
Growth, Seasonality, and Lizard Life Histories: Age and Size at Maturity
13
Nordic Society Oikos Growth, Seasonality, and Lizard Life Histories: Age and Size at Maturity Author(s): Stephen C. Adolph and Warren P. Porter Source: Oikos, Vol. 77, No. 2 (Nov., 1996), pp. 267-278 Published by: Blackwell Publishing on behalf of Nordic Society Oikos Stable URL: http://www.jstor.org/stable/3546065 Accessed: 08/07/2009 12:52 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/action/showPublisher?publisherCode=black. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit organization founded in 1995 to build trusted digital archives for scholarship. We work with the scholarly community to preserve their work and the materials they rely upon, and to build a common research platform that promotes the discovery and use of these resources. For more information about JSTOR, please contact [email protected]. Blackwell Publishing and Nordic Society Oikos are collaborating with JSTOR to digitize, preserve and extend access to Oikos. http://www.jstor.org
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Growth, Seasonality, and Lizard Life Histories: Age and Size at Maturity

Nordic Society Oikos
Growth, Seasonality, and Lizard Life Histories: Age and Size at MaturityAuthor(s): Stephen C. Adolph and Warren P. PorterSource: Oikos, Vol. 77, No. 2 (Nov., 1996), pp. 267-278Published by: Blackwell Publishing on behalf of Nordic Society OikosStable URL: http://www.jstor.org/stable/3546065Accessed: 08/07/2009 12:52
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/action/showPublisher?publisherCode=black.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit organization founded in 1995 to build trusted digital archives for scholarship. We work with thescholarly community to preserve their work and the materials they rely upon, and to build a common research platform thatpromotes the discovery and use of these resources. For more information about JSTOR, please contact [email protected].
Blackwell Publishing and Nordic Society Oikos are collaborating with JSTOR to digitize, preserve and extendaccess to Oikos.
http://www.jstor.org

OIKOS 77: 267-278. Copenhagen 1996
Growth, seasonality, and lizard life histories: age and size at maturity
Stephen C. Adolph and Warren P. Porter
Adolph, S. C. and Porter, W. P. 1966. Growth, seasonality, and lizard life histories: age and size at maturity. - Oikos 77: 267-278.
Temperature influences the activity seasons, reproductive phenology, survival rates, and growth rates of lizards. We present a model of lizard growth that predicts phenotypic patterns of age and size at reproductive maturity in different thermal environments (i.e. different activity seasons). The model predicts a threshold in length of activity season: above this threshold (long season), lizards can mature one year earlier, but at a smaller size, compared to populations with activity seasons below the threshold. This environmentally imposed pattern reflects the proximate consequences of temperature, together with simple rules about the timing of maturation. A key prediction of the model is that age and size at maturity can vary non-linearly with the length of the activity season, and with the timing and duration of egg laying and hatching. We tested these predictions with published data from field studies of the phrynosomatid lizard Sceloporus undulatus, which is geographically widespread and occupies a range of thermal environments. We estimated activity seasons for each population by modeling the links between climates, microclimates and lizard body temperatures using heat-transfer principles. Female age at maturity showed the predicted threshold in length of activity season, whereas female size at maturity did not show the predicted threshold, but instead was negatively correlated with length of activity season. Two prairie populations were exceptions to this pattern: females matured in one year despite their short activity seasons, and consequently matured at an unusually small size. Prairie populations may have evolved differences in growth response and reproductive timing. The thermal environment appears to be an important correlate of life history variation among populations of Sceloporus undulatus.
S. C. Adolph and W. P. Porter, Dept of Zoology, Univ. of Wisconsin-Madison, 1117 West Johnson St., Madison, WI 53706, USA (present address of SCA: Dept of Biology, Harvey Mudd College, 301 E. Twelfth St., Claremont, CA 91711, USA (stephen_ adolph@hmc .edu)).
A central question in life history research is whether variation among populations or species represents envi- ronmental or genetic variation (Ballinger 1983, Stearns 1989). Historically, this question has received less atten- tion than the separate issue of whether life history variation is adaptive. However, an increasing number of studies have examined proximate environmental infl- uences on the expression of ectotherm life history traits (Ballinger 1977, 1979, Dunham 1978, Berven et al. 1979, Ferguson and Brockman 1980, Berven and Gill
1983, Porter and Tracy 1983, Jones and Ballinger 1987, Jones et al. 1987, Sinervo and Adolph 1989, 1994, Grant and Dunham 1990, Sinervo 1990, Grant and Porter 1992, Ferguson and Talent 1993, Niewiarowski and Roosenburg 1993, James and Whitford 1994, Curtin 1995). Many of these studies address the role of temperature, which frequently exerts strong proximate effects on ectotherm life histories (Levinton and Mona- han 1983, Conover and Present 1990, Sinervo and Doyle 1990, Adolph and Porter 1993, Bernardo 1994).
Accepted 10 January 1996
Copyright ? OIKOS 1996 ISSN 0030-1299 Printed in Ireland - all rights reserved
OIKOS 77:2 (1996) 267
The thermal environment restricts the growing sea- son for many organisms. Organisms at low elevations or low latitudes can grow and be active much of the year, whereas the growing season is curtailed at high elevations or high latitudes. Thermal effects on growth can greatly alter life histories. A common consequence is that organisms in cold environments exhibit delayed maturity, postponing reproduction until they are chronologically older (Tinkle et al. 1970). Although age at maturity is an important life history parameter (Cole 1954, Roff 1992, Stearns 1992), relatively little attention has been paid to the evolution of age and size at maturity among populations experiencing different thermal environments (Stearns and Koella 1986, Berri- gan and Charnov 1994). A first step is to quantify how these life history traits would be expressed in different environments in the absence of other differences (e.g., genetic composition, food resources) among popula- tions (Stearns 1989). This can serve as a physiologically based null hypothesis for life history variation, useful both for evaluating field data and for constructing models of life history evolution that incorporate proxi- mate effects (e.g., Beuchat and Ellner 1987).
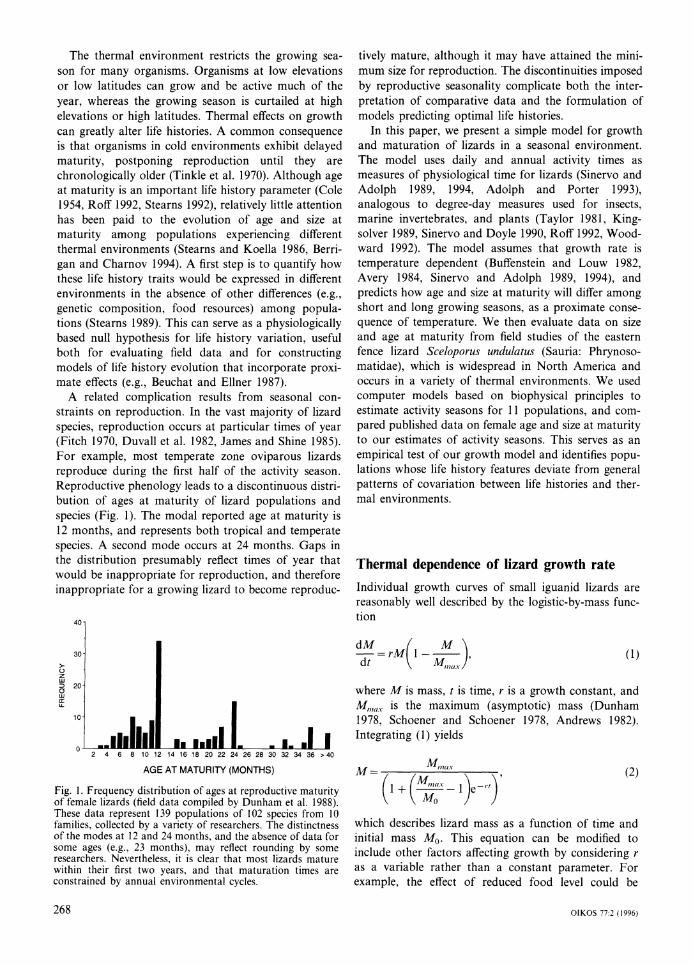
A related complication results from seasonal con- straints on reproduction. In the vast majority of lizard species, reproduction occurs at particular times of year (Fitch 1970, Duvall et al. 1982, James and Shine 1985). For example, most temperate zone oviparous lizards reproduce during the first half of the activity season. Reproductive phenology leads to a discontinuous distri- bution of ages at maturity of lizard populations and species (Fig. 1). The modal reported age at maturity is 12 months, and represents both tropical and temperate species. A second mode occurs at 24 months. Gaps in the distribution presumably reflect times of year that would be inappropriate for reproduction, and therefore inappropriate for a growing lizard to become reproduc-
40-
0 z w
LC
30-
20-
10- I E.
tively mature, although it may have attained the mini- mum size for reproduction. The discontinuities imposed by reproductive seasonality complicate both the inter- pretation of comparative data and the formulation of models predicting optimal life histories.
In this paper, we present a simple model for growth and maturation of lizards in a seasonal environment. The model uses daily and annual activity times as measures of physiological time for lizards (Sinervo and Adolph 1989, 1994, Adolph and Porter 1993), analogous to degree-day measures used for insects, marine invertebrates, and plants (Taylor 1981, King- solver 1989, Sinervo and Doyle 1990, Roff 1992, Wood- ward 1992). The model assumes that growth rate is temperature dependent (Buffenstein and Louw 1982, Avery 1984, Sinervo and Adolph 1989, 1994), and predicts how age and size at maturity will differ among short and long growing seasons, as a proximate conse- quence of temperature. We then evaluate data on size and age at maturity from field studies of the eastern fence lizard Sceloporus undulatus (Sauria: Phrynoso- matidae), which is widespread in North America and occurs in a variety of thermal environments. We used computer models based on biophysical principles to estimate activity seasons for 11 populations, and com- pared published data on female age and size at maturity to our estimates of activity seasons. This serves as an empirical test of our growth model and identifies popu- lations whose life history features deviate from general patterns of covariation between life histories and ther- mal environments.
Thermal dependence of lizard growth rate Individual growth curves of small iguanid lizards are reasonably well described by the logistic-by-mass func- tion
dM M ( dt r Ml I- )
I 0 I1Illllli . ... 1. m.. .11
2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 > 40
AGE AT MATURITY (MONTHS)
Fig. 1. Frequency distribution of ages at reproductive maturity of female lizards (field data compiled by Dunham et al. 1988). These data represent 139 populations of 102 species from 10 families, collected by a variety of researchers. The distinctness of the modes at 12 and 24 months, and the absence of data for some ages (e.g., 23 months), may reflect rounding by some researchers. Nevertheless, it is clear that most lizards mature within their first two years, and that maturation times are constrained by annual environmental cycles.
(1)
where M is mass, t is time, r is a growth constant, and M,..ax is the maximum (asymptotic) mass (Dunham 1978, Schoener and Schoener 1978, Andrews 1982). Integrating (1) yields
M= M_-X
+ -M .I.e-,)' + \ 0Mo
(2)
which describes lizard mass as a function of time and initial mass Mo. This equation can be modified to include other factors affecting growth by considering r as a variable rather than a constant parameter. For example, the effect of reduced food level could be
268 OIKOS 77:2 (1996)

modeled by choosing a smaller value of r. Similarly, thermal influences on growth rate can be modeled by making r a function of body temperature, Th. Because rt appears as a product in eq. 2, modifying r can be viewed as rescaling chronological time t in terms of physiological time.
Here, we are interested in the effect of the thermal environment on growth rate. For temperate zone lizards, temperature is likely to influence growth in two ways. First, lizards only grow during their activity season, which is jointly determined by the availability of suitable microclimates, temperature-dependent phys- iology and behavior, and sometimes additional consid- erations (e.g., food and moisture availability). During winter inactivity, lizards lose some body mass (e.g., Blair 1960). Thus, annual growth potential will be greater in environments with a longer activity season, all else being equal (Pianka 1970, Tinkle 1972a, Ballinger 1979, 1983, James and Shine 1988, Grant and Dunham 1990). Second, growth rates during different parts of the activity season are likely to vary due to seasonal variation in temperature, water, food availability, and energy expenditure (Andrews 1982).
Daily growth rates during the activity season are likely to be a function of the amount of time the thermal environment permits activity at high T,b, i.e., by "thermal opportunity" (Sinervo and Adolph 1994). This assumption is supported by several laboratory studies on lizards and snakes. Growth rates of juvenile Lacerta vivipara, Sceloporus occidentalis, Sceloporus graciosus, and Thamnophis elegans increase with daily activity time (i.e., access to high T,, via heat lamps; Avery 1984, Sinervo and Adolph 1989, 1994, Sinervo 1990, Peterson and Arnold, cited in Peterson et al. 1993). Thermally induced variation in growth rates probably results from several mechanisms acting in concert (Sinervo and Adolph 1989, Adolph and Porter 1993). Lizards with longer activity times spend more time at high Th, at which rates of physiological pro- cesses underlying growth (e.g., metabolism, digestion) are higher (Bennett and Dawson 1976, Skoczylas 1978, Huey 1982). Longer activity times also permit longer feeding periods (Avery 1984, Karasov and Anderson 1984, Waldschmidt et al. 1986). Thus, growth rates should often be positively correlated with thermal op- portunity, whether growth is limited by processing rates or by resource harvesting rates (Congdon 1989).
Following the above laboratory studies, we assume that daily growth rates increase linearly with daily activity time h according to
r = rmax ch, (3)
where r,,ax is maximum relative growth rate and c is a constant relating growth rate to activity time. Labora- tory experiments with juvenile Sceloporus occidentalis and S. graciosus indicate that a linear growth model is
MONTH
A 24 J , F,M,A,M ,J,J,A,S,0 ,N,D
$ 18- - 1 A 0 - 2d O 12 - 2y L. - \ / E 6- v
0 , . . ,0 ,0 -150 -100 -50 50 100 150
DAY OF ACTIVITY SEASON, i
B
0
0 w
-
C
0
0 IL
DAY OF ACTIVITY SEASON
Fig. 2. A) Seasonal variation in potential activity time of a diurnal lizard, as determined by the thermal environment and thermal physiology of the lizard. Within the ellipse thermal conditions permit activity; outside the ellipse lizards would be inactive. Here, we present activity season characteristic of many diurnal temperate zone lizards. Seasonal activity pat- terns in deserts and lowland tropical areas may be better described by other shapes, such as rectangles or ellipses with midday periods of inactivity (Adolph and Porter 1993). B) Activity ellipse for a warm environment. Dates of oviposition (dark area) and egg hatching (hatched area) are indicated. C) Activity ellipse for a cool environment, including dates of oviposition and hatching.
reasonably accurate for day lengths up to 12 h (Sinervo and Adolph 1989, 1994). In Appendix A we show how a curvilinear growth function can be incorporated into the model. A curvilinear growth response does not alter the general results.
Substituting (3) into (1) gives the following expres- sion for the dependence of daily growth rate on temper- ature, via the effect of temperature on activity time:
dM (M M
dt IIX
Ml(t-A^ ) (4)
Seasonal variation in activity times and growth rate
In the temperate zone, potential daily activity times for diurnal lizards are typically long in the summer and short in the spring and fall (Porter et al. 1973, Porter and Tracy 1983, Sinervo and Adolph 1994). This pat- tern of seasonal change in daily activity time can be conveniently approximated as an ellipse (Fig. 2), where the length of the activity season = 2y d, and the length
OIKOS 77:2 (1996) 269

of the maximum activity day = 2d h (Adolph and Porter 1993). For an elliptical activity season the poten- tial number of hours of activity per day is then given by
h = 2d1- ) (5)
where j represents the Julian day of the year, rescaled so that j = 0 at the middle of the activity season, where h is maximal. The area of the ellipse rT y d equals the cumulative potential hours of activity per year.
Diurnal lizards are primarily active when the thermal environment permits them to reach a T. within their preferred range (Cowles and Bogert 1944, Bartlett and Gates 1967, Norris 1967, Huey 1982). Values of y and d reflect these thermal constraints, which are deter- mined both by environmental characteristics (mainly air temperature and solar radiation) and by the lizard's thermal physiology and morphology (Porter et al. 1973, Porter and James 1979, Porter and Tracy 1983, Bakken 1989, 1992, Grant 1990, Adolph and Porter 1993). As discussed in Adolph and Porter (1993), shapes other than ellipses might be more appropriate for certain thermal environments, such as deserts or tropical low- lands. Here, we will restrict our analysis to elliptical activity seasons, but our model can be applied readily to any seasonal activity pattern.
Substituting expression (5) into eq. (4) yields an expression for growth rate as a function of time of year:
dt (6)
Eq. (6) is a modified version of eq. (1), with additional factors that rescale daily growth rate according to thermal opportunity. To determine how body mass changes we need to integrate eq. (6) over the appropri- ate time period. This yields
_f(j) 1- 1 1_
f(io) _M M x _ Mm ' (
where Mo = body mass at the beginning of the time period, J0 = initial date, and
f(j) = exp{rma /y2 x2 + y sin- (} (8)
For example, for the first (partial) growing season, Mo represents hatchling (or neonate) mass and Jo the date of hatching (or birth). Eq. (7), evaluated at j = y, gives the size reached by juveniles when their first growing season ends.
Environmental seasonality and age and size at maturity
Eq. (7) can be used to examine how age and size at maturity will vary as a proximate consequence of differ- ent thermal environments. This requires additional in- formation on hatchling or neonate size, minimum reproductive size, and reproductive phenology. Sizes at hatching and sexual maturity can be estimated from interspecific allometric relationships, if data are un- available (Appendix B).
We assume that a growing lizard will become repro- ductively mature if it has reached the minimum repro- ductive size and it is the appropriate time of year. This rule seems reasonable for lizards, although different rules may apply for other ectotherms (Bernardo 1993). Temperature probably plays the most important role in determining the time window for physiological decisions regarding maturation in temperate zone lizards, both as a proximate factor and a selective agent (Licht 1984, Lofts 1978, Marion 1970, 1982, Duvall et al. 1982). The latest possible date at which a maturation decision is feasible for females is likely to be determined by the total length of time required to mobilize resources for ovarian development, to produce eggs, for the eggs to incubate (or gestate), and for hatchlings (or neonates) to prepare for the onset of the inactive season. A longer activity season should permit an earlier date for the beginning of the maturation window, as reflected by the earlier reproduction frequently observed at lower alti- tudes and latitudes (Fitch 1970, Goldberg 1974). In addition, longer activity seasons often allow multiple clutches by a single female (e.g., McCoy and Hodden- bach 1966, Fitch 1970, Pianka 1970, Pianka and Parker 1972, Goldberg 1974, Parker and Pianka 1975, Ballinger 1983, Jones et al. 1987, James and Shine 1988), suggest- ing that window duration should increase with the length of the activity season.
A variety of mechanisms could be responsible for the timing and duration of the maturation window in natu- ral populations. For the purposes of this model, it is not necessary to identify the mechanism(s) operating in nature, but it is useful to specify a quantitative relation- ship between the maturation window and the length of the activity season that is consistent with observed patterns. Here, we assume that the maturation window spans a constant proportion of the activity season length (in d), with the initial date given by j = -0.75y and the final date by j= -0.4y. Thus, the maturation window will begin at an earlier calendar date given a longer activity season, and its length will increase linearly (with a slope of 0.35) with the length of the activity season.
Fig. 3 shows an example of a female lizard's growth to reproductive maturity in a cool environment with a short activity season (180 d; y = 90; 1,696 h/yr) and a warm environment with a long activity season (280 d; y = 140; 2,639 h/yr). Both seasons have maximum daily
OIKOS 77:2 (1996) 270

SHORT ACTIVITY SEASON
LL
.0
0C <
J F M AM J J A SN D 24- -
18
12-
6 hatching date
maturation window
20 -YR
1 5 YR 2
minimum / reproductive size
5
?n .. I . I...
-180 -120 -60 0 60 120
RELATIVE DATE (DAYS)
LONG ACTIVITY SEASON
AJ F M A M J
18"
12-
6-
J A S O N D
hatching date maturation
window
20- 1 - =' YR 2
15"
0) 10m /
~2 > - reproductive size
YR I
-180 -120 -60 0 60 120 180
RELATIVE DATE (DAYS)
Fig. 3. Examples of growth curves for a lizard with a short activity season (upper panels) and a lizard with a long activity season (lower panels), calculated using eq. 7. We used the same growth parameters for both examples: r,,,.. = 0.02, c = 0.0833, Mhatchling = 0.75 g, M,,,a- = 20 g. Shaded regions indicate matu- ration windows, during which a lizard must reach the minimum reproductive size (8.0 g, based on allometric relationships in Appendix B) in order to become reproductively mature that season. In this example, we assumed the maturation window included days affording 8-11 h activity. In the cool environ- ment (short season), the lizard does not reach the minimum size until after the maturation window, so it continues to grow until the end of the activity season. It matures in its second full growing season, at nearly two years of (chronological) age, and at a relatively large size. In contrast, the lizard with the long. growing season is able to reach minimum reproductive size at the end of its first maturation window, and therefore matures at nearly one year of age and a small size.
activity periods of 12 h/d (d= 6). Growth follows eq. (7), with parameter values given in the figure legend. We assumed the same growth parameters, hatchling size, and minimum size at maturity for both cases, so that growth differences were due solely to environmen- tal differences. In the short activity season, the lizard hatchling has 50 d to grow before the end of the activity season. During its second growing season, it does not reach minimum reproductive size during the maturation window, and continues to grow until the end of the season. By the time it reaches the maturity window in its third growing season, it is well above the minimum size for maturation. Its size at maturity is 12 g, and its chronological age at maturity is 20.5 months. In the long growing season, the hatchling has 120 d to grow during the remainder of the growing season. In its second year, the lizard reaches minimum reproductive size towards the end of the maturation window, and therefore matures at a small size (8 g) and a young age (9.4 months). After they mature, female lizards would probably not follow the illustrated growth trajectories because energy is diverted towards reproduction.
The long and short activity seasons in the above example (Fig. 3) represent points along a continuum of possible activity seasons. In Fig. 4 we show the average ages and sizes at maturity obtained for a range of activity seasons, using a particular set of growth parameters (see the figure caption for parameter val- ues). For this example, we assumed a uniform distribu- tion of hatching dates, beginning 100 d after the first date of the maturation window and ending 100 d after the last date of the maturation window. Thus, the duration of the hatching period was equal to the dura- tion of the maturation window, which increased with the length of the activity season. For each hatching date, we determined the age and size at which a lizard would reach reproductive maturity, based on the growth and maturation rules outlined above. We then computed the mean size and age at maturity (averaged over all hatching dates) for each activity season.
For short activity seasons, the model predicts an average age at maturity around 20 months, whereas for long activity seasons lizards mature around 8 months of age. For intermediate activity seasons, average age at maturity reflects a combination of some lizards who mature in their first full summer and some who mature in their second full summer, depending on hatching date. As season length increases, a larger proportion of hatching dates would allow maturation in under one year, so the average age at maturity declines. Size at maturity varies with length of activity season in a more complicated way. Initially, as season length increases, average size at maturity increases, because of the greater amount of time available for growth between hatching and the maturation window. Once the season is long enough for the earliest hatchlings to mature in their first year, however, average size at maturity de-
OIKOS 77:2 (1996)
U- LL LEL 0 w 2
24 , Ii . .... l
271

of a single species. The eastern fence lizard, Sceloporus undulatus (Sauria: Phrynosomatidae), is an excellent
20- . ....... candidate (Jones and Ballinger 1987). Sceloporus undu- latus is widespread in the United States, occurring
15- in montane, woodland, prairie and desert habitats (Smith 1946). Field studies by a number of researchers
10- have demonstrated extensive geographic variation in a number of life history traits. We used data from
1000 1500 2000 2500 3000 11 populations (Fig. 5) collected by several different researchers; these data are summarized and analyzed
20 in Tinkle and Dunham (1986) and Dunham et al. (1988).
15- / X Data on duration of activity season were not re- ported for most of these populations. Therefore, we
,10 estimated potential activity seasons for each population using computer models that calculate microclimates and animal Tb based on heat transfer principles (Porter
0 , , , et al. 1973, Porter and Tracy 1983, Porter 1989, see 1000 1500 2000 2500 3000 Adolph and Porter 1993 for further details). In brief,
ACTIVITY SEASON (HOURS) we used climate data for each site for each year of the
ample of model predictions of age (A) and size (B) field study (United States Weather Bureau) together y versus length of activity season (total potential with geographic data (latitude and altitude; obtained activity per year). For this example, we used the from United States Geological Survey maps in conjunc- parameter values: r = 0.025, M,, .....= 20 g, Mwith,, author=
-,,,,,, ,= 8.0 g, c = 0.0833, and d- 6 h. We assumed tion wth authors descriptions of the study sites) as distribution of hatching dates, beginning and ending input to the model. The model calculated solar radia- * the beginning and end of the maturation window. tion using program SOLRAD (McCullough and Porter represents the average value obtained for all hatch-
or a given activity season. 1971, see Adolph and Porter 1993) assuming clear sky 'or a given activity season. conditions. The microclimate simulations were coupled to a second computer model that calculated the equi-
ecause these lizards are maturing at a smaller librium Tb, attainable by S. undulatus, and determined Is, average size at maturity decreases with the times of day that a 10-g individual could reach its length of activity season over this intermedi- preferred body temperature range of 32?-37?C (Bogert of season lengths. Finally, for seasons suffi- 1949, Avery 1982, Crowley 1985, Grant and Porter
ciently long that all lizards can mature in under one year, average size at maturity should again increase as season length increases.
Thus, a key prediction of the model is that age and size at maturity can vary nonlinearly, and sometimes non-monotonically, with the length of the activity sea- son. The exact forms of these relationships depend on the values of growth parameters, duration of matura- tion window, and distribution of hatching dates. How- ever, the general form of these relationships is fairly robust for a variety of parameter values that are realis- tic for small-to-medium-sized lizards. Average age at maturity is predicted to show a series of plateaus (Fig. 4A), whereas average size at maturity is predicted to show a series of peaks (Fig. 4B), as a function of length of activity season.
Testing the model: analyzing field data from Sceloporus undulatus
Stearns (1980) suggested that hypotheses about life history variation are ideally tested among populations
1 1000 KM \I f
Fig. 5. Locations of life history field studies of the eastern fence lizard Sceloporus undulatus in the United States. Abbre- viations of states (and authors of studies) are: AZ (Arizona; Tinkle and Dunham 1986); CO (Colorado; Tinkle and Ballinger 1972); GA (Georgia; Crenshaw 1955); KS (Kansas; Ferguson et al. 1980); NM1 and NM2 (New Mexico; Vinegar 1975); NE (Nebraska; Ballinger et al. 1981); OH (Ohio; Tinkle and Ballinger 1972); SC (South Carolina; Tinkle and Ballinger 1972); TX (Texas; Tinkle and Ballinger 1972); UT (Utah; Tinkle 1972b).
OIKOS 77:2 (1996)
A
z o
F-
C:
w 1t- tU
B
a
LL Nr (0
Fig. 4. Ex< at maturit] hours of a following I 0.75 g, M,,, a uniform ( 100 d after Each point ing dates f
creases, b size. Thu increasing ate range
272

1992) in acceptable microhabitats. These daily potential activity periods were summed over the year to give an estimate of the total number of hours of potential activity. Estimated annual activity seasons ranged from 1707 h/yr for an Ohio population to 3012 h/yr for a population in Texas, nearly a two-fold range. In gen- eral, populations at low latitudes had longer estimated activity seasons (compare Figs 5 and 6).
Life history traits of most S. undulatus populations varied consistently with estimated length of activity season. Female age at maturity varied approximately as a step function of season length (Fig. 6A): lizards in environments with estimated activity seasons <2300 h/yr matured in their second full activity season, at ~20 months of age; lizards with activity seasons >2400 h/yr matured at 12 months. A New Mexico montane population, with an estimated season length of 2330 h/yr, had a mean age of maturity of 18 months, because some individuals matured in their first repro- ductive season and others in their second (Vinegar 1975). However, two populations from prairie habitats in Kansas and Nebraska were exceptions to this pat- tern: females matured during their first reproductive season, despite having short estimated activity seasons (% 2000 h/year; Fig. 6A). Female size at maturity de- creased with increasing season length (Fig. 6B), except for the two prairie populations, in which females ma- tured at a smaller than expected size because they matured a year early (Fig. 6B). The number of clutches laid per year increased with activity season length (Fig. 6C; r = 0.678); this supports the idea that the length of the maturation window should covary with the length of the activity season.
Discussion
The effect of season length on lizard life histories is well known to field researchers, but is rarely included in analyses of life history variation. Our model can be used to make quantitative predictions about how life- history phenotypes should vary as a proximate conse- quence of different thermal environments, given simple rules about reproductive phenology and the thermal dependence of growth. It shows that seasonal reproduc- tion can lead to discontinuous variation in age and size at maturity along latitudinal and altitudinal tempera- ture gradients in nature. In an earlier paper, we mod- eled the influence of activity season length on annual survival rate and reproductive output (Adolph and Porter 1993). Predictions of these models are summa- rized in Table 1. The combinations of life history traits predicted by "null" physiological models are the same as those predicted by evolutionary models: populations experiencing high adult mortality should have early maturity and/or high fecundity, relative to populations
I z 0
cn
- U'
cc
<
NU CO)
25-
20-
15-
10-
A
_-----------______.
0~~~~
KS
* NE
1500 2000 2500 3000
70- B
0
60- ?
KS *
NE 40 , , , ,
1500 2000 2500
4-
oc
CO LU I
D C)
3-
2-
1500
3000
* NM1 C
UT NM2
AZ TX
OH / SC GA
* 0 S NE CO
KS
2000 2500 3000
ANNUAL ACTIVITY SEASON (HOURS)
Fig. 6. Female size at maturity (snout-vent length; A), age at maturity (B), and average number of clutches laid per year (C) versus length of activity season for populations of the iguanid lizard Sceloporus undulatus in the United States. Activity sea- sons were calculated using microclimate simulations. Field data are from published studies summarized in Dunham et al. (1988); letters indicate location of study site (see Fig. 5). Prairie populations from Kansas (KS) and Nebraska (NE ) deviate from the general patterns. Omitting the KS and NE populations, size at maturity is negatively correlated with annual activity hours (r= -0.83; P < 0.01).
experiencing low mortality (reviews in Roff 1992 and Stearns 1992). This finding is consistent with Shine and Charnov's (1992) analysis of published data on sur- vival, growth and maturation in lizards and snakes, which showed that adult survival rate was positively correlated with age at maturity. Similarly, Ballinger and Congdon (1981) commented that "Lizards occurring in
OIKOS 77:2 (1996)
1 , ,.
*----__-_ --_
3
273

environments with long growing seasons typically pro- duce multiple clutches, have high reproductive efforts, and have short life expectancies. Conversely, lizards occurring in environments with short growing seasons are longer-lived with low reproductive efforts". Ob- served patterns of variation in life history traits among wild lizard populations could reflect proximate environ- mental influences, adaptive genetic differentiation, or a combination of both.
Our model also highlights the functional relation- ships among sizes, growth rates, seasonality, and the duration of different components of reproduction (e.g., egg laying, hatching). Changing any one of these fac- tors will usually affect the population mean size and/or age at maturity. This effect will be particularly strong when activity seasons are near the threshold length: a small change in growth rate, hatchling mass, or incuba- tion time could have a large effect on age and size at maturity. This suggests that selection on age or size at maturity could act indirectly on these functionally con- nected traits. For example, selection for early maturity could result in the evolution of shorter incubation time, or higher growth rate. Thus, seasonality can result in "leveraged" effects of growth-related traits on major fitness components. Similarly, a population experienc- ing directional climate change could experience a dis- proportionate change in one or more life history traits, which could have both demographic and evolutionary consequences.
Life history variation in Sceloporus undulatus
A number of researchers have examined intraspecific variation in life history traits of S. undulatus (Tinkle and Ballinger 1972, Ferguson et al. 1980, Ballinger et al. 1981, Tinkle and Dunham 1986, Dunham et al. 1988, Gillis and Ballinger 1992, Niewiarowski 1994). Neither habitat type nor phylogenetic relatedness pro- vides a satisfactory explanation for the observed pat- tern of geographic variation (Tinkle and Dunham 1986, Gillis and Ballinger 1992). Gillis and Ballinger (1992) suggested that climatic and biophysical considerations might improve our understanding of life history pat- terns in this species.
Table 1. Lizard life history patterns predicted as a proximate consequence of different thermal environments (this study and Adolph and Porter 1993).
Life history Long activity Short activity characteristic season season
Annual survival low high rate
Annual reproductive high low output
Age at maturity early delayed Size at maturity small large
Our analysis indicates that the thermal environment, through its effects on activity time, growth energetics, and reproductive seasonality, may explain some of the variation in age at maturity in S. undulatus. In addition, variation in size at maturity may be strongly related to length of activity season; however, our model predicted a different form for this relationship. Previously, we found that annual adult survival rate was negatively correlated with length of activity season (Adolph and Porter 1993). We also showed that size-corrected an- nual egg mass production (a measure of reproductive investment) was positively correlated with length of activity season, and that body size and length of activ- ity season together explained 87% of the variation among populations in annual egg mass.
Together, these results are consistent with the hy- pothesis that the proximate effects of climate govern much of the life history variation among populations of S. undulatus. Populations with longer estimated activity seasons matured earlier and at a smaller size, produced more clutches and more eggs per year, and had lower annual survival rates, compared to populations living in cool environments. Most of these trends can be inter- preted as simple proximate consequences of tempera- ture, via its influence on daily and seasonal activity time, growth energetics and reproductive phenology. However, female size at maturity varied with annual activity time in a manner different from that predicted by our physiological model, suggesting that the as- sumptions of our model are not met by these popula- tions. For example, populations could differ in resource availability, which can also affect growth rates in nature (Grant and Dunham 1990).
Alternatively, these populations could differ geneti- cally in their growth or size parameters, such as r,n,
initial (hatchling) size or maximum size. Indeed, both egg size and adult size vary substantially among these populations (Dunham et al. 1988). Aspects of size are also strongly correlated with one another; for example, female SVL at maturity is positively correlated with both mean egg mass (r = 0.72) and mean female SVL (r = 0.94) (omitting the two outlier populations in Kan- sas and Nebraska). Hence, like female SVL at maturity, mean egg mass and mean female SVL are also nega- tively correlated with length of activity season. Hatch- ling size is also likely to covary with egg and adult size; hatchling mass is positively correlated with egg mass among individuals of S. undulatus (Ferguson and Brockman 1980) and of the related species Sceloporus occidentalis and S. graciosus (Sinervo and Adolph 1989, Sinervo and Huey 1990, S. C. Adolph and B. Sinervo unpubl.).
Several common-garden studies have revealed differ- ences in growth rates among populations of S. undula- tus that may represent genetic differences. Ferguson and Talent (1993) raised lizards from eggs to maturity in the laboratory, and found that Utah and Oklahoma
OIKOS 77:2 (1996) 274

populations differed in growth rate, age and size at maturity, clutch size, and egg size. Lizards from both populations matured in just 4-5 months in the labora- tory, compared to 1-2 yr in the field. This clearly demonstrates that age at maturity is phenotypically plastic in this species (see also Porter and Tracy 1983). Ferguson and Talent (1993) also found that size at maturity differed between the laboratory and the field: in the lab, Utah lizards were smaller at maturity, but Oklahoma lizards were larger at maturity, compared to lizards in their respective field populations. Ferguson and Brockman (1980) found interpopulational differ- ences in growth rates of laboratory-reared juveniles of three populations of S. undulatus. Niewiarowski and Roosenburg (1993) performed reciprocal field trans- plants of juvenile S. undulatus and found differences in growth response between lizards from Nebraska and New Jersey populations: lizards from both populations grew at similar rates when grown in New Jersey, but lizards from Nebraska grew faster in Nebraska. These results may reflect genetic differences in plasticity of growth rate in response to environments differing in thermal opportunity (see also Sinervo and Adolph 1994). Growth rates of juvenile S. occidentalis in the laboratory also indicate possible genetic variation within and between populations (Sinervo and Adolph 1989, 1994, Sinervo 1990). Thus, a number of studies on S. undulatus and other Sceloporus species indicate that genetically based differences in egg size, adult size, and growth rate may be common.
Together, size and temperature appear to explain much of the phenotypic life history variation among populations of S. undulatus. Our analysis suggests that proximate effects of the thermal environment could be responsible for some of the variation in size at maturity among populations. However, variation in egg size, hatchling size, and maximum body size are not readily explained by proximate effects of temperature. Because these measures of size are highly correlated among populations of S. undulatus, (see above) they are proba- bly not free to evolve independently, but instead would evolve in a correlated manner. The tendency for eggs, hatchlings and adults to be smaller where activity sea- sons are longer could reflect adaptation to different climates. For example, body size influences the ability of ectotherms to thermoregulate: all else being equal, large lizards can maintain a higher equilibrium Th than small lizards, because of their larger boundary layers (Porter and Gates 1969). However, this difference would be small (<2?C) over the size range encom- passed by populations of S. undulatus (Stevenson 1985), and therefore would probably have little effect on activ- ity times. Another possibility is that longer seasons somehow favor smaller lizards via the life history corre- lates of body size. These possibilities could be addressed through field measurements of natural selection (e.g., Ferguson and Fox 1984, Sinervo et al. 1993), or by
developing an optimization model that includes the proximate effects of temperature (e.g., Beuchat and Ellner 1987) along with size- and season-specific mor- tality schedules. Still another possibility is the interac- tion between thermal opportunity and food availability (Grant and Dunham 1990, Dunham 1993). For exam- ple, intermittent or chronic food shortages, which might be more common for populations with long activity seasons, would reduce growth potential, and possibly cause selection for slower growth rates and/or smaller body size.
Why are the prairie populations exceptions to the general patterns? Life history theory predicts that early maturation is likely to evolve when adult mortality is high (Roff 1992, Stearns 1992). However, the Kansas and Nebraska populations have among the highest adult annual survival rates of all S. undulatus popula- tions (Jones and Ballinger 1987, Dunham et al. 1988). Thus, neither an evolved response to age-specific mor- tality patterns nor the proximate effects of temperature offer a simple explanation for early maturation of fe- males in prairie populations. This suggests that selec- tion on size per se, rather than reproductive timing, could be involved.
Ecological and evolutionary implications
Simple physiologically based models predict that the proximate effects of temperature on lizard life history traits can be automatically compensatory: thermal envi- ronments resulting in reduced survival rates also permit higher annual reproductive output and earlier maturity (Table 1; Adolph and Porter 1993). This phenomenon has implications for both population ecology and the evolution of life history traits. On an ecological time scale, lizard population dynamics could be partially buffered against temporal (or geographic) variation in climate, as population mean life history phenotypes respond to the proximate effects of temperature (but see Dunham 1993). The fact that many species such as Sceloporus undulatus can exist in a variety of thermal environments indicates one possible advantage of hav- ing a phenotypically plastic life history. Indeed, wide- spread species that are not genetically adapted to local thermal conditions may be better able to cope with temporal climatic changes; lack of local adaptation may indicate that phenotypic flexibility is sufficient.
Similarly, from an evolutionary perspective, one might expect natural selection to have favored patterns of phenotypic response of the appropriate shape to maximize fitness across a range of environmental condi- tions (Stearns and Koella 1986, Sinervo and Adolph 1994). The physiological mechanism outlined in our models would facilitate the evolutionary adjustment of norms of reaction of life history traits, because the proximate effects of temperature, acting via activity
OIKOS 77:2 (1996) 275

time, are already in the appropriate direction. Thus, adaptive evolution of the norms of reaction of lizard life history traits could involve fine-tuning of an already existing response, rather than wholesale adjustments.
Acknowledgements - We thank S. Beaupre, J. Jaeger, G. C. Mayer, R. T. M'Closkey, D. K. Padilla, P. S. Reynolds, R. Shine, B. Sinervo, and B. Wilson for helpful discussion or comments on the manuscript, and B. Feeny for help with the United States map. This research was supported by the Office of Health and Environmental Research, US Dept of Energy, through contract DE-FG02-88ER60633 to WPP, and a Guyer Fellowship from the Dept of Zoology, UW, to SCA. Manuscript preparation was supported by a faculty research grant from Harvey Mudd College to SCA.
References Adolph, S. C. and Porter, W. P. 1993. Temperature, activity,
and lizard life histories. - Am. Nat. 142: 273-295. Andrews, R. M. 1982. Patterns of growth in reptiles. - In:
Gans, C. and Pough, F. H. (eds), Biology of the Reptilia, Vol. 13, Physiology D. Physiological ecology. Academic Press, New York, pp. 273-320.
Avery, R. A. 1982. Field studies of body temperatures and thermoregulation. - In: Gans, C. and Pough, F. H. (eds), Biology of the Reptilia, Vol. 12, Physiology C. Physiologi- cal ecology. Academic Press, New York, pp. 93-166.
- 1984. Physiological aspects of lizard growth: the role of thermoregulation. - Symp. Zool. Soc. Lond. 52: 407-424.
Bakken, G. S. 1989. Arboreal perch properties and the opera- tive temperature experienced by small animals. - Ecology 70: 922-930.
- 1992. Measurement and application of operative and stan- dard operative temperatures in ecology. - Am. Zool. 32: 194-216.
Ballinger, R. E. 1977. Reproductive strategies: food availabil- ity as a source of proximal variation in a lizard. - Ecology 58: 628-635.
- 1979. Intraspecific variation in demography and life history of the lizard, Sceloporus jarrovi, along an altitudinal gradi- ent in southeastern Arizona. - Ecology 60: 901-909.
- 1983. Life-history variations. - In: Huey, R. B., Pianka, E. R. and Schoener, T. W. (eds), Lizard ecology: studies of a model organism. Harvard Univ. Press, Cambridge, MA, pp. 241-260.
- and Congdon, J. D. 1981. Population ecology and life history strategy of a montane lizard (Sceloporus scalaris) in Southeastern Arizona. - J. Nat. Hist. 15: 213-222.
- , Droge, D. L. and Jones, S. M. 1981. Reproduction in a Nebraska sandhills population of the northern prairie lizard Sceloporus undulatus garmani. - Am. Midl. Nat. 106: 157-164.
Bartlett, P. N. and Gates, D. M. 1967. The energy budget of a lizard on a tree trunk. - Ecology 48: 315-322.
Bennett, A. F. and Dawson, W. R. 1976. Metabolism. - In: Gans, C. and Dawson, W. R. (eds), Biology of the Rep- tilia, Vol. 5, Physiology A. Academic Press, New York, pp. 127-223.
Bernardo, J. 1993. Determinants of maturation in animals. - Trends Ecol. Evol. 8: 166-173.
- 1994. Experimental analysis of allocation in two divergent, natural salamander populations. - Am. Nat. 143: 14- 38.
Berrigan, D., and Charnov, E. L. 1994. Reaction norms for age and size at maturity in response to temperature: a puzzle for life historians. - Oikos 70: 474-478.
Berven, K. A. and Gill, D. E. 1983. Interpreting geographic variation in life-history traits. - Am. Zool. 23: 85-97.
- , Gill, D. E., and Smith-Gill, S. J. 1979. Countergradient
selection in the green frog, Rana clamitans. - Evolution 33: 609-623.
Beuchat, C. A. and Ellner, S. 1987. A quantitative test of life history theory: thermoregulation by a viviparous lizard. - Ecol. Monogr. 57: 45-60.
Blair, W. F. 1960. The rusty lizard. A population study. - Univ. of Texas Press, Austin, TX.
Bogert, C. M. 1949. Thermoregulation and eccritic body tem- peratures in Mexican lizards of the genus Sceloporus. - An. Inst. Biol. Univ. Nac. Auton. M6x. 20: 415-426.
Buffenstein, R. and Louw, G. 1982. Temperature effects on bioenergetics of growth, assimilation efficiency and thyroid activity in juvenile varanid lizards. - J. Therm. Biol. 7: 197-200.
Cole, L. C. 1954. The population consequences of life-history phenomena. - Q. Rev. Biol. 29: 103-137.
Congdon, J. D. 1989. Proximate and evolutionary constraints on energy relations of reptiles. - Physiol. Zool. 62: 356- 373.
Conover, D. 0. and Present, T. M. C. 1990. Countergradient variation in growth rate: compensation for length of the growing season among Atlantic silversides from different latitudes. - Oecologia 83: 316-324.
Cowles, R. B. and Bogert, C. M. 1944. A preliminary study of the thermal requirements of desert reptiles. - Bull. Am. Mus. Nat. Hist. 83: 261-296.
Crenshaw, J. W., Jr. 1955. The life history of the southern spiny lizard, Sceloporus undulatus undulatus Latreille. - Am. Midi. Nat. 54: 257-298.
Crowley, S. R. 1985. Thermal sensitivity of sprint-running in the lizard Sceloporus undulatus: support for a conservative view of thermal physiology. - Oecologia 66: 219-225.
Curtin, C. G. 1995. Latitudinal compensation in ornate box turtles: implications for species response to shifting land use and climate. - Dissertation. Univ. of Wisconsin, Madison, WI.
Dunham, A. E. 1978. Food availability as a proximate factor influencing individual growth rates in the iguanid lizard Sceloporus merriami. - Ecology 59: 770-778.
- 1993. Population responses to environmental change: oper- ative environments, physiologically structured models, and population dynamics. - In: Kareiva, P. M., Kingsolver, J. G. and Huey, R. B. (eds), Biotic interactions and global change. Sinauer, Sunderland, MA, pp. 95-119.
- , Miles, D. B. and Reznick, D. N. 1988. Life history patterns in squamate reptiles. - In: Gans, C. and Huey, R. B. (eds), Biology of the Reptilia, Vol. 16, Ecology B, Defense and life history. Liss, New York, pp. 441-522.
Duvall, D., Guillette, L. J. Jr. and Jones, R. E. 1982. Environ- mental control of reptilian reproductive cycles. - In: Gans, C. and Pough, F. H. (eds), Biology of the Reptilia, Vol. 13, Physiology D, Physiological ecology. Academic Press, New York, pp. 201-231.
Ferguson, G. W. and Brockman, T. 1980. Geographic differ- ences of growth rate of Sceloporus lizards (Sauria: Iguanidae). - Copeia 1980: 259-264.
- and Fox, S. F. 1984. Annual variation of survival advan- tage of large juvenile side-blotched lizards Uta stansburi- ana: its causes and evolutionary significance. - Evolution 38:342-349.
- and Talent, L. G. 1993. Life-history traits of the lizard Sceloporus undulatus from two populations raised in a common laboratory environment. - Oecologia 93: 88-94.
- , Bohlen, C. H. and Woolley, H. P. 1980. Sceloporus undulatus: comparative life history and regulation of a Kansas population. - Ecology 61: 313-322.
Fitch, H. S. 1970. Reproductive cycles in lizards and snakes. - Univ. Kan. Mus. Nat. Hist. Misc. Publ. 52: 1-247.
Gillis, R. and Ballinger, R. E. 1992. Reproductive ecology of red-chinned lizards (Sceloporus undulatus erythrocheilus) in Southcentral Colorado: comparisons with other popula- tions of a wide-ranging species. - Oecologia 89: 236- 243.
276 OIKOS 77:2 (1996)

Goldberg, S. R. 1974. Reproduction in mountain and lowland populations of the lizard Sceloporus occidentalis. - Copeia 1974: 176-182.
Grant, B. W. 1990. Trade-offs in activity time and physiologi- cal performance for thermoregulating desert lizards, Sceloporus merriami. - Ecology 71: 2323-2333.
- and Dunham, A. E. 1990. Elevational covariation in envi- ronmental constraints and life histories of the desert lizard Sceloporus merriami. - Ecology 71: 1765-1776.
- and Porter, W. P. 1992. Modeling global macroclimatic constraints on ectotherm energy budgets. - Am. Zool. 32: 154-178.
Huey, R. B. 1982. Temperature, physiology, and the ecology of reptiles. - In: Gans, C. and Pough, F. H. (eds), Biology of the Reptilia, Vol. 12, Physiology C, Physiological ecol- ogy. Academic Press, New York, pp. 25-91.
James, C. and Shine, R. 1985. The seasonal timing of repro- duction: a tropical-temperate comparison in Australian lizards. - Oecologia 67: 464-474.
- and Shine, R. 1988. Life-history strategies of Australian lizards: a comparison between the tropics and the temper- ate zone. - Oecologia 75: 307-316.
- and Whitford, W. G. 1994. An experimental study of phenotypic plasticity in the clutch size of a lizard. - Oikos 70: 49-56.
Jones, S. M. and Ballinger, R. E. 1987. Comparative life histories of Holbrookia maculata and Sceloporus undulatus in western Nebraska. - Ecology 68: 1828-1838.
-, Ballinger, R. E. and Porter, W. P. 1987. Physiological and environmental sources of variation in reproduction: prairie lizards in a food rich environment. - Oikos 48: 325-335.
-, Waldschmidt, S. R. and Potvin, M. A. 1987. An experi- mental manipulation of food and water: growth and time- space utilization of hatchling lizards (Sceloporus undulatus). - Oecologia 73: 53-59.
Karasov, W. H. and Anderson, R. A. 1984. Interhabitat differences in energy acquisition and expenditure in a lizard. - Ecology 65: 235-247.
Kingsolver, J. G. 1989. Weather and the population dynamics of insects: integrating physiology and population ecology. - Physiol. Zool. 62: 314-334.
Levinton, J. S. and Monahan, R. K. 1983. The latitudinal compensation hypothesis: growth data and a model of latitudinal growth differentiation based upon energy bud- gets. II. Intraspecific comparisons between subspecies of Ophryotrocha puerilis (Polychaeta: Dorvillidae). - Biol. Bull. 165: 699-707.
Licht, P. 1984. Reptiles. - In: Lamming, E. (ed.), Marshall's physiology of reproduction. Academic Press, New York, pp. 206-282.
Lofts, B. 1978. Reptilian reproductive cycles and environmen- tal regulators. - In: Assenmacher, I. and Farner, D. S. (eds), Environmental endocrinology. Springer, New York, pp. 37-43.
Marion, K. R. 1970. Temperature as the reproductive cue for the female fence lizard Sceloporus undulatus. - Copeia 1970: 562-564.
- 1982. Reproductive cues for gonadal development in tem- perate reptiles: temperature and photoperiod effects on the testicular cycle of the lizard Sceloporus undulatus. - Her- petologica 38: 26-39.
McCoy, C. J. and Hoddenbach, G. A. 1966. Geographic variation in ovarian cycles and clutch size in Cnemidopho- rus tigris (Teiidae). - Science 154: 1671-1672.
McCullough, E. C. and Porter, W. P. 1971. Computing clear day solar radiation spectra for the terrestrial ecological environment. - Ecology 52: 1008-1015.
Niewiarowski, P. H. 1994. Understanding geographic life- history variation in lizards. - In: Vitt, L. J. and Pianka, E. R. (eds), Lizard ecology: historical and experimental perspectives. Princeton Univ. Press, Princeton, NJ, pp. 31- 49.
- and Roosenburg, W. M. 1993. Reciprocal transplant re-
veals sources of variation in growth rates of the lizard Sceloporus undulatus. - Ecology 74: 1992-2002.
Norris, K. S. 1967. Color adaptation in desert reptiles and its thermal relationships. - In: Milstead, W. W. (ed.), Lizard ecology: a symposium. Univ. of Missouri Press, Columbia, MO, pp. 162-229.
Parker, W. S. and Pianka, E. R. 1975. Comparative ecology of populations of the lizard Uta stansburiana. - Copeia 1975: 615-632.
Peterson, C. R., Gibson, A. R. and Dorcas, M. E. 1993. Snake thermal ecology: the causes and consequences of body- temperature variation. - In: Siegel, R. A. and Collins, J. T. (eds), Snakes: ecology and behavior. McGraw-Hill, New York, pp. 241-314.
Pianka, E. R. 1970. Comparative autecology of the lizard Cnemidophorus tigris in different parts of its geographic range. - Ecology 51: 703-720.
- and Parker, W. S. 1972. Ecology of the iguanid lizard Callisaurus draconoides. - Copeia 1972: 493-508.
Porter, W. P. 1989. New animal models and experiments for calculating growth potential at different elevations. - Phys- iol. Zool. 62: 286-313.
- and Gates, D. M. 1969. Thermodynamic equilibria of animals with environment. - Ecol. Monogr. 39: 227-244.
- and James, F. C. 1979. Behavioral implications of mechanis- tic ecology II: the African rainbow lizard, Agama agama. - Copeia 1979: 594-619.
- and Tracy, C. R. 1983. Biophysical analyses of energetics, time-space utilization, and distributional limits. - In: Huey, R. B., Pianka, E. R. and Schoener, T. W. (eds), Lizard ecology: studies of a model organism. Harvard Univ. Press, Cambridge, MA, pp. 55-83.
-, Mitchell, J. W., Beckman, W. A. and DeWitt, C. B. 1973. Behavioral implications of mechanistic ecology. - Oecolo- gia 13: 1-54.
Roff, D. A. 1992. The evolution of life histories: theory and analysis. - Chapman and Hall, New York.
Schoener, T. W. and Schoener, A. 1978. Estimating and interpreting body-size growth in some Anolis lizards. - Copeia 1978: 390-405.
Shine, R. and Charnov, E. L. 1992. Patterns of survival, growth and maturation in snakes and lizards. - Am. Nat. 139: 1257-1269.
Sinervo, B. 1990. Evolution of thermal physiology and growth rate between populations of the western fence lizard (Sceloporus occidentalis). - Oecologia 83: 228-237.
- and Adolph, S. C. 1989. Thermal sensitivity of growth rate in hatchling Sceloporus lizards: environmental, behavioral and genetic aspects. - Oecologia 78: 411-419.
- and Doyle, R. W. 1990. Life-history analysis in "physiolog- ical" compared with "sidereal" time: an example with an amphipod (Gammarus lawrencianus) in a varying environ- ment. - Mar. Biol. 107: 129-139.
- and Huey, R. B. 1990. Allometric engineering: an experi- mental test of the causes of interpopulational differences in performance. - Science 248: 1106-1109.
- and Adolph, S. C. 1994. Growth plasticity and thermal opportunity in Sceloporus lizards. - Ecology 75: 776-790.
-, Doughty, P., Huey, R. B. and Zamudio, K. 1993. Allo- metric engineering: a causal analysis of natural selection on offspring size. - Science 258: 1927-1930.
Skoczylas, R. 1978. Physiology of the digestive tract. - In: Gans, C. and Gans, K. A. (eds), Biology of the Reptilia, Vol. 8, Physiology B. Academic Press, New York, pp. 589-717.
Smith, H. M. 1946. Handbook of lizards. - Comstock, Ithaca, NY.
Stearns, S. C. 1980. A new view of life history evolution. - Oikos 35: 266-281.
- 1989. The evolutionary significance of phenotypic plastic- ity. - BioScience 39: 436-445.
- 1992. The evolution of life histories. - Oxford Univ. Press, Oxford.
OIKOS 77:2 (1996) 277

- and Koella, J. C. 1986. The evolution of phenotypic plas- ticity in life-history traits: predictions of reaction norms for age and size at maturity. - Evolution 40: 893-913.
Stevenson, R. D. 1985. The relative importance of behavioral and physiological adjustments controlling body tempera- ture in terrestrial ectotherms. - Am. Nat. 126: 362-386.
Taylor, F. 1981. Ecology and evolution of physiological time in insects. - Am. Nat. 117: 1-23.
Tinkle, D. W. 1972a. The role of environment in the evolution of life history differences within and between lizard species. - In: Allen, R. T. and James, F. C. (eds), A symposium on ecosystematics. Univ. Ark. Mus. Occas. Pap. No. 4, pp. 77-100.
- 1972b. The dynamics of a Utah population of Sceloporus undulatus. - Herpetologica 28: 351-359.
and Ballinger, R. E. 1972. Sceloporus undulatus: a study of the intraspecific comparative demography of a lizard. - Ecology 53: 570-584.
- and Dunham, A. E. 1986. Comparative life histories of two syntopic sceloporine lizards. - Copeia 1986: 1-18.
- , Wilbur, H. M. and Tilley, S. G. 1970. Evolutionary strategies in lizard reproduction. - Evolution 24: 55-74.
Vinegar, M. B. 1975. Life history phenomena in two popula- tions of the lizard Sceloporus undulatus in southwestern New Mexico. - Am. Midl. Nat. 93: 388-402.
Waldschmidt, S. R., Jones, S. and Porter, W. P. 1986. The effect of body temperature and feeding regime on activity, passage time, and digestive coefficient in the lizard Uta stansburiana. - Physiol. Zool. 59: 376-383.
Woodward, F. I. 1992. A review of the effects of climate on vegetation: Ranges, competition, and composition. - In: Peters, R. L., and Lovejoy, T. E. (eds), Global warming and biological diversity. Yale Univ. Press, New Haven, CT, pp. 105-123.
Appendix A. Derivation of growth expression when daily growth rate varies non-linearly with potential activity time.
Substituting expression (10) into eq. (4) yields an ex- pression for growth rate as a function of time of year:
dM = M )M -s-- =r,,U.AMl - 1 M1
x L2cd l- 2)-4kd(2l-(j )) 2)) (11)
Integrating eq. (11) over the growth period yields the following expression for body size as a function of time of year:
f(j) 1 1 -- M(j) = () - + M
fYo)_Mo M,nc.x- Mnax -. (12)
where Mo is body mass at the beginning of the time period and
f(/) = exp r,,, -- j/y2 _j2 +y2 sin-l Y -
- j3 -
+4kd2 J3- 'K - 3y _
This model is a more general case of the model de- scribed in the text, which is equivalent to the case k = 0.
Appendix B. Allometry of hatchling size, minimum re- productive size, and time to maturity in lizards.
(13)
Several empirical studies (Sinervo and Adolph 1989, 1994) have shown that daily growth rates may show diminishing returns as a function of daily activity time. We can model this effect as follows:
r= r, (ch - kh2), (9)
choosing a value of k such that r increases monotoni- cally with h over the range of activity times experienced in nature. Values for k between 0 and 0.04c yield a range of responses similar to those observed in the laboratory. The advantage of using a quadratic form to describe diminishing returns, versus an exponential or some other expression, is that (9) leads to analytically tractable solutions.
Substituting (9) into (1) gives the following expres- sion for the dependence of daily growth rate on activity time:
d-=r,, ,x(ch-kh2)M(l-M ) (10) dt M^J
Andrews (1982) provides tables of size data for a number of lizard species, including hatchling size, size at reproductive maturity (females), and maximum body size. We converted these data (snout-vent lengths in mm) to mass (g) using the interspecific allometric rela- tionship SVL = 29.98M0333 (Porter and Tracy 1983), and used least-squares linear regression of log-trans- formed data to derive the following allometric relation- ships for female lizards:
Mlltlchil= 0.0866M?, l.' (14)
and
M, .t,,,ty =0.623M? ,843 ......l'tt Illa . I (15)
These equations can be used to estimate hatchling and minimum reproductive sizes when only maximum size is available. Shine and Charnov (1992) present addi- tional allometric analysis of size at maturity and maxi- mum size in lizards and snakes.
278 OIKOS 77:2 (1996)
(Y)_




















