
Captured (and disturbing for some): A last look at the abortionist
Upload
unipamplonaCategory
view
1download
0
S(
Ca
b
c
9
a
ARRA
KRGFCA
1
oidl3t
trmK2
bAs
0d
Fisheries Research 110 (2011) 207–216
Contents lists available at ScienceDirect
Fisheries Research
journa l homepage: www.e lsev ier .com/ locate / f i shres
ex ratios, size at sexual maturity, and spawning seasonality of dolphinfishCoryphaena hippurus) captured in the Gulf of Tehuantepec, Mexico
armen Alejo-Plataa,∗, Píndaro Díaz-Jaimesb, Isaías H. Salgado-Ugartec
Universidad del Mar, Ciudad Universitaria, Puerto Ángel, San Pedro Pochutla, C.P. 70902, Oaxaca, MexicoInstituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Apartado Postal 70-305, C.P. 04510, D.F., MexicoFacultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México, Av. 5 de Mayo y Fuerte de Loreto, Col. Ejército de Oriente, Iztapalapa Apdo, Postal 09230, C.P.-020, D.F., Mexico
r t i c l e i n f o
rticle history:eceived 11 November 2010eceived in revised form 12 April 2011ccepted 12 April 2011
eywords:eproductionulf Tehuantepececundity
a b s t r a c t
Aspects of the reproductive biology of dolphinfish (Coryphaena hippurus) in the Gulf of Tehuantepec,Mexico, were analysed. A total of 7371 fish were obtained from catches of artisanal fisheries duringthe period of December 2004–2007. The reproductive activity was assessed using gonad histology,oocyte development, maximum whole oocyte size, maturity, and gonadosomatic index. The sex ratio(males:females) was 1:1, except in April–May (1:1.5, P < 0.05) and November (1:0.5, P < 0.05). The sexratio at different size classes showed a significant bias towards females at smaller sizes (<75 cm FL),whereas the males were predominant in the larger size classes (>100 cm FL). The estimated size-at-50%-maturity for females was 48.38 ± 0.84 cm, whereas for males it was 50.57 ± 2.16 cm. The batch fecundity
oryphaena hippurusrtisanal fishery
for female fish between 49 and 129 cm FL was 45,022–1,930,245 hydrated oocytes per female. The aver-age interval between spawns was 2.0 days. The size distribution of oocytes, with at least two groups ofoocytes in the ovaries, suggests that C. hippurus is a multiple spawner with an extended spawning seasonin the study area. The results suggest a pre-spawning migration and evidence of two main spawningevents occurring in May–July and November–January as well as intense utilisation of the coastal zone ofthe Gulf of Tehuantepec as an area for reproduction, with aggregation behaviour during spawning.
. Introduction
The dolphinfish (Coryphaena hippurus Linnaeus, 1758) is anceanic epipelagic fish found worldwide in tropical and subtrop-cal waters. It is a fast-swimming top-level predator, with a highispersal capability via large-scale migrations. This species is short
ived and fast growing, reaching lengths greater than 2 m by agewith a weight up to 30 kg (Palko et al., 1982). The dolphinfish is
argeted by both recreational and commercial fisheries.Studies thus far on the dolphinfish in different fisheries, in both
he Atlantic and Pacific, have shown a clear seasonality in catchates and peaks of abundance. These data suggest a pre-spawningigration (Massutí and Morales-Nin, 1995; Arocha et al., 1999;
raul, 1999; Lasso and Zapata, 1999; Oxenford, 1999; Wu et al.,001; Schwenke and Buckel, 2008).
In the Mexican Pacific, large abundances of dolphinfish have
een reported around Los Cabos and into the Gulf of California.second main area of abundance has also been recognised inouthern Mexico in the Gulf of Tehuantepec (Santana-Hernández,
∗ Corresponding author. Tel.: +52 958 584 3057; fax: +52 958 584 3708.E-mail addresses: [email protected], car [email protected] (C. Alejo-Plata).
165-7836/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.fishres.2011.04.008
© 2011 Elsevier B.V. All rights reserved.
2001). A significant catch is taken along the Pacific coast of Mexicoby artisanal fisheries, even though the area within 80 km of thecoastline is reserved by law for sport fishing (Diario Oficial de laFederación, 1995). Although this species has been considered apotential resource to be exploited by commercial fishing cooper-atives, the scarcity of basic biological information and of catcheshas been the main limitation for the development of an adequatefishery-management plan.
In the Pacific area of Mexico, some aspects of the biology of thedolphinfish have been studied extensively to date. These includeaspects of fishery and catch statistics (Zúniga et al., 2008) on pop-ulation dynamics (Madrid and Beltran-Pimienta, 2001), feeding(Aguilar-Palomino et al., 1998), larval development (Sánchez-Reyes, 2008), and population genetic structure assessed by mtDNAsequences (Díaz-Jaimes et al., 2006; Rocha-Olivares et al., 2006).While no genetic heterogeneity has been detected, significant dif-ferences in allele frequency were observed in samples from the Gulfof California (Tripp-Valdez et al., 2010). Thus, information aboutreproductive biology and reproductive strategies in main abun-
dance areas should add to the knowledge regarding dolphinfishpopulation management units.The aim of this study is to examine aspectsof the reproductivebiology of C. hippurus in the Gulf of Tehuantepec. For this purpose,
208 C. Alejo-Plata et al. / Fisheries Res
F2
tsyb
2
2
Mebmtwrt(IBf
2
sDiabiw(ln(pum
0g(2t
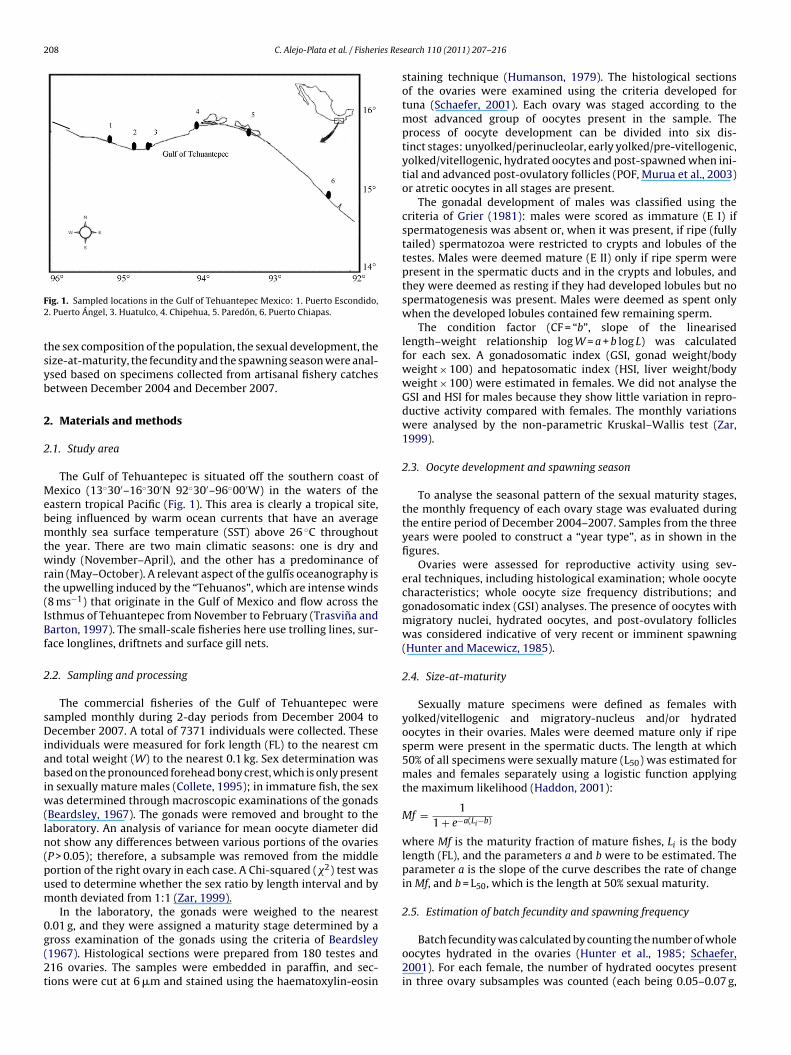
ig. 1. Sampled locations in the Gulf of Tehuantepec Mexico: 1. Puerto Escondido,. Puerto Ángel, 3. Huatulco, 4. Chipehua, 5. Paredón, 6. Puerto Chiapas.
he sex composition of the population, the sexual development, theize-at-maturity, the fecundity and the spawning season were anal-sed based on specimens collected from artisanal fishery catchesetween December 2004 and December 2007.
. Materials and methods
.1. Study area
The Gulf of Tehuantepec is situated off the southern coast ofexico (13◦30′–16◦30′N 92◦30′–96◦00′W) in the waters of the
astern tropical Pacific (Fig. 1). This area is clearly a tropical site,eing influenced by warm ocean currents that have an averageonthly sea surface temperature (SST) above 26 ◦C throughout
he year. There are two main climatic seasons: one is dry andindy (November–April), and the other has a predominance of
ain (May–October). A relevant aspect of the gulfıs oceanography ishe upwelling induced by the “Tehuanos”, which are intense winds8 ms−1) that originate in the Gulf of Mexico and flow across thesthmus of Tehuantepec from November to February (Trasvina andarton, 1997). The small-scale fisheries here use trolling lines, sur-
ace longlines, driftnets and surface gill nets.
.2. Sampling and processing
The commercial fisheries of the Gulf of Tehuantepec wereampled monthly during 2-day periods from December 2004 toecember 2007. A total of 7371 individuals were collected. These
ndividuals were measured for fork length (FL) to the nearest cmnd total weight (W) to the nearest 0.1 kg. Sex determination wasased on the pronounced forehead bony crest, which is only present
n sexually mature males (Collete, 1995); in immature fish, the sexas determined through macroscopic examinations of the gonads
Beardsley, 1967). The gonads were removed and brought to theaboratory. An analysis of variance for mean oocyte diameter didot show any differences between various portions of the ovariesP > 0.05); therefore, a subsample was removed from the middleortion of the right ovary in each case. A Chi-squared (�2) test wassed to determine whether the sex ratio by length interval and byonth deviated from 1:1 (Zar, 1999).In the laboratory, the gonads were weighed to the nearest
.01 g, and they were assigned a maturity stage determined by a
ross examination of the gonads using the criteria of Beardsley1967). Histological sections were prepared from 180 testes and16 ovaries. The samples were embedded in paraffin, and sec-ions were cut at 6 �m and stained using the haematoxylin-eosinearch 110 (2011) 207–216
staining technique (Humanson, 1979). The histological sectionsof the ovaries were examined using the criteria developed fortuna (Schaefer, 2001). Each ovary was staged according to themost advanced group of oocytes present in the sample. Theprocess of oocyte development can be divided into six dis-tinct stages: unyolked/perinucleolar, early yolked/pre-vitellogenic,yolked/vitellogenic, hydrated oocytes and post-spawned when ini-tial and advanced post-ovulatory follicles (POF, Murua et al., 2003)or atretic oocytes in all stages are present.
The gonadal development of males was classified using thecriteria of Grier (1981): males were scored as immature (E I) ifspermatogenesis was absent or, when it was present, if ripe (fullytailed) spermatozoa were restricted to crypts and lobules of thetestes. Males were deemed mature (E II) only if ripe sperm werepresent in the spermatic ducts and in the crypts and lobules, andthey were deemed as resting if they had developed lobules but nospermatogenesis was present. Males were deemed as spent onlywhen the developed lobules contained few remaining sperm.
The condition factor (CF = “b”, slope of the linearisedlength–weight relationship log W = a + b log L) was calculatedfor each sex. A gonadosomatic index (GSI, gonad weight/bodyweight × 100) and hepatosomatic index (HSI, liver weight/bodyweight × 100) were estimated in females. We did not analyse theGSI and HSI for males because they show little variation in repro-ductive activity compared with females. The monthly variationswere analysed by the non-parametric Kruskal–Wallis test (Zar,1999).
2.3. Oocyte development and spawning season
To analyse the seasonal pattern of the sexual maturity stages,the monthly frequency of each ovary stage was evaluated duringthe entire period of December 2004–2007. Samples from the threeyears were pooled to construct a “year type”, as in shown in thefigures.
Ovaries were assessed for reproductive activity using sev-eral techniques, including histological examination; whole oocytecharacteristics; whole oocyte size frequency distributions; andgonadosomatic index (GSI) analyses. The presence of oocytes withmigratory nuclei, hydrated oocytes, and post-ovulatory follicleswas considered indicative of very recent or imminent spawning(Hunter and Macewicz, 1985).
2.4. Size-at-maturity
Sexually mature specimens were defined as females withyolked/vitellogenic and migratory-nucleus and/or hydratedoocytes in their ovaries. Males were deemed mature only if ripesperm were present in the spermatic ducts. The length at which50% of all specimens were sexually mature (L50) was estimated formales and females separately using a logistic function applyingthe maximum likelihood (Haddon, 2001):
Mf = 11 + e−a(Li−b)
where Mf is the maturity fraction of mature fishes, Li is the bodylength (FL), and the parameters a and b were to be estimated. Theparameter a is the slope of the curve describes the rate of changein Mf, and b = L50, which is the length at 50% sexual maturity.
2.5. Estimation of batch fecundity and spawning frequency
Batch fecundity was calculated by counting the number of wholeoocytes hydrated in the ovaries (Hunter et al., 1985; Schaefer,2001). For each female, the number of hydrated oocytes presentin three ovary subsamples was counted (each being 0.05–0.07 g,

C. Alejo-Plata et al. / Fisheries Research 110 (2011) 207–216 209
Table 1Sex ratio by size for dolphinfish captured in Gulf of Tehuantepec.
Size (cm) Frequency Sex-ratio (m:f) �2
Males Females
26–35 6 19 1:3.16 6.836–45 62 165 1:2.66 46.7446–55 366 657 1:1.80 82.7856–65 458 783 1:1.71 85.1166–75 274 273 1:1 076–85 167 169 1:1.01 0.0186–95 283 278 1:0.98 0.0596–105 820 652 1:0.79 19.17106–115 848 423 1:0.50 142.11116–125 451 61 1:1.14 297.07
ms
2tleswMattt(
oat(
3
3
3fi2bTvml
ft(FNo
(ionw(
Fig. 2. Size-specific sex ratios of C. hippurus used to evaluate sex and size com-
fication scheme for the female fish (Table 2 and Fig. 4). The femalesused to construct the oocyte size frequency distributions ranged insize from 49 to 129 cm FL. The resulting distribution (Kernel densityestimates, EDK with bandwidth = 0.0425) indicated the presence
126–135 121 10 1:0.08 94.06136–145 20 3 1:0.15 12.61
easured to the nearest 0.01 mg). The mean count of the threeubsamples was used as the final batch fecundity for each fish.
To determine the size distribution of oocytes within the ovary,00 oocytes with a diameter greater than 0.2 mm were measured tohe nearest 0.001 mm from each subsample (<0.2 mm perinucleo-ar oocytes are considered the permanent stock of oocytes, Huntert al., 1985). The measurements were made using a Zeiss stereo-copic microscope equipped with a digital camera in conjunctionith a program for image analysis (Motic Images Plus, version 2.0L, Multi Language, Motic Group Corporation). Because oocytes
re basically spherical, the orientation of oocytes with respecto the line of measurement was random. Kernel density estima-ors (KDEs) were used to separate groups of oocytes according toheir diameter using the method described by Salgado-Ugarte et al.1997).
The spawning frequency (batch interval) was estimated basedn the frequency of ovaries containing hydrated oocytes (Hunternd Macewicz, 1985). The inverse of the spawning fraction yieldshe average expected interval in days between spawning eventsMurua et al., 2003).
. Results
.1. Size distribution and sex ratio
Samples from a total of 7371 dolphinfish were collected during7 months of study, including 3494 females and 3,877 males. Thesh sampled ranged in size from 20.5 to 129 cm FL for females and5.5–152 cm FL for males. Changes in the sex ratio and size distri-ution were observed during the studied period (Table 1 and Fig. 2).he catches presented a multimodal distribution with a wide inter-al of sizes, although the fishing activity particularly impacted theodes of 50 and 100 cm FL. The modes of 80 and 90 cm FL were the
east abundant in the artisanal catches.Dolphinfish are dioecious organisms with sexual dimorphism, a
act that was evident in specimens larger than 35 cm FL. The propor-ion of both sexes during the studied period was approximately 50%Fig. 3), except for the April–May period, when females (size < 75 cmL) were clearly predominant (1 M:1.5 F; �2 = 51.67, P < 0.05) andovember–December, when a significant dominance of males wasbserved (1 M:0.5 F, P < 0.05) at sizes above 100 cm FL.
Females appeared to be the most favoured in smaller size classes<50 cm FL), with proportions reaching 70%. In specimens rangingn size from 55 to 100 cm FL, the proportion of females levelledff at approximately 65%. Thereafter, a significant reduction in the
umber of females for size classes beyond > 100 cm FL (Table 1)as observed, thus favouring a higher proportion of larger males>120 cm FL) throughout the year.
position of catches from the artisanal fishery from the Gulf of Tehuantepec. Datacollected from December 2004 to December 2007.
3.2. Maturity classification and oocyte development
Using the criteria of Grier (1981), immature males (E I, 22%)were present in catches throughout the year, with the exception ofSeptember, and they had the greatest abundance from May to July.Mature males (E II, 88%) were found in each month of sampling,with an important decrease in May and June. Males classified asspent or resting by the presence of few remaining sperm in thegonads were observed in catches from April to June and in Decem-ber.
The histological characteristics of the ovaries and the mostadvanced group of oocytes were used to develop a maturity classi-
Fig. 3. Monthly sex ratio, the number inside the bars corresponds to the number ofsampled fish. The data were collected from December 2004 to December 2007.

210 C. Alejo-Plata et al. / Fisheries Research 110 (2011) 207–216
Table 2Maturity classification of female dolphinfish ovaries from samples collected in the Gulf of Tehuantepec.
Stage Maturityclassification
Macroscopic description Microscopic description GSI ± S.D. (samplesize)
E I Immature Bright/dark orange, firm ovaries,neither eggs nor blood vessels visible
Histologically, ovaries contain densely packed unyolked/perinucleolaroocytes (Fig. 4a). Whole oocytes <0.2 mm in diameter, appeartransparent with several vacuoles present in cytoplasm
0.46 ± 0.18 (117)
E II Maturing Orange, firm ovaries, eggs and smallveins visible
Unyolked/perinucleolar oocytes present, most oocytes are early yolkedpre-vitellogenic (Fig. 4b). Whole oocytes appear spherical andtranslucent with lipid droplets locate around the outer periphery ofthe oocyte (0.2–0.4 mm)
0.95 ± 0.26 (48)
E III Mature Bright yellow/orange, turgid ovaries.Eggs and large blood vessels visible
The most advanced group of oocytes in the ovaries containyolked/vitellogenic oocytes. (a) Unyolked/perinucleolar and earlyvitellogenic are also present; (b) advanced vitellogenic oocytes(Fig. 4c). The most advanced group of whole oocytes measure0.4–1.0 mm in diameter, yolk appear dense with several transparentlipid globules surrounding the nucleus (not visible)
3.67 ± 2.04 (237)
E IV Spawning Pale orange/yellow, soft swollenovaries. Large translucent eggs visiblethrough ovary wall, with large bloodvessels surrounding the ovaries
Migratory nucleus oocytes (1.0–1.2 mm) and hydrated oocytes(>1.3 mm in diameter) (Fig. 4d) are the largest group present. Wholehydrated oocytes appear translucent with a single droplet. Lateyolked/vitellogenic oocytes present. No post-ovulatory folliclespresent
4.41 ± 2.36 (310)
E V Spawned Dull red and discolored, particularly atthe posterior end; numerous bloodclots large flaccid ovaries. Few eggsvisible
Ovaries contain visible and abundant post-ovulatory follicles (Fig. 4e),residual hydrated oocytes present, yolked/vitellogenic oocytes and anincreased number of pre-vitellogenic oocytes become visible
0.94 ± 0.29 (33)
E VI Recovering Ovaries completely flaccid, andreduced in diameter. Blood vessels
Perinucleolar oocytes predominate. yolked/vitellogenic oocytes withsigns of atresia present. Few whole yolked/vitellogenic oocytes
ed wi
0.36 ± 0.15 (8)
oddwaiwctesfmsotghafow
batAe
3
1DwMcM
present imbedd
f six modes, which were characterised by Bhattacharya’s proce-ure (Salgado-Ugarte et al., 2005) as six Gaussian components withiameters ranging, as a whole, from 0.1 to 1.99 mm. The first groupas formed by perinucleolar oocytes (<0.2 mm in diameter), which
re transparent and are considered the permanent stock of oocytesn the gonad. The second group (0.2–0.4 mm, mode at 0.29 mm)
as formed by pre-vitellogenic oocytes, which appear spheri-al and translucent with numerous small lipid droplets aroundhe outer periphery of the oocyte. The third group consisted ofarly vitellogenic oocytes (0.4–0.6 mm, mode at 0.47 mm), withmall lipid droplets completely covering the oocyte interior. Theourth group included advanced vitellogenic oocytes (0.7–1.0 mm,
ode at 0.94 mm), which appear with a few large lipid dropletsurrounding the nucleus (not visible). The fifth group consistedf fully developed oocytes (1.0–1.2 mm, mode at 1.11 mm), withhe nucleus migrating towards the oocyte’s animal pole and withradually coalescing lipid droplets. The sixth group consisted ofydrated oocytes (>1.3 mm, mode at 1.42 mm) with a translucentppearance and with a single droplet and the lack of post-ovulatoryollicles, a status that indicates imminent spawning (Fig. 5). Webserved that the diameter of the hydrated oocytes did not increaseith the size of the female.
The monthly changes in oocyte diameter distribution fromimodality to multimodality can be observed in Fig. 6. A group withdiameter greater than 1.0 mm was observed in each month, and
he largest oocytes were present in June and December (>1.3 mm).correspondence between high GSI values and oocytes with diam-
ters greater than 1.0 mm was also observed.
.3. Spawning season
The ovarian maturation indicated that immature females (E I,.9% capture) with furcal lengths less than 25 cm were caught fromecember to August. Early maturing females (E II, 7% capture)
ere observed throughout the year and were most abundant fromay to August. Mature females (E III, 58.6% capture) appeared inatches throughout the year, with two peaks of intense activity,arch–April and October–November, which coincided with rel-
thin the compacting tissue of the ovary
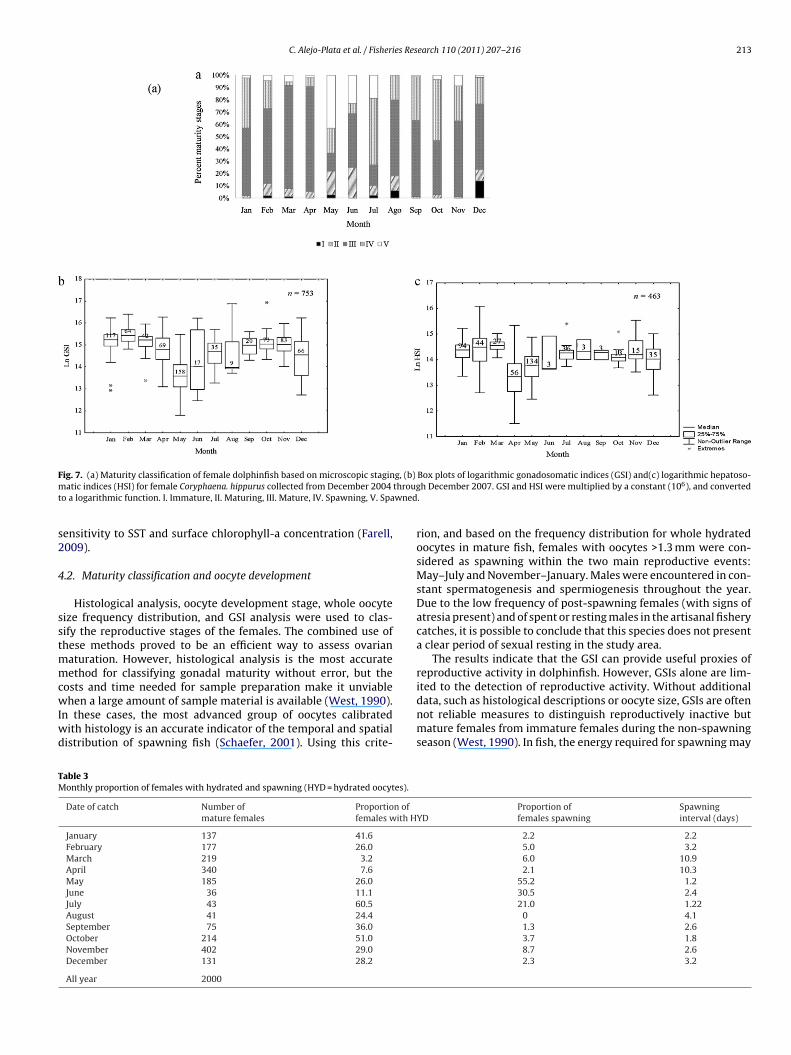
atively high GSI values (>4). Spawning females appeared in thecatches throughout the year (E IV; 22.4% capture), with peaksof intense activity, May–July and November–January. Spawnedfemales (E V; 9.2%) and recovering stage females (E VI, 1%) appearedin the catches from May to July and November (Fig. 7a).
Although the reproductive activity of female dolphinfish canbe classified based on the values of the GSI, the prediction accu-racy improves when a measure of ovarian maturation (as themost advanced group of oocytes present or histological analy-sis) is included. The GSI values presented significant differencesthroughout the year (H(11, 732) = 610.6666, P < 0.01), with the high-est values from September to November and February to April andwith markedly low values in May (Fig. 7b). The HSI of femalesshowed slight fluctuations paralleling changes in GSI; however, itdid not show any specific trend in relation to the spawning state(Fig. 7c). The condition factor (CF) of both sexes did not show anysignificant differences. The CF for females ranged from 2.1 in Mayto 3.1 in November, whereas it ranged from 2.7 in December to3.3 in November for males. The condition factors, regardless ofsex, peaked during the pre-spawning pulses. Males, however, werefatter than females at FL larger than 90 cm.
The combined information of the seasonal distribution ofmature females (Table 3) and GSI results indicate that the dolphin-fish spawned throughout the year with intense activity and massin two peaks. The first peak occurred from May to July, when smallfemales predominated (FL < 80 cm), and the second peak occurredfrom November to January, when there was an abundance of largefemales (FL > 100 cm).
3.4. Size-at-maturity
The estimated size at sexual maturity (L50) for females was48.38 ± 0.84 (95% Confidence Interval, CI) cm FL, and 100% weremature at approximately 64 cm FL. For males, the L50 was
50.57 ± 2.16 (95% CI) cm FL, with 100% maturity reached at 67 cmFL (Fig. 8). The L50 of females was slightly smaller than for males,but the confidence limits for this parameter overlap with those ofthe males.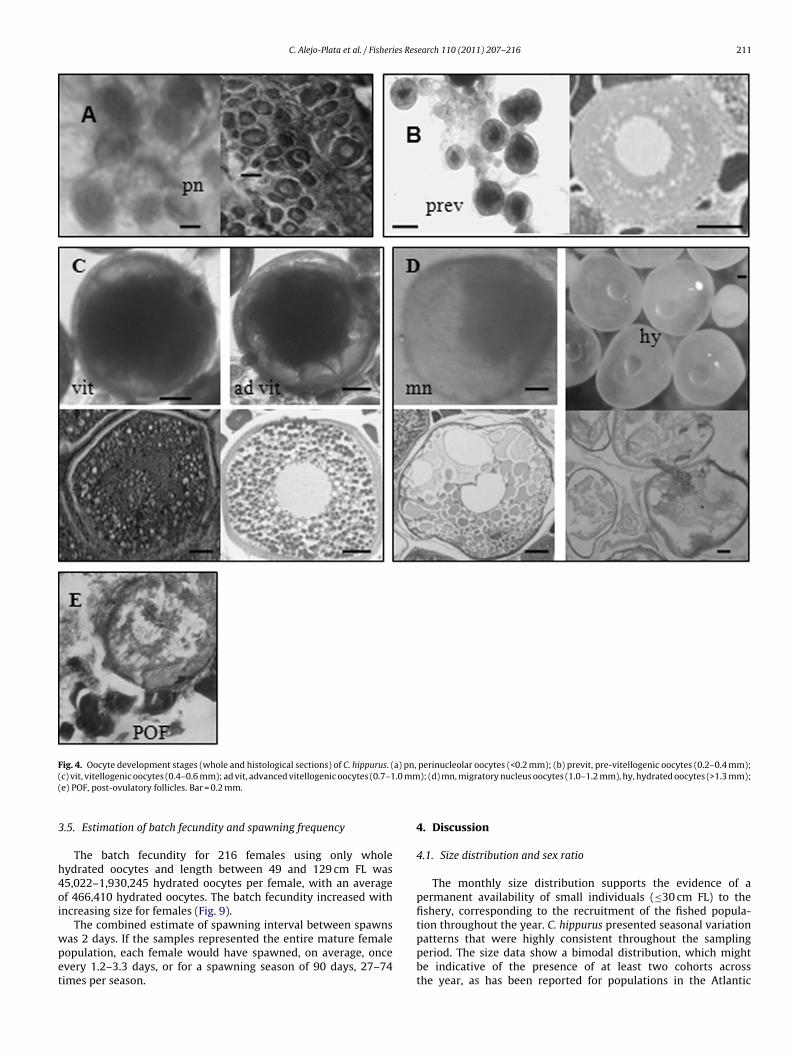
C. Alejo-Plata et al. / Fisheries Research 110 (2011) 207–216 211
Fig. 4. Oocyte development stages (whole and histological sections) of C. hippurus. (a) pn, perinucleolar oocytes (<0.2 mm); (b) previt, pre-vitellogenic oocytes (0.2–0.4 mm);( .0 mm(
3
h4oi
wpet
c) vit, vitellogenic oocytes (0.4–0.6 mm); ad vit, advanced vitellogenic oocytes (0.7–1e) POF, post-ovulatory follicles. Bar = 0.2 mm.
.5. Estimation of batch fecundity and spawning frequency
The batch fecundity for 216 females using only wholeydrated oocytes and length between 49 and 129 cm FL was5,022–1,930,245 hydrated oocytes per female, with an averagef 466,410 hydrated oocytes. The batch fecundity increased withncreasing size for females (Fig. 9).
The combined estimate of spawning interval between spawns
as 2 days. If the samples represented the entire mature femaleopulation, each female would have spawned, on average, oncevery 1.2–3.3 days, or for a spawning season of 90 days, 27–74imes per season.); (d) mn, migratory nucleus oocytes (1.0–1.2 mm), hy, hydrated oocytes (>1.3 mm);
4. Discussion
4.1. Size distribution and sex ratio
The monthly size distribution supports the evidence of apermanent availability of small individuals (≤30 cm FL) to thefishery, corresponding to the recruitment of the fished popula-tion throughout the year. C. hippurus presented seasonal variation
patterns that were highly consistent throughout the samplingperiod. The size data show a bimodal distribution, which mightbe indicative of the presence of at least two cohorts acrossthe year, as has been reported for populations in the Atlantic
212 C. Alejo-Plata et al. / Fisheries Res
Fig. 5. Size distribution of whole oocyte stages in ovaries of dolphinfish by usingband width (h = 0.0425) as suggested by the Silverman test. The dashed line repre-sents the group of perinucleolar oocytes and the permanent stock of undevelopedoocytes (<0.2 mm), second group (0.2–0.4 mm, mode at 0.29 mm), third group(0.4–0.6 mm, mode at 0.47 mm), fourth group (0.7–1.0 mm, mode at 0.94 mm), fifthgroup (1.0–1.2 mm, mode at 1.11 mm), sixth group (>1.3 mm, mode 1.42 mm). I(n = 66), II (n = 244), III (n = 2047), IV (n = 783), V (n = 389).
Fig. 6. Violin plots showing monthly variations in the diameter of oo
earch 110 (2011) 207–216
(Massutí and Morales-Nin, 1995) and the North Pacific (Kraul,1999).
Large fish such as C. hippurus also tend to associate with floatingobjects, whether natural or artificial, such as logs, branches, ropes,buoys and kelp (Kojima, 1961; Rose and Hassler, 1974). Kojima(1961) and Manooch et al. (1984) reported that dolphinfish feedon fish that use floating objects as shelter and that they move awayfrom them only when there is a chance of catching larger prey. Inthe Gulf of Tehuantepec, most of these floating objects are land veg-etation transported by rivers flowing into the ocean, mainly duringthe rainy season, which coincides with the presence of the highestdemersal fish biomass values (Tapia-García, 1998). This factor maypartially explain the presence of dolphinfish close to the coast.
The sex ratio showed differences by length, with the largestproportion of females (70%) was found in size classes <65 cm FLthroughout the year and the largest proportion of males was foundin sizes >120 cm FL. An increase in the proportion of males with sizehas been observed in other geographical regions (Rose and Hassler,1974; Massutí and Morales-Nin, 1997; Oxenford, 1999; Wu et al.,2001). Rose and Hassler (1974) found different proportions of thesexes according to size, and they attributed this to the hypothesisof differential schooling and feeding patterns between large andsmall dolphinfish. The differences could also be caused by segrega-tion because of the need for males to supply their energy demands,which are apparently higher than for females. Males consume ahigher proportion of free-swimming pelagic species than females,which is consistent with the suggestion that they spend more timeaway from flotsam. As the size increases, these fish increase theirenergy requirements and consumption rates (Olson and Galván-Magana, 2002).
The results given here show that temporal differences in thecatches of both male and female dolphinfish occur only duringpeaks of spawning. In the Gulf of Tehuantepec, the predomi-nance of females throughout the year and of spawning females inMay–July and November–January combined with the large males(>100 cm FL) outnumbering females in November suggests thatthe differences in sex ratio observed may be due to the reproduc-tive behaviour of dolphinfish. The temporal patterns of changesin sex ratios are therefore consistent with sexual differences inmigratory, spawning, or other reproductive behaviours, such asthe greater propensity for males to aggregate in spawning groupsfor pelagic species (Hunter and Macewicz, 1985), which can also
constitute an explanation for the observed temporal genetic dif-ferences (Tripp-Valdez et al., 2010). Tagging studies indicate thatdolphinfish also occur seasonally in some areas, with strong spatialcytes. Data collected from December 2004 to December 2007.
C. Alejo-Plata et al. / Fisheries Research 110 (2011) 207–216 213
Fig. 7. (a) Maturity classification of female dolphinfish based on microscopic staging, (b) Box plots of logarithmic gonadosomatic indices (GSI) and(c) logarithmic hepatoso-m througt wned
s2
4
sstmmcwIwd
TM
atic indices (HSI) for female Coryphaena. hippurus collected from December 2004o a logarithmic function. I. Immature, II. Maturing, III. Mature, IV. Spawning, V. Spa
ensitivity to SST and surface chlorophyll-a concentration (Farell,009).
.2. Maturity classification and oocyte development
Histological analysis, oocyte development stage, whole oocyteize frequency distribution, and GSI analysis were used to clas-ify the reproductive stages of the females. The combined use ofhese methods proved to be an efficient way to assess ovarian
aturation. However, histological analysis is the most accurateethod for classifying gonadal maturity without error, but the
osts and time needed for sample preparation make it unviable
hen a large amount of sample material is available (West, 1990).n these cases, the most advanced group of oocytes calibratedith histology is an accurate indicator of the temporal and spatialistribution of spawning fish (Schaefer, 2001). Using this crite-
able 3onthly proportion of females with hydrated and spawning (HYD = hydrated oocytes).
Date of catch Number ofmature females
Proportion offemales with H
January 137 41.6February 177 26.0March 219 3.2April 340 7.6May 185 26.0June 36 11.1July 43 60.5August 41 24.4September 75 36.0October 214 51.0November 402 29.0December 131 28.2
All year 2000
h December 2007. GSI and HSI were multiplied by a constant (106), and converted.
rion, and based on the frequency distribution for whole hydratedoocytes in mature fish, females with oocytes >1.3 mm were con-sidered as spawning within the two main reproductive events:May–July and November–January. Males were encountered in con-stant spermatogenesis and spermiogenesis throughout the year.Due to the low frequency of post-spawning females (with signs ofatresia present) and of spent or resting males in the artisanal fisherycatches, it is possible to conclude that this species does not presenta clear period of sexual resting in the study area.
The results indicate that the GSI can provide useful proxies ofreproductive activity in dolphinfish. However, GSIs alone are lim-ited to the detection of reproductive activity. Without additional
data, such as histological descriptions or oocyte size, GSIs are oftennot reliable measures to distinguish reproductively inactive butmature females from immature females during the non-spawningseason (West, 1990). In fish, the energy required for spawning mayYDProportion offemales spawning
Spawninginterval (days)
2.2 2.25.0 3.26.0 10.92.1 10.3
55.2 1.230.5 2.421.0 1.22
0 4.11.3 2.63.7 1.88.7 2.62.3 3.2

214 C. Alejo-Plata et al. / Fisheries Res
Fig. 8. Proportion of mature specimens by length in male and female dolphinfishco
bvchitwM
caaovfHpevoas
Ff
aught in Gulf of Tehuantepec at 5 cm length intervals. Fitted curve illustrate theptimal logistic maturation ogive fitted by maximum likelihood. (a) female, (b) male.
e derived from liver oil reserves (Diana and Mackay, 1979) or theiscera (Rajasilta, 1992); in this work, the HSI values showed nolear trend related to spawning. This species is voracious, with aigh feeding activity (Palko et al., 1982) that does not decrease dur-
ng the spawning period (Olson and Galván-Magana, 2002). Thus,he required energy for spawning might be derived from the diet,ithout any significant use of their energy reserves (Massutí andorales-Nin, 1997).Ovaries in the phase of nucleus migration and lipid globule
oalescence represent a stage prior to oocyte hydration, which isrecurring stage throughout the sampling period in females from
ll locations. This characteristic may be considered an indicatorf constant spawning because once the migration of the germinalesicles is initiated, hydration of the oocytes can occur within aew hours of meiosis re-initiation (Wallace and Selman, 1981).istological analysis was necessary to describe the spawningattern. C. hippurus presents an asynchronous spawning strat-gy that requires the constant presence of pre-vitellogenic anditellogenic oocytes, as was confirmed in almost all of the gonads
f the females analysed, which also presented hydrated oocytesnd post-ovulatory follicles. The simultaneous presence of at leastix types of clearly differentiated oocytes, both morphologicallyig. 9. Relationship between batch fecundity and fork length of C. hippurus collectedrom December 2004 through December 2007 in Gulf of Tehuantepec.
earch 110 (2011) 207–216
and quantitatively (diameter), represented six partial potentialspawnings that would sustain such a strategy.
Descriptions of reproductive strategies and the assessment offecundity are fundamental topics in the study of the biology andpopulation dynamics of fish species (Schaefer, 2003). The repro-ductive aspects studied show sexual maturity at 48–55 cm FL in thefirst year of life (Alejo-Plata et al., in press), a high fecundity, and anasynchronous reproductive strategy. This reproductive behaviourhas also been observed for dolphinfish in the Atlantic (Arochaet al., 1999; Oxenford, 1999) and the Mediterranean (Massutí andMorales-Nin, 1997), and it is typical of sub-tropical and tropicalfishes (Burt et al., 1988). This strategy is usually considered adap-tive to changes in environmental conditions, minimising the risk ofhaving a single expulsion during the spawning season (Ditty et al.,1994) and probably taking advantage of favourable environmentalconditions for larval survival and recruitment.
4.3. Spawning season
In tropical areas such as the Gulf of Tehuantepec, where dol-phinfish are present year round, the maturation of the gonads canbe followed through a complete annual cycle. The spawning sea-son for dolphinfish is defined and identified by a combination ofseveral factors including: (1) high GSI values; (2) the identifica-tion of females at imminent spawning (with hydrated oocytes); and(3) ovaries containing visible and abundant post-ovulatory folliclesand residual hydrated oocytes.
The spawning behaviour presented by dolphinfish appears tobe congruent with an opportunistic reproductive strategy in whichindividuals spawn at times when favourable conditions for preyavailability are present in the area. The Gulf of Tehuantepec is ahighly productive area with highly complex biological cycles. Dis-tinct seasonal changes in primary and secondary production havebeen observed. These changes are mainly influenced by periodicevents of wind upwelling that occur from November to Februaryand during the rainy season (June–October) (Lara-Lara et al., 1998).Maximum phytoplankton production occurs during the season ofnortherly winds, or “Tehuanos” (Trasvina and Barton, 1997), sug-gesting that this highly productive zone is an important feedingground for the fishes that migrate through it. According to Olsonand Galván-Magana (2002), this is an area of high prey diversity fortop-level predators such as tuna, sharks, billfishes and dolphinfish,which primarily prey on crustaceans, small fishes, and squid.
The dolphinfish extensively uses the coastal zone of the Gulf ofTehuantepec as an area of reproduction, exhibiting marked aggre-gation behaviour during the spawning season. The results suggesta pre-spawning migration, and they appear to show different sizegroups arriving at different time periods. The first group com-prised smaller specimens (50–80 cm FL) and was formed mostlyby females that will spawn during the beginning of the rainyseason, arriving in March–April. Newly hatched dolphinfish inMay–July will ensure survival based on the biological productionoriginated by the fluvio-lagoonal systems, which discharge con-siderable amounts of nutrients into the Gulf. The second groupcomprised specimens larger than 100 cm FL and with a larger frac-tion of males than females, arriving during October–November. Themost intense spawning period is from November to January, dur-ing the most intense upwelling season, when it is possible to finda sufficient food supply for gonad development that will ensuresuccessful spawning. Similar observations on pre-spawning migra-tion have been reported in the Caribbean (Oxenford, 1999), Ecuador(Campos et al., 1993), the Mediterranean (Massutí and Morales-Nin,
1995), Venezuela (Arocha et al., 1999), and in Colombian waters(Lasso and Zapata, 1999).These pulses in spawning activity can be explained by thehypothesis of seasonal coast-sea movement proposed by Kraul

es Res
(dAeeiiwaoothaSecJbddm
pifioo2tc
4
m(apaCtbFtmtrsuatom
nddptbshfid2
C. Alejo-Plata et al. / Fisheri
1999) and can be further analysed through changes in the con-ition of the dolphinfish, as proposed Uchiyama and Boggs (2006).similar movement of seasonal spawning was suggested by Arocha
t al. (1999) in waters off of Venezuela. In the study area, the high-st CF values occurred in pulses of pre-spawning migration, thats, October–November and March–April, which coincides with anncrease in coastal catches. Well-conditioned fish enter nearshore
aters from the open ocean and engage in intense reproductivectivities, resulting in weight loss. C. hippurus then returns to thepen ocean in an emaciated state. In the June–August period, webserved the lowest condition values, and the dolphinfish essen-ially disappeared from the artisanal fishery, perhaps because theyad migrated offshore to feed and improve their condition duringnon-reproductive period. When the dolphinfish returned in late
eptember and October, their condition was at the highest level,vidence of which can be found in the abundance of fish in theatch of the longline fishery off the Gulf of Tehuantepec duringune to September (Santana-Hernández, 2001). This reproductiveehaviour may represent spawning migration circuits such thatolphinfish exhibit a more complex stock structure than has beenemonstrated, making a re-examination of population structure byeans of genetic data necessary (Oxenford, 1999).The location of dolphinfish spawning in the Pacific Ocean is
oorly documented because of the lack of ichthyoplanktonic stud-es, but it is presumably widespread based on the presence of ripesh. Sánchez-Reyes (2008) contends that the highest abundancef dolphinfish larvae in pre-flexion along the North Pacific Coastf México is present within the 28 ◦C isotherm in summer and the4 ◦C isotherm in spring. In the present study, mature females withhe most advanced group of oocytes (>1.3 mm diameters) wereonsidered imminent spawners.
.4. Size-at-maturity
For the Mexican Pacific, there are no previous reports withaturity estimations on the length L50 for dolphinfish. Our results
L50 = 48.38 and 50.57 cm FL for females and males, respectively)re nearly identical to those reported by Wu et al. (2001) for dol-hinfish populations off Taiwan (L50 = 51 cm FL for both sexes)nd by Schwenke and Buckel (2008) for dolphinfish off Northarolina (L50 = 45.8 and 47.6 cm FL for females and males, respec-ively). However, our results were different from those reportedy Beardsley (1967) for the Florida straits (L50 = 35 and 45 cmL for females and males, respectively), by Oxenford (1999) forhe western Atlantic (L50 = 84 cm and 80.5 cm FL for females and
ales, respectively), and by Massutí and Morales-Nin (1997) forhe Mediterranean (L50 = 54.5 and 61.8 cm FL for females and males,espectively). However, these analyses were derived from macro-copic assessments of gonads without histological calibration. These of histological techniques and an assessment of the mostdvanced group of oocytes in the ovaries to assess maturity reduceshe possibility of including post-spawning females and immaturer inactive females and has proven to have greater precision foraturity at size estimates (DeMartini et al., 2000).Likewise, these L50 estimates for dolphinfish in the Atlantic may
ot be completely comparable to ours because the studies mightiffer in whether the estimates were applied to fish caught onlyuring spawning periods. The present study included mature dol-hinfish of relatively small sizes caught throughout the year. Inhe Gulf of Tehuantepec, the gonad maturity of dolphinfish occursefore one year of age, and the peak of spawning should corre-pond with the first year of age. In other latitudes, a few authors
ave reported that the gonad maturity of C. hippurus occurs in therst year of life, although spawning peaks are not present until theolphinfish reach two years of age (Palko et al., 1982; Wu et al.,001).earch 110 (2011) 207–216 215
4.5. Estimation of batch fecundity and spawning frequency
In species with an asynchronous oocyte development patternand indeterminate fecundity, batch fecundity estimates are com-monly made from counts of non-ovulated hydrated oocytes toavoid biases due to the release of eggs (Hunter et al., 1985; Muruaet al., 2003). However, some authors have used migratory nucleusstage oocytes in batch fecundity estimates of highly migratoryspecies (Schaefer, 2001). However, migratory nucleus stage oocytescan be accurately identified in histological sections, because wholeoocytes are difficult to recognise (Hunter et al., 1985). Therefore, toavoid overestimating the values of fecundity by confusing oocytesin the ultimate phase of vitellogenesis with oocytes in the stage ofnucleus migration, our estimations of batch fecundity were basedon counts of whole hydrated oocytes. Reports with varying esti-mates of batch fecundity in dolphinfish at other latitudes exist inthe literature (Beardsley, 1967; Massutí and Morales-Nin, 1997;Oxenford, 1999; Wu et al., 2001). A comparison of estimates offecundity in this study with those of other authors is not feasi-ble due to the different methodologies used (e.g., their estimatesincluded counts of vitellogenic oocytes, differences in size rangesampled, and oocyte size thresholds).
The present study is the first report on batch fecundity for C.hippurus in the Mexican Pacific based on counts of whole hydratedoocytes. The batch fecundity for females between 49 and 129 cm FLwas 450,222–1,930,245 whole hydrated oocytes per female. Fecun-dity was strongly correlated with size; in small females between49 and 129 cm FL, the estimated average fecundity was 52,700hydrated oocytes.
In species with an indeterminate fecundity pattern, the spawn-ing fraction is probably the most difficult to produce because of thelow number of samples available. Most of the work on estimatingspawning fractions in highly migratory species comes from stud-ies on tuna species (Schaefer, 2001; Schaefer, 2003). The preferredmethod has been the histological identification of post-ovulatoryfollicles (Hunter and Macewicz, 1985). Because of the limitationsduring the present study (small sample size and limited histologi-cal sections), the spawning frequency estimates for the dolphinfishwere based on the presence of hydrated oocytes. This methodhas restrictions due to the limited time period in which hydratedoocytes are present in the ovary prior to spawning, and it mayoverestimate the spawning fraction.
A microscopic examination of histological slides of ovarian andtesticular tissue provides the greatest precision for the classifica-tion of reproductive states of C. hippurus. The results of the presentstudy established an understanding of several key aspects of thereproductive biology of C. hippurus in the Gulf of Tehuantepec. Dol-phinfish obtained directly from the catches of artisanal vessels,where they were caught by different types of fishing gear and at var-ious stages of development, were exceptionally useful for analysinga much greater temporal and spatial distribution of spawning fishthan has been reported so far. Consequently, accurate estimationscould be made of the spawning season, size at maturity, and thebatch fecundity. The results identified the Gulf of Tehuantepec asan important area for C. hippurus spawning in the eastern tropicalPacific. In terms of spawning strategy, there is an obvious need fordetailed information on spawning locations from larval collectionsand on the temporal and spatial oceanographic conditions in theGulf of Tehuantepec.
Acknowledgements
The first author acknowledges the fishermen’s cooperative soci-eties of Oaxaca and Chiapas for the information provided. Wegreatly appreciate the support of our colleagues in the ‘Labora-torio de ictiología y biología pesquera’ at Universidad del Mar.

2 es Res
TfiiPts
R
A
A
A
B
B
C
C
D
D
D
D
D
F
G
H
H
H
H
K
K
L
16 C. Alejo-Plata et al. / Fisheri
his research was carried out as a part of the PhD studies of therst author in the Posgrado en Ciencias Biológicas UNAM. Fund-
ng was provided by the Comisión Nacional de Acuacultura yesca (CONAPESCA 2IR0502), SAGARPA-CONACYT-225-2003 andhe State Governments of Oaxaca. The manuscript benefited fromuggestions and recommendations of the anonymous reviewers.
eferences
lejo-Plata, M.C., Gómez-Márquez, J.L., Salgado-Ugarte, I., 2011. Edad y crecimientodel dorado Coryphaena hippurus en el Golfo de Tehuantepec, México. Rev. Biol.Mar. Ocean (in press).
guilar-Palomino, B., Galván-Magana, F., Abitia-Cardenas, A., Muhlia-Melo, A.,Rodríguez-Romero, J., 1998. Aspectos alimentarios del dorado Coryphaena hip-purus Linnaeus, 1758 en cabo San Lucas, Baja California Sur, México. Cienc. Mar.24, 253–265.
rocha, F., Marcano, L., Lárez, A., Altuve, D., Alio, J., 1999. The fishery, demographicsize structure and oocyte development dolphinfish, Coryphaena hippurus, inVenezuela and adjacent waters. Sci. Mar. 63, 401–409.
eardsley, G., 1967. Age, growth and reproduction of the dolphinfish, Coryphaenahippurus, in the Straits of Florida. Copeia 1967, 441–451.
urt, A., Kramer, D.L., Nakatsura, K., Spry, C., 1988. The tempo of reproduction inHyphessobrycon pulchripinis (Characidae), with a discussion of multiple spawn-ing in fishes. Environ. Biol. Fish. 22, 15–27.
ampos, J., Segura, A., Lizano, O., Madrigal, E., 1993. Ecología básica de Coryphaenahippurus (Pisces: Coryphaenidae) y abundancia de otros grandes pelágicos en elPacifico de Costa Rica. Rev. Bio. Trop. 41, 783–790.
ollete, B.B., 1995. Coryphaenidae. In: Fisher, W., Krupp, F., Schneider, W., Sommer,C., Carpenter, K.E., Niem, V.H. (Eds.), Guía FAO para la identificación de especiespara los fines de la Pesca. Pacífico Centro-Oriental, volumen II. Vertebrados-Parte1. FAO Roma, pp. 1036–1038.
eMartini, E.E., Uchiyama, J.H., Williams, H.A., 2000. Sexual maturity, sex ratio,and size composition of swordfish, Xiphias gladius, caught by the Hawaii-basedpelagic longline fishery. Fish. Bull. 98, 489–506.
iana, J.S., Mackay, W.C., 1979. Timing and magnitude of energy deposition and lossin the body, liver and gonads of northern pike (Esox lucius). J. Fish Res. Boar Can.36, 481–487.
iario Oficial de la Federación, 1995. Norma oficial mexicana NOM-017-PESC-1994.Para regular las actividades de Pesca deportiva en las Aguas de JurisdicciónFederal de los Estados Unidos Mexicanos. Tomo No. 15-19. México, D.F.
íaz-Jaimes, P., Uribe-Alcocer, M., Ortega-García, S., Durand, J.D., 2006. Spatialand temporal mitochondrial DNA genetic homogeneity of dolphinfish popu-lations (Coryphaena hippurus) in the eastern central Pacific. Fish. Res. 80, 333–338.
itty, J., Shaw, R., Grimes, C., Cope, J., 1994. Larval development, distribution, andabundance of common dolphin, Coryphaena hippurus, and pompano dolphin C.equiselis (Fam. Coryphaenidae), in the northern Gulf of Mexico. Fish. Bull. 92,275–291.
arell, E., 2009. The Habitat, Movements, and Management of Dolphin, Coryphaenahippurus, in the Western North Atlantic, Caribbean, and Gulf of Mexico. DukeUniversity, USA, pers. commn.
rier, H.J., 1981. Cellular organization of the testis and spermatogenesis in fishes.Am. Zool. 21, 345–367.
addon, M., 2001. Modeling and Quantitative Methods in Fisheries. Chapman andHall/CRC, FL, 406 p.
umanson, G.L., 1979. Animal Tissue Techniques. W.H. Freeman and Co, San Fran-cisco, 344 p.
unter, J., Macewicz, B., 1985. Measurement of spawning frequency in multiplespawning fishes. In: Lasker, R. (Ed.), An Egg Production Method for Estimat-ing Spawning Biomass of Pelagic Fish: Application to the Northern Anchovy,Engraulis mordax. U.S. National Marine Fisheries Service, National Oceanic andAtmospheric Administration Technical Report 36, pp. 79–94.
unter, J.R., Lo, N.C., Leong, R.J., 1985. Batch fecundity in multiple spawning fishes.In: Lasker, R. (Ed.), Egg Production Method for Estimating Spawning Biomassof Pelagic Fish: Application to the Northern Anchovy, Engraulis mordax. U.S.National Marine Fisheries Service, National Oceanic and Atmospheric Admin-istration Technical Report 36, pp. 67–78.
ojima, S., 1961. Studies of dolphin fishing conditions in the Western Sea of Japan.III. On the stomach contents of dolphin. Bull. Jpn. Soc. Sci. Fish. 27, 625–629.
raul, S., 1999. Seasonal abundance of the dolphinfish, Coryphaena hippurus, in
Hawaii and the tropical Pacific Ocean. Sci. Mar. 63, 261–266.ara-Lara, J.R., Robles-Jarero, E.G., Bazán-Guzmán, M.C., Millán-Nunez, E., 1998. Pro-ductividad del fitoplancton. In: Tapia-Garcia, M. (Ed.), El Golfo de Tehuantepec:el ecosistema y sus recursos. Universidad Autónoma Metropolitana-Iztapalapa,México, pp. 51–58.
earch 110 (2011) 207–216
Lasso, J., Zapata, L., 1999. Fisheries and biology of Coryphaena hippurus (pises:Coryphaenidae) in the Pacific coast of Colombia and Panama. Sci. Mar. 63,387–399.
Madrid, J.V., Beltran-Pimienta, R., 2001. Length, weight and sex of the dolphin fishCoryphaena hippurus (Perciformes: Coryphaenidae), of the littoral of Sinaloa,Nayarit and Baja California Sur, Mexico. Rev. Biol. Trop. 49, 931–938.
Manooch, C.S., Mason, D.L., Nelson, R.S., 1984. Food and gastrointestinal parasites ofdolphin, Coryphaena hippurus, collected along the southeastern and gulf coastsof the United States. Bull. Jpn. Soc. Fish. 50, 1511–1525.
Massutí, E., Morales-Nin, B., 1997. Reproductive biology of dolphinfish (Coryphaenahippurus L) off the island of Majorca (western Mediterranean). Fish. Res. 30,57–65.
Massutí, E., Morales-Nin, B., 1995. Seasonality and reproduction of dolphinfish(Coryphaena hippurus) in the western Mediterranean. Sci. Mar. 59, 357–364.
Murua, H., Kraus, G., Saborido-Rey, F., Witthames, P.R., Thorsen, A., Junquera, S.,2003. Procedures to estimate fecundity of marine fish species in relation to theirreproductive strategy. J. Northw. Atl. Fish. Sci. 33, 33–54.
Olson, R.J., Galván-Magana, F., 2002. Food habits and comparison rates of commondolphinfish (Coryphaena hippurus) in the eastern Pacific Ocean. Fish. Bull. 100,279–298.
Oxenford, H., 1999. Biology of the dolphinfish (Coryphaena hippurus) in the westerncentral Atlantic: a review. Sci. Mar. 63, 277–301.
Palko, B., Beardsley, G., Richards, W., 1982. Synopsis of biological data on dolphin-fishes Coryphaena hippurus and Coryphaena equiselis L. U.S. Dep. Commer. NOAATech. Rep. NMFS Cir. 443, 28 pp.
Rajasilta, M., 1992. Relationship between food, fat, sexual maturation and spawningtime of Baltic herring (Clupea harengus membras) in the Archipelago Sea. Can. J.Fish Aquat. Sci. 49, 644–654.
Rocha-Olivares, A., Bobadilla-Jiménez, M., Ortega-García, S., Saavedra-Sotelo, N.,Sandoval-Castillo, J.R., 2006. Variabilidad mitocondrial del dorado Coryphaenahippurus en poblaciones del Pacífico. Cienc. Mar. 32, 569–578.
Rose, C.D., Hassler, W.W., 1974. Food habits and sex ratios of the dolphin Coryphaenahippurus captured in the western Atlantic Ocean off Hatteras, North Carolina.Trans. Am. Fish. Soc. 103, 94–100.
Salgado-Ugarte, I.H., Gómez-Márquez, J.L., Pena-Mendoza, B., 2005. Métodos actu-alizados para el análisis de datos biológicos-Pesqueros. Escuela Nacional deEstudios Profesionales Zaragoza, UNAM, 240 p.
Salgado-Ugarte, I.H., Shimizu, M., Taniuchi, T., 1997. Nonparametric assessment ofmultimodality for univariate data. Stata Tech. Bull. 38, 27–35.
Sánchez-Reyes, N., 2008. Distribución de larvas de dorado Coryphaena hippurus(Linnaeus, 1758) y Coryphaena equiselis (Linnaeus, 1758) en el Pacífico Orien-tal mexicano. Centro Interdisciplinario de Ciencias Marinas-IPN, La Paz, B.C.S.Mexico, pers. commn.
Santana-Hernández, H., 2001. Estructura de la comunidad de pelágicos mayorescapturados con palangre en el Pacífico mexicano (1983–1996) y su relación conla temperatura superficial del mar. Ph.D. Thesis. Universidad de Colima, Mexico.
Schaefer, K.M., 2003. Estimation of the maturity and fecundity of tunas. In: Kjesbu,O., Hunter, J.R., Witthames, P.R. (Eds.), Modern Approaches to Assess Maturityand Fecundity of Warm and Cold Water Fish and Squids. Institute of MarineResearch, Bergen-Norway, pp. 117–124.
Schaefer, K., 2001. Reproductive biology of tunas. In: Block, B., Stevens, E.D. (Eds.),Tuna Physiology, Ecology, and Evolution. Fish Physiology, vol. 19. AcademicPress, New York, pp. 225–270.
Schwenke, K., Buckel, J., 2008. Age, growth, and reproduction of dolphinfish(Coryphaena hippurus) caught off the coast of North Carolina. Fish. Bull. 106,82–92.
Tapia-García, M., 1998. Evaluación ecológica de la ictiofauna demersal. In: Tapia-García, M. (Ed.), El Golfo de Tehuantepec: el ecosistema y sus recursos.Universidad Autónoma Metropolitana-Iztapalapa, México, pp. 103–128.
Trasvina, A., Barton, E.D., 1997. Los Nortes del Golfo de Tehuantepec: Dinámica delocéano costero. In: Lavin, M.F. (Ed.), Contribuciones a la Oceanografía Física enMéxico, Monografía No. 3. Unión Geofísica Mexicana, pp. 25–46.
Tripp-Valdez, M.A., García de León, F.J., Ortega-García, S., Lluch-Cota, D., López-Martínez, J., Cruz, P., 2010. Population genetic structure of dolphinfish(Coryphaena hippurus) in the Gulf of California, using microsatellite loci. Fish.Res. 105, 172–177.
Uchiyama, J., Boggs, C., 2006. Length-weight relationships of dolphinfish, Coryphaenahippurus, and wahoo, Acanthocybium solandri: seasonal effects of spawning andpossible migration in the central north pacific. Mar. Fish. Rev. 68, 19–29.
Wallace, R.A., Selman, K., 1981. Cellular and dynamic aspects of oocyte growth inteleosts. Am. Zool. 21, 325–343.
West, G., 1990. Methods of assessing ovarian development in fishes: a review. Aust.J. Mar. Freshw. Res. 41, 199–222.
Wu, C.C., Su, W.C., Kawasaki, T., 2001. Reproductive biology of the dolphin fish
Coryphaena hippurus on the east coast of Taiwan. Fish. Sci. 67, 784–793.Zar, J., 1999. Biostatistical Analysis. Prentice Hall, NJ, 718 p.Zúniga, M.S., Ortega-García, S., Klett-Traulsen, A., 2008. Interannual and seasonal
variation of dolphinfish (Coryphaena hippurus) catch rates in the southern Gulfof California, Mexico. Fish. Res. 94, 13–17.
Copyright © 2022 FDOKUMEN








![Batteries In A Portable Wold [captured website] WW.pdf](https://static.fdokumen.com/doc/165x107/631bc4c07051d371800f2b67/batteries-in-a-portable-wold-captured-website-wwpdf.jpg)











