
Life cycle of the temporary fish parasite, gnathia africana (crustacea: isopoda: gnathiidae)
Upload
worldagroforestryCategory
view
0download
0
oneOnewththis
sae,hasomthee tohes04).eanlianrateees
s in
nes)
the
are
rier
Faso
cies.
rea.
ther
tem
was
thin
ased
ith
eds
y be
ved.
G Model
FORECO-11100; No of Pages 8
Growth and survival of Prosopis africana provenances tested in Niger andrelated to rainfall gradients in the West African Sahel
John C. Weber a,*, Mahamane Larwanou b, Tougiani A. Abasse c, Antoine Kalinganire a
a World Agroforestry Centre (ICRAF), BP 320, Bamako, Malib Universite Abdou Moumouni, Niamey, Nigerc Institut National de la Recherche Agronomique du Niger, Maradi, Niger
1. Introduction
Tree improvement programs must understand patterns ofintraspecific variation in tree species in order to effectively manageand conserve their genetic resources. In regions with pronouncedenvironmental gradients in temperature and rainfall, tree popula-tions may exhibit continuous variation in growth, phenology,survival and other adaptive traits in relation to these gradients(Langlet, 1971). This clinal variation is generally assumed to reflectthe result of natural selection along the environmental gradient.Clines have been reported in many conifer and hardwood speciesfrom the temperate zone (Morgenstern, 1996), but there have beenvery few published reports of clinal variation in tropical hard-woods. The West African Sahel is a semi-arid transitional zonebetween the relatively humid savannah woodlands to the southand the Sahara Desert to the north. In general, mean annual rainfall
decreases from south to north and from west to east in this z(Sivakumar and Gnoumou, 1987; Sivakumar et al., 1993).would expect, therefore, the existence of clinal variation in groand survival of native tree species distributed throughoutregion.
Prosopis africana (Guill., Perrott. and Rich.) Taub. (Leguminosub-family Mimosoideae) is the only native Prosopis in Africa. Ita natural distribution from Senegal to Ethiopia in the north, frGuinea to Cameroon in the south, and from Uganda to Egypt ineast; but it has disappeared from extensive parts of its range duover-exploitation, such as excessive cutting of stems and brancresulting in limited natural regeneration (Pasiecznik et al., 20In West Africa, it extends throughout the Sudanian and Guinecozones in the southern part of its range and into the Saheecozone in the northern part of its range. It does not tolehabitually dry sites, preferring 600–1500 mm annual rainfall. Tr
Forest Ecology and Management xxx (2008) xxx–xxx
A R T I C L E I N F O
Article history:
Received 10 January 2008
Received in revised form 30 April 2008
Accepted 6 May 2008
Keywords:
Adaptation
Cline
Drought
Conservation
Reforestation
A B S T R A C T
Prosopis africana (Guill., Perrott. and Rich.) Taub. is important for farming and pastoralist communitie
the West African Sahel. It provides a range of essential products (wood, fuel, food, fodder and medici
and services (soil fertility improvement), and is one of many native tree species maintained in
traditional agroforesty system, known as the parkland. However, this and other valuable species
disappearing in many parts of the region due to over-exploitation and, perhaps, due to increasingly d
conditions in the region. A provenance/progeny test (275 families from 28 provenances in Burkina
and Niger) was established at one location in Niger to investigate geographic variation in the spe
Mean annual rainfall decreases from south to north and from west to east in the seed collection a
Given these gradients, one would expect clinal variation in growth and survival of P. africana and o
tree species that occur naturally in the region. Tree growth (height, number of primary branches, s
diameter at 20 cm and 1.3 m) and survival were analyzed at 11 years. Variation due to provenances
significant for growth variables and almost significant for survival. Variation due to families wi
provenances was not significant for any variable. Provenance means for growth and survival incre
from the more humid to the drier parts of the sample region. The clines in growth and survival w
regional rainfall patterns suggest that tree improvement and conservation programs should collect se
from populations in the drier zones for planting and conservation in the West African Sahel. This ma
particularly important when one considers the trend in global warming.
� 2008 Elsevier B.V. All rights reser
Contents lists available at ScienceDirect
Forest Ecology and Management
journal homepage: www.elsevier .com/locate/ foreco
0 msts.ay
ing* Corresponding author. Tel.: +223 2223375; fax: +223 2228683.
E-mail address: [email protected] (J.C. Weber).
0378-1127/$ – see front matter � 2008 Elsevier B.V. All rights reserved.
doi:10.1016/j.foreco.2008.05.004
Please cite this article in press as: Weber, J.C. et al., Growth and survrainfall gradients in the West African Sahel, Forest Ecol. Manage. (
produce a deep taproot, grow slowly but can attain a height of 2in natural stands, and can be coppiced for successive harveTrees in natural stands generally have an erect form, but they mbe multi-stemmed due to forking in the lower trunk. The breed
ival of Prosopis africana provenances tested in Niger and related to2008), doi:10.1016/j.foreco.2008.05.004

systcroset apollafric
browdrythetheidistalongstrustudwernurs
P
comis mBurktermconsfarm(Pasblacleavgoatsom(Barso thFayerootKali(Haltrad
BnitieparkAfricexamareafacttionareapasthott
IprogdisctestNigenumandmajvarirelaesis,
2. M
2.1.
TNigeandobta
2
G Model
FORECO-11100; No of Pages 8
Plerai
em has not been studied, but it is assumed to be primarily out-sing, as reported for other species in the genus (e.g., Bessegal., 2000; Dhillon et al., 2003). Prosopis species are insectinated (e.g., Toro, 2002), but the guild of pollinators of P.
ana has not been investigated. Seeds are naturally dispersed bysing animals, such as camels, cattle and goats at the end of the
season (Tybirk, 1991), and perhaps also by humans who collectpods to feed to their animals and collect cow dung to fertilizer fields (A. Bationo, personal communication). Seed dispersalnces could vary, therefore, from relatively short to relativelydistances, and this would affect the population genetic
cture (Hamrick et al., 1992). There has been only one publishedy of genetic variation in P. africana: significant differencese observed in height among four Nigerian provenances in aery test (Akinnagbe and Oni, 2007).. africana is very important for farming and pastoralistmunities in the West African Sahel (Agboola, 2004). The woododerately dense (basic density = 687 kg/m3 at one site inina Faso: Nygard and Elving, 2000) and very resistant toites and fungi (Gerardin et al., 2004), making it useful fortruction poles and planks, mortars, pestles, and handles forimplements. It is also highly valued for firewood and charcoal
iecznik et al., 2004), and is the preferred species by manyksmiths in the region (J.C. Weber, personal observations). Itses, succulent branches and pods provide fodder for cattle ands, which is essential during the 9-month long dry season. Ine areas, people use the fermented seeds as a food condimentminas et al., 1998; Aremu et al., 2006; Kalinganire et al., 2007),ere is a thriving market for the seeds in these areas (M. Dieye, personal communication). The leaves, branches, bark ands are used for several traditional medicines (Arbonnier, 2002;nganire et al., 2007). In addition, trees fix atmospheric nitrogenliday, 1984) and, therefore, can improve soil fertility in theitional parkland agroforestry system.ecause it has many uses for farming and pastoralist commu-s, there is intensive extraction pressure on P. africana inland agroforests and state-controlled forests in the Westan Sahel. Excessive branch lopping and pod harvesting, forple, have seriously reduced the natural regeneration in some
s (J.C. Weber, personal observations). This, together with thethat few communities protect and manage natural regenera-, has dramatically reduced the abundance of P. africana in manys (J.C. Weber, personal observations). In addition, farmers andoralists state that many trees are dying due to increasinglyer, drier conditions in the region (ICRAF-IFAD, 2006).n this paper, results are presented from the first provenance/eny test of P. africana, and some practical implications are
ussed for tree improvement and conservation programs. Theincludes 275 families from 28 provenances in Burkina Faso andr, and was established at one site in Niger. Tree growth (height,ber of primary branches, stem diameter at 20 cm and 1.3 m)survival were evaluated 11 years after establishment. The
or objectives were to determine (a) if growth and survivaled significantly due to provenances, and (b) if variation wasted to provenance latitude and/or longitude and, by hypoth-
regional gradients in mean annual rainfall.
aterials and methods
m.org/), described by Hijmans et al. (2005), and used to constructmean annual rainfall isohyets for the sample region (Fig. 1). Meanannual rainfall decreases from west to east (�750–350 mm) andfrom south to north (�750–650 mm in the west,�450–350 mm inthe east). Mean annual temperature varies from approximately 28to 29 8C, being slightly higher in the north than in the south(Sivakumar and Gnoumou, 1987; Sivakumar et al., 1993). Soils aregenerally very sandy and infertile: they are classified as arenosolsthroughout most of the sample region, with some regosols andvertisols in Burkina Faso (FAO, 2007).
Fruit pods were collected in March–April 1993 from 320 mothertrees located in 28 natural stands in the sample region (Fig. 1). Thestands are referred to hereafter as provenances, using this term inthe broad sense as the source of seed from a defined geographicarea (Morgenstern, 1996). Trees were selected if they had sufficientpods and no obvious external disease symptoms. To reduce thechance of sampling siblings, at least 100 m was maintainedbetween any two selected trees. Latitude, longitude and elevationwere recorded for each selected tree, using a GPS receiver.
2.2. Seedling production, experimental design and management of the
provenance/progeny test
Seed processing and seedling production were carried out at theInternational Crops Research Institute for the Semi-Arid Tropics(ICRISAT) Sahelian Centre, located 40 km southeast of Niamey,Niger (Fig. 1). Pods were maintained in separate bags for each tree,and air-dried in the shade for 2 months. Seeds were extracted byhand, scarified with sand paper, soaked in water for 48 h, and thensown in plastic nursery bags.
Only 275 of the 320 mother trees produced sufficient plants inthe nursery for the provenance/progeny test. The number ofmother trees per provenance varied from 3 to 15, with a median ofapproximately 11 (Table 1). Seedlings from these trees werearranged in the nursery according to the experimental design usedin the field (described below), and grown for 12 months. The plantswere partially shaded with millet stems, watered daily, and rootsand shoots were regularly pruned to control growth.
The provenance/progeny test was established in July 1994 atthe ICRISAT Sahelian Centre (1381403500N, 281701400E, 230-melevation). Mean annual rainfall and temperature are 539 mmand 29 8C, respectively (I. Maıkano, personal communication), andthe soils are very sandy and infertile (arenosols). Annual rainfall isquite variable at the test site, ranging from 372 to 794 mm since1994.
The experimental design was a randomized complete block,with eight blocks (replications) and 275 families. In eachreplication, the families were randomly assigned to 275 single-tree plots. Spacing was 3 m within and among rows. A row ofborder trees was planted around each replication, using a mixtureof seedlings from 25 families that had abundant viable seeds. Nofertilizers or insecticides were applied at any time. Dead trees werereplanted in August–September 1994 and the replications wereweeded during the first year, but not thereafter.
2.3. Data collection and statistical analyses
Tree growth was evaluated in 2005, 11 years after trialestablishment. Tree height was measured to the nearest cm using
J.C. Weber et al. / Forest Ecology and Management xxx (2008) xxx–xxx
Sample region
he sample region extends from central Burkina Faso to centralr, covering an area approximately 1200 km from west to east,50–200 km from south to north (Fig. 1). Rainfall data wereined from the WorldClim database (http://www.worldcli-
ase cite this article in press as: Weber, J.C. et al., Growth and surnfall gradients in the West African Sahel, Forest Ecol. Manage.
a telescopic measuring pole. The number of primary branches wascounted. Stem diameter was measured to the nearest mm at 20 cm(basal diameter, BasDia) and at 1.3 m (diameter at breast height,DBH) above ground level using diameter tape. For multi-stemmedtrees, DBH was measured on the stem with the largest diameter.Replants were recorded as dead trees for the estimation of survival,
vival of Prosopis africana provenances tested in Niger and related to(2008), doi:10.1016/j.foreco.2008.05.004

rateu-
e tobutded
wasre).reeere
tingde,ing
rate, sothis
tedees
toand
ily.
. The
3
G Model
FORECO-11100; No of Pages 8
and they were not included in the analyses of tree growth. Thenumber of living trees was recorded for each family andprovenance across the eight replications, and used to calculatethe proportion of living trees at the family and provenance levels(referred to hereafter as survival).
The SAS1 statistical package Version 9.1 (SAS Institute Inc.,2004) was used for all statistical analyses. Departure from thenormal distribution and homogeneity of variance were tested forthe residuals using statistics provided by the UNIVARIATEprocedure. Data transformations for height, BasDia, DBH andsurvival were not considered necessary to satisfy the assumptionsof analysis of variance, but the square-root transformation wasused for the number of branches. The significance level for all testswas a � 0.05.
Analyses of variance of individual tree growth and survival atthe family level were carried out using the MIXED procedure(restricted maximum likelihood method). The model for growthvariables was: Yijkl = m + ai + ßj + gk(j) + aßij + eijkl, where Yijkl = treel
in treatment combinationijk, m = the grand mean, ai = replicationi,ßj = provenancej, gk(j) = familyk nested within provenancej,aßij = interaction between replicationi and provenancej, andeijkl = residual error. The model for survival was: Yij = m + ßi + eij,where Y = proportion in treatment , m = the grand mean,
minimum sample size of 40 is recommended to obtain accuestimates of variance components (N. Mandel, personal commnication): the number of families (275) is more than adequatobtain an accurate estimate for family nested in provenance,the number of provenances (28) is lower than the recommenminimum.
Clinal variation in provenance growth and survivalinvestigated using multiple linear regression (REG proceduModels were selected using the backward technique: thindependent variables (latitude, longitude and elevation) wentered in a full model, and this was reduced by eliminavariables that were not significant at a � 0.05. Mean latitulongitude and elevation were computed for each provenance usthe location of the trees sampled in the provenance. Accurainfall data were not available for all provenance locationsmultiple regressions could not be carried out directly withclimatic variable.
Pearson (r) linear correlation coefficients were calculaamong the four growth variables, using data from individual tr(CORR procedure). Correlation coefficients were also useddetermine if there was a linear relationship between survivalgrowth: in this case, correlations were calculated between fammeans for growth variables and the proportion of living trees
Fig. 1. Geographic location of 28 Prosopis africana provenances sampled in Burkina Faso and Niger, and mean annual rainfall isohyets (mm) in the sample region
provenance/progeny test is located at the ICRISAT Sahelian Centre, approximately 40 km southeast of Niamey, Niger.
J.C. Weber et al. / Forest Ecology and Management xxx (2008) xxx–xxx
answthore,
ij j i
ßi = provenancei, and eij = residual error (families nested inprovenances). All factors were considered random, and Z-testswere used to determine if variance components were significantlygreater than zero. Each variance component was expressed as apercentage of the total variance (VC%), where the total varianceequals the sum of all variance components in the model. A
Please cite this article in press as: Weber, J.C. et al., Growth and survrainfall gradients in the West African Sahel, Forest Ecol. Manage. (
3. Results
Sample sizes were very small for estimating family me(maximum of eight trees) and a few provenance means for grovariables (<20 trees for provenances 8 and 14, Table 2). Theref
ival of Prosopis africana provenances tested in Niger and related to2008), doi:10.1016/j.foreco.2008.05.004
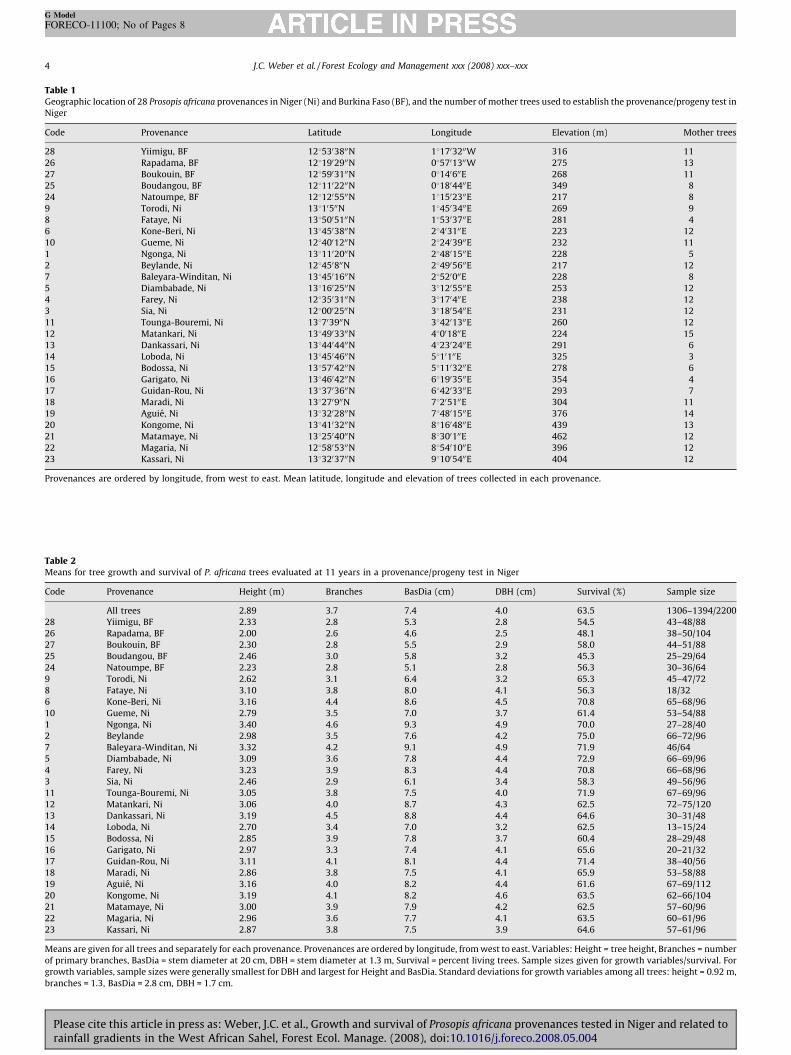
Table 1Geographic location of 28 Prosopis africana provenances in Niger (Ni) and Burkina Faso (BF), and the number of mother trees used to establish the provenance/progeny test in
Niger
Code Provenance Latitude Longitude Elevation (m) Mother trees
28 Yiimigu, BF 1285303800N 181703200W 316 11
26 Rapadama, BF 1281902900N 085701300W 275 13
27 Boukouin, BF 1285903100N 08140600E 268 11
25 Boudangou, BF 1281102200N 081804400E 349 8
24 Natoumpe, BF 1281205500N 181502300E 217 8
9 Torodi, Ni 13810500N 184503400E 269 9
8 Fataye, Ni 1385005100N 185303700E 281 4
6 Kone-Beri, Ni 1384503800N 28403100E 223 12
10 Gueme, Ni 1284001200N 282403900E 232 11
1 Ngonga, Ni 1381102000N 284801500E 228 5
2 Beylande, Ni 128450800N 284905600E 217 12
7 Baleyara-Winditan, Ni 1384501600N 28520000E 228 8
5 Diambabade, Ni 1381602500N 381205500E 253 12
4 Farey, Ni 1283503100N 38170400E 238 12
3 Sia, Ni 1280002500N 381805400E 231 12
11 Tounga-Bouremi, Ni 138703900N 384201300E 260 12
12 Matankari, Ni 1384903300N 48001800E 224 15
13 Dankassari, Ni 1384404400N 482302400E 291 6
14 Loboda, Ni 1384504600N 5810100E 325 3
15 Bodossa, Ni 1385704200N 581103200E 278 6
16 Garigato, Ni 1384604200N 681903500E 354 4
17 Guidan-Rou, Ni 1383703600N 684203300E 293 7
18 Maradi, Ni 138270900N 78205100E 304 11
19 Aguie, Ni 1383202800N 784801500E 376 14
20 Kongome, Ni 1384103200N 881604800E 439 13
21 Matamaye, Ni 1382504000N 88300100E 462 12
22 Magaria, Ni 1285805300N 885401000E 396 12
23 Kassari, Ni 1383203700N 981005400E 404 12
Provenances are ordered by longitude, from west to east. Mean latitude, longitude and elevation of trees collected in each provenance.
Table 2Means for tree growth and survival of P. africana trees evaluated at 11 years in a provenance/progeny test in Niger
Code Provenance Height (m) Branches BasDia (cm) DBH (cm) Survival (%) Sample size
All trees 2.89 3.7 7.4 4.0 63.5 1306–1394/2200
28 Yiimigu, BF 2.33 2.8 5.3 2.8 54.5 43–48/88
26 Rapadama, BF 2.00 2.6 4.6 2.5 48.1 38–50/104
27 Boukouin, BF 2.30 2.8 5.5 2.9 58.0 44–51/88
25 Boudangou, BF 2.46 3.0 5.8 3.2 45.3 25–29/64
24 Natoumpe, BF 2.23 2.8 5.1 2.8 56.3 30–36/64
9 Torodi, Ni 2.62 3.1 6.4 3.2 65.3 45–47/72
8 Fataye, Ni 3.10 3.8 8.0 4.1 56.3 18/32
6 Kone-Beri, Ni 3.16 4.4 8.6 4.5 70.8 65–68/96
10 Gueme, Ni 2.79 3.5 7.0 3.7 61.4 53–54/88
1 Ngonga, Ni 3.40 4.6 9.3 4.9 70.0 27–28/40
2 Beylande 2.98 3.5 7.6 4.2 75.0 66–72/96
7 Baleyara-Winditan, Ni 3.32 4.2 9.1 4.9 71.9 46/64
5 Diambabade, Ni 3.09 3.6 7.8 4.4 72.9 66–69/96
4 Farey, Ni 3.23 3.9 8.3 4.4 70.8 66–68/96
3 Sia, Ni 2.46 2.9 6.1 3.4 58.3 49–56/96
11 Tounga-Bouremi, Ni 3.05 3.8 7.5 4.0 71.9 67–69/96
12 Matankari, Ni 3.06 4.0 8.7 4.3 62.5 72–75/120
13 Dankassari, Ni 3.19 4.5 8.8 4.4 64.6 30–31/48
14 Loboda, Ni 2.70 3.4 7.0 3.2 62.5 13–15/24
15 Bodossa, Ni 2.85 3.9 7.8 3.7 60.4 28–29/48
16 Garigato, Ni 2.97 3.3 7.4 4.1 65.6 20–21/32
17 Guidan-Rou, Ni 3.11 4.1 8.1 4.4 71.4 38–40/56
18 Maradi, Ni 2.86 3.8 7.5 4.1 65.9 53–58/88
19 Aguie, Ni 3.16 4.0 8.2 4.4 61.6 67–69/112
20 Kongome, Ni 3.19 4.1 8.2 4.6 63.5 62–66/104
21 Matamaye, Ni 3.00 3.9 7.9 4.2 62.5 57–60/96
22 Magaria, Ni 2.96 3.6 7.7 4.1 63.5 60–61/96
23 Kassari, Ni 2.87 3.8 7.5 3.9 64.6 57–61/96
Means are given for all trees and separately for each provenance. Provenances are ordered by longitude, from west to east. Variables: Height = tree height, Branches = number
of primary branches, BasDia = stem diameter at 20 cm, DBH = stem diameter at 1.3 m, Survival = percent living trees. Sample sizes given for growth variables/survival. For
growth variables, sample sizes were generally smallest for DBH and largest for Height and BasDia. Standard deviations for growth variables among all trees: height = 0.92 m,
branches = 1.3, BasDia = 2.8 cm, DBH = 1.7 cm.
J.C. Weber et al. / Forest Ecology and Management xxx (2008) xxx–xxx4
G Model
FORECO-11100; No of Pages 8
Please cite this article in press as: Weber, J.C. et al., Growth and survival of Prosopis africana provenances tested in Niger and related torainfall gradients in the West African Sahel, Forest Ecol. Manage. (2008), doi:10.1016/j.foreco.2008.05.004
rierudewas
ithare
659and
Thetwoest
5)
VC%
–
6.7
–
–
93.3
.3 m,
75 –
d in
nent
964
0241
009
369
0092
003
964
0741
028
795
0449
017
0045
002
cm),
west
ient.
in a
H
305)
5
G Model
FORECO-11100; No of Pages 8
results focus on variation due to provenances, rather than at thefamily level.
3.1. Variation in tree growth and survival
Mean growth and survival of P. africana trees were relativelylow at 11 years in this test (Table 2). However, there were relativelylarge differences among provenances. The range in provenancemeans was 1.4 m for tree height, 2 for the number of primarybranches, 4.7 cm for basal stem diameter (BasDia), 2.4 cm for stemDBH, and 29.7% for survival. The largest means were observed inprovenance #1 for growth traits and provenance #2 for survival,and the corresponding smallest means were observed in prove-nance #26 and #25, respectively.
Analysis of variance confirmed that there was significantvariation in tree growth due to provenances, while variation insurvival was almost significant (Table 3). Provenances accountedfor approximately 13–17% of the total variation in growth variables(VC% lowest for DBH and highest for BasDia), and about 7% of thevariation in survival. Variation due to families within provenanceswas not significant for any growth variable. Most variation wasresidual (VC% generally >80%).
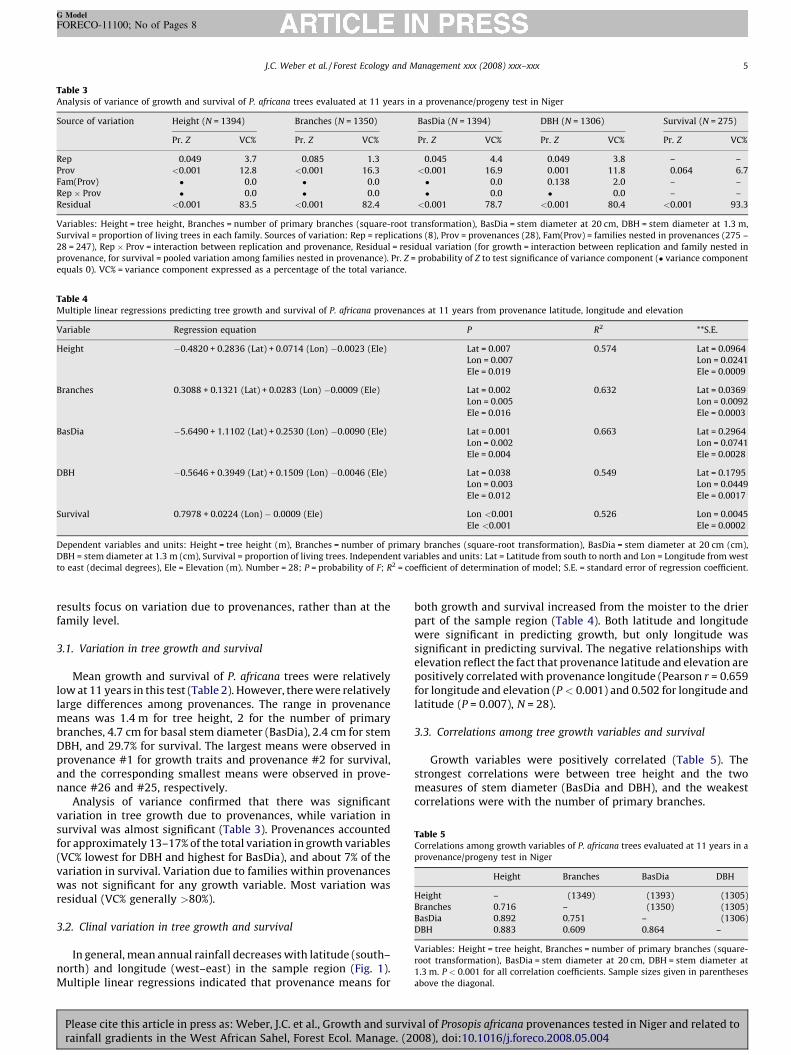
both growth and survival increased from the moister to the dpart of the sample region (Table 4). Both latitude and longitwere significant in predicting growth, but only longitudesignificant in predicting survival. The negative relationships welevation reflect the fact that provenance latitude and elevationpositively correlated with provenance longitude (Pearson r = 0.for longitude and elevation (P < 0.001) and 0.502 for longitudelatitude (P = 0.007), N = 28).
3.3. Correlations among tree growth variables and survival
Growth variables were positively correlated (Table 5).strongest correlations were between tree height and themeasures of stem diameter (BasDia and DBH), and the weakcorrelations were with the number of primary branches.
Table 3Analysis of variance of growth and survival of P. africana trees evaluated at 11 years in a provenance/progeny test in Niger
Source of variation Height (N = 1394) Branches (N = 1350) BasDia (N = 1394) DBH (N = 1306) Survival (N = 27
Pr. Z VC% Pr. Z VC% Pr. Z VC% Pr. Z VC% Pr. Z
Rep 0.049 3.7 0.085 1.3 0.045 4.4 0.049 3.8 –
Prov <0.001 12.8 <0.001 16.3 <0.001 16.9 0.001 11.8 0.064
Fam(Prov) � 0.0 � 0.0 � 0.0 0.138 2.0 –
Rep � Prov � 0.0 � 0.0 � 0.0 � 0.0 –
Residual <0.001 83.5 <0.001 82.4 <0.001 78.7 <0.001 80.4 <0.001
Variables: Height = tree height, Branches = number of primary branches (square-root transformation), BasDia = stem diameter at 20 cm, DBH = stem diameter at 1
Survival = proportion of living trees in each family. Sources of variation: Rep = replications (8), Prov = provenances (28), Fam(Prov) = families nested in provenances (2
28 = 247), Rep � Prov = interaction between replication and provenance, Residual = residual variation (for growth = interaction between replication and family neste
provenance, for survival = pooled variation among families nested in provenance). Pr. Z = probability of Z to test significance of variance component (� variance compo
equals 0). VC% = variance component expressed as a percentage of the total variance.
Table 4Multiple linear regressions predicting tree growth and survival of P. africana provenances at 11 years from provenance latitude, longitude and elevation
Variable Regression equation P R2 **S.E.
Height �0.4820 + 0.2836 (Lat) + 0.0714 (Lon) �0.0023 (Ele) Lat = 0.007 0.574 Lat = 0.0
Lon = 0.007 Lon = 0.
Ele = 0.019 Ele = 0.0
Branches 0.3088 + 0.1321 (Lat) + 0.0283 (Lon) �0.0009 (Ele) Lat = 0.002 0.632 Lat = 0.0
Lon = 0.005 Lon = 0.
Ele = 0.016 Ele = 0.0
BasDia �5.6490 + 1.1102 (Lat) + 0.2530 (Lon) �0.0090 (Ele) Lat = 0.001 0.663 Lat = 0.2
Lon = 0.002 Lon = 0.
Ele = 0.004 Ele = 0.0
DBH �0.5646 + 0.3949 (Lat) + 0.1509 (Lon) �0.0046 (Ele) Lat = 0.038 0.549 Lat = 0.1
Lon = 0.003 Lon = 0.
Ele = 0.012 Ele = 0.0
Survival 0.7978 + 0.0224 (Lon) � 0.0009 (Ele) Lon <0.001 0.526 Lon = 0.
Ele <0.001 Ele = 0.0
Dependent variables and units: Height = tree height (m), Branches = number of primary branches (square-root transformation), BasDia = stem diameter at 20 cm (
DBH = stem diameter at 1.3 m (cm), Survival = proportion of living trees. Independent variables and units: Lat = Latitude from south to north and Lon = Longitude from
to east (decimal degrees), Ele = Elevation (m). Number = 28; P = probability of F; R2 = coefficient of determination of model; S.E. = standard error of regression coeffic
Table 5Correlations among growth variables of P. africana trees evaluated at 11 years
provenance/progeny test in Niger
Height Branches BasDia DB
Height – (1349) (1393) (1
J.C. Weber et al. / Forest Ecology and Management xxx (2008) xxx–xxx
Branches 0.716 – (1350) (1305)
BasDia 0.892 0.751 – (1306)
DBH 0.883 0.609 0.864 –
Variables: Height = tree height, Branches = number of primary branches (square-
root transformation), BasDia = stem diameter at 20 cm, DBH = stem diameter at
1.3 m. P < 0.001 for all correlation coefficients. Sample sizes given in parentheses
above the diagonal.
3.2. Clinal variation in tree growth and survival
In general, mean annual rainfall decreases with latitude (south–north) and longitude (west–east) in the sample region (Fig. 1).Multiple linear regressions indicated that provenance means for
Please cite this article in press as: Weber, J.C. et al., Growth and survival of Prosopis africana provenances tested in Niger and related torainfall gradients in the West African Sahel, Forest Ecol. Manage. (2008), doi:10.1016/j.foreco.2008.05.004

Tbetwstrothe nP = 0
4. D
Radapimpspec(Tchandanth
20072004BergKalkSchu
Tin ginititrialcont(e.g.trialevalet acontvaritivenilot
Wol(DiaAno
Cord
CheWan199exce
Addhardinstnot
TresuInfegrapandtestmayafric
Tprovdifferegicallyvarirepoandet aRouSincLuuk
6
G Model
FORECO-11100; No of Pages 8
Plerai
here was a very weak positive relationship at the family leveleen the proportion of living trees and growth variables. The
ngest correlation was with BasDia, and the weakest was withumber of primary branches (r = 0.223, P = 0.001 and r = 0.150,.013, respectively, N = 275).
iscussion
esearch on patterns of natural variation in commercially andtively important traits is essential in order to develop tree
rovement and conservation strategies for native hardwoodies in Africa. This is particularly urgent for P. africana
oundjeu et al., 1997) and other species that are over-exploitedthreatened with local or even regional extinction [e.g., Khaya
otheca (Welw.) C. DC., Khaya ivorensis A. Chev. (Ofori et al.,); Khaya senegalensis (Desr.) A. Juss. (Diallo et al., 2000; Arnold,); Milicia excelsa (Welw.) C.C. Berg., Milicia regia (A. Chev.) C.C.. (Ofori and Cobbinah, 2007); Prunus africana (Hook f.)man (Dawson and Powell, 1999); Triplochiton scleroxylon K.m. (Bowen et al., 1977)].here has been little systematic research on genetic variation
rowth and survival of native hardwood species in Africa. FAOated, and the Danida Forest Seed Centre analyzed severals to identify some superior provenances from Africa and otherinents for reforestation in arid and semi-arid zones in Africa, Ræbild et al., 2003a,b). In addition, numerous provenances of several exotic species have been established anduated (e.g., Khasa et al., 1995; Ræbild et al., 2003c,d; Andrewl., 2004; Langat and Kariuki, 2004; Pedersen et al., 2007). Inrast, there are relatively few published reports of genetication in growth, survival and other commercially or adap-ly important traits of native African hardwoods [e.g., Acacia
ica subsp. tomentosa (L.) Willd. ex Delile (Diallo et al., 2000;de-Mieskel and Sinclair, 2000); Acacia senegal (L.) Willd.llo et al., 2000; Raddad and Luukkanen, 2006; Raddad, 2007);geissus leiocarpus (DC.) Guill. and Perr. (Diallo et al., 2000);ia africana Lam. (Loha et al., 2006); Faidherbia albida (del.) A.
v. (Sniezko and Stewart, 1989; Billand and De Framond, 1993;yancha et al., 1994; Bastide and Diallo, 1996; Dangasuk et al.,
7; Roupsard et al., 1998); K. senegalensis (Diallo et al., 2000); M.
lsa (Appiah, 2003); M. regia (Ofori and Cobbinah, 2007)].itional studies of genetic variation in these and other nativewoods have probably been conducted by national research
itutes in some African countries, but the results generally arewidely disseminated.his is the first provenance/progeny test of P. africana, and thelts cannot be extrapolated to the entire West African Sahel.rences about provenance variation are limited to the geo-hical region covered by the 28 provenances in Burkina FasoNiger. Furthermore, results and conclusions are based on oneat a single site in Niger. Nevertheless, the results of this studyhave important practical implications for management of P.
ana genetic resources in the West African Sahel.here was significant variation in tree growth due toenances in this test, indicating that there are geneticrences among natural populations of P. africana in the sample
on. Therefore, tree improvement programs can select geneti-more productive provenances for reforestation. Genetic
ation in growth among natural populations has also been
Growth and survival of P. africana provenances appears to berelated to rainfall gradients in the sample region. Provenances fromdrier parts of the region had better growth and survival thanprovenances from more humid parts of the region, when tested in arelatively dry site in Niger. To our knowledge, these are the firstreported clines in growth and survival of a hardwood species fromthe West African Sahel. Clinal, ecotypic and/or regional variation ingrowth have been reported among provenances of a few otherAfrican hardwood species (Sniezko and Stewart, 1989; Dangasuket al., 1997; Raddad, 2007).
The clines suggest that natural populations of P. africana fromthe drier parts of the sample region are genetically better adaptedto drought, compared with populations from the more humid partsof the region. This appears to be an intuitively reasonable patternof variation maintained by differences in natural selectionpressures along the rainfall gradients (Langlet, 1971). This is ahypothesis, however, and there is no direct evidence to confirmthat the variation patterns are primarily the result of naturalselection. Patterns of variation within and among naturalpopulations of timber tree species probably reflect the combinedhistorical effects of natural selection, gene flow, genetic drift,mutation (Hu and Li, 2002), forest fragmentation, inbreeding,stand density and selective logging (Murawski et al., 1994;Hardner et al., 1996; Nason and Hamrick, 1997; Cascante et al.,2002).
Assuming that the clines do reflect adaptive variation, andconsidering the fact that some computer models project a drierclimate in the Sahel during the 21st century (Held et al., 2005), itwould be prudent for tree improvement and conservationprograms to collect P. africana seed from the drier parts of theregion for planting and conservation in the West African Sahel.Specifically, we recommend that transfers of germplasm shouldonly be made from north to south, and from east to west: transfersfrom the more humid to the drier parts of the region should beavoided. This strategy is currently being used in a project to rebuildand diversify the parkland agroforests in Niger, Burkina Faso, Maliand Senegal (ICRAF-IFAD, 2006); and it was developed with 45villages in response to their observation that parkland trees need tobe better adapted to the long dry season.
Based on linear correlations, P. africana families with greaterabove-ground growth tended to have better survival at 11 years.This relationship may reflect variation in taproot growth amongfamilies, but this hypothesis requires testing. Trees that produce adeeper taproot could access soil moisture at greater depths,allowing them to grow for a longer period of time during the dryseason and thereby produce greater above-ground biomass.Moreover, they could have a selective advantage during years ofextreme drought (Pasiecznik et al., 2001). In this test, for example,there were 2 years when annual rainfall was less than 400 mm (I.Maıkano, personal communication).
More genetics research is needed in order to develop acomprehensive tree improvement and conservation program forP. africana in the West African Sahel. Since it is highly valued forboth timber and energy, studies of variation in wood properties(density, strength, stiffness, shrinkage and heat content), and theircorrelations (both phenotypic and genetic) with tree growth arenecessary. Wood with higher density tends to be stronger andstiffer, but it has greater tangential, radial and volumetricshrinkage compared with lower-density wood (Tsoumis, 1991).
J.C. Weber et al. / Forest Ecology and Management xxx (2008) xxx–xxx
rted in a few other native hardwood species in Africa (SniezkoStewart, 1989; Billand and De Framond, 1993; Wanyanchal., 1994; Bastide and Diallo, 1996; Dangasuk et al., 1997;psard et al., 1998; Diallo et al., 2000; Wolde-Mieskel andlair, 2000; Appiah, 2003; Loha et al., 2006; Raddad andkanen, 2006; Raddad, 2007).
ase cite this article in press as: Weber, J.C. et al., Growth and surnfall gradients in the West African Sahel, Forest Ecol. Manage.
If, for example, tree growth and wood density are positivelycorrelated, then selecting faster growing provenances couldincrease the strength and stiffness of the wood, but it may resultin excessive wood shrinkage (e.g., Sotelo Montes et al., 2007a,b). Inaddition, the correlation between tree growth and wood densityshould be investigated at the provenance level, as it may vary
vival of Prosopis africana provenances tested in Niger and related to(2008), doi:10.1016/j.foreco.2008.05.004

the2.
geny. Afr.
ancei. 16,
turalty of
e depourHN),
andm. 5,
d itskles,nta-–21,orth
eeds.
a enenilecs analeoret
ating132,
yme. In:des
aris,
hitonrcesited
oresttree
dlingwar,
unus
and, 71–
ivitelig-
e etde
e enorettion
ited28
004.ood.
therp. 2,
ls of
out-
7
G Model
FORECO-11100; No of Pages 8
among provenances (e.g., Weber and Sotelo Montes, 2008). In thistest, family means were poorly estimated due to small samplesizes: variation in growth, wood properties and survival needs tobe investigated in provenance/progeny tests that include largersample sizes for estimating family means and trait heritability.These tests should include a systematic sample of P. africana
provenances from the entire West African Sahel and be establishedat multiple sites in the region. These tests should be designed toquantify clinal variation related to rainfall gradients in the region;determine patterns of ecotypic variation related to soil types, asobserved in other African hardwood species (e.g., Dangasuk et al.,1997; Raddad and Luukkanen, 2006), and quantify their effects ontraits of adaptive and commercial importance; and allowcomparison of the local provenance with provenances from othersites in the region. Patterns of natural variation observed in thesetests should then be used to refine guidelines for germplasmtransfers, and develop dynamic strategies to conserve the geneticresources of this species (Hu and Li, 2002).
Studies such as this provide useful technical recommenda-tions for tree improvement and conservation programs, butsustainable management of tree genetic resources requires asupportive social and political environment. In the West AfricanSahel, farming and pastoralist communities are not involved indeveloping policies related to land and tree tenure, and thisproduces some dysfunctional policies that actually prevent ruralcommunities from sustainably managing their forest resources(Yatich et al., 2008). In addition, there are often conflictsbetween farming and pastoralist communities related to the useand management of the native trees and shrubs due touncertainties about land and tree tenure. Local governments,farming and pastoralist communities need to resolve theseissues and create a supportive social and political environmentfor sustainable management and conservation of native treespecies in the region.
5. Conclusion
The clines in growth and survival of P. africana provenancesprovide basic information for tree improvement and conservationprograms in the West African Sahel, but more research is necessaryfor this species. In addition, studies should be initiated toinvestigate genetic variation among and within natural popula-tions of other native tree species in the region, especially thosethreatened with local or regional extinction. This research providestechnical recommendations for tree improvement and conserva-tion strategies, but social and political changes must also befostered in order to facilitate the sustainable management andconservation of native tree species by farming and pastoralistcommunities in the region.
Acknowledgements
Seed collection and trial establishment were supported bygenerous grants to ICRAF from the International Fund forAgricultural Development (IFAD). The authors thank IFAD fortheir financial support; Boube Aw and Hama Moumouni fromICRAF for coordinating the seed collection and trial establishment;Idrissa Habi from ICRISAT for assistance in data collection; I.
References
Agboola, D.D., 2004. Prosopis africana (Mimosaceae): stems, roots and seeds ineconomy of the savannah areas of Nigeria. Econ. Bot. 58 (Suppl.), S34–S4
Akinnagbe, A., Oni, O., 2007. Quantitative variations in the growth of proseedlings of Prosopis africana (Guill., Perrott. and Rich.) plus trees in NigeriaJ. Biotechnol. 6, 359–363.
Andrew, S.M., Maliondo, S.M.S., Mtika, J., Msanga, H.P., 2004. Growth performof Azadirachta indica provenances in Morogoro, Tanzania. J. Trop. For. Sc328–335.
Appiah, M., 2003. Domestication of an indigenous tropical forest tree: silviculand socio-economic studies on Iroko (Melicia excelsa) in Ghana. UniversiHelsinki Tropical Forestry Reports 25, Helsinki.
Arbonnier, M., 2002. Arbres Arbustes et Lianes des Zones Seches d’Afriqul’Ouest. Centre de Cooperation Internationale en Recherche Agronomiquele Developpement (CIRAD)/Museum National d’Histoire Naturelle (MNMontpellier/Paris.
Aremu, M.O., Olonisakin, A., Atolaye, B.O., Ogbu, C.F., 2006. Some nutritionalfunctional studies of Prosopis africana. Electron. J. Environ. Agric. Food Che1640–1648.
Arnold, R., 2004. Khaya senegalensis—current use from its natural range anpotential in Sri Lanka and elsewhere in Asia. in: Bevege, D.I., Bristow, M., NiD.G., Skelton, D.J., (Eds.), Prospects for High-value Hardwood Timber Plations in the Dry Tropics of Northern Australia, Proc. Workshop October 192004, Mareeba, Queensland. Published as CD-ROM. Private Forestry NQueensland Association Inc., Kairi.
Barminas, J.T., Maina, H.M., Ali, J., 1998. Nutrient content of Prosopis africana sPlant Foods Hum. Nutr. 52, 325–328.
Bastide, B., Diallo, B., 1996. Comparaison de provenances de Faidherbia albidplantations au Burkina Faso. Taux de survie et vitesse de croissance juvdans les zones nord et sud-soudanienne. In: Peltier, R. (Ed.), Les parFaidherbia. Cahier Scientifique No. 12. Centre de Cooperation Internatioen Recherche Agronomique pour le Developpement, Departement F(CIRAD-Foret), Montpellier, pp. 259–268.
Bessega, C., Ferreyra, L., Julio, N., Montoya, S., Saidman, B., Vilardi, J.C., 2000. Msystem parameters in species of genus Prosopis (Leguminosae). Hereditas19–27.
Billand, A., De Framond, H., 1993. Variabilite genetique d’Acacia albida (synonFaidherbia albida) en essais comparatifs de provenance au Burkina FasoRiedacker, A., Dreyer, E., Pafadnam, C., Joly, H., Bory, G. (Eds.), PhysiologieArbres et Arbustes en Zones Arides et Semi-Arides. John Libbey Eurotext, Ppp. 235–248.
Bowen, M.R., Howland, P., Last, F.T., Leakey, R.R.B., Longman, K.A., 1977. Triplocscleroxylon: its conservation and future improvement. Forest Genetic ResouInformation No. 6, Food and Agriculture Organization (FAO) of the UnNations, Rome.
Cascante, A., Quesada, M., Lobo, J.A., Fuchs, E.J., 2002. Effects of dry tropical ffragmentation on the reproductive success and genetic structure of theSamanea saman. Conserv. Biol. 16, 137–147.
Dangasuk, O.G., Seurei, P., Gudu, S., 1997. Genetic variation in seed and seetraits of 12 African provenances of Faidherbia albida (del.) A. Chev. at LodKenya. Agrofor. Syst. 37, 133–141.
Dawson, I.K., Powell, W., 1999. Genetic variation in the afromontane tree Prafricana, an endangered medicinal species. Mol. Ecol. 8, 151–156.
Dhillon, R.S., Hooda, M.S., Chopra, D., Arya, S., 2003. Studies on floral biologybreeding behaviour of Prosopis cineraria (L.) Druce (Khejri). Forrage Res. 2975.
Diallo, B.O., Sanou, J., Some Dao, M., Cao, T.V., Asimi, S., 2000. Rapport d’actfinal. Projet FAC N894/CD/78/BKA Volet 1: Amelioration genetique desneux soudano-saheliens. Centre National de la Recherche ScientifiquTechnologique (CNRST); Institut National de l’Environnement etRecherches Agricoles (INERA); Centre de Cooperation InternationalRecherche Agronomique pour le Developpement, Departement F(CIRAD-Foret); Cooperation Francais, Mission de Cooperation et d’AcCulturelle, Ouagadougou.
FAO, 2007. FAO soil map. Food and Agriculture Organization (FAO) of the UnNations. www.mapjourney.com/sahel/zoom/zoom_003_.htm (accessedNovember 2007).
Gerardin, P., Neya, B., Dumarcay, S., Petrisanns, M., Serraj, M., Huber, F., 2Contribution of gums to natural durability of Prosopis africana heartwHolzforschung 58, 39–44.
Halliday, J., 1984. Register of nodulation reports for leguminous trees and oarboreal genera with nitrogen fixing members. Nitrogen Fixing Tree Res. Re38–56.
Hamrick, J.L., Godt, M.J., Sherman Broyles, S.L., 1992. Factors influencing levegenetic diversity in woody plant species. New For. 6, 95–124.
Hardner, C.M., Vaillancourt, R.E., Potts, B.M., 1996. Stand density influences
J.C. Weber et al. / Forest Ecology and Management xxx (2008) xxx–xxx
ulus.
ahel891–
solu-. 25,
Maıkano from ICRISAT for preparing a summary of rainfall data atthe ICRISAT Sahelian Centre; Meshack Nyabenge from ICRAF forpreparing the map used in Fig. 1; Nancy Mandel from the USDAForest Service for reviewing the statistical methods; and theanonymous reviewers for their useful comments on an earlierversion of this manuscript.
Please cite this article in press as: Weber, J.C. et al., Growth and survrainfall gradients in the West African Sahel, Forest Ecol. Manage. (
crossing rate and growth of open-pollinated families of Eucalyptus globSilvae Genet. 45, 226–228.
Held, I.M., Delworth, T.L., Lu, J., Findell, K.L., Knutson, T.R., 2005. Simulation of Sdrought in the 20th and 21st centuries. Proc. Natl. Acad. Sci. U.S.A. 102, 1717896.
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.J., Jarvis, A., 2005. Very high retion interpolated climate surfaces for global land areas. Int. J. Climatol1965–1978.
ival of Prosopis africana provenances tested in Niger and related to2008), doi:10.1016/j.foreco.2008.05.004

Hu, Xo
ICRAtAt
KalinhInttW
KhasRZ
Langd1
LangLoha
inL
MorgAV
MuraloL
Nasofr2
Nygas1
Oforim2
OforisM
PasieMg
PasieA
Pedea1
RaddfiS
RaddcB
Ræbip
8
G Model
FORECO-11100; No of Pages 8
Plerai
.S., Li, B., 2002. Linking evolutionary quantitative genetics to the conservationf genetic resources in natural forest populations. Silvae Genet. 51, 177–183.F-IFAD, 2006. Renforceme nt des strategies de subsistance a travers une utilisa-ion et une gestion ameliorees des parcs agroforestiers au Sahel. In: Rapportnnuel du Projet ICRAF TAG 799, World Agroforestry Centre (ICRAF)/Interna-
ional Fund for Agricultural Development (IFAD), Bamako/Rome.ganire, A., Weber, J.C., Uwamariya, A., Kone, B., 2007. Improving rural liveli-oods through domestication of indigenous fruit trees in parklands of the Sahel.: Akinnifesi, F.K., Leakey, R.R.B., Ajayi, O.C., Sileshi, G., Tchoundjeu, Z., Ma-
acala, P., Kwesiga, F.R. (Eds.), Indigenous Fruit Trees in the Tropics: Domes-ication, Utilization and Commercialization. CAB International Publishing,
allingford, Oxfordshire, pp. 186–203.a, P.D., Li, P., Vallee, G., Magnussen, S., Bousquet, J., 1995. Early evaluation ofacosperma auriculiforme and R. mangium provenance trials on four sites inaire. For. Ecol. Manage. 78, 99–113.at, D.K., Kariuki, J.G., 2004. Variations in heartwood proportions and basicensity between provenances of Eucalyptus grandis in Kenya. J. Trop. For. Sci. 16,79–186.let, O., 1971. Two hundred years of genecology. Taxon 20, 653–722., A., Tigabu, M., Teketay, D., Lundkvist, K., Fries, A., 2006. Provenance variation
seed morphometric traits, germination and seedling growth of Cordia africanaam. New For. 32, 71–86.enstern, E.K., 1996. Geographic Variation in Forest Trees—Genetic Basis andpplication of Knowledge in Silviculture. University of British Columbia Press,ancouver.wski, D.A., Gunatilleke, I.A.U.N., Bawa, K.S., 1994. The effects of selectivegging and inbreeding on Shorea megistrophylla (Dipterocarpaceae) from Sri
anka. Conserv. Biol. 8, 997–1002.n, J.D., Hamrick, J.L., 1997. Reproductive and genetic consequences of forestagmentation: two case studies of neotropical canopy trees. J. Hered. 88, 264–76.rd, R., Elving, B., 2000. Stem basic density and bark proportion of 45 woodypecies in young savanna coppice forests in Burkina Faso. Ann. For. Sci. 57, 143–53., D.A., Cobbinah, J.R., 2007. Integrated approach for conservation and manage-ent of genetic resources of Milicia species in West Africa. For. Ecol. Manage.
38, 1–6., D.A., Opuni-Frimpong, E., Cobbinah, J.R., 2007. Provenance variation in Khayapecies for growth and resistance to shoot borer Hypsipyla robusta. For. Ecol.anage. 242, 438–443.
cznik, N.M., Felker, P., Harris, P.J.C., Harsh, L.N., Cruz, G., Tewari, J.C., Cadoret, K.,aldonado, L.J., 2001. The Prosopis juliflora–Prosopis pallida complex: A Mono-
raph. Henry Doubleday Research Association (HDRA), Coventry.cznik, N.M., Harris, P.J.C., Smith, S.J., 2004. Identifying Tropical Prosopis Species:Field Guide. Henry Doubleday Research Association (HDRA), Coventry.
rsen, A.P., Hansen, J.K., Mtika, J.M., Msangi, T.H., 2007. Growth, stem quality andge–age correlations in a teak provenance trial in Tanzania. Silvae Genet. 56,42–148.ad, E.Y., 2007. Ecophysiological and genetic variation in seedling traits and inrst-year field performance of eight Acacia senegal provenances in the Blue Nile,udan. New For. 34, 207–222.ad, E.Y., Luukkanen, O., 2006. Adaptive genetic variation in water-use effi-iency and gum yield in Acacia senegal provenances grown on clay soil in thelue Nile region, Sudan. For. Ecol. Manage. 226, 219–229.ld, A., Diallo, B.O., Graudal, L., Dao, M., Sanou, J., 2003a. Evaluation of arovenance trial of Acacia senegal at Djibo, Burkina Faso. Trial no. 5 in the arid
zone series. Results and Documentation No. 7. Danida Forest Seed Centre,Humlebaek.
Ræbild, A., Diallo, B.O., Graudal, L., Dao, M., Sanou, J., 2003b. Evaluation of aprovenance trial of Acacia nilotica and A. tortilis at Gonse, Burkina Faso. Trialno. 11 in the arid zone series. Results and Documentation No. 10. Danida ForestSeed Centre, Humlebaek.
Ræbild, A., Diallo, B.O., Graudal, L., Dao, M., Sanou, J., 2003c. Evaluation of a speciesand provenance trial of Prosopis cineraria, P. pallida and Acacia senegal at Djibo,Burkina Faso. Trial no. 6 in the arid zone series. Results and Documentation No.8. Danida Forest Seed Centre, Humlebaek.
Ræbild, A., Diallo, B.O., Graudal, L., Dao, M., Sanou, J., 2003d. Evaluation of a speciesand provenance trial of Prosopis at Gonse, Burkina Faso. Trial no. 14 in the aridzone series. Results and Documentation No. 11. Danida Forest Seed Centre,Humlebaek.
Roupsard, O., Joly, H.I., Dreyer, E., 1998. Variability of initial growth, water-useefficiency and carbon isotope discrimination in seedlings of Faidherbia albida(Del.) A. Chev., a multipurpose tree of semi-arid Africa. Provenance and droughteffects. Ann. Sci. For. 55, 329–348.
SAS Institute Inc., 2004. SAS/STAT Users’ Guide, Version 9.1. SAS Institute Inc., Cary.Sivakumar, M.V.K., Gnoumou, F., 1987. Agroclimatologie de l’Afrique de l’Ouest: le
Burkina Faso Bulletin d’information No. 23. International Crops Research Insti-tute for the Semi-Arid Tropics (ICRISAT), Patancheru.
Sivakumar, M.V.K., Maidoukia, A., Stern, R.D., 1993. Agroclimatologie de l’Afrique del’Ouest: le Nıger Bulletin d’information No. 5. International Crops ResearchInstitute for the Semi-Arid Tropics (ICRISAT), Patancheru.
Sniezko, R.A., Stewart, H.T.L., 1989. Range-wide provenance variation in growth andnutrition of Acacia albida seedlings propagated in Zimbabwe. For. Ecol. Manage.27, 179–197.
Sotelo Montes, C., Beaulieu, J., Hernandez, R.E., 2007a. Genetic variation in woodmechanical properties of Calycophyllum spruceanum at an early age in thePeruvian Amazon. Wood Fiber Sci. 39, 578–590.
Sotelo Montes, C., Beaulieu, J., Hernandez, R.E., 2007b. Genetic variation in woodshrinkage, and its correlations with tree growth and wood density of Calyco-phyllum spruceanum at an early age in the Peruvian Amazon. Can. J. For. Res. 37,966–976.
Tchoundjeu, Z., Weber, J., Guarino, L., 1997. Germplasm collections of endangeredagroforestry tree species: the case of Prosopis africana in the semi-arid lowlandsof West Africa. Agrofor. Syst. 39, 91–100.
Toro, H., 2002. Pollination of Prosopis tamarugo in the Atacama desert with remarkson the roles of associated plants. In: Kevan, P., Imperatriz Fonseca, V.L. (Eds.),Pollinating Bees—The Conservation Link Between Agriculture and Nature.Ministry of Environment, Brasilia, pp. 267–273.
Tsoumis, G.T., 1991. Science and Technology of Wood: Structure, Properties,Utilization. Van Nostrand Reinhold, New York.
Tybirk, K., 1991. Regeneration of Woody Legumes in Sahel. Aarhus University Press,Aarhus.
Wanyancha, J.M., Mills, W.R., Gwaze, D.P., 1994. Genetic variation in Acacia albida(Faidherbia albida) and its agroforestry potential in Zimbabwe. For. Ecol. Man-age. 64, 127–134.
Weber, J.C., Sotelo Montes, C., 2008. Variation in tree growth and wood density ofGuazuma crinita Mart. in the Peruvian Amazon. New For. 36, 29–52.
Wolde-Mieskel, E., Sinclair, F.L., 2000. Growth variability in a Senegalese prove-nance of Acacia nilotica subsp. tomentosa. Agrofor. Syst. 48, 207–213.
Yatich, T., Kalinganire, A., Alinon, K., Weber, J.C., Dakouo, J.-M., Samake, O., Sangare,S., 2008. Moving beyond forestry laws in Sahelian countries. Policy brief, WorldAgroforestry Centre (ICRAF), Nairobi.
J.C. Weber et al. / Forest Ecology and Management xxx (2008) xxx–xxx
ase cite this article in press as: Weber, J.C. et al., Growth and survival of Prosopis africana provenances tested in Niger and related tonfall gradients in the West African Sahel, Forest Ecol. Manage. (2008), doi:10.1016/j.foreco.2008.05.004
Copyright © 2022 FDOKUMEN




















