).
Diagenetically altered specimens may have surfaces that arepitted, dissolved or recrystallised, although not all alteration can beidentified by eye. Several techniques have been used to assessdiagenesis in Recent ostracods. For example, a visual assessment ofpreservation under light microscopy (Dwyer et al., 2002; Keatingset al., 2002) and surface recrystallisation using scanning electronmicroscopy images (Mischke et al., 2008) are both routinely used foryounger (Quaternary) material.
If a pristine ostracod carapace is preserved, a wealth ofpalaeoenvironmental information can be obtained. For example, theoxygen isotope composition (coupledwith other proxies) can provideinformation on global ice volume, salinity and temperature (Chivaset al., 1986a,b; Dwyer and Chandler, 2009). On a global scale the sizeof the polar ice caps significantly affects the δ18O composition of theoceans. For example during “icehouse” climatic periods, isotopicallynegative water is locked at the poles, resulting in more positive oceanδ18O. Glacial and interglacial episodes occurred during the lateMississippian (Lower Carboniferous) (Fielding et al., 2008). Morepositive oxygen isotope values measured from brachiopod calcite areinterpreted as indicative of glacial conditions, in the early to mid

151C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
Mississippian, whilemore negative δ18O are interpreted as interglacialconditions, in the latest Mississippian (Mii et al., 1999). On localscales, for example in restricted marine basins, the ocean waters mayevaporate and become more saline, 16O is preferentially lost and thusδ18Oseawater increases. Temperature affects the fractionation of oxygenisotopes into carbonate, with a decrease in δ18Ocarbonate correspondingto an increase in temperature of about −0.2‰/°C (Chivas et al.,1986a).
Studies on living lacustrine ostracods have shown that biologicaleffects can also be important to δ18O values. Vital effects (nonequilibrium fractionation based on experimental data) can be morethan 3‰ for δ18O in some ostracod species (von Grafenstein et al.,1999; Griffiths and Holmes, 2000; Decrouy, 2009). Isotopic vitaleffects and Sr and Mg partition coefficients are variable betweenspecies (von Grafenstein et al., 1999), and even within individuals ofthe same species (Wansard et al., 1998). The concentration of traceamounts of Sr and Mg in the carapaces is controlled by temperature,salinity and the Mg/Ca ratio of the host water (Chivas et al., 1986a,b;Wansard et al., 1998; De Deckker et al., 1999; Dwyer et al., 2002). TheMg/Ca ratio of ostracod carapace calcite has been successfully used asa palaeotemperature proxy (Chivas et al., 1986a,b; Dwyer et al., 1995;Cronin et al., 2003, 2005; Dwyer and Chandler, 2009).
The effect of diagenetic overprinting on low-magnesium calcitebrachiopod shells has beenwell documented. Diagenesis tends to leadto lower carbon and oxygen isotope values (Banner and Kaufman,1994; Mii et al., 2001), although the types of fluid responsible fordiagenetic alteration result in different isotope compositions. InCarboniferous sediments, diagenetic alteration by later meteoricwater tends to lead to lower carbon and oxygen isotope compositionscompared to alteration by seawater (Immenhauser et al., 2002). Theburial depth of the sediment is a critical factor during diagenesis, asincreasing temperature with depth can lead to lower δ18O. This hasbeen shown in studies of benthic foraminifera and bulk carbonatefrom multiple Cenozoic ocean drill cores, where diagenesis leads tolower oxygen isotope composition with increasing depth (Schrag etal., 1995).
Previous stable isotope studies on Palaeozoic ostracods are rarebut include material from the Welsh Ordovician (Brenchley et al.,2003), where diagenetic alteration was assumed to be minimal, and astudy of specimens from the Carboniferous Ballagan Formation ofScotland (Williams et al., 2006). In the study of the BallaganFormation material the oxygen isotope compositions of the ostracodswere combined with associated palaeontological and sedimentolog-ical data, to support a hypothesis of brackish water conditions, andthus identify a step in the colonisation of non-marine environmentsby ostracods (Williams et al., 2006).
Here we succeed the study of Williams et al. (2006) by comparingthe ultrastructure, mineralogy and petrology of a range of Recent andCarboniferous ostracods as a means of assessing preservationcondition prior to palaeoenvironmental interpretation. We alsocompare the alteration of the ostracods to the alteration of macro-
Table 1Locality information for Recent podocopid ostracods studied. Specimens were preserved inethanol. National Grid Reference locations are given in brackets for British specimens.
Recent ostracod species Locali
Aurila woutersi Horne, 1986 PorlocBradleystrandesia fuscata (Jurine, 1820) A shalCandona candida (Müller, 1776) The riCandonopsis tenuis (Brady, 1886) Lake BCyprideis torosa (Jones, 1850) A saltCythere lutea Müller, 1875 A saltwEucypris virens (Jurine, 1820) A shalFabaeformiscandona japonica (Okubo, 1990) Lake BHemicythere villosa (Sars, 1866) Blue APseudocandona sucki (Hartwig, 1901) The ri
fossils found in the same succession. The six diagenetic stagesdescribed here are based on crystal texture, structure andmineralogy.We hypothesise that for the Carboniferous Ballagan Formation andStrathclyde Group fossils, the dominant signal preserved in the stableisotope record is that of diagenesis, leaving little palaeoenvironmentalinformation. However, some material which preserves early shallowburial alteration only may contain a remnant environmental signalsince the pore water composition and temperature were probablysimilar in the shallow sediments and the overlying water body.
2. Geological background and collection localities
Living Podocopida ostracods were collected in vivo from a range ofmarine and non-marine environments (Table 1). Approximately 60Recent specimens representing 10 species were analysed in order tocompare the ultrastructure and preservation of the carapace to that ofCarboniferous ostracods. The Recent specimens were not subject toisotopic analysis.
Mississippian sediments of the Midland Valley of Scotlandrepresent deposition in a range of environments from marine tonon-marine, and contain a high diversity of ostracods and macro-fossils (Forsyth and Chisholm, 1977; Wilson, 1989; Browne et al.,1999; Williams et al., 2006; Bennett, 2008; Bennett et al., in press).However, as a result of numerous Carboniferous igneous intrusionsduring the Pennsylvanian (Dean and Turner, 1995), hydrothermalalteration has significantly affected the Midland Valley sediments andfossil preservation. In particular, many of the limestones of theStrathclyde Group are dolomitised (Searl and Fallick, 1990; Searl,1991). In addition to hydrothermal alteration, a secondary re-mobilisation of carbonate in pore fluids and the dissolution of fossilshells may have taken place at the time of maximum sediment burial,at the end Triassic, when there was a 26 km overburden (Dean andTurner, 1995).
Carboniferous fossil ostracods were collected and studied fromMidland Valley boreholes and field sections (Fig. 1A). During theMississippian theMidland Valleywas situated on the southern edge ofLaurussia, on the margins of a restricted marine seaway (Fig. 1B).Samples were obtained from the Ballagan Formation, InverclydeGroup of Ayrshire and the Anstruther to Pathhead formations, of theStrathclyde Group of Fife. Hundreds of ostracod specimens wereanalysed from 20 Carboniferous species (Appendix 1), in order tocompare their preservation with Recent ostracods and to assess anyisotopic differences in carapaces from palaeoenvironments rangingfrom marine to non-marine. A range of macrofossils were sampled inthin section (Appendix 2). The following macrofossils were sampledfor isotopic analysis: Productus brachiopod, Leiptera bivalve, anindeterminate bivalve and Lithostrotion junceum coral. The age ofthe sediments is established on geochronological studies of volcanicrocks and palynomorph biostratigraphy (Stephenson et al., 2004;Monaghan and Parrish, 2005; Owens et al., 2005). The dominantpalaeoenvironmental signature of the formations studied is detailed
ethanol, then picked and stored dry in microfossil slides, or in test tubes filled with
ty information
k, Severn Estuary; donated by Dr David J Horne (Queen Mary University of London)low temporary pond, Flatford Mill, Suffolk (TM 077 332)ver Stour, Flatford Mill, Suffolk (TM 081 327)iwa, Japan; donated by Dr Robin J Smith (Lake Biwa Museum)marsh at Pegwell Bay, Isle of Thanet, Kent (TR 344 635)ater rock pool, Foreness Point, Kent (TR 384 716)
low temporary pond, Flatford Mill, Suffolk (TM 077 332)iwa, Japan; donated by Dr Smithnchor, Severn Estuary; donated by Dr Hornever Stour, Flatford Mill, Suffolk (TM 081 327)

Fig. 1. A:Mississippian sections studied in theMidland Valley of Scotland. Borehole material (1964–1980 drilling) and some field section samples are housed at the BGS in Edinburgh.Field samples were collected (CEB) between 2005 and 2008. B: Mississippian palaeogeography (c.340 Ma) of the Midland Valley of Scotland (modified from Prof. Blakely's websitehttp://jan.ucc.nau.edu/~rcb7).
152 C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
in Fig. 2. The sampled sedimentary rocks are identified as non-marine,marine or marginal marine, based on their faunal assemblages ofostracods and macrofossils (Wilson, 1989; Bennett, 2008; Bennettet al., in press).
3. Methodology
In order to assess diagenesis three techniques were used: 1, visualanalysis of Recent and fossil ultrastructure using scanning electronmicroscopy (SEM), using a range of magnification (×60 to ×3200); 2,visual analysis of diagenetic cements and mineralisation of polishedthin sections of fossils under plane polarised light (PPL), SEM andcathodoluminescence (CL); and 3, geochemical analysis of fossilsusing SEM and the electron microprobe. Stage 1 only applies toostracods, while stages 2 and 3 also apply to macrofossils.
The fossil ostracods were removed from the sediment with aneedle, which avoids contamination from other carbonate compo-nents (Keatings et al., 2006; Mischke et al., 2008), and mounted onstubs. Carapaces and polished thin sections were analysed under highand partial vacuum using a Hitachi S-3600N SEM at the University ofLeicester. Carapaces were also embedded in araldite and sectioned at160 μm and 100 μm thicknesses, in order to reveal different areas ofthe carapace. Polished thin sections of the sedimentary rock were cut30 μm thick and were selected to encompass a range of lithologies,environments and faunal content (Appendix 2). Twenty four thinsections of Carboniferous ostracods were analysed and compared toone thin section containing nine species of Recent ostracodsembedded in araldite. A back-scattered electron (BSE) detector wasused for identifying changes in composition, the element mapping ofcarbon coated sections (as in Martill et al., 1992) and for producingsemi-quantitative compositional data. Over 250 compositions of arange of diagenetic products were analysed using SEM and theelectron microprobe.
CL analysis used an optical microscope, cold cathode vacuumsetup, with a 15 kV accelerating voltage and 300–400 μm beamcurrent, at the University of Liverpool. The electron microprobe (JEOLJXA-8600S) was used with an accelerating voltage of 15 kV, probecurrent of 30 nA and a beam diameter of 15 μm. The quantitativeresults were calibrated to standards used for carbonate analysis, withan analytical error of 0.1 wt.%.
Ostracod carapaces were analysed for oxygen and carbon isotopesat the NERC Isotope Geosciences Laboratory. A weight of 40–100 μgwas required for each analysis and, where the specimens were largeenough, this comprised one single ostracod valve. Only single valves
of supposed adult ostracods were analysed, because closed carapacesmay enclose sediment or diagenetic contaminants and juveniles canbe magnesium-enriched (Chivas et al., 1986b). The analyses wereperformed on a GV IsoPrime dual inlet mass spectrometer, withmultiprep device set at 90 °C and a 15 min reaction time. The sampleswere run alongside both calcite (KCM) and dolomite (TDS) standards.δ18O was calculated from both calcite or dolomite (assuming singlemineralogy), using the fractionation factors of Rosenbaum andSheppard (1986). Isotope values are reported in the usual deltanotation, as per mille values, on the VPDB scale. Replicate analysis ofthe standards gave reproducibility around 0.1‰ for both carbon andoxygen.
4. Results
4.1. The preservation of Recent ostracods
The calcified part of Recent ostracod carapaces, the outer lamellacuticle, or procuticle, is between 20 and 45 μm thick (Sylvester-Bradley and Benson, 1971). The procuticle is covered by a continuouscuticular integument, composed of a thin (sub-μm thick) chitinousand organic membrane. The outer layer of this integument is theepicuticle, through which pores and sensory setae emerge (Fig. 3A).The procuticle crystals are tabulate to elongate, less than 0.5 μm longand aligned parallel to the margins of the carapace (Fig. 3B, C). TheRecent ostracod carapaces are non-luminescent under CL and arecomposed of low-magnesium calcite. Studies of Recent ostracods alsoshow a procuticle with sub-μm calcite crystals, and a foliated orlaminated structure (Sylvester-Bradley and Benson, 1971; Bate andEast, 1972; Okada, 1982; Sohn and Kornicker, 1988; Keyser, 1995).The epicuticle decays when the carapace is fossilised (Dép che, 1982).
4.2. Diagenetic stages of Carboniferous ostracods
Diagenetic minerals occur replacing whole ostracod carapaces andmacrofossil shells, in fossil cavities, and as crystals in the host sediment.Diagenetic assessment has revealed six diagenetic stages, from themost“pristine” material (Stage 1) to the most altered (Stage 6). The bestpreserved specimens comprise neomorphic calcite. Beyond the neo-morphic calcite there is a range of diagenetic cements and minerals;pyrite, ferroan calcite, ferroan dolomite, siderite, barite and sphalerite(Fig. 4). The occurrence of these diagenetic minerals and cements inostracods, macrofossils and in voids (carapace fills and sediment porespaces) in thin sections are given in Appendix 2. In the limited number

Fig. 2. Stratigraphy and palaeoenvironments of Mississippian sections studied. Thestratigraphy and palaeoenvironmental information are based on Browne et al. (1999),Bennett (2008) and Bennett et al. (in press). The location of the borehole and fieldsections is shown in Fig. 1.
153C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
of diagenetically assessed thin sections there is no correlation betweenthe sediment type, stratigraphic age of the material, fossil group,ostracod species, or thediagenetic stage (Appendix2).However, there isan association between certain diagenetic stages of 1) pyritewith calciteand 2) siderite with ferroan dolomite. Dissolution, pitting anddegradation of ostracod carapaces occur throughout the diagenetichistory. The timing of these six stages has been distinguished relative toone another using petrographic evidence such as cross-cutting relation-ships (Fig. 4).
4.2.1. Stage 1: neomorphic calciteMany of the Carboniferous ostracod specimens are superficially
well preserved, with apparently pristine composition of low-magne-sium calcite and with fine original surface ornamentation. However,the carapace ultrastructure of the Carboniferous ostracods has beenaltered compared to that of Recent ostracods.
The carapaces of the Carboniferous specimens are composed of acrystalline ultrastructure in the form of euhedral low-magnesiumcalcite crystals, 1–3 μm in diameter (Fig. 3D, E). The ultrastructure isdistinguished from an overgrowth texture as it incorporates surfacefeatures such as pores, spines and tubercles (Fig. 3D). The calcite issimilar to the neomorphic recrystallisation product described bySexton et al. (2006). Sub-micron size biogenic calcite has beenreplaced by inorganic calcite crystals of a larger size. Calcite crystalsare interlocking, with clear faces; growth lines and ridges areobserved (Fig. 3E). In some specimens the crystal edges are smoothand there are gaps between the crystals due to partial dissolution(Fig. 3F). Degradation or pitting on an ultrastructural level is also fairlycommon (Fig. 3G). This ultrastructure extends throughout theprocuticle, with larger crystals present on the outer margins (1–3 μm diameter) compared to those in the interior (1–2 μm diameter)(Fig. 3I). There is no apparent relationship between the size of thecrystals of the procuticle and the age of the sample, the representedspecies, the size of the carapace, or the host lithology (Appendix 3).
The crystal alignment in cross-section through the procuticleappears random in some specimens (Fig. 3I). However, to some extentthe neomorphic calcite preserves the original multi-laminate procu-ticle structure (Fig. 3H, J, K), as seen in 12 of the 16 ostracod-bearingpolished thin sections, when viewed under PPL, CL or SEM. Laminaeare parallel to the carapace outer margins, densely packed together(between 5 and 9 laminae per 10 μm carapace thickness), and appearbrightly luminescent under CL. Elemental mapping of the multi-laminate carapace structure under SEM does not reveal anycompositional differences between the laminae. Carapace laminaewere also analysed under the electron microprobe to elucidate anytrace compositional differences. A specimen with clear multi-laminae(Fig. 3K) did not have any significant chemical variation across thelaminae (Table 2): trace amounts of magnesium, iron and manganesepresent are not confined by or related to the laminae. Anotherspecimen, which does not have clear multi-laminae (Fig. 3L) has aninnermost procuticle layer that luminesces brightly under CL. Electronmicroprobe analysis shows that this inner region contained thehighest concentrations of iron and magnesium, with a maximum of0.8 wt.% MgO and 1.2 wt.% FeO (Table 2), but with trace amounts ofmanganese only.
4.2.2. Stage 2Euhedral pyrite grows in the position of dissolved ostracod
carapaces and macrofossil shells, in voids (Fig. 4, Stage 2), and isalso commonly disseminated within the sediment. Framboidal pyriteis most commonly associated with marine sediments. In one marineostracod specimen (Fig. 4, Stages 1–3), framboids on the innercarapace margin are overprinted by a ferroan calcite carapace fillingcement (Stage 3). Both euhedral and framboidal pyrite are predom-inantly found in association with Stage 1 calcitic ostracods, ratherthan those which have undergone further diagenetic alteration(Appendix 2).
4.2.3. Stage 3Ferroan calcite has sub-micron size crystals and does not preserve
the Stage 1 crystalline calcitic structure. Despite this, a remnant of theoriginal ostracod multi-laminar procuticle structure remains untilStage 5. Ferroan calcite carapace replacement is fairly rare (Appen-dix 2), and is often associated with ferroan dolomite (Stage 4) void-fills. As mentioned above, ferroan calcite void-fills overprint pyriteminerals (Fig. 4, Stage 3).
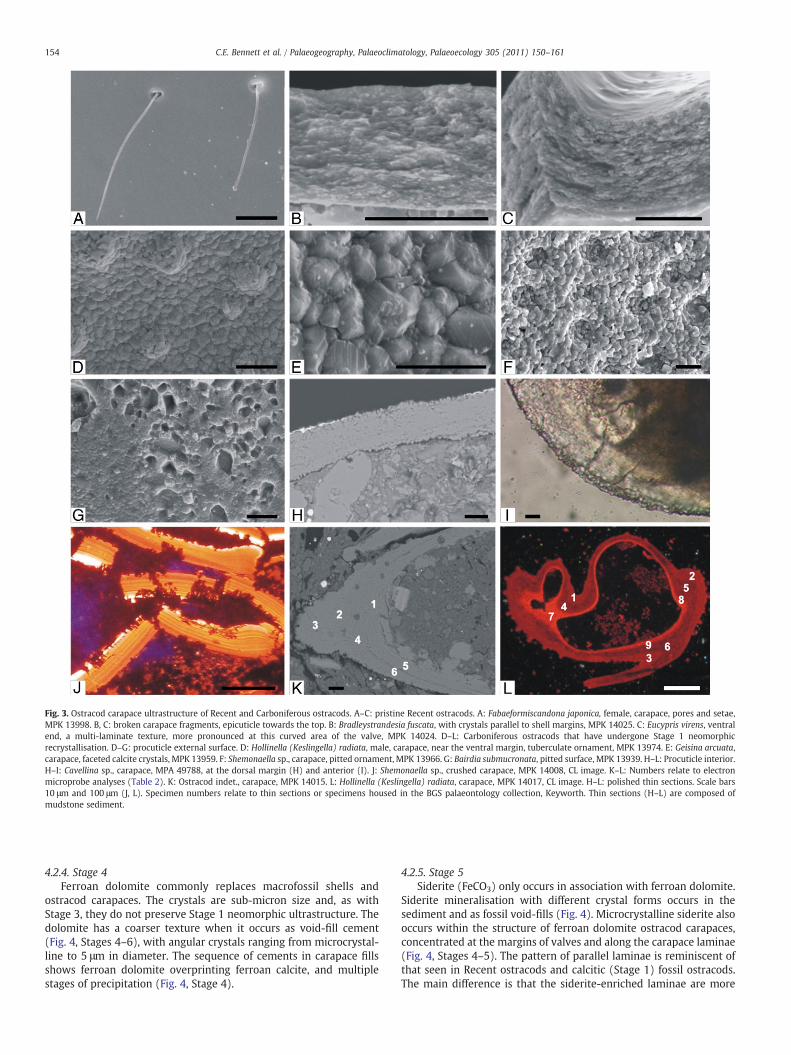
Fig. 3. Ostracod carapace ultrastructure of Recent and Carboniferous ostracods. A–C: pristine Recent ostracods. A: Fabaeformiscandona japonica, female, carapace, pores and setae,MPK 13998. B, C: broken carapace fragments, epicuticle towards the top. B: Bradleystrandesia fuscata, with crystals parallel to shell margins, MPK 14025. C: Eucypris virens, ventralend, a multi-laminate texture, more pronounced at this curved area of the valve, MPK 14024. D–L: Carboniferous ostracods that have undergone Stage 1 neomorphicrecrystallisation. D–G: procuticle external surface. D: Hollinella (Keslingella) radiata, male, carapace, near the ventral margin, tuberculate ornament, MPK 13974. E: Geisina arcuata,carapace, faceted calcite crystals, MPK 13959. F: Shemonaella sp., carapace, pitted ornament, MPK 13966. G: Bairdia submucronata, pitted surface, MPK 13939. H–L: Procuticle interior.H–I: Cavellina sp., carapace, MPA 49788, at the dorsal margin (H) and anterior (I). J: Shemonaella sp., crushed carapace, MPK 14008, CL image. K–L: Numbers relate to electronmicroprobe analyses (Table 2). K: Ostracod indet., carapace, MPK 14015. L: Hollinella (Keslingella) radiata, carapace, MPK 14017, CL image. H–L: polished thin sections. Scale bars10 μm and 100 μm (J, L). Specimen numbers relate to thin sections or specimens housed in the BGS palaeontology collection, Keyworth. Thin sections (H–L) are composed ofmudstone sediment.
154 C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
4.2.4. Stage 4Ferroan dolomite commonly replaces macrofossil shells and
ostracod carapaces. The crystals are sub-micron size and, as withStage 3, they do not preserve Stage 1 neomorphic ultrastructure. Thedolomite has a coarser texture when it occurs as void-fill cement(Fig. 4, Stages 4–6), with angular crystals ranging from microcrystal-line to 5 μm in diameter. The sequence of cements in carapace fillsshows ferroan dolomite overprinting ferroan calcite, and multiplestages of precipitation (Fig. 4, Stage 4).
4.2.5. Stage 5Siderite (FeCO3) only occurs in association with ferroan dolomite.
Siderite mineralisation with different crystal forms occurs in thesediment and as fossil void-fills (Fig. 4). Microcrystalline siderite alsooccurs within the structure of ferroan dolomite ostracod carapaces,concentrated at the margins of valves and along the carapace laminae(Fig. 4, Stages 4–5). The pattern of parallel laminae is reminiscent ofthat seen in Recent ostracods and calcitic (Stage 1) fossil ostracods.The main difference is that the siderite-enriched laminae are more
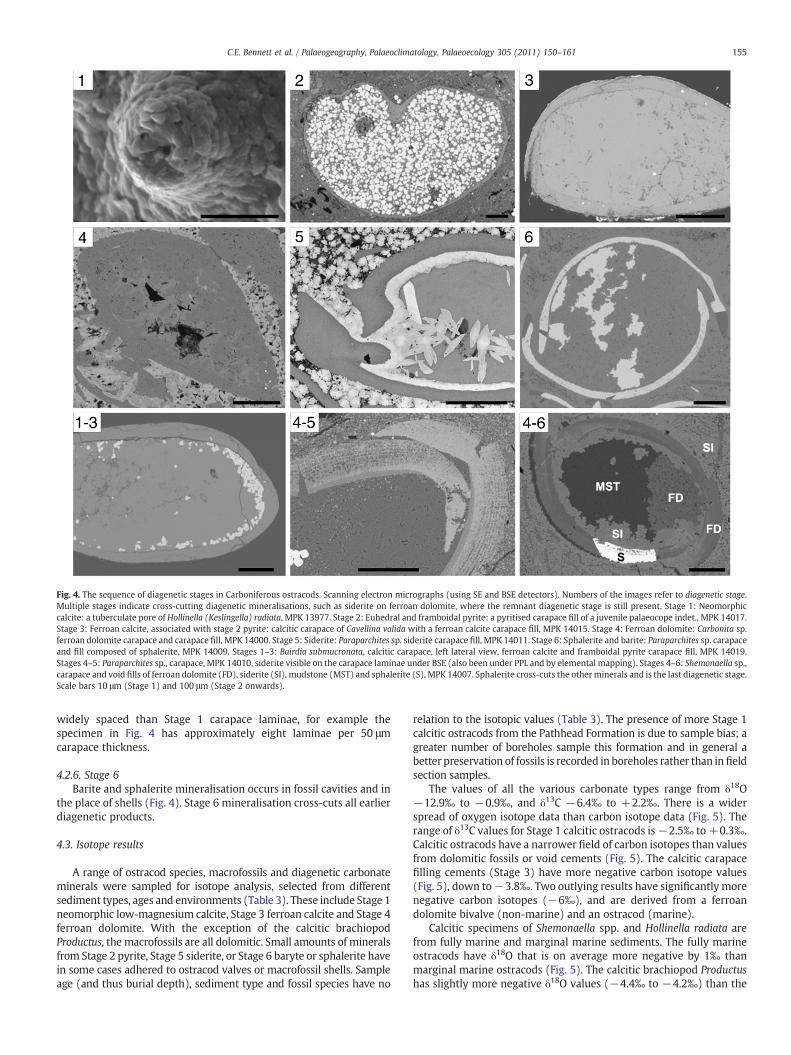
Fig. 4. The sequence of diagenetic stages in Carboniferous ostracods. Scanning electron micrographs (using SE and BSE detectors). Numbers of the images refer to diagenetic stage.Multiple stages indicate cross-cutting diagenetic mineralisations, such as siderite on ferroan dolomite, where the remnant diagenetic stage is still present. Stage 1: Neomorphiccalcite: a tuberculate pore of Hollinella (Keslingella) radiata, MPK 13977. Stage 2: Euhedral and framboidal pyrite: a pyritised carapace fill of a juvenile palaeocope indet., MPK 14017.Stage 3: Ferroan calcite, associated with stage 2 pyrite: calcitic carapace of Cavellina valida with a ferroan calcite carapace fill, MPK 14015. Stage 4: Ferroan dolomite: Carbonita sp.ferroan dolomite carapace and carapace fill, MPK 14000. Stage 5: Siderite: Paraparchites sp. siderite carapace fill, MPK 14011. Stage 6: Sphalerite and barite: Paraparchites sp. carapaceand fill composed of sphalerite, MPK 14009. Stages 1–3: Bairdia submucronata, calcitic carapace, left lateral view, ferroan calcite and framboidal pyrite carapace fill, MPK 14019.Stages 4–5: Paraparchites sp., carapace, MPK 14010, siderite visible on the carapace laminae under BSE (also been under PPL and by elemental mapping). Stages 4–6: Shemonaella sp.,carapace and void fills of ferroan dolomite (FD), siderite (SI), mudstone (MST) and sphalerite (S), MPK 14007. Sphalerite cross-cuts the otherminerals and is the last diagenetic stage.Scale bars 10 μm (Stage 1) and 100 μm (Stage 2 onwards).
155C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
widely spaced than Stage 1 carapace laminae, for example thespecimen in Fig. 4 has approximately eight laminae per 50 μmcarapace thickness.
4.2.6. Stage 6Barite and sphalerite mineralisation occurs in fossil cavities and in
the place of shells (Fig. 4). Stage 6 mineralisation cross-cuts all earlierdiagenetic products.
4.3. Isotope results
A range of ostracod species, macrofossils and diagenetic carbonateminerals were sampled for isotope analysis, selected from differentsediment types, ages and environments (Table 3). These include Stage 1neomorphic low-magnesium calcite, Stage 3 ferroan calcite and Stage 4ferroan dolomite. With the exception of the calcitic brachiopodProductus, the macrofossils are all dolomitic. Small amounts of mineralsfrom Stage 2 pyrite, Stage 5 siderite, or Stage 6 baryte or sphalerite havein some cases adhered to ostracod valves or macrofossil shells. Sampleage (and thus burial depth), sediment type and fossil species have no
relation to the isotopic values (Table 3). The presence of more Stage 1calcitic ostracods from the Pathhead Formation is due to sample bias; agreater number of boreholes sample this formation and in general abetter preservation of fossils is recorded in boreholes rather than in fieldsection samples.
The values of all the various carbonate types range from δ18O−12.9‰ to −0.9‰, and δ13C −6.4‰ to +2.2‰. There is a widerspread of oxygen isotope data than carbon isotope data (Fig. 5). Therange of δ13C values for Stage 1 calcitic ostracods is−2.5‰ to +0.3‰.Calcitic ostracods have a narrower field of carbon isotopes than valuesfrom dolomitic fossils or void cements (Fig. 5). The calcitic carapacefilling cements (Stage 3) have more negative carbon isotope values(Fig. 5), down to−3.8‰. Two outlying results have significantly morenegative carbon isotopes (−6‰), and are derived from a ferroandolomite bivalve (non-marine) and an ostracod (marine).
Calcitic specimens of Shemonaella spp. and Hollinella radiata arefrom fully marine and marginal marine sediments. The fully marineostracods have δ18O that is on average more negative by 1‰ thanmarginal marine ostracods (Fig. 5). The calcitic brachiopod Productushas slightly more negative δ18O values (−4.4‰ to −4.2‰) than the

Table 2Carboniferous ostracod electron microprobe data for the specimens in Fig. 5K and L.Values are reported in weight percent, and numbers refer to the spot analysis position(Fig. 5K, L).
Thin section MPK 14017 contains Hollinella (Keslingella) radiataCarapace area samples: outer (1–3), middle (4–6), inner (7–9)
1 2 3 4 5 6 7 8 9
CaO 54.5 54.3 54.4 54.9 54.8 54.6 47.3 46.3 54.1MgO 0.6 0.6 0.5 0.7 0.5 0.5 0.8 0.8 0.8FeO 0.3 0.2 0.2 0.1 0.3 0.1 1.1 1.2 0.4MnO 0.1 0.1 0.1 0.1 0.1 0.0 0.5 0.4 0.2SrO 0.3 0.3 0.3 0.3 0.3 0.3 0.1 0.1 0.2BaO 0.1 0.0 0.1 0.1 0.1 0.0 0.1 0.0 0.0CO2 44.2 44.4 44.4 43.8 43.9 44.5 50.2 51.2 44.2Total 100.0 100.0 100.0 100.0 100.0 100.0 100.0 100.0 100.0
Thin section MPK 14015 contains ostracods indet
1 2 3 4 5 6
CaO 53.9 54.1 55.7 49.3 54.8 44.0MgO 0.3 0.4 0.1 0.3 0.2 0.4FeO 0.6 0.9 0.7 0.5 0.2 1.4MnO 0.1 0.3 0.5 0.3 0.1 0.2SrO 0.1 0.0 0.0 0.1 0.2 0.1BaO 0.0 0.0 0.1 0.0 0.0 0.1CO2 45.0 44.2 42.9 49.6 44.5 53.8Total 100.0 100.0 100.0 100.0 100.0 100.0
156 C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
calcitic ostracod carapaces, but more positive δ13C values (+0.8‰ to+1.3‰) (Fig. 5). Prior analysis of Productus valves from a polishedthin section of this sample did not reveal any evidence ofluminescence under CL.
The isotopic composition of the ostracods and macrofossils plottedagainst their diagenetic stages (Fig. 5) illustrates that dolomiticostracods and macrofossils typically have more negative δ18O valuesthan calcitic specimens. The calcitic ostracods have the highest δ18O(−3.7‰ to −0.9‰), while the dolomitic ostracods and void fillingcements occupy a wider range, but with on average much lowervalues (−12.9‰ to −2.9‰).
5. Interpretation
5.1. Petrographic and compositional interpretation of diagenetic stages
5.1.1. Stage 1Ostracods have a neomorphic carapace ultrastructure, formed by
the process of recrystallisation. The luminescence of calciticfossil ostracods under CL also indicates diagenesis. The pristinepreservation of low-magnesium calcite fossils is commonly indicatedby a lack of luminescence, for example in brachiopod shells (Bannerand Kaufman, 1994; Mii et al., 1999, 2001). In the fossil ostracods, theluminescent carapace laminae do not have different manganese oriron concentrations (that would affect luminescence). The multi-laminate structure revealed by luminescence is a remnant of theoriginal ultrastructure seen in Recent ostracods, and is retaineddespite the growth of neomorphic calcite. The trace elements that arepresent in the neomorphic calcite (iron, magnesium and manganese)are likely to be the result of contamination with pyrite-bearing claysediments.
The calcitic fossil ostracods of this study appear to have beenrecrystallised on the sediment surface or at very shallow burialdepths, on the basis of three factors: 1, the neomorphic calcite is thefirst stage of diagenesis, before framboidal pyrite growth in the topfew metres of sediment; 2, the fidelity of ornamental preservationimplies an early replacement, while the original cellular template wasstill intact; and 3, the ultrastructure is comparable with that seen inshallow diagenesis foraminifera (Sexton et al., 2006).
The recrystallisation process of original calcite dissolution andrecrystallisation into larger neomorphic crystals is poorly understood.The degradation of the organo-chitinous integument may haveallowed the nucleation of larger neomorphic crystals. The originalcrystals may not have completely dissolved, but instead just grown insize. A key question is whether diagenesis took place prior to burial (inthe same water the ostracods lived in) or post-burial, with differentfluid types, such as meteoric water?We suggest that the recrystallisa-tion happened pre-burial or relatively soon after burial, mainly on thebasis of the high fidelity of preservation and the low-magnesiumcalcite carapace composition.
Diagenetically altered planktonic foraminifera from the Cenozoichave a similar ultrastructure to that described here for Stage 1ostracods (Pearson et al., 2001; Sexton et al., 2006; Pearson andBurgess, 2008). Pristine foraminifera have a sub-micron low-magne-sium calcite ultrastructure that is replaced by 1–2 μm diameter size ofdiagenetic neomorphic calcite crystals. The neomorphic calcite isinterpreted to have grown by dissolution of the test and recrystallisa-tion at shallow burial depths, of less than 300 m (Pearson et al., 2001).The effect of this diagenetic recrystallisation was to significantly alterthe oxygen isotope signal from that of the sea surface (where theplanktonic foraminifera lived) to bottom water (where the recrys-tallisation occurred; see Pearson et al., 2001; Williams et al., 2005a,b,2007).
5.1.2. Stage 2The growth of euhedral and framboidal pyrite occurs at an early
diagenetic stage, when sediment is at the sediment–water interface,or shallow burial depths (of a few metres). The sulphate-reducingbacterial reactions to produce pyrite occur due to contact with salinewaters (Raiswell, 1982; Raiswell and Berner, 1985; Brett and Baird,1986; Wilkin et al., 1996; Taylor and Macquaker, 2000). Pyritemorphologies and the degree of pyritisation are affected by manyfactors and are not always linked to sediment redox conditions(Roychoudhury et al., 2003).
5.1.3. Stage 3In marine mudstones the formation of ferroan calcite occurs at a
few metres burial depth; below the sulphate-reduction (pyritegrowth) zone and the methanogenic (siderite growth) zone, in thedecarboxylation zone (Macquaker et al., 1997). This shallow depth offormation is evidenced by the fact that most ferroan calcite isassociated with Stage 1 calcite or Stage 2 pyrite.
5.1.4. Stage 4Coarse or void-filling dolomite crystals often represent a later
diagenetic stage than microcrystalline crystal growth, for examplethat related to deep burial or hydrothermal alteration (Gregg andShelton, 1990; Al-Aasm and Packard, 2000; Gregg et al., 2001). Here,the void-fills are interpreted to have formed after the carapacereplacement. The ferroan dolomite was precipitated from hydrother-mal fluids, which were responsible for the dolomitisation of thelimestones of Fife (Searl and Fallick, 1990; Searl, 1991). Thedestruction of the ostracod carapace ultrastructure indicates thedissolution of the neomorphic calcitic carapace before dolomiteprecipitation, possibly by hot hydrothermal fluids.
5.1.5. Stage 5Only a remnant of the original multi-laminate carapace structure
remains, and is seen by siderite mineralisation. The remnant structureindicates that the recrystallisation of the ostracod carapace may nothave destroyed the carapace template of chitinous-organic material.The location of siderite within the structure of the ostracod carapacereveals its growth from the dedolomitisation of ferroan dolomite byacidic pore waters (Searl and Fallick, 1990).
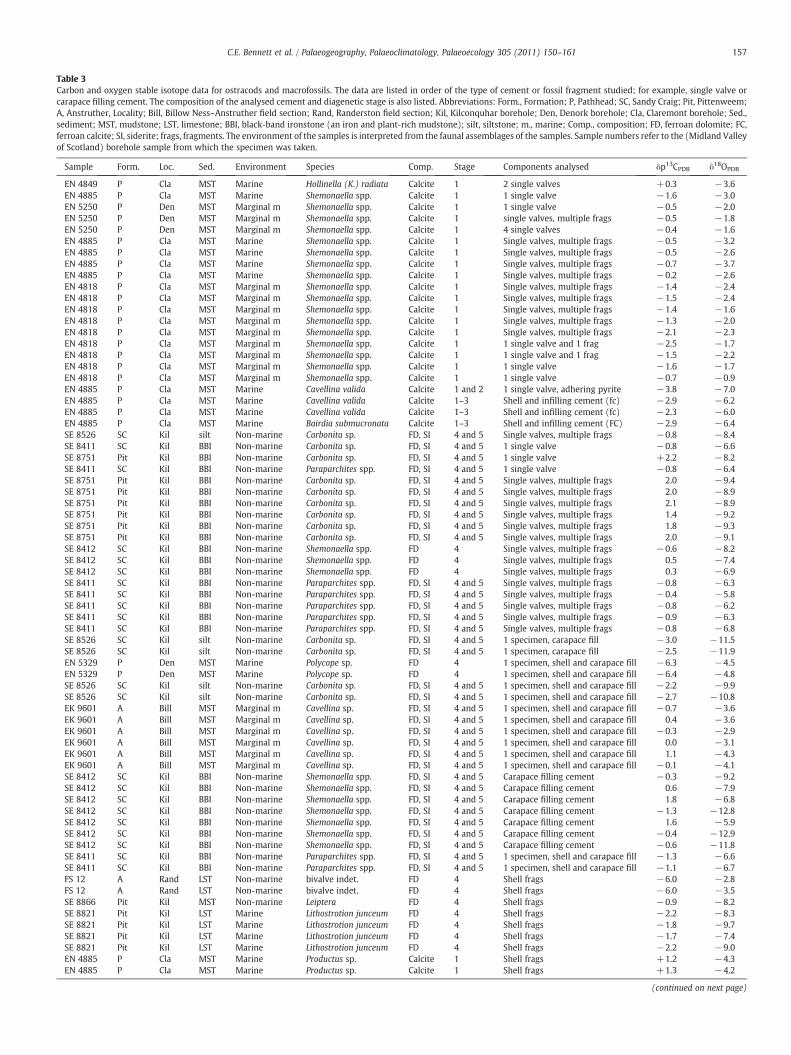
Table 3Carbon and oxygen stable isotope data for ostracods and macrofossils. The data are listed in order of the type of cement or fossil fragment studied; for example, single valve orcarapace filling cement. The composition of the analysed cement and diagenetic stage is also listed. Abbreviations: Form., Formation; P, Pathhead; SC, Sandy Craig; Pit, Pittenweem;A, Anstruther, Locality; Bill, Billow Ness–Anstruther field section; Rand, Randerston field section; Kil, Kilconquhar borehole; Den, Denork borehole; Cla, Claremont borehole; Sed.,sediment; MST, mudstone; LST, limestone; BBI, black-band ironstone (an iron and plant-rich mudstone); silt, siltstone; m., marine; Comp., composition; FD, ferroan dolomite; FC,ferroan calcite; SI, siderite; frags, fragments. The environment of the samples is interpreted from the faunal assemblages of the samples. Sample numbers refer to the (Midland Valleyof Scotland) borehole sample from which the specimen was taken.
Sample Form. Loc. Sed. Environment Species Comp. Stage Components analysed δp13CPDB δ18OPDB
EN 4849 P Cla MST Marine Hollinella (K.) radiata Calcite 1 2 single valves +0.3 −3.6EN 4885 P Cla MST Marine Shemonaella spp. Calcite 1 1 single valve −1.6 −3.0EN 5250 P Den MST Marginal m Shemonaella spp. Calcite 1 1 single valve −0.5 −2.0EN 5250 P Den MST Marginal m Shemonaella spp. Calcite 1 single valves, multiple frags −0.5 −1.8EN 5250 P Den MST Marginal m Shemonaella spp. Calcite 1 4 single valves −0.4 −1.6EN 4885 P Cla MST Marine Shemonaella spp. Calcite 1 Single valves, multiple frags −0.5 −3.2EN 4885 P Cla MST Marine Shemonaella spp. Calcite 1 Single valves, multiple frags −0.5 −2.6EN 4885 P Cla MST Marine Shemonaella spp. Calcite 1 Single valves, multiple frags −0.7 −3.7EN 4885 P Cla MST Marine Shemonaella spp. Calcite 1 Single valves, multiple frags −0.2 −2.6EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 Single valves, multiple frags −1.4 −2.4EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 Single valves, multiple frags −1.5 −2.4EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 Single valves, multiple frags −1.4 −1.6EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 Single valves, multiple frags −1.3 −2.0EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 Single valves, multiple frags −2.1 −2.3EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 1 single valve and 1 frag −2.5 −1.7EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 1 single valve and 1 frag −1.5 −2.2EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 1 single valve −1.6 −1.7EN 4818 P Cla MST Marginal m Shemonaella spp. Calcite 1 1 single valve −0.7 −0.9EN 4885 P Cla MST Marine Cavellina valida Calcite 1 and 2 1 single valve, adhering pyrite −3.8 −7.0EN 4885 P Cla MST Marine Cavellina valida Calcite 1–3 Shell and infilling cement (fc) −2.9 −6.2EN 4885 P Cla MST Marine Cavellina valida Calcite 1–3 Shell and infilling cement (fc) −2.3 −6.0EN 4885 P Cla MST Marine Bairdia submucronata Calcite 1–3 Shell and infilling cement (FC) −2.9 −6.4SE 8526 SC Kil silt Non-marine Carbonita sp. FD, SI 4 and 5 Single valves, multiple frags −0.8 −8.4SE 8411 SC Kil BBI Non-marine Carbonita sp. FD, SI 4 and 5 1 single valve −0.8 −6.6SE 8751 Pit Kil BBI Non-marine Carbonita sp. FD, SI 4 and 5 1 single valve +2.2 −8.2SE 8411 SC Kil BBI Non-marine Paraparchites spp. FD, SI 4 and 5 1 single valve −0.8 −6.4SE 8751 Pit Kil BBI Non-marine Carbonita sp. FD, SI 4 and 5 Single valves, multiple frags 2.0 −9.4SE 8751 Pit Kil BBI Non-marine Carbonita sp. FD, SI 4 and 5 Single valves, multiple frags 2.0 −8.9SE 8751 Pit Kil BBI Non-marine Carbonita sp. FD, SI 4 and 5 Single valves, multiple frags 2.1 −8.9SE 8751 Pit Kil BBI Non-marine Carbonita sp. FD, SI 4 and 5 Single valves, multiple frags 1.4 −9.2SE 8751 Pit Kil BBI Non-marine Carbonita sp. FD, SI 4 and 5 Single valves, multiple frags 1.8 −9.3SE 8751 Pit Kil BBI Non-marine Carbonita sp. FD, SI 4 and 5 Single valves, multiple frags 2.0 −9.1SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD 4 Single valves, multiple frags −0.6 −8.2SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD 4 Single valves, multiple frags 0.5 −7.4SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD 4 Single valves, multiple frags 0.3 −6.9SE 8411 SC Kil BBI Non-marine Paraparchites spp. FD, SI 4 and 5 Single valves, multiple frags −0.8 −6.3SE 8411 SC Kil BBI Non-marine Paraparchites spp. FD, SI 4 and 5 Single valves, multiple frags −0.4 −5.8SE 8411 SC Kil BBI Non-marine Paraparchites spp. FD, SI 4 and 5 Single valves, multiple frags −0.8 −6.2SE 8411 SC Kil BBI Non-marine Paraparchites spp. FD, SI 4 and 5 Single valves, multiple frags −0.9 −6.3SE 8411 SC Kil BBI Non-marine Paraparchites spp. FD, SI 4 and 5 Single valves, multiple frags −0.8 −6.8SE 8526 SC Kil silt Non-marine Carbonita sp. FD, SI 4 and 5 1 specimen, carapace fill −3.0 −11.5SE 8526 SC Kil silt Non-marine Carbonita sp. FD, SI 4 and 5 1 specimen, carapace fill −2.5 −11.9EN 5329 P Den MST Marine Polycope sp. FD 4 1 specimen, shell and carapace fill −6.3 −4.5EN 5329 P Den MST Marine Polycope sp. FD 4 1 specimen, shell and carapace fill −6.4 −4.8SE 8526 SC Kil silt Non-marine Carbonita sp. FD, SI 4 and 5 1 specimen, shell and carapace fill −2.2 −9.9SE 8526 SC Kil silt Non-marine Carbonita sp. FD, SI 4 and 5 1 specimen, shell and carapace fill −2.7 −10.8EK 9601 A Bill MST Marginal m Cavellina sp. FD, SI 4 and 5 1 specimen, shell and carapace fill −0.7 −3.6EK 9601 A Bill MST Marginal m Cavellina sp. FD, SI 4 and 5 1 specimen, shell and carapace fill 0.4 −3.6EK 9601 A Bill MST Marginal m Cavellina sp. FD, SI 4 and 5 1 specimen, shell and carapace fill −0.3 −2.9EK 9601 A Bill MST Marginal m Cavellina sp. FD, SI 4 and 5 1 specimen, shell and carapace fill 0.0 −3.1EK 9601 A Bill MST Marginal m Cavellina sp. FD, SI 4 and 5 1 specimen, shell and carapace fill 1.1 −4.3EK 9601 A Bill MST Marginal m Cavellina sp. FD, SI 4 and 5 1 specimen, shell and carapace fill −0.1 −4.1SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD, SI 4 and 5 Carapace filling cement −0.3 −9.2SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD, SI 4 and 5 Carapace filling cement 0.6 −7.9SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD, SI 4 and 5 Carapace filling cement 1.8 −6.8SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD, SI 4 and 5 Carapace filling cement −1.3 −12.8SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD, SI 4 and 5 Carapace filling cement 1.6 −5.9SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD, SI 4 and 5 Carapace filling cement −0.4 −12.9SE 8412 SC Kil BBI Non-marine Shemonaella spp. FD, SI 4 and 5 Carapace filling cement −0.6 −11.8SE 8411 SC Kil BBI Non-marine Paraparchites spp. FD, SI 4 and 5 1 specimen, shell and carapace fill −1.3 −6.6SE 8411 SC Kil BBI Non-marine Paraparchites spp. FD, SI 4 and 5 1 specimen, shell and carapace fill −1.1 −6.7FS 12 A Rand LST Non-marine bivalve indet. FD 4 Shell frags −6.0 −2.8FS 12 A Rand LST Non-marine bivalve indet. FD 4 Shell frags −6.0 −3.5SE 8866 Pit Kil MST Non-marine Leiptera FD 4 Shell frags −0.9 −8.2SE 8821 Pit Kil LST Marine Lithostrotion junceum FD 4 Shell frags −2.2 −8.3SE 8821 Pit Kil LST Marine Lithostrotion junceum FD 4 Shell frags −1.8 −9.7SE 8821 Pit Kil LST Marine Lithostrotion junceum FD 4 Shell frags −1.7 −7.4SE 8821 Pit Kil LST Marine Lithostrotion junceum FD 4 Shell frags −2.2 −9.0EN 4885 P Cla MST Marine Productus sp. Calcite 1 Shell frags +1.2 −4.3EN 4885 P Cla MST Marine Productus sp. Calcite 1 Shell frags +1.3 −4.2
(continued on next page)
157C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
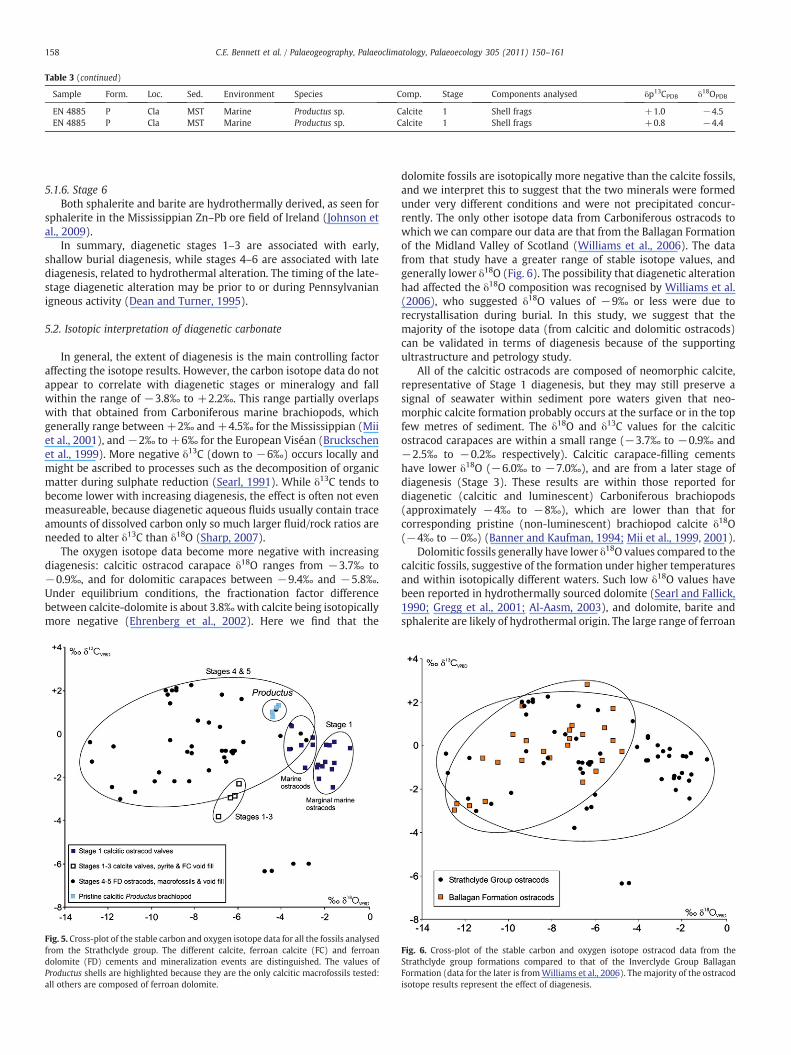
Table 3 (continued)
Sample Form. Loc. Sed. Environment Species Comp. Stage Components analysed δp13CPDB δ18OPDB
EN 4885 P Cla MST Marine Productus sp. Calcite 1 Shell frags +1.0 −4.5EN 4885 P Cla MST Marine Productus sp. Calcite 1 Shell frags +0.8 −4.4
158 C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
5.1.6. Stage 6Both sphalerite and barite are hydrothermally derived, as seen for
sphalerite in the Mississippian Zn–Pb ore field of Ireland (Johnson etal., 2009).
In summary, diagenetic stages 1–3 are associated with early,shallow burial diagenesis, while stages 4–6 are associated with latediagenesis, related to hydrothermal alteration. The timing of the late-stage diagenetic alteration may be prior to or during Pennsylvanianigneous activity (Dean and Turner, 1995).
5.2. Isotopic interpretation of diagenetic carbonate
In general, the extent of diagenesis is the main controlling factoraffecting the isotope results. However, the carbon isotope data do notappear to correlate with diagenetic stages or mineralogy and fallwithin the range of −3.8‰ to +2.2‰. This range partially overlapswith that obtained from Carboniferous marine brachiopods, whichgenerally range between +2‰ and +4.5‰ for the Mississippian (Miiet al., 2001), and−2‰ to +6‰ for the European Viséan (Bruckschenet al., 1999). More negative δ13C (down to −6‰) occurs locally andmight be ascribed to processes such as the decomposition of organicmatter during sulphate reduction (Searl, 1991). While δ13C tends tobecome lower with increasing diagenesis, the effect is often not evenmeasureable, because diagenetic aqueous fluids usually contain traceamounts of dissolved carbon only so much larger fluid/rock ratios areneeded to alter δ13C than δ18O (Sharp, 2007).
The oxygen isotope data become more negative with increasingdiagenesis: calcitic ostracod carapace δ18O ranges from −3.7‰ to−0.9‰, and for dolomitic carapaces between −9.4‰ and −5.8‰.Under equilibrium conditions, the fractionation factor differencebetween calcite-dolomite is about 3.8‰with calcite being isotopicallymore negative (Ehrenberg et al., 2002). Here we find that the
Fig. 5. Cross-plot of the stable carbon and oxygen isotope data for all the fossils analysedfrom the Strathclyde group. The different calcite, ferroan calcite (FC) and ferroandolomite (FD) cements and mineralization events are distinguished. The values ofProductus shells are highlighted because they are the only calcitic macrofossils tested:all others are composed of ferroan dolomite.
dolomite fossils are isotopically more negative than the calcite fossils,and we interpret this to suggest that the two minerals were formedunder very different conditions and were not precipitated concur-rently. The only other isotope data from Carboniferous ostracods towhich we can compare our data are that from the Ballagan Formationof the Midland Valley of Scotland (Williams et al., 2006). The datafrom that study have a greater range of stable isotope values, andgenerally lower δ18O (Fig. 6). The possibility that diagenetic alterationhad affected the δ18O composition was recognised by Williams et al.(2006), who suggested δ18O values of −9‰ or less were due torecrystallisation during burial. In this study, we suggest that themajority of the isotope data (from calcitic and dolomitic ostracods)can be validated in terms of diagenesis because of the supportingultrastructure and petrology study.
All of the calcitic ostracods are composed of neomorphic calcite,representative of Stage 1 diagenesis, but they may still preserve asignal of seawater within sediment pore waters given that neo-morphic calcite formation probably occurs at the surface or in the topfew metres of sediment. The δ18O and δ13C values for the calciticostracod carapaces are within a small range (−3.7‰ to −0.9‰ and−2.5‰ to −0.2‰ respectively). Calcitic carapace-filling cementshave lower δ18O (−6.0‰ to −7.0‰), and are from a later stage ofdiagenesis (Stage 3). These results are within those reported fordiagenetic (calcitic and luminescent) Carboniferous brachiopods(approximately −4‰ to −8‰), which are lower than that forcorresponding pristine (non-luminescent) brachiopod calcite δ18O(−4‰ to −0‰) (Banner and Kaufman, 1994; Mii et al., 1999, 2001).
Dolomitic fossils generally have lower δ18O values compared to thecalcitic fossils, suggestive of the formation under higher temperaturesand within isotopically different waters. Such low δ18O values havebeen reported in hydrothermally sourced dolomite (Searl and Fallick,1990; Gregg et al., 2001; Al-Aasm, 2003), and dolomite, barite andsphalerite are likely of hydrothermal origin. The large range of ferroan
Fig. 6. Cross-plot of the stable carbon and oxygen isotope ostracod data from theStrathclyde group formations compared to that of the Inverclyde Group BallaganFormation (data for the later is fromWilliams et al., 2006). The majority of the ostracodisotope results represent the effect of diagenesis.

159C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
dolomite oxygen isotope values and variation between differentformations perhaps suggests multiple stages of hydrothermal activitythat affected different localities to varying degrees.
6. Discussion
6.1. Palaeoenvironmental signals
The neomorphic calcite in Stage 1 ostracod carapaces preserve apalaeoenvironmental signal incorporated during the precipitation ofthe carapace (with some possible early burial alteration) providinginformation on the environment in which the ostracods lived. Theonly specimen analysed that we consider to preserve pristine calcite isa specimen of a Productus brachiopod. Productus shells composed oflow-magnesium calcite are non-luminescent and show no ultrastruc-tural indications of diagenesis such as recrystallisation. Importantly,this was a fully marine organism, so the isotope composition of itscalcite can be used to interpret seawater conditions in the Mississip-pian (assuming no vital effects, Brand et al., 2003). The Productus δ18Ois −4.3‰, similar to (but at the lower end of) published estimates ofpristine Carboniferous marine carbonate (−4‰). The Stage 1neomorphic ostracods δ18O (−3.7‰ to −0.9‰) sit within thedocumented range of Mississippian pristine brachiopod shell calcite(−4‰ to 0) (Mii et al., 2001: including data from Popp et al., 1986;Grossman, 1994; Bruckschen et al., 1999; Mii et al., 1999). Thissuggests that the ostracod neomorphic calcite preserves a seawatersignal even though theremay have been some shallow alteration soonafter burial.
The only other study to which we can compare ultrastructure andisotopes is a planktonic foraminiferal study which showed thatplanktonic specimens recrystallised their tests at the ocean bottom (indeep water), thus altering the isotope signal from the original surfacewaters (Pearson et al., 2001). In contrast, ostracods in this study aremostly benthic and shallow marine so pore waters are likely to besimilar to the waters in which the ostracods calcified their carapace.Williams et al. (2006) suggested that some of the Ballagan Formationostracods with low δ18O were as a result of the ostracods living in amore brackish environment, and that low δ18O values were due to amixing of seawater with freshwater. While we cannot discount this,none of our nemorphic calcite ostracods have δ18O values that arelower than that previously published for Carboniferous marinecarbonate.
6.2. The effect of palaeogeography and palaeoenvironment
We can use our “pristine” δ18O range to estimate palaeotempera-tures of ocean waters using an empirical temperature equation. Here,we use O'Neil et al. (1969), where:
T = 16:9–4:38 δc–δwð Þ + 0:10 δc–δwð Þ2
and δc=δ18OPDB carbonate, and δw=δ18OSMOW.Estimates of seawater (δ18Oseawater) from the Mississippian have
been previously derived from brachiopod data, and range from −1‰to +2.5‰ for a fixed palaeotemperature of +28 °C (Mii et al., 1999,2001) and as low as −6‰ for an average palaeotemperature of+25 °C (Bruckschen et al., 1999). Tropical Viséan sea surfacetemperatures are estimated at between +25 and +30 °C (Mii et al.,1999). Using these values, Mii et al. (1999) suggested that seawaterδ18O of −1‰ was the most probable for the Mississippian.
From the marine calcitic fossils (including Productus) in this study(δ18O −4.3‰ to −2.6‰), and using the open ocean δ18O value of−1‰ (Mii et al., 1999) the calculated tropical sea surface temperaturevalues range from +23 to +32 °C using the above equation. Thisrange of temperature is suitable for a tropical region, but the locationof the Midland Valley of Scotland in a restricted marine basin may
have impacted on the oxygen isotopic composition of the seawater. Itis well known that the δ18O of surface seawater varies greatly in theRecent ocean as a function of evaporation versus precipitation (P–E)effects, especially in shallow waters. For example, Recent restrictedbasins such as the Mediterranean have more positive seawateroxygen isotope values (δ18O +1.5‰: Schmidt et al., 1999) than theopen ocean (δ18O +0.5‰ in equatorial latitudes: Schmidt et al.,1999). Increased evaporation in this Mississippian restricted basinwould have also driven the seawater composition to more positiveoxygen isotopes. The Ballagan Formation of the Inverclyde Group,Midland Valley of Scotland, contains hypersaline sediments andevaporites (Williams et al., 2006). If we assume that the marginalmarine calcitic fossils in this study lived in a restricted basin withseawater δ18O of +1.5‰; then our range in neomorphic calcitesuggests that the ostracods lived in water temperatures of between+27 and +34 °C (using the equation of O'Neil et al., 1969). The everchanging nature of the palaeoenvironment and variation in the openversus restricted nature of the basin means that a range of seawaterδ18O between −1‰ and +1.5‰ is most likely. The effect of aMississippian glaciation (Fielding et al., 2008) on global ice-volumechanges may also have altered oxygen isotope values in the restrictedPalaeotethys seaway, leading to higher temperatures or moreevaporated waters (Mii et al., 1999). The closing of the Palaeotethysocean gateway and start of the Variscan orogeny occurred during themid-Carboniferous, and would have substantially changed oceancirculation patterns and ocean water (Mii et al., 1999).
Putative palaeoenvironmental signatures are present in the calciticostracods from different environments, if the isotope signal isinterpreted as retaining the composition of pre-burial waters. Early-diagenetic calcitic ostracods are present from marine and marginalmarine environments. Marginal marine ostracods have more positiveoxygen isotope values (on average by δ18O +1‰) than marineostracods, which may indicate that they lived in waters that weresubject to more evaporation, such as an embayment or lagoon withelevated salinities. This is further evidence to suggest early-stagereplacement of the carapace by neomorphic calcite precipitated fromsaline versus meteoric waters. Increased run off from the land shouldaffect Stage 1 marginal marine ostracods by lowering their δ13Csignature, compared to marine specimens, due to the influence ofmore negative δ13C derived from land plants and soils. However, thereverse trend is seen, marginal marine specimens have more positiveδ13C than the marine Productus, which may indicate that evaporationis more important than freshwater input.
However, there are many variables that could affect the isotopicsignal besides salinity, such as temperature variation, evaporation,precipitation (the run-off from rain precipitated on higher groundwould have more negative δ18O), and ostracod vital effects. Definingthe environment for these fairly well-preserved Carboniferousostracods is not without difficulties. Due to the general poor state offossil preservation, only low numbers of the Hollinella species (whichis typically marine; Athersuch et al., 2009) were sampled, while themore common genera Shemonaella, is thought to be adaptable to agreater range of salinities (Williams et al., 2006). A wider comparisonof more fresh, brackish, marginal and marine calcitic ostracods in thefuture may help to resolve some of these palaeoenvironmental issues.
7. Conclusions
• A study of the carapace ultrastructure of Carboniferous ostracodsreveals diagenetic alteration that cannot otherwise be detected bycompositional analysis. A protocol is proposed for the examinationof diagenetic alteration of carapaces. The methodology is alsoapplied to macrofossils, and it can be used for other fossil groupsprior to isotope analysis. The protocol involves a visual assessmentof specimens under a binocular microscope; study of carapaceultrastructure under PPL, SEM and CL; testing for diagenetic

160 C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
cements using element mapping and compositional analysis usingSEM and the electron microprobe.
• The carapace ultrastructure of Carboniferous ostracods from centralScotland is compared to Recent ostracods. The procuticle of Recentostracods comprises an arrangement of sub-micron size (less than0.5 μm diameter), equant to elongate calcite crystals. In the Carbon-iferous ostracods the procuticle is recrystallised byneomorphic calciteto produce equant calcite crystals of 1–3 μm diameter, similar to thatknown in recrystallised fossil planktonic foraminifera.
• The reconstructed diagenetic history is: Early, shallow diagenesis: 1,replacement of the original carapace with neomorphic calcite; 2,framboidal and euhedral pyrite growth in voids and in the carapace;3, ferroan calcite replacement of carapaces and growth in cavities.Late diagenesis by hydrothermal alteration: 4, ferroan dolomitereplacement of fossils and void fills; 5, growth of siderite from thededolomitisation of ferroan dolomite; 6, replacement of someferroan dolomite and siderite by sphalerite and barite.
• Carbon and oxygen stable isotope values for a range of ostracod andmacrofossil carapaces are δ13C=−6.4‰ to +2.2‰ and δ18O=−12.9‰ to −0.9‰. The carbon isotope data do not show a clearsignal in terms of diagenesis or palaeoenvironment, suggesting thatthey may preserve a pristine signal of seawater at very shallowburial depths. The δ18O results closely correspond to the six stages ofdiagenesis, with the most negative values for dolomite representinglate stage hydrothermal alteration.
• The petrography and isotope results of Stage 1 neomorphic calciticostracods infer that the environment of diagenesis was very shallowburial or even pre-burial. The oxygen isotope values for neomorphiccalcite ostracod valves are similar to other Carboniferous shellsthought to be unaltered. A rare marine pristine macrofossil(Productus) also sits within the isotopic range of previouslydetermined pristine Carboniferous carbonates.
• Some of the marine macrofossils and ostracods described here giveisotope signals that suggest that the fossils are pristine or haveundergone only early-stage diagenesis, and thus preserve a signal ofCarboniferous seawater, with δ18O values within previously de-scribed values for the Mississippian. This supports the notion thatthe protocol for assessing the diagenetic alteration of ostracodcarapaces accurately categorises pristine carbonates.
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.palaeo.2011.02.028.
Acknowledgements
The Natural Environment Research Council (NERC) funded theproject, grant number NER/S/A/2005/13368. The British GeologicalSurvey (BGS) provided additional CASE award funding through theBritish Universities Funding Initiative. BGS officers are thanked forproviding access to samples and geological information relating to theMidland Valley: Drs Mike Browne, Maxine Akhurst, Sarah Arkley,Mike Howe and Mark Dean. All figured specimens are housed in theBGS palaeontology collections, Keyworth. Dr. Jim Hendry read anearlier version of this manuscript and provided insightful suggestions.Prof. Jim Marshall kindly allowed access to use of the CL equipment atThe University of Liverpool; Dr. David Horne and Dr. Robin Smithdonated Recent ostracods; Prof. Paul Pearson, Dr. Phil Wilby, Prof.Patrick De Deckker, Dr. Jonathan Holmes, Prof. Tõnu Meidla and Dr.Thijs Vandenbroucke contributed to discussions about ostracodcarapace ultrastructure and diagenesis. Two authors (MJL and IPW),publish with permission of the Executive Director of the BGS (NERC).
References
Al-Aasm, I., 2003. Origin and characterization of hydrothermal dolomite in theWesternCanada Sedimentary Basin. Journal of Geochemical Exploration 78–79, 9–15.
Al-Aasm, I., Packard, J.J., 2000. Stabilization of early-formed dolomite: a tale ofdivergence from two Mississippian dolomites. Sedimentary Geology 131, 97–108.
Athersuch, J., Gooday, A.J., Pollard, J.E., Riley, N.J., 2009. Carboniferous. In: Whittaker, J.E., Hart, M.B. (Eds.), Ostracods in British Stratigraphy. The MicropalaeontologicalSociety Special Publications, London, pp. 111–153.
Banner, J.L., Kaufman, J., 1994. The isotopic record of ocean chemistry and diagenesispreserved in non-luminescent brachiopods from Mississippian carbonate rocks,Illinois and Missouri. GSA Bulletin 106, 1074–1082.
Bate, R.H., East, B.A., 1972. The structure of the ostracode carapace. Lethaia 5, 177–194.Bennett, C.E., 2008. A review of the Carboniferous colonisation of non-marine
environments by ostracods. Senckenbergiana lethaea 88, 37–46.Bennett, C.E., Siveter, D.J., Davies, S.J., Williams, M., Wilkinson, I.P., Browne, M., Miller, C.G.,
in press. Ostracods from freshwater and brackish environments of the Carboniferousof the Midland Valley of Scotland: the early colonisation of terrestrial water bodies.Geological Magazine.
Brand, U., Logan, A., Hiller, N., Richardson, J., 2003. Geochemistry of Recentbrachiopods: applications and implications for oceanography and paleoceanogra-phy. Chemical Geology 198, 305–334.
Brenchley, P.J., Carden, G.A., Hints, L., Kaljo, D., Marshall, J.D., Martma, T., Meidla, T.,Nolvak, J., 2003. High-resolution stable isotope stratigraphy of Upper Ordoviciansequences: constraints on the timing of bioevents and environmental changesassociated with mass extinction and glaciation. Geological Society of AmericaBulletin 115, 89–104.
Brett, C.E., Baird, G.C., 1986. Comparative taphonomy: a key to palaeoenvironmentalinterpretation based on fossil preservation. Palaios 1, 207–227.
Browne, M.A.E., Dean, M.T., Hall, I.H.S., McAdam, A.D., Monro, S.K., Chisholm, J.I., 1999. Alithostratigraphical framework for the Carboniferous rocks of the Midland Valley ofScotland. BGS Research Report RR/99/07.
Bruckschen, P., Oesmann, S., Veizer, J., 1999. Isotope stratigraphy of the EuropeanCarboniferous: proxy signals for ocean chemistry, climate and tectonics. ChemicalGeology 161, 127–163.
Chivas, A.R., De Deckker, P., Carapaceey, J.M.G., 1986a. Magnesium content of non-marine ostracod carapaces: a new palaeosalinometer and palaeothermometer.Palaeogeography, Palaeoclimatology, Palaeoecology 54, 43–61.
Chivas, A.R., De Deckker, P., Carapaceey, J.M.G., 1986b. Magnesium and strontium innon-marine ostracod carapaces as indicators of palaeosalinity and palaeotempera-ture. Hydrobiologia 143, 135–142.
Cronin, T.M., Dwyer, G.S., Kamiya, T., Schwede, S., Willard, D.A., 2003. Medieval WarmPeriod, Little Ice Age and 20th century temperature variability from ChesapeakeBay. Global and Planetary Change 36, 17–29.
Cronin, T.M., Thurnell, R., Dwyer, G.S., Saenger, C., Mann, M.E., Vann, C., 2005.Multiproxy evidence of Holocene climate variability from estuarine sediments,eastern North America. Palaeoceanography 20, 1–21.
De Deckker, P., Chivas, A.R., Carapaceey, J.M.G., 1999. Uptake of Mg and Sr in theeuryhaline ostracod Cyprideis determined from in vitro experiments. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 148, 105–116.
Dean, M.T., Turner, N., 1995. Conodont Colour Alteration Index (CAI) values for theCarboniferous of Scotland. Transactions of the Royal Society of Edinburgh EarthSciences 85, 211–220.
Decrouy, L., 2009. Environmental and biological controls on the geochemistry (δ18O,δ13C, Mg/Ca, and Sr/Ca) of living ostracods from Lake Geneva. Unpublished PhDthesis, University of Lausanne.
Dép che, F., 1982. Ultrastructure of the wall of two living ostracods, Herpetocyprischevreuxi (Sars) and Pontocythere elongata (Brady), in comparison with fossilostracods from the Middle Jurassic of Normandy. In: Bate, R.H., Robinson, E.,Sheppard, L.M. (Eds.), Fossil and Recent Ostracods. Ellis Horwood, Chichester, UK,pp. 61–74.
Dwyer, G.S., Chandler, M.A., 2009. Mid-Pliocene sea level and continental ice volumebased on coupled benthic Mg/Ca palaeotemperatures and oxygen isotopes.Philosophical transactions of the Royal Society of London, A 367, 157–168.
Dwyer, G.S., Cronin, T.S., Baker, P.A., Raymo, M.E., Buzas, J.S., Corrège, T., 1995. NorthAtlantic deepwater temperature change during late Pliocene and late Quaternaryclimatic cycles. Science 270, 1347–1351.
Dwyer, G.S., Cronin, T.M., Baker, P.A., 2002. Trace elements in marine ostracodes. TheOstracoda: Applications in Quaternary Research Geophysical Monograph 131,205–225.
Ehrenberg, S.N., Pickard, N.A.H., Svånå, T.A., Oxtoby, N.H., 2002. Cement geochemistryof photozoan carbonate strata (Upper Carboniferous-Lower Permian), Finnmarkcarbonate platform, Barents Sea. Journal of Sedimentary Research 72, 95–115.
Fielding, C.R., Frank, T.D., Isbell, J.L., 2008. The late Paleozoic ice age — a review ofcurrent understanding and synthesis of global climate patterns. The GeologicalSociety of America Special Paper 441, 343–354.
Forsyth, I.H., Chisholm, M.A., 1977. The Geology of East Fife. Natural EnvironmentResearch Council, Institute of Geological Sciences, Scotland, Memoirs of theGeological Survey of Great Britain.
Gregg, J.M., Shelton, K.L., 1990. Dolomitization and dolomite neomorphism in the backreef facies of the Bonneterre and Davis formations (Cambrian), SoutheasternMissouri. Journal of Sedimentary Petrology 60, 549–562.
Gregg, J.M., Shelton, K.L., Johnson, A.W., Somerville, I.D., Wright, W.R., 2001.Dolomitization of the Waulsortian Limestone (Lower Carboniferous) in the IrishMidlands. Sedimentology 48, 745–766.
Griffiths, H.I., Holmes, J.A., 2000. Non-Marine Ostracods and Quaternary Palaeoenvir-onments: Quaternary Research Association, technical guide number 8, London.
Grossman, E.L., 1994. The carbon and oxygen isotope record during the evolution ofPangea: Carboniferous to Triassic. Geological Society of America Special Paper 288,207–228.

161C.E. Bennett et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 150–161
Immenhauser, A., Kenter, J.A.M., Ganssen, G., Bahamonde, J.R., Van Vilet, A., Saher, M.H.,2002. Origin and significance of isotope shifts in Pennslyvanian carbonates(Asturias, NW Spain). Journal of Sedimentary Research 72, 82–94.
Johnson, A.W., Shelton, K.L., Gregg, J.M., Somerville, I.D., Wright, W.R., Nagy, Z.R., 2009.Regional studies of dolomites and their included fluids: recognizing multiplechemically distinct fluids during the complex diagenetic history of LowerCarboniferous (Mississippian) rocks of the Irish Zn–Pb ore field. Mineral Petrology96, 1–18.
Keatings, K.W., Heaton, T.H.E., Holmes, J.A., 2002. The effects of diagenesis on the traceelement and stable isotope geochemistry of non-marine ostracod valves. Journal ofPaleolimnology 28, 245–252.
Keatings, K.W., Holmes, J.A., Heaton, T.H.E., 2006. Effects of pre-treatment on ostracodvalve chemistry. Chemical Geology 235, 250–261.
Keyser, D., 1995. Structural elements on the surface of ostracod carapaces. In: Riha, J.(Ed.), Ostracoda and Biostratigraphy. A. Balkema, Rotterdam, pp. 5–10.
Kornicker, L.S., 1969. Relationship between the free and attached margins of themyodocopid ostracod shell. In: Neale, J.W. (Ed.), The Taxonomy, Morphology andEcology of Recent Ostracoda. Edinburgh, Oliver and Boyd, pp. 109–135.
Macquaker, J.H.S., Curtis, C.D., Coleman, M.L., 1997. The role of iron in mudstonediagenesis: comparison of Kimmeridge Clay Formation mudstone from onshoreand offshore (UKCS) localities. Journal of Sedimentary Research 67, 871–878.
Martill, D.M., Wilby, P.R., Williams, N., 1992. Elemental mapping: a technique forinvestigating delicate phosphatized fossil soft tissues. Palaeontology 35, 869–874.
Mii, H.-S., Grossman, E.L., Yancey, T.E., 1999. Carboniferous isotope stratigraphies ofNorth America: implications for Carboniferous paleoceanography and Mississip-pian glaciation. GSA Bulletin 111, 960–973.
Mii, H.-S., Grossman, E.L., Yancey, T.E., Chuvashov, B., Egorov, A., 2001. Isotopic recordsof brachiopod shells from the Russian Platform — evidence for the onset of mid-Carboniferous glaciation. Chemical Geology 175, 133–147.
Mischke, S., Zhang, C., Borner, A., 2008. Bias of ostracod stable isotope data caused bydrying of sieve residues from water. Journal of Paleolimnology 40, 567–575.
Monaghan, A.A., Parrish, R.R., 2005. Geochronology of Carboniferous–Permianmagmatism in the Midland Valley of Scotland: implications for regionaltectonomagmatic evolution and the numerical time scale. Journal of the GeologicalSociety, London 162, 1–15.
O'Neil, J.R., Clayton, R.N., Mayeda, T.K., 1969. Oxygen isotope fractionation in divalentmetal carbonates. Journal of Chemistry and Physics 51, 5547–5548.
Okada, Y., 1982. Structure and cuticle formation of the reticulated carapace of theostracode Bicornucythere bisanensis. Lethaia 15, 85–101.
Owens, B., McLean, D., Simpson, K.R.M., Shell, P.M.J., Robinson, R., 2005. Reappraisal ofthe Mississippian palynostratigraphy of the east Fife coast, Scotland, UnitedKingdom. Palynology 29, 23–47.
Pearson, P.N., Burgess, C.E., 2008. Foraminifer test preservation and diagenesis:comparison of high latitute Eocene sites. Geological Society, London, SpecialPublications 303, 59–72.
Pearson, P.N., Ditchfield, P.W., Singano, J., Harcourt-Brown, K.G., Nicholas, C.J., Olsson, R.K.,Shackleton, N.J., Hall, M.A., 2001. Warm tropical sea surface temperatures in the LateCretaceous and Eocene epochs. Nature 413, 481–487.
Popp, B.N., Anderson, T.F., Sandberg, P.A., 1986. Brachiopods as indicators of originalisotopic compositions in some Paleozoic limestones. GSA Bulletin 97, 1262–1269.
Raiswell, R., 1982. Pyrite texture, isotopic composition and the availability of iron.American Journal of Science 282, 1244–1263.
Raiswell, R., Berner, R.A., 1985. Pyrite formation in euxinic and semi-euxinic sediments.American Journal of Science 285, 710–724.
Rosenbaum, J., Sheppard, S.M.F., 1986. An isotopic study of siderites, dolomites andankerites at high temperatures. Geochimica et Cosmochimica Acta 50, 1147–1150.
Roychoudhury, A.N., Kostka, J.E., Van Cappellen, P., 2003. Pyritization: a palaeoenvir-onmental and redox proxy reevaluated. Estuarine, Coastal and Shelf Science 57,1183–1193.
Schmidt, G.A., Bigg, G.R., Rohling, E.J., 1999. Global Seawater Oxygen-18 Database.http://data.giss.nasa.gov/o18data/.
Schrag, D.P., DePaolo, D.J., Richter, F.M., 1995. Reconstructing past sea surfacetemperatures: correcting for diagenesis of bulk marine carbonate. Geochimica etCosmochimica Acta 59, 2265–2278.
Searl, A., 1991. Early Dinantian dolomites from East Fife: hydrothermal overprinting ofearly diagenetic fabrics? Journal of the Geological Society, London 148, 737–747.
Searl, A., Fallick, A.E., 1990. Dinantian dolomites from East Fife: hydrothermaloverprinting of early mixing-zone stable isotopic and Fe/Mn compositions. Journalof the Geological Society, London 147, 623–638.
Sexton, P.F., Wilson, P.A., Pearson, P.N., 2006. Microstructural and geochemicalperspectives on planktic foraminiferal perservation: “glassy” versus “frosty”.Geochemistry, Geophysics, Geosystems 7, Q12P19. doi:10.1029/2006GC001291.
Sharp, Z., 2007. Principles of Stable Isotope Geochemistry. Pearson Education, Inc.Sohn, I.G., Kornicker, L.S., 1988. Ultrastructure ofMyodocopid Carapaces (Ostracoda). In:
Hanai, T., Ikeya, N., Ishizaki, K. (Eds.), Evolutionary Biology of Ostracoda, itsFundamentals and Applications. : Proceedings of the ninth international sympo-siumonostracoda, Shizuoka, Japan, 1985. Elsevier, Tokyo, Amsterdam, pp. 243–257.
Stephenson, M.H., Williams, M., Monaghan, A.A., Arkley, S., Smith, R.A., Dean, M.,Browne, M.A.E., Leng, M.J., 2004. Palynomorph and ostracod biostratigraphy of theBallagan Formation, Midland Valley of Scotland, and elucidation of intra-Dinantianunconformities. Proceedings of the Yorkshire Geological Society 55, 131–143.
Sylvester-Bradley, P.C., Benson, R., 1971. Terminology for surface features in ornateostracodes. Lethaia 4, 249–286.
Taylor, K.G., Macquaker, J.H.S., 2000. Early diagenetic pyrite morphology in amudstone-dominated succession: the Lower Jurassic Cleveland Ironstone Forma-tion, eastern England. Sedimentary Geology 131, 77–86.
Von Grafenstein, U., Erlernkeuser, H., Trimborn, P., 1999. Oxygen and carbon isotopes inRecent fresh-water ostracod valves: assessing vital offsets and autecological effectsof interest for palaeoclimate studies. Palaeogeography, Palaeoclimatology, Palaeoe-cology 148, 133–152.
Wansard, G., De Deckker, P., Julia, R., 1998. Variability in ostracod partition coefficientsD(Sr) and D(Mg): implications for lacustrine palaeoenvironmental reconstruc-tions. Chemical Geology 146, 39–54.
Wilkin, R.T., Barnes, H.L., Brantley, S.L., 1996. The size distribution of framboidal pyritein Recent sediments: an indicator of redox conditions. Geochimica et Cosmochi-mica Acta 60, 3897–3912.
Williams, M., Haywood, A.M., Hillenbrand, C.D., Wilkinson, I.P., 2005a. Efficacy of δ18Odata from Pliocene planktonic foraminifer calcite for spatial sea surface temper-ature reconstruction: comparisonwith a fully coupled ocean–atmosphere GCM andfossil assemblage data for the mid-Pliocene. Geological Magazine 142, 399–417.
Williams, M., Haywood, A.M., Taylor, S.P., Valdes, P.J., Sellwood, B.W., Hillenbrand, C.-D.,2005b. Evaluating the efficacy of planktonic foraminifer calcite δ18O data for seasurface temperature reconstruction for the Late Miocene. Geobios 38, 843–863.
Williams, M., Leng, M.L., Stephenson, M.H., Andrews, J.E., Wilkinson, I.P., Siveter, D.J.,Horne, D.J., Vannier, J.M.C., 2006. Evidence that Early Carboniferous ostracodscolonised coastal flood plain brackish water environments. Palaeogeography,Palaeoclimatology, Palaeoecology 230, 299–318.
Williams, M., Haywood, A.M., Vautravers, M., Sellwood, B.W., Hillenbrand, C.D.,Wilkinson, I.P., Miller, C.G., 2007. Relative effect of taphonomy on calcificationtemperature estimates from fossil planktonic foraminifera. Geobios 40, 861–874.
Wilson, R.B., 1989. A study of the Dinantian marine macrofossils of central Scotland.Transactions of the Royal Society of Edinburgh: Earth Sciences 80, 91–126.
Yamada, S., Tsukagoshi, A., Ikeya, N., 2005. Carapace formation of the podocopidostracode Semicytherura species (Crustacea: Ostracoda). Lethaia 38, 323–325.



















