
Heat- and light-induced detachment of the light harvesting complex from isolated photosystem I...
9
Heat- and light-induced detachment of the light-harvesting antenna complexes of photosystem I in isolated stroma thylakoid membranes S.B. Krumova a,1 , Zs. Várkonyi a , P.H. Lambrev a , L. Kovács a , S.J. Todinova b , M.C. Busheva b , S.G. Taneva b , G. Garab a,⇑ a Institute of Plant Biology, Biological Research Centre, Hungarian Academy of Sciences, Temesvári krt. 62, H-6726 Szeged, Hungary b Institute of Biophysics and Biomedical Engineering, Bulgarian Academy of Sciences, Acad. G. Bontchev Str., Bl. 21, 1113 Sofia, Bulgaria article info Article history: Received 3 December 2013 Received in revised form 22 April 2014 Accepted 25 April 2014 Available online 16 May 2014 abstract The multisubunit pigment–protein complex of photosystem I (PSI) consists of a core and peripheral light- harvesting antenna (LHCI). PSI is thought to be a rather rigid system and very little is known about its structural and functional flexibility. Recent data, however, suggest LHCI detachment from the PSI super- complex upon heat and light treatments. Furthermore, it was suggested that the splitting off of LHCI acts as a safety valve for PSI core upon photoinhibition (Alboresi et al., 2009). In this work we analyzed the heat- and light-induced reorganizations in isolated PSI vesicles (stroma membrane vesicles enriched in PSI). Using differential scanning calorimetry we revealed a stepwise dis- assembly of PSI supercomplex above 50 °C. Circular dichroism, sucrose gradient centrifugation and 77 K fluorescence experiments identified the sequence of events of PSI destabilization: 3 min heating at 60 °C or 40 min white light illumination at 25 °C resulted in pronounced Lhca1/4 detachment from the PSI supercomplex, which is then followed by the degradation of Lhca2/3. The similarity of the main structural effects due to heat and light treatments supports the notion that thermo-optic mechanism, structural changes induced by ultrafast local thermal transients, which has earlier been shown to be responsible for structural changes in the antenna system of photosystem II, can also regulate the assembly and functioning of PSI antenna. Ó 2014 Elsevier B.V. All rights reserved. 1. Introduction The multisubunit pigment–protein complex of photosystem I (PSI) is incorporated in the stroma thylakoid membranes and the end membranes of grana. The overall function of PSI involves har- vesting photons and using their energy for electron transfer through a series of redox centers. Although laterally separated, the functioning of photosystem II (PSII) and PSI is coupled via an electron transport chain involving plastoquinoine, cytochrome b 6 f and plastocyanine molecules, which finally direct the electrons toward ferredoxin and lead to NADP reduction and ATP synthesis [1]. PSI is also able to adjust the ratio between ATP synthesis and NADP reduction to the needs of the cells, via cyclic transport of electrons [2]. The crystal structure of plant PSI supercomplex has been solved and in higher plants it is found to be in a monomeric state [3–6]. The reaction center consists of up to 14 subunits. In higher plants, typically four different light-harvesting membrane proteins (LHCIs), Lhca1-4, assembled in a half-moon shape, are asymmetri- cally attached to it [3,6]; they are organized in two dimer types in equal amounts – Lhca1/4 and Lhca2/3 [3,5–7]. LHCI proteins are unique among the chlorophyll (Chl) a/b binding proteins in their red-shifted absorbance and in the formation of dimers [7]. Two other Lhca genes, Lhca5 and Lhca6, have also been identified [8], however they seem to be only minor components of the PSI antenna and probably play a role under specific environmental conditions. PSI is also able to interact (via transient attachment) with the major light-harvesting complex of PSII, LHCII, upon state transitions, thus enlarging its effective antenna [9,10]. In contrast to the supercomplex of PSII and its associated light harvesting antenna system, which form large ordered domains and possess a remarkable structural flexibility associated with sev- eral regulatory processes [11–13], PSI is thought to be rather rigid system and very little is known about its structural and functional flexibility [14]. In higher plants it does not change its oligomeriza- tion state and the docking sites for the individual subunits are highly specific [15]. It was established that the interaction between Lhca subunits and the PSI core is strongly cooperative and when one subunit is missing, the whole LHCI system is destabilized http://dx.doi.org/10.1016/j.jphotobiol.2014.04.029 1011-1344/Ó 2014 Elsevier B.V. All rights reserved. ⇑ Corresponding author. Tel.: +36 62 599 709; fax: +36 62 433 434. E-mail address: [email protected] (G. Garab). 1 Permanent address: Institute of Biophysics and Biomedical Engineering, Bulgarian Academy of Sciences, Acad. G. Bontchev Str., Bl. 21, 1113 Sofia, Bulgaria. Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12 Contents lists available at ScienceDirect Journal of Photochemistry and Photobiology B: Biology journal homepage: www.elsevier.com/locate/jphotobiol
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Heat- and light-induced detachment of the light harvesting complex from isolated photosystem I...

Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12
Contents lists available at ScienceDirect
Journal of Photochemistry and Photobiology B: Biology
journal homepage: www.elsevier .com/locate / jphotobiol
Heat- and light-induced detachment of the light-harvesting antennacomplexes of photosystem I in isolated stroma thylakoid membranes
http://dx.doi.org/10.1016/j.jphotobiol.2014.04.0291011-1344/� 2014 Elsevier B.V. All rights reserved.
⇑ Corresponding author. Tel.: +36 62 599 709; fax: +36 62 433 434.E-mail address: [email protected] (G. Garab).
1 Permanent address: Institute of Biophysics and Biomedical Engineering, BulgarianAcademy of Sciences, Acad. G. Bontchev Str., Bl. 21, 1113 Sofia, Bulgaria.
S.B. Krumova a,1, Zs. Várkonyi a, P.H. Lambrev a, L. Kovács a, S.J. Todinova b, M.C. Busheva b, S.G. Taneva b,G. Garab a,⇑a Institute of Plant Biology, Biological Research Centre, Hungarian Academy of Sciences, Temesvári krt. 62, H-6726 Szeged, Hungaryb Institute of Biophysics and Biomedical Engineering, Bulgarian Academy of Sciences, Acad. G. Bontchev Str., Bl. 21, 1113 Sofia, Bulgaria
a r t i c l e i n f o
Article history:Received 3 December 2013Received in revised form 22 April 2014Accepted 25 April 2014Available online 16 May 2014
a b s t r a c t
The multisubunit pigment–protein complex of photosystem I (PSI) consists of a core and peripheral light-harvesting antenna (LHCI). PSI is thought to be a rather rigid system and very little is known about itsstructural and functional flexibility. Recent data, however, suggest LHCI detachment from the PSI super-complex upon heat and light treatments. Furthermore, it was suggested that the splitting off of LHCI actsas a safety valve for PSI core upon photoinhibition (Alboresi et al., 2009).
In this work we analyzed the heat- and light-induced reorganizations in isolated PSI vesicles (stromamembrane vesicles enriched in PSI). Using differential scanning calorimetry we revealed a stepwise dis-assembly of PSI supercomplex above 50 �C. Circular dichroism, sucrose gradient centrifugation and 77 Kfluorescence experiments identified the sequence of events of PSI destabilization: 3 min heating at 60 �Cor 40 min white light illumination at 25 �C resulted in pronounced Lhca1/4 detachment from the PSIsupercomplex, which is then followed by the degradation of Lhca2/3.
The similarity of the main structural effects due to heat and light treatments supports the notion thatthermo-optic mechanism, structural changes induced by ultrafast local thermal transients, which hasearlier been shown to be responsible for structural changes in the antenna system of photosystem II,can also regulate the assembly and functioning of PSI antenna.
� 2014 Elsevier B.V. All rights reserved.
1. Introduction
The multisubunit pigment–protein complex of photosystem I(PSI) is incorporated in the stroma thylakoid membranes and theend membranes of grana. The overall function of PSI involves har-vesting photons and using their energy for electron transferthrough a series of redox centers. Although laterally separated,the functioning of photosystem II (PSII) and PSI is coupled via anelectron transport chain involving plastoquinoine, cytochromeb6f and plastocyanine molecules, which finally direct the electronstoward ferredoxin and lead to NADP reduction and ATP synthesis[1]. PSI is also able to adjust the ratio between ATP synthesis andNADP reduction to the needs of the cells, via cyclic transport ofelectrons [2].
The crystal structure of plant PSI supercomplex has been solvedand in higher plants it is found to be in a monomeric state [3–6].The reaction center consists of up to 14 subunits. In higher plants,
typically four different light-harvesting membrane proteins(LHCIs), Lhca1-4, assembled in a half-moon shape, are asymmetri-cally attached to it [3,6]; they are organized in two dimer types inequal amounts – Lhca1/4 and Lhca2/3 [3,5–7]. LHCI proteins areunique among the chlorophyll (Chl) a/b binding proteins in theirred-shifted absorbance and in the formation of dimers [7]. Twoother Lhca genes, Lhca5 and Lhca6, have also been identified [8],however they seem to be only minor components of the PSIantenna and probably play a role under specific environmentalconditions. PSI is also able to interact (via transient attachment)with the major light-harvesting complex of PSII, LHCII, upon statetransitions, thus enlarging its effective antenna [9,10].
In contrast to the supercomplex of PSII and its associated lightharvesting antenna system, which form large ordered domainsand possess a remarkable structural flexibility associated with sev-eral regulatory processes [11–13], PSI is thought to be rather rigidsystem and very little is known about its structural and functionalflexibility [14]. In higher plants it does not change its oligomeriza-tion state and the docking sites for the individual subunits arehighly specific [15]. It was established that the interaction betweenLhca subunits and the PSI core is strongly cooperative and whenone subunit is missing, the whole LHCI system is destabilized

S.B. Krumova et al. / Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12 5
[16]. Nevertheless, it was shown that the composition of PSIperipheral antenna varies with long-term changes of the lightintensity [17]. Furthermore, there are data strongly suggesting thatLHCI is able to disconnect energetically from the core complexupon illumination [18] and heating to 57–60 �C [19]. Heat-inducedenergetic disconnection of LHCI at 55–70 �C was found also inleaves [20] and isolated chloroplasts [21].
This work explores the structural changes and aims at identify-ing the sequence of events occurring in isolated PSI stroma thyla-koid vesicles upon elevating the temperature as well as underwhite light irradiation at 25 �C. In the accompanying paper, similarexperiments are carried out on isolated PSI–LHCI supercomplexes.
2. Materials and methods
2.1. Isolation of PSI stroma thylakoid vesicles
PSI-enriched (129,000 g) subchloroplast fraction (PSI vesiclesformed by stroma membranes devoid of PSII components) was iso-lated by digitonin digestion of spinach chloroplasts, followed bydifferential centrifugation as in Peters et al. [22], and stored at�20 �C until use. For all measurements PSI vesicles were resus-pended in a reaction medium containing 20 mM tricine (pH 7.8),25 mM KCl, 25 mM NaCl, 5 mM MgCl2 and 250 mM sorbitol. TheChl a/b ratio was about 10 (determined by the method of Porraet al. [23] in all preparations). This value agrees well with the Chla/b ratio of 9.7 given for wild type PSI–LHCI supercomplex [15].It can be calculated that with LHCII trimer associated with every10th and every 5th supercomplex, this ratio would decrease to8.9 and 8.1, respectively. Assuming an up to 10% error in the absor-bance measurement, one can rule out the presence of significantamounts of LHCII in our stroma thylakoid membrane preparation.
2.2. Heating protocols
Two heating protocols were used: (i) gradual heating from 25 to90 �C in 5 �C steps, combined with 3 min incubation at the desiredtemperature (this heating protocol is similar to the linear heatingin DSC experiments described in Section 2.5), and (ii) short heattreatment, where aliquots of stock solution were kept at 25 �C andincubated for 5 min at the given temperature between 30 and90 �C. For each treatment aliquots of 100 ll suspension of PSIvesicles were incubated at desired temperatures in a block heater.
2.3. Illumination
Thin layer (�3.5 mm) of PSI vesicles suspension at a concentra-tion of 200 lg Chl/ml was illuminated at room temperature in a0.6 cm diameter glass tube with heat-filtered white light (1150lmol photons m�2 s�1 at the surface, �150 lmol photons m�2 s�1
average intensity) and constantly stirred during the illumination.
2.4. Sucrose gradient centrifugation
Seven-step discontinuous sucrose gradient (0.1–0.96 M) wasused. Two hundred ll aliquots of PSI vesicles at a Chl concentrationof 200 lg Chl/ml were incubated at the given temperatures usingthe heating protocols described in Section 2.2. The samples were sol-ubilized for 20 min with 1% dodecylmaltoside and loaded on thesucrose gradient. The gradients were centrifuged in a Sorvall swingout, TH-641 rotor at 180,000g for 17 h. After centrifugation, severalbands were discernible and the chlorophyll content in each bandwas estimated from digital photographic images of the centrifugetubes. To this end, raw camera images were processed withoutgamma correction. Red-channel brightness values from the area
inside the tubes were integrated horizontally using ImageJ(www.imagej.net) or MATLAB (MathWorks) and converted to opti-cal density as OD = �logRy/R0 where Ry is brightness along the ver-tical coordinate and R0 is baseline brightness. Bands in the obtaineddensitograms were then integrated using Gaussian fit. The methodwas tested by measuring the densities of acetone extracts ofpigments at different concentrations and comparing with spectro-photometric measurements ensuring that the density estimatedfrom the photographs depends linearly on pigment concentration.
2.5. DSC measurements
DSC scans in the temperature range between 20 and 100 �Cwere measured at a heating rate of 1 �C/min with a high-sensitivityscanning microcalorimeter DASM4 (Biopribor, Pushchino, Russia).The concentration of the PSI vesicles was 1 mg Chl/ml. The calori-metric transition temperatures (the temperature at the maximaof the excess heat capacity curve), was determined by mathemat-ical deconvolution of the thermogram by applying Gaussian fittingas in Dobrikova et al. [24]. For each scan a buffer–buffer scan wassubtracted, followed by subtraction of a linear-baseline fit in orderto mathematically deconvolve the thermograms. Four independentmeasurements were performed and the standard error for eachtransition temperature is estimated.
2.6. Circular dichroism measurements
Circular dichroism (CD) spectra were measured between 400and 750 nm in a Jobin-Yvon CD6 dichrograph equipped with athermostable sample holder. The Chl content of the suspensionof PSI vesicles was adjusted to 20 lg/ml. The CD spectra of the dif-ferent sucrose gradient fractions are normalized to the maximumof the red-most absorption band (672 nm for the upper-most gra-dient fraction, and 680 nm for the two lower fractions, respec-tively). The optical pathlength of the cell was 1 cm, and thedistance of the sample from the photomultiplier was 5 cm. Thespectra were recorded in 0.5 nm steps with an integration timeof 0.1 s and a bandpass of 1 nm.
The temperature and light dependence of the selected CDbands, at (�)491/(+)510 nm, (�)650 nm and (�)684 nm, weredetermined by measuring the difference in the CD amplitudes at491 and 510 nm, and for the case of (�)650 nm and (�)681 nmCD bands – the difference between these wavelengths and the ref-erence wavelengths at 620 and 730 nm, respectively. The transi-tion temperature (Tm) was determined as the temperature atwhich the selected CD band had half intensity.
2.7. Low temperature fluorescence spectroscopy
100 ll of PSI vesicles pretreated for 3 min at different tempera-tures or illuminated for different time periods at 25 �C were loadedin a homemade sample holder and were frozen in liquid nitrogen.The sample thickness was 1 mm. The 77 K emission spectra wererecorded on a Jobin-Yvon Horiba Fluorolog-3 spectrofluorimeterusing excitation and emission slits of 7 and 5 nm, respectively,and were corrected for spectral sensitivity of the detection system.The presented spectra are excited at 436 nm and normalized to theabsorption at 680 nm.
3. Results
3.1. Thermal stability of PSI vesicles
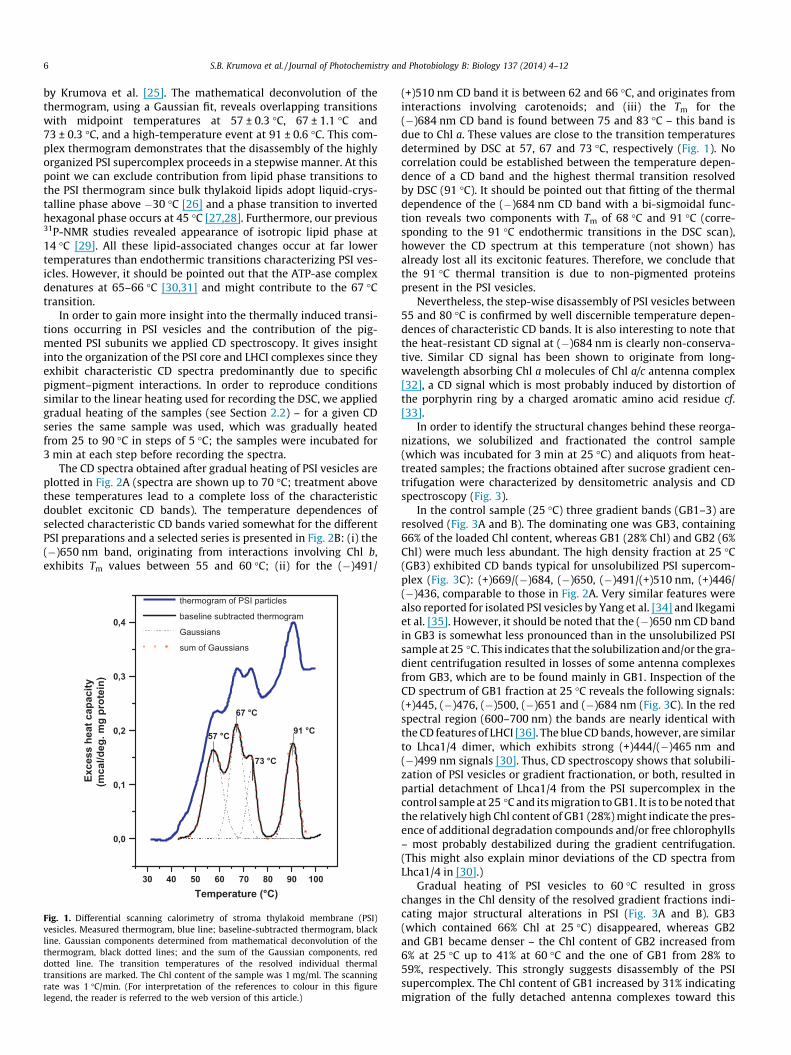
3.1.1. Gradual heatingThe thermogram of PSI vesicles (Fig. 1) reveals several endo-
thermic transitions between 40 and 100 �C, similar to that reported
6 S.B. Krumova et al. / Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12
by Krumova et al. [25]. The mathematical deconvolution of thethermogram, using a Gaussian fit, reveals overlapping transitionswith midpoint temperatures at 57 ± 0.3 �C, 67 ± 1.1 �C and73 ± 0.3 �C, and a high-temperature event at 91 ± 0.6 �C. This com-plex thermogram demonstrates that the disassembly of the highlyorganized PSI supercomplex proceeds in a stepwise manner. At thispoint we can exclude contribution from lipid phase transitions tothe PSI thermogram since bulk thylakoid lipids adopt liquid-crys-talline phase above �30 �C [26] and a phase transition to invertedhexagonal phase occurs at 45 �C [27,28]. Furthermore, our previous31P-NMR studies revealed appearance of isotropic lipid phase at14 �C [29]. All these lipid-associated changes occur at far lowertemperatures than endothermic transitions characterizing PSI ves-icles. However, it should be pointed out that the ATP-ase complexdenatures at 65–66 �C [30,31] and might contribute to the 67 �Ctransition.
In order to gain more insight into the thermally induced transi-tions occurring in PSI vesicles and the contribution of the pig-mented PSI subunits we applied CD spectroscopy. It gives insightinto the organization of the PSI core and LHCI complexes since theyexhibit characteristic CD spectra predominantly due to specificpigment–pigment interactions. In order to reproduce conditionssimilar to the linear heating used for recording the DSC, we appliedgradual heating of the samples (see Section 2.2) – for a given CDseries the same sample was used, which was gradually heatedfrom 25 to 90 �C in steps of 5 �C; the samples were incubated for3 min at each step before recording the spectra.
The CD spectra obtained after gradual heating of PSI vesicles areplotted in Fig. 2A (spectra are shown up to 70 �C; treatment abovethese temperatures lead to a complete loss of the characteristicdoublet excitonic CD bands). The temperature dependences ofselected characteristic CD bands varied somewhat for the differentPSI preparations and a selected series is presented in Fig. 2B: (i) the(�)650 nm band, originating from interactions involving Chl b,exhibits Tm values between 55 and 60 �C; (ii) for the (�)491/
30 40 50 60 70 80 90 100
0,0
0,1
0,2
0,3
0,4
thermogram of PSI particles
baseline subtracted thermogram
Gaussians
sum of Gaussians
67 °C
57 °C
Exce
ss h
eat c
apac
ity(m
cal/d
eg. m
g pr
otei
n)
Temperature (°C)
73 °C
91 °C
Fig. 1. Differential scanning calorimetry of stroma thylakoid membrane (PSI)vesicles. Measured thermogram, blue line; baseline-subtracted thermogram, blackline. Gaussian components determined from mathematical deconvolution of thethermogram, black dotted lines; and the sum of the Gaussian components, reddotted line. The transition temperatures of the resolved individual thermaltransitions are marked. The Chl content of the sample was 1 mg/ml. The scanningrate was 1 �C/min. (For interpretation of the references to colour in this figurelegend, the reader is referred to the web version of this article.)
(+)510 nm CD band it is between 62 and 66 �C, and originates frominteractions involving carotenoids; and (iii) the Tm for the(�)684 nm CD band is found between 75 and 83 �C – this band isdue to Chl a. These values are close to the transition temperaturesdetermined by DSC at 57, 67 and 73 �C, respectively (Fig. 1). Nocorrelation could be established between the temperature depen-dence of a CD band and the highest thermal transition resolvedby DSC (91 �C). It should be pointed out that fitting of the thermaldependence of the (�)684 nm CD band with a bi-sigmoidal func-tion reveals two components with Tm of 68 �C and 91 �C (corre-sponding to the 91 �C endothermic transitions in the DSC scan),however the CD spectrum at this temperature (not shown) hasalready lost all its excitonic features. Therefore, we conclude thatthe 91 �C thermal transition is due to non-pigmented proteinspresent in the PSI vesicles.
Nevertheless, the step-wise disassembly of PSI vesicles between55 and 80 �C is confirmed by well discernible temperature depen-dences of characteristic CD bands. It is also interesting to note thatthe heat-resistant CD signal at (�)684 nm is clearly non-conserva-tive. Similar CD signal has been shown to originate from long-wavelength absorbing Chl a molecules of Chl a/c antenna complex[32], a CD signal which is most probably induced by distortion ofthe porphyrin ring by a charged aromatic amino acid residue cf.[33].
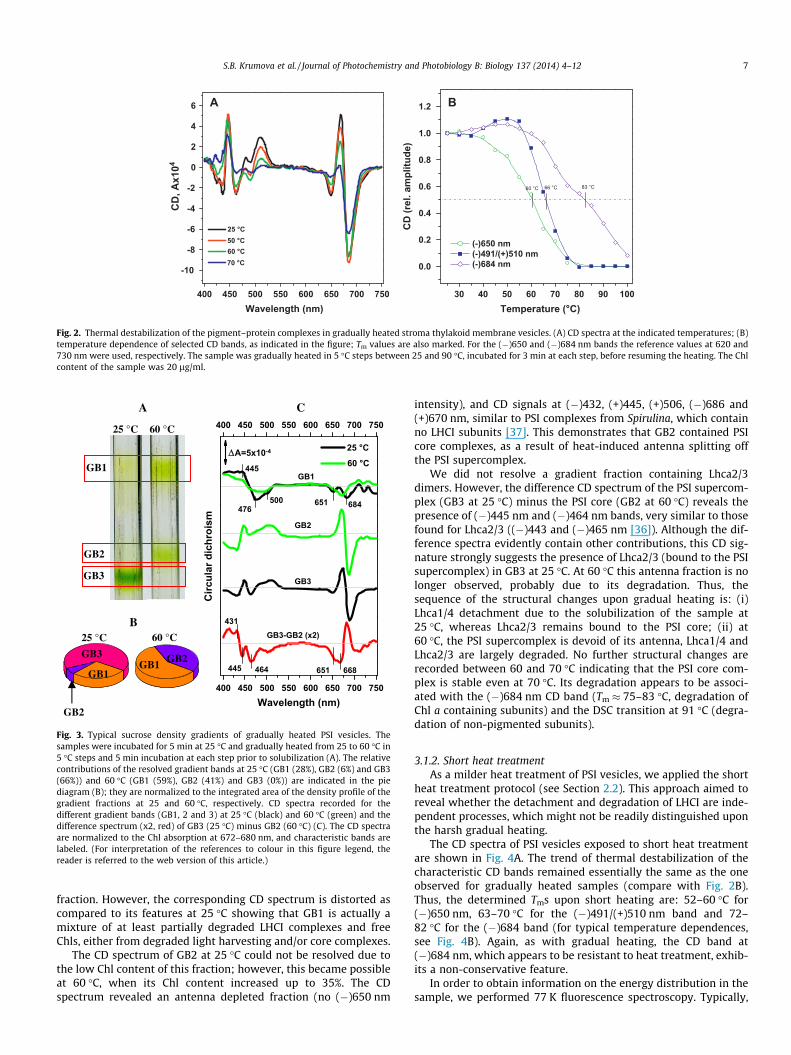
In order to identify the structural changes behind these reorga-nizations, we solubilized and fractionated the control sample(which was incubated for 3 min at 25 �C) and aliquots from heat-treated samples; the fractions obtained after sucrose gradient cen-trifugation were characterized by densitometric analysis and CDspectroscopy (Fig. 3).
In the control sample (25 �C) three gradient bands (GB1–3) areresolved (Fig. 3A and B). The dominating one was GB3, containing66% of the loaded Chl content, whereas GB1 (28% Chl) and GB2 (6%Chl) were much less abundant. The high density fraction at 25 �C(GB3) exhibited CD bands typical for unsolubilized PSI supercom-plex (Fig. 3C): (+)669/(�)684, (�)650, (�)491/(+)510 nm, (+)446/(�)436, comparable to those in Fig. 2A. Very similar features werealso reported for isolated PSI vesicles by Yang et al. [34] and Ikegamiet al. [35]. However, it should be noted that the (�)650 nm CD bandin GB3 is somewhat less pronounced than in the unsolubilized PSIsample at 25 �C. This indicates that the solubilization and/or the gra-dient centrifugation resulted in losses of some antenna complexesfrom GB3, which are to be found mainly in GB1. Inspection of theCD spectrum of GB1 fraction at 25 �C reveals the following signals:(+)445, (�)476, (�)500, (�)651 and (�)684 nm (Fig. 3C). In the redspectral region (600–700 nm) the bands are nearly identical withthe CD features of LHCI [36]. The blue CD bands, however, are similarto Lhca1/4 dimer, which exhibits strong (+)444/(�)465 nm and(�)499 nm signals [30]. Thus, CD spectroscopy shows that solubili-zation of PSI vesicles or gradient fractionation, or both, resulted inpartial detachment of Lhca1/4 from the PSI supercomplex in thecontrol sample at 25 �C and its migration to GB1. It is to be noted thatthe relatively high Chl content of GB1 (28%) might indicate the pres-ence of additional degradation compounds and/or free chlorophylls– most probably destabilized during the gradient centrifugation.(This might also explain minor deviations of the CD spectra fromLhca1/4 in [30].)
Gradual heating of PSI vesicles to 60 �C resulted in grosschanges in the Chl density of the resolved gradient fractions indi-cating major structural alterations in PSI (Fig. 3A and B). GB3(which contained 66% Chl at 25 �C) disappeared, whereas GB2and GB1 became denser – the Chl content of GB2 increased from6% at 25 �C up to 41% at 60 �C and the one of GB1 from 28% to59%, respectively. This strongly suggests disassembly of the PSIsupercomplex. The Chl content of GB1 increased by 31% indicatingmigration of the fully detached antenna complexes toward this
30 40 50 60 70 80 90 100
0.0
0.2
0.4
0.6
0.8
1.0
1.2
(-)650 nm (-)491/(+)510 nm (-)684 nm
B
CD
(rel
. am
plitu
de)
Temperature (°C)
60 °C
400 450 500 550 600 650 700 750
-10
-8
-6
-4
-2
0
2
4
6
25 °C
A
CD
, Ax1
04
Wavelength (nm)
50 °C 60 °C 70 °C
66 °C 83 °C
Fig. 2. Thermal destabilization of the pigment–protein complexes in gradually heated stroma thylakoid membrane vesicles. (A) CD spectra at the indicated temperatures; (B)temperature dependence of selected CD bands, as indicated in the figure; Tm values are also marked. For the (�)650 and (�)684 nm bands the reference values at 620 and730 nm were used, respectively. The sample was gradually heated in 5 �C steps between 25 and 90 �C, incubated for 3 min at each step, before resuming the heating. The Chlcontent of the sample was 20 lg/ml.
A C 400 450 500 550 600 650 700 750
GB3-GB2 (x2)
GB3
GB2
25 °C
Circ
ular
dic
hroi
sm
ΔA=5x10-4
Wavelength (nm)
445
476500 651 684
431
445 464 651 668
GB1GB1
GB2
GB3
25 °C 60 °C
B
GB1
GB2
GB2 GB3 GB1
400 450 500 550 600 650 700 750
25 °C 60 °C
60 °C
Fig. 3. Typical sucrose density gradients of gradually heated PSI vesicles. Thesamples were incubated for 5 min at 25 �C and gradually heated from 25 to 60 �C in5 �C steps and 5 min incubation at each step prior to solubilization (A). The relativecontributions of the resolved gradient bands at 25 �C (GB1 (28%), GB2 (6%) and GB3(66%)) and 60 �C (GB1 (59%), GB2 (41%) and GB3 (0%)) are indicated in the piediagram (B); they are normalized to the integrated area of the density profile of thegradient fractions at 25 and 60 �C, respectively. CD spectra recorded for thedifferent gradient bands (GB1, 2 and 3) at 25 �C (black) and 60 �C (green) and thedifference spectrum (x2, red) of GB3 (25 �C) minus GB2 (60 �C) (C). The CD spectraare normalized to the Chl absorption at 672–680 nm, and characteristic bands arelabeled. (For interpretation of the references to colour in this figure legend, thereader is referred to the web version of this article.)
S.B. Krumova et al. / Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12 7
fraction. However, the corresponding CD spectrum is distorted ascompared to its features at 25 �C showing that GB1 is actually amixture of at least partially degraded LHCI complexes and freeChls, either from degraded light harvesting and/or core complexes.
The CD spectrum of GB2 at 25 �C could not be resolved due tothe low Chl content of this fraction; however, this became possibleat 60 �C, when its Chl content increased up to 35%. The CDspectrum revealed an antenna depleted fraction (no (�)650 nm
intensity), and CD signals at (�)432, (+)445, (+)506, (�)686 and(+)670 nm, similar to PSI complexes from Spirulina, which containno LHCI subunits [37]. This demonstrates that GB2 contained PSIcore complexes, as a result of heat-induced antenna splitting offthe PSI supercomplex.
We did not resolve a gradient fraction containing Lhca2/3dimers. However, the difference CD spectrum of the PSI supercom-plex (GB3 at 25 �C) minus the PSI core (GB2 at 60 �C) reveals thepresence of (�)445 nm and (�)464 nm bands, very similar to thosefound for Lhca2/3 ((�)443 and (�)465 nm [36]). Although the dif-ference spectra evidently contain other contributions, this CD sig-nature strongly suggests the presence of Lhca2/3 (bound to the PSIsupercomplex) in GB3 at 25 �C. At 60 �C this antenna fraction is nolonger observed, probably due to its degradation. Thus, thesequence of the structural changes upon gradual heating is: (i)Lhca1/4 detachment due to the solubilization of the sample at25 �C, whereas Lhca2/3 remains bound to the PSI core; (ii) at60 �C, the PSI supercomplex is devoid of its antenna, Lhca1/4 andLhca2/3 are largely degraded. No further structural changes arerecorded between 60 and 70 �C indicating that the PSI core com-plex is stable even at 70 �C. Its degradation appears to be associ-ated with the (�)684 nm CD band (Tm � 75–83 �C, degradation ofChl a containing subunits) and the DSC transition at 91 �C (degra-dation of non-pigmented subunits).
3.1.2. Short heat treatmentAs a milder heat treatment of PSI vesicles, we applied the short
heat treatment protocol (see Section 2.2). This approach aimed toreveal whether the detachment and degradation of LHCI are inde-pendent processes, which might not be readily distinguished uponthe harsh gradual heating.
The CD spectra of PSI vesicles exposed to short heat treatmentare shown in Fig. 4A. The trend of thermal destabilization of thecharacteristic CD bands remained essentially the same as the oneobserved for gradually heated samples (compare with Fig. 2B).Thus, the determined Tms upon short heating are: 52–60 �C for(�)650 nm, 63–70 �C for the (�)491/(+)510 nm band and 72–82 �C for the (�)684 band (for typical temperature dependences,see Fig. 4B). Again, as with gradual heating, the CD band at(�)684 nm, which appears to be resistant to heat treatment, exhib-its a non-conservative feature.
In order to obtain information on the energy distribution in thesample, we performed 77 K fluorescence spectroscopy. Typically,
A
GB1
GB2
GB3
C
GB3-GB2 (x2)
GB3
GB2
25 °C
686651
493
Circ
ular
dic
hroi
sm
445
470
Wavelength (nm)
ΔA=5x10-4
432
444 463 656 678
GB1
GB1 GB2 GB3 GB2
GB1
B
GB3
60 °C70 °C
450 500 550 600 650 700
450 500 550 600 650 700
60 °C 70 °C
60 °C 70 °C
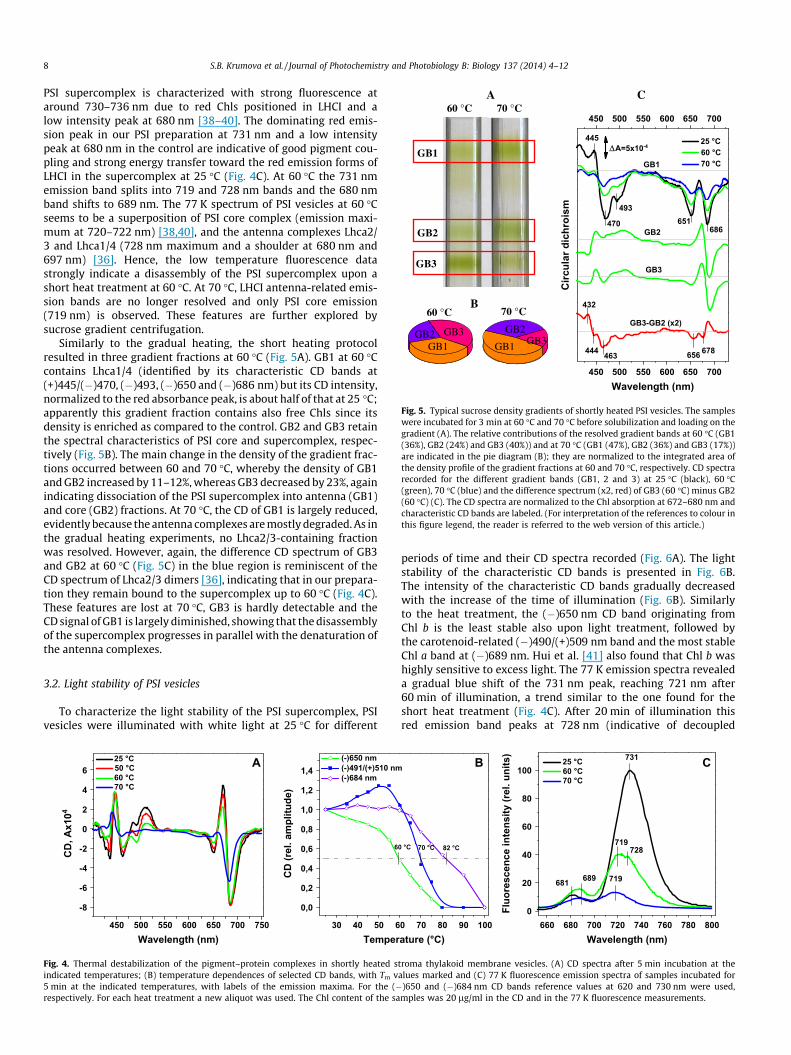
Fig. 5. Typical sucrose density gradients of shortly heated PSI vesicles. The sampleswere incubated for 3 min at 60 �C and 70 �C before solubilization and loading on thegradient (A). The relative contributions of the resolved gradient bands at 60 �C (GB1(36%), GB2 (24%) and GB3 (40%)) and at 70 �C (GB1 (47%), GB2 (36%) and GB3 (17%))are indicated in the pie diagram (B); they are normalized to the integrated area ofthe density profile of the gradient fractions at 60 and 70 �C, respectively. CD spectrarecorded for the different gradient bands (GB1, 2 and 3) at 25 �C (black), 60 �C(green), 70 �C (blue) and the difference spectrum (x2, red) of GB3 (60 �C) minus GB2(60 �C) (C). The CD spectra are normalized to the Chl absorption at 672–680 nm andcharacteristic CD bands are labeled. (For interpretation of the references to colour inthis figure legend, the reader is referred to the web version of this article.)
8 S.B. Krumova et al. / Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12
PSI supercomplex is characterized with strong fluorescence ataround 730–736 nm due to red Chls positioned in LHCI and alow intensity peak at 680 nm [38–40]. The dominating red emis-sion peak in our PSI preparation at 731 nm and a low intensitypeak at 680 nm in the control are indicative of good pigment cou-pling and strong energy transfer toward the red emission forms ofLHCI in the supercomplex at 25 �C (Fig. 4C). At 60 �C the 731 nmemission band splits into 719 and 728 nm bands and the 680 nmband shifts to 689 nm. The 77 K spectrum of PSI vesicles at 60 �Cseems to be a superposition of PSI core complex (emission maxi-mum at 720–722 nm) [38,40], and the antenna complexes Lhca2/3 and Lhca1/4 (728 nm maximum and a shoulder at 680 nm and697 nm) [36]. Hence, the low temperature fluorescence datastrongly indicate a disassembly of the PSI supercomplex upon ashort heat treatment at 60 �C. At 70 �C, LHCI antenna-related emis-sion bands are no longer resolved and only PSI core emission(719 nm) is observed. These features are further explored bysucrose gradient centrifugation.
Similarly to the gradual heating, the short heating protocolresulted in three gradient fractions at 60 �C (Fig. 5A). GB1 at 60 �Ccontains Lhca1/4 (identified by its characteristic CD bands at(+)445/(�)470, (�)493, (�)650 and (�)686 nm) but its CD intensity,normalized to the red absorbance peak, is about half of that at 25 �C;apparently this gradient fraction contains also free Chls since itsdensity is enriched as compared to the control. GB2 and GB3 retainthe spectral characteristics of PSI core and supercomplex, respec-tively (Fig. 5B). The main change in the density of the gradient frac-tions occurred between 60 and 70 �C, whereby the density of GB1and GB2 increased by 11–12%, whereas GB3 decreased by 23%, againindicating dissociation of the PSI supercomplex into antenna (GB1)and core (GB2) fractions. At 70 �C, the CD of GB1 is largely reduced,evidently because the antenna complexes are mostly degraded. As inthe gradual heating experiments, no Lhca2/3-containing fractionwas resolved. However, again, the difference CD spectrum of GB3and GB2 at 60 �C (Fig. 5C) in the blue region is reminiscent of theCD spectrum of Lhca2/3 dimers [36], indicating that in our prepara-tion they remain bound to the supercomplex up to 60 �C (Fig. 4C).These features are lost at 70 �C, GB3 is hardly detectable and theCD signal of GB1 is largely diminished, showing that the disassemblyof the supercomplex progresses in parallel with the denaturation ofthe antenna complexes.
3.2. Light stability of PSI vesicles
To characterize the light stability of the PSI supercomplex, PSIvesicles were illuminated with white light at 25 �C for different
-8
-6
-4
-2
0
2
4
625 °C A
CD
, Ax1
04
Wavelength (nm)450 500 550 600 650 700 750 30 40 50 6
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4(-)650 nm(-)491/(+)510 nm(-)684 nm
CD
(rel
. am
plitu
de)
Temper
50 °C 60 °C 70 °C
6
Fig. 4. Thermal destabilization of the pigment–protein complexes in shortly heated sindicated temperatures; (B) temperature dependences of selected CD bands, with Tm v5 min at the indicated temperatures, with labels of the emission maxima. For the (�respectively. For each heat treatment a new aliquot was used. The Chl content of the sa
periods of time and their CD spectra recorded (Fig. 6A). The lightstability of the characteristic CD bands is presented in Fig. 6B.The intensity of the characteristic CD bands gradually decreasedwith the increase of the time of illumination (Fig. 6B). Similarlyto the heat treatment, the (�)650 nm CD band originating fromChl b is the least stable also upon light treatment, followed bythe carotenoid-related (�)490/(+)509 nm band and the most stableChl a band at (�)689 nm. Hui et al. [41] also found that Chl b washighly sensitive to excess light. The 77 K emission spectra revealeda gradual blue shift of the 731 nm peak, reaching 721 nm after60 min of illumination, a trend similar to the one found for theshort heat treatment (Fig. 4C). After 20 min of illumination thisred emission band peaks at 728 nm (indicative of decoupled
0
20
40
60
80
100
Fluo
resc
ence
inte
nsity
(rel
. uni
ts)
Wavelength (nm)
C731
728719
719689681
660 680 700 720 740 760 780 8000 70 80 90 100
B
ature (°C)
25 °C 60 °C 70 °C
0 °C 70 °C 82 °C
troma thylakoid membrane vesicles. (A) CD spectra after 5 min incubation at thealues marked and (C) 77 K fluorescence emission spectra of samples incubated for)650 and (�)684 nm CD bands reference values at 620 and 730 nm were used,mples was 20 lg/ml in the CD and in the 77 K fluorescence measurements.
GB3-GB2 (x2)
GB3
GB2
dark 20 min light 40 min light
Circ
ular
dic
hroi
sm
443
474498
ΔA=5x10-4
Wavelength (nm)
GB1
432
444466 502 654 670
651
651684
40 min dark
GB1
GB2
GB3
A C
GB1
GB2
GB3
dark 40 min B
23.23% GB1 GB2
GB3
450400 500 550 600 650 750700
450400 500 550 600 650 750700
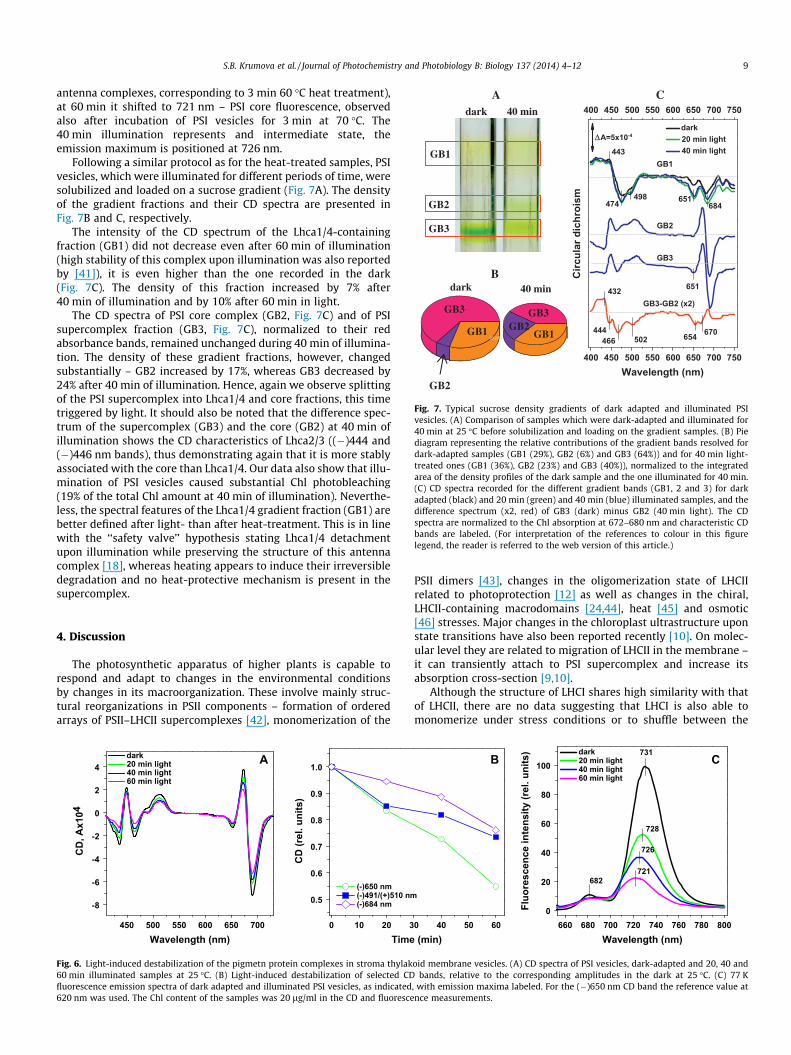
Fig. 7. Typical sucrose density gradients of dark adapted and illuminated PSIvesicles. (A) Comparison of samples which were dark-adapted and illuminated for40 min at 25 �C before solubilization and loading on the gradient samples. (B) Piediagram representing the relative contributions of the gradient bands resolved fordark-adapted samples (GB1 (29%), GB2 (6%) and GB3 (64%)) and for 40 min light-treated ones (GB1 (36%), GB2 (23%) and GB3 (40%)), normalized to the integratedarea of the density profiles of the dark sample and the one illuminated for 40 min.(C) CD spectra recorded for the different gradient bands (GB1, 2 and 3) for darkadapted (black) and 20 min (green) and 40 min (blue) illuminated samples, and thedifference spectrum (x2, red) of GB3 (dark) minus GB2 (40 min light). The CDspectra are normalized to the Chl absorption at 672–680 nm and characteristic CDbands are labeled. (For interpretation of the references to colour in this figurelegend, the reader is referred to the web version of this article.)
S.B. Krumova et al. / Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12 9
antenna complexes, corresponding to 3 min 60 �C heat treatment),at 60 min it shifted to 721 nm – PSI core fluorescence, observedalso after incubation of PSI vesicles for 3 min at 70 �C. The40 min illumination represents and intermediate state, theemission maximum is positioned at 726 nm.
Following a similar protocol as for the heat-treated samples, PSIvesicles, which were illuminated for different periods of time, weresolubilized and loaded on a sucrose gradient (Fig. 7A). The densityof the gradient fractions and their CD spectra are presented inFig. 7B and C, respectively.
The intensity of the CD spectrum of the Lhca1/4-containingfraction (GB1) did not decrease even after 60 min of illumination(high stability of this complex upon illumination was also reportedby [41]), it is even higher than the one recorded in the dark(Fig. 7C). The density of this fraction increased by 7% after40 min of illumination and by 10% after 60 min in light.
The CD spectra of PSI core complex (GB2, Fig. 7C) and of PSIsupercomplex fraction (GB3, Fig. 7C), normalized to their redabsorbance bands, remained unchanged during 40 min of illumina-tion. The density of these gradient fractions, however, changedsubstantially – GB2 increased by 17%, whereas GB3 decreased by24% after 40 min of illumination. Hence, again we observe splittingof the PSI supercomplex into Lhca1/4 and core fractions, this timetriggered by light. It should also be noted that the difference spec-trum of the supercomplex (GB3) and the core (GB2) at 40 min ofillumination shows the CD characteristics of Lhca2/3 ((�)444 and(�)446 nm bands), thus demonstrating again that it is more stablyassociated with the core than Lhca1/4. Our data also show that illu-mination of PSI vesicles caused substantial Chl photobleaching(19% of the total Chl amount at 40 min of illumination). Neverthe-less, the spectral features of the Lhca1/4 gradient fraction (GB1) arebetter defined after light- than after heat-treatment. This is in linewith the ‘‘safety valve’’ hypothesis stating Lhca1/4 detachmentupon illumination while preserving the structure of this antennacomplex [18], whereas heating appears to induce their irreversibledegradation and no heat-protective mechanism is present in thesupercomplex.
4. Discussion
The photosynthetic apparatus of higher plants is capable torespond and adapt to changes in the environmental conditionsby changes in its macroorganization. These involve mainly struc-tural reorganizations in PSII components – formation of orderedarrays of PSII–LHCII supercomplexes [42], monomerization of the
0.5
0.6
0.7
0.8
0.9
1.0
(-)650 nm (-)491/(+)510 nm (-)684 nm
CD
(rel
. uni
ts)
Time0 10 20 3450 500 550 600 650 700
-8
-6
-4
-2
0
2
4dark 20 min light 40 min light 60 min light
A
CD
, Ax1
04
Wavelength (nm)
Fig. 6. Light-induced destabilization of the pigmetn protein complexes in stroma thylak60 min illuminated samples at 25 �C. (B) Light-induced destabilization of selected CDfluorescence emission spectra of dark adapted and illuminated PSI vesicles, as indicated620 nm was used. The Chl content of the samples was 20 lg/ml in the CD and fluoresce
PSII dimers [43], changes in the oligomerization state of LHCIIrelated to photoprotection [12] as well as changes in the chiral,LHCII-containing macrodomains [24,44], heat [45] and osmotic[46] stresses. Major changes in the chloroplast ultrastructure uponstate transitions have also been reported recently [10]. On molec-ular level they are related to migration of LHCII in the membrane –it can transiently attach to PSI supercomplex and increase itsabsorption cross-section [9,10].
Although the structure of LHCI shares high similarity with thatof LHCII, there are no data suggesting that LHCI is also able tomonomerize under stress conditions or to shuffle between the
B
(min)
0
20
40
60
80
100dark 20 min light 40 min light 60 min light
Wavelength (nm)
Fluo
resc
ence
inte
nsity
(rel
. uni
ts)
C731
728
726
721682
0 40 50 60 660 680 700 720 740 760 780 800
oid membrane vesicles. (A) CD spectra of PSI vesicles, dark-adapted and 20, 40 andbands, relative to the corresponding amplitudes in the dark at 25 �C. (C) 77 K
, with emission maxima labeled. For the (�)650 nm CD band the reference value atnce measurements.

10 S.B. Krumova et al. / Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12
two photosystems. In general PSI is regarded as a conservativesupramolecular complex. With very few exceptions in green plantsit is organized in monomers [3] containing the core and twodimeric antenna complexes, Lhca1/4 and Lhca2/3 [3,5], and nohigher order structures of PSI supercomplexes are found so far.The lack of only one LHCI subunit destabilizes the whole structure[15,16]. Thus, it was suggested that there is a strong cooperativitybetween the PSI supercomplex subunits in terms of their reactiontoward stress factors and degradation. The DSC data presented inthis paper (Fig. 1) however clearly show the presence of severalthermally-induced events in PSI supercomplex demonstrating thatits disassembly proceeds in several steps with distinct thermody-namic properties. Complementary biochemical and spectroscopicdata helped us to distinguish and characterize the core andantenna denaturation.
4.1. LHCI detachment and degradation
The first thermally-induced structural change in PSI supercom-plex at 57 �C resolved by DSC nicely correlates with a process, withsimilar transition temperature, revealed by CD under comparableconditions, i.e. gradual heating (Fig. 2B, (�)650 nm CD band,Tm � 55–60 �C). These data, together with sucrose gradient centri-fugation of heat-treated samples (Fig. 3) demonstrate that thistransition is correlated with LHCI degradation. This is concludedfrom the following facts: (i) this transition involves Chl b molecules((�)650 nm CD band), which are found only in the PSI antenna andnot in the core complex [3,5] and (ii) the density of the LHCI con-taining sucrose gradient fraction increases after incubation at60 �C, however, its CD features are largely diminished (Fig. 3C).Since under these experimental conditions LHCI degraded, wecould detect neither energetic disconnection nor physical dissocia-tion of structurally intact LHCI from the PSI core reported to occurat this temperature [19,47]. However, when we used a somewhatmilder heat treatment (3 min incubation of PSI vesicles at asequence of increasing temperatures in the range 25–90 �C, fol-lowed by sucrose gradient centrifugation) the Lhca1/4 detachmentand degradation could be distinguished. On the contrary, evenunder these conditions we did not resolve heat-induced Lhca2/3detachment from the core complex. This conclusion is largelybased on CD data on the different fraction of sucrose density gradi-ent centrifugation of heat-treated samples. Similar conclusionswere reached by an immunoblot analysis of the protein composi-tion of fractions obtained from heat-treated PSI–LHCI supercom-plexes (Nelleapalli et al., accompanying paper).
Our results also demonstrate Lhca1/4 detachment upon illumi-nation with white light that proceeded proportionally to theabsorbed quanta (time of illumination, Figs. 6 and 7). Although thisprocess is accompanied by gradual photobleaching of Chls, thecharacteristic Lhca1/4 and PSI core CD features are retained up to40 min of illumination (Fig. 7). In contrast to the heat treatment,the illumination conditions used in this study did not induce pro-nounced Lhca1/4 degradation; the Lhca1/4 CD spectrum (GB1,Fig. 7C) of the dark and those illuminated for 20 min and 40 minsamples showed similar features; the same holds true also forthe PSI core complex (Fig. 7C). Thus, our data are in line with thosereported by Alboresi et al. [18] showing that LHCI detachmentupon illumination serves as a ‘‘safety valve’’ and preserves the corecomplex from overexcitation and oxidation damage. Our resultsfurther add that, at least at 25 �C, only Lhca1/4 detaches from thecore, whereas Lhca2/3 retains its localization (Fig. 7C). LHCI func-tional decoupling from the PSI core is also observed in Chlamydo-monas upon iron deficiency and might represent a majormechanism for light stress adaptation of the photosyntheticorganisms [48].
It appears that Lhca1/4 is loosely bound to the core since part ofit detaches even after the detergent solubilization performedbefore loading the PSI sample on the sucrose gradient (Fig. 3).Apparently the recently identified protein–protein contactsbetween the core and antenna subunits and the suggested role oflipid molecules and gap chlorophylls [5] differ in strength forLhca1/4 and Lhca2/3. The light-induced splitting off of Lhca1/4 isassociated with significant photobleaching, which however didnot affect either the PSI core structure or that of Lhca1/4. In allour experiments (heat and light treatment) Lhca2/3 remainedbound to the core until its denaturation. This heterodimer interactswith PsaA, PsaK and is probably stabilized by the lumen-exposedPsaN subunit [5]. These interactions are probably keeping Lhca2/3 closely associated to the core.
4.2. PSI core complex degradation
The combination of DSC and CD spectroscopy showed that grad-ually heated antenna complexes are degrading at 60 �C whichmeans that the calorimetric transitions at 67, 73 and 91 �C mustbe due to components of the PSI core complex. Alboresi et al.[18] revealed that upon photoinhibition at 4 �C PsaA and PsaB sub-units are less stable than PsaD and PsaF. To our knowledge thereare no detailed electrophoretic and immunoblotting studies ofthe thermal stability of PSI components, but Lipova et al. [19]showed that PsaA and PsaB are the most thermally stable pig-ment–protein complexes in barley thylakoids, detectable even at80 �C when all other complexes are degraded. Since PsaA and PsaBbind the majority of the Chls and carotenoids in PSI [3,5], they mostprobably contribute significantly to the characteristic CD bandsoriginating from carotenoids ((�)490/(+)506 nm, Tm � 62–66 �C)and Chl a ((�)681 nm, Tm � 75–81 �C) and correspondingly to thethermal events recorded by DSC at 67 and 73 �C in our PSIpreparations. The non-pigmented PsaD and PsaF subunits probablycontribute to the highest thermal transition at 91 �C.
The PSI core is also stable upon photoinhibition, prolonged illu-mination at 25 �C resulted in gradual photobleaching and splittingof a substantial part of Lhca1/4 off the supercomplex. It appearsthat as long as the Lhca1/4 complexes are able to direct the excesslight energy to red Chls, the core is preserved from overexcitation.
4.3. Thermo-optic effect
Heat and light-induced structural changes were identified by uspreviously in isolated LHCII, isolated grana and intact thylakoidmembranes. Elevated temperatures and excess light were shownto exert very similar effects in the macroorganization of thepigment–protein complexes in the thylakoid membrane and theoligomerization state (trimer vs monomer) of LHCII. The light-induced structural changes were thus proposed to be driven bythermo-optic mechanism – elementary structural changes inducedby ultrafast thermal transients in the close vicinity of the site ofdissipation. These elementary structural changes are accumulatingduring illumination of samples with excess light and are manifest-ing themselves in macroscopically detectable structural changes[12,24,44,49]. In isolated LHCII, as well as in thylakoid membranesin vitro and in vivo, thermo-optic mechanism has been proposed tobe responsible for the regulation of LHCII phosphorylation at thesubstrate level [50,51]. Gruszecki et al. [52], by means of resonanceRaman scattering technique, observed that upon strong illumina-tion, violaxanthin changed its molecular configuration from themore twisted to the more relaxed one, an effect which might alsobe related to the thermo-optic reorganizations in LHCII [53]. Ener-getic decoupling of the phycobilisome antenna, induced by highlight, could also be mimicked by heat-treatments and wereattributed to thermo-optic effect [54,55].

S.B. Krumova et al. / Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12 11
The results presented in this paper demonstrate for the firsttime that heat, above 50 �C, and prolonged white light treatmentat 25 �C induce very similar structural changes in PSI, and thusthe data are fully consistent with the notion that they are inducedby thermo-optic effect.
5. Conclusions
Our data demonstrate the ability of Lhca1/4 to detach from thePSI supercomplex upon heat- and light treatments. This event isfollowed by the degradation or detachment and immediate degra-dation of Lhca2/3 dimer. The extent of splitting is determined bythe heating rate and the applied elevated temperature, or by theamount absorbed quanta at 25 �C – strongly suggesting the roleof thermo-optic effect in PSI cf. [44]. Our data support the hypoth-esis of Alboresi et al. [18] that LHCI plays a role in the photoprotec-tion of PSI. It is concluded that, similarly to PSII, PSI antenna alsopossesses a structural flexibility, which probably plays a significantrole in photoprotection of the photosynthetic apparatus.
Abbreviations
CD
circular dichroism Chl chlorophyll DSC differential scanning calorimetry LHCI light harvesting antenna of photosystem I LHCII major light harvesting complex of photosystem II PSI photosystem I PSII photosystem II Tm transition temperatureAcknowledgements
This work was supported by bilateral projects in the frame ofBulgarian and Hungarian Academy of Sciences, by HungarianResearch Fund OTKA-CNK-80345, TÁMOP-4.2.2.A-11/1/KONV-2012-0060 and by A*STAR, TET_10_SG_STAR_RBIO-FOTOSZIN jointresearch Project to G.G.
References
[1] C. Bengis, N. Nelson, Purification and properties of the photosystem I reactioncenter from chloroplasts, J. Biol. Chem. 250 (1975) 2783–2788.
[2] P. Manna, P.R. Chitnis, Function and molecular genetics of photosystem I, in:G.S. Singhal, G. Renger, S.K. Sapory, K.-D. Irrgang, Govindjee (Eds.), Conceptsin Photobiology: Photosynthesis and Photomorphogenesis, Kluwer, Dordrecht,1999, pp. 212–251.
[3] A. Ben-Shem, F. Frolow, N. Nelson, Crystal structure of plant photosystem I,Nature 426 (2003) 630–635.
[4] A. Amunts, O. Drory, N. Nelson, The structure of a plant photosystem Isupercomplex at 3.4 Å resolution, Nature 447 (2007) 58–63.
[5] A. Amunts, H. Toporik, A. Borovikova, N. Nelson, Structure determination andimproved model of plant photosystem I, J. Biol. Chem. 285 (2010) 3478–3486.
[6] A. Busch, M. Hippler, The structure and function of eukaryotic photosystem I,Biochim. Biophys. Acta 2011 (1807) 864–877.
[7] R. Croce, T. Morosinotto, S. Castelletti, J. Breton, R. Bassi, The Lhca antennacomplexes of higher plants photosystem I, Biochim. Biophys. Acta 1556 (2002)29–40.
[8] S. Jansson, A guide to the Lhc genes and their relatives in Arabidopsis, TrendsPlant Sci. 4 (1999) 236–240.
[9] J.F. Allen, J. Forsberg, Molecular recognition in thylakoid structure andfunction, Trends Plant Sci. 6 (2001) 317–326.
[10] M. Tikkanen, M. Nurmi, M. Suorsa, R. Danielsson, F. Mamedov, S. Styring, E.-M.Aro, Phosphorylation-dependent regulation of excitation energy distributionbetween the two photosystems in higher plants, Biochim. Biophys. Acta 1777(2008) 425–432.
[11] J.P. Dekker, E.J. Boekema, Supramolecular organization of thylakoid membraneproteins in green plants, Biochim. Biophys. Acta 1706 (2005) 12–39.
[12] G. Garab, Z. Cseh, L. Kovács, S. Rajagopal, Z. Várkonyi, M. Wentworth, L.Mustárdy, A. Der, A.V. Ruban, E. Papp, A. Holzenburg, P. Horton, Light-induced trimer to monomer transition in the main light-harvesting antenna
complex of plants: thermo-optic mechanism, Biochemistry 41 (2002)15121–15129.
[13] G. Garab, H. van Amerongen, Linear dichroism and circular dichroism inphotosynthesis research, Photosynth. Res. 101 (2009) 135–146.
[14] A. Amunts, N. Nelson, Functional organization of a plant photosystem I:evolution of a highly efficient photochemical machine, Plant Physiol. Biochem.46 (2008) 228–237.
[15] E. Wientjes, G.T. Oostergetel, S. Jansson, E.J. Boekema, R. Croce, The role of Lhcacomplexes in the supramolecular organization of higher plant photosystem I, J.Biol. Chem. 284 (2009) 7803–7810.
[16] T. Morosinotto, M. Ballottari, F. Klimmek, S. Jansson, R. Bassi, The association ofthe antenna system to photosystem I in higher plants, J. Biol. Chem. 280 (2005)31050–31058.
[17] S. Bailey, R.G. Walters, S. Jansson, P. Horton, Acclimation of Arabidopsisthaliana to the light environment: the existence of separate low light and highlight responses, Planta 213 (2001) 794–801.
[18] A. Alboresi, M. Ballottari, R. Hienerwadel, G.M. Giacometti, T. Morosinotto,Antenna complexes protect photosystem I from photoinhibition, BMC PlantBiol. 9 (2009) 71.
[19] L. Lípová, P. Krchnák, J. Komenda, P. Ilík, Heat-induced disassembly anddegradation of chlorophyll-containing protein complexes in vivo, Biochim.Biophys. Acta 1797 (2010) 63–70.
[20] J. Nauš, L. Dvorak, R. Kuropatwa, M. Maslan, Transitions in the thylakoidmembranes of barley leaves studied by chlorophyll fluorescence temperaturecurve, Photosynthetica 27 (1992) 563–570.
[21] Y. Yamane, Y. Kashino, H. Koike, K. Satoh, Increases in the fluorescence Fo
level and reversible inhibition of photosystem II reaction center by high-temperature treatments in higher plants, Photosynth. Res. 52 (1997) 57–64.
[22] F.A.J. Peters, J.E. van Wielink, W.F. Sang, S. Pevries, R. Krayenhof, Studies onwell coupled photosystem I – enriched subchloroplast vesicles – content andredox properties of electron-transfer components, Biochim. Biophys. Acta 722(1983) 460–470.
[23] R.J. Porra, W.A. Thompson, P.E. Kriedeman, Determination of accurateextinction coefficients and simultaneous-equations for assaying chlorophyll-a and chlorophyll-b extracted with 4 different solvents – verification of theconcentration of chlorophyll standards by atomic absorption spectroscopy,Biochim. Biophys. Acta 975 (1989) 384–394.
[24] A.G. Dobrikova, Z. Várkonyi, S.B. Krumova, L. Kovács, G.K. Kostov, S.J. Todinova,M.C. Busheva, S.G. Taneva, G. Garab, Structural rearrangements in chloroplastthylakoid membranes revealed by differential scanning calorimetry andcircular dichroism spectroscopy. Thermo-optic effect, Biochemistry 42(2003) 11272–11280.
[25] S.B. Krumova, S.J. Todinova, A.G. Dobrikova, S.G. Taneva, Differential scanningcalorimetry of photosynthetic membranes: resolving contributions of themajor photosynthetic complexes to the sequential thermal transitions, TrendsPhotochem. Photobiol. 12 (2010) 37–51.
[26] W.P. Williams, P.J. Quinn, The phase behavior of lipids in photosyntheticmembranes, J. Bioenerg. Biomembr. 19 (1987) 605–624.
[27] K. Gounaris, A.P.R. Brain, P.J. Quinn, W.P. Williams, Structural and functionalchanges associated with heat-induced phase-separations of non-bilayer lipidsin chloroplast thylakoid membranes, FEBS Lett. 153 (1983) 47–52.
[28] Z. Kóta, B. Szalontai, M. Droppa, G. Horváth, T. Páli, Fourier transform infraredand electron paramagnetic resonance spectroscopic studies of thylakoidmembranes, J. Mol. Str. 480–481 (1999) 395–400.
[29] S.B. Krumova, C. Dijkema, P. de Waard, H.V. As, G. Garab, H. van Amerongen,Phase behavior of phosphatidylglycerol in spinach thylakoid membranes asrevealed by 31P-NMR, Biochim. Biophys. Acta 1778 (2008) 997–1003.
[30] W.G. Nolan, H.P. Hopkins Jr., S.A. Kalini, Differential scanning calorimetricinvestigation of pea chloroplast thylakoids and thylakoid fractions, Arch.Biochem. Biophys. 297 (1992) 19–27.
[31] K.A. Smith, B.K. Ardelt, P.S. Low, Identification of the soluble coupling factortransition in calorimetric scans of chloroplast membranes, Biochemistry 25(1986) 7099–7105.
[32] C. Büchel, G. Garab, Organization of the pigment molecules in the chlorophylla/c light-harvesting complex of Pleurochloris meiringensis (Xanthophyceae).Characterization with circular dichroism and absorbance spectroscopy, JPhotochem. Photobiol. B 37 (1997) 118–124.
[33] R.M. Pearlstein, Theoretical interpretation of antenna spectra, in: H. Scheer(Ed.), Chlorophylls, CRC Press, Boca Raton, 1991, pp. 1047–1078.
[34] Z. Yang, X. Su, F. Wu, Y. Gong, T. Kuang, Effect of phosphatidylglycerol onmolecular organization of photosystem I, Biophys. Chem. 115 (2005) 19–27.
[35] I. Ikegami, S. Itoh, Chlorophyll organization in P-700-enriched particlesisolated from spinach chloroplasts. CD and absorption spectroscopy,Biochim. Biophys. Acta 851 (1986) 75–85.
[36] E. Wientjes, R. Croce, The light-harvesting complexes of higher-plantPhotosystem I: Lhca1/4 and Lhca2/3 form two red-emitting heterodimers,Biochem. J. 433 (2011) 477–485.
[37] V.V. Shubin, V.L. Tsuprun, I.N. Bezsmertnaya, N.V. Karapetyan, Trimeric formsof the photosystem I reaction center complex pre-exist in the membranes ofthe cyanobacterium Spirulina platensis, FEBS Lett. 334 (1993) 79–82.
[38] J.E. Mullet, J.J. Burke, C.J. Arntzen, Chlorophyll proteins of photosystem I, PlantPhysiol. 65 (1980) 814–822.
[39] B.W. Dreyfuss, P. Thornber, Organization of the light-harvesting complex ofphotosystem I and its assembly during plastid development, Plant Physiol. 106(1994) 841–848.

12 S.B. Krumova et al. / Journal of Photochemistry and Photobiology B: Biology 137 (2014) 4–12
[40] V.H. Schmid, S. Potthast, M. Wiener, V. Bergauer, H. Paulsen, S. Storf, Pigmentbinding of photosystem I light-harvesting proteins, J. Biol. Chem. 277 (2002)37307–37314.
[41] Y. Hui, W. Jie, R. Carpentier, Degradation of the photosystem I complex duringphotoinhibition, Photochem. Photobiol. 72 (2000) 508–512.
[42] E.J. Boekema, J.F.L. van Breemen, H. van Roon, J.P. Dekker, Arrangement ofphotosystem II supercomplexes in crystalline macrodomains within thethylakoid membrane of green plant chloroplasts, J. Mol. Biol. 301 (2000)1123–1133.
[43] E.-M. Aro, M. Suorsa, A. Rokka, Y. Allahverdiyeva, V. Paakkarinen, A. Saleem, N.Battchikova, E. Rintamaeki, Dynamics of photosystem II: a proteomic approachto thylakoid protein complexes, J. Exp. Botany 56 (2005) 347–356.
[44] Z. Cseh, A. Vianelli, S. Rajagopal, S. Krumova, L. Kovács, E. Papp, V. Barzda, R.Jennings, G. Garab, Thermo-optically induced reorganizations in the main lightharvesting antenna of plants. I. Non-Arrhenius type of temperaturedependence and linear light-intensity dependencies, Photosynth. Res. 86(2005) 263–273.
[45] Y. Tang, X. Wen, Q. Lu, Z. Yang, Z. Cheng, C. Lu, Heat stress induces anaggregation of the light-harvesting complex of photosystem II in spinachplants, Plant Physiol. 143 (2007) 629–638.
[46] L. Gáspár, E. Sárvári, I. Molnár, L. Stéhli, M. Molnár-Láng, G. Galiba, Structuralchanges of the photosynthetic apparatus under osmotic stress in differentTriticum aestivum and Aegilops biuncialis genotypes, Acta Biol. Szeged. 46(2002) 91–93.
[47] Z.-H. Hu, Y.-N. Xu, G.-Z. Jiang, T.-Y. Kuang, Degradation and inactivation ofphotosystem I complexes during linear heating, Plant Sci. 166 (2004) 1177–1183.
[48] J.L. Moseley, T. Allinger, S. Herzog, P. Hoerth, E. Wehinger, S. Merchant, M.Hippler, Adaptation to Fe-deficiency requires remodeling of thephotosynthetic apparatus, EMBO J. 21 (2002) 6709–6720.
[49] Z. Cseh, S. Rajagopal, T. Tsonev, M. Busheva, E. Papp, G. Garab, Thermoopticeffect in chloroplast thylakoid membranes. Thermal and light stability ofpigment arrays with different levels of structural complexity, Biochemistry 39(2001) 15250–15257.
[50] H. Zer, M. Vink, N. Keren, H.G. Dilly-Hartwig, H. Paulsen, R.G. Herrmann, B.Andersson, I. Ohad, Regulation of thylakoid protein phosphorylation at thesubstrate level: reversible light-induced conformational changes expose thephosphorylation site of the light-harvesting complex II, Proc. Natl. Acad. Sci.USA 96 (1999) 8277–8282.
[51] H. Zer, M. Vink, S. Shochat, R.G. Herrmann, B. Andersson, I. Ohad, Light affectsthe accessibility of the thylakoid light harvesting complex II, (LHCII)phosphorylation site to the membrane protein kinase(s), Biochemistry 42(2003) 728–738.
[52] W.I. Gruszecki, M. Gospodarek, W. Grudzinski, R. Mazur, K. Gieczewska, M.Garstka, Light-induced change of configuration of the LHCII-boundxanthophyll (tentatively assigned to violaxanthin): a resonance Ramanstudy, J. Phys. Chem. 113 (2009) 2506–2512.
[53] M. Zubik, R. Luchowski, M. Puzio, E. Janik, J. Bednarska, W. Grudzinski, W.I.Gruszecki, The negative feedback molecular mechanism which regulatesexcitation level in the plant photosynthetic complex LHCII: towardsidentification of the energy dissipative state, Biochim. Biophys. Acta 2013(1827) 355–364.
[54] K. Stoitchkova, O. Zsiros, T. Jávorfi, T. Páli, A. Andreeva, Z. Gombos, G. Garab,Heat- and light-induced reorganizations in the phycobilisome antenna ofSynechocystis sp. PCC 6803. Thermo-optic effect, Biochim. Biophys. Acta 1767(2007) 750–756.
[55] E. Tamary, V. Kiss, R. Nevo, Z. Adam, G. Bernát, S. Rexroth, M. Rögner, Z. Reich,Structural and functional alterations of cyanobacterial phycobilisomesinduced by high-light stress, Biochim. Biophys. Acta 2012 (1817) 319–327.



















