Land Snail Models in Island Biogeography: A Tale of Two Snails

ORIGINALARTICLE
Habitats of relict terrestrial snails insouthern Siberia: lessons for thereconstruction of palaeoenvironmentsof full-glacial Europe
Michal Horsak1*, Milan Chytry1, Beata M. Pokryszko2, Jirı Danihelka1,3,
Nikolai Ermakov4, Michal Hajek1,3, Petra Hajkova1,3, Katerina Kintrova1,
Martin Kocı1, Svatava Kubesova1,5, Pavel Lustyk1, Zdenka Otypkova1,
Barbora Pelankova1,3 and Milan Valachovic6
1Department of Botany and Zoology, Masaryk
University, Kotlarska 2, CZ-611 37 Brno,
Czech Republic, 2Museum of Natural History,
Wrocław University, Sienkiewicza 21, PL-50-
335 Wrocław, Poland, 3Institute of Botany,
Academy of Sciences of the Czech Republic,
Porıcı 3a, CZ-603 00 Brno, Czech Republic,4Central Siberian Botanical Garden, Russian
Academy of Sciences, Zolotodolinskaya 101,
Novosibirsk, 630090, Russia, 5Department of
Botany, Moravian Museum, Hviezdoslavova
29a, CZ-627 00 Brno, Czech Republic,6Institute of Botany, Slovak Academy of
Sciences, Dubravska cesta 14, SK-845 23
Bratislava, Slovakia
*Correspondence: Michal Horsak, Department
of Botany and Zoology, Masaryk University,
Kotlarska 2, CZ-611 37 Brno, Czech Republic.
E-mail: [email protected]
ABSTRACT
Aim Shells of fossil molluscs are important for palaeoecological reconstructions.
However, the habitat requirements of snail species typical of central European
full-glacial loess sediments are poorly known because most of them became very
rare or extinct in Europe. The recent discovery of an almost complete extant
assemblage of such species in mountainous regions of central Asia enables more
precise characterization of their habitats, which may significantly improve
reconstructions of Pleistocene environments.
Location Altai Mountains, Russia.
Methods Terrestrial snail assemblages, vegetation composition and selected
environmental variables were recorded at 118 sites along a gradient of climatic
continentality in the Russian Altai. Habitat characteristics of sites where species
typical of the full-glacial period occurred were described using a classification
tree.
Results Seven of the eight species that are typical of central European full-glacial
loess sediments were found in the study area. They were confined to cool areas
with January mean temperatures below )17 �C, but occurred mainly in sheltered
habitats with a warmer microclimate, such as scrub or open woodland. Pupilla
loessica and Vallonia tenuilabris had the broadest habitat range, occurring from
woodland to dry steppe. Unexpectedly, Columella columella, Pupilla alpicola,
Vertigo genesii, V. parcedentata and V. pseudosubstriata were found mainly in
wooded fens and shrubby tundra rather than in open steppe. Most of these seven
species were recorded in base-rich wooded fens. Very dry open steppe habitats
usually supported no snails.
Main conclusions Habitat ranges of the studied snails in the Altai indicate that
the full-glacial landscapes of central European lowlands that harboured these
species were not completely dominated by open and dry loess steppe. Most
probably they contained a significant component of shrubby vegetation, patches
of wet habitats, and probably also areas of woodland at sites with a favourable
mesoclimate.
Keywords
Altai Mountains, climate, Gastropoda, habitat preferences, index species, loess
steppe, palaeoreconstruction, Pleistocene analogue, Russia, vegetation.
Journal of Biogeography (J. Biogeogr.) (2010) 37, 1450–1462
1450 www.blackwellpublishing.com/jbi ª 2010 Blackwell Publishing Ltddoi:10.1111/j.1365-2699.2010.02280.x

INTRODUCTION
Ecological studies of extant faunas and floras that are similar to
species assemblages found in the Pleistocene fossil record
provide important lessons for an understanding of glacial
environments (Anderson et al., 1989; Rousseau, 1991; Moine
& Rousseau, 2002; Schofield et al., 2007; Fløjgaard et al.,
2009). Landscapes of the modern boreal and arctic zones of
northern Europe suggest some similarities to the full-glacial
landscape of central Europe, because they harbour many
species of the central European full glacial, such as Scots pine
(Pinus sylvestris), Norway spruce (Picea abies), dwarf birch
(Betula nana) (Meusel et al., 1965–1992; Willis et al., 2000;
Willis & van Andel, 2004), reindeer (Rangifer tarandus),
muskox (Ovibos moschatus), wolverine (Gulo gulo), Arctic fox
(Alopex lagopus), Norway lemming (Lemmus lemmus) (Ho-
racek & Lozek, 1988; Mitchell-Jones et al., 1999; Musil, 2003;
Pazonyi, 2004; Sommer & Nadachowski, 2006) and several
species of snails (e.g. Columella columella and Vertigo genesii;
Lozek, 1964, 2001; Kerney et al., 1983). However, the northern
European climate, with its low summer temperatures, low
temperature sums of the growing season, and a low ratio of
summer temperatures and precipitation (Rivas-Martınez et al.,
2004), may not be the best modern analogue of the full-glacial
climates for the subcontinental south-eastern part of central
Europe (Frenzel et al., 1992; Guthrie, 2001; Barron & Pollard,
2002; Jost et al., 2005). Indeed, Fennoscandia lacks many
species of the central European full glacial, such as Swiss pine
(Pinus cembra) and European larch (Larix decidua) (Willis
et al., 2000; Willis & van Andel, 2004), the rodents Allactaga
major, Cricetulus migratorius, Lagurus lagurus and Microtus
gregalis (Horacek & Lozek, 1988; Pazonyi, 2004; Sommer &
Nadachowski, 2006) and some species of land snails that are
regarded as index (i.e. indicator) fossils of full-glacial loess
sediments in central Europe, for example Pupilla loessica and
Vallonia tenuilabris (Lozek, 1964, 2001).
The modelled full-glacial climate of central Europe (Barron
& Pollard, 2002; Jost et al., 2005) corresponds closely to the
modern climate in Siberia (Fløjgaard et al., 2009). This
climatic analogue has recently been supported by biological
data. Kunes et al. (2008) demonstrated a close similarity
between full- and late-glacial pollen samples from central
Europe and modern surface-pollen spectra from the Western
Sayan Mountains in southern Siberia. Similar spectra have also
been found in surface pollen from the adjacent Russian Altai
Mountains (Pelankova & Chytry, 2009). These mountain
ranges contain several vegetation types that are dominated by
sibling species (or subspecies) of the trees and shrubs that were
widespread across full-glacial central Europe (Willis & van
Andel, 2004) but retreated to the mountains after the Holocene
climate amelioration. These species include Siberian pine
(Pinus sibirica, related to European P. cembra), Siberian larch
(Larix sibirica, related to European L. decidua) and dwarf birch
(Betula rotundifolia, related to B. nana). Moreover, in the
Altai-Sayan region, vegetation types of taiga, open woodland
and tundra dominated by these species occur in close
proximity to the steppes (Kuminova, 1960), as they did in
full-glacial central Europe (Willis et al., 2000; Jankovska &
Pokorny, 2008).
The Altai Mountains also hold mammal assemblages that
are very similar to the fossil fauna of the European ‘mammoth
steppe’, including Equus spp., Rangifer tarandus, Saiga tatarica,
Gulo gulo, Lepus timidus, Ochotona spp., Allactaga major,
Cricetulus migratorius, Lagurus lagurus, Marmota spp. and
Microtus gregalis (Yudin et al., 1979; Horacek & Lozek, 1988;
Musil, 2003; Pazonyi, 2004; Agadjanian & Serdyuk, 2005;
Ricankova, 2008). This evidence suggests that the Altai-Sayan
region is probably the most significant modern refugium of the
central European full-glacial biota. Its refugial character is also
supported by the fact that fossil pollen spectra from the Altai
and adjacent regions indicate little difference between modern
biomes of this region and those reconstructed for the Last
Glacial Maximum (Tarasov et al., 2000). There was a rapid
spread of coniferous trees at the Pleistocene/Holocene bound-
ary, and an increase in forested area at the expense of treeless
formations (Blyakharchuk et al., 2004). However, there is no
evidence in the available fossil pollen data for the complete
disappearance or new emergence of any major vegetation type
in the Altai-Sayan region. The environment of this region can
be thus considered as rather conservative and stable, at least in
comparison with central Europe.
Recently, the view of the Altai and adjacent mountain ranges
as a refugium of European glacial biota has been confirmed by
a discovery in this area of extant snail assemblages that contain
nearly all the index species from full- and late-glacial central
Europe, including Columella columella, Pupilla alpicola,
P. loessica, Vallonia tenuilabris, Vertigo genesii, V. parcedentata
and V. pseudosubstriata (Pokryszko & Horsak, 2007; Meng,
2008, 2009; Meng & Hoffmann, in press).
Because terrestrial snails are common fossils in all types of
calcium-rich sediments, particularly loess (Lozek, 1964, 2001;
Rousseau, 1990), they are frequently used for the reconstruc-
tion of Quaternary palaeoenvironments (Krolopp, 1983;
Horacek & Lozek, 1988; Rousseau, 1991; Willis et al., 2000;
Moine & Rousseau, 2002; White et al., 2008). Snail fossils have
several important advantages over other types of fossil
material. First, snail shells can be easily identified to species,
whereas plant pollen is often identifiable only to genus or
family. Second, each shell represents one individual, which
enables an accurate estimation of population density. Third,
unlike pollen or vertebrate remains, snail fossils are usually
deposited in places where they lived, thus enabling a fine
spatial resolution of the resulting palaeoenvironmental recon-
struction. Fourth, snail fossils are well preserved in conditions
that are usually poor in other fossils, especially in dry and
calcium-rich sediments.
However, the use of a species for palaeoenvironmental
reconstruction can only be successful if its habitat range and
environmental affinities are sufficiently well known. So far this
has not been the case for several of the index snail species from
European full-glacial loess sediments, because modern ecolog-
ical studies either were confined to restricted parts of their
Snails of glacial Europe extant in southern Siberia
Journal of Biogeography 37, 1450–1462 1451ª 2010 Blackwell Publishing Ltd

range in central European mountains and Fennoscandia (e.g.
Columella columella, Pupilla alpicola, Vertigo genesii and V.
parcedentata; e.g. Kerney et al., 1983; Pokryszko, 1993; Horsak
et al., 2007) or have never been undertaken (e.g. Pupilla
loessica, Vallonia tenuilabris and Vertigo pseudosubstriata).
Some of these species were first described from the fossil
record and extant populations were discovered only some
decades later (e.g. Pokryszko, 1993; Pokryszko & Horsak, 2007;
Meng, 2009; Meng & Hoffmann, in press). The first reports on
the habitats in which some of these species (Vallonia tenui-
labris and Vertigo spp.) live in central Asia have become
available only recently (Meng, 2008, 2009; White et al., 2008).
In this study we attempt to identify and describe the habitats
that harbour the index snail species found in central European
full-glacial loess sediments in their modern refugium in the
Russian Altai Mountains. We describe the vegetation types,
climate and environmental conditions associated with the
occurrence of these species in order to obtain information
necessary for palaeoecological reconstructions of landscapes,
environments and vegetation during the cold periods of the
Pleistocene.
MATERIALS AND METHODS
Study area
This study was undertaken along an approximately 300-km-
long NNW–SSE transect (49�56¢–52�19¢ N, 85�35¢–88�31¢ E,
260–2570 m a.s.l.) running across the Russian part of the Altai
Mountains in the Altai Republic, part of the Russian Feder-
ation, in southern Siberia (Fig. 1). The mountains are formed
of Proterozoic to Devonian siliceous bedrocks such as mica
and chlorite schist and slate, together with local beds of
limestone and some intrusions of granite and other plutonic
rocks (Nekhoroschev, 1966). The high ranges of the Altai
Mountains strongly affect the distribution of precipitation,
which comes mostly with fronts from the north-west. In the
northern Altai foothills, mean annual precipitation locally
exceeds 800 mm, but it decreases sharply with every sub-
sequent mountain range crossed when moving south or south-
east (Gidrometeoizdat, 1966–1970; Polikarpov et al., 1986).
Particularly dry conditions occur in the intermontane basins in
the south-eastern part of the Russian Altai, which receive less
than 180 mm of annual precipitation (Beresneva, 2006). Most
precipitation falls in the summer, but in the south-eastern
basins it comes very irregularly with thunderstorms, and
therefore long periods of drought occur. Temperatures are
generally higher in the northern Altai foothills, and decrease
towards the south-east. Most of the study area is covered with
natural vegetation, which is weakly influenced by humans,
except for grazing in steppes and forest-steppes. Vegetation
varies strongly in response to the NW–SE climatic gradient
(Kuminova, 1960; Walter, 1974).
The northern part of the studied transect includes the Biya
river basin in the northern foothills of the Altai at elevations of
300–700 m. This is the most oceanic part of the study area,
with an annual precipitation of 450–800 mm and mean
January and July temperatures of )14 to )10 �C and 15 to
21 �C, respectively. It is dominated by taiga and hemiboreal
forests, with Abies sibirica, Betula pendula, Picea obovata, Pinus
sibirica, P. sylvestris and Populus tremula. Treeless areas are
covered by mesic meadows. Bogs and mineral-poor fens are
more common in this region than elsewhere in the study area.
The north-western part of the transect includes the Katun’
river valley and adjacent mountain ranges up to the Seminskii
Range in the south. At lower elevations in the Altai foothills
(250–1000 m), annual precipitation is 400–700 mm and mean
temperatures for January and July are )16 to )7 �C and 14 to
21 �C, respectively. There is a forest-steppe landscape consist-
ing of a mosaic of hemiboreal forests with Pinus sylvestris and
Betula pendula and meadow steppes. At higher elevations
(1400–2100 m), annual precipitation is 1200–1400 mm and
mean temperatures for January and July are )24 to )21 �C
and 7 to 10 �C, respectively. Mountain ranges are covered
mainly by sparse Pinus sibirica taiga and, above the timberline,
by shrubby tundra dominated by Betula rotundifolia.
The central part of the transect includes the Katun’ river
valley south of the Seminskii range and the lower part of the
Chuya valley. Owing to its leeward position, annual precip-
itation is only 250–400 mm in the valley bottoms and on the
lower slopes (at 700–1000 m a.s.l.) and about 800–900 mm at
higher elevations (1300–1400 m). Temperatures are )18 to
)15 �C in January and 11 to 15 �C in July. The landscape is a
forest-steppe consisting of a mosaic of Larix sibirica hemibo-
real forests and short-grass steppe, with Picea obovata
occurring in wetter sites of valley bottoms and on north-
facing slopes, and Pinus sibirica at higher elevations, where
it is associated with alpine tundra dominated by Betula
rotundifolia.
Figure 1 Studied sites in the Altai Republic (Russia). Black
squares indicate sites with at least one of the full-glacial index snail
species; white squares are other studied sites.
M. Horsak et al.
1452 Journal of Biogeography 37, 1450–1462ª 2010 Blackwell Publishing Ltd

The south-eastern part of the transect includes the large
intermontane basins along the Chuya River and their adjacent
mountain ranges. The bottom of the Kurai Basin (c. 1500 m
a.s.l.) receives about 150–180 mm of annual precipitation, and
the bottom of the Chuya Basin (c. 1750 m a.s.l.) less than
150 mm. We sampled sites at elevations of 1300–2600 m,
where annual precipitation was 150)550 mm and mean
temperatures of January and July were )23 to )18 �C, and 5
to 12 �C, respectively. There are generally the same vegetation
types as in the central part of the transect, but the area of forest
decreases and that of steppe increases towards drier regions.
The bottoms of the Kurai and Chuya basins are extensive
treeless areas with a short-grass steppe. On some mountain
slopes around the Chuya Basin, forest is entirely absent and
steppe borders directly on alpine tundra.
Sampling
We sampled snail assemblages, vegetation and environmental
variables at 118 sites along the environmental transect of
increasing climate continentality across the Russian Altai
Mountains in August 2005 and July 2006. The sites were
selected to include the whole regional range of habitat types,
determined by physiognomy and dominant plant species.
Sampling plots of 10 · 10 m2 were located in the central parts
of physiognomically and ecologically homogeneous vegetation
stands. Places affected by recent disturbance, for example early
stages of post-fire succession or ruderal vegetation near human
settlements, were not sampled. Multiple sampling of a single
habitat was avoided at nearby sites with similar elevation, slope
and aspect in order to avoid pseudo-replications.
Within each plot, snails were searched for by eye and
collected in appropriate microhabitats for 2 h. In addition, a
5-L sample of leaf litter, moss polsters and topsoil was collected
within each plot from several patches, especially those
considered most favourable for snails. The samples were
washed through a sieve with a 0.6 · 0.6 mm mesh in the field
immediately after sampling, using the wet-sieving method
(Horsak, 2003). Live snails and shells were extracted during
washing and preserved in 80% ethanol or dried out.
Sites were classified into the following 13 habitat types
according to vegetation structure, plant species composition
and abiotic environmental factors. The number of sampled
sites (in brackets) approximately reflects the frequency of a
particular habitat in the landscape. Taiga (n = 11): mesic to
wet coniferous forest with a species-poor herb layer and a well-
developed moss layer, usually dominated by Abies sibirica,
Larix sibirica, Picea obovata or Pinus sibirica. Hemiboreal
forest (n = 22): dry to mesic coniferous or deciduous forest
with a species-rich herb layer and a sparse moss layer, usually
dominated by Betula pendula, Larix sibirica or Pinus sylvestris.
Wooded fen (n = 2): wet woodland with accumulation of
base-rich organic sediment, dominated by Picea obovata.
Treeless fen (n = 4): open base-rich fen with sedges and
mosses. Acidic mire (n = 7): bogs and mineral-poor fens.
Alluvial scrub (n = 3): riverine woody vegetation with willows
(Salix spp.). Tall-forb vegetation (n = 5): tall herbaceous
vegetation in wet places, usually along mountain streams.
Alpine grassland (n = 3): short herbaceous vegetation above
the timberline. Shrubby tundra (n = 8): low-shrub vegetation
above the timberline, dominated mainly by Betula rotundifolia
(= B. nana s.l.), in places also by Dryas oxyodonta. Steppe
(n = 36): both short-grass steppe and tall-grass (meadow)
steppe, in places with low shrubs such as Caragana or Spiraea.
Saline grassland (n = 4): grassland on soils with elevated salt
concentration, usually in shallow depressions in steppe land-
scapes. Meadow (n = 8): mesic grassland used for regular or
occasional hay-making. Scree (n = 5): treeless talus slopes.
In each plot, all species of vascular plants and ground-
dwelling bryophytes and lichens were recorded, with their
cover-abundances estimated on the Braun-Blanquet scale (van
der Maarel, 1979). The following environmental variables were
compiled for each plot (Table 1). (1) Elevation, measured with
a GPS receiver (Garmin 60CSx) in the field. (2–7) Climatic
data, including annual, winter (November–March) and su-
mmer (April–October) precipitation sums and mean annual,
January and July temperatures, were taken from a spatially
explicit climatic model (Chytry et al., 2007) based on climate-
station data interpolated over a digitized 1:200,000 map using
the ArcGIS 8.2 Geographic Information System (ESRI, 2003).
Temperature interpolations considered local topography,
using an adiabatic lapse rate of 0.65 �C per 100 m of altitude.
Precipitation was computed from the precipitation-elevation
charts compiled for each of the aridity-humidity sectors of the
Altai-Sayan region (Polikarpov et al., 1986). Summer and
winter precipitation were strongly correlated (r = 0.992,
P < 0.001), as were January and July temperatures (r =
0.710, P = 0.001). (8) Slope inclination was measured in the
field with a clinometer (Clino Master, Silva, Sollentuna,
Sweden). (9) Radiation index was calculated from slope
inclination and aspect according to McCune & Keon (2002,
eq. 3). The values of this index are high on steep south-facing
slopes, low on steep north-facing slopes and medium on flat
land, gentle slopes, and steep east- or west-facing slopes. (10–
12) Potential radiation, accounting for shading by adjacent
topographic features and forest canopy, was calculated from
hemispheric photographs taken in the vertical direction at a
height of 1.5 m above ground in each plot, using a Nikon
CoolPix 4500 digital camera with a fish-eye lens. Radiation was
estimated as a daily sum of direct, diffuse, and total (= direct +
diffuse) radiation for 21 June in mol m)2 day)1. (13–16)
Percentage covers of trees, shrubs, herbs and bryophytes/
lichens were estimated by eye in the field. (17–19) Depths of
soil, peat and litter layers were measured using a metal rod.
(20) Percentage cover of litter was estimated by eye. (21–22)
Soil pH and electrical conductivity were measured in a 2:5
soil:water solution of mixed soil samples taken at four places in
each plot from a depth of 5–10 cm or less if the soil was very
shallow. A GPRT 1400 AN pH-meter (Greisinger Electronic
GmbH, Regenstauf, Germany) and a CM113 conductivity
meter (Snail Instruments, Beroun, Czech Republic) were used,
respectively. Conductivity caused by H+ ions was subtracted
Snails of glacial Europe extant in southern Siberia
Journal of Biogeography 37, 1450–1462 1453ª 2010 Blackwell Publishing Ltd
according to Sjors (1952). (23–24) Numbers of tree species and
vascular plant species were recorded for each plot.
Data analysis
To define indicator snail species of the full-glacial landscapes
of central Europe we adopted Lozek’s classification of fossil
molluscs and his concept of index species (Lozek, 2001). The
vast majority of fossil records of full-glacial index species are
confined to loess sediments, and at present these species
are either extinct or survive in remote areas with a climate
different from the modern climate of central Europe (Lozek,
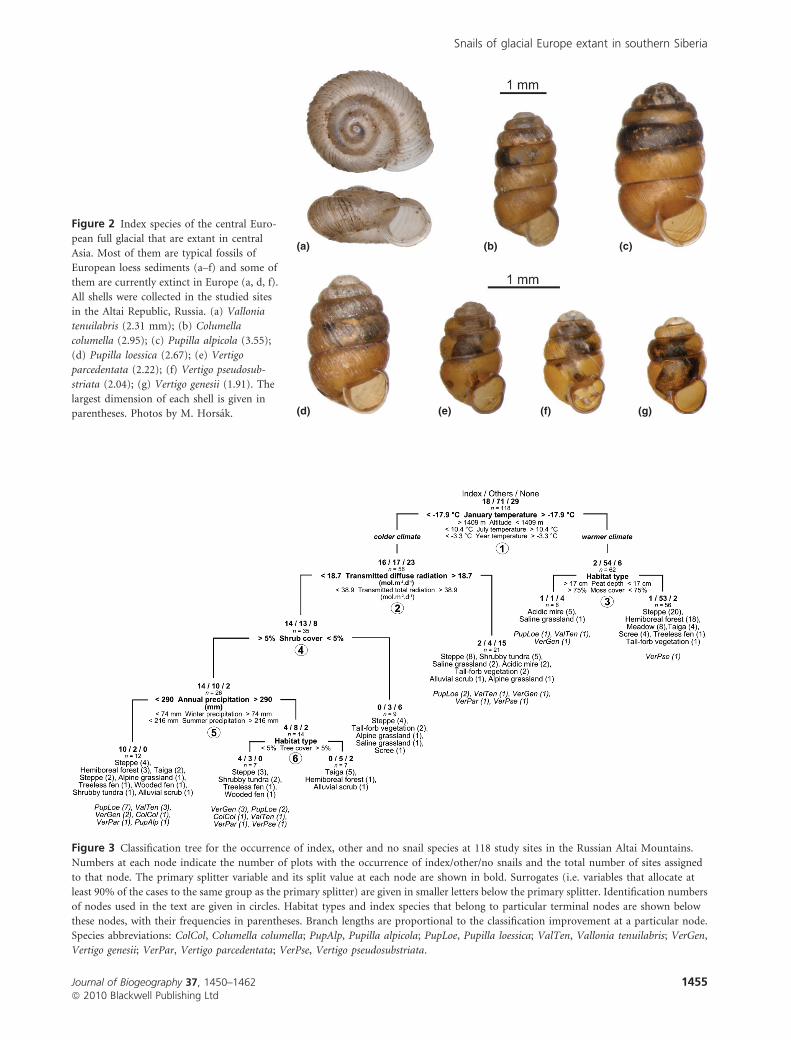
2001). We considered the following seven species as full-glacial
index species: Columella columella, Pupilla alpicola, P. loessica,
Vallonia tenuilabris, Vertigo genesii, V. parcedentata and
V. pseudosubstriata (Fig. 2). Vertigo genesii was included, even
though it is very rare in loess sediments (Lozek, 2001);
however, it became more common during the Late Glacial
period (Lozek, 1964). Its rarity in the dry full-glacial
landscapes was probably linked to the rarity of suitable
habitat, namely calcareous marshes.
The 118 plots were classified into three categories: (1) plots
containing at least one of the seven index species; (2) plots
containing other snails only; and (3) plots containing no snails.
Hereafter, these categories of plots are termed Index, Others
and None.
The relationships between these three categories, habitat
types and environmental variables were analysed using classi-
fication trees (CART; Breiman et al., 1984; De’ath & Fabricius,
2000). This method is suitable for exploratory analysis
of complex unbalanced data sets that involve higher-order
interactions and nonlinear relationships between predictor
variables. It reveals ecologically meaningful relationships that
might remain hidden if standard techniques, such as linear
models, were used. To select the optimal tree size (with the
optimal number of nodes) we used a complexity parameter (cp)
that weighs both the tree complexity and the sum of errors in
the classification of a new set of sites (i.e. a test data set). This is
usually computed in a cross-validation process that enables the
utilization of all information available. We worked with a
cross-validation function of the rpart package, R 2.9.2
software (R Development Core Team, 2009) with 10 subsets,
and built the tree with constraints on the minimal number of
sites to be divided (10) and the minimal number of sites in
terminal nodes (5). The cross-validation process was repeated
500 times with random data subsets in each repetition, and the
complexity parameter for every node of all 500 trees was
recorded. The complexity level of the tree that most often
reached the lowest cp-value was chosen. For each node of the
tree, apart from the primary splitter variable, surrogates (i.e.
variables allocating at least 90% of the cases similarly to the
primary splitter) were identified.
RESULTS
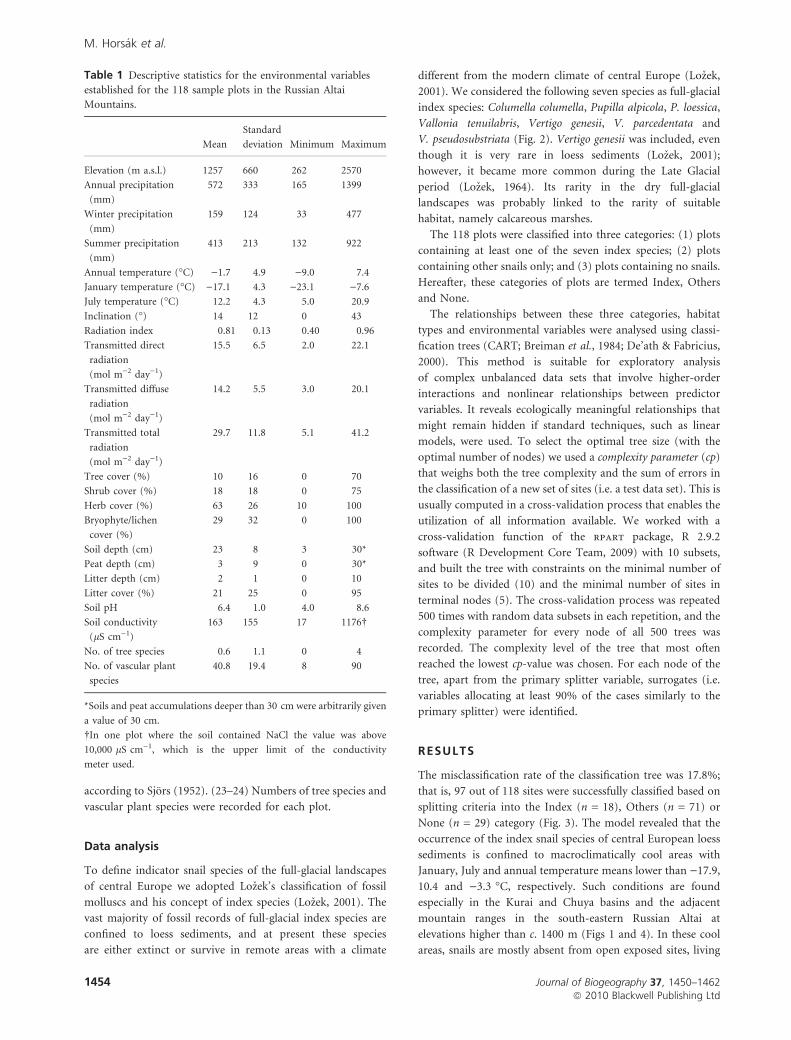
The misclassification rate of the classification tree was 17.8%;
that is, 97 out of 118 sites were successfully classified based on
splitting criteria into the Index (n = 18), Others (n = 71) or
None (n = 29) category (Fig. 3). The model revealed that the
occurrence of the index snail species of central European loess
sediments is confined to macroclimatically cool areas with
January, July and annual temperature means lower than )17.9,
10.4 and )3.3 �C, respectively. Such conditions are found
especially in the Kurai and Chuya basins and the adjacent
mountain ranges in the south-eastern Russian Altai at
elevations higher than c. 1400 m (Figs 1 and 4). In these cool
areas, snails are mostly absent from open exposed sites, living
Table 1 Descriptive statistics for the environmental variables
established for the 118 sample plots in the Russian Altai
Mountains.
Mean
Standard
deviation Minimum Maximum
Elevation (m a.s.l.) 1257 660 262 2570
Annual precipitation
(mm)
572 333 165 1399
Winter precipitation
(mm)
159 124 33 477
Summer precipitation
(mm)
413 213 132 922
Annual temperature (�C) )1.7 4.9 )9.0 7.4
January temperature (�C) )17.1 4.3 )23.1 )7.6
July temperature (�C) 12.2 4.3 5.0 20.9
Inclination (�) 14 12 0 43
Radiation index 0.81 0.13 0.40 0.96
Transmitted direct
radiation
(mol m)2 day)1)
15.5 6.5 2.0 22.1
Transmitted diffuse
radiation
(mol m)2 day)1)
14.2 5.5 3.0 20.1
Transmitted total
radiation
(mol m)2 day)1)
29.7 11.8 5.1 41.2
Tree cover (%) 10 16 0 70
Shrub cover (%) 18 18 0 75
Herb cover (%) 63 26 10 100
Bryophyte/lichen
cover (%)
29 32 0 100
Soil depth (cm) 23 8 3 30*
Peat depth (cm) 3 9 0 30*
Litter depth (cm) 2 1 0 10
Litter cover (%) 21 25 0 95
Soil pH 6.4 1.0 4.0 8.6
Soil conductivity
(lS cm)1)
163 155 17 1176�
No. of tree species 0.6 1.1 0 4
No. of vascular plant
species
40.8 19.4 8 90
*Soils and peat accumulations deeper than 30 cm were arbitrarily given
a value of 30 cm.
�In one plot where the soil contained NaCl the value was above
10,000 lS cm)1, which is the upper limit of the conductivity
meter used.
M. Horsak et al.
1454 Journal of Biogeography 37, 1450–1462ª 2010 Blackwell Publishing Ltd
(a)
(d) (e) (f) (g)
(b) (c)
Figure 2 Index species of the central Euro-
pean full glacial that are extant in central
Asia. Most of them are typical fossils of
European loess sediments (a–f) and some of
them are currently extinct in Europe (a, d, f).
All shells were collected in the studied sites
in the Altai Republic, Russia. (a) Vallonia
tenuilabris (2.31 mm); (b) Columella
columella (2.95); (c) Pupilla alpicola (3.55);
(d) Pupilla loessica (2.67); (e) Vertigo
parcedentata (2.22); (f) Vertigo pseudosub-
striata (2.04); (g) Vertigo genesii (1.91). The
largest dimension of each shell is given in
parentheses. Photos by M. Horsak.
Figure 3 Classification tree for the occurrence of index, other and no snail species at 118 study sites in the Russian Altai Mountains.
Numbers at each node indicate the number of plots with the occurrence of index/other/no snails and the total number of sites assigned
to that node. The primary splitter variable and its split value at each node are shown in bold. Surrogates (i.e. variables that allocate at
least 90% of the cases to the same group as the primary splitter) are given in smaller letters below the primary splitter. Identification numbers
of nodes used in the text are given in circles. Habitat types and index species that belong to particular terminal nodes are shown below
these nodes, with their frequencies in parentheses. Branch lengths are proportional to the classification improvement at a particular node.
Species abbreviations: ColCol, Columella columella; PupAlp, Pupilla alpicola; PupLoe, Pupilla loessica; ValTen, Vallonia tenuilabris; VerGen,
Vertigo genesii; VerPar, Vertigo parcedentata; VerPse, Vertigo pseudosubstriata.
Snails of glacial Europe extant in southern Siberia
Journal of Biogeography 37, 1450–1462 1455ª 2010 Blackwell Publishing Ltd
mainly in places protected by tree canopy or adjacent
topographic features (Fig. 3, node 2) and containing at least
5% of shrub cover (node 4). The index species prevailed in
areas with mean annual precipitation lower than 290 mm or
outside woodlands in areas with higher precipitation (nodes 5
and 6), whereas other species were more common in
woodlands in areas of higher precipitation (node 6).
Wooded fens dominated by Picea obovata, although repre-
sented by only two sites, harboured all the index species except
Pupilla loessica (Table 2, Fig. 4). Four of the seven index
species were also recorded in tundra with low shrubs such as
Betula rotundifolia and in places also B. humilis, Dryas
oxyodonta, Pentaphylloides fruticosa and Salix spp. The other
habitats contained only 1–3 index species. Pupilla loessica was
the most frequent index species, recorded at 13 sites and in
seven habitats. Two other index species, Vertigo genesii (8 sites)
and Vallonia tenuilabris (7 sites), were also relatively common,
found in four and five habitats, respectively. The remaining
four species were recorded at fewer than four sites and in fewer
than three habitats.
Full-glacial index species were most often found in wood-
lands with Larix sibirica or Picea obovata as well as in shrubby
habitats with the above-mentioned tundra shrubs, or in steppe
landscapes, with drought-adapted shrubs such as Caragana
Table 2 Habitat types and ranges of environmental variables associated with the occurrence of the studied index snail species in the Russian
Altai Mountains. For each species and habitat the range of the number of individuals found at particular sites is given. For snail species
authorities see Appendix S1.
Columella
columella
Pupilla
alpicola
Pupilla
loessica
Vallonia
tenuilabris
Vertigo
genesii
Vertigo
parcedentata
Vertigo
pseudosubstriata
Total no. of sites/individuals 2/83 1/33 13/133 7/90 8/650 3/23 3/28
Habitat type (number of sites/total number of all snail species)
Shrubby tundra (3/11) 0 0 3–7 0 1–4 2–17 23
Taiga (1/8) 0 0 1–8 2 4 0 0
Hemiboreal forest (3/9) 0 0 3–18 29 0 0 0
Wooded fen (3/11) 21–62 33 0 15–18 37–410 4 4
Alpine grassland (1/7) 0 0 6 3 0 0 0
Treeless fen (2/3) 0 0 0 0 73–116 0 0
Meadow (1/7) 0 0 0 0 0 0 1
Steppe (3/13) 0 0 1–53 9 0 0 0
Saline grassland (1/4) 0 0 8 14 5 0 0
Environmental variable
Elevation (m a.s.l.) 1354–1731 1731 1342–2490 1342–2059 1330–1998 1731–1994 1354–1994
Annual precipitation (mm) 196–522 195 205–879 195–522 195–880 195–821 522–821
Winter precipitation (mm) 41–164 40.9 45–308 44–164 41–308 41–308 164–283
Summer precipitation (mm) 154–358 154 160–572 154–358 154–572 154–538 358–586
Annual temperature (�C) )5.3–()3.9) )5.3 )8.2–()0.7) )7.8–()3.1) )8.2–()3.1) )8.2–()5.3) )7.5–1.0
January temperature (�C) )19.5–()18.0) )19.5 )22.2–()17.1) )21.9–()17.1) )22.1–()17.1) )22.1–()19.5) )21.4–()18.0)
July temperature (�C) 8.3–10.1 8.3 5.7–11.1 6.1–10.8 5.7–10.8 5.7–8.3 6.4–11.4
Inclination (�) 1–2 1 0–33 0–33 0–19 1–19 2–19
Radiation index 0.86–0.87 0.86 0.45–0.95 0.45–0.90 0.84–0.88 0.84–0.87 0.82–0.87
Transmitted direct radiation
(mol m)2 day)1)
5.3–13.4 13.4 4.4–21.7 4.4–21.3 5.3–21.2 13.4–21.2 5.3–21.2
Transmitted diffuse radiation
(mol m)2 day)1)
5.7–11.6 11.6 5.0–19.9 5.0–19.9 5.7–19.2 11.6–19.2 5.7–19.2
Transmitted total radiation
(mol m)2 day)1)
10.9–25.0 25 9.4–40.4 9.4–39.6 10.9–40.4 25.0–40.4 11.0–40.4
Tree cover (%) 15–20 15 0–35 0–35 0–20 0–15 0–20
Shrub cover (%) 25–40 25 0–70 0–50 0–70 25–70 5–70
Herb cover (%) 30–60 60 10–95 10–80 10–95 60–95 30–100
Bryophyte/lichen cover (%) 70–80 70 5–90 5–90 5–90 40–70 40–80
Soil depth (cm) ‡30 ‡30 5 to ‡30 5 to ‡30 5 to ‡30 10 to ‡30 10 to ‡30
Peat depth (cm) 0 to ‡30 ‡30 0–12 0 to ‡30 0 to ‡30 0 to ‡30 15 to ‡30
Litter depth (cm) 3–15 3 1–10 1–15 1–15 1–3 1–15
Litter cover (%) 10–30 30 1–80 1–30 1–30 3–30 3–15
Soil pH 6.8–7.5 6.8 5.4–8.6 6.1–8.6 5.4–8.6 5.4–6.8 5.8–7.5
Soil conductivity (lS cm)1) 318–408 408 40–1176 40–1176 56–1176 56–408 17–318
No. tree species 2–3 3 0–2 0–3 0–3 0–3 0–2
No. vascular plant species 34–35 35 25–58 33–56 25–55 35–43 34–45
M. Horsak et al.
1456 Journal of Biogeography 37, 1450–1462ª 2010 Blackwell Publishing Ltd

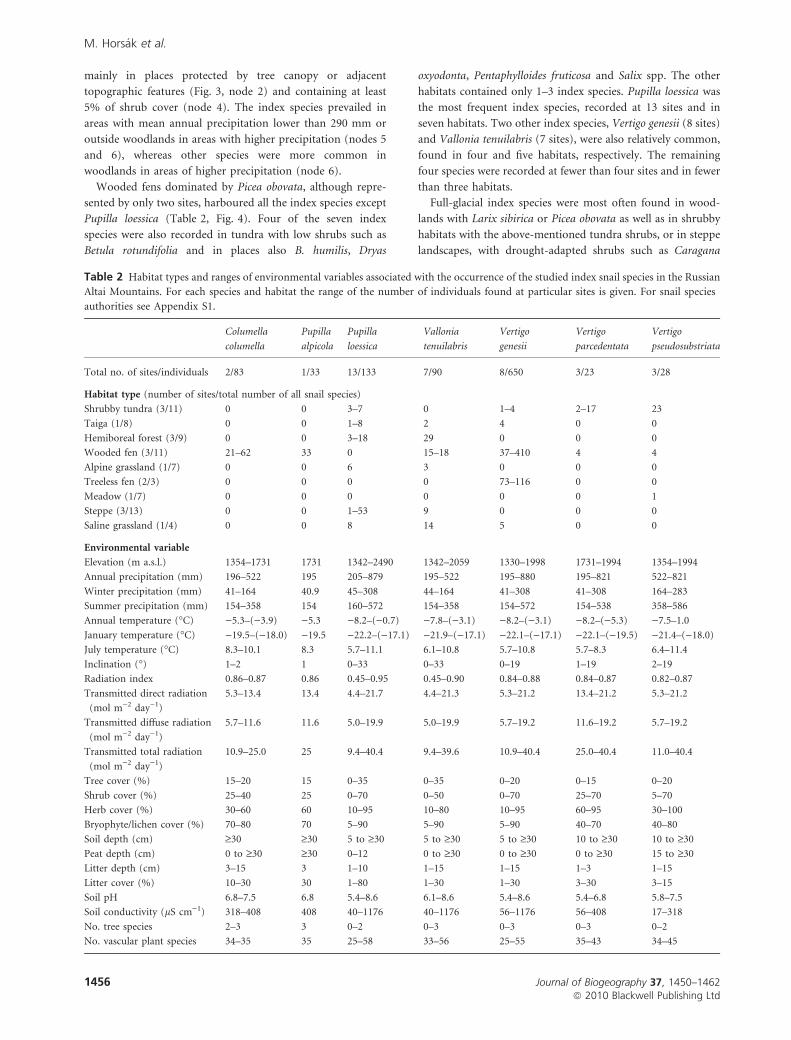
Figure 4 Habitats harbouring the studied index snail species in the Russian Altai Mountains. 1, taiga; 2, hemiboreal forest; 3, wooded
fen; 4, treeless fen; 5, alpine grassland; 6, shrubby tundra; 7, steppe; 8, saline grassland; 9, meadow. Photos by P. Hajkova (1–5, 7, 8),
Z. Otypkova (6) and M. Chytry (9). A detailed description and a list of snail species recorded at each site are available in Appendix S1 in the
Supporting Information.
Snails of glacial Europe extant in southern Siberia
Journal of Biogeography 37, 1450–1462 1457ª 2010 Blackwell Publishing Ltd

arborescens, Cotoneaster uniflorus, Rosa pimpinellifolia and
Spiraea media (68% of all records, see Fig. 4). Only two
species, Pupilla loessica and Vallonia tenuilabris, were also
frequently recorded in treeless habitats, although they were still
more frequent in woodlands and scrub. At two sites we
sampled both a wooded fen and an adjacent open fen lacking
trees and shrubs. In both cases, wooded fens were richer in
snail species and contained several index species, whereas the
treeless areas of the same fen contained fewer than four species
of land snails, of which only one, Vertigo genesii, was an index
species. However, this species occurred there in high densities.
DISCUSSION
Our data confirm that the Altai Mountains are an important
refugium for full-glacial snail faunas, as recently documented
by Pokryszko & Horsak (2007), Meng (2008, 2009) and Meng
& Hoffmann (in press). Extant populations of index snail
species of the central European full glacial are significantly
associated with a cold macroclimate in the southern part of the
Russian Altai. Of the 18 sites where these species were found,
16 had a mean January temperature below )17.9 �C and for
the other two it was below )17.1 �C. None of these sites had
mean July and annual temperatures higher than 11.4 and
1.0 �C, respectively (Table 2). These values correspond to
those modelled for the central European full-glacial period
(Barron & Pollard, 2002). The temperature limitation of the
full-glacial index snail species suggests that their niches, like
those of plants (Martınez-Meyer & Peterson, 2006), mammals
(Martınez-Meyer et al., 2004) and probably other animal
groups, remained relatively stable over several millennia.
However, their affinity to cool environments contrasts with
the fact that land snails, as poikilotherms, are generally
favoured by warmer conditions (e.g. Bishop, 1977; Cameron
& Greenwood, 1991) because of their sensitivity to superco-
oling. It is known that small snail species are able to survive
under cold conditions better that those with large bodies
(Ansart & Vernon, 2003), but both groups need specific shelter
to survive. This raises the question why these species were not
also found in warmer areas of the Altai Mountains in spite of
the fact that suitable habitats and source populations were
available in adjacent areas and that sites from warmer areas
were well represented in our samples. This pattern can be
explained either by physiological constraints on the species
fundamental niche, or by biotic interactions. Physiological
constraints that exclude these species from warmer areas may
have evolved in the full-glacial landscapes, which probably
contained few microclimatically warmer or otherwise favour-
able habitats. Competitive exclusion of the index snail species
by other snails in warmer areas is unlikely, because snail
assemblages of warmer areas, although richer in species than
those of cold areas, are still distinctly poorer in species than the
snail assemblages of temperate Europe (e.g. Martin & Sommer,
2004; Sulikowska-Drozd & Horsak, 2007) and include species
that would hardly be able to outcompete all the index species of
the region. However, further biotic mechanisms could control
this distributional pattern, for example competitive pressure of
snail predators (Barker, 2004) occurring only in warmer areas.
An important finding for the reconstruction of full-glacial
environments is the preference of index snail species for wet or
microclimatically favourable wooded or shrubby habitats.
Meng (2008, 2009) observed a similar pattern in various
mountain ranges of central Asia, where 70% of all records of
Vallonia tenuilabris and most records of Vertigo genesii and V.
parcedentata were from wet habitats. Vallonia tenuilabris has
also been previously reported from various forest habitats in
southern Siberia (Schileyko, 1984, p. 169; White et al., 2008).
Although it has traditionally been believed that trees were
virtually absent in central European full-glacial landscapes with
loess accumulation, the above-mentioned data indicate the
existence of patches of open woodland or scrub in the loess
sites from which fossils of the index snails are known. This is in
accordance with recent palaeobotanical evidence of several tree
species occurring in the full-glacial landscapes of eastern-
central Europe (Rybnıckova & Rybnıcek, 1991; Willis et al.,
2000; Jankovska et al., 2002; Willis & van Andel, 2004; Birks &
Willis, 2008; Jankovska & Pokorny, 2008). Modelling exercises
also indicated the climatic suitability of full-glacial central
Europe for coniferous trees and other species with modern
boreal distributions (Alfano et al., 2003; Svenning et al., 2008;
Fløjgaard et al., 2009). We found the index snail species to
occur in Altaian woodlands dominated by Larix sibirica and
Picea obovata, that is, by sibling species of European L. decidua
and P. abies, respectively, which are both reported from central
European full-glacial fossil pollen and charcoal. Thus, our data
on the Altai snail assemblages provide further evidence of
northern tree refugia in full-glacial Europe. They are consistent
with the reconstruction of the full-glacial landscape in central
Europe as a mosaic of loess steppes, shrubby tundra, and
woodland patches in mesoclimatically suitable pockets (Rud-
ner & Sumegi, 2001; Kunes et al., 2008).
Shrubby or woodland vegetation with a favourable micro-
climate in the undergrowth may have been the main full-
glacial habitat of all snail species included in the present study
except Vertigo genesii. In the full-glacial sediments this snail is
known only from calcareous fens and very wet habitats (Lozek,
1964). The same pattern was observed in the extant snail
assemblages in the Altai, where the most abundant populations
of this species were encountered in treeless, calcium-rich fens.
In northern Europe this species is also characteristic of base-
rich treeless fens (Speight et al., 2003), but it is also reported
from wooded mires (Valovirta, 2003). Preferences for wet,
base-rich fens are also known for northern European popu-
lations of Columella columella and Vertigo parcedentata,
although the former occurs also in wet subarctic woodlands,
grey willow thickets and alpine meadows (Kerney et al., 1983;
Pokryszko, 1990, 1993). In central European high mountains,
C. columella can also inhabit grasslands on limestone outcrops
above 1500 m a.s.l. (Lozek, 1964; Pokryszko, 1990).
Another full-glacial index snail species that may not
necessarily have been linked with woody vegetation is Pupilla
loessica. This species was described from the central European
M. Horsak et al.
1458 Journal of Biogeography 37, 1450–1462ª 2010 Blackwell Publishing Ltd

fossil record (Lozek, 1954) and its extant populations were
reliably recognized only recently in central Asia (Meng, 2009;
Meng & Hoffmann, in press). It seems to be the species most
tolerant of cold conditions, which are unfavourable for the
other index snail species. Pupilla loessica was more often
recorded at open steppe sites. This observation is in accor-
dance with the fossil record, because this snail is most
abundant in loess sediments from the Last Glacial Maximum
(e.g. Lozek, 1964, 2001). The areas and sites in the Altai
Mountains with the exclusive occurrence of this snail have a
fossil analogue in the drier phases of full-glacial loess steppes
(e.g. Lozek, 1989). We also found that this species can occupy a
broad range of habitats, including all types of habitats
supporting the other index species except fens. This is in
accordance with its abundant occurrence in most loess
sediments from central Europe (Lozek, 1964, 2001). A
similarly broad range of habitats was found for Vallonia
tenuilabris, which is another very common species associated
with loess sediments (Table 2). The broad niche of both
species in cool climatic conditions could be the reason why
they survived glacial periods of the European Pleistocene. They
also occurred in drier phases, which were characterized by
species-poor assemblages dominated by Pupilla spp. (Lozek,
1964; Willis et al., 2000). This Pupilla-rich fauna is conse-
quently regarded as aridity-tolerant. In contrast, the other
index species were mostly characteristic of the youngest loess
deposits (30,000–14,000 yr bp; Lozek, 2001). The basal part of
this thick depositional sequence includes several horizons of
pale grey tundra gley soils, indicating a wetter climate (Klıma,
1958) than in previous glacial periods. This basal part is
characterized by rich snail assemblages with all the loess index
species (Lozek, 2001). This corresponds perfectly to the
modern situation observed in the Altai, where dry steppe
habitats are inhabited mostly by species-poor Pupilla/Vallonia
faunas. However, the driest steppes at exposed sites, which
contain no or an insignificant cover of shrubs, are usually
devoid of all snail species. This indicates that the environ-
mental conditions at the bottoms of the driest basins in the
south-eastern Russian Altai (Kurai and Chuya Basins) are
probably harsher than those of many full-glacial landscapes of
central Europe.
It should be noted that there are some differences between
the studied snail faunas and the full-glacial snail assemblages of
central Europe (see Lozek, 2001 and lists of species recorded in
the Altai in Appendix S1 in the Supporting Information).
There are several species unique to each of these two contexts;
however, most differences can be easily explained by a limited
distribution or different biogeography of the higher taxa to
which these species belong. For example, one of the most
abundant species of European loess sediments was Helicopsis
striata, which has never occurred outside Europe. Indeed, there
are about 5000 km between central Europe and the Altai – a
distance that could cause differences in species composition
even within the environmentally homogeneous loess steppe
biome. However, an important finding is that, except for one
index species of the full-glacial loess sediments of central
Europe, Catinella antiqua, all index species were found extant
in the study area.
In conclusion, the occurrence of relict snail assemblages,
coupled with relict assemblages of plants and mammals,
confirms that the landscape of the Altai Mountains can be
regarded as the best known modern analogue of the last full-
glacial period of central Europe. Even within this region of
continental climate, the index snail species of the full-glacial
loess sediments are limited to colder areas. In these areas their
current habitat ranges suggest that the full-glacial landscapes of
central European lowlands very probably contained a signif-
icant component of shrub vegetation, patches of wet habitats,
and probably also patches of woodland at sites with a
favourable mesoclimate (Fig. 4). These results add to the
growing body of literature demonstrating that full-glacial
landscapes of the central European lowlands were not com-
pletely treeless as had traditionally been believed.
ACKNOWLEDGEMENTS
We thank the local staff of our expeditions for their varied
support, the employees of the Kamlak field station for
accommodation and a warm welcome, Vojen Lozek for
inspiring discussions, Katrin Karimova for processing the
hemispheric photographs, Ondrej Hajek for preparing the map
and Denis Popov for developing the climatic models. Very
useful comments and editorial work from John Birks, Pim van
der Knaap, Richard Preece and Vera Ricankova are greatly
appreciated. This research was funded by the Ministry of
Education of the Czech Republic (MSM0021622416) and the
Academy of Sciences of the Czech Republic (IAA6163303,
AVOZ60050516).
REFERENCES
Agadjanian, A.K. & Serdyuk, N.V. (2005) The history of
mammalian communities and paleogeography of the Altai
Mountains in the Paleolithic. Paleontological Journal, 39
(Suppl. 6), S645–S821.
Alfano, M.J., Barron, E.J., Pollard, D., Huntley, B. & Allen,
J.R.M. (2003) Comparison of climate model results with
European vegetation and permafrost during oxygen isotope
stage three. Quaternary Research, 59, 97–107.
Anderson, P.M., Bartlein, P.J., Brubaker, L.B., Gajewski, K. &
Ritchie, J.C. (1989) Modern analogues of late-Quaternary
pollen spectra from the western interior of North America.
Journal of Biogeography, 16, 573–596.
Ansart, A. & Vernon, P. (2003) Cold hardiness in molluscs.
Acta Oecologica, 24, 95–102.
Barker, G.M. (Ed.) (2004) Natural enemies of terrestrial mo-
lluscs. CABI Publishing, Wallingford, UK.
Barron, E. & Pollard, D. (2002) High-resolution climate sim-
ulations of Oxygen Isotope Stage 3 in Europe. Quaternary
Research, 58, 296–309.
Beresneva, I.A. (2006) Klimaty aridnoi zony Azii [Climates of
the arid zone of Asia]. Nauka, Moskva (in Russian).
Snails of glacial Europe extant in southern Siberia
Journal of Biogeography 37, 1450–1462 1459ª 2010 Blackwell Publishing Ltd

Birks, H.J.B. & Willis, K.J. (2008) Alpines, trees, and refugia in
Europe. Plant Ecology and Diversity, 1, 147–160.
Bishop, M.J. (1977) The habitats of Mollusca in the central
Highlands of Scotland. Journal of Conchology, 29, 189–197.
Blyakharchuk, T.A., Wright, H.E., Borodavko, P.S., van der
Knaap, W.O. & Ammann, B. (2004) Late Glacial and
Holocene vegetational changes on the Ulagan high-
mountain plateau, Altai Mountains, southern Siberia.
Palaeogeography, Palaeoclimatology, Palaeoecology, 209, 259–
279.
Breiman, L., Friedman, J.H., Olshen, R.A. & Stone, C.G. (1984)
Classification and regression trees. Wadsworth International
Group, Belmont, CA.
Cameron, R.A.D. & Greenwood, J.J.D. (1991) Some montane
and forest molluscan faunas from eastern Scotland: effects of
altitude, disturbance and isolation. Proceedings of the 10th
International Malacological Congress (ed. by C. Meier-
Brook), pp. 437–442. Unitas Malacologica, Tubingen.
Chytry, M., Danihelka, J., Ermakov, N., Hajek, M., Hajkova,
P., Kocı, M., Kubesova, S., Lustyk, P., Otypkova, Z., Popov,
D., Rolecek, J., Reznıckova, M., Smarda, P. & Valachovic, M.
(2007) Plant species richness in continental southern Sibe-
ria: effects of pH and climate in the context of the species
pool hypothesis. Global Ecology and Biogeography, 16, 668–
678.
De’ath, G. & Fabricius, K.E. (2000) Classification and regre-
ssion trees: a powerful yet simple technique for ecological
data analysis. Ecology, 81, 3178–3192.
ESRI (2003) ArcGIS 8.3. Environmental Systems Research
Institute, Redlands. Available at: http://www.esri.com.
Fløjgaard, C., Normand, S., Skov, F. & Svenning, J.C. (2009)
Ice age distributions of European small mammals: insights
from species distribution modelling. Journal of Biogeography,
36, 1152–1163.
Frenzel, B., Pecsi, M. & Velichko, A.A. (eds) (1992) Atlas of
paleoclimates and paleoenvironments of the Northern Hemi-
sphere. Geographical Research Institute, Budapest and
Gustav Fischer Verlag, Stuttgart.
Gidrometeoizdat (1966–1970) Spravochnik po klimatu SSSR
[Reference books on the climate of the USSR]. Gidro-
meteoizdat, Leningrad (in Russian).
Guthrie, R.D. (2001) Origin and causes of the mammoth
steppe: a story of cloud cover, woolly mammal tooth pits,
buckles, and inside-out Beringia. Quaternary Science
Reviews, 20, 549–574.
Horacek, I. & Lozek, V. (1988) Palaeozoology and the Mid-
European Quaternary past: scope of the approach and
selected results. Rozpravy Ceskoslovenske akademie ved,
Rada matematickych a prırodnıch ved, 98/4, 1–103.
Horsak, M. (2003) How to sample mollusc communities in
mires easily. Malacologica Bohemoslovaca, 2, 11–14.
Horsak, M., Hajek, M., Tichy, L. & Jurickova, L. (2007) Plant
indicator values as a tool for land mollusc autecology
assessment. Acta Oecologica, 32, 161–171.
Jankovska, V., Chromy, P. & Niznianska, M. (2002) Safarka –
first palaeobotanical data of the character of Last Glacial
vegetation and landscape in the West Carpathians (Slova-
kia). Acta Palaeobotanica, 42, 39–50.
Jankovska, V. & Pokorny, P. (2008) Forest vegetation of the
last full-glacial period in the Western Carpathians (Slovakia
and Czech Republic). Preslia, 80, 307–324.
Jost, A., Lunt, D., Kageyama, M., Abe-Ouchi, A., Peyron, O.,
Valdes, P.J. & Ramstein, G. (2005) High-resolution simu-
lations of the last glacial maximum climate over Europe:
a solution to discrepancies with continental palaeoclimatic
reconstructions? Climate Dynamics, 24, 577–590.
Kerney, M.P., Cameron, R.A.D. & Jungbluth, J.H. (1983) Die
Landschnecken Nord- und Mitteleuropas. Verlag Paul Parey,
Hamburg & Berlin.
Klıma, B. (1958) Prıspevek ke stratigrafii nejmladsıho
sprasoveho pokryvu [A contribution to the stratigraphy of
the youngest loess deposits]. Anthropozoikum, 7, 111–143
(in Czech).
Krolopp, E. (1983) Biostratigraphic division of Hungarian
Pleistocene formations according to their molluscan fauna.
Acta Geologica Academiae Scientiarum Hungaricae, 26, 69–
82.
Kuminova, A.V. (1960) Rastitel’nyi pokrov Altaya [Vegetation
cover of the Altai]. Izdatel’stvo AN SSSR, Sibirskoe otdelenie,
Novosibirsk (in Russian).
Kunes, P., Pelankova, B., Chytry, M., Jankovska, V., Pokorny,
P. & Petr, L. (2008) Interpretation of the last-glacial vege-
tation of eastern-central Europe using modern analogues
from southern Siberia. Journal of Biogeography, 35, 2223–
2236.
Lozek, V. (1954) Novı mekkysi ceskoslovenskeho pleistocenu
[New molluscs of the Czechoslovak Pleistocene]. Vertigo
pseudosubstriata sp. n., Pupilla muscorum densegyrata ssp.
n. a Pupilla loessica sp. n. Anthropozoikum, 3, 327–342
(in Czech).
Lozek, V. (1964) Quartarmollusken der Tschechoslowakei.
CSAV, Prag.
Lozek, V. (1989) Postglacialnı souvrstvı v previsu na zapadnım
svahu Kobyly u Koneprus [Postglacial sedimentary fill of the
rock-shelter on the western slope of Kobyla Hill near
Koneprusy]. Cesky kras, 40, 57–72 (in Czech).
Lozek, V. (2001) Molluscan fauna from the loess series of
Bohemia and Moravia. Quaternary International, 76/77,
141–156.
van der Maarel, E. (1979) Transformation of cover-abundance
values in phytosociology and its effects on community
similarity. Vegetatio, 39, 97–114.
Martin, K. & Sommer, M. (2004) Relationships between land
snail assemblage patterns and soil properties in temperate-
humid forest ecosystems. Journal of Biogeography, 31, 531–
545.
Martınez-Meyer, E. & Peterson, A.T. (2006) Conservatism of
ecological niche characteristics in North American plant
species over the Pleistocene-to-Recent transition. Journal of
Biogeography, 33, 1779–1789.
Martınez-Meyer, E., Peterson, A.T. & Hargrove, W.W. (2004)
Ecological niches as stable distributional constraints on
M. Horsak et al.
1460 Journal of Biogeography 37, 1450–1462ª 2010 Blackwell Publishing Ltd

mammal species, with implications for Pleistocene extinc-
tions and climate change projections for biodiversity. Global
Ecology and Biogeography, 13, 305–314.
McCune, B. & Keon, D. (2002) Equations for potential annual
direct incident radiation and heat load. Journal of Vegetation
Science, 13, 603–606.
Meng, S. (2008) Neue Daten zur Verbreitung der Vertiginidae
(Gastropoda: Pulmonata) in Zentralasien. Mollusca, 26,
207–219.
Meng, S. (2009) Rezente zentralasiatische und pleistozane
mitteleuropaische Faunen mit Vallonia tenuilabris (A.
Braun, 1843). Mollusca, 27, 61–82.
Meng, S. & Hoffmann, M.H. (in press) Pupilla loessica Lozek
1954 (Gastropoda: Pulmonata: Pupillidae) – ‘‘a living fossil’’
in Central Asia? Quaternary Science Journal.
Meusel, H., Jager, E.J., Weinert, E. & Rauschert, S. (1965–
1992) Vergleichende Chorologie der zentraleuropaischen Flora
I–III. Gustav Fischer Verlag, Jena.
Mitchell-Jones, A.J., Amori, G., Bogdanowicz, W., Krystufek,
B., Reijnders, P.J.H., Spitzenberger, F., Stubbe, M., This-
sen, J.B.M., Vohralık, V. & Zima, J. (1999) The atlas of
European mammals. T. & A.D. Poyser Natural History,
London.
Moine, O. & Rousseau, D.D. (2002) Terrestrial molluscs and
temperature: a new quantitative transfer function. Comptes
Rendus Palevol, 1, 145–151.
Musil, R. (2003) The Middle and Upper Palaeolithic game
suite in Central and Southeastern Europe. Neanderthals and
modern humans in the European landscape during the last
glaciation (ed. by T.H. van Andel and S.W. Davies), pp. 167–
190. McDonald Institute for Archaeological Research,
Cambridge.
Nekhoroschev, V.P. (1966) Geologie des Altai. Akademie
Verlag, Berlin.
Pazonyi, P. (2004) Mammalian ecosystem dynamics in the
Carpathian Basin during the last 27,000 years. Palaeo-
geography, Palaeoclimatology, Palaeoecology, 212, 295–
314.
Pelankova, B. & Chytry, M. (2009) Surface pollen–vegetation
relationships in the forest-steppe, taiga and tundra land-
scapes of the Russian Altai Mountains. Review of Palaeobo-
tany and Palynology, 157, 253–265.
Pokryszko, B.M. (1990) The Vertiginidae of Poland (Gastro-
poda: Pulmonata: Pupilloidea) – a systematic monograph.
Annales Zoologici, 43/8, 134–256.
Pokryszko, B. & Horsak, M. (2007) Pupilloidea (Pupiliidae,
Vertiginidae, Valloniidae, Gastrocoptinae) of the Altay – a
travel in space and time. World Congress of Malacology,
Antwerp, Belgium, 15–20 July 2007, Abstracts (ed. by K.
Jordeans, N. Van Houtte, J. Van Goethem and T. Backeljau),
p. 170. Unitas Malacologica, Antwerp.
Pokryszko, B.M. (1993) Fen malacocenoses in Dovrefjell
(S. Norway). Fauna Norvegica, Series A, 14, 27–38.
Polikarpov, N.P., Chebakova, N.M. & Nazimova, D.I. (1986)
Klimat i gornye lesa Sibiri [Climate and mountain forests of
Siberia]. Nauka, Novosibirsk (in Russian).
R Development Core Team (2009) R: a language and envi-
ronment for statistical computing. R Foundation for Statis-
tical Computing, Vienna.
Ricankova, V. (2008) Ecology and paleobiogeography of Altai-
Sayan mammals. PhD Thesis, University of South Bohemia,
Ceske Budejovice.
Rivas-Martınez, S., Penas, A. & Dıaz, T.E. (2004) Bioclimatic
map of Europe. University of Leon, Leon.
Rousseau, D.D. (1990) Statistical analyses of loess molluscs for
paleoecological reconstructions. Quaternary Research, 7–8,
81–89.
Rousseau, D.D. (1991) Climatic transfer function from Qua-
ternary molluscs in European loess deposits. Quaternary
Research, 36, 195–209.
Rudner, Z.E. & Sumegi, P. (2001) Recurring taiga forest-steppe
habitats in the Carpathian Basin in the Upper Weichselian.
Quaternary International, 76–77, 177–189.
Rybnıckova, E. & Rybnıcek, K. (1991) The environment of the
Pavlovian – palaeoecological results from Bulhary, South
Moravia. Palaeovegetational development in Europe and
regions relevant to its palaeofloristic evolution (ed J. Kovar-
Eder), pp. 73–79. Museum of Natural History, Vienna.
Schileyko, A.A. (1984) Nazemnye mollyuski podotryada
Pupillina fauny SSSR (Gastropoda, Pulmonata, Geophila)
[Land molluscs of the suborder Pupillina of the USSR fauna
(Gastropoda, Pulmonata, Geophila)]. Fauna SSSR, Mollyuski,
3. Akademiya Nauk SSSR, Leningrad (in Russian).
Schofield, J.E., Edwards, K.J. & McMullen, J.A. (2007)
Modern pollen–vegetation relationships in subarctic
southern Greenland and the interpretation of fossil pollen
data from the Norse landnam. Journal of Biogeography, 34,
473–488.
Sjors, H. (1952) On the relation between vegetation and
electrolytes in north Swedish mire waters. Oikos, 2, 241–258.
Sommer, R.S. & Nadachowski, A. (2006) Glacial refugia of
mammals in Europe: evidence from fossil records. Mammal
Review, 36, 251–265.
Speight, M.C.D., Moorkens, E. & Falkner, G. (eds) (2003)
Proceedings of the Workshop on Conservation Biology of
European Vertigo Species (Dublin, April 2002). Heldia, 5,
1–183.
Sulikowska-Drozd, A. & Horsak, M. (2007) Woodland mollusc
faunas along environmental gradients in the East Carpa-
thians. Biologia, 62, 201–209.
Svenning, J.-C., Normand, S. & Kageyama, M. (2008) Glacial
refugia of temperate trees in Europe: insights from species
distribution modelling. Journal of Ecology, 96, 1117–1127.
Tarasov, P.E., Volkova, V.S., Webb, T., III, Guiot, J., Andreev,
A.A., Bezusko, L.G., Bezusko, T.V., Bykova, G.V., Doro-
feyuk, N.I., Kvavadze, E.V., Osipova, I.M., Panova, N.K. &
Sevastyanov, D.V. (2000) Last glacial maximum biomes
reconstructed from pollen and plant macrofossil data from
northern Eurasia. Journal of Biogeography, 27, 609–620.
Valovirta, I. (2003) The habitat and status of Vertigo angustior,
V. genesii, and V. geyeri in Finland and nearby Russian
Karelia. Heldia, 5, 85–94.
Snails of glacial Europe extant in southern Siberia
Journal of Biogeography 37, 1450–1462 1461ª 2010 Blackwell Publishing Ltd

Walter, H. (1974) Die Vegetation Osteuropas, Nord- und
Zentralasiens. Gustav Fischer Verlag, Stuttgart.
White, D., Preece, R.C., Shchetnikov, A.A., Parfitt, S.A. &
Dlussky, K.G. (2008) A Holocene molluscan succession
from floodplain sediments of the upper Lena River (Lake
Baikal region), Siberia. Quaternary Science Reviews, 27, 962–
987.
Willis, K.J. & van Andel, T.H. (2004) Trees or no trees?
The environments of central and eastern Europe during
the Last Glaciation. Quaternary Science Reviews, 23, 2369–
2387.
Willis, K.J., Rudner, E. & Sumegi, P. (2000) The full-glacial
forests of central and southeastern Europe. Quaternary
Research, 53, 203–213.
Yudin, B.S., Galkina, L.I. & Potapkina, A.F. (1979) Mlekopi-
tayushchie Altae-Sayanskoi gornoi strany [Mammals of
the Altai-Sayan mountain region]. Nauka, Novosibirsk
(in Russian).
SUPPORTING INFORMATION
Additional Supporting Information may be found in the
online version of this article:
Appendix S1 Detailed location and species data of the 18
study sites in the Altai Mountains, with occurrences of index
snail species of full-glacial central Europe.
As a service to our authors and readers, this journal
provides supporting information supplied by the authors.
Such materials are peer-reviewed and may be re-organized for
online delivery, but are not copy-edited or typeset. Technical
support issues arising from supporting information (other
than missing files) should be addressed to the authors.
BIOSKETCHES
Michal Horsak and Beata Pokryszko are malacologists,
Katerina Kintrova is a statistician, Barbora Pelankova
is a palynologist, and the other team members are botanists
and plant ecologists. In a project of Masaryk University, Brno,
Czech Republic, coordinated by Milan Chytry, this team
studied the diversity of natural ecosystems in the Altai and
Western Sayan Mountains of southern Siberia in 2003–2006.
This research focused on patterns of species richness, their
determinants, and on the interpretation of full-glacial envi-
ronments of eastern-central Europe using modern analogues
from these continental areas. Particular attention was paid to
the calibration of the fossil record using modern pollen
deposition and modern assemblages of land snails.
Author contributions: M. Horsak and M. Chytry conceived
the ideas and led the writing; M. Horsak collected and
identified all snail samples; B. Pokryszko helped with the
identification of some problematic snail species; K. Kintrova
analysed the data using a classification tree; and the other co-
authors collected and processed environmental and botanical
data and commented on the manuscript.
Editor: John Birks
M. Horsak et al.
1462 Journal of Biogeography 37, 1450–1462ª 2010 Blackwell Publishing Ltd

1
M. Horsák et al.: Habitats of relict terrestrial snails in southern Siberia: lessons for the reconstruction of palaeoenvironments of full-glacial Europe
Appendix S1 Detailed description of the 18 study sites in the Russian Altai Mountains with occurrences of index snail species of full-glacial loess sediments of central Europe. All recorded snail species and plant species with cover > 5% are given for each site. Plants are ordered by vegetation layers (tree, shrub, herb and moss layer), with layer separated by semicolons and species within layers ranked by decreasing cover. The taxonomy and nomenclature of vascular plants, bryophytes and lichens follow Cherepanov (1995), Ignatov & Afonina (1992) and Andreev et al. (1996), respectively. The first nine sites are shown in Figure 4; their numbers match the figure numbers.
1 – 50°07’06”N; 87°48’18”E; Kosh-Agach distr., Kurai: Northern Chuya Ridge, Bol’shoi Akturu river valley 14 km SSW of the village, valley bottom with open subalpine taiga, 1950 m a.s.l., 5 Aug 2005, field no. PH121. Snails: Columella intermedia Schileyko et Almuhambetova, 1984, Deroceras altaicum (Simroth, 1886), Euconulus fulvus (O.F. Müller, 1774), Vallonia tenuilabris (Al. Braun, 1842), Vertigo genesii (Gredler, 1856), Vitrina rugulosa Martens, 1874, Pupilla loessica Ložek, 1954, P. muscorum (Linné, 1758). Dominant plants: Larix sibirica; Betula rotundifolia; Hylocomium splendens, Aulacomnium palustre, Cladonia arbuscula, Dicranum spadiceum. 2 – 50°10’01”N; 87°48’37”E, Kosh-Agach distr., Kurai: SW margin of the Kurai steppe 12 km SW of the settlement, hemiboreal forest on a N facing slope, 1676 m, 4 Aug 2005, field no. PH120. Snails: Vertigo alpestris Alder, 1838, V. ronnebyensis (Westerlund, 1871), Euconulus fulvus (O.F. Müller, 1774), Pupilla loessica Ložek, 1954. Dominant plants: Larix sibirica; Caragana arborescens, Cotoneaster uniflorus; Bromopsis inermis, Iris ruthenica; Rhytidium rugosum. 3 – 50°09’04”N; 88°18’10”E, Kosh-Agach distr., Chagan-Uzun: Chuya river valley near the mouth of the Kuyuktanar valley 2 km NW of the settlement, wooded fen on a valley bottom, 1731 m, 3 Aug 2005, field no. PH117. Snails: Columella columella (G. v. Martens, 1830), Euconulus fulvus (O.F. Müller, 1774), Oxyloma sarsii (Esmark, 1886), Pupilla alpicola (Charpentier, 1837), Vallonia tenuilabris (Al. Braun, 1842), Vertigo genesii (Gredler, 1856), V. parcedentata (Al. Braun, 1847). Dominant plants: Picea obovata; Carex enervis, C. nigra, Equisetum pratense, E. scirpoides, Bryum pseudotriquetrum, Drepanocladus aduncus, Limprichtia cossoni, Sanionia uncinata. 4 – 50°09’16”N; 88°18’30”E; Kosh-Agach distr., Chagan-Uzun: Chuya river valley near the mouth of the Kuyuktanar valley 2 km NW of the settlement, base-rich fen, 1740 m, 2 Aug 2005, field no. PH113. Snails: Deroceras altaicum (Simroth, 1886), Oxyloma sarsii (Esmark, 1886), Vertigo genesii (Gredler, 1856). Dominant plants: Carex curaica, C. enervis, C. nigra, Equisetum variegatum, Juncus triglumis, Salix divaricata, Triglochin palustre; Hamatocaulis vernicosus, Aulacomnium palustre, Campylium stellatum. 5 – 50°06’02”N; 87°47’48”E; Kosh-Agach distr., Kurai: Northern Chuya Ridge, Bol’shoi Akturu river valley 17 km SSW of the village, wet alpine meadow on a gentle SSE
slope near the valley bottom, 2059 m, 5 Aug 2005, field no. PH123. Snails: Euconulus fulvus (O.F. Müller, 1774), Fruticicola schrenkii (Middendorff, 1851), Novisuccinea altaica (Martens, 1879), Perpolita hammonis (Ström, 1765), Pupilla loessica Ložek, 1954, Vallonia tenuilabris (Al. Braun, 1842), Vitrina pellucida (O.F. Müller, 1774). Dominant plants: Bupleurum longifolium, Carex pediformis s. l., Festuca sphagnicola, Pentaphylloides fruticosa, Trollius asiaticus. 6 – 50°28’32”N; 87°37’54”E; Ulagan distr., Ulagan: Ulagan plateau near Uzunkel’ lake, 28 km SW of the village, shrubby tundra, 1998 m, 18 Jul 2006, field no. MC202. Snails: Columella intermedia Schileyko et Almuhambetova, 1984, Euconulus fulvus (O.F. Müller, 1774), Novisuccinea altaica (Martens, 1879), Perpolita hammonis (Ström, 1765), Pupilla loessica Ložek, 1954, Vertigo genesii (Gredler, 1856), V. parcedentata (Al. Braun, 1847). Dominant plants: Betula rotundifolia; Aulacomnium palustre, Cladonia rangiferina, Polytrichum commune, Sphagnum warnstorfii, Tomenthypnum nitens. 7 – 50°09’52”N; 87°48’31”E; Kosh-Agach distr., Kurai: SW margin of the Kurai steppe 12 km SW of the settlement, steppe on a steep ESE-facing slope on limestone bedrock, 1693 m, 4 Aug 2005, field no. PH119. Snails: Pupilla alluvionica Meng et Hoffmann, 2008, P. altaica Meng et Hoffmann, 2008, P. loessica Ložek, 1954, P. muscorum (Linné, 1758), Vallonia costata (O.F. Müller, 1774). Dominant plants: Potentilla acaulis, Thymus serpyllum s. l. 8 – 50°18’27”N; 87°38’46”E; Ulagan distr., Aktash: Menka river valley 2 km E of the settlement, saline meadow grazed by horses, 1342 m, 6 Aug 2005, field no. PH126. Snails: Deroceras altaicum (Simroth, 1886), Pupilla loessica Ložek, 1954, Vallonia tenuilabris (Al. Braun, 1842), Vertigo genesii (Gredler, 1856). Dominant plants: Carex enervis, Cirsium esculentum, Deschampsia cespitosa, Glaux maritima, Halerpestes salsuginosa, Potentilla anserina. 9 – 50°59’06”N; 85°40’30”E; Ongudai distr., Zaisan’skaya Elan’: Seminskii Ridge, above the right bank of the Tuekta creek 2 km NW of the settlement, irregularly mown meadow, 1349 m, 12 Aug 2005, field no. MC150. Snails: Chilanodon bicallosa (L. Pfeiffer, 1853), Euconulus fulvus (O.F. Müller, 1774), Lindholmomneme altaica (Westerlund, 1896), Perpolita petronella (L. Pfeiffer, 1853), P. hammonis (Ström, 1765), Vertigo pseudosubstriata Ložek, 1954, V. substriata (Jeffreys, 1833).

2
Dominant plants: Deschampsia cespitosa, Cirsium heterophyllum, Geranium pseudosibiricum; Brachythecium salebrosum, Rhytidiadelphus triquetrus. 10 – 49°56’40”N; 88°00’47”E; Kosh-Agach distr., Chuya steppe, Bel’tir: Imele brook valley (right-side tributary of the Taldura river) 12 km WSW of the village, alpine tundra on a N facing slope, 2450 m, 1 Aug 2005, field no. PH106. Snails: Pupilla loessica Ložek, 1954. Dominant plants: Dryas oxyodonta, Betula rotundifolia, Salix alpina, S. arctica, S. glauca; Abietinella abietina, Aulacomnium palustre, A. turgidum, Dicranum sp., Tortula ruralis. 11 – 49°57’03”N; 88°01’06”E; Kosh-Agach distr., Chuya steppe, Bel’tir: slopes above the right bank of the Taldura river near Oimok settlement, subalpine forest on a steep NNW facing slope near the timberline, 2490 m, 1 Aug 2005, field no. PH110. Snails: Pupilla loessica Ložek, 1954. Dominant plants: Larix sibirica; Arctous alpina, Dryas oxyodonta, Hedysarum neglectum, Pyrola rotundifolia, Abietinella abietina, Aulacomnium turgidum, Rhytidium rugosum, Sanionia uncinata, Tortula ruralis. 12 – 50°08’56”N; 87°48’58”E; Kosh-Agach distr., Kurai: SW margin of the Kurai steppe 12 km SW of the settlement, shruby steppe, 1726 m, 4 Aug 2005, field no. PH118. Snails: Euconulus fulvus (O.F. Müller, 1774), Pupilla loessica Ložek, 1954, Vallonia kamtschatica Likharev, 1963, V. tenuilabris (Al. Braun, 1842), Vertigo ronnebyensis (Westerlund, 1871). Dominant plants: Caragana arborescens, Rosa pimpinellifolia, Spiraea media; Artemisia sericea, Carex pediformis, Festuca valesiaca, Iris ruthenica; Rhytidium rugosum. 13 – 50°14’57”N; 87°41’32”E; Ulagan distr., Chuya valley, Aktash: right bank of the Chuya river 9.5 km SE of the village, spruce forest with a well-developed shrub layer on a steep NNW facing slope with open boulder patches, 1472 m, 30 Jul 2005, field no. MC102. Snails: Euconulus fulvus (O.F. Müller, 1774), Pupilla alluvionica Meng et Hoffmann, 2008, P. altaica Meng et Hoffmann, 2008, P. loessica Ložek, 1954, Vallonia costata (O.F. Müller, 1774), V. ladacensis (Nevill, 1878), V. tenuilabris (Al. Braun, 1842), Vertigo alpestris Alder, 1838, V. ronnebyensis (Westerlund, 1871). Dominant plants: Picea obovata; Caragana arborescens; Carex macroura; Rhytidium rugosum, Abietinella abietina. 14 – 49°57’14”N; 88°02’17”E; Kosh-Agach distr., Chuya steppe, Bel’tir: above the right bank of the Taldura river 10 km W of the village, near Oimok settlement, larch forest on a steep N facing slope, 2199 m, 31 Jul 2005, field no. MC105.
Snails: Pupilla loessica Ložek, 1954, Vallonia ladacensis (Nevill, 1878). Dominant plants: Larix sibirica; Abietinella abietina, Rhytidium rugosum. 15 – 50°26’50”N; 87°36’28”E; Ulagan distr., Aktash: near the road to Ulagan 16 km N of the town, south of the Ulagan pass, species-rich grassland on a steep slope, 1976 m, 19 Jul 2006, field no. MC209. Snails: Novisuccinea altaica (Martens, 1879), Perpolita hammonis (Ström, 1765), Pupilla loessica Ložek, 1954, P. muscorum (Linné, 1758), Vallonia kamtschatica Likharev, 1963, V. ladacensis (Nevill, 1878), Vertigo sp. (undescribed species). Dominant plants: Artemisia laciniata, Aster alpinus, Astragalus austrosibiricus, Carex pediformis s. l., Iris ruthenica, Pentaphylloides fruticosa, Saussurea controversa. 16 – 50°18’27”N; 87°38’22”E; Ulagan distr., Aktash: Menka river valley 2 km E of the settlement, treeless calcareous fen in a river floodplain, 1330 m, 6 Aug 2005, field no. PH125. Snails: Oxyloma sarsii (Esmark, 1886), Vertigo genesii (Gredler, 1856). Dominant plants: Blysmus rufus, Carex enervis, C. nigra, Eleocharis quinqueflora, Primula nutans; Limprichtia cossoni, Bryum pseudotriquetrum, Campylium stellatum, Pseudocalliergon turgescens. 17 – 50°28’36”N; 87°37’48”E; Ulagan distr., Ulagan: plateau near Uzunkel’ lake 28 km SW of the village, shrubby tundra, 1994 m, 18 Jul 2006, field no. PH201. Snails: Columella intermedia Schileyko et Almuhambetova, 1984, Deroceras altaicum (Simroth, 1886), Euconulus fulvus (O.F. Müller, 1774), Novisuccinea altaica (Martens, 1879), Pupilla loessica Ložek, 1954, P. muscorum (Linné, 1758), Vertigo sp. (undescribed species), V. genesii (Gredler, 1856), V. parcedentata (Al. Braun, 1847), V. pseudosubstriata Ložek, 1954. Dominant plants: Betula humilis, B. rotundifolia, Pentaphylloides fruticosa; Lagotis integrifolia; Sanionia uncinata. 18 – 50°18’25”N; 87°38’55”E; Ulagan distr., Aktash: Menka river valley 2 km E of the settlement, base-rich wooded fen on a valley bottom, 1354 m, 6 Aug 2005, field no. PH127. Snails: Columella columella (G. v. Martens, 1830), Deroceras altaicum (Simroth, 1886), Pupilla muscorum (Linné, 1758), Punctum pygmaeum (Draparnaud, 1801), Vallonia tenuilabris (Al. Braun, 1842), Vertigo genesii (Gredler, 1856), V. pseudosubstriata Ložek, 1954. Dominant plants: Picea obovata; Salix divaricata, S. pyrolifolia; Comarum palustre, Deschampsia cespitosa, Equisetum scirpoides; Helodium blandowii, Plagiomnium affine, Tomenthypnum nitens.
REFERENCES Andreev, M., Kotlov, Yu. & Makarova, I. (1996) Checklist of lichens and lichenicolous fungi of the Russian Arctic.
Bryologist, 99, 137–169. Cherepanov, S.K. (1995) Sosudistye rasteniya Rossii i sopredel’nykh gosudarstv (Vascular plants of Russia and adjacent
countries). Mir i sem’ya-95, Sankt-Peterburg. Ignatov, M.S. & Afonina, O.M. (Eds) (1992) Check-list of mosses of the former USSR. Arctoa, 1, 1–58.
Copyright © 2022 FDOKUMEN