
Hemorrhagic, fibrinogenolytic and edema-forming activities of the venom of the colubrid snake...
12
Toslcon Vol . 30, No. 4, pp. 42738, 1992. 0041-0101/92 f5 .00+ .00 Printed in Great Britain. ® 1992 Pergrmoo Prey Ltd HEMORRHAGIC, FIBRINOGENOLYTIC AND EDEMA- FORMING ACTIVITIES OF THE VENOM OF THE COLUBRID SNAKE PHILODRYAS OLFERSII (GREEN SNAKE) MARINA T . ASSAKURA, I MARiA DA GRAÇA SALO11Iw0,2 GIUSEPPE PUORT0 2 and FAIGA R . MANDELBAUM I 'Serviço de Bioquimica, and ZSeçso de Herpetologia, Instituto Butantan, Sâo Paulo, S .P ., Brazil (Received 8 August 1991 ; accepted 3 December 1991) M . T . ASSAKURA, M . DA GRAçA $ALOMÂO, G . PUORTO and F . R . MANDELBAUM . Hemorchagic, fibrinogenolytic and edema-forming activities of the venom of the colubrid snake Philodryas olfersü (green snake) . Toxicon 30, 42738, 1992 .-The venom of P. olfersü has high hemorchagic, edema- inducing and fibrin(ogen)olytic activities . It is devoid of thrombin-like, procoagulant, phospholipase A 2 and platelet aggregating enzymes. The main activities are metalloproteinases inhibited by metal chelators (EDTA and 1,10-phenanthroline) and sulfhydryl compounds (DTT and cysteine) . The hemorchagic and fibrinogenolytic enzymes were partially purified by gel filtration on HPLC . The hemorchagic activity of the venom was neutralized by commercial horse antivenoms to Bothrops species, as well as by rabbit antisera specific for hemorchagic factors isolated from these Bothrops venoms . No immunoprecipitin reactions were obtained, indicating that the few epitopes of the P . olfersü hemorchagin are involved in these neutralization reactions. The fibrinogenolytic enzyme cleaves A a-chain more quickly than the B ß-chain of human fibrinogen . The venom also solubilizes fibrin . This solubilization appears to occur from the hydrolysis of unpolymerized a~hain and cross- linked y-y dimer. The fibrin peptide products are distinct from those produced by plasmin . INTRODUCTiON REPORTS On toxic saliva in colubrid snakes are few and the chances of being bitten by colubrid snakes are rare, because of its non-aggressive behavior and the anatomical disadvantage of having fangs situated behind its other teeth . Nevertheless, some species have caused severe reactions in humans. Paralysis, respiratory failure, hemorchage and death were observed . The venom of the African boomslang (Dispholidus typos) and the bird snake (Thelotornis kirtlandi) have proven lethal to man (MACKAY et al., 1969; VLSSER and CHAPMAN, 1978). Also, it is worthwhile to point out that non-clotting blood and hemorchage, usually associated with viperid envenomations, are the most striking clinical Author to whom correspondcnce should be addressed . 42T
Transcript of Hemorrhagic, fibrinogenolytic and edema-forming activities of the venom of the colubrid snake...

Toslcon Vol . 30, No. 4, pp. 42738, 1992.
0041-0101/92 f5 .00+ .00Printed in Great Britain.
® 1992 Pergrmoo Prey Ltd
HEMORRHAGIC, FIBRINOGENOLYTIC AND EDEMA-FORMING ACTIVITIES OF THE VENOM OF THE COLUBRID
SNAKE PHILODRYAS OLFERSII (GREEN SNAKE)
MARINA T. ASSAKURA, I MARiA DA GRAÇA SALO11Iw0,2 GIUSEPPE PUORT02 and FAIGA R.MANDELBAUMI
'Serviço de Bioquimica, and ZSeçso de Herpetologia, Instituto Butantan, Sâo Paulo, S.P ., Brazil
(Received 8 August 1991 ; accepted 3 December 1991)
M . T . ASSAKURA, M. DA GRAçA $ALOMÂO, G. PUORTO and F. R .MANDELBAUM . Hemorchagic, fibrinogenolytic and edema-forming activities ofthe venom of the colubrid snake Philodryas olfersü (green snake) . Toxicon 30,42738, 1992.-The venom of P. olfersü has high hemorchagic, edema-inducing and fibrin(ogen)olytic activities . It is devoid of thrombin-like,procoagulant, phospholipase A2 and platelet aggregating enzymes. The mainactivities are metalloproteinases inhibited by metal chelators (EDTA and1,10-phenanthroline) and sulfhydryl compounds (DTT and cysteine). Thehemorchagic and fibrinogenolytic enzymes were partially purified by gelfiltration on HPLC. The hemorchagic activity of the venom was neutralized bycommercial horse antivenoms to Bothrops species, as well as by rabbit antiseraspecific for hemorchagic factors isolated from these Bothrops venoms . Noimmunoprecipitin reactions were obtained, indicating that the few epitopes ofthe P. olfersü hemorchagin are involved in these neutralization reactions. Thefibrinogenolytic enzyme cleaves A a-chain more quickly than the B ß-chain ofhuman fibrinogen . The venom also solubilizes fibrin . This solubilizationappears to occur from the hydrolysis of unpolymerized a~hain and cross-linked y-y dimer. The fibrin peptide products are distinct from those producedby plasmin.
INTRODUCTiON
REPORTS On toxic saliva in colubrid snakes are few and the chances of being bitten bycolubrid snakes are rare, because of its non-aggressive behavior and the anatomicaldisadvantage of having fangs situated behind its other teeth. Nevertheless, some specieshave caused severe reactions in humans. Paralysis, respiratory failure, hemorchage anddeath were observed . The venom of the African boomslang (Dispholidus typos) and thebird snake (Thelotornis kirtlandi) have proven lethal to man (MACKAY et al., 1969; VLSSERand CHAPMAN, 1978). Also, it is worthwhile to point out that non-clotting blood andhemorchage, usually associated with viperid envenomations, are the most striking clinical
Author to whom correspondcnce should be addressed .
42T

428
M. T. ASSAKURA et al.
manifestations of colubrid envenomations (VEST, 1978 ; VISSER and CHAPMAN, 1978 ; SILVAand BUONONATO, 1983/84) .While there is active research on venom constituents of snakes belonging to the families
of the Elapidae and Viperidae, because of the small quantity of secretion of theDuvernoy's gland, little has been done to elucidate the composition and biologicalactivities of colubrid venoms. Some biochemical and pharmacological studies on thesevenoms have been published (NAH~S et al., 1976; FERLAN et al., 1983 ; SnKnl et al., 1984;SAKAI and SAWAI, 1984; Snwnl et al ., 1985 ; VEST, 1978). Only a few components,principally coagulant and hemorrhagic, were isolated (ICORNALIK et al ., 1978 ; HIESTANDand HIESTAND, 1979 ; Ovnnln, 1987 ; MORrrA et al., 1988) .
Philodryas olfersü, a Colubridae-Xenodontinae, is an opisthoglyphous snake with awell-developed Duvernoy's gland connected with a grooved tooth. This snake is found inSouth America, from Amazonas to Patagonia . Accidents with this snake are rare and notlethal . Envenomations, however, are usually painful followed by edema and hemorrhageof the limb (M~RTnvs, Ph.D . Thesis, Sâo Paulo, 1916 ; NICKERSON and HENDERSON, 1976 ;Sn.v~ and BuoxoxnTO, 1983/84). In this report, the composition and biological activitiesof this venom were studied . Two principal components were partially isolated and theirproperties determined .
MATERIALS AND METHODS
Venom of Philodryas olfersüVenom was extracted from adult snakes measuring 90-120 crn. The snakes were milked by introducing a
100-p1 micropipet over each fang following the procedure of FFrtuN et al. (1983) . The samples were kept in afrozen state at -20°C . For studies, the venom was diluted with 0.9% NaCI or 0.05 M Tris-HCl buffer (pH 7 .5) .
Estimation of proteinProtein was measured by the method of LowxY et al. (1951) using bovine serum albumin as standard, or by
spectrophotometric absorption at 280 nm. An absorbance of I .0 at 280 nm of the venom solution correspondedto 1 mg/ml of protein.
Hemorrhagic activityHemorrhagic activity was quantitatively determined by the skin method of Koxno et al. (1960). Test solutions
(50 pl) were injected into the skin of the back of mice . The skins were removed 2 hr Iater and the diameters of thehemorchagic spots were measured on the inside surfaces. The minimum amount of protein that produced ahemorrhagic spot of 10 mm= was taken as one minimum hemorchagic dose (MHD).
Neutralization of the hemorrhagic activity by immune seraTo 10 !rg of venom in 125 pl of 0.9% NaCI (corresponding to 20 MHD) was added 25 pl of antiserum or
antiserum diluted I : 2 or 1 : 4, and 100 pl of 0.9% NaCI . After 2 hr at room temperature, aliquots of 50 p1 ofthese mixtures containing 2 lrg of venom (4 MHD) were injected intradermally into the backs of mice (fouranimals for each mixture) . In these tests we used horse antisera (anti-Bothrops jararaca, anti-Bothrops neuwiedi,anti-Bothrops moojent) produced by the Hutantan Institute . Rabbit specific antisera for the hemorchagic factorsHF Z and NHF, .b, isolated from the venoms of Bothrops jararaca and Bothrops neuwiedi, respectively, were alsoused (M~rmer.s~uM and Ass~ua~, 1988). To estimate the perroentage of inhibition, these hemorchages werecompared to those of control animals which received l, 2 or 4 MHD without antiserum .
Caseinolytic activityActivity was estimated using casein by a modification of the method of KuNrrz (1946) as described by
M~xner.H~uM et al. (1982). One unit is defined as the amount of enzyme yielding an increase in absorbance of!.0 per min at 750 nar. Specific activity is expressed as units/mg protein .

Philodryas oljersit Venom Compontnts
429
Bloo~clotting activityActivity was measured on citrated horse plasma [1 ml of trisodium citrate 3.8% (w/v) to 10 ml of blood] or
human fibrinogen (4 mg/ml) in 0.9% NaCI . To 200 pl of plasma or fibrinogen were added aliquots of 50 lrl ofvenom solution or HPLC fractions, and the clotting time was measured at 37°C . The delay of the clotting time ofthrombin was tested with human fibrinogen previously incubated at 37°C with the venom or HPLC fractions atvarious concentrations . After 3 min of incubation, 50 pl of thrombin (10 NIH units/ml) was added and theclotting time recorded .
Phospholipase A2 activityPhosphatide-aryl-hydrolaxe was tested using egg-yolk lecithin as substrate, as described by KORNALIK and
M~stEre (1964) . Lysolecithin released as a result of enzyme activity was assayed by means of hemolytic effect onwashed horse red blood cells . The phospholipase AZ activity was expressed as pg/ml of the sample causing 50%hemolytis .
Polyacrylamtde gel (basic and acidic) electrophoresisPolyacryhunide 10% slab gel electrophoresis in Tris-glycine buffer at pH 8.5 was performed according to the
method of M~rzet. (1969) with a 2.5% gel spacer, following the method of Dw~s (1964) . Runs were of 5 hr with20 mA, and bromophenol blue was used as an indicator. Polyacrylamide gel electrophoresis at pH 4.3 wasperformed with 7.5% gel and 1.25% gel spacer, following the method described by RnsWet.n et al. (1962) . Runswere of 90 min with I25 mA . Methyl green was used as a tracking dye.
SDSpolyacrylamide gel electrophoresisElectrophoresis was performed on 5-15, 5-20 or 10-20% gradient polyacrylamide slab gels following the
method of L~~w (1970) using a 2.5% gel spacer according to the method of Dwts (1964) . Runs were of4-5 hr with 20 mA. Bromophenol blue was used as an indicator.
Speck hydrolysis offibrinogen andfibrinSpecific cleavage of fibrinogen and fibrin by the venom was shown by electrophoresis on 5-20%
polyacrylamide gels. Seven hundred microliterx of human fibrinogen (KabiVitrum, Sweden) at a concentrationof 4 mg/ml in 0.9% NaCI, wax incubated with 56 pg of venom in l70 pl of 0.1 M Tris-HCl buffer (pH 8.0),corresponding to a ratio of 50 : 1 (w/w). At various time intervals, aliquots of 20pl were withdrawn from thedigestion mixture and the reaction was stopped by addition of 20 pl of denaturing solution (2% SDS and 2%2-mercaptoethanol). The samples were denatured and reduced by boiling for 5 min prior the run on SDS-polyac-rylamide gel . Fibrin was prepared following the procedure ofWtr.ras and Tu (1988). Philodryas olfersit venom orplasmin was added to the clot and incubated at 37°C for various time intervals .When activity wax determined in the presence of inhibitors, the venom was incubated, for 2 hr at room
temperature, with the inhibitor prior to the addition of fibrinogen or fibrin . The ratio of the substrate: venomwas 50 : 1 . The reaction was stopped by boiling with denaturing solution, as was done with fibrinogen control,and run on SDS-polyacrylamide gel .
Edemaforming activity (Ytiucew~ et al ., 1976)Mice weighing 18-20 g (groups of siz) were injected in the right foot-pads with 25 pl doses ofthree dilutions of
the venom. For controls, left pads were injected with 25 pl of 0.9% NaCI. Throe hours after injection the micewere killed by ether inhalation. The pads were removed with a scalpel and weighed. The minimum edema dose(MED) was defined as the least quantity ofvenom causing 30% increase in the weight, compared to the control .
Assay on arginine synthetic substratesHydrolyxis of a-N-tosyl-rrarginine methyl ester (TAME) was determined at room temperature by the pH-stat
in a Radiometer titrator TTT-1 . Venom samples were added to a 3.0 ml solution of 20 mM substrate in 0.1 MKCl at pH 8.5 and titrated with 50 mM NaOH . The specific esterase activity was recorded as lrmoles/TAMEhydrolyzed/min/mg protein .
Amidolytic activity on acetyl-t.-phenylalanyl-t.-arginine-p-NA (Ao-Phe-Arg-pNA) was measured at 37°C in abuffer containing 50 pl Of 0.1 M Tris-HCl (pH 8.0), 0.02 M CaCI=, 20-50 pl of venom samples, 0.15 M NaCI(added up to a ßnal volume of 140 pl) and 10 lrl of 5mM chromogenic substrate. The amount ofp-nitroanilineformed was determined photometrically (e,~ =8900 M'' cm'') The specific amidolytic activity was defined asmoles ofp-nitroaniline released/min/mg protein .

43 0
M. T. ASSAKURA et al.
Platelet aggregating measurementHuman platelet rich plasma (PRP) was prepared by collecting blood in I/l0 volume of 0 .1 M sodium citrate,
and centrifuged at 250 g for 4 min at room temperature . The aggregation was measured photometricallyfollowing the procedure of BoxN and CROSS (1963), using a 'Payton Scientific' Minigator II, linked to a `Linear'registrator . In the tests we used 350 pl of PRP preparation, 100 pl of venom fraction and 50 ~1 of ADP (finalconcentration l-3 pM) .
PuriJicatian of venom prateinvVenom (200 hg) in 100 ~1 of0.05 M Tris-HCl buffer pH 7.0 containing O.15 M NaCI, t mM CxCI Z and 0.05%
NaN, was loaded on to a TSK-3000 SW (7 .5 mm x 30 cm x 2) column, previously equilibrated with the samebuffer solution . The flow rate was 0.5 ml/min, and the proteins were detected at 280 nm . The peaks of proteinwere collected, analyzed on SDS-PAGE and assayed on several substrates .
A
RESULTS
The protein composition of the venom of P. olfersü, as analyzed by electrophoresis onpolyacrylamide gel, at pH 8.5, showed that the components with low electrophoresicmobility were relatively poorly separated (Fig . lA) . This could be attributed to thepresence of mucous material and lipids that hindered the mobility of these proteins .Polyacrylamide gel electrophoresis at pH 4.3 also gave a poor resolution . A cluster of
/68,000-50,000~43,000
23,000
13,700
F1G I . PROTEIN BANDS OF PllilOClryaS OIjerSi! VENOM ON POLYACRYLAMIUE GEL (A) ANDSDS-POLYACRYLAMIDE (IO-2O% GRADIENT) ELECTROPHOREST:S (B).
(A) crude venom 25 l+g, pH 8 .5 ; (B) crude venom (25 hg) after denaturation with SDS andreduction with 2-mercaptcethanol ; (C) mixture of denatured and reduced bovine serum albumin(68,000), ovalbumin (43,000), human y-globulin (H- and L~hains, 50,000 and 23,000, respectively)and ribonuclease (13,700), as standards for mol . wts . Electrophoresis was carried out as described
in Materials and Methods .

Philodryas olferaü Venom Components
431
FIG . Z . HEMORRHAGIC ACTIVITY OF PhiIOdryas Olfersü VENOM AND THE EFFECT OF INHIBITORS.(A) Venom solutions (2, 1 and 0.5 l+g in 50 pl of 0.9% NaCI) were injected in mice. (B) Venom waspreviously incubated with EDTA, cysteine or I,10 phenanthroline . Two hours later, the mice wereinjected with 50 ~1 aliquots containing 2 l+g (4 MHD) of venom and 1 mM inhibitor . For control
50 ~1 of 0.9% NaCI was injected .
protein bands ran to the cathode. A good separation was only achieved on SDS-PAGEwith the venom denatured by SDS and reduced by 2-mercaptoethanol . In this conditionthe venom showed a simple compostion, four major protein bands of mol. wts between20,000 and 58,000, and three minor components, one of 70,000 and the other two near10,000 (Fig . 1 B) .The hemorrhagic activity of the venom, as tested in mice using a serial dilution in 0.9%
NaCI, gave aminimum hemorrhagic dose (MHD) of0.5 pg . The proteolytic activity of thevenom, assayed on casein, was 4.47 units/mg protein. These two activities were inhibitedby metal chelating compounds, such as EDTA and 1,10-phenanthroline . Changing thepHof the venom solution from neutral to acidic decreased both activities . At pH 6.0 a 25%loss of activity was observed, and at pH lower than 4.0 the loss of activity was 100% .Theactivity lost, either by metal chelation or by an acidic solution, was not recovered bydialysis, addition of CaZ+ or neutralization . While 1 mM DTT inhibited completely bothhemorrhagic and caseinolytic activities, the same concentration of cysteine was able toabolish only the hemorrhagic activity . The proteolytic activity on casein was almost thesame and 1 mM cysteine reduced it only 17% . These data show no parallelism betweenhemorrhagic and caseinolytic activities, suggesting that besides the hemorrhagic com-ponent, the venom has other proteinases active on casein . On the other hand, thehemorrhagic proteinase might have low activity on this substrate and, in contrast to theother metalloproteinases of the venom, be easily inhibited by cysteine . The hemorrhagicactivity of the venom and its inhibition in the presence of 1 mM EDTA, 1 mM1,10-phenanthroline and 1 mM cysteine are shown in Fig. 2. The high content ofmetalloproteinases (hemorrhagic and non-hemorrhagic) in the venom of P. olfersü, is anindication of some similarity, although remote, to the venoms of the Crotalinae and

432 M . T . ASSAKURA et al.
T~erE 1 . H~roruutwarc ecrtvrr~r oP P. olfersü vexotK Nevrtut.rz® aYx~oxse sEw+ exn-Bothrops vwo~ erm xeasrr s~crntc ~ ~rrnHE~rORRHAßrC FACI~ORS (NHF, b pot~rl~ rno~t Bothrops neuwiedi ~rtn
HFz rsora~ xttoa Bothrops jararaca)
Equal volumes of venom in 0.9'/. NaCI and antisera diluted in the samesaline were mixed 2 hr before injections. The tests were performed asdescribed in Materials and Methods. The data are mean values of fourmice. For values under 100% neutralvation the S.D . values were f 10-15%. Not tested (-).
Viperinae subfamilies . Because of the possible similarities of this colubrid venom tocrotalid venoms we compared it to venoms of the Bothrops species. We were interestedmainly in the hemorrhagec principle of this venom in order to determine whether it hasany analogy with other snake hemorrhagins . Initially, we tested the ability of thecommercial horse antivenoms from the Butantan Institute to cross-react with the venomof P. oifersü. In these assays we used antivenoms to B. jararaca, B . neuwiedi andB. moojeni. Immuncelectrophoresis showed the presence of several common antigens .While anti-B. moojeni gave four immunoprecipitin lines, the others were each only able torecognize two components. However, when tested with rabbit antisera specific forhemorrhagec factors isolated from these venoms no cross-reaction was observed .
Ouchterlony double diffusion showed no immunoprecipitin line, either with anti-NHF,,bprepared to the factor NHF, .b isolated from B. neuwiedi, or with anti-HFZ, prepared to thefactor HFZ isolated from the venom of B. jararaca (MANDELBAUM and ASSARURA, 1988) .
Although the cross-reactivity was poor or negative, these antisera were tested in orderto determine whether they would neutralize the hemorrhagec activity of this venom. Theneutralization reactions were assayed in the mouse and estimated as percentage of thehemorrhages observed in the control animals, which received the same amount of venomwithout serum (Table 1) . Horse and rabbit normal sera, used as controls, did notneutralize the venom. These tests showed that the antivenoms were able to abolishcompletely or reduce significantly the hemorrhages elicited by 4MHD of the venom. Therabbit antisera specific for hemorrhagec factors isolated from the venoms of B. jararaca
Serum Dilution
NeutralizationVenom of P. olfersü2 PS 1 kg
(4 MHD) (2 MHD)
Anti-Bothrops 1 :10 100jaratwca 1 :20 80(horse) 1 :40 40
Anti-Bothrops I :10 100moojeni 1 :20 100(horse) 1 :40 70
Anti-Bothrops 1 :10 90neuwiedi 1 :20 80(horse) 1 :40 0
Anti-NHFRb 1 :2 I00 100(rabbit) 1 :5 40 100
( :10 0 0Anti-HFZ I : 2 100 100
(rabbit) 1 :5 40 1001 :10 0 0

Pkitodryas otferaü Venom Components
433
10 30 60 TOPROTEIN,M9
FIa. 3. THE DELAY IN 111E TIfllO1~IN CT.01'IßdO Tn1E OF F®RINOCIEN PREVIOUSLY INCUITATED WITHTHE VENOM OF P.OIjCrJ(( OR A PARTIALLY PUItII+® FRACTION (PEAa IIb PROSi HPLC
cxiTOMAroTSRAPHY) .Tests were performed as described in Materials and Methods . The clotting time of fibrinogen bythrombin without venom or fraction corresponded to 30 sec. Fibrinogen plus venom (~-~) or
plus peak IIb (0--0) incubated for 3 min prior to the addition of thrombin .
and B. neuwiedi failed to precipitate with the P. olfersü hemorrhagin. However, in highconcentrations they neutralized its hemorrhagc activity (Table 1) . This suggests that theP. olfersü hemorrhgin might have few epitopes similar to those of Bothrops species.These few epitopes are involved in the neutralization reactions .
Besides hemorrhage, the venom of P. olfersü induces local swelling . Compared tomouse foot pads injected with 0.9% NaCI, a 30% increase in the weight was produced byinjecting 0.25 hg of the venom. This edema-producing activity is three-fold higher than theedema caused by injecting B. moojeni venom, which among the Bothrops venoms has ahigh edematogenic activity .
Lethality was assessed by i.p . injection of various concentrations of venom in mice . TheLn~ of P. olfersü venom was 2.79f0.58 mg/kg (meantS.D.), being less potent than thevenom ofB. jararaca (mow = 1 .23 t0.08 mg,/kg) (S~LOMXo, M. G., Ph.D . Thesis, Univ. ofSao Paulo, 1991).Using egg-yolk lecithin as substrate and horse red blood cells, no phospholipase AZ
activity could be detected in the venom, even when high doses of 130 hg were used.The venom of P. olfersü does not clot citrated horse plasma or human fibrinogen, even
in 15 min. Addition of Cat+ to the plasma caused no modification of the incoagulability .These results indicate absence of thrombin-like and procoagulant enzymes in the venom.On the other hand, the clotting time of human fibrinogen by thrombin or batrozobin (athrombin-like enzyme isolated from Bothrops atrox) was delayed in the presence of thevenom of P. olfersü. This delay was proportional to the amount of venom in theincubation mixture (Fig . 3) . To confirm that the venom, without clotting, is hydrolyzingfibrinogen, it was incubated with this substrate and analyzed by SDS-gel electrophoresis
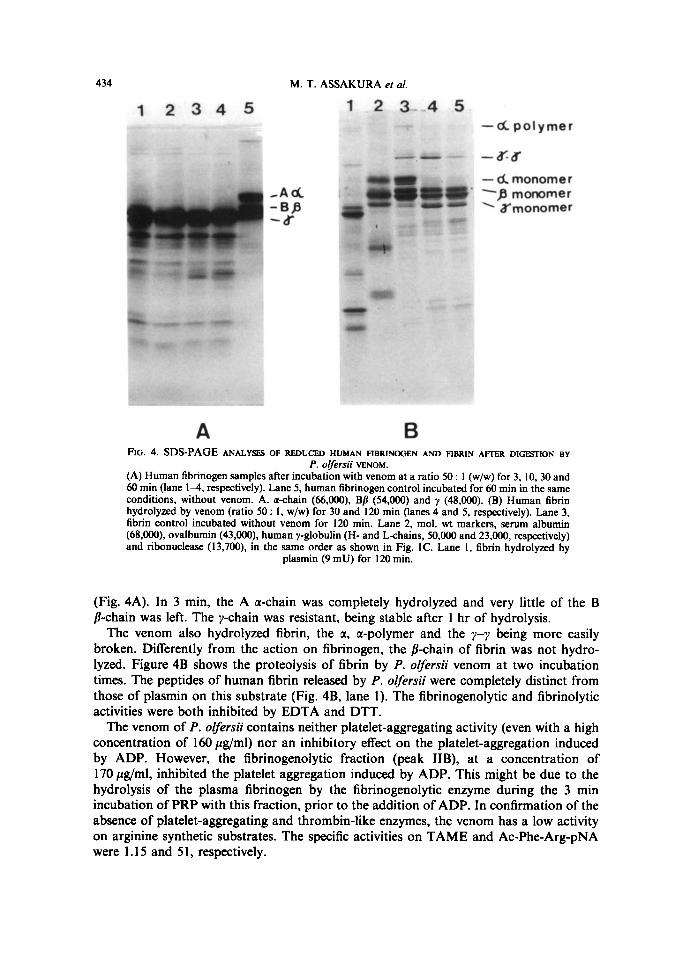
434
M. T. ASSAKURA et al.
1 2 3 4 5
.Ad-BA
d"
-d polymer
-dmonomerp monomer
~ ä"monomer
FIG. 4. SDS-PAGE ANALYSES OF REDUCED HUMAN FIBRINOGEN AND FIBRIN AFTER DIGESTION BYP. OfelS%% VENOM.
(A) Human fibrinogen samples after incubation with venom at a ratio 50 : I (w/w) for 3, 10, 30 and60 min (lane lam, respectively) . Lane 5, human fibrinogen control incubated for 60 min in the sameconditions, without venom . A. a-chain (66,000), Bß (54,000) and y (48,000). (B) Human fibrinhydrolyzed by venom (ratio 50 : l, w/w) for 30 and I20 min (lanes 4 and 5, respectively) . Lane 3,fibrin control incubated without venom for 120 min . Lane 2, mol . wt markers, serum albumin(68,000), ovalbumin (43,000), human y-globulin (H- and L~hains, 50,000 and 23,000, respectively)and ribonuclease (13,700), in the same order as shown in Fig. IC . Lane l, fibrin hydrolyzed by
plasmin (9 mU) for 120 min .
(Fig . 4A) . In 3 min, the A a-chain was completely hydrolyzed and very little of the Bß-chain was left. The y-chain was resistant, being stable after 1 hr of hydrolysis .The venom also hydrolyzed fibrin, the a, a-polymer and the y-~ being more easily
broken . Differently from the action on fibrinogen, the ß-chain of fibrin was not hydro-lyzed . Figure 4B shows the proteolysis of fibrin by P . olfersü venom at two incubationtimes . The peptides of human fibrin released by P . olfersü were completely distinct fromthose of plasmin on this substrate (Fig . 4B, lane 1) . The fibrinogenolytic and fibrinolyticactivities were both inhibited by EDTA and DTT.The venom of P. olfersü contains neither platelet-aggregating activity (even with a high
concentration of 160 iegfml) nor an inhibitory effect on the platelet-aggregation inducedby ADP. However, the fibrinogenolytic fraction (peak IIB), at a concentration of170 ~g/ml, inhibited the platelet aggregation induced by ADP. This might be due to thehydrolysis of the plasma fibrinogen by the fibrinogenolytic enzyme during the 3 minincubation of PRP with this fraction, prior to the addition of ADP. In confirmation of theabsence of platelet-aggregating and thrombin-like enzymes, the venom has a low activityon arginine synthetic substrates . The specific activities on TAME and Ac-Phe-Arg-pNAwere 1 .15 and 51, respectively .

Pkilodryas olfersii Venom Components
Attempts have been made to isolate the two principal enzymes; hemorrhagic andfibrinogenolytic . By chromatography on DEAE-cellulose DE-32 and SP-Sephadex C-50,several protein peaks were obtained, but hemorrhagic activity was almost lost by theseprocedures . However, gel filtration chromatography (HPLC) separated the venom intothree major protein peaks (Fig . SA). All the hemorrhagic and the edema-producingactivity was eluted in the first peak (RT 35 .61). The fibrinogenolytic activity was
20 40 60RETENTION TIME, min .
Fic . 5 . GEL F1LT'aAT10N OF Philodryas olfersü vPStoM wrro SDS-e~ecrtsoPxoat~ OF rm: FancnoNS.(A) HPLC gel filtration on TSK-3000 SW (7.5 mm x 30 cm x 2) . Amount applied to the column,200 hg in 100~l . Elution with 0.05 M Tris-HCl buffer pH 7,0 containing 0.15 M NaCI, 1 mMCaC12 and 0.05% NaN3 , with a $ow of 0 .5 ml/min. Detector u .v . 280 nm (0,200 AUFS) .(B) Electrophoredc pattern on SDS-PAGE (10-20% gradient) of the reduced protein fractionscollected from HPLC . Amount applied 5 to 10 l+g of protein .
43 5

436
M. T. ASSAKURA et al.
concentrated in the peak II B (RT 42.12) . The activity of this fraction was five-fold higherthan the crude venom (Fig. 3) . The caseinolytic activity was distributed between the twofirst peaks . SDS-PAGE of the HPLC fractions, after denaturation and reduction by SDSand 2-mercaptoethanol, is shown in Fig . SB . The hemorrhagic fraction, peak I, containedtwo protein bands of mol . wt 58,000 (the major protein) and 70,000 . The fibrinogenolyticfraction (peak II B) presented only a protein band of 47,000 . The third major peak, havinga mol . wt of 20,000, possessed none of the tested activities in the venom .
DISCUSSION
The venom of P . olfersü has high hemorrhagic, edematogenic, fibrinogenolytic andfibrinolytic activities . It hydrolyzes casein rapidly, whereas its activity is low on argininesynthetic substrates, such as TAME and Ac-Phe-Arg-pNA . The venom is devoid ofcoagulant, procoagulant, phospholipase A~ and platelet aggregating enzymes . The mainactivities of the venom are inhibited by metal chelating compounds (such as EDTA and1,10-phenanthroline) and sulfhydryl agents (such as cysteine and DTT) suggesting thatthese venom components are metalloproteinases and their activities rely on the integrity oftheir disulfide bridges . Like other snake venom metalloproteinases, they are labile to heatand acidic medium.MINTON arid WEINSTEIN (1987) showed that the Duvernoy's secretions of colubrid
snakes possess as complex activities as venoms of many proteroglyphous snakes . Only thenumber of proteins on electrophoresis were fewer than in most viperids, and their patternsdiffered markedly from one another. The many activities of the venom of P . olfersü andthe low number of protein bands in SDS-PAGE corroborate the findings of theseauthors .BoQuET and SnINT-GIRONS (1972) and MINTON and WEINSTEIN (1987) using antivenoms
against venoms of elapid, viperid and colubrid snakes showed that there is a widespreadsharing of venom antigens between colubrid and elapid (except the Micrurus genus)snakes and also with some vipers. Only Dispholidus typus was neither related to colubridvenoms, nor to the venoms from snakes of other families . SAKAI and SAWAI (1984)reported that antiserum prepared against the secretion of Rhabdophis t . tigrinus neutral-ized the lethal hemorrhagic and coagulant activities of its own secretion, as well as that ofR. s. subminiatus. Anti-Echis carinatus venom was also able to abolish the lethal toxicity ofR. t . tigrinus . From our results, it appears that the venom of P. olfersü shares fewcommon antigens with the Bothrops species . On the other hand, the hemorrhagic activityof P. olfersü was neutralized not only by the antivenoms against Bothrops venoms but alsoby their specific anti-hemorrhagins. The absence of immunoprecipitin lines with theBothrops anti-hemorrhagins indicates that the few recognized epitopes, which are insufiï-cient to precipitate, are involved in the neutralization reactions . Such antigenic similaritywas observed with Atractaspis engaddensis hemorrhagin and a viperid factor from Viperapalaestinae venom (OVADIA, 1987) . The P. olfersü and A . engaddensis hemorrhagins mightbe closely related considering their properties, mol wts and activities .
Coagulation is the other striking activity in the colubrid venoms . This is still acontroversial matter involving the components affecting hemostasis and fibrinolysis .MACKAY et al. (1969) demonstrated that the venom of boomslang (Dispholidus typus)produces thrombin in vitro . In disagreement with these results, HIESTAND and HIESTAND

Philodryas olfersü Venom Components 437
(1979) separated from the venom of Dispholidus typos five coagulant fractions . The maincoagulant component converted fibrinogen into fibrin and had no prothrombin activatingactivity . However, BRADLOW et al . (1980) showed that the effect ofthis venomwas mainlydue to its ability to activate prothrombin, and it was also able to activate factor X andpossibly IX, confirming therefore the results of MACKAY et al . (1969) . KoRxnLlx andTnHOIZSK~ (1978) demonstrated also that the venom of Thelotornis kirtlandi has factors IIand X activators and no thrombin-like enzyme . In the same year, KORNALIK et al. (1978)isolated from this venom only a procoagulant enzyme, which directly converted pro-thrombin to thrombin . While Nnxas et al. (1976) suggested that the venom of Rhabdophist . tigrinus had a factor X activator, the studies of SAWAI et al. (1985) led to the conclusionthat the strong coagulant activity of this venom was due to a prothrombin activator.However, without studying the hemorrhagic activity of this venom, they suggested thatthe extensive hemorrhage as a result of envenomation was promoted by the bloodcoagulant abnormality due to hypofibrinogenemia induced by the venom. RecentlyMORITA et al. (1988) isolated a prothrombin activator from this venom, confirming thefinding of Snwnl et al . (1985) .The venom of P. olfersü does not clot plasma or fibrinogen . Therefore it has neither
fibrinogenase activity which converts fibrinogen into fibrin, nor procoagulant activity,which produces thrombin . On the contrary, it degrades fibrinogen in vitro rendering ituncoagulable to thrombin . This potent fibrinogenolytic enzyme cleaves the A a-chainfaster than the B ß~hain . The venom also has a strong fibrinolytic activity. The fibrinsolubilization appears to occur primarily from the hydrolysis of the a-polymer andunpolymerized a-chain and later y-y dimers . This shows that the venom of P. olfersü, likethe other colubrid venoms has no thrombin-like enzyme . On the other hand however, thevenom is devoid of procoagulant enzymes. Moreover, it has a potent fibrinogenolyticactivity, which until now had not being described in colubrid venoms.
Acknowledgements-We are grateful t0 Neusw DE Ltstw, Mwxu Aspwxac,~w StQuirtxw and Mwnttsw nw Stt .vw forskilful technical help .
REFERENCES
HOQi1E1', P. and Swttvx-Gtttot~, H . (1972) Etude immunologique dea glandes salivaires du vestibule buccal dequelques Colubtidae opistoglyphes. Toxlcan 10, 635-644.
Haw, G . V . R. and CAOes, M. J . (1%3) The aggt+egation of the blood platelets . J. Physiol. 168, 178-195.Bxwut,ow, B . A ., A~txttvsoN, P. M., Gontretrls, E . D . and Gwlt,t .wxn, M . C. (1980) Studies on the coagulant
effect of boomshlng (Dirpholidus typos) venom. Clin. Lab. Hacmatol. 2, 317-331 .Dwvls, B . J . (1964) Disc electrophoresis-II . Method and application to human serum proteins . Ann . N. Y. Acad.
Sci. 121, 404-427 .F~tu.wN, L, Fetu.wx, A., KING, T. and Russet., F . E. (1983) Preliminary studies on the venom of the colubridsnake Rhabdophis subminiatlrs (red-naked keelback). Toxicon 21, 570-574.
Httsrwxn, P . C. and Htt~rwxn, R. R. (1979) D[tpholidws typos (boomslang) snake venom : purification andproperties of the coagulant principle . Toxicon 17, 489498 .
Korroo, H., Kolvno, S., Itcezwww, H ., MtJRATA, R. and Ottgwtcw, A . (1960) Studies on the quantitative methodfor determination of hemorrhagic activity of Habu snake venom. Jpn. J. Afed. Sci. Bèol. 13, 43-51 .
KOANAWK, F . and Mws~tEte, R . W, P . (1964) A comparative examination of yellow and white venoms of Vipersammodytes. Toxicon 2, 109-I11 .
KORNALIK, F. and TAeoxslcl., E . (1978) Procoagulant and defibrinating potency ofthe venom gland extract from7Tlelotornis kirtlandi. 77tromb . Res . 12, 991-(001 .
KOtwwLttr, F., T~(aoastcd, E . and Mlms, D . (1978) Pharmacological and biochemical properties ofa venom glandextract from the snake Thelotornis kirtlattdl. Toxicon 16, 535-542 .

438
M. T. ASSAKURA et al .
KUNITZ, M. (1946) Crystalline soybean trypsin inhibitor. II . General properties . J. gen. Physiol. 30, 291-310.LAEMAiI.t, U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4 .
Nature 217, 680-685.LOWRY, O. H., R06EBR000H, N. J., FARR, A. L. and RANUALL, R. J. (1951) Protein measurement with the Folinphenol reagent. J. biol. Chem. 193, 265-275.
MACKAY, N., FI~tGUSON, J. C., ASHE, J., BAGSHAwE, A., FORItFSIER, A. T. T. and McNtcoL, G. P. (1969) Thevenom of the boomslang (Dispholidus typos) : in viva and in vitro studies . 77eromb. Diath. haemorrh . 21,234-244.
MAIZEL, J. V., JR (1969) Acrylamide gel electrophoresis of proteins and nucleic acids. In : FundamentalTechniques in Virology, p. 334 (HABEL, K. and SALZMAx, N. P., Eds) . New York : Academic Press.
MANDELBAUM, F. R. and AssAxuRw, M. T. (1988) Antigenic relationship of hemorrhagic factors and professesisolated from the venoms of three species of Bothrops snakes. Toxicon 26, 379-385.
MANDELBAUMt, F. R., REICHL, A. P. and ASSAKURA, M. T. (1982) Isolation and characterization of a proteolyticenzyme from the venom of the snake Bothropsjararaca (jararaca) . Toxicon 20, 955-972.
Mtxrox, S. A. and WEIx3TElx, S. A. (1987) Colubrid snake venoms: immunologic relationships, electrophoresicpatterns. Copeia 1987, 993-1000 .
MORrrA, T., MA~rsurdo'ro, H., IWANAGA, S. and SAKAI, A. (1988) A prothrombrin activator found in Rhabdophistigrinus tigrinus (Yamakagashi snake) venom. In : Hemostasis and Animal Venoms, pp . 556 (PtRtctE, H. andMARKLAND, F. S., JR, F.ds) . New York : Mar+cel Dekker .
NAHAS, L., KAMIGUTt, A. S., HOGS, A. R. and Goats, R. C. (1976) Characterization of the coagulant activity ofthe venom of aglyphous Rhabduphis tigrinus snake. In : Animal. Plant, and Microbial Toxins, Vol. I,pp . 159-170 (OHSAKA, A., HAYASHt, K. and $AWAI, Y., Eds) . New York : Plenum Press.
NICKERSOx, M. A. and Htarot~tsox, R. W. (1976) A case of envenomation by the South American colubrid,Philodryas olfersü. Herpetologica 32, 197-198.
Ovwou, M. (1987) Isolation and characterization of a hemorrhagic factor from the venom of the snakeAtractaspis engaddensis (Atractaspidae). Toxicon 25, 62130.
REtseELn, R. A., LEwts, W. J. and WILLIAMS, D. F. (1962) Disk electrophoresis of basic proteins and peptides onpolyacrylamide gels . Nature 195, 281-283.
SAKAI, A. and SAWAI, Y. (1984) Studies on the pathogenesis of envenomation of yamakagashi, Rhabduphistigrinus tigrinus (Boie) . 2. Immunogenicity of the venom. Snake 16, 21-26.
SAtut, A., HONMA, M. and SAWAt, Y. (1984) Study of the toxicity ofvenoms extracted from Duvernoy's gland ofcertain Asian colubrid snakes . Snake 16, 16-20.
SAWAI, Y., SAKAI, A. and HOxMA, M. (1985) Study on pathogenesis ofenvenomation due to Rhabduphis tigrinustigrinus (Bole) a colubrid . Toxicon 23, 607.
SILVA, M. V. and BUONONATO, M. A. (1983/84) Relato clinico de envenenamcnto humano per Philodryas oljersü .Mem. lnst. Butantan 47/48, 121-126.
Vtsr, D. K. (1978) Some effects and properties of Duvernoy's gland secretion from Hvp.siglena turquata texana(Texas night snake) . Toxicon 26, 41719.
VtssPrt, l. and C~upatAx, D. S. (1978) In: Snakes and Snake Bite . Venomous Snakes and Management oJSnakeBite in Southern Africa, pp . 61-131 (PuRtveLL, Mc.D ., Ed.). Johannesburg.
WILLIS, T. W. and Tu, A. T. (1988) Purification and biochemical characterization of atroxase, a nonhemorrhagicfibrinolytic protease from western diamondback rattlesnake venom. Biochemistry 27, 4769777.
YAAtARAWA, M., NozAKI, M. and HOKAStA, Z. (1976) Fractionation of Sakishima habu (Trimeresurus elegans)venom and lethal, hemorrhagic, and edema-forming activities of the fractions . In : Animal, Plant, andMicrobial Toxins Vol. l, pp . 97-(09 (ONSAICA, A., HAYASHI, K. and $AWAt, Y., F.ds) . New York : PlenumPress.




















