
Helicobacter pylori induce neutrophil transendothelial migration: Role of the bacterial HP-NAP
9
Helicobacter pylori induce neutrophil transendothelial migration: Role of the bacterial HP-NAP Mikael Brisslert a,b, * , Karin Enarsson a , Samuel Lundin a , Anna Karlsson b , Johannes G. Kusters c , Ann-Mari Svennerholm a , Steffen Backert d , Marianne Quiding-Ja ¨rbrink a a Department of Medical Microbiology and Immunology, Go ¨ teborg University, Sweden b Department of Rheumatology and Inflammation Research, The Sahlgrenska Academy, Go ¨ teborg University, Guldhedsgatan 10A, 413 45 Go ¨ teborg, Sweden c Department of Gastroenterology and Hepatology, Erasmus MC-University Medical Center, Rotterdam, The Netherlands d Department of Medical Microbiology, Otto von Guericke University, Magdeburg, Germany Received 12 April 2005; received in revised form 2 June 2005; accepted 2 June 2005 First published online 22 June 2005 Edited by A.H.M. van Vliet Abstract Continuous recruitment of neutrophils into the inflamed gastric mucosal tissue is a hallmark of Helicobacter pylori infection in humans. In this study, we examined the ability of H. pylori to induce transendothelial migration of neutrophils using a transwell system consisting of a cultured monolayer of human endothelial cells as barrier between two chambers. We showed for the first time that live H. pylori, but not formalin-killed bacteria, induced a significantly increased transendothelial migration of neutrophils. H. pylori con- ditioned culture medium also induced significantly increased transendothelial migration, whereas heat-inactivated culture filtrates had no effect, suggesting that the chemotactic factor was proteinaceous. Depletion of H. pylori-neutrophil activating protein (HP-NAP) from the culture filtrates resulted in significant reduction of the transmigration. Culture filtrates from isogenic HP-NAP deficient mutant bacteria also induced significantly less neutrophil migration than culture filtrates obtained from wild-type bacteria. HP-NAP did not induce endothelial cell activation, suggesting that HP-NAP acts directly on the neutrophils. In conclusion, our results demonstrate that secreted HP-NAP is one of the factors resulting in H. pylori induced neutrophil transendothelial migration. We pro- pose that HP-NAP contributes to the continuous recruitment of neutrophils to the gastric mucosa of H. pylori infected individuals. Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: HP-NAP; Helicobacter pylori; Transmigration; Endothelial cells; Inflammation 1. Introduction Infection with Helicobacter pylori gives rise to active chronic gastritis in virtually all infected subject. In some individuals, the infection eventually leads to the devel- opment of ulcer disease or gastric adenocarcinoma. Sev- eral bacterial and host factors seem to play important roles in determining the severity of the inflammation and the outcome of the infection [1,2]. Continuous recruitment of neutrophils into the inflamed gastric mucosal tissue is a hallmark in human H. pylori infections. A strong correlation exists between gastric neutrophil infiltration, mucosal damage, and develop- ment of duodenal ulcer (DU) disease in H. pylori 0378-1097/$22.00 Ó 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsle.2005.06.008 * Corresponding author. Tel.: +46 0 31 3424021; fax: +46 0 31 823925. E-mail address: [email protected] (M. Brisslert). www.fems-microbiology.org FEMS Microbiology Letters 249 (2005) 95–103
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Helicobacter pylori induce neutrophil transendothelial migration: Role of the bacterial HP-NAP

www.fems-microbiology.org
FEMS Microbiology Letters 249 (2005) 95–103
Helicobacter pylori induce neutrophil transendothelialmigration: Role of the bacterial HP-NAP
Mikael Brisslert a,b,*, Karin Enarsson a, Samuel Lundin a, Anna Karlsson b,Johannes G. Kusters c, Ann-Mari Svennerholm a, Steffen Backert d,
Marianne Quiding-Jarbrink a
a Department of Medical Microbiology and Immunology, Goteborg University, Swedenb Department of Rheumatology and Inflammation Research, The Sahlgrenska Academy, Goteborg University, Guldhedsgatan 10A,
413 45 Goteborg, Swedenc Department of Gastroenterology and Hepatology, Erasmus MC-University Medical Center, Rotterdam, The Netherlands
d Department of Medical Microbiology, Otto von Guericke University, Magdeburg, Germany
Received 12 April 2005; received in revised form 2 June 2005; accepted 2 June 2005
First published online 22 June 2005
Edited by A.H.M. van Vliet
Abstract
Continuous recruitment of neutrophils into the inflamed gastric mucosal tissue is a hallmark of Helicobacter pylori infection in
humans. In this study, we examined the ability ofH. pylori to induce transendothelial migration of neutrophils using a transwell system
consisting of a cultured monolayer of human endothelial cells as barrier between two chambers. We showed for the first time that live
H. pylori, but not formalin-killed bacteria, induced a significantly increased transendothelial migration of neutrophils. H. pylori con-
ditioned culture medium also induced significantly increased transendothelial migration, whereas heat-inactivated culture filtrates had
no effect, suggesting that the chemotactic factor was proteinaceous. Depletion of H. pylori-neutrophil activating protein (HP-NAP)
from the culture filtrates resulted in significant reduction of the transmigration. Culture filtrates from isogenic HP-NAP deficient
mutant bacteria also induced significantly less neutrophil migration than culture filtrates obtained from wild-type bacteria.
HP-NAP did not induce endothelial cell activation, suggesting that HP-NAP acts directly on the neutrophils. In conclusion, our results
demonstrate that secreted HP-NAP is one of the factors resulting inH. pylori induced neutrophil transendothelial migration. We pro-
pose that HP-NAP contributes to the continuous recruitment of neutrophils to the gastric mucosa of H. pylori infected individuals.
� 2005 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: HP-NAP; Helicobacter pylori; Transmigration; Endothelial cells; Inflammation
1. Introduction
Infection with Helicobacter pylori gives rise to active
chronic gastritis in virtually all infected subject. In some
individuals, the infection eventually leads to the devel-
0378-1097/$22.00 � 2005 Federation of European Microbiological Societies
doi:10.1016/j.femsle.2005.06.008
* Corresponding author. Tel.: +46 0 31 3424021; fax: +46 0 31
823925.
E-mail address: [email protected] (M. Brisslert).
opment of ulcer disease or gastric adenocarcinoma. Sev-
eral bacterial and host factors seem to play important
roles in determining the severity of the inflammation
and the outcome of the infection [1,2]. Continuous
recruitment of neutrophils into the inflamed gastricmucosal tissue is a hallmark in human H. pylori
infections. A strong correlation exists between gastric
neutrophil infiltration, mucosal damage, and develop-
ment of duodenal ulcer (DU) disease in H. pylori
. Published by Elsevier B.V. All rights reserved.

Table 1
Characterization of H. pylori strains used in the study
Strain cag PAIa VacAb Origin
Hel 312 + s1/m1c ASd
Hel333 � s2/m2 DUe
96 M. Brisslert et al. / FEMS Microbiology Letters 249 (2005) 95–103
infections [3,4]. There is also strong evidence that H. py-
lori is able to activate neutrophils; in some studies, the
strains isolated from DU patients were shown to be
more potent activators of neutrophils than other isolates
[3,5–9]. One of the bacterial factors shown to mediate
neutrophil activation is the H. pylori neutrophil activat-ing protein (HP-NAP) [5]. Evans et al. [5,9] first charac-
terized the HP-NAP protein and showed that it
stimulated the release of reactive oxygen species by hu-
man neutrophils. HP-NAP is dodecameric and is able
to bind iron. It has sequence similarities with iron-bind-
ing proteins in other bacterial species [10,11]. HP-NAP
is a cytosolic protein expressed by virtually all H. pylori
isolates [12]. It is released into the supernatant by autol-ysis, upon which a fraction of HP-NAP is then bound to
carbohydrates on the external surface of the outer mem-
brane [5,13,14].
While the mechanisms behind the continuous recruit-
ment of neutrophils to H. pylori infected tissues in vivo
remain to be fully understood, endothelial cell function
and activation appear to be key components in this pro-
cess. Endothelial cells are involved in a wide range ofphysiological reactions including blood coagulation,
blood pressure regulation, and inflammatory responses.
One important function is to direct circulating leuko-
cytes to sites of inflammation via a series of steps that
are independent of the stimulatory agent [15]. During
the inflammatory process, or upon in vitro stimulation
with TNF-a, IL-1 or bacterial products, endothelial cellsrespond rapidly (within a few hours) by upregulation ofadhesion molecules such as VCAM-1, ICAM-1, and E-
selectin as well as production of proinflammatory cyto-
kines and chemokines [16], all of which contribute to
leukocyte recruitment.
We have shown in a previous study that endothelial
cells are strongly activated by H. pylori, resulting in in-
creased production of neutrophil recruiting chemokines
and expression of adhesion molecules [17]. To furthercharacterize this response, we investigated the effect of
H. pylori on the migration of neutrophils through cul-
tured endothelial monolayers. We showed that live bac-
teria, secreted bacterial proteins containing HP-NAP,
and purified HP-NAP all resulted in an increased trans-
endothelial migration of neutrophils. In contrast, cul-
ture filtrates depleted of HP-NAP, as well as culture
filtrates produced by isogenic HP-NAP deficient mu-tants were less effective in inducing neutrophil transen-
dothelial migration.
P1 + s1/m2 NUDfP12 + s1/m1 DU
SS1 + s2/m2 DU, mouse adapted
a cag PAI, cytotoxin-associated gene pathogenicity island.b VacA, vacuolating toxin A.c s, sequence region; m, middle region.d AS, asymptomatic carrier.e DU, duodenal ulcer patient.f NUD, non-ulcer dyspepsia patient.
2. Materials and methods
2.1. H. pylori strains
H. pylori strains were isolated from the duodenum of
adults suffering from DU (strain Hel 333, P12, Table 1),
from the stomach of adult asymptomatic subject (strain
Hel 312, Table 1) or from a patient suffering from a non-
ulcer dyspepsia (strain P1, Table 1), as previously de-
scribed [3,18]. Isolates were stored in a freeze-drying
medium (tryptic soy broth with 15% glycerol) at
�80 �C. The mouse adapted strain SS1 was originallyprovided by Dr. Adrian Lee, Sydney, Australia [19].
Characteristics of these strains [12,17] are summarized
in Table 1.
2.2. Generation of isogenic HP-NAP deficient mutants
Isogenic P1D nap and P12D nap knock-out mutants
were constructed by insertion of a chloramphenicolresistance gene cassette (CmR, 1 kb fragment of plasmid
pTnMax1) or kanamycin resistance gene cassette (Aph-
3, 1.4 kb fragment of plasmid pILL-600) according to a
standard protocol [18]. For this purpose, we amplified a
fragment of 1.8 kb containing the nap gene and flanking
regions with the following primers: 179F 5 0-TTT-
TTGAAGGGCCAATCTTAGAA and 179R 5 0-AGC-
GACGAAGGGTTTTTTG. This product was ligatedinto pCR2.1 vector (Invitrogen, Karlsruhe, Germany)
and the correct integration of the CmR or Aph-3 cas-
settes in the nap gene was checked by standard PCR.
Both mutants were created with terminatorless cassettes,
and are non-polar.
2.3. H. pylori culture conditions
H. pylori were cultured routinely on Columbia-iso
agar plates at 37 �C in a microaerophilic milieu (3%
O2, 5% CO2 and 92% N2). After four days of incubation,
the bacteria were resuspended in 3–5 ml of PBS. The
optical density (OD) of the suspension was determined
(Shimadzu UV-1201, Lambda Polynom, Stockholm,
Sweden), and adjusted to a final OD600nm = 1.0 in
PBS, corresponding to approximately 5 · 109 H. pylori
bacteria ml�1 (5 · 109 CFUs ml�1). In some experi-
ments, PBS resuspended bacteria inactivated by treat-
ment with 0.01 M Formaldehyde at 37 �C for 2 h and
then at room temperature over night, were used.

M. Brisslert et al. / FEMS Microbiology Letters 249 (2005) 95–103 97
2.4. Preparation of H. pylori secreted proteins (culture
filtrate)
H. pylori strains were grown and resuspended as de-
scribed above. Bacterial suspension (2.5 ml) was added
to 48 ml of M200 medium (Cascade Biologics Inc. OR,USA) containing Brucella broth (Difco Laboratories,
Detroit, MA, USA), 5% new born calf serum (Bio-
chrome AG, Berlin Germany), 10lg ml�1 Vancomycin,
5 lg ml�1 Trimothoprim, and 20 U ml�1 Polymyxin B,
and the mixture was then incubated for 6.5 h at 37 �Cin 5% CO2. The time point was chosen based on our pre-
vious study showing that human umbilical vein endothe-
lial cells were strongly stimulated by secreted H. pylori
proteins produced by a 6.5 h culture [17]. The cultures
were sterile filtered, and one aliquot of each culture
filtrate was heat-inactivated at 85 �C for 35 min. All
filtrates were then stored at �80 �C until use.
2.5. Depletion of HP-NAP from H. pylori culture filtrate
The culture filtrate was depleted of HP-NAP usingmagnetic beads (Dynal, Norway) coupled to IgG1
monoclonal antibody (mAb) against HP-NAP (NAP
6:1), the preparation of which has been described previ-
ously [12]. Sixty milligrams of beads coated with sheep
antibodies to mouse IgG1 were incubated with 120 lgIgG1 mAb NAP 6:1 for 4 h, washed on a magnet, resus-
pended in 2 ml of the culture filtrate and incubated for
4 h at 4 �C on a waddle table. The beads were removedwith a magnet and the HP-NAP depleted culture filtrate
was stored at �80 �C until use. Beads incubated with an
IgG1 mAb reacting with cholera toxin were used as con-
trols for unspecific binding. HP-NAP depletion from
culture filtrates was confirmed by Western blot analysis
using the same NAP-mAb. In brief, samples were sepa-
rated on a 18% Tris–Glycine SDS–PAGE, and blotted
onto a PVDF membrane using standard procedures.After blocking in 2% milk powder diluted in blotting
buffer (25 mM Tris–HCl, pH 7.4, 150 mM NaCl, 0.1%
Tween 20), the membrane was probed with monoclonal
mouse anti-NAP [12] diluted 1:5 in blotting buffer over
night at 4 �C. Thereafter the membrane was washed
and mouse immunoglobulins were detected with biotin-
ylated rabbit anti-mouse Ig (1:2000, DAKO, E0413),
followed by a combination of HRP-conjugated donkeyanti-rabbit Ig (1:2000, Amersham, NA9340) and Strep
ABComplex/HRP (DAKO). Peroxidase activity was
subsequently detected by ECL chemiluminescense
(Amersham) according to the manufacturer�s instruc-
tions and exposure for 15 min on Fuji NP100 film
(Fig. 3B).
The distribution of immunogenic H. pylori proteins
in the culture filtrates was examined by Western blottingusing an H. pylori-reactive polyclonal mouse serum pool
(data not shown). With the exception of HP-NAP, the
relative distribution of other filtrate proteins remained
unchanged during incubation with the beads.
2.6. Production of rNAP
rNAP was produced in E. coli as described and pro-vided by Dr. Susanne Nystrom [12]. Briefly, E. coli cells
were transformed with an expression vector containing
the H. pylori nap gene under the control of the T7 pro-
moter. After disruption of the cells, the soluble proteins
were applied to a Q-Sepharose column (Pharmacia Bio-
Tech, Uppsala, Sweden) in 25 mM Tris–HCl and
50 mM NaCl (pH 8.0). rNAP was found in the break-
through. The fractions containing rNAP were concen-trated and then purified by gel filtration through a
Superdex 200 column (Pharmacia). rNAP eluted as a
single peak. Purity of the protein was confirmed by
SDS–PAGE and Coomassie staining as well as immuno-
blotting, using mAbs specific for rNAP [20].
2.7. Isolation of neutrophils
Human neutrophils were isolated from buffy coats, or
alternatively, from heparinised whole blood obtained
from healthy blood donors. After dextran sedimentation
at 1g, hypotonic lysis of the remaining erythrocytes, and
subsequent centrifugation in a Ficoll-Paque gradient
[21], the neutrophils were washed twice and resuspended
to a final volume of 1 · 107 ml�1 in Krebs–Ringer phos-
phate buffer (KRG) containing glucose (10 mM), Ca2+
(1 mM), and Mg2+ (1.5 mM), and kept on ice up to
1.5 h until use.
2.8. Sub-culturing and stimulation of endothelial cells
Human umbilical vein endothelial cells (HUVEC),
passage 1, were purchased from Cascade Biologics Inc.
Cells were grown in 75 cm2 plastic bottles (Nunc, Ros-kilde, Denmark) in M200 medium supplemented with
low serum growth supplement, (LSGS) and penicillin–
streptomycin–amphotericin (PSA) (all from Cascade
Biologics Inc.). Medium was changed every 48 h during
culturing at 37 �C, 5% CO2. The medium used in this
study contained routinely less than 0.03 EU ml�1 endo-
toxin as confirmed by Limulus test.
Confluent monolayers were trypsinized using0.25 mg ml�1 trypsin/EDTA (Cascade Biologics Inc.)
and split at a ratio of 1:3. At passage 4, the cells were
frozen in fetal calf serum containing 10% DMSO
(Sigma) at a density of 106 cells ml�1. HUVEC within
passage 4–7 were used throughout the experiments [22]
to minimize spontaneous morphological or functional
changes as a result of continuous passaging.
HUVEC were grown for four days, after which themedium was replaced with fresh antibiotic-free
medium containing 10 lg ml�1 rHP-NAP or 5 · 108 live

Fig. 1. FACS-analysis of migrating neutrophils. 105 cells were
collected after transendothelial migration against culture medium
(A), 10�8 M fMLF (B) or 5 · 108 live H. pylori bacteria (C) and
enumerated by flow cytometry. Axes show forward (FSC-H) and side
scatter (SSC-H) characteristics. In all figures, TrueCount� beads can
be seen as indicated by arrow and text in (B). Following encounter
with bacteria (C), neutrophils increase their side scatter signal. The
figure shows one representative example of all FACS plots analyzed in
this study.
98 M. Brisslert et al. / FEMS Microbiology Letters 249 (2005) 95–103
wild-type P1 or P1Dnap. After 6.5 h incubation, the cul-
ture medium was collected and HUVEC were detached
from the plate using trypsin. The endothelial expression
of ICAM-1, E-selectin and VCAM-1 was determined by
flow cytometry, and the IL-8 concentration in the cul-
ture medium analyzed by ELISA, as previously de-scribed [17].
2.9. Transendothelial migration of neutrophils
HUVEC (3 · 105 cells) were added to collagen-coated
nets of Transwell chambers (Costar, Badhoevedorp, The
Netherlands) with a growth area of 4.7 cm2, and allowed
to grow to confluence. When HUVEC reached a conflu-ent monolayer after 24 h as confirmed bymicroscopy, the
medium was changed and incubation continued for an-
other hour at 37 �C. This creates a system with two sep-
arate chambers, where transport through the
endothelial monolayer is the only means of communica-
tion between the chambers, thereby allowing a chemotac-
tic gradient of the substances added to the lower chamber
to be established. Stimuli were placed in the lower cham-ber in 1 ml of M200 medium, and purified neutrophils
(106 cells in 100 ll KRG medium) were added in the
upper chamber to a final volume of 1 ml. The addition
of bacteria was carried out 1 h before the addition of neu-
trophils to allow a gradient of secreted products to be
established, whereas all other stimulatory agents were
added at the same time as the neutrophils. The chemo-
attractant formyl-methionyl-leucyl-phenylalanine (fMLF)was used at a concentration of 10�8 M as a positive con-
trol in all experiments and was added in a volume of
100 ll. All stimuli (including bacteria) used were present
in the lower chamber throughout the experiments.
After 90 min (optimal duration for reducing sponta-
neous migration and yet achieving maximum migration
of attracted cells) of transendothelial migration, the neu-
trophil containing medium in the lower chamber was re-moved and collected in polypropylene plastic tubes on
ice. Meanwhile, 2 ml of 0.25% Trypsin/EDTA solution
was added in order to detach neutrophils that were
adhering to the plastic well or to the bottom of the filter.
The cells harvested were sedimented, transferred to a 96-
well polypropylene plate and stained for the expression
of the complement receptor 3 (CR3) by adding
10 lg ml�1 of the PE-conjugated anti-CR3 antibody(BD, Erembodegem, Belgium) in a total volume of
100 ll PBS containing 1.46 g l�1 EDTA, 2.5 g l�1 bovine
serum albumin, 0.2 g l�1 NaN3 and 2% AB+ human ser-
um (FACS-wash), and incubated on ice for 35 min.
After washing the pellet twice in FACS-wash buffer
the transmigrated cells were fixed in Cell Fix� (BD)
and enumerated by flow cytometry (Facs-Calibur, BD)
using True Count� beads (BD). Neutrophils were iden-tified by their forward and side scatter characteristics, as
illustrated in Fig. 1. Since these parameters vary with the
agent used for stimulation, expression of CR3, which is
upregulated during transendothelial migration, was used
to confirm that gated events were indeed neutrophils.
2.10. Statistical analysis
Statistical evaluations were performed using the
Mann–Whitney test and the Prism software.

M. Brisslert et al. / FEMS Microbiology Letters 249 (2005) 95–103 99
3. Results
3.1. Live H. pylori and culture filtrates induce neutrophil
transendothelial migration
Neutrophil transendothelial migration towardsH. py-
lori bacteria was analyzed in the Transwell system and
quantified using flow cytometry. Five well-characterised
H-pylori strains (Hel 312, Hel 333, SS1, P1 and P12;
Table 1) were used in our experiments and gave similar
results. One representative FACS-dot plot is shown in
Fig. 1. Titration experiments demonstrated a dose
dependent migration of neutrophils towards H. pylori,
with 5 · 108 of bacteria giving a strong and consistent re-sponse (data not shown). This amount of bacteria was
used in all further experiments. H. pylori strain Hel 312
carrying the cag pathogenicity island (cag PAI+) and
vacA s1/m1 allele induced transendothelial migration of
significantly (p < 0.01) more neutrophils than the
unstimulated control (approximately 25% of the added
neutrophils compared to about 5% in the control) (Fig.
2). The H. pylori strain Hel 333 (cag PAI�, vacA s2/m2allele) also induced a substantially increased neutrophil
transendothelial migration, where approximately 70%
of the total added neutrophils had migrated (p < 0.01,
data not shown). In contrast, the same number of forma-
lin-inactivated Hel 312 induced significantly less
(p < 0.05) migration compared to live Hel 312 (Fig. 2).
Since live but not dead bacteria could induce neutro-
phil migration, we sought to test if the effect was caused
Fig. 2. Induction of neutrophil transendothelial migration by culture
filtrates from H. pylori. 5 · 108 live or formalin killed H. pylori
bacteria, culture filtrate, or heat-inactivated culture filtrate, were added
to the lower wells of Transwell cultures and 106 neutrophils to the
upper wells. fMLF (10�8 M) was used as a positive control. Bars
indicate geometric mean + SEM of percentage of transmigrated
neutrophils. Statistical evaluation between negative control and
stimulated samples was performed by the Mann–Whitney test.
***p < 0.001, **p < 0.01, and *p < 0.05. Data are obtained in tripli-
cates from at least three independent experiments.
by secretion of neutrophil chemotactic factors by bacte-
ria into the culture filtrates. We chose to use strain Hel
312, since Hel 333 grows poorly and since the presence
of the CagPAI did not seem to influence induction of
migration. In addition, H. pylori induced chemokine
and adhesion molecule upregulation on endothelial cellsis independent of the CagPAI (16). Indeed, when culture
filtrate from Hel 312 bacteria was used as stimulatory
agent, we could detect a significant level of approxi-
mately 20% neutrophil transendothelial migration
(p < 0.01), whereas the positive control fMLF induced
approximately 40% migration (p < 0.001) (Fig. 2). Heat
inactivation of the H. pylori filtrate at 85 �C for 35 min
significantly abolished the capacity of the culture super-natant to induce transendothelial migration (Fig. 2),
indicating that the stimulatory agents are of proteina-
ceous nature. Similar results were obtained from culture
filtrates of strain Hel 333. As a negative control, culture
medium treated in the same way but without the addi-
tion of bacteria did not affect neutrophil transendothe-
lial migration (data not shown).
3.2. HP-NAP is a potent chemotactic factor for
neutrophils in culture filtrates of H. pylori
Since H. pylori culture filtrates induced neutrophil
transendothelial migration to the same extent as bacte-
ria, we examined if HP-NAP, a well known neutrophil
activating factor [5,7,8], is responsible for the induction
of neutrophil migration. rNAP was added to the lowerchambers at a concentration of 10 lg ml�1 and neutro-
phil transendothelial migration was measured. This con-
centration was selected based on in vitro titration
experiments in our model system to give the highest
induction of transendothelial migration (data not
shown). When rNAP was added to the assay, almost
25% (p < 0.01) of the added neutrophils transmigrated
(Fig. 3A). Again, the positive control fMLF inducedthe highest degree of transendothelial migration, attract-
ing about 35% (p < 0.001) of the added neutrophils in
this set of experiments (Fig. 3A). To avoid the possibil-
ity of LPS contamination, 200 lg ml�1 rNAP was incu-
bated with 20 lg ml�1 of polymyxin B for 1 h at room
temperature. After incubation, 100 ll of the HP-NAP–
polymyxin B mixture was added to each well, giving a
final concentration of 10 lg ml�1 HP-NAP and1 lg ml�1 polymyxin B. Limulus endotoxin test per-
formed before and after polymyxin treatment showed
that the HP-NAP preparation contained less than
1.4 EU ml�1 of LPS corresponding to 70 pg ml�1 E. coli
LPS, far less than the LPS concentration needed for
induction of transendothelial migration (10,000–
25,000 pg ml�1) [23]. Polymyxin itself had no effect on
transendothelial migration (data not shown).Thus, our findings show that rNAP is a potent indu-
cer of transendothelial migration. We next sought to
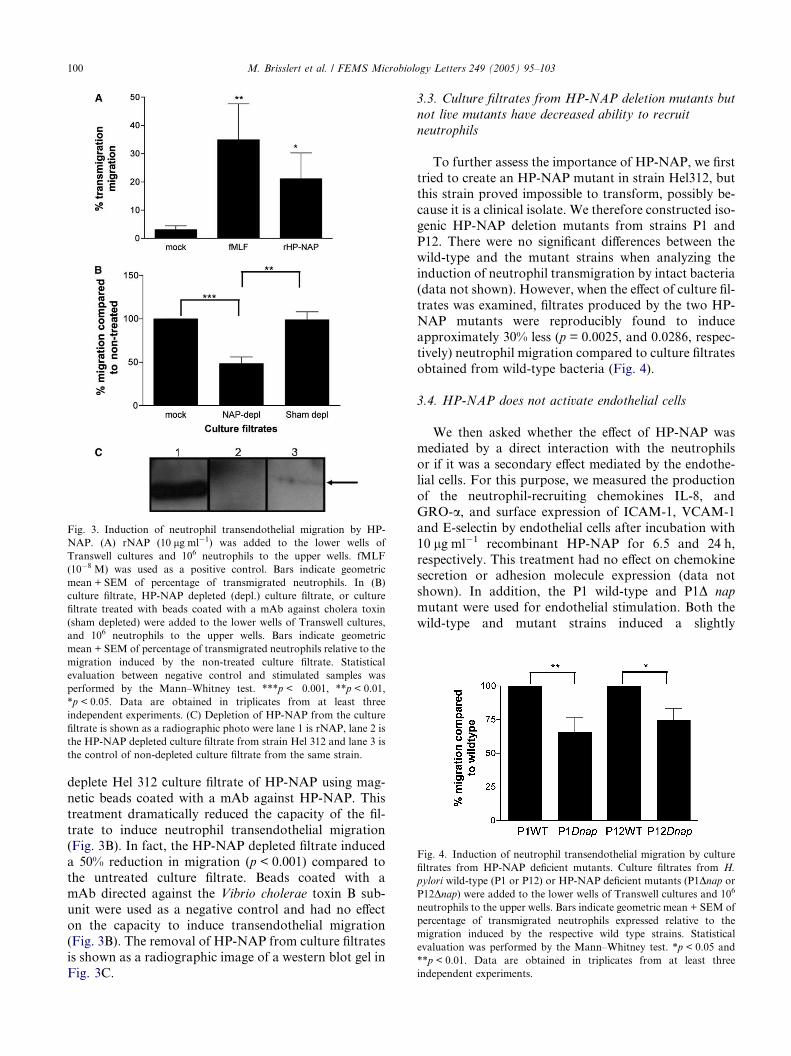
Fig. 3. Induction of neutrophil transendothelial migration by HP-
NAP. (A) rNAP (10 lg ml�1) was added to the lower wells of
Transwell cultures and 106 neutrophils to the upper wells. fMLF
(10�8 M) was used as a positive control. Bars indicate geometric
mean + SEM of percentage of transmigrated neutrophils. In (B)
culture filtrate, HP-NAP depleted (depl.) culture filtrate, or culture
filtrate treated with beads coated with a mAb against cholera toxin
(sham depleted) were added to the lower wells of Transwell cultures,
and 106 neutrophils to the upper wells. Bars indicate geometric
mean + SEM of percentage of transmigrated neutrophils relative to the
migration induced by the non-treated culture filtrate. Statistical
evaluation between negative control and stimulated samples was
performed by the Mann–Whitney test. ***p < 0.001, **p < 0.01,
*p < 0.05. Data are obtained in triplicates from at least three
independent experiments. (C) Depletion of HP-NAP from the culture
filtrate is shown as a radiographic photo were lane 1 is rNAP, lane 2 is
the HP-NAP depleted culture filtrate from strain Hel 312 and lane 3 is
the control of non-depleted culture filtrate from the same strain.
Fig. 4. Induction of neutrophil transendothelial migration by culture
filtrates from HP-NAP deficient mutants. Culture filtrates from H.
pylori wild-type (P1 or P12) or HP-NAP deficient mutants (P1Dnap or
P12Dnap) were added to the lower wells of Transwell cultures and 106
neutrophils to the upper wells. Bars indicate geometric mean + SEM of
percentage of transmigrated neutrophils expressed relative to the
migration induced by the respective wild type strains. Statistical
evaluation was performed by the Mann–Whitney test. *p < 0.05 and
**p < 0.01. Data are obtained in triplicates from at least three
independent experiments.
100 M. Brisslert et al. / FEMS Microbiology Letters 249 (2005) 95–103
deplete Hel 312 culture filtrate of HP-NAP using mag-
netic beads coated with a mAb against HP-NAP. This
treatment dramatically reduced the capacity of the fil-
trate to induce neutrophil transendothelial migration
(Fig. 3B). In fact, the HP-NAP depleted filtrate induced
a 50% reduction in migration (p < 0.001) compared to
the untreated culture filtrate. Beads coated with amAb directed against the Vibrio cholerae toxin B sub-
unit were used as a negative control and had no effect
on the capacity to induce transendothelial migration
(Fig. 3B). The removal of HP-NAP from culture filtrates
is shown as a radiographic image of a western blot gel in
Fig. 3C.
3.3. Culture filtrates from HP-NAP deletion mutants but
not live mutants have decreased ability to recruit
neutrophils
To further assess the importance of HP-NAP, we first
tried to create an HP-NAP mutant in strain Hel312, butthis strain proved impossible to transform, possibly be-
cause it is a clinical isolate. We therefore constructed iso-
genic HP-NAP deletion mutants from strains P1 and
P12. There were no significant differences between the
wild-type and the mutant strains when analyzing the
induction of neutrophil transmigration by intact bacteria
(data not shown). However, when the effect of culture fil-
trates was examined, filtrates produced by the two HP-NAP mutants were reproducibly found to induce
approximately 30% less (p = 0.0025, and 0.0286, respec-
tively) neutrophil migration compared to culture filtrates
obtained from wild-type bacteria (Fig. 4).
3.4. HP-NAP does not activate endothelial cells
We then asked whether the effect of HP-NAP wasmediated by a direct interaction with the neutrophils
or if it was a secondary effect mediated by the endothe-
lial cells. For this purpose, we measured the production
of the neutrophil-recruiting chemokines IL-8, and
GRO-a, and surface expression of ICAM-1, VCAM-1
and E-selectin by endothelial cells after incubation with
10 lg ml�1 recombinant HP-NAP for 6.5 and 24 h,
respectively. This treatment had no effect on chemokinesecretion or adhesion molecule expression (data not
shown). In addition, the P1 wild-type and P1D nap
mutant were used for endothelial stimulation. Both the
wild-type and mutant strains induced a slightly

Fig. 5. Effect of HP-NAP on endothelial cells. 5 · 108 live H. pylori P1 wild-type or HP-NAP deficient mutant P1Dnap were added to cultured
endothelial cells and incubated for 6 h whereafter IL-8 production (A) was analyzed using ELISA and the expression of ICAM-1 (B), E-selectin (C)
and VCAM-1 (D) was evaluated by flow cytometry. Data are obtained in triplicates from at least three independent experiments. Bars indicate
geometric mean + SEM of percentage of transmigrated neutrophils. Statistical evaluation was performed by the student�s t test. ***p < 0.001,
**p < 0.01, and *p < 0.05. There was no significant difference between wild-type and mutant bacteria in any of the assays.
M. Brisslert et al. / FEMS Microbiology Letters 249 (2005) 95–103 101
increased production of IL-8 (Fig. 5) and increased
expression of the adhesion molecules ICAM-1,
VCAM-1, and E-selectin compared to unstimulated
controls (Fig. 5). Since the effect of the HP-NAP defi-
cient mutant was indistinguishable from that of the
wild-type bacteria, the results indicate that HP-NAP ex-erts transmigratory effects on the neutrophils themselves
rather than on the endothelial cells.
4. Discussion
Endothelial cells have a key function in the recruit-
ment of neutrophils during inflammation due to theirability to produce cytokines and chemokines and
expressing adhesion molecules relevant for transmigra-
tion. Circulating neutrophils in turn are sensitive to che-
motactic factors such as bacterial peptides and
chemokines produced at the site of inflammation. When
encountering such stimuli, the neutrophils attach to the
endothelium, leave the blood stream, and migrate to-
wards increasing amounts of stimulus. During H. pylori
infection, a pronounced and continuous recruitment of
neutrophils to the inflamed gastric mucosa has been ob-
served [1,2]. Indeed, neutrophil infiltration has been sug-
gested to be one of the factors leading to mucosal
damage and DU formation [3,4]. In a recent study, we
showed that live H. pylori was able to activate endothe-
lial cells to express several adhesion molecules and neu-
trophil recruiting CXC-chemokines [17]. In this study,
using the transwell system where cultured endothelial
cells creates two separate compartments allowing pas-
sage from one side to another only through the endothe-
lial monolayer, we extend these findings by showing
that live H. pylori as well as bacterial culture filtrates
and purified HP-NAP can induce substantial transendo-thelial migration of human neutrophils in a manner
independent of a functional cag PAI or VacA. Further-
more, the chemotactic effect occurred without prior trig-
gering or other stimulatory signals.
Broom et al. [24] previously showed that H. pylori
can synthesize and secrete formylated peptides into cul-
ture medium. In addition, a study by Craig et al. [25]
showed that H. pylori secrete heat-stable and acid resis-tant neutrophil chemotactic factors into the culture
medium, which have a molecular weight of less than
3 kDa, indicating that they might be the same peptides
as those detected by Broom et al. [24]. In addition, Niel-
sen and Andersen [6] showed that sonicates from H. py-
lori contain chemotactic proteins with a molecular
weight between 25 and 35 kDa. The latter results are
in agreement with our finding that H. pylori culture fil-trates induce extensive neutrophil transendothelial
migration, and that heat treatment inhibits this activity,
indicating that the stimulatory factor(s) are of proteina-
ceous nature. Previous studies [6] have shown chemotac-
tic activity of H. pylori sonicates or water extracts, but
this is the first study to demonstrate the actual release
of chemotactic proteins from H. pylori to the
environment.

102 M. Brisslert et al. / FEMS Microbiology Letters 249 (2005) 95–103
HP-NAP has been shown to be chemotactic to hu-
man neutrophils [9], and we therefore depleted HP-
NAP from the Hel 312 culture filtrate. This treatment
substantially decreased their chemotactic effect. Further-
more, purified HP-NAP induced a large increase in neu-
trophil transendothelial migration. However, when weassessed the importance of HP-NAP using intact bacte-
ria, there was no significant difference in the induction of
migration between wild-type and HP-NAP mutant
strains, whereas culture filtrates from the knock-out
strains showed significantly reduced ability to attract
neutrophils as compared to culture filtrates from wild-
type H. pylori. These seemingly contradictory findings
might stem from the fact that intact bacteria are morecomplex than culture filtrate in terms of protein compo-
sition – the former might contain other chemotactic fac-
tors in addition to HP-NAP. Furthermore, strain
differences may explain why HP-NAP exhibits high lev-
els of chemotactic activity in Hel 312 derived culture fil-
trates but only a lower level of activity in the P1 and P12
culture filtrates. We therefore postulate that P1 and P12
may release other chemotactic factors, which are notproduced by Hel 312. It is interesting to note that a cor-
responding difference has recently been observed when
analyzing the effect of VacA blocking of NF-AT in T
cells [26]. In this system, while supernatants from wild-
type bacteria and not vacA mutants inhibited T cell pro-
liferation, no difference was observed with either live
wild-type H. pylori or vacA mutants.
In the original article, Evans et al. [5] describe in-creased adherence of neutrophils to HUVEC after incu-
bation with H. pylori water extracts, an effect that could
partly be blocked by antibodies directed against HP-
NAP. However, we could not demonstrate that purified
HP-NAP activates the HUVEC to express adhesion mol-
ecules or chemokines. Furthermore, there was no differ-
ence in the induction of endothelial activation
molecules between wild-type andHP-NAPmutantH. py-
lori in our system. These findings suggest that HP-NAP is
transported across the endothelial monolayer and acts di-
rectly on the neutrophils, which could in turn influence
the endothelial cells to promote transmigration. We have
thus extended previous studies on HP-NAP-induced che-
motaxis [9] showing that this protein is highly effective in
inducing transendothelial migration of neutrophils. In-
deed, our findings indicate thatHP-NAP is one of thema-jor inducers of transendothelial migration of neutrophils
present in H. pylori culture filtrates. However, we do not
discount the possibility that additional chemotactic H.
pylori products could act in parallel, or in synergy with
HP-NAP inmediating neutrophil chemotaxis in vivo. Be-
sides, chemokines such as IL-8 and ENA-78, which have
been detected in H. pylori positive individuals, are also
likely to contribute to neutrophil recruitment [27,28].Our results also showed that sufficient amounts of
HP-NAP to induce transendothelial migration were re-
leased into the culture medium by live H. pylori within
only 6 h of incubation. It is not known if HP-NAP
can gain access to the lamina propria during infection
in vivo. Given that H. pylori urease is released in a sim-
ilar fashion, and can be detected in the infected gastric
tissues, we propose that HP-NAP might be able to pen-etrate into the lamina propria. In addition, active trans-
port of HP-NAP across epithelial monolayers has
recently been demonstrated in vitro [29]. Our laboratory
has recently shown that expression of HP-NAP is much
higher in vivo than in vitro, suggesting that HP-NAP is
an important factor in H. pylori induced gastritis [30].
Furthermore, no clinical isolate of H. pylori lacking
the gene encoding HP-NAP has been reported, furthersupporting the postulation that this protein confers a
selective advantage for the bacteria in vivo.
In conclusion, H. pylori have the ability to induce sig-
nificant transendothelial migration of neutrophils
through a monolayer of human endothelial cells. Live
bacteria and bacterial filtrates may induce neutrophil
migration in different manners. HP-NAP thus may play
a significant role in neutrophil migration when presentin bacterial filtrates. Therefore, we suggest that in vivo
where live bacteria in the lamina propria are scarce,
HP-NAP may be a major factor contributing to the con-
tinuous recruitment of neutrophils to the gastric mucosa
in H. pylori-associated gastritis.
Acknowledgements
We thank Dr. Susanne Nystrom for kindly providing
the rNAP used in this study, Dr. Mattias Magnusson for
help with western blot analyses, and Dr. Terry Kwok for
critical reading of the manuscript. This study was sup-
ported by grants from the Swedish Science Council
(Grant No. 06X-13428) and the Faculty of Medicine
at Goteborg University.
References
[1] Go, M.F. (1997) What are the host factors that place an
individual at risk for Helicobacter pylori-associated disease?.
Gastroenterology 113, S15–20.
[2] Del Giudice, G., Covacci, A., Telford, J.L., Montecucco, C. and
Rappuoli, R. (2001) The design of vaccines against Helicobacter
pylori and their development. Annu. Rev. Immunol. 19, 523–
563.
[3] Hamlet, A., Thoreson, A.C., Nilsson, O., Svennerholm, A.M. and
Olbe, L. (1999) Duodenal Helicobacter pylori infection differs in
cagA genotype between asymptomatic subjects and patients with
duodenal ulcers. Gastroenterology 116, 259–268.
[4] Davies, G.R., Simmonds, N.J., Stevens, T.R., Sheaff, M.T.,
Banatvala, N., Laurenson, I.F., Blake, D.R. and Rampton, D.S.
(1994) Helicobacter pylori stimulates antral mucosal reactive
oxygen metabolite production in vivo. Gut 35, 179–185.
[5] Evans Jr., D.J., Evans, D.G., Takemura, T., Nakano, H.,
Lampert, H.C., Graham, D.Y., Granger, D.N. and Kvietys,

M. Brisslert et al. / FEMS Microbiology Letters 249 (2005) 95–103 103
P.R. (2001) Characterization of a Helicobacter pylori neutrophil-
activating protein. Infect. Immun. 63, 2213–2220.
[6] Nielsen, H. and Andersen, L.P. (1992) Chemotactic activity of
Helicobacter pylori sonicate for human polymorphonuclear leu-
cocytes and monocytes. Gut 33, 738–742.
[7] Nielsen, H. and Andersen, L.P. (1995) Activation of phagocytes
by Helicobacter pylori correlates with the clinical presentation of
the gastric infection. Scand. J. Infect. Dis. 27, 347–350.
[8] Rautelin, H., Blomberg, B., Fredlund, H., Jarnerot, G. and
Danielsson, D. (1993) Incidence of Helicobacter pylori strains
activating neutrophils in patients with peptic ulcer disease. Gut
34, 599–603.
[9] Satin, B., Del Giudice, G., Della Bianca, V., Dusi, S., Laudanna,
C., Tonello, F., Kelleher, D., Rappuoli, R., Montecucco, C. and
Rossi, F. (2000) The neutrophil-activating protein (HP-NAP) of
Helicobacter pylori is a protective antigen and a major virulence
factor. J. Exp. Med. 191, 1467–1476.
[10] Tonello, F., Dundon, W.G., Satin, B., Molinari, M., Tognon, G.,
Grandi, G., Del Giudice, G., Rappuoli, R. and Montecucco, C.
(1999) The Helicobacter pylori neutrophil-activating protein is an
iron-binding protein with dodecameric structure. Mol. Microbiol.
34, 238–246.
[11] Zanotti, G., Papinutto, E., Dundon, W., Battistutta, R., Seveso,
M., Giudice, G., Rappuoli, R. and Montecucco, C. (2002)
Structure of the neutrophil-activating protein from Helicobacter
pylori. J. Mol. Biol. 323, 125–130.
[12] Thoreson, A.C., Hamlet, A., Celik, J., Bystrom, M., Nystrom, S.,
Olbe, L. and Svennerholm, A.M. (2000) Differences in surface-
exposed antigen expression between Helicobacter pylori strains
isolated from duodenal ulcer patients and from asymptomatic
subjects. J. Clin. Microbiol. 38, 3436–3441.
[13] Namavar, F., Sparrius, M., Veerman, E.C., Appelmelk, B.J. and
Vandenbroucke-Grauls, C.M. (1998) Neutrophil-activating pro-
tein mediates adhesion of Helicobacter pylori to sulfated carbo-
hydrates on high-molecular-weight salivary mucin. Infect.
Immun. 66, 444–447.
[14] Teneberg, S., Miller-Podraza, H., Lampert, H.C., Evans Jr., D.J.,
Evans, D.G., Danielsson, D. and Karlsson, K.A. (1997) Carbo-
hydrate binding specificity of the neutrophil-activating protein of
Helicobacter pylori. J. Biol. Chem. 272, 19067–19071.
[15] Muller, W.A. (1999) Leukocyte-endothelial cell adhesion mole-
cules in transendothelial migration In: Inflammation: Basic
Principles and Clinical Correlates (Gallin, J. and Snyderman,
R., Eds.), 3rd edn. Lippincott Williams & Wilkins, Philadelphia.
[16] Silverstein, R. (2000) The vascular endothelium In: Inflammation:
Basic Principles and Clinical Correlates (Gallin, J. and Snyder-
man, R., Eds.), 3rd edn, pp. 207–227. Lippincott Williams &
Wilkins, Philadelphia.
[17] Innocenti, M., Thoreson, A.C., Ferrero, R.L., Stromberg, E.,
Bolin, I., Eriksson, L., Svennerholm, A.M. and Quiding-Jarbrink,
M. (2002) Helicobacter pylori-induced activation of human
endothelial cells. Infect. Immun. 70, 4581–4590.
[18] Backert, S., Ziska, E., Brinkmann, V., Zimny-Arndt, U., Fau-
connier, A., Jungblut, P.R., Naumann, M. and Meyer, T.F.
(2000) Translocation of the Helicobacter pylori CagA protein in
gastric epithelial cells by a type IV secretion apparatus. Cell
Microbiol. 2, 155–164.
[19] Lee, A., O�Rourke, J., De Ungria, M.C., Robertson, B., Dask-
alopoulos, G. and Dixon, M.F. (1997) A standardized mouse
model of Helicobacter pylori infection: introducing the Sydney
strain. Gastroenterology 112, 1386–1397.
[20] Lindholm, C., Osek, J. and Svennerholm, A.M. (1997) Quanti-
fication of conserved antigens in Helicobacter pylori during
different culture conditions. Infect. Immun. 65, 5376–5380.
[21] Boyum, A. (1968) Isolation of mononuclear cells and granulo-
cytes from human blood. Isolation of monuclear cells by one
centrifugation, and of granulocytes by combining centrifugation
and sedimentation at 1g. Scand. J. Clin. Lab. Invest. Suppl. 97,
77–89.
[22] Klein, C.L., Bittinger, F., Kohler, H., Wagner, M., Otto, M.,
Hermanns, I. and Kirkpatrick, C.J. (1995) Comparative studies
on vascular endothelium in vitro. 3. Effects of cytokines on the
expression of E-selectin, ICAM-1 and VCAM-1 by cultured
human endothelial cells obtained from different passages. Patho-
biology 63, 83–92.
[23] Shen, Y., Sultana, C., Arditi, M., Kim, K.S. and Kalra, V.K.
(1998) Endotoxin-induced migration of monocytes and PECAM-
1 phosphorylation are abrogated by PAF receptor antagonists.
Am. J. Physiol. 275, E479–486.
[24] Broom, M.F., Sherriff, R.M., Munster, D. and Chadwick, V.S.
(1992) Identification of formyl Met-Leu-Phe in culture filtrates of
Helicobacter pylori. Microbios 72, 239–245.
[25] Craig, P.M., Territo, M.C., Karnes, W.E. and Walsh, J.H. (1992)
Helicobacter pylori secretes a chemotactic factor for monocytes
and neutrophils. Gut 33, 1020–1023.
[26] Gebert, B., Fischer, W., Weiss, E., Hoffmann, R. and Haas, R.
(2003) Helicobacter pylori vacuolating cytotoxin inhibits T lym-
phocyte activation. Science 301, 1099–1102.
[27] Shimoyama, T., Everett, S.M., Dixon, M.F., Axon, A.T. and
Crabtree, J.E. (1998) Chemokine mRNA expression in gastric
mucosa is associated with Helicobacter pylori cagA positivity and
severity of gastritis. J. Clin. Pathol. 51, 765–770.
[28] Rieder, G., Einsiedl, W., Hatz, R.A., Stolte, M., Enders, G.A. and
Walz, A. (2001) Comparison of CXC chemokines ENA-78 and
interleukin-8 expression inHelicobacter pylori-associated gastritis.
Infect. Immun. 69, 81–88.
[29] Montemurro, P., Nishioka, H., Dundon, W.G., de Bernard, M.,
Del Giudice, G., Rappuoli, R. and Montecucco, C. (2002) The
neutrophil-activating protein (HP-NAP) of Helicobacter pylori is
a potent stimulant of mast cells. Eur. J. Immunol. 32, 671–676.
[30] Blom, K., Lundin, B.S., Bolin, I. and Svennerholm, A. (2001)
Flow cytometric analysis of the localization of Helicobacter pylori
antigens during different growth phases. FEMS Immunol. Med.
Microbiol. 30, 173–179.




















