
Unfolding the political capacities of design. With Uriel Fogue

1 23
Biotechnology Letters ISSN 0141-5492 Biotechnol LettDOI 10.1007/s10529-011-0710-9
Hairy roots cultures from differentSolanaceous species have varyingcapacities to produce E. coli B-subunitheat-labile toxin antigen
Giorgio De Guzman, AmandaM. Walmsley, Diane E. Webster & JohnD. Hamill

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media B.V.. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

ORIGINAL RESEARCH PAPER
Hairy roots cultures from different Solanaceous specieshave varying capacities to produce E. coli B-subunitheat-labile toxin antigen
Giorgio De Guzman • Amanda M. Walmsley •
Diane E. Webster • John D. Hamill
Received: 20 June 2011 / Accepted: 15 July 2011
� Springer Science+Business Media B.V. 2011
Abstract The gene encoding enterotoxigenic
Escherichia coli B-subunit heat-labile toxin (LTB)
antigen was co-transformed into hairy root cultures of
Nicotiana tabacum (tobacco), Solanum lycopersicum
(tomato) and Petunia parodii (petunia) under the
CaMV35S promoter. Tobacco and petunia roots
contained *65–70 lg LTB g-1 tissue whilst hairy
roots of tomato contained *10 lg LTB g-1. Antigen
at *600 ng ml-1 was detected in growth medium of
tobacco and petunia. Tobacco roots with higher LTB
levels showed growth retardation of *80% whereas
petunia hairy roots with similar levels of LTB showed
only *35% growth retardation, relative to vector
controls. Regeneration of plants from LTB-containing
tobacco hairy roots was readily achieved and re-
initiated hairy roots from greenhouse-grown plants
showed similar growth and LTB production charac-
teristics as the original hairy root cultures.
Keywords Hairy root � E. coli toxin � Molecular
pharming � Plant-made vaccine � Rhizosecretion
Introduction
Improved viral vector systems have increased the
efficacy of transient systems to allow large amounts
of immunogenic proteins to be produced in plants
(Gleba et al. 2005; Hefferon 2010). Despite the many
advantages of these approaches, they still have some
limitations with on-going requirements for re-infec-
tion/infiltration of plants and extensive batch-to-batch
testing for accurate dosage purposes. The use of
stable transgenic plants as a potential alternative for
vaccine production is an attractive proposition but the
technology has been slow to become established, due
in part to relatively low levels of vaccine production
in vivo and also the need for high levels of bio-
containment to avoid inadvertent dispersal of trans-
genic plants. Rhizosecretion of foreign proteins from
roots has also been noted as being a potentially
valuable feature of transgenic plants and has been
shown to be possible for both small (e.g. 4E10) and
large (e.g. Guy’s 13) functional proteins at relatively
high levels, particularly when roots of transgenic
plants are treated with the phytohormone auxin
(Drake et al. 2009).
G. De Guzman � A. M. Walmsley � D. E. Webster �J. D. Hamill (&)
School of Biological Sciences, Monash University,
Melbourne, VIC, Australia
e-mail: [email protected]
G. De Guzman
e-mail: [email protected]
A. M. Walmsley
e-mail: [email protected]
D. E. Webster
e-mail: [email protected]
D. E. Webster
Faculty of Medicine, Nursing & Health Sciences, Monash
University, Melbourne, VIC, Australia
123
Biotechnol Lett
DOI 10.1007/s10529-011-0710-9
Author's personal copy

Axenic hairy root cultures represent a genetically
stable transgenic tissue culture system which grows
rapidly in vitro in simple and inexpensive media,
without a requirement for exogenous phytohormones
(Hamill and Lidgett 1997; Guillon et al. 2006). Of
particular relevance for foreign protein production,
hairy roots are non-photosynthetic and thus easily
bio-contained and can be scaled up to produce large
amounts of biomass in industrial scale bio-reactors
(Wilson 1997; Boehm 2007; Shih and Doran 2009).
Recombinant proteins, such as GFP and SEAP, are
secreted into the surrounding media raising the
prospect of low-cost downstream purification proto-
cols for valuable therapeutic proteins produced by
hairy roots (Gaume et al. 2003).
LTB production by transgenic plants has been
reported (Mason et al. 1998; Walmsley et al. 2003).
The holotoxin consists of two subunits, LTA
(28 kDa) and LTB (11.6 kDa) (Sixma et al. 1993).
LTB forms a *60 kDa pentamer which facilitates
binding to the GM1 ganglioside receptors on host
epithelial cells allowing the LTA to endocytose and
cause toxicity (Mason et al. 1998). In the present
study, we quantify the capacity of hairy roots from
three species in the Solanaceous family to produce
and secrete LTB protein. We also explore the ability
of hairy roots to be regenerated into transgenic plants
from which LTB-producing hairy roots can be re-
initiated, thereby providing a low-technology alter-
native to repeated subculturing or more expensive
techniques such as cryopreservation for long-term
perpetuation of hairy root lines with desirable anti-
genic protein producing properties.
Materials and methods
Introduction of expression constructs
into Agrobacterium rhizogenes
The pBin? 35sLTB construct was made using the
pTH210 vector (Mason et al. 1998) where the
expression cassette was ligated into appropriate sites
of the pBinPlus binary vector described by van
Engelen et al. (1995). The native pBinPlus and the
pBI121 vector (containing the GUS gene) were used
as controls (Jefferson et al. 1987). Each construct was
separately transformed into Agrobacterium rhizoge-
nes strain LBA9402 by high voltage electroporation
(Wen-Jun and Forde 1989). Bacteria were selected on
YMB agar containing appropriate antibiotics and
single colonies screened by PCR to confirm the
presence of the genes of interest (Hamill et al. 1991;
Hamill and Lidgett 1997).
Establishment of hairy root cultures
Nicotiana tabacum var. NC 95 (tobacco; Cane et al.
2005), Solanum lycopersicum var. Roma (tomato;
Yates, Australia) and Petunia parodii (petunia; a gift
from Prof E.C. Cocking and Dr J.B. Power, Notting-
ham University, UK) were grown in a greenhouse
under a 16 h photoperiod as described previously by
Cane et al. (2005). Young leaves were infected with
A. rhizogenes containing pBin ? 35SLTB, pBI121 or
vector control, and clonal hairy root cultures selected
as described previously by Hamill and Lidgett
(1997). After *2 weeks, DNA from healthy prolif-
erating roots was extracted and PCR-screened for
the presence of LTB sequence using primers
FWD 50-GCCATGGTGAAGGTGAAGTGCTA-30 REV
50-CCATGGTGAAGGTGAAGTGCTA-30. RolB and
VirD primers were also used to show that the hairy
roots contained A.rhizogenes T-DNA without residual
Agrobacterium in the cultures (Hamill et al. 1991).
Similar PCR-screening was performed on regener-
ated plant tissues to confirm their transgenic status.
Growth
Culture flasks were seeded with 0.1 g healthy roots
using C5 selected independent clonal hairy root lines
of each species. Triplicate root cultures for each
independent line were harvested. Blotted dry weights
(BW) were recorded after repeated blotting to remove
extracellular moisture (Chintapakorn and Hamill
2003) as preliminary experiments (data not shown)
showed that levels of pentameric LTB protein
extracted from dried root material were greatly
reduced when compared to that of blotted plant
material. Separate analysis showed that the ratio
between BW and dry weight was 10:1 (P \ 0.05) for
roots of all species.
Extraction of total soluble protein
Total soluble protein (TSP) extractions of root
(100 lg BW) and leaf (500 lg) material were
Biotechnol Lett
123
Author's personal copy

performed as described by Wang et al. (2003) with
addition of 29 Roche Complete Protease Inhibitor
Cocktail solution to extraction buffer and homogeni-
sation with a 3 mm tungsten bead for 1 min at
28 Hz/s in a Qiagen Mixer Mill. Concentrations of
TSP were determined using a Bradford based com-
mercial kit method (Bio-Rad Protein Assay).
SDS PAGE—Western blot
Equal amounts of TSP (100 lg) were used for SDS-
PAGE (Walmsley et al. 2003). The primary antibody
used was a rabbit anti-LTB monoclonal antibody
(1/5,000 in 0.5% PBSTM) (Benchmark Biolabs) and
the secondary antibody was an anti-rabbit fluorescent
antibody ALEXA 700 (Rockland inc.) (1/5,000 in
0.5% PBSTM). Blot was detected on a Licor Odyssey
imaging system.
Quantification of LTB antigen
Equal amounts of TSP (25 lg) from each test sample
were analysed by ELISA using the GM1 ELISA
protocol (Walmsley et al. 2003). Standard curves
were produced using recombinant bacterial (rLTB)
protein (Benchmark Biolabs) in twofold serial dilu-
tions starting at 5 ng. Plates were incubated overnight
at 4�C. The primary antibody used was anti-LTB-
rabbit (Benchmark Biolabs) at 1/2,000 dissolved in
0.5% skim milk PBST and the secondary antibody
(goat anti rabbit—HRP) (Sigma Aldrich) was used at
a dilution of 1/15,000 in 0.5% skim milk PBST.
Tetra-methyl-benzedine (TMB) (Bio-Rad Laborato-
ries) was used for detection at 620 nm on a ‘Labsys-
tems Multiscan’ plate reader. Data obtained was
standardised to the rLTB standard curve.
Re-initiation of hairy roots from regenerated
transgenic plants
Regeneration of plants from selected hairy root
cultures of tobacco and transfer of plants to soil for
growth under greenhouse conditions was undertaken
as reported previously (Chintapakorn and Hamill
2003). Re-initiation of hairy roots was achieved by
culture of *2 cm pieces of surface sterilised upper
leaves from non-flowering plants grown on standard
MS agar medium containing 30 g l-1 sucrose. Roots
typically formed within 1–2 weeks of culture and
were easily transferred to liquid culture and grown
and per original hairy roots as noted above.
Results
Following selection in media containing kanamycin,
hairy roots of all species produced by A. rhizogenes
containing pBin? 35sLTB contained LTB using
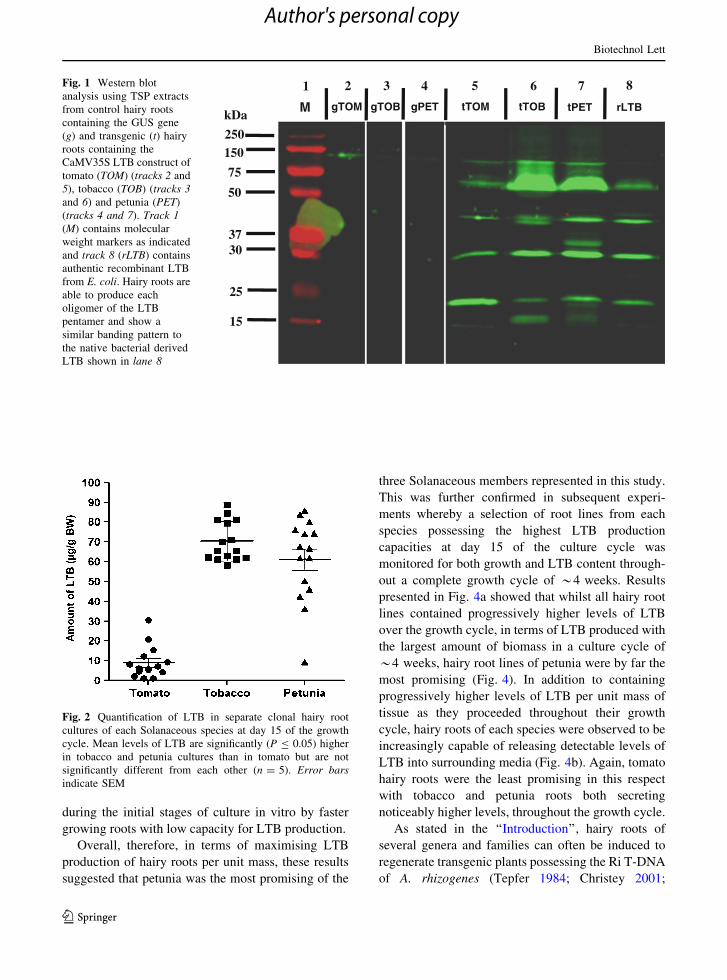
western analysis (Fig. 1). Oligomer sizes detected in
these extracts reflect that of the known LTB oligomer
sizes of the crystallised pentamer (Sixma et al. 1993).
Fifteen independently derived hairy root cultures were
assessed for each species at the mid-point in a typical
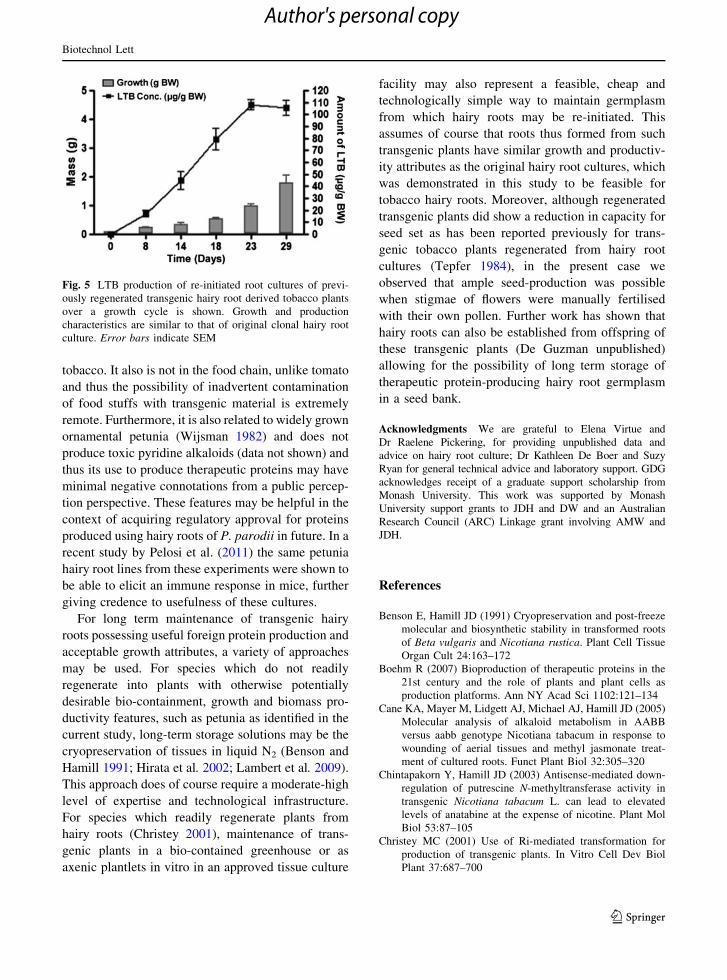
culture cycle. Mean LTB levels for each species are
shown in Fig. 2. To determine the extent to which
growth of individual root lines in each species varied
in relation to capacity for LTB production, C5
cultures representing high (above mean) and also
median levels of LTB within that species, were
monitored at regular time periods over a typical hairy
root culture growth cycle of *4 weeks (Fig. 3a, b, c).
Lines containing GUS protein served as controls, as
the presence of GUS did not alter growth relative to
lines transformed with A. rhizogenes alone (Fig. 3d).
However, hairy roots containing GUS protein typi-
cally produced more biomass per unit time than
comparable hairy roots of each species containing
LTB. Furthermore, a pattern was observed in each
species whereby roots containing high levels (above
mean) of LTB produced less biomass over time than
those which contained median levels of LTB. This
observation was most noticeable for hairy roots of
tobacco, showing *80% growth retardation relative
to the controls (Fig. 3a).
In the case of petunia, deleterious effects of LTB
production upon biomass produced per unit time were
noticeably less than in tobacco, with only *35%
growth retardation relative to their vector controls
(Fig. 3b). In the case of tomato hairy root cultures,
lines containing LTB grew only slightly slower than
their equivalent GUS controls (Fig. 3c). However, it is
important to note that the maximum level of LTB
observed in tomato hairy roots was 5–6 times lower
than the mean level observed in either petunia or
tobacco hairy roots. It is possible that any tomato hairy
root lines possessing high levels of LTB were either not
viable or grew so slowly that they were out-competed
Biotechnol Lett
123
Author's personal copy
during the initial stages of culture in vitro by faster
growing roots with low capacity for LTB production.
Overall, therefore, in terms of maximising LTB
production of hairy roots per unit mass, these results
suggested that petunia was the most promising of the
three Solanaceous members represented in this study.
This was further confirmed in subsequent experi-
ments whereby a selection of root lines from each
species possessing the highest LTB production
capacities at day 15 of the culture cycle was
monitored for both growth and LTB content through-
out a complete growth cycle of *4 weeks. Results
presented in Fig. 4a showed that whilst all hairy root
lines contained progressively higher levels of LTB
over the growth cycle, in terms of LTB produced with
the largest amount of biomass in a culture cycle of
*4 weeks, hairy root lines of petunia were by far the
most promising (Fig. 4). In addition to containing
progressively higher levels of LTB per unit mass of
tissue as they proceeded throughout their growth
cycle, hairy roots of each species were observed to be
increasingly capable of releasing detectable levels of
LTB into surrounding media (Fig. 4b). Again, tomato
hairy roots were the least promising in this respect
with tobacco and petunia roots both secreting
noticeably higher levels, throughout the growth cycle.
As stated in the ‘‘Introduction’’, hairy roots of
several genera and families can often be induced to
regenerate transgenic plants possessing the Ri T-DNA
of A. rhizogenes (Tepfer 1984; Christey 2001;
250150
75
50
kDaM gTOM tTOM rLTBtTOB tPETgTOB gPET
1 2 3 4 5 6 7 8
3730
25
15
Fig. 1 Western blot
analysis using TSP extracts
from control hairy roots
containing the GUS gene
(g) and transgenic (t) hairy
roots containing the
CaMV35S LTB construct of
tomato (TOM) (tracks 2 and
5), tobacco (TOB) (tracks 3and 6) and petunia (PET)
(tracks 4 and 7). Track 1(M) contains molecular
weight markers as indicated
and track 8 (rLTB) contains
authentic recombinant LTB
from E. coli. Hairy roots are
able to produce each
oligomer of the LTB
pentamer and show a
similar banding pattern to
the native bacterial derived
LTB shown in lane 8
Fig. 2 Quantification of LTB in separate clonal hairy root
cultures of each Solanaceous species at day 15 of the growth
cycle. Mean levels of LTB are significantly (P B 0.05) higher
in tobacco and petunia cultures than in tomato but are not
significantly different from each other (n = 5). Error barsindicate SEM
Biotechnol Lett
123
Author's personal copy

Chintapakorn and Hamill 2003). In the current study,
although repeated attempts employing a large range of
phytohormone combinations were unsuccessful in
regenerating plantlets from all hairy root lines of
petunia and tomato, hairy roots of tobacco expressing
LTB readily regenerated into plantlets in vitro when
cultured on basic medium containing a high cytoki-
nin:auxin ratio. Following transfer to compost, these
hairy root-derived transgenic tobacco plants were
observed to possess the slightly wrinkled leaf pheno-
type typical of plants containing Ri-T-DNA (Tepfer
1984; Chintapakorn and Hamill 2003) but overall
were healthy and contained LTB in both roots and
leaves (data not shown). Plants were maintained in a
healthy state in a bio-containment greenhouse for
more than 2 years. Culture of surface-sterilised leaf
material from these transgenic plants resulted in
prolific adventitious root formation when tissue was
cultured in vitro on standard medium lacking phyto-
hormones. Following removal from parental leaf
tissue, axenic hairy root cultures were easily estab-
lished which possessed growth rates and LTB con-
tents comparable to the parental hairy root culture
from which their source plants were derived (Fig. 5).
Discussion
Results noted above illustrate the potential value of
analysing a range of species to maximise potential for
vaccine production in cultured hairy roots. In the
current study, we focussed on species belonging to
three genera within the family Solanaceae and
observed that whilst all were highly receptive to
transformation with A. rhizogenes, Petunia parodii
(petunia) is the most capable of producing healthy
and rapidly growing hairy roots also expressing
high levels of LTB protein. LTB protein has been
produced at various levels in a range of other trans-
genic plant systems. For example transgenic carrots
contained 3 lg LTB g-1 FW (Rosales-Mendoza
et al. 2008), transgenic potato tubers contained up
to 17 lg g-1 FW (Mason et al. 1998), fruits from
transgenic tomato plants contained 38 lg g-1 DW
(Walmsley et al. 2003) and transiently-transformed
leaf tissues of N. benthamiana contained 75 lg g-1
FW (Wagner et al. 2004) Levels of LTB in hairy
roots produced in the current study compare favour-
ably therefore with those observed in other plant
systems.
0 5 10 15 20 25 300
1
2
3
4
5
Tom GusPet GusTob GusTob NegPet NegTom Neg
Time (Days)
Wei
gh
t (g
)0 5 10 15 20 25 30
0
1
2
3
4
5
Tom High LTB
Tom Medium LTB
Tom Gus (Neg)
Time (Days)
Wei
gh
t (g
)
0 5 10 15 20 25 300
1
2
3
4
5
Pet High LTB
Pet Medium LTB
Pet Gus (Neg)
Time (Days)
Wei
gh
t (g
)
0 5 10 15 20 25 300
1
2
3
4
Tob Medium LTBTob High LTB
Tob Gus (Neg)
Time (Days)
Wei
gh
t (g
)
A B
C D
Fig. 3 Growth of representative hairy root cultures of tobacco
(a), tomato (b) and petunia (c) expressing LTB compared to
GUS producing control hairy root cultures of each species.
LTB production has a negative effect on the growth capacity of
roots of each species, particularly in tobacco. GUS expression
within a species does not significantly have an effect on root
growth compared to roots containing only empty vector
T-DNA (d). Error bars indicate SEM
Biotechnol Lett
123
Author's personal copy

Rapidly growing hairy roots can be readily estab-
lished for species belonging to a wide range of
dicotyledonous species (Hamill and Lidgett 1997)
thus opening up the possibility of yet higher total
yields than have been observed using petunia as
outlined in the current study. From a bio-containment
point of view however, use of this species for
production of foreign proteins by hairy roots cultured
in vitro has the potential advantage that plantlets do
not readily regenerate from hairy roots, unlike
Fig. 4 a LTB produced by hairy roots of tobacco (panel A),
petunia (panel B) and tomato (panel C) harvested at different
stages of the growth cycle is shown for all species. Highest
levels of LTB in roots of each species at the point of harvest
were observed to be *3 weeks after subculture. Petunia
cultures are able to produce greater biomass while expressing
LTB levels at similar levels to tobacco hairy roots (P C 0.05).
Tomato hairy roots show rapid growth capacity but much lower
LTB levels. b LTB detected in media of tobacco, petunia and
tomato hairy root cultures harvested at discrete stages of the
growth cycle is shown for all species. Concentration of LTB in
the media was seen to be highest *3 weeks into the growth
cycle (*600 ng ml-1), with petunia and tobacco cultures
secreting greater amounts than that of tomato. Error barsindicate SEM
Biotechnol Lett
123
Author's personal copy
tobacco. It also is not in the food chain, unlike tomato
and thus the possibility of inadvertent contamination
of food stuffs with transgenic material is extremely
remote. Furthermore, it is also related to widely grown
ornamental petunia (Wijsman 1982) and does not
produce toxic pyridine alkaloids (data not shown) and
thus its use to produce therapeutic proteins may have
minimal negative connotations from a public percep-
tion perspective. These features may be helpful in the
context of acquiring regulatory approval for proteins
produced using hairy roots of P. parodii in future. In a
recent study by Pelosi et al. (2011) the same petunia
hairy root lines from these experiments were shown to
be able to elicit an immune response in mice, further
giving credence to usefulness of these cultures.
For long term maintenance of transgenic hairy
roots possessing useful foreign protein production and
acceptable growth attributes, a variety of approaches
may be used. For species which do not readily
regenerate into plants with otherwise potentially
desirable bio-containment, growth and biomass pro-
ductivity features, such as petunia as identified in the
current study, long-term storage solutions may be the
cryopreservation of tissues in liquid N2 (Benson and
Hamill 1991; Hirata et al. 2002; Lambert et al. 2009).
This approach does of course require a moderate-high
level of expertise and technological infrastructure.
For species which readily regenerate plants from
hairy roots (Christey 2001), maintenance of trans-
genic plants in a bio-contained greenhouse or as
axenic plantlets in vitro in an approved tissue culture
facility may also represent a feasible, cheap and
technologically simple way to maintain germplasm
from which hairy roots may be re-initiated. This
assumes of course that roots thus formed from such
transgenic plants have similar growth and productiv-
ity attributes as the original hairy root cultures, which
was demonstrated in this study to be feasible for
tobacco hairy roots. Moreover, although regenerated
transgenic plants did show a reduction in capacity for
seed set as has been reported previously for trans-
genic tobacco plants regenerated from hairy root
cultures (Tepfer 1984), in the present case we
observed that ample seed-production was possible
when stigmae of flowers were manually fertilised
with their own pollen. Further work has shown that
hairy roots can also be established from offspring of
these transgenic plants (De Guzman unpublished)
allowing for the possibility of long term storage of
therapeutic protein-producing hairy root germplasm
in a seed bank.
Acknowledgments We are grateful to Elena Virtue and
Dr Raelene Pickering, for providing unpublished data and
advice on hairy root culture; Dr Kathleen De Boer and Suzy
Ryan for general technical advice and laboratory support. GDG
acknowledges receipt of a graduate support scholarship from
Monash University. This work was supported by Monash
University support grants to JDH and DW and an Australian
Research Council (ARC) Linkage grant involving AMW and
JDH.
References
Benson E, Hamill JD (1991) Cryopreservation and post-freeze
molecular and biosynthetic stability in transformed roots
of Beta vulgaris and Nicotiana rustica. Plant Cell Tissue
Organ Cult 24:163–172
Boehm R (2007) Bioproduction of therapeutic proteins in the
21st century and the role of plants and plant cells as
production platforms. Ann NY Acad Sci 1102:121–134
Cane KA, Mayer M, Lidgett AJ, Michael AJ, Hamill JD (2005)
Molecular analysis of alkaloid metabolism in AABB
versus aabb genotype Nicotiana tabacum in response to
wounding of aerial tissues and methyl jasmonate treat-
ment of cultured roots. Funct Plant Biol 32:305–320
Chintapakorn Y, Hamill JD (2003) Antisense-mediated down-
regulation of putrescine N-methyltransferase activity in
transgenic Nicotiana tabacum L. can lead to elevated
levels of anatabine at the expense of nicotine. Plant Mol
Biol 53:87–105
Christey MC (2001) Use of Ri-mediated transformation for
production of transgenic plants. In Vitro Cell Dev Biol
Plant 37:687–700
Fig. 5 LTB production of re-initiated root cultures of previ-
ously regenerated transgenic hairy root derived tobacco plants
over a growth cycle is shown. Growth and production
characteristics are similar to that of original clonal hairy root
culture. Error bars indicate SEM
Biotechnol Lett
123
Author's personal copy

Drake PMW, Barbi T, Sexton A, McGowan E, Stadlmann J,
Navarre C, Paul MJ, Ma JK (2009) Development of
rhizosecretion as a production system for recombinant
proteins from hydroponic cultivated tobacco. FASEB J
23:3581–3589. doi:10.1096/fj.09-131771
Gaume A, Komarnytsky S, Borisjuk N, Raskin I (2003)
Rhizosecretion of recombinant proteins from plant hairy
roots. Plant Cell Rep 21:1188–1193
Gleba Y, Klimyuk V, Marillonnet S (2005) Magnifection—a
new platform for expressing recombinant vaccines in
plants. Vaccine 23:2042–2048. doi:10.1016/j.vaccine.
2005.01.006
Guillon S, Tremouillaux-Guiller J, Kumar-Pati P, Rideau M,
Gantet P (2006) Harnessing the potential of hairy roots:
dawn of a new era. Trends Biotech 24:403–409
Hamill JD, Lidgett AJ (1997) Hairy root cultures–opportunities
and key protocols for studies in metabolic engineering. In:
Doran PM (ed) Hairy roots: culture and application.
Overseas Publishers Association, Amsterdam, pp 1–29
Hamill JD, Rounsley S, Spencer A, Todd G, Rhodes MJC
(1991) The use of the polymerase chain reaction in plant
transformation studies. Plant Cell Rep 10:221–224. doi:
10.1007/BF00232562
Hefferon KL (2010) Biopharmaceuticals in plants—toward the
next century of medicine. CRC Press Taylor & Francis
Group, Boca Raton
Hirata K, Mukai M, Goda S, Ishio-Kinugasa M, Yoshida K,
Sakai A, Miyamoto K (2002) Cryopreservation of hairy
root cultures of Vinca minor (L.) by encapsulation-dehy-
dration. Biotechnol Lett 24:371–376
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions:
b-glucuronidase as sensitive and versatile gene fusion
marker in higher plants. EMBO J 6:3901–3907
Lambert E, Goosens A, Panis B, Van Labele MC, Geelan D
(2009) Cryopreservation of hairy root cultures of Maesalanceolata and Medicago trunculata. Plant Cell Tissue
Organ Cult 3:289–296. doi:10.1007/s11240-008-9486-6
Mason HS, Haq TA, Clements JD, Arntzen CJ (1998) Edible
vaccine protects mice against Escherichia coli heat-labile
enterotoxin (LT): potatoes expressing a synthetic LTB
gene. Science 16:336–1343
Pelosi A, Shepherd R, De Guzman G, Hamill JD, Meeusen E,
Sanson G, Walmsley AM (2011) The release and induced
immune responses of a plant-made and delivered antigen
in the mouse gut. Curr Drug Deliv (in press)
Rosales-Mendoza S, Soria-Guerra RE, Lopez-Revilla R,
Moreno-Fierros L, Alpuche-Solı AG (2008) Ingestion of
transgenic carrots expressing the Escherichia coli heat-
labile enterotoxin B subunit protects mice against cholera
toxin challenge. Plant Cell Rep 2008:79–84
Shih SM, Doran PM (2009) Foreign protein production using
plant cell and organ cultures: advantages and limitations.
Biotechnol Adv 27:1036–1042
Sixma TK, Van Zanten BAM, Dauter Z, Hol WGJ (1993)
Refined structure of Escherichia coli heat-labile entero-
toxin, a close relative of cholera toxin. J Mol Biol
230:890. doi:10.1006/jmbi.1993.1209
Tepfer D (1984) Transformation of several species of higher
plants by Agrobacterium rhizogenes: sexual transmission
of the transformed genotype and phenotype. Cell 37:959
van Engelen FA, Molthoff JW, Conner JW, Nap JP, Pereira A,
Stiekema WJ (1995) pBINPLUS—an improved plant
transformation vector based on pBIN19. Transgenic Res
4:288–290
Wagner B, Hufnagl K, Radauer C, Wagner S, Baier K,
Scheiner O, Wiedermann U, Breiteneder H (2004)
Expression of the B subunit of the heat-labile enterotoxin
of Escherichia coli in tobacco mosaic virus-infected
Nicotiana benthamiana plants and its characterization as
mucosal immunogen and adjuvant. J Immunol Methods
287:203–215. doi:10.1016/j.jim.2004.02.001
Walmsley AM, Alvarez ML, Jin Y, Kirk DD, Lee SM,
Pinkhasov J, Arntzen CJ, Rigano MM, Mason HS (2003)
Expression of the B subunit of Escherichia coli heat-labile
enterotoxin as a fusion protein in transgenic tomato. Plant
Cell Rep 21:1020–1026
Wang W, Scali M, Vignani R, Spadafora A, Sensi E, Mazzuca
S, Cresti M (2003) Protein extraction for two-dimensional
electrophoresis from olive leaf, a plant tissue containing
high levels of interfering compounds. Electrophoresis
24:2369–2375. doi:10.1002/elps.200305500
Wen-jun, Forde B (1989) Efficient transformation of Agro-bacterium spp. by high voltage electroporation. Nucleic
Acids Res 17:838–850
Wijsman HJW (1982) On the interrelationships of certain
species of Petunia—taxonomic notes on the parental
species of Petunia hybrida. Acta Bot Neerl 31:478Wilson PD (1997) The pilot-cultivation of transformed hairy
roots. In: Doran PM (ed) Hairy roots: culture and appli-
cation. Overseas Publishers Association, Amsterdam
Biotechnol Lett
123
Author's personal copy
Copyright © 2022 FDOKUMEN




















