
Drought resistance and recovery in mature Bituminaria bituminosa var. albomarginata
Upload
independentCategory
view
0download
0
ORIGINAL ARTICLE
Hepatoprotective effect of limonin, a natural limonoidfrom the seed of Citrus aurantium var. bigaradia ,on D-galactosamine-induced liver injury in rats
Mona F. Mahmoud & Dalia I. Hamdan & Michael Wink &
Assem M. El-Shazly
Received: 1 July 2013 /Accepted: 30 October 2013 /Published online: 21 November 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Toll-like receptors have been implicated in inflam-mation and injury in various tissues and organs including theliver.We have investigated the effects of limonin isolated fromthe dichloromethane fraction of the seeds of bittersweet or-ange (Citrus aurantium var. bigaradia ) in two dose levels (50and 100 mg/kg) against D-galactosamine (D-GalN)-inducedliver toxicity in comparison with standard silymarin treatmenton Toll-like receptors expression and hepatic injury, using awell-established rat model of acute hepatic inflammation. Thelimonoids in the seeds of bittersweet orange were identified.Oral administration of limonin before D-GalN injection, sig-nificantly attenuated markers of hepatic damage (elevatedliver enzyme activities and total bilirubin) and hepatic inflam-mation (TNF-α, infiltration of neutrophils), oxidative stressand expression of TLR-4 but not TLR-2 in D-GalN-treatedrats. Limonin effects were similar in most aspects to that of thelignan silymarin. The higher dose of limonin (100 mg/kg)performed numerically better for AST and bilirubin, and bothdoses yielded similar results for ALT and GGT. While the
lower dose of limonin (50 mg/kg) performed better againstoxidative stress and liver structural damage as compared to thehigher dose. Limonin exerts protective effects on liver toxicityassociated with inflammation and tissue injury via attenuationof inflammation and reduction of oxidative stress.
Keywords Bittersweet orange . Limonin . D-galactosamine .
Hepatoprotective . Toll-like receptors
Introduction
Inflammation represents a common pathological process as-sociated with several diseases. It is a major element of thewound healing process and the development of fibrosis. Toll-like receptors (TLRs) are a group of receptors that are in-volved in innate immunity and inflammation. TLRs are in-volved in liver fibrosis, caused by many common liver path-ogens, such as viral, parasitic and toxin-induced hepatitis(Yang and Seki 2012). Ten functional TLRs have been iden-tified in humans and 12 in mice (Takeuchi and Akira 2010),but TLR2 and TLR4 are among the most intensively studiedTLRs.
Both TLR2 and TLR4 exert a marked effect on fibrosis inorgans with higher exposure to bacterial TLR ligands, such asthe liver. Previous studies showed that hepatic TLR2 andTLR4 mRNA and protein expression were increased in pa-tients with hepatitis and cirrhosis (Berzsenyi et al. 2011;Soares et al. 2012). Both TLR2 and TLR4 have also beenimplicated in viral hepatitis; lipopeptides derived from hepa-titis C-stimulated TLR2 and TLR4 (Duesberg et al. 2002) andhepatitis C Core protein-stimulated TLR2 (Dolganiuc et al.2004). TLR4 is found mainly in macrophages such as Kupffercells of the liver. TLR4 mediates inflammation and fibrosis inbile duct ligation and toxin-induced models of liver fibrosis(Seki et al. 2007). It plays an important role in recognizing and
M. F. MahmoudDepartment of Pharmacology and Toxicology, Faculty of Pharmacy,Zagazig University, Zagazig 44519, Egypt
D. I. Hamdan :A. M. El-ShazlyDepartment of Pharmacognosy, Faculty of Pharmacy, ZagazigUniversity, Zagazig 44519, Egypt
M. WinkInstitute of Pharmacy and Molecular Biotechnology, HeidelbergUniversity, Im Neuenheimer Feld 364, 69120 Heidelberg, Germany
M. F. Mahmoud (*)Department of Pharmacology, Faculty of Pharmacy, University ofZagazig, Zagazig 44519, Egypte-mail: [email protected]
M. F. Mahmoude-mail: [email protected]
Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261DOI 10.1007/s00210-013-0937-1

mediating macrophage activation and the release of pro-inflammatory cytokines mainly tumor necrosis factor-alpha(TNF-α), which is apparently involved in the progression ofsevere liver injury (Kitazawa et al. 2010).
Furthermore, previous reports showed that TLR2 is impor-tant for the development of liver fibrosis following bile ductligation in mice and without this receptor, mice developed lessfibrosis (Hartmann et al. 2012). Thus interference with TLRsignaling at the ligand or receptor level represents an impor-tant and novel strategy to prevent the process of maladaptivewound healing and fibrosis in organs like liver (Huebener andSchwabe 2013).
The prevalence of viral hepatitis varies throughout theworld, with the highest prevalence reported in Egypt (WHO1999). Previous studies showed that D-GalN-induced liverinjury in animals, which is similar to that of acute viralhepatitis (Chaung et al. 2003; Nakagiri et al. 2003), is com-monly used for the screening of anti-hepatotoxic and/or hepa-toprotective activities of drugs. The toxicity of D-GalN resultsfrom an inhibition of RNA and protein synthesis in the liverthrough depletion of cellular uridine pools. The precise mech-anism of D-GalN-induced hepatitis has not yet been elucidat-ed. Recent investigations demonstrated that D-GalN-inducedhepatotoxicity in rats involved the increase of expression ofTLR4 mRNA (Zheng et al. 2012).
Silymarin is a plant flavonolignan obtained from milkthistle (Silybum marianum). It is used for hepatoprotectionas it possesses antioxidant, antifibrotic and anti-inflammatoryeffects. Silymarin showed hepatoprotective effects in acet-aminophen, carbon tetrachloride, ethanol, D-GalN andAmanita phalloides toxin-induced liver diseases in experi-mental animals (Pradhan and Girish 2006). Furthermore,in vitro studies have suggested that silymarin, promoted he-patocyte regeneration and survival (Wang et al. 2005). So,silymarin is used as a reference standard in experimentalstudies with newly discovered biologically active substanceswith unknown mechanism(s) of action.
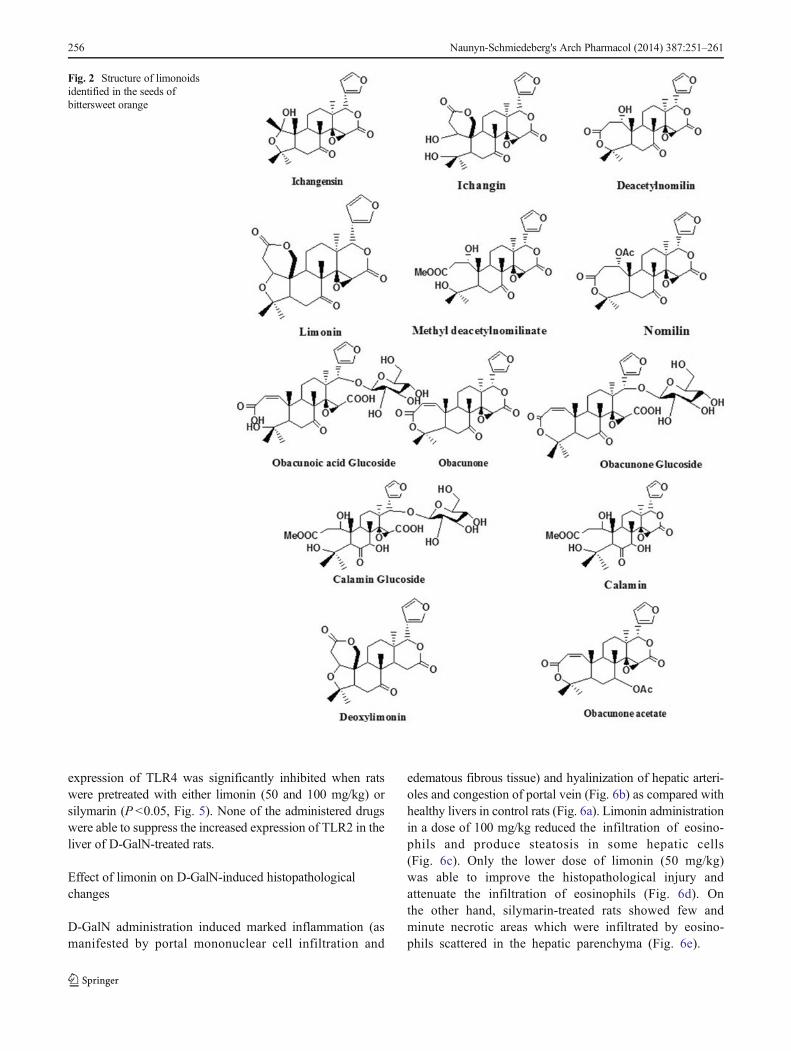
Citrus fruits (family, Rutaceae) are rich in biologicallyimportant classes of secondary metabolites, such as limo-noids, essential oils and flavonoids. Limonin (a typical limo-noid) is one of the main components which belongs to afamily of highly oxygenated triterpenoids responsible forbitterness in citrus fruits (Roy and Saraf 2006; Manners2007). Limonin has antibacterial, antiviral, antinociceptive,anti-inflammatory, anti-carcinogenic in Aflatoxin B1-induced hepatocellular carcinoma, anti-feedant and inhibit p-glycoprotein (Miller et al. 1992; El-Readi et al. 2010;Langeswaran et al. 2012). In addition, limonin exhibits abor-tifacient (Piyachafurawat et al. 1985) and chemopreventiveproperties (Lam et al. 1994). Previous studies showed thatlimonoids derived from Trichilia emetica (Meliaceae) appearto be responsible for hepatoprotective activity in chloroform-induced damage of rat hepatocytes (Germanò et al. 2005).
However, to our knowledge a hepatoprotective effect of purelimonin has not been investigated. Therefore, the aim ofpresent study was to examine the possible hepatoprotectiveeffect of limonin in comparison with plant lignan silymarin (astandard treatment) and illustrate the mechanism of this pro-tective effect.
Materials and methods
Phytochemical study
Plant material
The dried seeds of bittersweet orange (Citrus aurantium var.bigaradia Hook f., Citrus bigaradia Risso & Poit, C.bigaradia Loisel; Rutaceae) were collected from theResearch Center of El-Qanater El-Khireya, province ofKalubeya, Egypt in December 2011. The identity of the plantswas confirmed by Dr. B. Houlyel, Prof. of Pomology, Facultyof Agriculture, Benha University, Egypt. Avoucher specimenwas deposited in the herbarium of the Faculty of Pharmacy,Zagazig University (accession number P-80, 81).
Extraction procedure
The seeds of this species were collected from ripe fruits,cleaned, dried and powdered. The dried powder (450 g) wasdefatted with light petroleum and extracted with methanol.The concentrated methanolic extract (120 g) was suspended inH2O and extracted with CH2Cl2. The solvent was evaporatedunder vacuum to yield 10 g of brownish yellow oily residue.
Isolation of the compounds
Dry mixed initial zone of the dichloromethane fraction (8 g)was chromatographed over a silica gel column (150×4 cm,150 g). The column was eluted with cyclohexane and thepolarity was increased gradually using ethyl acetate followedby methanol. Fractions 250 mL each were collected, concen-trated under vacuum and monitored by TLC using solventsystems (chloroform–methanol, 9:1, 8:2, 6:4 v /v, and cyclo-hexane–ethyl acetate, 2:3 v /v ). TLC was visualized byspraying with 10 % H2SO4 and heated in an oven. Repeatedcrystallization and/or preparative TLC afforded three limo-noid aglycones.
Spectral analysis
Mass spectra were recorded on a MAT 8200 instrument withelectron energy 70 eV. 3-Nitrobenzyl alcohol was used as amatrix in FAB-MS. NMR spectra (1H and 13C) were recordedon VARIAN 500 at 500 MHz for 1H and 125 MHz for 13C.
252 Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261

CDCL3 was used as solvents. Experimental data were evalu-ated using MestRe-C software. The chemical structures of theisolated compounds were identified by interpretation of theirspectral data including MS and NMR experiments (APT,COSY, HSQC, HMBC and NOESY) and comparison withliterature data as well as authentic substances from ourlaboratory.
Liquid chromatography electrospray mass spectrometry(LC-ESI/MS) for limonoids
For identification of the minor limonoids in the dichlorometh-ane fraction, chromatographic separation was carried outusing RP C-18 LiChro CART (Merck- Darmstadt) column[250×4 mm (5 μm)]. The column was eluted at a flow rate1 ml/min with a linear gradient of the solvent mixture, solventA: water–formic acid (99.5:0.5 v /v ) and solvent B: acetoni-trile with the gradient program as follows: 0–5 min 0 % B, 5–110 min from 0 % to 70 % B, 110–115 min from 70 % to100 % B, 115–140 min 100 % B. The analysis was performedunder the following conditions: acquisition mode, ESI posi-tive; nebulizer gas, N2, 0.25 L/min; capillary 3.00 KV, HVlens −0.50 KV; cone, −35 V; source temp., 120˚C; RF lens,−0.2 V; skimmer, −1.5 V; mass scan range, 200–800 m/z ;interval 0.1 s. MS data were acquired and processed usingMassLynx V4.0 software. The purity of limonin used in thecurrent study was tested by HPLC and it was >97 %.
In vivo study of the hepatoprotective activity of limonin
Animals
Thirty adult male Wistar rats (180–200 g; Zagazig University,Zagazig, Egypt) were used in the present study. All the ani-mals were maintained under standard husbandry conditionwith food and water ad libitum. The experimental procedureswere approved by the Institutional Animal Ethics Committeeof the Faculty of Pharmacy, Zagazig University (approvalnumber P2-6, 2012) and animals were handled following theInternational Animal Ethics Guidelines, ensuring minimumanimal suffering.
Study protocol
Animals were randomly divided into five experimental groups(six animals each): control, D-GalN, limonin (50 mg/kg),limonin (100 mg/kg) and silymarin (100 mg/kg). Controlgroup received only the vehicle. D-GalN group received D-GalN, 800mg/kg dissolved in normal saline and given i.p. as asingle dose. Limonin and silymarin (suspended in gum acaciasolution, 10 mg/mL saline w /v ) were given separately to ratsby orogastric gavage. One hour later, rats of these groups wereinjected i.p. with D-GalN. Twenty-four hours after D-GalN
injection, rats were anaesthetized with subcutaneous injectionof 1.2 g/kg urethane (Sigma, St. Louis, MO, USA), blood wascollected from the retro-orbital plexus and centrifuged(3,000×g , 4 °C, 20 min) for separation of serum. The obtainedserum was used to analyse liver enzymes and total bilirubin.Thereafter, animals were sacrificed, livers were separatelydissected and blood was washed off with cold saline; liverswere divided into two parts; one part was immediately flashfrozen in liquid nitrogen and kept at −80 °C for measurementof tissue parameters and the other part was kept in 10 %formalin for histopathological examination.
Biochemical analysis
The serum alanine aminotransferase (ALT), aspartate amino-transferase (AST), gamma-glutamyltransferase (GGT) en-zyme activities and total bilirubin level were measured usingcommercially available analytical kit (Biodiagnostic Co,Egypt).
Determination of oxidative stress
The generation of reactive oxygen species (ROS) in responseto D-GalN-induced liver injury was evaluated in liver tissuesby the measurement of the lipid peroxidation productmalondialdehyde (MDA) and reduced glutathione content(GSH). MDA and GSH were measured in tissue homogenatephotometrically (spectrophotometer, Jenway®, England, UK)according to Wills (1987) and Ellman (1959), respectively.
Tumor necrosis factor alpha (TNF-α)
TNF-α level in liver homogenate was detected by sandwichELISA according to method of Maskos et al. (1998) usingELISA kit (Quantikine, R&D system USA).
Detection of the expression of TLR2 and TLR4 by RT-PCR
RNA was extracted from liver tissue homogenates usingRNeasy purification reagent (Qiagen, Valencia, CA) accord-ing to manufacturer’s instruction. RNA was photometricallyquantified at 260/280 nm. cDNAwas generated from 5 μg oftotal RNA using 20 pmol oligo dT and superscript AMVreverse transcriptase for 60 min at 37 °C. For PCR, cDNAwere incubated with dNTPs (10 mM), 10× PCR buffer, 2.5 UTaq DNA polymerase and 10 pmol of each primer. Thereaction mixture was subjected to 40 cycles of PCR as fol-lows: Denaturation at 95 °C for 1 min, annealing at 60 °C for1 min and extension at 72 °C for 2 min. Ten microlitres of thePCR mixture was electrophoresed on a 2 % agarose gel,stained with ethidium bromide and visualized by ultravioletlight. For semiquantitative analysis, a gel documentation sys-tem was used (BioDO, Analyser, Biometra, Göttingen,
Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261 253

Germany). According to the amplification procedure, relativeexpression of each studied gene (R) was calculated accordingto the following the formula: densitometrical units of eachstudied gene/densitometrical units of beta actin (a house keep-ing gene). Primer sequences are shown in Table 1. The aver-age value of beta actin ranges between 0.91 and 1 in all studiedsamples.
Histopathological study
Liver tissues were fixed in 10 % formalin and embedded inparaffin. Sections of 5–6 μm thickness were stained withhematoxylin and eosin (H & E) then examined under lightmicroscope for determination of histopathological changes.The histological analysis was performed by a person blindedfor treatment.
Ex vivo assay of limonin on cultured hepatocytes
Cell culture
HepG2, a human hepatoma cell line, was first purchased fromVACSERA, Giza, Egypt, and thenmaintained in tissue culturefacility. Cells were grown in RPMI medium supplementedwith heat-inactivated fetal bovine serum (10 %), penicillin G(100 IU/mL) and streptomycin (100 μg/mL). Cells weremaintained at 37 °C in a 5 % CO2 atmosphere with 95 %humidity. Cell culture reagents were obtained from Lonza(Basel, Switzerland).
Hepatotoxicity and treatment
The hepatoprotective activity of limonin was tested in-vitro attwo different concentrations (5 and 10 μM) and compared tothe standard hepatoprotective agent silymarin at 10 μM.HepG2 monolayer culture after attachment was pretreatedwith the assigned sample for 1 h. D-GalN was added at10 μM final concentration and incubation was continued foranother 2 h. The supernatant medium and cell lysate were thencollected and stored at −20 °C until analysis. Positive controlwas a set of cells maintained in culture medium and treatedonly with D-GalN (10 μM), while the negative control was aset of cells maintained in phosphate-buffered saline. The
levels of ALT, AST, MDA and SOD in addition to totalantioxidant capacity (TAC) were assessed in the supernatantusing commercially available kits according to the manufac-turer instructions (Biodiagnostic, Cairo, Egypt).
DPPH radical scavenging activity of limonin
The free radical scavenging activity of limonin was measuredby the 1, 1-diphenyl-2-picryl hydrazyl (DPPH) assay (Yu et al.2005). A 10-μM solution of DPPH in methanol was prepared.Different concentrations of limonin (10–90 μM) were sepa-rately mixed with DPPH solution. The mixtures were vigor-ously mixed and allowed to stand in the dark for 30 min at25 °C. The absorbance of the resulting solution was measuredusing a spectrophotometer at 517 nm and percent radicalscavenging activity was determined in comparison with meth-anol control. Rutin (10 μM) was used as reference standard.The scavenging activity was calculated from the followingequation:
DPPHscavengingeffect %ð Þ ¼ Ac−As=Ac� 100
where Ac = absorbance of control, As = absorbance ofsample.
Statistical analysis
All data are expressed as mean±SEM. Statistical analysis wasperformed by ANOVA followed by Tukey post hoc test usinga computer-based curve fitting program (prism 5, Graphpad,CA, USA). A P value <0.05 was considered statisticallysignificant.
Drugs and chemicals
Chemicals were purchased from AppliChem (Darmstadt,Germany), Fluka (Buchs, Switzerland) and Sigma AldrichGmbH (Sternheim, Germany). D-galactosamine andsilymarin were supplied by Sigma Chemicals (ST. Louis.Mo, USA). All solvents for extraction and separation wereof analytical grade.
Results
Phytochemistry
Various HPLC techniques such as microbore normal-phaseand reversed-phase chromatography coupled to a mass spec-trometer (operating in a positive ion atmospheric pressurechemical ionization and electrospray ionization modes) werefound to be rapid, selective and sensitive for the analysisof citrus limonoids (Manners 2007; Manners et al. 2003;
Table 1 Sequences of PCR primers (after Fan et al. 2009)
Gene Primer sequences
TLR2 Forward 5_-GAA AGATGC GCT TCC TGA AC-3_
Reverse 5_-CGC CTA AGA GCA GGATCA AC-3
TLR4 Forward 5_-GCC GGA AAG TTATTG TGG TGG T-3_
Reverse 5_-ATG GGT TTTAGG CGC AGA GTT T-3_
254 Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261
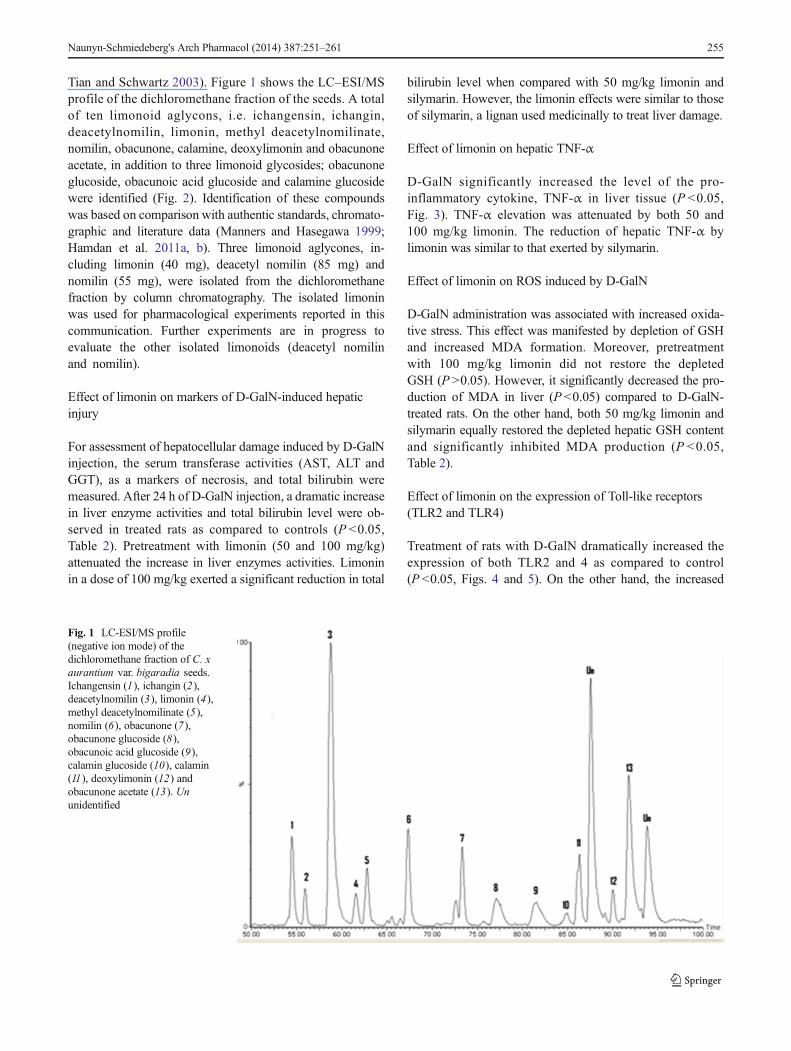
Tian and Schwartz 2003). Figure 1 shows the LC–ESI/MSprofile of the dichloromethane fraction of the seeds. A totalof ten limonoid aglycons, i.e. ichangensin, ichangin,deacetylnomilin, limonin, methyl deacetylnomilinate,nomilin, obacunone, calamine, deoxylimonin and obacunoneacetate, in addition to three limonoid glycosides; obacunoneglucoside, obacunoic acid glucoside and calamine glucosidewere identified (Fig. 2). Identification of these compoundswas based on comparison with authentic standards, chromato-graphic and literature data (Manners and Hasegawa 1999;Hamdan et al. 2011a, b). Three limonoid aglycones, in-cluding limonin (40 mg), deacetyl nomilin (85 mg) andnomilin (55 mg), were isolated from the dichloromethanefraction by column chromatography. The isolated limoninwas used for pharmacological experiments reported in thiscommunication. Further experiments are in progress toevaluate the other isolated limonoids (deacetyl nomilinand nomilin).
Effect of limonin on markers of D-GalN-induced hepaticinjury
For assessment of hepatocellular damage induced by D-GalNinjection, the serum transferase activities (AST, ALT andGGT), as a markers of necrosis, and total bilirubin weremeasured. After 24 h of D-GalN injection, a dramatic increasein liver enzyme activities and total bilirubin level were ob-served in treated rats as compared to controls (P <0.05,Table 2). Pretreatment with limonin (50 and 100 mg/kg)attenuated the increase in liver enzymes activities. Limoninin a dose of 100 mg/kg exerted a significant reduction in total
bilirubin level when compared with 50 mg/kg limonin andsilymarin. However, the limonin effects were similar to thoseof silymarin, a lignan used medicinally to treat liver damage.
Effect of limonin on hepatic TNF-α
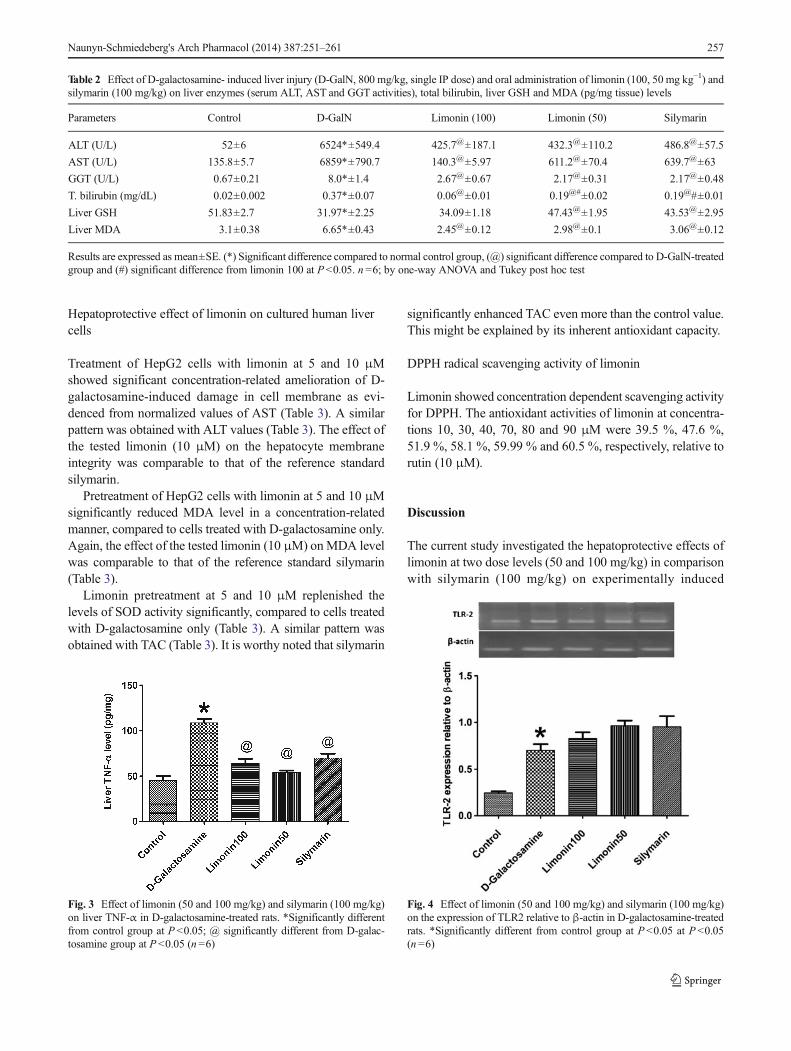
D-GalN significantly increased the level of the pro-inflammatory cytokine, TNF-α in liver tissue (P <0.05,Fig. 3). TNF-α elevation was attenuated by both 50 and100 mg/kg limonin. The reduction of hepatic TNF-α bylimonin was similar to that exerted by silymarin.
Effect of limonin on ROS induced by D-GalN
D-GalN administration was associated with increased oxida-tive stress. This effect was manifested by depletion of GSHand increased MDA formation. Moreover, pretreatmentwith 100 mg/kg limonin did not restore the depletedGSH (P >0.05). However, it significantly decreased the pro-duction of MDA in liver (P <0.05) compared to D-GalN-treated rats. On the other hand, both 50 mg/kg limonin andsilymarin equally restored the depleted hepatic GSH contentand significantly inhibited MDA production (P <0.05,Table 2).
Effect of limonin on the expression of Toll-like receptors(TLR2 and TLR4)
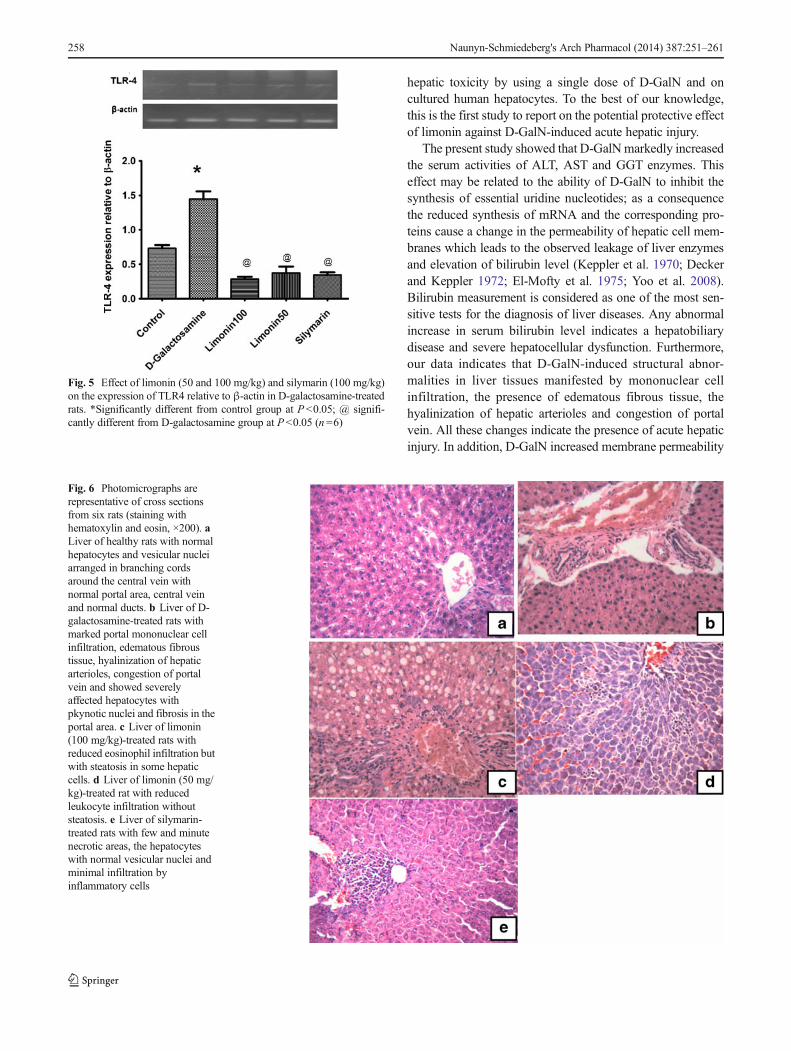
Treatment of rats with D-GalN dramatically increased theexpression of both TLR2 and 4 as compared to control(P <0.05, Figs. 4 and 5). On the other hand, the increased
Fig. 1 LC-ESI/MS profile(negative ion mode) of thedichloromethane fraction of C. xaurantium var. bigaradia seeds.Ichangensin (1), ichangin (2),deacetylnomilin (3), limonin (4),methyl deacetylnomilinate (5),nomilin (6), obacunone (7),obacunone glucoside (8),obacunoic acid glucoside (9),calamin glucoside (10), calamin(11), deoxylimonin (12) andobacunone acetate (13). Ununidentified
Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261 255
expression of TLR4 was significantly inhibited when ratswere pretreated with either limonin (50 and 100 mg/kg) orsilymarin (P <0.05, Fig. 5). None of the administered drugswere able to suppress the increased expression of TLR2 in theliver of D-GalN-treated rats.
Effect of limonin on D-GalN-induced histopathologicalchanges
D-GalN administration induced marked inflammation (asmanifested by portal mononuclear cell infiltration and
edematous fibrous tissue) and hyalinization of hepatic arteri-oles and congestion of portal vein (Fig. 6b) as compared withhealthy livers in control rats (Fig. 6a). Limonin administrationin a dose of 100 mg/kg reduced the infiltration of eosino-phils and produce steatosis in some hepatic cells(Fig. 6c). Only the lower dose of limonin (50 mg/kg)was able to improve the histopathological injury andattenuate the infiltration of eosinophils (Fig. 6d). Onthe other hand, silymarin-treated rats showed few andminute necrotic areas which were infiltrated by eosino-phils scattered in the hepatic parenchyma (Fig. 6e).
Fig. 2 Structure of limonoidsidentified in the seeds ofbittersweet orange
256 Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261
Hepatoprotective effect of limonin on cultured human livercells
Treatment of HepG2 cells with limonin at 5 and 10 μMshowed significant concentration-related amelioration of D-galactosamine-induced damage in cell membrane as evi-denced from normalized values of AST (Table 3). A similarpattern was obtained with ALT values (Table 3). The effect ofthe tested limonin (10 μM) on the hepatocyte membraneintegrity was comparable to that of the reference standardsilymarin.
Pretreatment of HepG2 cells with limonin at 5 and 10 μMsignificantly reduced MDA level in a concentration-relatedmanner, compared to cells treated with D-galactosamine only.Again, the effect of the tested limonin (10 μM) onMDA levelwas comparable to that of the reference standard silymarin(Table 3).
Limonin pretreatment at 5 and 10 μM replenished thelevels of SOD activity significantly, compared to cells treatedwith D-galactosamine only (Table 3). A similar pattern wasobtained with TAC (Table 3). It is worthy noted that silymarin
significantly enhanced TAC even more than the control value.This might be explained by its inherent antioxidant capacity.
DPPH radical scavenging activity of limonin
Limonin showed concentration dependent scavenging activityfor DPPH. The antioxidant activities of limonin at concentra-tions 10, 30, 40, 70, 80 and 90 μM were 39.5 %, 47.6 %,51.9 %, 58.1 %, 59.99 % and 60.5 %, respectively, relative torutin (10 μM).
Discussion
The current study investigated the hepatoprotective effects oflimonin at two dose levels (50 and 100 mg/kg) in comparisonwith silymarin (100 mg/kg) on experimentally induced
Table 2 Effect of D-galactosamine- induced liver injury (D-GalN, 800 mg/kg, single IP dose) and oral administration of limonin (100, 50 mg kg−1) andsilymarin (100 mg/kg) on liver enzymes (serum ALT, AST and GGT activities), total bilirubin, liver GSH and MDA (pg/mg tissue) levels
Parameters Control D-GalN Limonin (100) Limonin (50) Silymarin
ALT (U/L) 52±6 6524*±549.4 425.7@±187.1 432.3@±110.2 486.8@±57.5
AST (U/L) 135.8±5.7 6859*±790.7 140.3@±5.97 611.2@±70.4 639.7@±63
GGT (U/L) 0.67±0.21 8.0*±1.4 2.67@±0.67 2.17@±0.31 2.17@±0.48
T. bilirubin (mg/dL) 0.02±0.002 0.37*±0.07 0.06@±0.01 0.19@#±0.02 0.19@#±0.01
Liver GSH 51.83±2.7 31.97*±2.25 34.09±1.18 47.43@±1.95 43.53@±2.95
Liver MDA 3.1±0.38 6.65*±0.43 2.45@±0.12 2.98@±0.1 3.06@±0.12
Results are expressed as mean±SE. (*) Significant difference compared to normal control group, (@) significant difference compared to D-GalN-treatedgroup and (#) significant difference from limonin 100 at P<0.05. n =6; by one-way ANOVA and Tukey post hoc test
Fig. 3 Effect of limonin (50 and 100 mg/kg) and silymarin (100 mg/kg)on liver TNF-α in D-galactosamine-treated rats. *Significantly differentfrom control group at P <0.05; @ significantly different from D-galac-tosamine group at P <0.05 (n=6)
Fig. 4 Effect of limonin (50 and 100 mg/kg) and silymarin (100 mg/kg)on the expression of TLR2 relative to β-actin in D-galactosamine-treatedrats. *Significantly different from control group at P<0.05 at P<0.05(n =6)
Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261 257
hepatic toxicity by using a single dose of D-GalN and oncultured human hepatocytes. To the best of our knowledge,this is the first study to report on the potential protective effectof limonin against D-GalN-induced acute hepatic injury.
The present study showed that D-GalNmarkedly increasedthe serum activities of ALT, AST and GGT enzymes. Thiseffect may be related to the ability of D-GalN to inhibit thesynthesis of essential uridine nucleotides; as a consequencethe reduced synthesis of mRNA and the corresponding pro-teins cause a change in the permeability of hepatic cell mem-branes which leads to the observed leakage of liver enzymesand elevation of bilirubin level (Keppler et al. 1970; Deckerand Keppler 1972; El-Mofty et al. 1975; Yoo et al. 2008).Bilirubin measurement is considered as one of the most sen-sitive tests for the diagnosis of liver diseases. Any abnormalincrease in serum bilirubin level indicates a hepatobiliarydisease and severe hepatocellular dysfunction. Furthermore,our data indicates that D-GalN-induced structural abnor-malities in liver tissues manifested by mononuclear cellinfiltration, the presence of edematous fibrous tissue, thehyalinization of hepatic arterioles and congestion of portalvein. All these changes indicate the presence of acute hepaticinjury. In addition, D-GalN increased membrane permeability
Fig. 5 Effect of limonin (50 and 100 mg/kg) and silymarin (100 mg/kg)on the expression of TLR4 relative to β-actin in D-galactosamine-treatedrats. *Significantly different from control group at P<0.05; @ signifi-cantly different from D-galactosamine group at P<0.05 (n =6)
Fig. 6 Photomicrographs arerepresentative of cross sectionsfrom six rats (staining withhematoxylin and eosin, ×200). aLiver of healthy rats with normalhepatocytes and vesicular nucleiarranged in branching cordsaround the central vein withnormal portal area, central veinand normal ducts. b Liver of D-galactosamine-treated rats withmarked portal mononuclear cellinfiltration, edematous fibroustissue, hyalinization of hepaticarterioles, congestion of portalvein and showed severelyaffected hepatocytes withpkynotic nuclei and fibrosis in theportal area. c Liver of limonin(100 mg/kg)-treated rats withreduced eosinophil infiltration butwith steatosis in some hepaticcells. d Liver of limonin (50 mg/kg)-treated rat with reducedleukocyte infiltration withoutsteatosis. e Liver of silymarin-treated rats with few and minutenecrotic areas, the hepatocyteswith normal vesicular nuclei andminimal infiltration byinflammatory cells
258 Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261

of cultured human hepatocytes, Hep G2 cells, as evidenced bymarked increase of both ALT and AST activities. This effectmay be attributed to the increased membrane lipid peroxida-tion whereMDAwas increased following D-GalN applicationand reduction in the total antioxidant capacity and SOD ac-tivity were also observed.
In the current study, limonin was administered beforeinjection of D-GalN as a preventive strategy. Severalprevious studies administered drugs as preventive strategy inD-GalN-induced hepatic toxicity model (Nakagiri et al. 2003;Lian et al. 2010; Yoo et al. 2008; Sheik Abdulazeez andThiruvengadam 2013). Pretreatment with limonin virtuallyinhibited the development of hepatic injury in D-GalN-treatedrats. The following findings can explain the ability of limonin tocounteract hepatic injury accompanying D-GalN administration:(a) limonin ameliorated the biochemical changes and suppressedthe infiltration of leukocytes to the hepatic tissues, (b) the in-creased TNF-α production and the elevated oxidative stressassociated with D-GalN administration were abrogated bylimonin pretreatment and (c) limonin suppressed the over ex-pression of TLR-4 in D-GalN-treated rats. These findings pro-vide convincing evidence that limonin offsets the acute hepaticinjury accompanying D-GalN administration through antioxi-dant and anti-inflammatory effects.
We reasoned that limonin administration would minimizeor even mitigate the acute hepatic injury induced by D-GalNadministration. This assumption was ascertained in this workbecause both doses of limonin used significantly inhibited theelevations in liver enzymes activities and bilirubin in animalstreated with D-GalN in a way comparable and even better thansilymarin. The obtained results confirm the previous studywith aflatoxin B1-treated animals and showed that limonindecreased the elevated serum activities of AST and ALT backto normal (Langeswaran et al. 2012). Similarly, Germanò et al.(2005) found that mixed limonoids derived from T. emeticaprotected the isolated rat hepatocytes from CCL4-inducedtoxicity and had antibacterial activity as well. However, un-expectedly, administration of the higher dose of limonin(100 mg/kg) was associated with the development of steatosisin liver. This effect may be attributed to the ability of limonin
to inhibit CYP3A4 activity (Iwata et al. 2005). Previousstudies indicated that hepatic steatosis is associated with de-creased hepatic CYP3A4 activity in humans via post-translational mechanism (Kolwankar et al. 2007).
We investigated the effect of limonin pretreatment on theexpression of Toll-like receptors because of its importance inthe development of liver disorders and in particular viralhepatitis (Dolganiuc et al. 2004). We have found increasedexpression of both TLR2 and TLR4 genes in hepatic tissues ofanimals treated with D-GalN, while both limonin andsilymarin prevented only the increased expression of TLR4without affecting the increased expression of TLR2. Thesefindings are supported by previous reports, which showedincreased levels of hepatic TLR2 and TLR4 mRNA andcorresponding proteins in patients with hepatitis and cirrhosis(Berzsenyi et al. 2011; Soares et al. 2012). Increased TLR2and TLR4 expression was also observed in various models ofexperimental hepatic injury such as partial hepatectomy andacute hepatic failure (Takayashiki et al. 2004; Kitazawa et al.2008; Tu et al. 2012). Results of the present study showed thatTLR-4 plays a more important role in D-galactosamine-induced rat liver injury than TLR-2 and the inhibition ofTLR-4 expression might attenuate functional and structuralchanges induced by D-GalN. These findings were notmatched with the previous study of Hartmann et al. (2012)who showed that TLR2-deficient mice showed less liverfibrosis and intestinal translocation of bacteria and bacterialproducts than wild-type mice following bile duct ligation.This difference may be attributed to the variation of animalspecies and the model of liver injury. Activation of the TLR4signal pathway initiates a cascade of events, including trans-location of nuclear factor kappa B (NF-κB) to the nucleus,leading to production and release of inflammatory cytokines(Cotroneo et al. 2012). Due to its strong reduction of TLR4expression, we suggest that limonin exhibits anti-inflammatoryactivity. Our results clearly demonstrated that limonin andsilymarin inhibited TNF-α production which is involved inTLR4 signaling (Lian et al. 2010). This anti-inflammatoryeffect may be responsible for the hepatoprotective effect oflimonin.
Table 3 Effect of D-galactosamine (D-GalN, 10 μM), limonin (5 and 10 μM) and silymarin (10 μM) on liver enzymes (ALT, AST activities),malondialdehyde (MDA), superoxide dismutase (SOD) and total antioxidant capacity (pg/mg tissue) in cultured human hepatocytes
Parameters Control D-GalN Limonin (5 μM) Limonin (10 μM) Silymarin (10 μM)
ALT (U/mL) 10.07±1.01 36.70*±1.75 25.47@#±1.12 16.73@±0.84 13.20@±1.07
AST (U/mL) 7.33±0.61 25.15*±0.75 17.45@#±1.21 10.78@±0.80 9.17@±0.79
MDA ((nmol/mL) 11.35±1.41 25.89*±0.27 16.60@±0.075 13.76@±0.41 13.31@±0.51
SOD (U/mL) 391.1±11.15 196.4*±28.1 280.4@±19.88 351.8@±11.70 380.4@±8.18
TAC (mmol/L) 0.82±0.017 0.12*±0.012 0.66@±0.035 0.78@±0.01 1.19@±0.10
Results are expressed as mean±SE. (*) Significant difference compared to control group, (@) significant difference compared to D-GalN-treated groupand (#) significant difference from limonin 10 at P<0.05. n =3; by one-way ANOVA and Tukey post hoc test
Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261 259

In the present work, we found increased oxidativestress in animals treated with D-GalN which may berelated to up regulation of TLR4 or direct effect of D-GalN. The first mechanism is confirmed by previousreport of Tsung et al. (2007) who suggested thatTLR4 induces ROS production in liver cells. In thecurrent study, we observed an increase in lipid peroxi-dation and depletion of GSH in liver of D-GalN-treatedrats as compared to control. On the other hand, limoninin a dose of 50 mg/kg and silymarin significantlyinhibited lipid peroxidation and restored the depletedGSH levels. In a previous report, it was observed thatlimonin had similar effects on liver of aflatoxin-treatedrats (Langeswaran et al. 2012). The effect of limonin onGSH may be attributed to limonin-induced increases in glu-tathione transferase activity (GST) which elevates GSH con-tent in different tissues including liver homogenates (Perezet al. 2009). Our findings showed that the effect of limoninon GSH was dose dependent. The lower dose (50 mg/kg)being more effective (restored depleted GSH) than thehigher dose (100 mg/kg). Further studies are required toinvestigate the effect of different doses of limonin on GSTactivity. Furthermore, our study showed direct antioxidanteffect of limonin when compared with rutin in vitro whichwas concentration-dependent effect.
The second mechanism was confirmed by the increase ofMDA concentration and the reduction of both TAC and SODactivity in cultured human hepatocytes after incubation withD-GalN. Pretreatment of cultured human hepatocytes withlimonin at 5 and 10 μM significantly reduced MDA leveland increased both TAC and SOD activity in a concentration-related manner, compared to cells treated with D-GalN alone.The effect of limonin (10 μM) was comparable to that of thereference standard silymarin. These findings confirmed theobtained in vivo results.
Thus, it can be concluded that the C. aurantium var.bigardia derived limonin was effective as a hepatoprotectiveagent in D-GalN-induced hepatic injury, probably mediatedvia direct and indirect protective mechanisms. The directprotective mechanism is through antioxidant effects, whilethe indirect mechanism is through the inhibition of TNF-αproduction and subsequent suppression of eosinophils infil-tration in liver of D-GalN-treated rats. Our results also supportthat C. aurantium derived limonin has a more potent thera-peutic potential for protection against acute hepatic injury thanS. marianum derived silymarin.
Acknowledgments This work was funded by Zagazig University, pro-ject 5K2/M4 (to AE). The authors would like to thank Prof. L. Rashed(Biochemistry Department, Faculty of Medicine, Cairo University,Egypt) for assistance in performing PCR technique. Dr. A. Ahmed,Faculty of Veterinary Medicine, Zagazig University for histopathologicalexamination of liver and A. Tahrani, Institute of Pharmacy andMolecularBiotechnology, Heidelberg University, Germany for running LC-MS.
Conflict of interest The authors declare no conflict of interest.
References
Berzsenyi MD, Roberts SK, Preiss S, Woollard DJ, Beard MR, SkinnerNA, Bowden DS, Visvanathan K (2011) Hepatic TLR2 and TLR4expression correlates with hepatic inflammation and TNF-α in HCV& HCV/HIV infection. J Viral Hepat 18:852–860
Chaung SS, Lin CC, Lin J, Yu KH, Hsu YF, Yen MH (2003) Thehepatoprotective effects of Limonium sinense against carbon tetra-chloride and beta-d-galactosamine intoxication in rats. PhytotherRes 17:784–791
Cotroneo TM, Nemzek-Hamlin JA, Bayliss J, Su GL (2012)Lipopolysaccharide binding protein inhibitory peptide alters hepaticinflammatory response post-hemorrhagic shock. Innate Immun 18:866–875
Decker K, Keppler D (1972) Galactosamine induced liver injury. In:Popper H, Schaffner F (Eds) Progress in liver disseases. Grune &Stratton, New York 4: 183–199
Dolganiuc A, Oak S, Kodys K, Golenbock DT, Finberg RW, Kurt-JonesE, Szabo G (2004) Hepatitis C core and nonstructural 3 proteinstrigger toll-like receptor 2-mediated pathways and inflammatoryactivation. Gastroenterology 127:1513–1524
Duesberg U, von dem Bussche A, Kirschning C, Miyake K, SauerbruchT, Spengler U (2002) Cell activation by synthetic lipopeptides of thehepatitis C virus (HCV)-core protein is mediated by toll like recep-tors (TLRs) 2 and 4. Immunol Lett 8:89–95
Ellman GL (1959) Tissue sulfhydryl groups. Arch Biochem Biophys 82:70–77
El-Mofty SK, Scrutton MC, Serroni A, Nicolini C, Farber JL (1975)Early reversible plasma membrane injury in galactosamine-inducedliver cell death. Am J Pathol 79:579–596
El-Readi MZ, Hamdan D, Farrag N, El-Shazly A, Wink M (2010)Inhibition of P-glycoprotein activity by limonin and other secondarymetabolites from Citrus species in human colon and leukaemia celllines. Eur J Pharmacol 626:139–145
Fan H, Li L, Zhang X, Liu Y, Yang C, Yang Y, Yin J (2009) Oxymatrinedown regulates TLR4, TLR2, MyD88, and NF-kappa B and pro-tects rat brains against focal ischemia. Mediators Inflamm 2009:704706. doi:10.1155/2009/704706
Germanò MP, D’Angelo V, Sanogo R, Catania S, Alma R, De PasqualeR, Bisignano G (2005) Hepatoprotective and antibacterial effects ofextracts from Trichil ia emetica Vahl. (Meliaceae). JEthnopharmacol 96:227–232
Hamdan D, El-Readi MZ, Tahrania A, Herrmann F, Kaufmann D, FarragN, El-Shazly A, Wink M (2011a) Secondary metabolites ofPonderosa Lemon (Citrus pyriformis) and their antioxidant, anti-inflammatory, and cytotoxic activities. Z Naturforsch C 66:385–393
Hamdan D, El-Readi M, Tahrani A, Herrmann F, Kaufmann D, Farrag N,El-Shazly A,WinkM (2011b) Chemical composition and biologicalactivity of Citrus jambhiri Lush. Food Chem 127:394–403
Hartmann P, Haimerl M, Mazagova M, Brenner DA, Schnabl B (2012)Toll-like receptor 2-mediated intestinal injury and enteric tumornecrosis factor receptor I contribute to liver fibrosis in mice.Gastroenterology 143:1330–1340
Huebener P, Schwabe RF (2013) Regulation of wound healing and organfibrosis by toll-like receptors. Biochim Biophys Acta 1832:1005–1017
Iwata H, Tezuka Y, Kadota S, Hiratsuka A,Watabe T (2005) Mechanism-based inactivation of human liver microsomal CYP3A4 byrutaecarpine and limonin from Evodia fruit extract. Drug MetabPharmacokinet 20:34–45
Keppler DO, Rudigier JF, Bischoff E, Decker KF (1970) The trapping ofuridine phosphates by D-galactosamine. D-glucosamine and 2-
260 Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261

deoxy-D-galactose. A study on the mechanism of galactosaminehepatitis. Eur J Biochem 17:246–253
Kitazawa T, Tsujimoto T, Kawaratani H, Fujimoto M, Fukui H (2008)Expression of Toll-like receptor 4 in various organs in rats with D-galactosamine-induced acute hepatic failure. J GastroenterolHepatol 23:e494–e498
Kitazawa T, Tsujimoto T, Kawaratani H, Fukui H (2010) Salvage effect ofE5564, Toll-like receptor 4 antagonist on d-galactosamine andlipopolysaccharide-induced acute liver failure in rats. JGastroenterol Hepatol 25:1009–1012
Kolwankar D, Vuppalanchi R, Ethell B, Jones DR, Wrighton SA, HallSD, Chalasani N (2007) Association between nonalcoholic hepaticsteatosis and hepatic cytochrome P-450 3A activity. ClinGastroenterol Hepatol 5:388–393
Lam LKT, Zang J, Hasegawa S (1994) Citrus limonoid reduction ofchemically induced tumorigenesis. Food Technol 48:104–108
Langeswaran K, Gowtham Kumar S, Revathy R, Balasubramanian MP(2012) Attenuation of aflatoxin B1 induced primaryhepatocarcinogenesis by a citrus bioactive compound-limonin. JPharm Res 5:177–182
Lian LH, Jin X,Wu YL, Cai XF, Lee JJ, Nan JX (2010) Hepatoprotectiveeffects of Sedum sarmentosum on D-galactosamine/lipopolysaccha-ride-induced murine fulminant hepatic failure. J Pharmacol Sci 114:147–157
Manners GD (2007) Citrus limonoids: analysis, bioactivity, and biomed-ical prospects. J Agric Food Chem 55:8285–8294
Manners GD, Hasegawa S (1999) A new normal phase liquid chromato-graphic method for the analysis of limonoids in citrus. PhytochemAnal 10:76–81
Manners GD, Breksa AP 3rd, Schoch TK, Hidalgo MB (2003) Analysisof bitter limonoids in citrus juices by atmospheric pressure chemicalionization and electrospray ionization liquid chromatography - massspectrometry. J Agric Food Chem 51:3709–3714
Maskos K, Fernandez-Catalan C, Huber R, Bourenkov GP, Bartunik H,Ellestad GA, Reddy P, Wolfson MF, Rauch CT, Castner BJ, DavisR, Clarke HRG, Petersen M, Fitzner JN, Cerretti DP, March CJ,Paxton RJ, Black RA, Bode W (1998) Crystal structure of thecatalytic domain of human tumor necrosis factor-alpha-convertingenzyme. Proc Natl Acad Sci U S A 95:3408–3412
Miller EG, Gonzales-Sanders AP, Couvillon AM, Binnie WH, HasegawaS, Lam LK (1992) Inhibition of hamster buccal pouch carcinogen-esis by limonin 17-beta-D-glucopyranoside. Nutr Cancer 17:1–7
Nakagiri R, Hashizume E, Kavahashi S, Sakai Y, Kamiya T (2003)Suppression by Hydrangeae dulcis folium of d-galactosamine-induced liver injury in vitro and in vivo . Biosci BiotechnolBiochem 67:2641–2643
Perez JL, Jayaprakasha GK, Valdivia V, Munoz D, Dandekar DV, AhmadH, Patil BS (2009) Limonin methoxylation influences the inductionof glutathione S-transferase and quinone reductase. J Agric FoodChem 57:5279–5286
Piyachafurawat P, Glinsukon T, Chanjarunee A (1985) Antifertility effectof Citrus hystrix DC. J Ethnopharmacol 13:105–110
Pradhan SC, Girish C (2006) Hepatoprotective herbal drug, silymarinfrom experimental pharmacology to clinical medicine. Indian J MedRes 124:491–504
Roy A, Saraf S (2006) Limonoids: overview of significant bioactivetriterpenes distributed in plant kingdom. Biol Pharm Bull 29:191–201
Seki E, De Minicis S, Osterreicher CH, Kluwe J, Osawa Y, Brenner DA,Schwabe RF (2007) TLR4 enhances TGF-beta signaling and hepat-ic fibrosis. Nat Med 13:1324–1332
Sheik Abdulazeez S, Thiruvengadam D (2013) Effect of lycopeneon oxidative stress induced during d-galactosamine/lipopoly-saccharide-sensitized liver injury in rats. Pharm Biol 51:1592–1599
Soares JB, Pimentel-Nunes P, Afonso L, Rolanda C, Lopes P, Roncon-AlbuquerqueR Jr, Gonçalves N, Boal-Carvalho I, Pardal F, Lopes S,Macedo G, Lara-Santos L, Henrique R, Moreira-Dias L, GonçalvesR, Dinis-Ribeiro M, Leite-Moreira AF (2012) Increased hepaticexpression of TLR2 and TLR4 in the hepatic inflammation-fibrosis-carcinoma sequence. Innate Immun 18:700–708
Takayashiki T, Yoshidome H, Kimura F, Ohtsuka M, Shimizu Y, Kato A(2004) Increased expression of Toll-like receptor 4 enhancesendotoxin-induced hepatic failure in partially hepatectomized mice.J Hepatol 41:621–628
Takeuchi O, Akira S (2010) Pattern recognition receptors and inflamma-tion. Cell 140:805–820
Tian Q, Schwartz SJ (2003) Mass spectrometry and tandem mass spec-trometry of Citrus limonoids. Anal Chem 75:5451–5460
Tsung A, Klune JR, Zhang X, Jeyabalan G, Cao Z, Peng X, Stolz DB,Geller DA, Rosengart MR, Billiar TR (2007) HMGB1 releaseinduced by liver ischemia involves Toll-like receptor 4 dependentreactive oxygen species production and calcium-mediated signaling.J Exp Med 204:2913–2923
Tu CT, Han B, Yao QY, Zhang YA, Liu HC, Zhang SC (2012) Curcuminattenuates concanavalin A-induced liver injury in mice by inhibitionof Toll-like receptor (TLR) 2, TLR4 and TLR9 expression. IntImmunopharmacol 12:151–157
Wang XJ, Wang ZB, Xu JX (2005) Effect of salvianic acid A on lipidperoxidation and membrane permeability in mitochondria. JEthnopharmacol 97:441–445
WHO (1999) Hepatitis C-global prevalence (update). Wkly EpidemiolRec 74:425–427
Wills ED (1987) Evaluation of lipid peroxidation in lipids and biologicalmembranes. In: Snell K,Mullock B (eds) Biochemical toxicology: apractical approach. IRL Press, Oxford, pp 127–152
Yang L, Seki E (2012) Toll-like receptors in liver fibrosis: cellularcrosstalk and mechanisms. Front Physiol 3:138. doi:10.3389/fphys.2012.00138
Yoo YM, Nam JH, Kim MY, Choi J, Park HJ (2008) Pectolinarin andpectolinarigenin of Cirsium setidens prevent the hepatic injury inrats caused by D-galactosamine via an antioxidant mechanism. BiolPharm Bull 31:760–764
Yu J, Wang L, Walzem RL, Miller ED, Pike LM, Patil BS (2005)Antioxidant activity of citrus limonoids, flavonoids, and coumarins.J Agric Food Chem 53:2005–2014
Zheng S, Sugita S, Hirai S, Egashira Y (2012) Protective effect of lowmolecular fraction of MGN-3, a modified arabinoxylan from ricebran, on acute liver injury by inhibition of NF-κB and JNK/MAPKexpression. Int Immunopharmacol 14:764–769
Naunyn-Schmiedeberg's Arch Pharmacol (2014) 387:251–261 261
Copyright © 2022 FDOKUMEN




















