
A Contribution to Enterprise Interoperability Maturity Assessment

J . Fish &of. (1987) 30,73 1-741
Size at maturity in male Gambusia heterochir
H. Y. YAN* Department of Zoology, The University of Texas a1 Austin, Austin, TX 78712, U.S.A
(Received 18 July 1986, Accepted 23 November 1986)
The size at maturity in many male poeciliid fish is believed to be controlled by ' social control mechanism ', and post-maturation growth is insignificant. In the Clear Creek gambusia. Gambu- sia heterochir. post-maturation growth in male fish is limited. Male G. heterochir matures at a fixed size, and age ofmaturation isdependent on environmental factors (e .g . temperature). In the presence of an adult male, the maturation size of a young male is largely determined by genetic factors rather than ' social control ' factors. A male can switch its status as a . dominant ' or ' subordinate ' male, depending on the relative size of rival males in mating competition. Mating success of a large (dominant) male reduces as the number of competing small (subordinate) males increases. These unique reproductive tactics may explain in part why ' social control mechanism ' does not play a key role in determining the maturation size of male G. hererochir.
I. INTRODUCTION
Males of the fish family Poeciliidae greatly reduce growth upon attainment of sexual maturity (Turner, 1941; Schultz, 1961; Hughes, 1985). It is believed that a maturing male that delayed masculinization would attain a large size and peck- order dominance, while individual males maturing at smaller body sizes would be locked into subordinate status in the local, size-dependent, social hierarchy (Constantz, 1975). Borowsky ( 1 9 7 3 ~ 4 demonstrated that in swordtail, Xipho- phorus variatus, the first male to initiate the maturation process is usually the largest of the immature males. The next one to mature is the next largest, and so forth. Given ample space and food, each successively maturing fish does not reach maturity until having exceeded the size of previously matured ones. In Gambusia manni, the size at maturity is functionally related to the presence of an adult male and maturation is delayed until a juvenile exceeds that adult male's size (Sohn, 1977~). In the platyfish, Xiphophorus maculatus, when two individuals are reared together the socially dominant individual delays the maturation of the subordinate (Sohn, 1977b).
Hubbs (1971) reported that the collections of Gambusia a#inis from Clear Creek in September contained males with partially formed gonopodia which were often larger than the largest mature males. Hubbs reasoned that larger size might give males maturing later in the year a competitive edge over already mature males. Hughes (1985) showed that, in southern Indiana, male mosquitofish, Gambusia affinis, born early in the breeding season matured rapidly at a small body size, while males born later in the breeding season delayed maturation and achieved larger body size, and the process was, at least in part, under social control.
*Present address: Port Aransas Marine Laboratory, Marine Science Institute, The University of Texas at Austin. Port Aransas, TX 78373, U.S.A.
0022-1 112/87/060731+ I I $03.00/0 73 I
0 1987 The Fisheries Society of the British Isles

732 H. Y. YAN
This evidence suggests that a ' social control mechanism ' (Borowsky, 1973a) determines the maturation size of male poeciliid fish. The hypothesis that socially dominant animals will experience higher reproductive success than subordinates has received widespread support (Robel, 1966; Zimmerman, 1966; Van den Assem, 1967; Gandolfi, 1971; Daly & Wilson, 1983). Ifa ' socialcontrol mechanism ' really can control the maturation size of poeciliid fish, then age should not be important in determining the onset of maturation. For the platyfish, X. maculatus, Sohn & Crews (1977) demonstrated that the timing of maturation is not genetically determined, but is a function of growth rate and initial size. They concluded that maturation in platyfish depends upon size rather than age.
The Clear Creek gambusia, Gambusia heterochir, is an endemic poeciliid fish which occurs only in headwaters of Clear Creek, Menard County, Texas (Hubbs, 1957). Several aspects of the reproductive biology of this fish have been investi- gated by Hubbs and his associates during the past three decades (see Yan 1986 for bibliographies). This study is intended to answer the following questions related to the size at maturity in male G. heterochir: (1) Does post-maturation growth occur in male fish? (2) Will young males in the presence of adult males delay their maturation until their size exceeds that of the adult males? (3) Does a male mature at a fixed size or fixed age? (4) Will the mating success of a large (dominant) male remain unchanged as the number of small (subordinate) males increases? ( 5 ) Can a male switch its role as ' dominant ' or ' subordinate ' depending on the relative size of rival males in the mating competition?
11. MATERIALS AND METHODS EXPERIMENT I
The purpose of this treatment was to compare the growth rate of both sexes before and after maturation. It was also intended to test if there is any significant post-maturation growth in both sexes. Twenty one adult males, i.e. with fully developed gonopodia, fourteen adult females (standard length, S.L., > 25.0 mm) and thirty young fish (sex unknown, S.L. < 17.8 mm) collected from Clear Creek were used in this study. Each fish was individually reared in a 2-1 finger bowl at 14L:lOD photoperiod and 20" C. The fish were fed live brine shrimp or blended calf liver. The experiment lasted 2 months. The daily specific growth rate (DSGR) of both sexes was calculated by the following formula: DSGR (%)=[(Log,(L,)-Log,(L,))/f,-f,] x 100% where L, is standard length at day 60 ( t z , end of the experiment), and L, is standard length at day 1 (I,, beginning of the experiment)(Weatherley, 1972).
EXPERIMENT I1 This treatment was designed to observe whether or not the maturation of a young male in
the presence of an adult male would be delayed until its size exceeded that of the adult male. Thirty young fish (S.L. < 15.0 mm) and 30 adult males collected from the wild were used in this experiment. In each 68-1 tank, a young fish (sex unknown at this length) was paired with an adult male. The fish were fed twice a day with blended calf liver and live brine shrimp. The experiment was conducted under 14L: 10D light regime and water temperature was maintained at 20 f 0.5" C. When the young fish in each tank had reached maturity (i.e. males with fully developed gonopodia), the experiment was terminated. The standard lengths of both newly matured males and the original adult males were measured.
EXPERIMENT 111 The purpose of this experiment was the same as that of Experiment 11, but laboratory-
produced siblings from the same brood of known parental origin were used instead of wild-

MATURATION SIZE OF MALE GAMBUSIA HETEROCHIR 733
collected juveniles, to avoid the possible bias in selecting genetically determined large or small size fish from wild. Five virgin females were selected from the stock tank and each was inseminated by a different male. When the female was close to parturition it was placed in a parturition tank until the juveniles were born. Siblings of each brood (Broods A, B, C, D and E) were first reared together for 16 days, then randomly divided into two subgroups (subgroup Land S). A large adult male was added to the subgroup L, and a small adult male was added to the subgroup S. Large and small sizes were designated on a relative scale, valid only within a given brood comparison. Each subgroup was reared in a 68-1 aquarium under 14L:lOD at 20" C conditions and fed twice daily with live brine shrimp and blended calf liver. Every month the standard length of each fish was measured and the developmental condition of the gonopodium of young fish was recorded. When all the young fish in a subgroup reached maturity, the experiment was terminated for the subgroup.
EXPERIMENT IV By controlling the growth rate of male fish through manipulation of water temperature,
this experiment was to determine whether a male fish matures at a fixed size or age. Five virgin females were selected from the stock tank and each was inseminated by a different male. When the female was close to parturition, each female was placed in a parturition tank until the brood was produced. Juveniles of the same brood, i.e. siblings, were then randomly and evenly divided into two groups: one group was reared at 20" C (Group L), and the other group reared at 28" C (Group H). Each group was kept in a 68-1 tank with a 14L: 10D light regime. The fish were fed with live brine shrimp and blended calf liver. Every week the fish were checked to see if any of the male fish had reached maturity. If a mature male was noticed, the standard length of the fish was recorded. The experiment was not terminated until all of the male fish had reached maturity.
EXPERIMENT V This experiment was to observe the variation of mating success of a large male in the
presence of various numbers of small males. Prior to this experiment, adult males were isolated in individual 2-1 aquaria for 1 week. In the first test, a large male (S.L. > 25.0 mm) in a 68-1 tank was first allowed to mate a receptive female (a female within 1-8 days of parturition, which has the highest receptivity; Peden, 1973). The mating frequency by the male in a 10-min period was recorded (successful mating was defined as the thrust of gonopodium into female body, followed by an abrupt movement of the male separating from female). In the second test, one large male and one small male (S.L. < 2 5 0 mm) were given a female to compete for mating. The mating frequencies by both males were recorded. In the subsequent five tests, one large male was allowed to compete for matings with two, three, four, five and six small males. The observations for each test had five replicates, but each fish was used only once in the test to avoid possible individual bias in mating frequency. The cumulate mating success made by both large and small males (in 50 min) was then analyzed by the G-statistic (Zar, 1984).
EXPERIMENT VI This experiment was to test the ' role switching ' hypothesis which states that a male can
switch its role as ' dominant ' or ' subordinate ' during mating competition, depending on the relative size of the rival males. Three large males (Ll-L3, mean S.L. = 23.1 mm), three medium males (MI-M3, mean S.L. = 21.2 mm) and three small males (SI-S3, mean S.L. = 19.0 mm) were selected from the holding tank. In Test 1, a large male (LI) was allowed to complete for mating to a receptive female with a medium male (M 1) for a 5-min period. The test was then repeated three times (Tests 2 4 ) with a 1-h interval between successive tests. The large male was then replaced by a small male (Sl) and the procedure for Tests 1 4 was again followed (Tests 5-8). The same protocol was used in sixteen subsequent tests (L2, M2 and S2 were used in Tests 9-16; L3, M3 and S3 in Tests 17-24).
COMPARISON OF MEAN STANDARD LENGTH BETWEEN ADULT AND MATURING MALES
Monthly collections of G. heterochir from its natural habitat were made from July 1983 to June 1984 (see Yan, 1986 for collection procedures). Standard lengths of adult males and

734 H. Y. YAN
TABLE 1. Duncan multiple range test on the mean daily specific growth rates (MDSGR) in young females (G), young males (Y), adult
females (F) and adult males (M)
Sex n MDSGR
G Y F M
13 0.729 1 * 17 0*5490* 14 0.3317* 21 0.056 1 *
*MDSGR values group those means which are signifi- cantly different from each other (P<0.05).
TABLE 11. Dunca_n_multiple range test on mean standard length (S.L.) of adult males (M), young males smaller than adult males (YS), and young
males larger than adult males (YL)
- Male n S.L. ~
M 17 25.61 * YS 10 24.75* YL 7 25.82*
*Means which are not significantly different from each other (P<O.OS) .
maturing males (i.e. male with partially developed gonopodium) were measured to the nearest 0.05mm. Monthly mean standard lengths of adult and maturing males were compared by the /-test.
HI. RESULTS EXPERIMENT I
After 2 months it was found that 13 of the 30 fish were females and the rest were males. One-way ANOVA showed that a significant difference in daily specific growth rates existed among adult males, adult females, young females, and young males ( F = 95.95, P < 0.0001). Duncan's multiple range test revealed four distinc- tive growth rates among the groups (Table I). Post-maturation growth was signifi- cant in adult male fish ( t = 7.82, P<O.OOl) although the mean daily specific growth rate was quite small (0.0561 %). Post-maturation growth in females was significant ( t = 16-73, P<O-OOl) and the rate was about 5.9 times of that of adult males. EXPERIMENT I1
Only seven among 17 young males exhibited maturation sizes larger than the adult males (Table 11). No significant difference in mean standard length among adult males and young males was observed (Table 11). This result is contradictory to the prediction of the ' social control mechanism '.

MATURATION SIZE OF MALE GAMBUSIA HETEROCHIR 735
TABLE 111. Standard length of adult males (M) and mean standard length of young males (Y) in each brood (A-E) of Experiment I11
L S Brood
M N n Y M N n Y t
A 25.10 3 1 24.62 21.40 4 4 24.53 0.12 N.S. 0 26.70 4 0 23.54 21.90 5 5 22.91 1.82 N.S. C 76.65 3 O 25.93 23'50 3 3 26.03 -0.70 N.S. D 24.45 4 0 2360 22.65 4 4 23.74 -0.53 N.S. E 25.50 4 0 21.99 22.30 3 3 22.03 -0.26 N.S.
L. Subgroup with large adult male; S, subgroup with small male; N , sample size of young males in each subgroup: n. number of young males with size larger than adult male; i. i-test value on mean standard length of young males between subgroups; N.S. non-significant.
TABLE IV. Effect of low (20" C) and high (28" C ) temperatures on mean age (in days) and mean standard length at maturity in five broods of male siblings
20" c 28" C Brood
Age S.L. Age s. L. -
1
3 4 5
7
~ ~ _ _ _ _ _
96.6(3) 22.19 56.0(4) 22.42 I I4.3(3) 2445 73.5(4) 24.69 96.3(4) 23.00 57.4(5) 23.07
108.5(4) 20.68 66.5(3) 20.74 126.0(7) 2 1 .oo 56.0(7) 2 1.04
Numbers in parentheses indicate sample size.
EXPERIMENT I11 Among five groups, only one young male in subgroup L of group A grew larger
than the large adult male. All the young males in subgroup S grew larger than the small adults. No significant difference in maturation size between siblings of each group was observed (Table 111).
EXPERIMENT IV Two-way ANOVA revealed (Table IV) that different broods (F= 4.94, P< O.Ol) ,
temperatures ( F = 228.27, P< 0.0001) and the interaction between the two factors (F=4.27, P<O,OI) had significant effects on the age at which the males reached maturity. Temperature had no significant effect on the size of male maturation (F= 0.34. P > 0.5). The size at maturity was significantly different among broods (F= 80.50, P<O.OOOl). The interaction of brood and temperature had no signifi- cant effect on the size of males at maturity (F=0.06, P>0*90).
Based on the results of broods 1-5, under 20" C and 14L:lOD conditions the mean age at maturity in male fish was 108.3 days, while at 28" C it was 61.9 days.
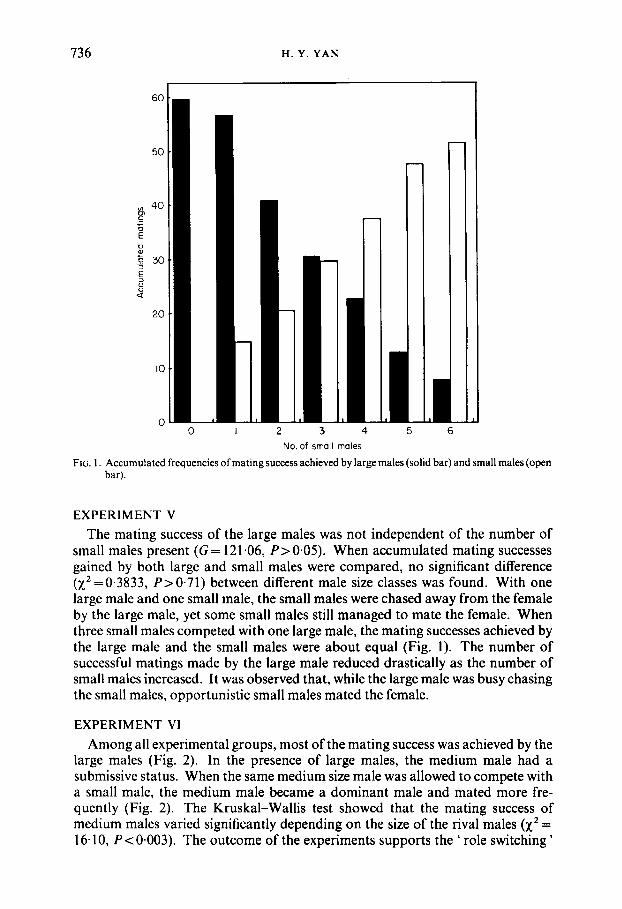
736 H. Y. YAN
60
50
40 0,
0 c E
- 30
+
u 0)
E, 2
20
10
0 1 0 I 2 3
i 4 5 6
No. of smoll moles
FIG. 1. Accumulated frequencies of mating success achieved by large males (solid bar) and small males (open bar).
EXPERIMENT V The mating success of the large males was not independent of the number of
small males present (G = 121.06, P> 0.05). When accumulated mating successes gained by both large and small males were compared, no significant difference (x2 =0.3833, P>0.71) between different male size classes was found. With one large male and one small male, the small males were chased away from the female by the large male, yet some small males still managed to mate the female. When three small males competed with one large male, the mating successes achieved by the large male and the small males were about equal (Fig. 1). The number of successful matings made by the large male reduced drastically as the number of small males increased. It was observed that, while the large male was busy chasing the small males, opportunistic small males mated the female.
EXPERIMENT VI Among all experimental groups, most of the mating success was achieved by the
large males (Fig. 2). In the presence of large males, the medium male had a submissive status. When the same medium size male was allowed to compete with a small male, the medium male became a dominant male and mated more fre- quently (Fig. 2). The Kruskal-Wallis test showed that the mating success of medium males varied significantly depending on the size of the rival males ( x 2 = 16.10, Pc0.003) . The outcome of the experiments supports the ' role switching '

MATURATION SIZE OF MALE GAMBUSIA HETEROCHIR
$ 6 - f 5 .
F 4 .
._ F 3 - p 2 .
r
I
0 -
737
c
c
-
I I 1 I I
81
I I 1
6 -
f 5 -
.- P 3 -
2 2 -
0 e .c
c
I -
0 '
c - - 4-I-
I 1 I I I I I
1 2 3 4 5 6 7 8
8 UI 7 -
6 - s 5 -
5 3 -
?
E .b- 4 - m
2- 2 -
I - -
- 9 l-JJLLL 20 21 22 23 24
Test
FIG. 2. Mating success achieved by large males (L, O) , medium males (M, a) and small males (S, Z) in (a) Tests 1-8; (b) Tests 9-16; (c) Tests 17-24.
hypothesis that a male's status as ' dominant ' or ' subordinate ' depends on the relative size of the rival male.
COMPARISON OF MEAN STANDARD LENGTH BETWEEN ADULT AND MATURING MALES
When the mean standard lengths of adult and maturing males in each monthly sample were compared (Fig. 3) no significant differences were found in the samples of July 1983 ( t = - 1.7951, P>0.08), August 1983 (z= 1.4046, P ~ 0 . 1 6 ) and June 1984 ( t = 0.3339, P>0.74), but, for the remaining nine monthly samples, the mean standard length of adult males was always significantly larger than the maturing males.
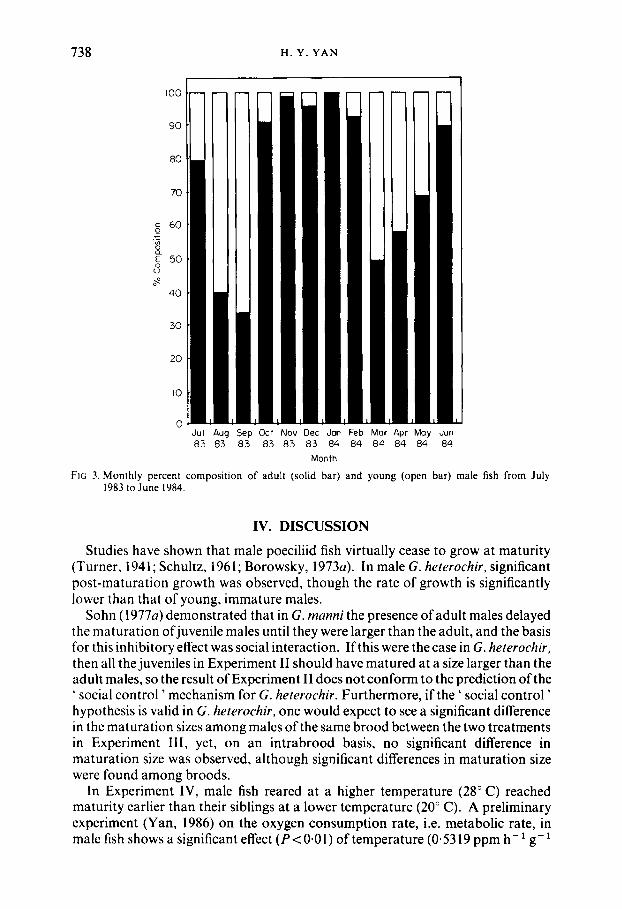
738 H. Y. Y A N
loo 90 In 00 '
70 '
c 60 . 0
s t -
a 50
0
8
Jul Bug Sep Oct Nov Dec Jan Feb Mar Apr May Jun 03 03 03 83 0 3 03 04 04 04 84 04 04
Month
FIG 3. Monthly percent composition of adult (solid bar) and young (open bar) male fish from July I983 to June 1984.
IV. DISCUSSION
Studies have shown that male poeciliid fish virtually cease to grow at maturity (Turner, 1941; Schultz, 1961; Borowsky, 1973~). In male G . heterochir, significant post-maturation growth was observed, though the rate of growth is significantly lower than that of young, immature males.
Sohn ( 1977~) demonstrated that in G. nzanni the presence of adult males delayed the maturation ofjuvenile males until they were larger than the adult, and the basis for this inhibitory effect was social interaction. If this were the case in G. heterochir, then all the juveniles in Experiment I1 should have matured at a size larger than the adult males, so the result of Experiment I1 does not conform to the prediction of the ' social control ' mechanism for G. heterochir. Furthermore, if the ' social control ' hypothesis is valid in G. heterochir, one would expect to see a significant difference in the maturation sizes among males of the same brood between the two treatments in Experiment 111, yet, on an intrabrood basis, no significant difference in maturation size was observed, although significant differences in maturation size were found among broods.
In Experiment IV, male fish reared at a higher temperature (28°C) reached maturity earlier than their siblings at a lower temperature (20" C ) . A preliminary experiment (Yan, 1986) on the oxygen consumption rate, i.e. metabolic rate, in male fish shows a significant effect (P<O.Ol) of temperature (0.5319 ppm h- g-

MATURATION SIZE OF MALE GAMBUSIA HETEROCHIR 739
at 28" C Y . 0-4098 ppm h- ' g- at 20" C) which may explain the positive effect of higher temperature on the younger age at maturity. It also indicates that the size at maturity is independent of physical environmental influence in male G . heterochir. Pitt (1975) reported that in plaice from the Grand Bank faster-growing fish matured earlier than, but at the same size as, slower-growing fish. Grainger (1953) described the same pattern in Artic char. Scott (1962), Bagenal (1969) and Wootton (1973) all reported decreased age at maturity in rainbow trout, brown trout and sticklebacks as food supply increased and their fish grew faster. Signifi- cant difference in maturation size of male G. heterochir among broods further suggests that genetic variation plays an important role. In male G. heterochir, on an intrabrood basis, maturation depends upon size rather than age, corroborating investigations with molluscs (Cammens, 1967), crustaceans (Lasenby & Langford, 1972), and fish (Sohn & Crews, 1977; Stearns, 1983). However, age at maturity in G . heterochir varied significantly among broods under different environmental treatments, suggesting that the age at maturity is influenced by variation in both genes and environment. Whether fish mature at a fixed size or at a fixed age has long been argued in the literature. Stearns & Crandall(l984) in their ' optimizable variation model ' suggested that age at maturity in fish is determined both by genes and by environment. The plasticity for age at sexual maturity observed in male G . heterochir in this study corroborates Stearns & Crandall's prediction. Further- more, four qualitatively different types of age-size maturation trajectories were predicted by the model. The trajectories revealed that organisms mature at neither a fixed size nor a fixed age: over some parts of such trajectories organisms appear to mature at a fixed size, whereas over other parts they appear to mature at a fixed age. Stearns & Crandall (1 984) suggested that only by considering maturation events occurring across a wide range of growth rates can the shape of the whole trajectory be perceived. Since only small ranges of growth rate based on two temperature treatments were addressed, the indication that male G. heterochir mature at a fixed size may represent only a small portion of the trajectories predicted by the optimizable variation model.
Based on collections of G. afinis in Indiana, Hughes (1985) reported that, in August samples, maturing males are abundant relative to adult males. In Clear Creek gambusia, the same pattern was observed in August and September samples (Fig. 3). Hughes (1985) claimed that his data suggested that many males were delaying maturation, and he believed that delayed maturation was under social control in G . ufinis. I contend that Hughes' interpretation of the data is misleading and that August is the time when young-of-the-year reach sexual maturity. In G . hererochir, maturing males in the early spawning season (February, March and April) are derived from the over-wintering juvenile cohort of the previous year. A high percentage of maturing males in the late spawning season (August, September) simply reflects that young-of-the-year are reaching maturity. The observation that the mean age at maturity at 20" C (the ambient water temperature of spring heads of Clear Creek) is 108 days (Table IV) further supports this argu- ment. The observed higher percentage of maturing males in the samples of August and September should not be interpreted as a process of delayed maturation under the control of social factors.
Hughes (1985) further reported that when two males differing in size are placed with a female, the larger is generally able to monopolize access to her. The same

740 H. Y. Y A N
phenomenon was observed in this study. However, his large v. small male exper- iment does not address the question as to how small size morphs are maintained in the population if mating is monopolized by the large males. The results in Experiment V show that the mating success of the large male declines as the number of rival small males increases. My field observations on courtship behaviour of male fish reveal that one large male and one small male rarely compete one on one for a female; generally, a receptive female in the wild is pursued by a group of males of various sizes. Multiple insemination occurs in Gambusia afirzis (Chesser et al., 1984), Poecilia reliculatus (Haskin et al., 1961), Poeciliopsis niorzacha (Leslie & Vrijenhoek, 1977) and Xiphophorus spp. (Borowsky & Kallman, 1976). demonstrated by the frequencies of enzyme phenotypes in mother-offspring combinations, and indicating that fertilization is not controlled by a single male.
Male G . heterochir can switch roles as ' dominant ' or ' subordinate ', depending on the relative size of the rival male. Subsequently the mating tactic may be contingent upon the status of other males in close proximity.
1 appreciate the guidance given by my supervising professor, Dr Clark Hubbs. Logistic support was provided by Department of Zoology, The University of Texas at Austin. Financial support for this research was provided in part by Office of Graduate Studies, The University of Texas at Austin and Sigma-Xi Grants-in-Aid award. Dr Judith C. Lang and an anonymous reviewer offered valuable suggestions to an earlier draft of manuscript.
References
Assem, J. van den, (1967). Territory in the three-spined stickleback Gasterosteus aculeatus L. Behaviour suppl. 16, 164 pp.
Bagenal, T. B. (1969). The relationship between food supply and fecundity in brown trout Salnzo trutta L. J . Fish Biol. 1, 167-182.
Borowsky, R. (1973~). Social control ofadult size in males of Xiphophorus variatus. Nature Lond. 245,332-335.
Borowsky, R. (19736). Relative size and the development of fish coloration in Xiphophorus variatus. Physiol. Zool. 46,22-28.
Borowsky, R. & Kallman, K. D. (1976). Patterns of mating in natural populations of Xiphophorus from Belize and Mexico. Evolution 30,693-706.
Cammens, J. J. (1967). Growth and reproduction in a tidal flat population of Macoma halthica. Neth. J . Sea Res. 3, 3-34.
Chesser, R. K. , Smith, M . W. &Smith, M. H. (1984). Biochemical genetics ofmosquitofish 111. Incidence and significance of multiple insemination. Geneticu 64,77-81.
Constantz, G . D. (1975). Behavioral ecology of mating in the male Gila topminnow, Poeciliopsis occidentalis (Cyprinodontiformes; Poeciliidae). Ecology 56,966973.
Daly, M . & Wilson, M. (1983). Sex, Evoluiion and Behavior. 2nd ed. Boston: Willard Grant Press.
Gandolfi, G. (1971). Sexual selection in relation to the social status of males in Poecilia reticulata (Te1eostei:Poeciliidae). Bull. Zool. 38, 3 W 8 .
Grainger, E. H. (1953). On the age, growth, migration, reproductive potential and feeding habits of the Arctic char (Salvelinus alpinus) of Frobisher Bay, Baffin Island. J . Fish Res. Bd Can. 10,326-370.
Haskin, C. P., Haskin, E. F., McLaughlin, J. J. A. & Hewitt, R. E. (1961). Polymorphism and population structure in Lebistes reticulatus (an ecological study). In Vertebrate Speciation (W. F. Blair, ed.). Austin: University Texas Press.
Hughes, A. L. (1985). Seasonal trends in body size of adult male mosquitofish, Gambusia ofinis, with evidence for their social control. Env. Biol. Fish. 14,251-258.
Hubbs, C. (1957). Gambusia heterochir, a new poeciliid fish from Texas, with an account of its hybridization with G . afinis. Tulane Stud. Zool. 5, 1-16.

MATURATION SIZE OF M A L E GAMBUSIA HETEROCHIR 74 1
Hubbs, C. (1971). Competition and isolation mechanisms in the Gambusia aflnis x G . heterochir hybrid swarm. Bull. Texas Mem. Mus. 19, 1 4 6 .
Lasenby, D. C. & Langford, R. R. (1972). Growth, life history and reproduction of Mysis relicta in an Arctic and temperate lake. J. Fish. Res. Bd Can. 29, 1701-1708.
Leslie, J. F. & Vrijenhoek, R. C. (1977). Genetic analysis of natural populations of Poeci- liopsismonacha. Allozyme inheritance and pattern ofmating. J. Hered. 68,301-306.
Peden, A. E. (1973). Variation in anal spot expression of gambusiine females and its effect on male courtship. Copeia 1973,250-263.
Pitt, T. K. (1975). Changes in abundance and certain biological characteristics of Grand Bank American plaice (Hippoglossoidesplatessoides). J. Fish. Res. Bd Can. 32,1383- 1398.
Robel, R. J. (1 966). Booming territory size and mating success of the greater prairie chicken (Tympanuchus cupidopinnatus). Anim. Behav. 14,328-33 1.
Schultz, R. J . (1961). Reproductive mechanism of unisexual and bisexual strains of the viviparous fish Poeciliopsis. Evolution 15,302-325.
Scott, D. P. ( 1 962). Effect of food quantity on fecundity of rainbow trout Salmo gairdneri. J. Fish. Res. Bd Can. 19,715-731.
Sohn, J . J . (1977~). The consequences of predation and competition upon the demography of Gambusia manni (Pisces: Poeciliidae). Copeia 1977,224-227.
Sohn, J . J. (1977b). Socially induced inhibition of genetically determined maturation in the platyfish, Xiphophorus maculatus. Science 195,199-20 1.
Sohn, J. J. & Crews, D. ( 1 977). Size-mediated onset of genetically determined maturation in the platy fish, Xiphophorus maculatus. Proc. natn. Acad. Sci. U.S.A. 74,45474548.
Stearns, S. C. (1983). The evolution of life-history traits in mosquitofish since their intro- duction to Hawaii in 1905: rates of evolution, heritabilities, and developmental plasticity. Am. Zool. 23,6>76.
Stearns, S. C. & Crandall, R. E. (1984). Plasticity for age and size at sexual maturity: a life- history response to unavoidable stress. In Fish Reproduction: Strategies and Tactics (G. W. Potts and R. J. Wootton, eds). London: Academic Press.
Turner, C. L. (1941). Morphogenesis of the gonopodium in Gambusia aflnis a#nis. J . Morph. 69, 161-185.
Weatherley. A. H. (1972). Growth and Ecology of Fish Populations. London: Academic Press.
Wootton, R. J. (1973). The effect of food ration on egg production in the female three- spined stickleback, Gasterosteus aculeatus L. J . Fish Biol. 5,89-96.
Yan, H. Y. (1986). Reproductive strategies of the Clear Creek gambusia, Gambusia hetero- chir. Dissert, The University of Texas at Austin. 178 pp.
Zar, J. H. (1984). Biostatistical Analysis, 2nd ed. Englewood Cliffs: Prentice-Hall. Zimmerman, J. L. (1966). Polygamy in the dickcissel. Auk 83,534-546.
Copyright © 2022 FDOKUMEN




















