
Key aspects of the biology of snail-killing Sciomyzidae flies
Upload
independentCategory
view
0download
0
Biological Journal of the Linnean Society (1997), 61: 267–279. With 3 figures
Growth rate differences between upper andlower shore ecotypes of the marine snailLittorina saxatilis (Olivi) (Gastropoda)
KERSTIN JOHANNESSON1, EMILIO ROLAN-ALVAREZ2 ANDJOHAN ERLANDSSON1
1Tjarno Marine Biological Laboratory, S-452 96 Stromstad, Sweden, 2Departamento deBiologıa Fundamental (Genetica), Universidad de Vigo, 36202 Vigo, Spain
Received 7 June 1996; accepted for publication 9 August 1996
Shell polymorphisms are widespread among those intertidal gastropods that lack a pelagicspreading stage. These polymorphisms may indicate diversifying selection in a heterogeneoushabitat, but to do this the variation must be at least partly inherited. Galician populationsof Littorina saxatilis (Olivi) living in exposed rocky shores are highly polymorphic in severalshell traits, e.g. ornamentation, banding and size. Mature snails of the upper-shore ridgedand banded (RB) morph is, for example, often twice as large as mature individuals of thelower-shore smooth and unbanded (SU) morph of the same shore.
We investigated the hypothesis that lower-shore snails grow more slowly and that differencesin growth rate were at least partly inherited and could be explained by diversifying selection.We released snails of different origin (upper, mid- and lower shore) and morph (RB, SUand hybrids) at different shore levels and compared their shell increment after one monthof growth. We found that despite considerable variation among individuals and amongreplicate samples (together about 53% of the total variation), average rates of growth differedbetween morphs. RB snails both from the upper and mid-shores grew at a high rate at allshore levels, SU snails grew considerably less, and hybrids grew at intermediate rates, at alllevels. Inherited difference among morphs explained about 34% of the total variation whileeffects of shore levels and the interaction morph × shore level explained only 5 and 7%,respectively. Thus a large part of the difference in growth rate leading to different adult sizesof the two morphs has probably evolved due to spatially varying selection favouring largesizes in upper-shore and small sizes in lower-shore environments.
1997 The Linnean Society of London
ADDITIONAL KEY WORDS:—shell increment – genetic differences – selection gradient– size differences – intraspecific variation – Galicia.
CONTENTS
Introduction . . . . . . . . . . . . . . . . . . . . . . . 268Material and methods . . . . . . . . . . . . . . . . . . . 269
Statistics . . . . . . . . . . . . . . . . . . . . . . 271Results . . . . . . . . . . . . . . . . . . . . . . . . 272
Correspondence to: Kerstin Johannesson. e-mail: [email protected]
2670024–4066/97/060267+13 $25.00/0/bj960122 1997 The Linnean Society of London

K. JOHANNESSON ET AL.268
Discussion . . . . . . . . . . . . . . . . . . . . . . . 276Acknowledgements . . . . . . . . . . . . . . . . . . . . 278References . . . . . . . . . . . . . . . . . . . . . . . 278
INTRODUCTION
Average adult shell size varies both within and between marine gastropod species.One pattern that seems to be common is that wave-exposed shores house smallergastropods than protected shores (Boulding & Van Alstyne, 1993), although this isnot invariably so (Reimchen, 1982). Part of the variation between individuals andpopulations is non-genetic. Poor food availability in dense populations, low ambienttemperatures, or other adverse physical conditions impede growth (e.g. Janson,1982; Boulding, Buckland-Nicks & Van Alstyne, 1993; Kemp & Bertness, 1984;Chow, 1987). However, growth rate and size may also be genetically and eco-phenotypically constrained. Differences between populations and individuals maybe due to either different genes in different populations controlling growth rate(genetic differences), or the same set of genes interacting differently with differentenvironments yielding different phenotypic responses (ecophenotypic differenceswithin the reaction norm of a certain genotype). It is essential to discriminatebetween environmentally-induced and heritable size differences between individuals.Genetic differences may result in population evolution while non-genetic variation,except adaptive plasticity, may not.
Differences in size both between adult individuals within a population and betweenpopulations, may have broad implications. For example, large snails are lessvulnerable to crushing by crabs (Elner & Raffaelli, 1980; Johannesson, 1986; BehrensYamada & Boulding, 1996). Small snails may, on the other hand, be favoured inwave exposed areas, where crabs are lacking, since small snails more easily findrefuges in crevices and cracks (Raffaelli & Hughes, 1978). Small individuals are alsoless physically affected by mechanical drag forces which may dislodge snails (Denny,Daniel & Koehl, 1985). In addition, large females are much more fecund than smallfemales ( Janson, 1985). More subtle, but still important effects of size may be relatedto mate choice strategies. For example, size assortative mating has been observedin populations of Littorina saxatilis (Olivi) (Saur, 1990; Erlandsson, 1996), and sexualselection on female size was found in L. littorea (L.) (Erlandsson & Johannesson,1994).
Like most marine invertebrates, snails have indeterminate growth, and usuallygrowth rate decreases with age (Hughes, 1980; Janson, 1982). Size differencesbetween snails of similar ages can only be due to different growth rates, while sizedifferences between populations may be due to different age-specific survival rates.
Growth rates have been measured in populations of planktonic dispersers andthe intraspecific differences found between populations are often suggested to benon-genetic due to an assumed high rate of genetic exchange (Kemp & Bertness,1984; Chow, 1987; Brown & Quinn, 1988, although see Struchsaker, 1968 for anexception).
Profound size differences are often present between populations of direct developersliving in different shore types (Reimchen, 1982; Sundberg, 1988; Etter, 1989;Boulding, 1990; Reid, 1996) or even on the same shore but in microhabitats ofdifferent types (Reimchen, 1981; Johannesson & Johannesson, 1990; Johannesson,

INTRASPECIFIC DIFFERENCES IN GROWTH OF LITTORINA 269
Johannesson & Rolan-Alvarez, 1993). Transplanted and laboratory raised popu-lations of poor dispersers indicate that at least some of this size variation is genetic( Janson, 1982; Behrens Yamada, 1987; Brown & Quinn, 1988; Johannesson &Johannesson, 1996).
Populations of the direct developing species Littorina saxatilis from wave exposedrocky areas of Galicia are strongly polymorphic for a number of characters, includingsize. Upper- and lower-shore snails differ in ornamentation, colour and size of adultshells ( Johannesson et al., 1993), embryo and radula characters (Rolan-Alvarez,Rolan & Johannesson, 1996), mating behaviours ( Johannesson, Rolan-Alvarez &Ekendahl, 1995), and tolerance of different types of physical stresses (Rolan-Alvarez,Johannesson & Erlandsson, 1997). Some differences, for example, in ornamentationand banding, are inherited ( Johannesson et al., 1993), and the morphs may beconsidered different ecotypes while living only 10 meters, or less, apart. The sizedifferences are profound, upper shore snails being on average 30–260% larger thanlower shore snails ( Johannesson et al., 1993). The upper shore ecotype has ridgesand bands (‘RB-morph’) and is easily distinguished from the lower shore ecotypewhich is smooth and unbanded (‘SU-morph’). The morphs overlap over a fewmeters of mid-shore zone where morphologically intermediate forms (‘mixed’ or‘hybrid’ morphs) are present (for details of distributions see Johannesson et al., 1993).
In this study we address the questions of whether size differences between theGalician ecotypes are due to differences in growth rate, and whether or not thesedifferences are in part inherited. Inherited differences would suggest that the differentsizes of the two ecotypes have evolved as adaptations to differential selection favouringlarge snails in the upper shore and small snails in the lower shore.
MATERIAL AND METHODS
The Atlantic coast of Galicia (NW Spain) has a tidal range of between 1 m (neaptide) and 4 m (spring tide) but the actual range of the littoral zone is much wider.Heavy wave action in combination with gently sloping rocks produces a splash zonetens of meters wide. The rocks are granite, mostly characterized by a complexmicrotopography of cracks and crevices. Littorina saxatilis is one of the main benthicgrazers of these rocks and occupies a wide range of the eulittoral and littoral fringezones, often at densities of hundreds per square meter. The eulittoral is characterizedby a zone of Mytilus galloprovincialis while in the littoral fringe the barnacle speciesChthamalus stellatus dominates. Thus L. saxatilis lives mainly in two types of habitats,the upper (barnacle) zone, and the lower (mussel) zone. In the mid shore, thebarnacles and mussels intergrade, forming a mosaic of barnacle and mussel patches.
In the upper shore L. saxatilis has shell ridges and dark continuous bands in thegrooves, while in the lower shore snails lack both these characters ( Johannesson etal., 1993). At mid-shore levels both upper and lower morphs are present as well asmorphologically intermediate snails (hybrids), snails with either ridges or bands butnot both. The group of intermediates probably contains true F1 hybrids but mayalso contain introgressed individuals. We have no evidence of F1 sterility, and itseems likely from estimates of gene flow ( Johannesson et al., 1993) and matingbehaviour ( Johannesson et al., 1995; Rolan-Alvarez, Rolan & Johannesson, 1996)that back-crossing and introgression occur frequently.
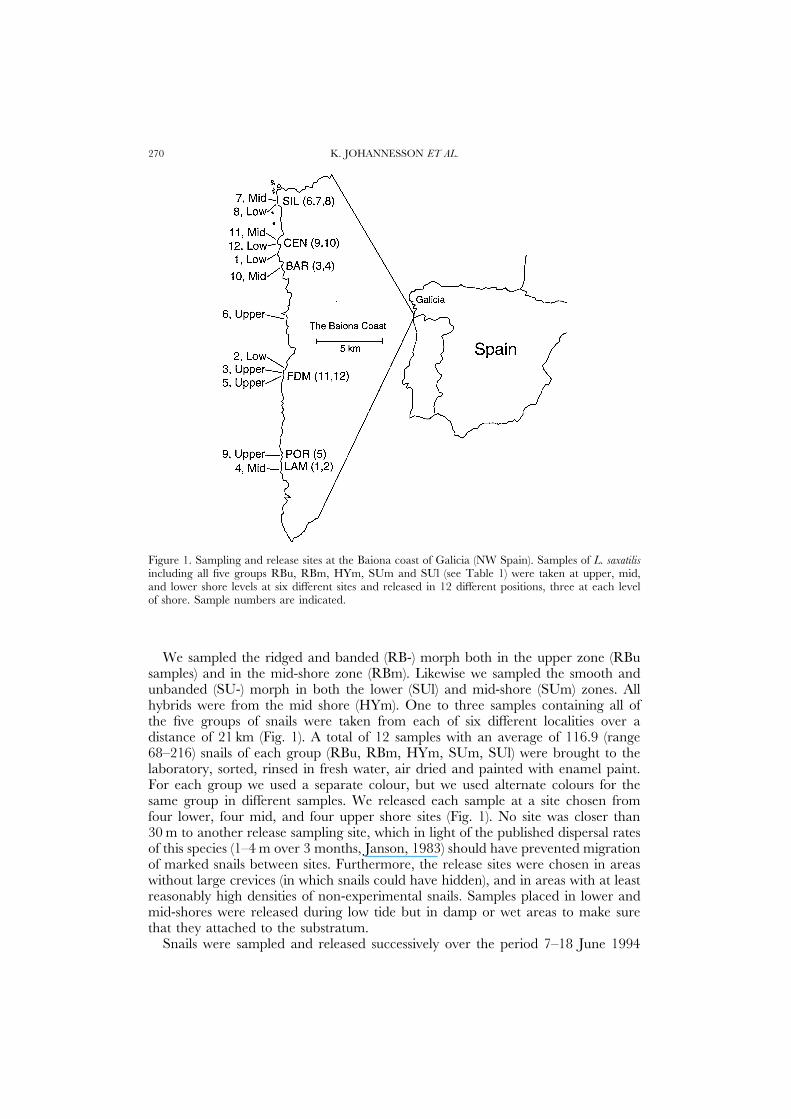
K. JOHANNESSON ET AL.270
Figure 1. Sampling and release sites at the Baiona coast of Galicia (NW Spain). Samples of L. saxatilisincluding all five groups RBu, RBm, HYm, SUm and SUl (see Table 1) were taken at upper, mid,and lower shore levels at six different sites and released in 12 different positions, three at each levelof shore. Sample numbers are indicated.
We sampled the ridged and banded (RB-) morph both in the upper zone (RBusamples) and in the mid-shore zone (RBm). Likewise we sampled the smooth andunbanded (SU-) morph in both the lower (SUl) and mid-shore (SUm) zones. Allhybrids were from the mid shore (HYm). One to three samples containing all ofthe five groups of snails were taken from each of six different localities over adistance of 21 km (Fig. 1). A total of 12 samples with an average of 116.9 (range68–216) snails of each group (RBu, RBm, HYm, SUm, SUl) were brought to thelaboratory, sorted, rinsed in fresh water, air dried and painted with enamel paint.For each group we used a separate colour, but we used alternate colours for thesame group in different samples. We released each sample at a site chosen fromfour lower, four mid, and four upper shore sites (Fig. 1). No site was closer than30 m to another release sampling site, which in light of the published dispersal ratesof this species (1–4 m over 3 months, Janson, 1983) should have prevented migrationof marked snails between sites. Furthermore, the release sites were chosen in areaswithout large crevices (in which snails could have hidden), and in areas with at leastreasonably high densities of non-experimental snails. Samples placed in lower andmid-shores were released during low tide but in damp or wet areas to make surethat they attached to the substratum.
Snails were sampled and released successively over the period 7–18 June 1994

INTRASPECIFIC DIFFERENCES IN GROWTH OF LITTORINA 271
so that no snails were more than 3 days (average 1.5 day) in the laboratory. Nosnails died during the laboratory treatments.
Twenty-seven to 35 days after the release at a particular site, surviving snailswere recaptured. We searched for snails during two successive days in each site,and in an approximately circular area (15 m radius) around the release spot. Of atotal of 8419 released snails, 1625 were recaptured. However, 85 of these had lostso much of the colour markings that the shell increment could not be conclusivelydetermined, leaving a total of 1540 snails which were measured. (Relative rates ofsurvival and dispersal among morphs and sites are described and analysed elsewhere,Rolan-Alvarez, Johannesson & Erlandsson, 1997 and Erlandsson, Johannesson &Rolan-Alvarez, subm.)
The outer lip of each shell had been painted with a 1–2 mm wide band indicatingthe size of the shell on the day of release.
During growth the snail added new (unpainted) shell, and shell increment wasmeasured as the width of the unpainted shell band. The actual period of growthdiffered, with up to 8 days between samples, but shell increment was standardizedto mm per month (30 days). Final shell length as well as shell length at the time ofrelease was recorded and mean size was taken as the average of these estimates.
Statistics
When comparing average growth rates for groups of snails the possibility of theirdependending on size must be considered. In general, but not invariably, absolutegrowth in Littorina is negatively correlated with size of an individual (e.g. Janson,1983; Roberts & Hughes, 1980). In our study 10 of 15 pooled samples revealednegative slopes of growth on size while ten indicated non-significant slopes. Breakingdown the total variation into components of variation would have required ananalysis of covariance with size being the covariate. However, heterogeneous slopes(P<0.05) of the different treatments prevented further analysis of the variation usingANCOVA. Taking size into account we instead compared growth rates betweenpairs of groups using a non-parametric method suggested by Tsutakawa & Hewett(1977). We plotted the increments against size of both groups in the same graph,and calculated the reduced major axis regression (Ricker, 1973) common to bothgroups. We used this type of Model II regression to describe growth in each sampleas this model recognizes that both variables are measured with error (Sokal & Rohlf,1981: 460). We then compared the number of individuals of each group above andbelow the common line with a 2×2 contingency table chi-square test. We correctedthe obtained probability values for multiple testing using Sidak’s multiplicativeinequality (see Sokal & Rohlf, 1981, p. 728 and Rohlf & Sokal, 1981, table 15)using the programme MULTTEST (Zaykin & Pudovkin, 1991).
The Tsutakawa & Hewett method, however, permitted only qualitative conclusionsto be made, and as we were also interested in quantitative estimates of the importanceof, for example, genetic and environmental components of variation, we performedan analysis of variation (ANOVA) with two orthogonal factors (GROUP: RBu,RBm, HYm, SUm, SUl, and SHORE LEVEL: upper, mid, and lower). We balancedthe data set by random exclusion of data to n=34 in all cells and obtained non-heterogeneous variances at a=0.01 level by square-root transformation (x′=8x+1).In this way the size effects of the variation were included in the residual variation,

K. JOHANNESSON ET AL.272
Figure 2. Shell increment over 30 days (one month) as a function of mean size in L. saxatilis. The toprow shows variation within and among replicate samples of the RBu morph translocated to four upper-shore sites. The bottom row shows three samples of the SUl morph released in three low-shore sites.(The fourth sample included less than five snails and was ignored.) Model II linear reagression linesare indicated.
but this proved to be a minor problem as the differences found between groupswere not generated by differences in average size (see below).
RESULTS
Growth rate, as indicated by shell increment over one month, varied substantiallybetween individuals of the same morph and original locality released at the samesite and at the same time. In general, large snails grew at a lower rate than-smallsnails, but most of the within-sample variation was due to variation betweenindividuals of similar sizes. In fact, rate of shell increment varied two-fold or moreamong similarly sized snails of identical treatments (Fig. 2). Growth rate sometimesdiffered between replicate samples. For example, the four samples of RBu snails,which were all from different upper-shore localities and released in different upper-shore sites, grew at different rates (Fig. 2; all pairwise comparisons of RBu samplesexcept Site 3 vs Site 5 are significantly different, P<0.05). The three SUl samplesreleased on lower shores were, however, not different, but this may at least partlybe due to small sample sizes (Fig. 2, P>0.05). The four RBu samples released inupper shores and the three SUl samples released in low shores represent merelytwo of 15 sets of pooled samples (see Fig. 3). Although there may be variationamong replicate samples in several of the 15 sets, we did not analyse further sets,as this variation may be due to a combination of genetic and environmental effectsand is not easily interpreted.
It was evident that there was both environmentally-induced variation (withingroups at differing shore levels) and inherited variation (at shore levels betweengroups). Disregarding the effect of size, it became evident that both these factorscontributed significantly to the total variation, as did the interaction between them

INTRASPECIFIC DIFFERENCES IN GROWTH OF LITTORINA 273
Figure 3. Shell increment over 30 days (one month) as a function of mean size in L. saxatilis. Growthrate of each of the five groups RBu, RBm, HYm, SUm, and SUl (see Table 1) at three different shorelevels are shown. Data is pooled from four replicate samples. Model II linear reagression lines areindicated.
(Table 1). One possibility would have been that the variation between groups wasdue to different average sizes of the groups. However, while growth rate declinedwith size in each group, the groups that grew most rapidly were the RB groups,which revealed a larger average size than hybrids and SU groups (Table 2). Thusthe ANOVA gave a conservative estimate of group difference. The estimated sizespermitted a quantitative conclusion of how much each individual factor and theinteraction between them contributed to the total variation in growth rate. Thus53% of the total variation was individual variation including effects of different sizesand replicate samples. Minor parts of the variation were attributable to differencesamong shore levels (5.6%) and the interaction of group and shore level (7%). Arelatively large part of the total variation (34%) was, however, explained by thesnails being of different groups (RBu, RBm, HYm, SUm and SUl).
We evaluated the differences in growth rate among groups and shore levels in

K. JOHANNESSON ET AL.274
T 1. Balanced (n=34) orthogonal two factor analysis of variance of mean growth rates (square-root transformed) of different groups of Littorina saxatilis (RBu – ridged and banded morph from uppershore, RBm – ridged and banded morph from mid shore, HYm – hybrids from mid shore, SUm –smooth and unbanded morph from mid shore, and SUl – smooth and unbanded morph from lowershore). Each group grew at three different shore levels (upper shore, mid shore and lower shore). Theeffect size indicate the contribution of each factor to the total variance. In this analysis we have notconsidered that growth rate changes with size of snail and among replicate sites (see text), both these
components of variation are thus included in the residual
Source of variation df Mean square F-value P-value Effect size
Group 4 13.038 66.68 0.0001 0.1259Shore level 2 3.680 18.82 0.0001 0.0205Group × Shore level 8 1.066 5.45 0.0001 0.0256Residual 495 0.196 0.1955
T 2. Means of growth rate and size for five groups of Littorina saxatilis (see Table 1 for abbreviations)grown at three different shore levels. The table covers the balanced set of data (n=34) used in the
analysis of variance (see Table 1), although here we give untransformed growth rates only
Upper shore Mid shore Lower shoreGrowth Size Growth Size Growth Size
(mm/mo) (mm) (mm/mo) (mm) (mm/mo) (mm)
RBu 2.93 6.22 1.62 6.73 2.32 5.54RBm 3.01 5.95 2.12 6.03 1.53 5.74HYm 1.77 5.73 1.65 5.67 0.71 5.45SUm 0.55 5.30 0.62 4.72 0.64 5.21SUl 0.56 5.86 0.70 4.62 0.42 5.33
T 3. Test of differences in growth rate between groups of L. saxatilis grown in upper and lowershore sites (see Table 1 for abbreviations). Differences between groups were evaluated both by theTsutakawa & Hewett’s (1977) chi-square method (see text) which takes size effects into account, andby the Student-Newman-Keul procedure in which size effects were not considered. In the SNK test
we used a balanced sub-set of square-root transformed data
Group NUPPER NLOWER v2 P1 NSNK PSNK
RBu 347 81 43.8 <0.001 34 n.sRBm 188 99 35.6 <0.001 34 <0.01HYm 45 51 18.1 <0.001 34 <0.01SUm 54 53 0.46 n.s. 34 n.s.SUl 34 60 3.51 n.s. 34 n.s.
1 Corrected for multiple testing by the programme MULTTEST (Zaykin & Pudovkin, 1991).
more detail both taking size effects into account (Tsutakawa & Hewett’s method)and in a more conservative way without considering size differences (Student-Newman-Keul test). Both approaches were in general consistent, although the SNKtest was, as expected, slightly more conservative in a few cases. Environmentally-induced variation was most evident among the RB morph and the hybrids. Bothupper and mid-shore ridged and banded snails, as well as the hybrids, grew onaverage faster in the upper than in the lower shore (Table 3). The smooth and

INTRASPECIFIC DIFFERENCES IN GROWTH OF LITTORINA 275
T 4. Tests of differences in growth rate between groups of L. saxatilis (see Table 1 for abbreviations).Each group grew at three different shore levels (upper, mid and lower) in four replicate sites. Differencesbetween groups were evaluated both by the Tsutakawa & Hewett’s (1977) chi-square method (see text)which takes size effects into account, and by the Student-Newman-Keul procedure in which size effectswere not considered. In the SNK test we used a balanced sub-set of square-root transformed data(n=34). ‘0’ indicates non-significant difference between groups, ‘+’ is P < 0.01 for the Tsutakawa &Hewett test corrected for multiple testing, and ‘#’ is P<0.01 for the SNK test. We only considered
P<0.01 being significant for the SNK test as the variances were homogeneous at a=0.01 only
Upper shore sitesRBu RBm HYm SUm
RBm +1/0HYm +/# +/#SUm +/# +/# +/#SUl +/# +/# +/# 0/0
Mid shore sitesRBu RBm HYm SUm
RBm 0/0 –HYm +/0 +/0 –SUm +/# +/# +/# –SUl +/# +/# +/# 0/0
Lower shore sitesRBu RBm HYm SUm
RBm 0/0 –HYm +/# +/# –SUm +/# +/# 0/0 –SUl +/# +/# 0/0 0/0
1 The highly significant difference between RBu and RBm in the upper shore became non-significant if snails8 mm and larger were excluded from the samples. (See text for further discussion.)
unbanded morph from mid-and lower shore, on the other hand, grew at similarrates at all shore levels (Table 3, Fig. 3).
The inherited part of the variation indicated by differences among groups seemedto be mostly due to differences among the RB ecotype, the SU ecotype, and thehybrids. With one exception we found no difference between RBu and RBm, orbetween SUm and SUl at any shore level (Table 4). The exception was the differencebetween RBu and RBm in the upper shore. This seemed, however, to be an artifactas a bias of large snails in the RBu sample tilted the regression line considerablyand caused significant differences between the two groups. After removal of all largesnails (>8 mm) the difference disappered completely (Table 4). Thus irrespective ofat which shore level they grew, the RB ecotype grew faster than the hybrids, andmuch faster than the SU ecotype, while the hybrids grew faster than the SU ecotypein upper and mid-shores (Fig. 3, Table 4). In addition to the presented analysis weused the Tsutakawa & Hewett method to test data from each of the four replicatesites at each shore level. These tests confirmed the results of the pooled tests.Furthermore, in the unpooled tests of RBu against RBm, none of the four comparisonswere significant, despite sample sizes of the smallest sample of the pair being mostlylarge (n= 60, 63, 16, 49). This supports the conclusion that the significant differenceof the pooled test was an artefact.

K. JOHANNESSON ET AL.276
DISCUSSION
The Galician populations of Littorina saxatilis revealed considerable variation ingrowth rate caused by several factors. Large parts of the total variation wereattributable to variation among individuals of the same group and replicate sample(Fig. 2). This supports earlier findings by Janson (1982) and Roberts & Hughes(1980) that individual variation is very large. Why this is so is not at all clear.Possibly, individuals have temporally variable growth, with asynchronized bursts ofintensive growth. A minor part of the difference between individuals may beexplained by differences between male and female growth rates since female L.saxatilis from Swedish populations grow at 10% higher average rates than do males( Johannesson & Johannesson, 1996). Part of the differences in increment betweenindividuals may be due to microscale differences in habitat which affect growth.Chapman (1994) describes patchy variation in size structure among populations ofthe planktonic developer Nodilittorina unifasciata in Australia. She reports part of thisvariation to be strongly density dependent.
Among organisms with indeterminate growth, there is usually a negative re-lationship between size and growth rate. In our samples ten out of 15 pooledregressions (one for each combination of morph and habitat) revealed a negativeslope, while five showed non-significant slopes. Hughes (1980) and Janson (1982)found that sometimes the relationship between size and growth in L. saxatilis maybe reversed, while in a related species, L. fabalis (=L. mariae see Reid, 1996) growthin size ceases rapidly when snails become sexually mature (Tatarenkov & Johannesson,in prep.). Possibly age, sex, sexual status (mature or immature), micro-niche position,snail density as well as genotype and additional factors, determined the actual growthrate of individual snails. Moreover, we cannot exclude the possibility that individualvariation in growth rate reflects differences in recovery from the lab treatment.
Variation among replicate samples of the same group of snails released at differentshore levels suggests a small component of environmentally-induced variation.Individuals of RB ecotype and hybrids grew, for example, at somewhat higher ratesin upper than in lower shore sites. As we lost a high number of individuals in thelow shore sites (88%), we cannot, however, exclude the possibility of size-relatedselection. For example, fast growing snails may have been at a disadvantage andtherefore lost at a higher rate than moderately or slow growing snails in the lowshore. It is, furthermore, impossible to interpret the variation between replicatesamples as purely non-genetic, as the samples were both originally from, as well asreleased in, different sites and the variation may thus be due to both genetic andnon-genetic components.
Growth rate of the RB ecotype and the hybrid changed with shore level, whilein the SU ecotype it did not (Table 3). This interaction explaining about 7% of thetotal variation seems likely to be an effect of inherited differences between themorphs.
A large part of the total variation (34%) was caused by differences between thefive experimental groups. However, as RBu and RBm snails, on the one hand, andSUm and SUl snails, on the other, grew at similar rates at all three shore levels thedifferences were between morphs rather than groups. From this we expected thehybrids to reveal intermediate levels of growth and this was also mostly true. ThusRB snails grew on average faster than the other snails in both upper, mid- andlower-shore sites while SU snails grew slower than both hybrids and RB forms.

INTRASPECIFIC DIFFERENCES IN GROWTH OF LITTORINA 277
The variation in growth among the morphs may be purely inherited. Alternativelythe snails were affected in some way by their native environment before they weretransplanted. For example, the scent of crabs induces higher growth rate in speciesof Nucella (Appleton & Palmer, 1988; Palmer, 1990). If habitat-triggering is the mainexplanation for growth rate differences among morphs of Galician L. saxatilis weexpect different morphs taken from the same shore level (e.g. RBm, HYm andSUm) to grow at similar rates when released at the same shore level. However, thiswas not the case (Table 4, Fig. 3). Moreover, we would not expect samples of thesame ecotype but from different shore levels to grow at similar rates, as was thecase. Mass-migration of RB ecotypes from the upper and SU ecotypes from thelower shore into the mid-shore may hypothetically explain different growth ratesof hybrids and parental ecotypes picked at the same shore level, still assumingenvironmental imprinting. However, RB and SU individuals from the mid-shoreare slightly more similar in allozymes to each other than are RB and SU ecotypesfrom upper and lower shore sites (Rolan-Alvarez et al., 1996) which suggests a mid-shore origin of at least most of the RB and SU individuals picked at mid-shorelevels.
Thus we conclude that the differences in growth rate between the ecotypes reflectinherited differences. These differences have most probably evolved and are probablystill maintained by divergent selection favouring small, slowly growing, snails in thelower shore and large, rapidly growing, snails in the upper shore. We haveearlier found evidence of assortative mating in the mid-shore, indicating a partialreproductive barrier between the RB and SU ecotypes and perhaps incipientspeciation ( Johannesson et al., 1995). However, the current level of gene flow, asassessed by allozyme screening, is currently large enough to prevent stochasticdivergence (and speciation) between ecotypes within shores ( Johannesson et al.,1993; Rolan-Alvarez et al., 1996), and the growth rate differences are thus not aconsequence of drift. Perhaps the size differences, however, contribute to the partialreproductive barrier between the ecotypes, as in some other species of snails(Erlandsson & Johannesson, 1994; Staub & Ribi, 1995; Yusa, pers. comm.).
We interpreted the ecotypic growth differences as due to differential selection atupper and lower shore levels. What could be the cause of these differences? Predatorycrabs attack small snails more readily, thus favouring rapid growth and large size(Elner & Raffaelli, 1980; Reimchen, 1982; Johannesson, 1986; Rolan-Alvarez et al.,1997). We observed a lot of crabs in the upper shore, but saw none in the lowershore where their presence is probably discouraged by heavy wave action. Thusselection resulting from crab predation may explain at least some of the sizedifferences. Large snails may similarly be constrained by limited availability of refugesites at the wave-affected lower shore levels (Emson & Faller-Fritsch, 1976; Raffaelli& Hughes, 1978). There are spatially variable survival rates in both parental ecotypesand hybrids. The RB ecotype survives best of all at upper shore levels while amonglarge snails the SU ecotype survives best at lower shore levels, with hybrids andparental phenotypes being equally fit in the mid-shore (Rolan-Alvarez et al., 1997).We do not currently know, however, how much the recorded differences in growthrate and size contribute to the total difference in survival rates.
On the west coast of Sweden, a small slowly growing ecotype of L. saxatilis isconfined to exposed cliffs, while a large, rapidly growing, ecotype is present only inboulder-strewn shores ( Janson, 1982).
Reciprocal transplants and animals bred in the laboratory confirm that the

K. JOHANNESSON ET AL.278
differences between the Swedish ecotypes are to a large part inherited ( Janson,1982; Johannesson & Johannesson, 1996). Furthermore, the phenotypic differencescause differential mortality of the two Swedish ecotypes ( Janson, 1983).
The similarity of the Galician and Swedish systems is promising and supports amodel of differentiation in size between ecotypes caused mainly by genetic differencesin growth rates responding to selection. After all, it may not be too surprising thatgenetic clines in, for example, growth rate are present in L. saxatilis which onlymoves a few meters per month ( Janson, 1983; Erlandsson et al., subm.), has directdevelopment, and lives in an environment with frequent horizontal and verticalhabitat shifts (see e.g. Endler, 1977).
ACKNOWLEDGEMENTS
This work was funded by the Swedish Natural Research Council and theCollianders Foundation (to KJ) and XUGA 2008B94 and DGICYT PB94–0649 (toER-A).
REFERENCES
Appleton RD, Palmer AR. 1988. Water-borne stimuli released by predatory crabs and damagedprey induce more predatory-resistant shells in a marine gastropod. Proceedings of the National Academyof Sciences, USA 85: 4387–4391.
Behrens Yamada S. 1987. Geographic variation in the growth rates of Littorina littorea and L. saxatilis.Marine Biology 96: 529–534.
Behrens Yamada S, Boulding EG. 1996. The role of highly mobile crab predators in the intertidalzonation of their gastropod prey. Journal of Experimental Marine Biology and Ecology 204: 59–83.
Boulding EG, Van Alstyne KL. 1993. Mechanisms of differential survival and growth of two speciesof Littorina on wave-exposed and on protected shores. Journal of Experimental Marine Biology and Ecology169: 139–166.
Boulding EG. 1990. Are the opposing selection pressures on exposed and protected shores sufficientto maintain genetic differentiation between gastropod populations with high intermigration rates?Hydrobiologia 193: 41–52.
Boulding EG, Buckland-Nicks J, Van Alstyne KL. 1993. Morphological and allozyme variationin Littorina sitkana and related Littorina species from the Northeastern Pacific. Veliger 36: 43–68.
Brown KM, Quinn JF. 1988. The effect of wave action on growth in three species of intertidalgastropods. Oecologia 75: 420–425.
Chapman MG. 1994. Small-scale patterns of distribution and size-structure of the intertidal littorinidLittorina unifasciata (Gastropoda: Littorinidae) in New South Wales. Australian Journal of Marine andFreshwater Research 45: 635–652.
Chow V. 1987. Patterns of growth and energy allocation in northern California populations of Littorina(Gastropoda: Prosobranchia). Journal of Experimental Marine Biology and Ecology 110: 69–89.
Denny MW, Daniel TL, Koehl MAR. 1985. Mechanical limits to size in wave-swept organisms.Ecological Monographs 55: 69–102.
Elner RW, Raffaelli DG. 1980. Interactions between two marine snails Littorina rudis Maton andLittorina nigrolineata Grey, a predator, Carcinus maenas (L.) and a parasite, Microphallus similis Jagerskiold.Journal of Experimental Marine Biology and Ecology 43: 151–160.
Emson RH, Faller-Fritsch RJ. 1976. An experimental investigation into the effect of creviceavailability on abundance and size structure in a population of Littorina rudis (Maton): Gastropoda:Prosobranchia. Journal of Experimental Marine Biology and Ecology 23: 285–297.
Endler JA. 1977. Geographic variation, speciation, and clines. Princeton: Princeton University Press.Erlandsson J. 1996. Mating behavioural patterns of rocky shore Littorina snails. Master’s thesis.
Goteborg University.

INTRASPECIFIC DIFFERENCES IN GROWTH OF LITTORINA 279
Erlandsson J, Johannesson K. 1994. Sexual selection on female size in a marine snail, Littorinalittorea (L). Journal of Experimental Marine Biology and Ecology 181: 145–157.
Etter RJ. 1989. Life history variation in the intertidal snail Nucella lapillus across a wave-exposuregradient. Ecology 70: 1857–1876.
Hughes RN. 1980. Population dynamics, growth and reproductive rates of Littorina nigrolineata Grayfrom a moderately sheltered locality in North Wales. Journal of Experimental Marine Biology and Ecology44: 211–228.
Janson K. 1982. Genetic and environmental effects on the growth rate of Littorina saxatilis. MarineBiology 69: 73–78.
Janson K. 1983. Selection and migration in two distinct phenotypes of Littorina saxatilis in Sweden.Oecologia 59: 58–61.
Janson K. 1985. Variation in the occurrence of abnormal embryos in females of the intertidalgastropod Littorina saxatilis Olivi. Journal of Molluscan Studies 51: 64–68.
Johannesson B. 1986. Shell morphology of Littorina saxatilis Olivi: the relative importance of physicalfactors and predation. Journal of Experimental Marine Biology and Ecology 102: 183–195.
Johannesson B, Johannesson K. 1990. Littorina neglecta Bean, a morphological form within thevariable species Littorina saxatilis (Olivi)? Hydrobiologia 193: 71–87.
Johannesson B, Johannesson K. 1996. Population differences in behaviour and morphology inthe snail Littorina saxatilis: Phenotypic plasticity or genetic differentiation? Journal of Zoology 240:475–493.
Johannesson K, Johannesson B, Rolan-Alvarez E. 1993. Morphological differentiation andgenetic cohesiveness over a microenvironmental gradient in the marine snail Littorina saxatilis.Evolution 47: 1770–1787.
Johannesson K, Rolan-Alvarez E, Ekendahl A. 1995. Incipient reproductive isolation betweentwo sympatric morphs of the intertidal snail Littorina saxatilis. Evolution 49: 1180–1190.
Kemp P, Bertness MD. 1984. Snail shape and growth rates: Evidence for plastic shell allometry inLittorina littorea. Proceedings of the National Academy of Sciences, USA 81: 811–813.
Palmer AR. 1990. Effects of crab effluent and scent of damaged conspecifics on feeding, growth,and shell morphology of the Atlantic dogwhelk Nucella lapillus (L.). Hydrobiologia 193: 155–182.
Raffaelli DG, Hughes RN. 1978. The effects of crevices size and availability on populations ofLittorina rudis and Littorina neritoides. Journal of Animal Ecology 47: 71–83.
Reid DG. 1996. Systematics and evolution in Littorina. London: Ray Society.Reimchen TE. 1981. Microgeographical variation in Littorina mariae Sacchi & Rastelli and a taxonomic
consideration. Journal of Conchology 30: 341–350.Reimchen TE. 1982. Shell size divergence in Littorina mariae and L. obtusata and predation by crabs.
Canadian Journal of Zoology 60: 687–695.Ricker WE. 1973. Linear regressions in fishery research. Journal of the Fisheries Research Board of Canada
30: 409–434.Roberts DJ, Hughes RN. 1980. Growth and reproductive rates of Littorina rudis from three contrasted
shores in North Wales, UK. Marine Biology 58: 47–54.Rolan-Alvarez E, Johannesson K, Erlandsson J. 1997. The maintenance of a cline in the marine
snail Littorina saxatilis: the role of home site advantage and hybrid fitness. Evolution (in press).Rolan-Alvarez E, Rolan E, Johannesson K. 1996. Differentiation in radular and embryonic
characters, and further comments on the gene flow, between two sympatric morphs of Littorinasaxatilis (Olivi). Ophelia 45: 1–15.
Rohlf FJ, Sokal RR. 1981. Statistical tables. San Francisco: Freeman.Saur M. 1990. Mate discrimination in Littorina littorea (L.) and L. saxatilis (Olivi) (Mollusca: Proso-
branchia). Hydrobiologia 193: 261–270.Sokal RR, Rohlf FJ. 1981. Biometry. San Francisco: Freeman.Staub R, Ribi G. 1995. Size assortative mating in a natural population of Viviparus ater (Gastropoda:
Prosobranchia) in Lake Zurich, Switzerland. Journal of Molluscan Studies 61: 237–247.Struhsaker JW. 1968. Selection mechanisms associated with intraspecific shell variation in Littorina
picta. Evolution 22: 459–480.Sundberg P. 1988. Microgeographic variation in shell characters of Littorina saxatilis Olivi – a question
mainly of size? Biological Journal of the Linnean Society 35: 169–184.Tsutakawa RK, Hewett JE. 1977. Quick test for comparing two populations with bivariate data.
Biometrics 33: 215–219.Zaykin DV, Pudovkin AI. 1991. The programme multitest: calculation indices of statistical significance
for multiple tests. Genetika 27: 2034–2038.
Copyright © 2022 FDOKUMEN




















