GPR39 Splice Variants Versus Antisense Gene LYPD1: Expression and Regulation in Gastrointestinal...
14
GPR39 Splice Variants Versus Antisense Gene LYPD1: Expression and Regulation in Gastrointestinal Tract, Endocrine Pancreas, Liver, and White Adipose Tissue Kristoffer L. Egerod, Birgitte Holst, Pia S. Petersen, Jacob B. Hansen, Jan Mulder, Tomas Ho ¨ kfelt, and Thue W. Schwartz Laboratory for Molecular Pharmacology, Department of Neuroscience and Pharmacology (K.L.E., B.H., P.S.P., T.W.S.) and Department of Biomedical Sciences (J.B.H.), University of Copenhagen, DK-2200 Copenhagen N, Denmark; Department of Neuroscience (J.M., T.H.), Karolinska Institutet, 171 77 Stockholm, Sweden; and 7TM Pharma A/S (T.W.S.), DK-2790 Hoersholm, Denmark G protein-coupled receptor 39 (GPR39) is a consti- tutively active, orphan member of the ghrelin re- ceptor family that is activated by zinc ions. GPR39 is here described to be expressed in a full-length, biologically active seven-transmembrane form, GPR39-1a, as well as in a truncated splice variant five-transmembrane form, GPR39-1b. The 3 exon of the GPR39 gene overlaps with an antisense gene called LYPD1 (Ly-6/PLAUR domain containing 1). Quantitative RT-PCR analysis demonstrated that GPR39-1a is expressed selectively throughout the gastrointestinal tract, including the liver and pan- creas as well as in the kidney and adipose tissue, whereas the truncated GPR39-1b form has a more broad expression pattern, including the central nervous system but with highest expression in the stomach and small intestine. In contrast, the LYPD1 antisense gene is highly expressed throughout the central nervous system as charac- terized with both quantitative RT-PCR and in situ hybridization analysis. A functional analysis of the GPR39 promoter region identified sites for the he- patocyte nuclear factors 1 and 4 (HNF-1 and -4) and specificity protein 1 (SP1) transcription factors as being important for the expression of GPR39. In vivo experiments in rats demonstrated that GPR39 is up-regulated in adipose tissue dur- ing fasting and in response to streptozotocin treat- ment, although its expression is kept constant in the liver from the same animals. GPR39-1a was expressed in white but not brown adipose tissue and was down-regulated during adipocyte differ- entiation of fibroblasts. It is concluded that the transcriptional control mechanism, the tissue ex- pression pattern, and in vivo response to physio- logical stimuli all indicate that the GPR39 receptor very likely is of importance for the function of a number of metabolic organs, including the liver, gastrointestinal tract, pancreas, and adipose tissue. (Molecular Endocrinology 21: 1685–1698, 2007) G PROTEIN-COUPLED RECEPTOR 39 (GPR39) was originally cloned in 1997 along with GPR38 as two novel orphan seven-transmembrane (7TM), G protein-coupled receptors showing high structural similarity to the GH secretagogue receptor (GHSR) (1), which later was shown to be the receptor for the gastrointestinal (GI) tract hormone ghrelin (2). The best established function for ghrelin is its action as an en- docrine hunger signal from the upper GI tract to the brainstem and hypothalamus to regulate food intake and energy homeostasis (3). GPR38 was shown to be the receptor for motilin, i.e. another gut hormone that mainly regulates interdigestive contractions and gut motility (4). Other members of the ghrelin receptor family are the two neuromedin U receptors and the two neurotensin receptors (Fig. 1A). Neuromedin U and neurotensin are regulatory peptides that, like gh- relin and motilin, both have been implicated in, for example, control of food intake and GI tract function (5, 6). For a decade, GPR39 remained a rather unnoticed orphan member of the ghrelin receptor family besides the fact that it was reported that GPR39, like the ghrelin and neurotensin R 2 receptors, signals with a rather high constitutive, ligand-independent activity and that it is First Published Online May 8, 2007 Abbreviations: BAT, Brown adipose tissue; CNS, central nervous system; Ct, cycle threshold; FABP4, fatty acid bind- ing protein 4; GHSR, GH secretagogue receptor; GI, gastro- intestinal; GPR, G protein-coupled receptor; HNF, hepato- cyte nuclear factor; MEF, mouse embryo fibroblast; MODY, maturity-onset diabetes of the young; PGC-1, peroxisome proliferator-activated receptor coactivator 1; QPCR, quantitative RT-PCR; RACE, rapid amplification of cDNA ends; Rb, retinoblastoma protein; SP1, specificity protein 1; STZ, streptozotocin; 5TM, five transmembrane; 7TM, seven transmembrane; UCP1, uncoupling protein 1; WAT, white adipose tissue. Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving the endocrine community. 0888-8809/07/$15.00/0 Molecular Endocrinology 21(7):1685–1698 Printed in U.S.A. Copyright © 2007 by The Endocrine Society doi: 10.1210/me.2007-0055 1685
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of GPR39 Splice Variants Versus Antisense Gene LYPD1: Expression and Regulation in Gastrointestinal...

GPR39 Splice Variants Versus Antisense GeneLYPD1: Expression and Regulation inGastrointestinal Tract, Endocrine Pancreas, Liver,and White Adipose Tissue
Kristoffer L. Egerod, Birgitte Holst, Pia S. Petersen, Jacob B. Hansen, Jan Mulder, Tomas Hokfelt,and Thue W. Schwartz
Laboratory for Molecular Pharmacology, Department of Neuroscience and Pharmacology (K.L.E.,B.H., P.S.P., T.W.S.) and Department of Biomedical Sciences (J.B.H.), University of Copenhagen,DK-2200 Copenhagen N, Denmark; Department of Neuroscience (J.M., T.H.), Karolinska Institutet,171 77 Stockholm, Sweden; and 7TM Pharma A/S (T.W.S.), DK-2790 Hoersholm, Denmark
G protein-coupled receptor 39 (GPR39) is a consti-tutively active, orphan member of the ghrelin re-ceptor family that is activated by zinc ions. GPR39is here described to be expressed in a full-length,biologically active seven-transmembrane form,GPR39-1a, as well as in a truncated splice variantfive-transmembrane form, GPR39-1b. The 3� exonof the GPR39 gene overlaps with an antisense genecalled LYPD1 (Ly-6/PLAUR domain containing 1).Quantitative RT-PCR analysis demonstrated thatGPR39-1a is expressed selectively throughout thegastrointestinal tract, including the liver and pan-creas as well as in the kidney and adipose tissue,whereas the truncated GPR39-1b form has a morebroad expression pattern, including the centralnervous system but with highest expression in thestomach and small intestine. In contrast, theLYPD1 antisense gene is highly expressedthroughout the central nervous system as charac-terized with both quantitative RT-PCR and in situhybridization analysis. A functional analysis of the
GPR39 promoter region identified sites for the he-patocyte nuclear factors 1� and 4� (HNF-1� and-4�) and specificity protein 1 (SP1) transcriptionfactors as being important for the expression ofGPR39. In vivo experiments in rats demonstratedthat GPR39 is up-regulated in adipose tissue dur-ing fasting and in response to streptozotocin treat-ment, although its expression is kept constant inthe liver from the same animals. GPR39-1a wasexpressed in white but not brown adipose tissueand was down-regulated during adipocyte differ-entiation of fibroblasts. It is concluded that thetranscriptional control mechanism, the tissue ex-pression pattern, and in vivo response to physio-logical stimuli all indicate that the GPR39 receptorvery likely is of importance for the function of anumber of metabolic organs, including the liver,gastrointestinal tract, pancreas, and adiposetissue. (Molecular Endocrinology 21: 1685–1698,2007)
G PROTEIN-COUPLED RECEPTOR 39 (GPR39)was originally cloned in 1997 along with GPR38
as two novel orphan seven-transmembrane (7TM), Gprotein-coupled receptors showing high structuralsimilarity to the GH secretagogue receptor (GHSR) (1),
which later was shown to be the receptor for thegastrointestinal (GI) tract hormone ghrelin (2). The bestestablished function for ghrelin is its action as an en-docrine hunger signal from the upper GI tract to thebrainstem and hypothalamus to regulate food intakeand energy homeostasis (3). GPR38 was shown to bethe receptor for motilin, i.e. another gut hormone thatmainly regulates interdigestive contractions and gutmotility (4). Other members of the ghrelin receptorfamily are the two neuromedin U receptors and thetwo neurotensin receptors (Fig. 1A). Neuromedin Uand neurotensin are regulatory peptides that, like gh-relin and motilin, both have been implicated in, forexample, control of food intake and GI tract function(5, 6).
For a decade, GPR39 remained a rather unnoticedorphan member of the ghrelin receptor family besidesthe fact that it was reported that GPR39, like the ghrelinand neurotensin R2 receptors, signals with a rather highconstitutive, ligand-independent activity and that it is
First Published Online May 8, 2007Abbreviations: BAT, Brown adipose tissue; CNS, central
nervous system; Ct, cycle threshold; FABP4, fatty acid bind-ing protein 4; GHSR, GH secretagogue receptor; GI, gastro-intestinal; GPR, G protein-coupled receptor; HNF, hepato-cyte nuclear factor; MEF, mouse embryo fibroblast; MODY,maturity-onset diabetes of the young; PGC-1�, peroxisomeproliferator-activated receptor � coactivator 1�; QPCR,quantitative RT-PCR; RACE, rapid amplification of cDNAends; Rb, retinoblastoma protein; SP1, specificity protein 1;STZ, streptozotocin; 5TM, five transmembrane; 7TM, seventransmembrane; UCP1, uncoupling protein 1; WAT, whiteadipose tissue.
Molecular Endocrinology is published monthly by TheEndocrine Society (http://www.endo-society.org), theforemost professional society serving the endocrinecommunity.
0888-8809/07/$15.00/0 Molecular Endocrinology 21(7):1685–1698Printed in U.S.A. Copyright © 2007 by The Endocrine Society
doi: 10.1210/me.2007-0055
1685

activated by zinc ions (7). In 2005, GPR39 was sug-gested to be the receptor for a peptide fragment from theghrelin precursor named obestatin, which was proposedto be a gut hormone having the opposite effects on foodintake and GI tract function as ghrelin (8). However, aseries of subsequent studies from a number of indepen-dent groups have not been able to demonstrate anybinding of obestatin to GPR39 or any stimulatory func-tion of the obestatin peptide on GPR39 (9–11). Thus, itcan today be concluded that GPR39 is not the receptorfor obestatin. [This is also acknowledged by A. Hsueh(40).]
Originally, GPR39 was, by Northern blot analysis,shown to be widely expressed throughout the wholebody (1), which probably dampened the interest intrying to deorphanize this receptor. However, this ex-pression analysis, which was performed with full-length GPR39 cDNA probes, could very well havebeen complicated or even invalidated by the occur-rence of a gene called LYPD1 (Ly-6/PLAUR domaincontaining 1), which, in a large-scale systematic
search for secreted proteins, was identified to be en-coded at the antisense DNA strand corresponding tothe genomic locus of GPR39 (1, 12). Moreover, severalmembers of the ghrelin receptor family have beenshown to be expressed in different splice variantforms, i.e. corresponding to both a full-length 7TMform and a truncated five-transmembrane (5TM) form(4, 13), which also could have complicated the originalexpression analysis. Variations in the control of ex-pression in response to physiological stimuli are par-ticularly interesting for constitutively active receptors,because, for example, an increased signaling activitywill be directly correlated to an increased receptorexpression, independent of the hormone or transmit-ter. This has been demonstrated recently for the con-stitutively active ghrelin receptor and the CB1 canna-binoid receptors, which both are closely regulated andfluctuate with physiological stimuli such as fasting (14–16). Importantly, this was not the case for nonconsti-tutively active, “normal” 7TM receptors, in which it isthe expression of their ligands, and not the receptor,
Fig. 1. The Ghrelin Receptor FamilyA shows a schematic phylogenic tree of the ghrelin receptor family. The constitutively active receptors are highlighted with red
circles. B shows a serpentine model of GPR39 in which residues highlighted in black indicate residues that are most highlyconserved within the family and residues highlighted in gray indicate residues that are generally conserved. The position of theintron, which is conserved throughout the ghrelin receptor family, is indicated by an orange line. NMU, Neuromedin U; NTS,neurotensin.
1686 Mol Endocrinol, July 2007, 21(7):1685–1698 Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target
that is regulated, i.e. cholecystokinin, neuropeptide Y,and orexin (16).
In the present study, we characterize the humanGPR39 gene, its promoter, and its expression patternin peripheral tissues and in the central nervous system(CNS) of rats and have studied changes in the expres-sion of GPR39 in vivo in response to starvation andstreptozotocin (STZ) treatment in liver and adiposetissue, as well as during white and brown adipocytedifferentiation of mouse fibroblasts.
RESULTS
Genomic Structure of GPR39
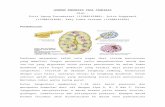
Analysis of the genomic structure of the humanGPR39 revealed that it consists of two exons sepa-rated by a very large intron of approximately 200 kb(Fig. 2A). Interestingly, an intron is found at the sameposition also in the genes for the ghrelin receptor andthe motilin receptor, which in both cases give rise totruncated, 5TM splice variant forms of these recep-
tors, i.e. GHSR-1b and motilin receptor 1b (4, 13).Within the intron of the GPR39 gene, we identified apotential alternative polyadenylation site, indicatingthat GPR39, like GHSR and the motilin receptor, couldpossibly be expressed in two splice variant forms. Onthe antisense strand corresponding to the 3�exon en-coding TM segments VI and VII of GPR39 an antisensegene, LYPD1, is expressed in concordance with arecent systematic search for secreted proteins (12).
Analysis of GPR39 Expression
PCR analysis verified the notion that GPR39 is ex-pressed in two splice variant forms, which we havenamed GPR39-1a, corresponding to the full-length7TM receptor, and GPR39-1b, corresponding to the5TM truncated form, in analogy with the nomenclatureused for the ghrelin receptor (Fig. 2A). The nucleotidesequence of the previously unrecognized GPR39-1bhas been submitted to Entrez GenBank under theaccession nos. BK005609 (Homo sapiens), BK005610(Rattus norvegicus), BK005608 (Mus musculus), andBK005612 (Gallus gallus).
Fig. 2. The Genomic Structure of the GPR39 Locus and Tissue-Specific Expression of the Three Gene ProductsA, The splice variation of GPR39 was predicted from the genomic sequence and later verified through QPCR. Therefore, GPR39
is now annotated as GPR39-1a for the full-length functional 7TM receptor (shown in dark green), whereas GPR39-1b representsan unspliced 5TM receptor form (shown in light green). In the same genomic locus, an antisense gene LYPD1 is expressed (shownin orange), giving rise to overlapping sense antisense transcripts of both gene 3� ends. B, The relative expression of GPR39-1a,GPR39-1b, and LYPD1 in peripheral organs as well as in various regions of the CNS. The expressions were measured relativeto �-actin and normalized to the tissue showing highest expression of the gene. BSTN, Bed nucleus of the stria terminalis.
Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target Mol Endocrinol, July 2007, 21(7):1685–1698 1687

When the GPR39 was originally cloned, the genomicstructure shown in Fig. 2A was not yet available, andthe interpretation of Northern blot analysis was thatthe orphan receptor was expressed throughout thebody, including multiple areas of the brain (1). How-ever, as determined by specific quantitative RT-PCR(QPCR), the full-length GPR39-1a is in fact not ex-pressed to any significant degree in the CNS (Fig. 2B).Instead, GPR39-1a is highly expressed in the liver, theGI tract from the stomach to the colon, the pancreas,the kidney, as well as in adipose tissue, and to a lowerdegree in the spleen, thyroid, lung, and heart. Thetruncated GPR39-1b form was found to be expressedmore widely but again especially in the stomach andthe small intestine (Fig. 2B). In contrast, the antisensegene LYPD1 showed a very different expression pat-tern because it was found highly expressed in all brainregions tested, with the highest levels observed inamygdala and septum. Although LYPD1 evidently wasexpressed to a much higher degree in the CNS thananywhere else (Fig. 2B), the expression in peripheraltissues was large enough to be readily detectedthrough QPCR, with the heart showing the highestexpression outside the CNS (Fig. 2B). As shown in Fig.3A, in which the QPCR analysis in selected tissueswas performed with the three gene products againsteach other instead of individually against �-actin, theexpression of GPR39-1a is approximately 100-foldhigher than the expression of LYPD1 in, for example,the liver and GI tract, whereas the expression ofLYPD1 is between 100- and 1000-fold higher than theexpression of GPR39-1a in various brain regions, thusrevealing a ratio shift between the two genes of ap-proximately 1 million in different tissues (Figs. 2B and3A). It should be noted that the expression ofGPR39-1a in some brain tissues is so low that it ap-proaches the detection limit for the method. A similaranalysis showed that the expression of the truncatedGPR39-1b is approximately 10-fold lower than theexpression of the full-length GPR39-1a in the liver and
kidney but relatively similar in the stomach and smallintestine, in which both gene products are high ex-pressed. In contrast, the expression of GPR39-1b isapproximately 10-fold higher than that of the 1a formin, for example, the hippocampus and amygdala(Fig. 3B).
In situ hybridization analysis using antisense probesselective for LYPD1 revealed expression of this genein many brain nuclei (Fig. 4). Strong labeling was foundin the lateral septum, islets of Calleja, basolateral,basomedial, and medial, intercalated amygdala nuclei,CA1 of the hippocampus, pyramidal cells in layer IVand neurons in layer VI of the cortex, as well as theparaventricular nucleus of the thalamus. Other areas,including the caudate putamen, globus pallidus, bednucleus of the stria terminalis, the arcuate nucleus,and several other hypothalamic nuclei, revealed aclear but moderate labeling for LYPD1. LYPD1 seemsto be expressed in a subpopulation of neurons, al-though expression in non-neuronal cells can not beexcluded. In the investigated brain areas, no specificlabeling for GPR39-1a could be observed (Fig. 4).
Analysis of the GPR39 Promoter Region
An automated, systematic analysis had indicated thata transcriptional start site is located 469 nucleotidesupstream from the translation initiating site in theGPR39 gene (17). Based on this, we performed ananalysis of the GPR39 promoter region through a se-ries of 5� deletion constructs using a luciferase re-porter assay and expression in both U-138 cells,which have been shown previously to express GPR39(18), and the liver cell line Hep G2.
In the U-138 cells, maximal promoter activity was ob-served in constructs encompassing the region from�673 to �14, whereas in the Hep G2 cells the constructcorresponding to the region from �573 to �14 showedmaximal promoter activity, indicating that the minimalpromoter varies between the two cell lines (Fig. 5). The
Fig. 3. Relative Expression Levels for GPR39-1a, GPR39-1b, and LYPD1Using a direct �Ct method (for details, see Materials and Methods), the ratio of GPR39-1a vs. LYPD1 expression (A) and
GPR39-1b vs. GPR39-1a expression (B) were determined in selected tissues: liver, stomach, kidney, small intestines, heart,hippocampus, hypothalamus, amygdala, and septum. The expression levels have been normalized to the expression in the heart,in which all three gene products are expressed to a reasonable level.
1688 Mol Endocrinol, July 2007, 21(7):1685–1698 Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target

results also indicated that the region �673 to �573holds negative regulatory elements that are used exclu-sively in Hep G2 cells and not in the U-138 cells, in whichno negative regulations were observed because the tran-scriptional activity just continued at a plateau when theconstructs exceed position �673 (Fig. 5).
As shown in Fig. 6, 5� rapid amplification of cDNAends (RACE) PCR surprisingly identified two alterna-tive transcriptional start sites located closer to thetranslational initiation site than expected, i.e. one atposition �196, which was used in both Hep G2 andU-138 cells, and one at position �396, which wasused exclusively in the Hep G2 cell line. The transcrip-tional start site proposed to be located in position�469 as part of the automated annotation performedon 21,243 genes (17) was not observed to be used inany of the cell lines.
The promoter region of GPR39 was analyzed insilico, using the programs Match (BIOBASE, Wolfen-buettel, Germany) and ConSite (Center for Genom-
ics and Bioinformatics, Karolinska Institute, Stock-holm, Sweden) to locate potential binding sites forregulating transcription factors. No classical TATAbox could be identified, which often is the casewhen the transcription is driven by, for example,specificity protein 1 (SP1) (19). In accordance withthis notion, regions of high guanine and cytosinecontent holding several SP1 binding sites were iden-tified. One of these SP1 sites located at aroundposition �598 is conserved between human androdent GPR39. Several potential binding sites formetabolism-related transcription factors were iden-tified (Fig. 7A), of which especially the sites for he-patocyte nuclear factor 1� (HNF-1�) and HNF-4�appeared interesting in view of the QPCR-basedtissue expression pattern for GPR39-1as presentedabove (Fig. 2). The HNF-1�, HNF-4�, and SP1 bind-ing sites were characterized experimentally throughmutagenesis in which nucleotides were mutated todisrupt the binding sites for the relevant transcrip-
Fig. 5. Functional Analysis of the Transcriptional Activity of GPR39 Promoter Region: Effect of 5� Deletion MutagenesisThe relative promoter activity was measured in a series of constructs containing a decreasing length of the 5� region of the
promoter. At least three independent assays were performed, and the luciferase activity was normalized to the basic luciferasevector containing no promoter elements.
Fig. 4. Autoradiograms of LYPD1 and GPR39-1a mRNA Expression in Brain SectionsLYPD1 at rostrocaudal levels 0.2 and �3.6 mm from bregma and GPR39-1a at rostrocaudal level �3.6 mm from bregma.
Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target Mol Endocrinol, July 2007, 21(7):1685–1698 1689

tion factor using the matrices from the TRANSFAC6public database.
As observed in the truncation analysis, certain dif-ferences were also observed between the two celllines concerning the effects of the point mutations inthe GPR39 promoter. Thus, whereas selective disrup-tion of the HNF-1� site around position �468 impaired
the transcriptional activity of the GPR39 promoter sig-nificantly in both U-138 and Hep G2 cells, mutation ofthe HNF-4� site around position �1253 only impairedthe promoter activity in the U-138 cells and not in theHep G2 cells (Fig. 7B). Interestingly, in the gene for theHNF-4� transcription factor, a conserved aspartic acidin the DNA binding region of HNF-4� is mutated into
Fig. 7. Functional Analysis of the Transcriptional Activity of GPR39 Promoter Region: Effect of Point Mutations in Proposed Sitefor Selected, “Metabolic” Transcription Factors
A, Overview of the promoter region of GPR39. Various transcription factor binding sites are shown on either sense or antisensestrand. Disruptive mutations were designed to demolish the consensus for transcription factor binding. Mutations are shown inred, and the potential transcription factor is indicated: HNF-1�, �; HNF-4�, ✦ ; and SP1, ¨. At least three independent assays wereperformed, and the luciferase activity was normalized to the construct pGL3-F7 encompassing the wild-type (WT) sequencecontaining no point mutations. PAX2, Paired box gene 2; C/EBP�, CCAAT/enhancer-binding protein �.
-469 -396 -196ATG+1
HepG2
U-138
5´-RACE PCR
1kb+
ladder
Determination of transcription start site
Fig. 6. Investigation of the Transcription Start Site for GPR395�RACE PCR was performed on cDNA obtained from Hep G2 cells (left) and human glioma cell line (U-138) MG cells (right).
A representative gel is shown. The locations of two identified major transcription start sites (TSS) are shown graphically, at 396and 196 bases upstream of the translation initiation codon ATG. There was no indication of the transcription start site at �469originally annotated by Ota et al. (17) in a large-scale analysis of multiple genes.
1690 Mol Endocrinol, July 2007, 21(7):1685–1698 Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target
an alanine in the Hep G2 cell line. The consequence ofthis mutation is somewhat unclear, but it has beensuggested to increase the transactivation potential ofHNF-4� (20). Nevertheless, this altered HNF-4� pro-tein to some degree disqualifies the Hep G2 cell line asa tool to investigate the actions of HNF-4�.
Concerning the three SP1 sites, which were studiedby mutational analysis, the one located around posi-tion �598 was clearly essential for the GPR39 pro-moter activity in both the U-138 and the Hep G2 cells(Fig. 7B). Promoter function was also significantly im-paired by mutations of the SP1 sites located especiallyat position �514 but also at �522 in the U-138 cells.However, smaller or no effect was observed in the HepG2 cell line with these mutations. Such differencesbetween two cell lines is to be expected because ofvariance in the transcriptional factor milieu, which alsois emphasized by the differences in transcriptionalstart sites (Fig. 6).
It is concluded that SP1, HNF-1�, and HNF-4� areimportant transcription factors involved in the regula-tion of the expression of the GPR39 receptor.
In Vivo Regulation of GPR39 Expression
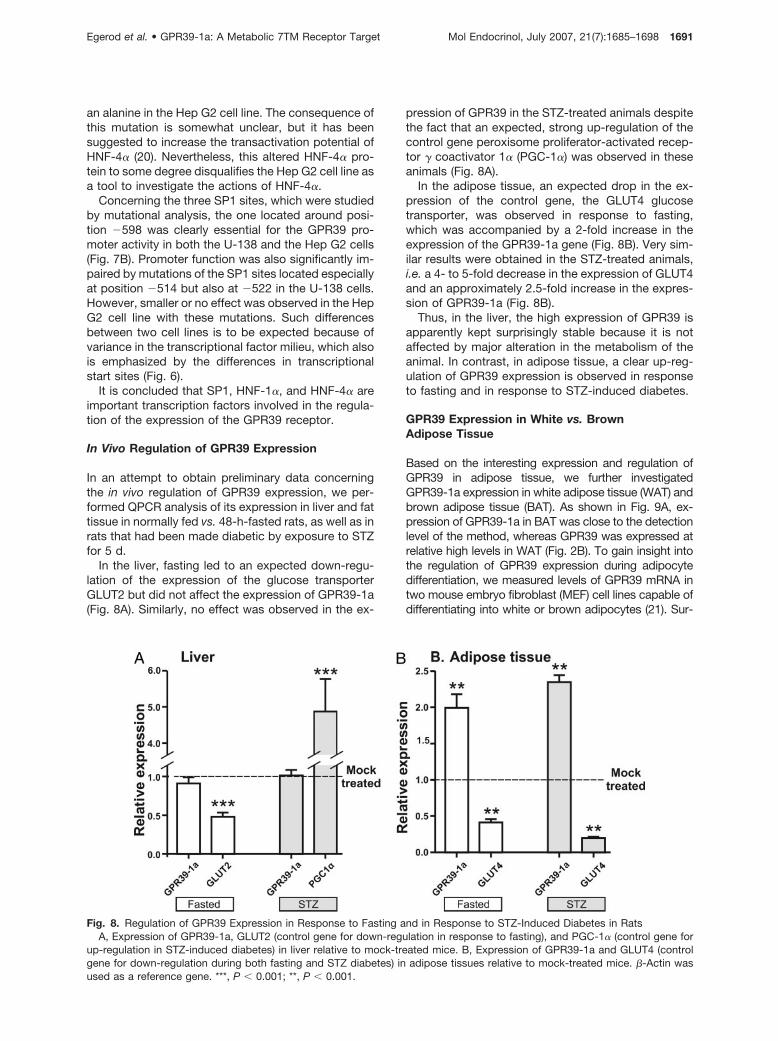
In an attempt to obtain preliminary data concerningthe in vivo regulation of GPR39 expression, we per-formed QPCR analysis of its expression in liver and fattissue in normally fed vs. 48-h-fasted rats, as well as inrats that had been made diabetic by exposure to STZfor 5 d.
In the liver, fasting led to an expected down-regu-lation of the expression of the glucose transporterGLUT2 but did not affect the expression of GPR39-1a(Fig. 8A). Similarly, no effect was observed in the ex-
pression of GPR39 in the STZ-treated animals despitethe fact that an expected, strong up-regulation of thecontrol gene peroxisome proliferator-activated recep-tor � coactivator 1� (PGC-1�) was observed in theseanimals (Fig. 8A).
In the adipose tissue, an expected drop in the ex-pression of the control gene, the GLUT4 glucosetransporter, was observed in response to fasting,which was accompanied by a 2-fold increase in theexpression of the GPR39-1a gene (Fig. 8B). Very sim-ilar results were obtained in the STZ-treated animals,i.e. a 4- to 5-fold decrease in the expression of GLUT4and an approximately 2.5-fold increase in the expres-sion of GPR39-1a (Fig. 8B).
Thus, in the liver, the high expression of GPR39 isapparently kept surprisingly stable because it is notaffected by major alteration in the metabolism of theanimal. In contrast, in adipose tissue, a clear up-reg-ulation of GPR39 expression is observed in responseto fasting and in response to STZ-induced diabetes.
GPR39 Expression in White vs. BrownAdipose Tissue
Based on the interesting expression and regulation ofGPR39 in adipose tissue, we further investigatedGPR39-1a expression in white adipose tissue (WAT) andbrown adipose tissue (BAT). As shown in Fig. 9A, ex-pression of GPR39-1a in BAT was close to the detectionlevel of the method, whereas GPR39 was expressed atrelative high levels in WAT (Fig. 2B). To gain insight intothe regulation of GPR39 expression during adipocytedifferentiation, we measured levels of GPR39 mRNA intwo mouse embryo fibroblast (MEF) cell lines capable ofdifferentiating into white or brown adipocytes (21). Sur-
Fig. 8. Regulation of GPR39 Expression in Response to Fasting and in Response to STZ-Induced Diabetes in RatsA, Expression of GPR39-1a, GLUT2 (control gene for down-regulation in response to fasting), and PGC-1� (control gene for
up-regulation in STZ-induced diabetes) in liver relative to mock-treated mice. B, Expression of GPR39-1a and GLUT4 (controlgene for down-regulation during both fasting and STZ diabetes) in adipose tissues relative to mock-treated mice. �-Actin wasused as a reference gene. ***, P � 0.001; **, P � 0.001.
Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target Mol Endocrinol, July 2007, 21(7):1685–1698 1691

prisingly, GPR39-1a was expressed already in the undif-ferentiated state (designated d 0), and this was observedin both the retinoblastoma protein positive (Rb�/�) andnegative (Rb�/�) fibroblasts, with expression levels be-ing approximately 2-fold higher in the former (Fig. 9B).The Rb�/� cells are capable of differentiating into whiteadipocytes, whereas Rb�/� cells differentiate into brownadipocytes, as was confirmed by measuring expressionlevels of the general adipocyte marker gene fatty acidbinding protein 4 (FABP4) as well as the brown adipo-cyte-specific gene uncoupling protein 1 (UCP1) (Fig. 9B).Interestingly, in both cell lines, GPR39-1a expressionwas completely down-regulated during the differentia-tion process (Fig. 9B, compare d 0 with d 8).
DISCUSSION
In the present study, we find that GPR39-1a, i.e. thefull-length, real GPR39 receptor, is expressed mainlyin peripheral, metabolic tissues such as the liver, theGI tract, the pancreas, the kidney, and adipose tissue.Focusing on the adipose tissue, it is found thatGPR39-1a is up-regulated in vivo by fasting and bySTZ-induced diabetes and that it is expressed in MEFsand is down-regulated during their differentiation intoadipocytes. Moreover, it is selectively expressed inWAT compared with BAT. The previous notion, thatGPR39 was broadly expressed, for example in theCNS, is shown to be based mainly on the detection ofan antisense gene, LYPD1, that is highly and ratherselectively expressed in several brain regions. More-over, like many other members of the ghrelin receptorfamily, GPR39 is in a number of tissues also expressedin a truncated splice variant version having only 5TM,coined GPR39-1b, the function of which remains to beshown. Analysis of the GPR39 promoter demonstratedthat transcription factors associated with metabolicactivity such as HNF-1�, HNF-4�, and SP1 are impor-tant for the expression of the GPR39 receptor.
GPR39 Is Found in Two Splice Variant Forms
A characteristic feature of several members of theghrelin receptor family is the occurrence of an intronlocated at a conserved site in the gene correspondingto the position in which where TM VI enters the mem-brane (Fig. 1). In GPR39, this intron is particularly large,and, for awhile, it led to the misconception that GPR39was a pseudogene. In mice, GPR39 is still wronglyannotated in the Entrez GenBank (accession no.71111), and this error was only recently corrected inthe ensemble database in which it is now shown cor-rectly (accession no. ENSRNOG00000021586). Inter-estingly the antisense gene LYPD1 is very conservedbetween all of these species, and, in fact, before theannotation of GPR39 was corrected in mice, etc., weidentified the second exon of GPR39 by locatingLYPD1.
For both the ghrelin receptor and the motilin recep-tor, splice variants corresponding to 5TM truncatedreceptor proteins have been described (4, 13).Through PCR analysis, we also found that GPR39 isexpressed as an alternative splice protein, GPR39-1b.Interestingly, the 5TM form of the ghrelin receptor hasbeen reported to be able to suppress the function ofthe full-length GHSR-1a receptor (22). A similar phe-nomenon has been observed in a few other cases inwhich 5TM splice variants are expressed, as for ex-ample in the GnRH receptor (23, 24). This could verywell be related to the proposed two-domain functionof 7TM receptors, in which TMs I–V, connectedthrough loops of relatively well conserved length, con-stitute an A-domain that is separated by a very vari-able and often very long intracellular loop 3 from aB-domain consisting of the closely associated TM VIand TM VII (Fig. 1) (25–28). According to the so-calledglobal toggle switch model, 7TM receptor activation isassociated with a vertical see-saw movement occur-ring in the kinked TM VI and TM VII of the B-domain inrelation to the relatively rigid A-domain (29).
Fig. 9. Regulation of GPR39 Expression in WAT and BAT and during Adipocyte Differentiation of FibroblastsA, Expression of GPR39-1a in mouse epididymal white fat and interscapular brown fat. Expression of the control genes FABP4
and UCP1 is included to verify the purity of the tissue samples. B, Expression of GPR39-1a in MEFs wild type (Rb�/�) or deficient(Rb�/�) for the retinoblastoma gene, before and after differentiation. RNA was isolated from Rb�/� and Rb�/� MEF cells before(d 0) and after (d 8) conversion into white and brown adipocytes, respectively. Expression of FABP4 and UCP1 is included ascontrol genes. TATA box binding protein was used as reference gene.
1692 Mol Endocrinol, July 2007, 21(7):1685–1698 Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target

In the case of the GPR39-1b, we find that this spliceform is rather widely expressed but with clearly higherlevels observed in the stomach and small intestine, inwhich the full-length GPR39-1a receptor is also highlyexpressed (Fig. 2B). The fact that the splice site is sur-prisingly well conserved within the ghrelin receptor familyindicates that these “5TM” proteins could serve a func-tion in the in vivo setting. If the truncated 1b receptorform does affect the signaling of the full-length receptors,this could be particularly interesting for constitutive ac-tive receptors such as GPR39 and the ghrelin receptor,because modulation of the expression of the 1b formwould be a novel, ligand-independent way of fine-tuningthe level of constitutive activity of the receptor. ForGPR39, the stomach and small intestine would then betissues in which GPR39-1b could be expected to servea physiological purpose.
Selective Expression of the Full-Length GPR39-1aReceptor in Metabolic Tissues
In the Northern blot analysis performed by McKee etal. in association with the original cloning of GPR39 (1),a cDNA construct containing the whole-coding regionfor GPR39 was used as a probe. This lead to thegeneral deception that GPR39 was widely expressedin both the CNS and in multiple peripheral tissuesbecause of the fact that this probe picked up not onlythe real receptor GPR39-1a but also the 5TM splicevariant GPR39-1b, as well as the antisense geneLYPD1. Through QPCR analysis using specific probesfor each of these three gene products, we demon-strate in the present study that the full-length, func-tional GPR39-1a receptor is mainly expressed in pe-ripheral, metabolic organs, whereas the antisensegene proved to be a CNS-specific gene. The truncatedGPR39-1b appeared to be more widely expressed, asdiscussed above. This picture of expression as well asthe sizes of the individual mRNAs fit very well with theoriginal Northern blot analysis, i.e. when it is inter-preted with our current knowledge that the probe usedcould anneal to three individual transcripts giving riseto three distinct bands on the Northern blot, becausethe blots displayed not one but three bands conceiv-ably corresponding to GPR39-1a, GPR39-1b, andLYPD1, respectively (1).
Our QPCR analysis of the tissue distribution ofGPR39-1a expression is in very good agreement witha recently published QPCR analysis from Moechars etal. (30), who, according to the probes they used, verylikely also detected GPR39-1a selectively. In bothstudies, the highest expression of GPR39 was foundthroughout the GI tract, including the liver and thepancreas, as well as in the kidney and with tissuessuch as lung, heart, and spleen showing clear butlower expression. We also found a high expression ofGPR39-1a in adipose tissue and a clear expression inthe thyroid, which however were not among the tis-sues tested by Moechars et al.
Debate Concerning GPR39 Expression in the CNS
In relation to the proposal that GPR39 could be thereceptor for obestatin, a peptide fragment from theghrelin precursor (see below), Zhang et al. (8) pre-sented QPCR data indicating that GPR39 was ex-pressed in the pituitary and hypothalamus at levelssimilar to those observed in the stomach and liver.However, subsequent QPCR analysis performed bytwo other groups showed very low, if any, expressionof GPR39 in these areas as well as in the CNS ingeneral, which is in agreement with the QPCR analysisfor GPR39-1a of the present study (Fig. 2B) (9, 11, 30).It is in fact likely that there are major methodologicaldifferences between the approach of Zhang et al. andthat of the other groups because Zhang et al. not onlydetected high levels of GPR39 expression in the hy-pothalamus and pituitary but also a low expression ofGPR39 in, for example, the kidney and the colon (8), inwhich all the other groups detect high levels (9, 11, 30)(Fig. 2B).
Through in situ hybridization, GPR39 expression hasbeen detected in the amygdala, hippocampus, andauditory cortex but explicitly not in the hypothalamus(31). In GPR39 knockout mice, who express LacZinstead of the GPR39 receptor, a signal was also de-tected in the septum-amygdala system in addition tothroughout the GI tract, including the endocrine pan-creas, but again not in the hypothalamus (30). It shouldbe noted that, in the same paper, the QPCR analysisdetected very low expression of GPR39 in septum andother brain areas in wild-type mice (30). It cannot atpresent be excluded that real GPR39-1a is expressedto a low degree in certain areas of the CNS such asperhaps specific nuclei in the septum, although it wasnot detected by our selective in situ hybridization (Fig.4). However, it is likely that at least some of the GPR39expression reported in various brain areas by use ofeither in situ hybridization or LacZ expression could beattributable to expression of the truncated, GPR39-1bform of the receptor (Fig. 2B). At least, all reports(except one) agree that GPR39 is not widely expressedin the CNS and in particular not in the hypothalamus.
The Antisense Gene LYPD1 Is Highly Expressedin the CNS
As shown in Fig. 2, LYPD1 is encoded on the anti-sense strand corresponding to the exon encoding TMVI and TM VII and the C-terminal segment of GPR39.In fact, GPR39 and LYPD1 have been used as a modelsystem in a study on the use of RNA interference tocontrol the expression of antisense genes (18). In thepresent study, we find that LYDC1 and GPR39 have areciprocal or inverse expression pattern, becauseLYPD1 is expressed highly selectively in the CNS andbecause the in situ hybridization analysis reveals adistinct cellular, presumably neuronal localization indistinct brain nuclei (Fig. 4). In contrast, GPR39, asdiscussed above, is expressed selectively in metabolic
Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target Mol Endocrinol, July 2007, 21(7):1685–1698 1693

tissues in the periphery (Fig. 2B). In certain peripheraltissues, there appears to be some overlap in the ex-pression pattern, for example, the heart and the lung(Fig. 2B). However, it remains to be demonstratedwhether this apparent coexpression really is a truecoexpression in the same cells or whether LYPD1 maybe expressed, for example, in neurons in these tis-sues. For many, but not all, pairs of antisense genes,it has been found that expression of one gene ex-cludes the expression of the other or at least that theyare expressed at opposite levels in the cell (32, 33).This is the pattern that we observe for GPR39 andLYPD1 in a large number of tissues throughout thebody (Fig. 2B).
LYPD1 appears to encode a highly interesting pro-tein. An amino-terminal signal peptide indicates thatthe LYPD1 gene product is a secreted protein. Thesignal peptide is followed by a sequence that appar-ently encodes an Ly-6 domain or module character-ized by a highly conserved pattern of Cys residuesforming up to five disulfide bridges in the folded pro-tein structure (34). Ly-6 domains or modules arefound, for example, in the urokinase-type plasminogenactivator receptor, in CD59, in C4.4A, in RGTR430, inLy-6 neurotoxin-like 1c, and in SLURP1 (secreted Ly-6/PLAUR domain containing 1) (34). In LYPD1, theLy-6 domain is followed by a potential proteolyticcleavage site (rich in Pros and with multiple basicresidues), which again is followed by a potential gly-cosylphosphatidylinositol anchor site. Thus, it is likelythat LYPD1 is expressed both as a glycosylphosphati-dylinositol-modified, membrane-associated proteinand as a secreted protein that could serve some sig-naling purpose.
Transcription Factors Controlling GPR39Expression
The mutational analysis of the promoter constructsusing the luciferase reporter assay clearly indicatedthat HNF-1�, HNF-4�, and SP1 are involved in thecontrol of GPR39 expression (Fig. 7). This also fits wellwith the observed expression pattern for the receptorin the GI tract, including the liver and pancreas, etc.HNF-1� and HNF-4� are strongly linked to early-onsettype 2 diabetes especially maturity-onset diabetes ofthe young (MODY), in which mutation in HNF-1� re-sults in MODY3 whereas mutations in HNF-4� resultsin MODY1 (35, 36). Thus, among carriers of a mutationin HNF-1�, approximately 63% will have diabetes bythe age of 25 yr and 95% by the age of 55 yr.
In a systematic analysis performed by microarraysand chromatin immunoprecipitation on 13,000 genes,it was found that, in hepatocytes, 1.7% of the genesare targeted by HNF-1� whereas 12% are targeted byHNF-4� (37). GPR39 was not discussed in the paper,but the receptor is found in the table listing the 158genes, which are characterized by potentially beingtargeted both by HNF-1� and by HNF-4� in a feed-forward, multicomponent loop manner (37). Interest-
ingly, in this group of genes, GPR39 and the soma-tostatin receptor SSTR-1 were the only 7TM receptorslisted (37). In the present study, we provide functionaldata that substantiates that the expression of GPR39is in fact regulated by both HNF-1� and HNF-4�. Thisobviously also fits well with the expression pattern forGPR39 in the liver, pancreas, and GI tract as such (Fig.2B) and point to a potential important role for GPR39in metabolism.
What Is the Physiological Role of GPR39?
It is obviously far too early to answer this question.However, in the present study, we found that GPR39 isup-regulated in adipose tissue during fasting and inresponse to STZ-induced diabetes in rats (Fig. 8B),which support the notion that this receptor is a meta-bolically interesting target. Somewhat surprisingly, inthe same animals, the expression of GPR39 in the liverwas held very stable. Although these are early andpreliminary studies, it could indicate that GPR39 hassome other, nonmetabolic function at least in the liver.The observation that GPR39 is expressed in MEFs andthat its expression changes during adipocyte differen-tiation suggest that the receptor could be expressedspecifically on preadipocytes and possibly play a rolein adipocyte differentiation. One speculative possibilitywould be that GPR39 could play a similar role in, forexample, the liver and the endocrine pancreas andperhaps the GI tract being involved also in tissue re-generation and differentiation.
Recently, the two first reports were published con-cerning GPR39 knockout mice (9, 30). These micebreed normally and do not display any overt pheno-typic characteristics. Moechars et al. (30) focused onthe GI function and presented evidence that gastricemptying and GI tract passage as such is enhanced inthe GPR39�/� mice conceivably mainly as a result ofan almost 3-fold increase in gastric fluid secretion. TheGPR39�/� mice had normal body weight and foodintake until wk 20, whereas very old animals showedincreased body weight. Most biochemical parameterswere normal, but the GPR39�/� animals did showincreased levels of plasma cholesterol (30). Tremblayand coworkers focused on the food intake and bodyweight issues, in which they found that the GPR39�/�
animals behaved just like the wild-type littermates inall respects. Both groups reported normal plasma glu-cose and insulin levels for the GPR39�/� animals (9,30). Although the GPR39�/� mice in many ways ap-pear to be rather normal, besides especially the gastricsecretion, it will be interesting to see how theGPR39�/� mice respond to various challenges of thefunction of the liver, pancreas, intestine, adipocytes,etc.
Current Status for GPR39
GPR39 is a constitutively active 7TM receptor that isactivated and or modulated by zinc ions and potentially
1694 Mol Endocrinol, July 2007, 21(7):1685–1698 Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target

by another as yet unidentified ligand. The receptor isexpressed throughout the GI tract, including the liver andpancreas as well as in adipose tissue, and its expressionis controlled in a rather unique way, at least among 7TMreceptors, by transcription factors such as HNF-1�,HNF-4a, and SP1, which points to an interesting functionof GPR39 in these metabolically important organs. Thegenomic arrangement of GPR39 together with themembrane-associated/secreted neuronally expressedLYPD1 protein as a sense-antisense gene pair points tointeresting possibilities in both relation to function andconnection with regulatory mechanisms such asgenomic imprinting (32, 33).
MATERIALS AND METHODS
Animals
Male Wistar rats weighing approximately 300 g were used.Rats were housed individually in plastic cages in a tempera-ture- and humidity-controlled environment with a 12-h light,12-h dark cycle. Standard pellet chow and water were avail-able ad libitum. Animals were randomly assigned to threegroups: STZ-treated diabetic, mock treated, and fasted. Ratswere rendered diabetic by a single ip injection of STZ [70mg/kg bodyweight dissolved in citrate buffer (pH 4.5)] (n � 8).Rats treated with a single ip injection of vehicle were used asmock controls (n � 8). The third group was fasted for 48 hwith free access to water (n � 8). The animals were killed 5 dafter STZ/vehicle administration or after the 48 h of fasting.Pancreas, liver, and adipose tissue were excised, rapidlyfrozen in liquid nitrogen, and stored at �80 C. All rats werecared for and handled according to the European Commu-nities Council Directives (86/609/ECC), and the present studywas approved by the Danish laboratory animal carecommittee.
PCR
PCR for constructs analyzing the promoter region was per-formed directly on chromosomal DNA, which were extractedfrom human blood using the QIAamp DNA Blood Mini kit(Qiagen, Hilden, Germany). The PCR program was 94 C for 3min, followed by 25 cycles at 94 C for 45 s, 53 C for 45 s, 72C for 2 min. Primers used are listed in Table 1.
Site-directed mutagenesis was performed using the overlapextension method (38) The same PCR program as describedabove was used. The primers used are listed in Table 1.
5�RACE PCR was performed using the SMART RACE cDNAAmplification kit (BD Biosciences, San Jose, CA) for investiga-tion of the transcription start site. Total RNA was isolated fromthe cell lines Hep G2 and U-138, using the RNeasy Lipid TissueMini kit (Qiagen) including a treatment with deoxyribonuclease I.The PCR program was 94 C for 3 min, then a 20-cycle touch-down, 94 C for 45 s, 63 C for 45 s decreasing 0.5 C per cycle,72 C for 2 min, followed by 25 cycles at 94 C for 45 s, 53 C for45 s, 72 C for 2 min. This was used for both the initial PCR andthe nested PCR. Primers used are listed in Table 1.
All of the above PCRs were run on Eppendorf Mastercyclermachines using the Pfu DNA polymerase (Promega, Madi-son, WI) and further analyzed on a 1.2% agarose gel. Prod-ucts were extracted using the NucleoSpin Extract kits (Ma-cherey-Nagel, Duren, Germany), restricted by the enzymesKpn1 and HindIII (New England Biolabs, Beverly, MA), andligated into the pGL3-basic vector (Promega) using theTaKaRa (Tokyo, Japan) DNA Ligation kit. The constructswere transformed into the XL-1 Blue Competent Cells (Strat-
agene, La Jolla, CA) and plated on LB agar containing 100�g/ml ampicillin. One clone was selected, and plasmid DNAwas extracted using the Qiagen Plasmid Maxi kit. Sequenc-ing was done by MWG Biotech (Ebersberg, Germany).
QPCR was performed using the Mx3000P (Stratagene)and the SYBR Premix Ex Taq (TaKaRa). Cycle threshold(Ct) values were obtained using Stratagene Mx3000P soft-ware, �-actin was used as a reference gene, and a cali-brator sample was included in each round of QPCR tonormalize between runs. The data were normalized bysetting the maximum expression value to 1, thus showingthe relative expression of the gene. To show the direct foldchange of the ratio of two genes (gene_A/gene_B), a direct�Ct method was used, basically using one of the genes asreference gene giving the following ratio: gene_A/gene_B � 2�(Ctgene_A�Ctgene_B). This ratio was arbitrary set to1 for the heart in which all three gene products are ex-pressed to a reasonable degree and thereby clearly showthe ratio change in the various other tissues. RNA fromtissue was extracted using the RNeasy Lipid Tissue Mini kit(Qiagen), followed by cDNA synthesis using the ImProm-IIReverse Transcriptase (Promega). Primers and probesused are listed in Table 1, and the specificity of the primerswas evaluated through both melting curve analyzes andsequencing.
In Situ Hybridization
A 200-bp cDNA fragment corresponding to a 200-bp regionof the 3� untranslated region of GPR39-1a and likewise a200-bp region of the 3� region of LYPD1 were amplified usingthe primers Fr39/LYPD1 and Rr39/LYPD1 listed in Table 1and ligated into the pSTP18 vector (Roche, Nutley, NJ). Theplasmid containing this cDNA fragment was linearized withappropriate restriction enzymes. [35S]Uridine-5�-triphos-phate-labeled RNA antisense (GPR39-1a) and sense (LYPD1)probes were transcribed in vitro using the appropriate tem-plate and phage RNA polymerase, purified using ProbeQuant G-50 microcolumns (GE Healthcare, Little Chalfont,UK), and checked in denaturing acrylamide gel.
Expression of GPR39-1a and LYPD1 mRNA in the rat brainwas investigated using in situ hybridization technique accordingto Diaz Heijtz and Castellanos (39). Briefly, brains were rapidlydissected and frozen on dry ice. Coronal sections (14 �m) wereprepared on a cryostat and stored at �80 C until used. Thefrozen tissue sections were fixed in cold 4% paraformaldehyde[in 0.1 M PB (pH 7.4)], deproteinated with 0.1 M HCl, incubatedin 0.25% acetic anhydride in 0.1 M triethanolamine (pH 8.0), anddehydrated. Sections were air dried and prehybridized [50%formamide, 50 mM Tris-HCl (pH 7.6), 25 mM EDTA (pH 8.0), 20mM NaCl, 0.25 mg/ml yeast tRNA, and 2.5� Denhardt’s solu-tion] for 4 h at 55 C followed by overnight (14–16 h) hybridizationin a humidified chamber at 55 C. For hybridization, labeledprobe was diluted to a final concentration of 0.5 � 106 cpm/200�l containing 50% deionized formamide (pH 5), 0.3 M NaCl, 20mM Tris-HCl (pH 7.6), 5 mM EDTA (pH 8.0), 10 mM PBS, 0.2 mM
dithiothreitol, 0.5 mg/ml yeast tRNA, 0.1 mg/ml poly-A-RNA,10% dextran sulfate, and 1� Denhardt’s solution. After hybrid-ization and rinsing, the sections were then treated with 1 �gribonuclease A (Roche) in ribonuclease buffer [0.5 M NaCl, 10mM Tris-HCl, and 5 mM EDTA (ph 8.0)] for 1 h at 37 C, dehy-drated in ascending alcohol series, and air dried. Sections wereplaced against ®-Max film (GE Healthcare) and stored at roomtemperature for 2–3 d.
Cell Cultures and Promoter Assay
The glioblastoma cell line U-138 (American Type Culture Col-lection, Manassas, VA), which has been reported to expressGPR39 (18), and the hepatocellular carcinoma cell line Hep G2(kindly provided by Prof. Jens Høiriis Nielsen, University ofCopenhagen, Copenhagen, Denmark), were both grown in
Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target Mol Endocrinol, July 2007, 21(7):1685–1698 1695

GIBCO DMEM/F-12 with glutamine and HEPES (Invitrogen,Carlsbad, CA). The following were added to the media:nonessential amino acids (0.1 mM), fetal bovine serum(5%), and penicillin-streptomycin (50 U/ml). LipofectamineReagent (Invitrogen) was used for transfection, and thepromoter activity was analyzed using the Firelite Dual Lu-minescence Reporter Gene Assay System (PerkinElmer,Wellesley, MA). This includes a measurement of Renillaluciferase to normalize for transfection efficiency, whichwas obtained by cotransfection with a pCDNA3.1(�) (In-vitrogen) construct carrying Renilla luciferase. The lumi-nescence was measured in a TopCount (PerkinElmer). Lu-minescences from all constructs were normalized to theempty vector pGL3-basic, thus giving a relative measure-ment for transcriptional activity. All assays were performedat least three times in five replicates.
Rb�/� and Rb�/� MEFs were propagated and differenti-ated as described previously (21). Three dishes were har-
vested for each cell line at each time point and further inves-tigated for gene expression by QPCR.
Sequence Analyzes and Software Programs
Vector NTI Advance 10 (Invitrogen) was used for generalhandling of sequence information. The TRANSFAC 6.0 publicdatabase was used for identifying consensus of transcriptionfactor binding sites, using the program Match (BIOBASE).The consensus of transcription factor binding sites were alsoused for designing disrupting point mutations. The programConSite (Center for Genomics and Bioinformatics, KarolinskaInstitute) was used to locate conserved binding sites betweenH. sapiens and M. musculus. PRISM version 4 (GraphPadSoftware, San Diego, CA) was used for data handling andgeneration of graphs. The data were normalized to show foldinduction relative to one chosen sample, and a t test (Mann-Whitney) was used to investigate significant variances.
Table 1. Sequences of Primers
Sequence
Primers for promoter constructsF8 AAAGGTACCTAACTGTTGTCCTGTGGCTGTATGF7 AAAGGTACCCATTGCTCATTCCCTCTGATTGF6 AAAGGTACCGGAAAGGCAGGGATTCTATAGTGF5 AAAGGTACCCAGAGCCCAGCTGTCCTTGF3 AAAGGTACCCACCTCCTTGGGACGCTTF2 AAAGGTACCGAGAGCCACGATCTGATTTCAGF1 AAAGGTACCCAATCCGCTGTGGTTAATGATTF0 AAAGGTACCTGGAAAGGAAGAGGAGTGCTGR2 TTTAAGCTTGTGAAGCCATGAGAAAGAGCAC
Primers for site-directed mutagenesisF-HNF-1� GCTGTGGTTAACGCTTAATCGCTGR-HNF-1� CAGCGATTAAGCGTTAACCACAGCF-HNF-4� CGAGGGAACCGGTCATGATCR-HNF-4� GATCATGACCGGTTCCCTCGF-SP1–598 AGGGTGGAACGGGGGTGGGTR-SP1–598 ACCCACCCCCGTTCCACCCTF-SP1–522 AAACCCCGAAGCTGGGCTGGR-SP1–522 CCAGCCCAGCTTCGGGGTTTFSP1–514 AAACCCCGCCGCTGAACTGGRSP1–514 CCAGTTCAGCGGCGGGGTTT
Primers for 5� RACE PCRR2 TTTAAGCTTGTGAAGCCATGAGAAAGAGCACR3 TTTAAGCTTCCAGGTCCAAAGACTTTCTCAACR4 TTTAAGCTTCTTGGCTGCAAACTCACCTGC
Primers for in situ hybridizationFr39/LYPD1 TTTAAGCTTAGTTGAAGAACAGGACGGTGRr39/LYPD1 TTTGAATTCATCATGTACCGGAAGTCGTG
Primers for QPCRFrGPR39-1a AGTGAGGAGAGCCGGACAGRrGPR39-1a CAGTCATGTTTGGGTTTTGCRrGPR39-1b CCTCGGTGAACTCAGGGTTTFr&�-actin TTCTACAATGAGCTGCGTGTGRr&�-actin GGGGTGTTGAAGGTCTCAAAFrLYPD1 AGAAGTGATGGAGCAAAGTGCRrLYPD1 GGACTGGTACCCAGCTGAAGFmFABP4 TGGAAGCTTGTCTCCAGTGARmFABP4 AATCCCCATTTACGCTGATGFmUCP1 GGCATTCAGAGGCAAATCAGCTRmUCP1 CAATGAACACTGCCACACCTCFmTBP TCAAACCCAGAATTGTTCTCCRmTBP GGTAGATGTTTTCAAATGCTTCA
F, Forward; R, reverse; TBP, TATA box binding protein; r, rat; m, mouse.
1696 Mol Endocrinol, July 2007, 21(7):1685–1698 Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target

Acknowledgments
We thank Mette Simons for expert technical help and Dr.Rochellys Diaz Heijtz for her help with the in situ hybridization.
Received January 26, 2007. Accepted April 30, 2007.Address all correspondence and requests for reprints to:
Thue W. Schwartz, Laboratory for Molecular Pharmacology,Department of Neuroscience and Pharmacology, The PanumInstitute, University of Copenhagen, Blegdamsvej 3, DK-2200Copenhagen, Denmark. E-mail: [email protected].
The study was supported by grants from the Danish Med-ical Research Council, the Novo Nordisk Foundation, and theSwedish Research Council. K.L.E. was the recipient of astipend from the Health Science Faculty of University ofCopenhagen.
Disclosure Statement: The authors have nothing todisclose.
REFERENCES
1. McKee KK, Tan CP, Palyha OC, Liu J, Feighner SD,Hreniuk DL, Smith RG, Howard AD, Van der Ploeg LH1997 Cloning and characterization of two human G pro-tein-coupled receptor genes (GPR38 and GPR39) relatedto the growth hormone secretagogue and neurotensinreceptors. Genomics 46:426–434
2. Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H,Kangawa K 1999 Ghrelin is a growth-hormone-releasingacylated peptide from stomach. Nature 402:656–660
3. Holst B, Schwartz TW 2004 Constitutive ghrelin receptoractivity as a signaling set-point in appetite regulation.Trends Pharmacol Sci 25:113–117
4. Feighner SD, Tan CP, McKee KK, Palyha OC, HreniukDL, Pong SS, Austin CP, Figueroa D, MacNeil D, CascieriMA, Nargund R, Bakshi R, Abramovitz M, Stocco R,Kargman S, O’Neill G, Van der Ploeg LH, Evans J, Patch-ett AA, Smith RG, Howard AD 1999 Receptor for motilinidentified in the human gastrointestinal system. Science284:2184–2188
5. Howard AD, Wang R, Pong SS, Mellin TN, Strack A, GuanXM, Zeng Z, Williams Jr DL, Feighner SD, Nunes CN,Murphy B, Stair JN, Yu H, Jiang Q, Clements MK, TanCP, McKee KK, Hreniuk DL, McDonald TP, Lynch KR,Evans JF, Austin CP, Caskey CT, Van der Ploeg LH, LiuQ 2000 Identification of receptors for neuromedin U andits role in feeding. Nature 406:70–74
6. Zhao D, Pothoulakis C 2006 Effects of NT on gastroin-testinal motility and secretion, and role in intestinal in-flammation. Peptides 27:2434–2444
7. Holst B, Holliday ND, Bach A, Elling CE, Cox HM,Schwartz TW 2004 Common structural basis for consti-tutive activity of the ghrelin receptor family. J Biol Chem279:53806–53817
8. Zhang JV, Ren PG, vsian-Kretchmer O, Luo CW, RauchR, Klein C, Hsueh AJ 2005 Obestatin, a peptide encodedby the ghrelin gene, opposes ghrelin’s effects on foodintake. Science 310:996–999
9. Tremblay F, Perreault M, Klaman LD, Tobin JF, Smith E,Gimeno RE 2007 Normal food intake and body weight inmice lacking the G protein-coupled receptor GPR39. En-docrinology 148:501–506
10. Lauwers E, Landuyt B, Arckens L, Schoofs L, Luyten W2006 Obestatin does not activate orphan G protein-cou-pled receptor GPR39. Biochem Biophys Res Commun351:21–25
11. Holst B, Egerod KL, Schild E, Vickers SP, Cheetham S,Gerlach LO, Storjohann L, Stidsen CE, Jones R, Beck-
Sickinger AG, Schwartz TW 2007 GPR39 signaling isstimulated by zinc ions but not by obestatin. Endocrinol-ogy 148:13–20
12. Clark HF, Gurney AL, Abaya E, Baker K, Baldwin D,Brush J, Chen J, Chow B, Chui C, Crowley C, Currell B,Deuel B, Dowd P, Eaton D, Foster J, Grimaldi C, Gu Q,Hass PE, Heldens S, Huang A, Kim HS, Klimowski L, JinY, Johnson S, Lee J, Lewis L, Liao D, Mark M, Robbie E,Sanchez C, Schoenfeld J, Seshagiri S, Simmons L, SinghJ, Smith V, Stinson J, Vagts A, Vandlen R, Watanabe C,Wieand D, Woods K, Xie MH, Yansura D, Yi S, Yu G,Yuan J, Zhang M, Zhang Z, Goddard A, Wood WI,Godowski P, Gray A 2003 The secreted protein discoveryinitiative (SPDI), a large-scale effort to identify novel hu-man secreted and transmembrane proteins: a bioinfor-matics assessment. Genome Res 13:2265–2270
13. Howard AD, Feighner SD, Cully DF, Arena JP, LiberatorPA, Rosenblum CI, Hamelin M, Hreniuk DL, Palyha OC,Anderson J, Paress PS, Diaz C, Chou M, Liu KK, McKeeKK, Pong SS, Chaung LY, Elbrecht A, Dashkevicz M,Heavens R, Rigby M, Sirinathsinghji DJ, Dean DC, MelilloDG, Patchett AA, Nargund R, Griffin PR, DeMartino JA,Gupta SK, Schaeffer JM, Smith RG, Van der Ploeg LH1996 A receptor in pituitary and hypothalamus that func-tions in growth hormone release. Science 273:974–977
14. Nogueiras R, Tovar S, Mitchell SE, Rayner DV, Archer ZA,Dieguez C, Williams LM 2004 Regulation of growth hor-mone secretagogue receptor gene expression in the ar-cuate nuclei of the rat by leptin and ghrelin. Diabetes53:2552–2558
15. Kim MS, Yoon CY, Park KH, Shin CS, Park KS, Kim SY,Cho BY, Lee HK 2003 Changes in ghrelin and ghrelinreceptor expression according to feeding status. Neuro-report 14:1317–1320
16. Burdyga G, Lal S, Varro A, Dimaline R, Thompson DG,Dockray GJ 2004 Expression of cannabinoid CB1 recep-tors by vagal afferent neurons is inhibited by cholecys-tokinin. J Neurosci 24:2708–2715
17. Ota T, Suzuki Y, Nishikawa T, Otsuki T, Sugiyama T, IrieR, Wakamatsu A, Hayashi K, Sato H, Nagai K, Kimura K,Makita H, Sekine M, Obayashi M, Nishi T, Shibahara T,Tanaka T, Ishii S, Yamamoto J, Saito K, Kawai Y, IsonoY, Nakamura Y, Nagahari K, Murakami K, Yasuda T,Iwayanagi T, Wagatsuma M, Shiratori A, Sudo H, HosoiriT, Kaku Y, Kodaira H, Kondo H, Sugawara M, TakahashiM, Kanda K, Yokoi T, Furuya T, Kikkawa E, Omura Y, AbeK, Kamihara K, Katsuta N, Sato K, Tanikawa M,Yamazaki M, Ninomiya K, Ishibashi T, Yamashita H, Mu-rakawa K, Fujimori K, Tanai H, Kimata M, Watanabe M,Hiraoka S, Chiba Y, Ishida S, Ono Y, Takiguchi S, Wa-tanabe S, Yosida M, Hotuta T, Kusano J, Kanehori K,Takahashi-Fujii A, Hara H, Tanase TO, Nomura Y, TogiyaS, Komai F, Hara R, Takeuchi K, Arita M, Imose N,Musashino K, Yuuki H, Oshima A, Sasaki N, Aotsuka S,Yoshikawa Y, Matsunawa H, Ichihara T, Shiohata N,Sano S, Moriya S, Momiyama H, Satoh N, Takami S,Terashima Y, Suzuki O, Nakagawa S, Senoh A, Mizogu-chi H, Goto Y, Shimizu F, Wakebe H, Hishigaki H, Wa-tanabe T, Sugiyama A, Takemoto M, Kawakami B,Yamazaki M, Watanabe K, Kumagai A, Itakura S, Fuku-zumi Y, Fujimori Y, Komiyama M, Tashiro H, Tanigami A,Fujiwara T, Ono T, Yamada K, Fujii Y, Ozaki K, Hirao M,Ohmori Y, Kawabata A, Hikiji T, Kobatake N, Inagaki H,Ikema Y, Okamoto S, Okitani R, Kawakami T, Noguchi S,Itoh T, Shigeta K, Senba T, Matsumura K, Nakajima Y,Mizuno T, Morinaga M, Sasaki M, Togashi T, Oyama M,Hata H, Watanabe M, Komatsu T, Mizushima-Sugano J,Satoh T, Shirai Y, Takahashi Y, Nakagawa K, Okumura K,Nagase T, Nomura N, Kikuchi H, Masuho Y, YamashitaR, Nakai K, Yada T, Nakamura Y, Ohara O, Isogai T,Sugano S 2004 Complete sequencing and characteriza-tion of 21,243 full-length human cDNAs. Nat Genet 36:40–45
Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target Mol Endocrinol, July 2007, 21(7):1685–1698 1697

18. Hu X, Hipolito S, Lynn R, Abraham V, Ramos S, Wong-Staal F 2004 Relative gene-silencing efficiencies of smallinterfering RNAs targeting sense and antisense tran-scripts from the same genetic locus. Nucleic Acids Res32:4609–4617
19. Crawford DL, Segal JA, Barnett JL 1999 Evolutionaryanalysis of TATA-less proximal promoter function. MolBiol Evol 16:194–207
20. Lausen J, Thomas H, Lemm I, Bulman M, BorgschulzeM, Lingott A, Hattersley AT, Ryffel GU 2000 Naturallyoccurring mutations in the human HNF4� gene impairthe function of the transcription factor to a varying de-gree. Nucleic Acids Res 28:430–437
21. Hansen JB, Jorgensen C, Petersen RK, Hallenborg P, DeMR, Boye HA, Petrovic N, Enerback S, Nedergaard J,Cinti S, te Riele H, Kristiansen K 2004 Retinoblastomaprotein functions as a molecular switch determiningwhite versus brown adipocyte differentiation. Proc NatlAcad Sci USA 101:4112–4117
22. Chan CB, Cheng CH 2004 Identification and functionalcharacterization of two alternatively spliced growth hor-mone secretagogue receptor transcripts from the pitu-itary of black seabream Acanthopagrus schlegeli. MolCell Endocrinol 214:81–95
23. Grosse R, Schoneberg T, Schultz G, Gudermann T 1997Inhibition of gonadotropin-releasing hormone receptorsignaling by expression of a splice variant of the humanreceptor. Mol Endocrinol 11:1305–1318
24. Wang L, Oh DY, Bogerd J, Choi HS, Ahn RS, Seong JY,Kwon HB 2001 Inhibitory activity of alternative splicevariants of the bullfrog GnRH receptor-3 on wild-typereceptor signaling. Endocrinology 142:4015–4025
25. Bakker RA, Dees G, Carrillo JJ, Booth RG, Lopez-Gime-nez JF, Milligan G, Strange PG, Leurs R 2004 Domainswapping in the human histamine H1 receptor. J Phar-macol Exp Ther 311:131–138
26. Maggio R, Vogel Z, Wess J 1993 Coexpression studieswith mutant muscarinic/adrenergic receptors provide ev-idence for intermolecular “cross-talk” between G-pro-tein-linked receptors. Proc Natl Acad Sci USA 90:3103–3107
27. Nielsen SM, Elling CE, Schwartz TW 1998 Split-receptorsin the tachykinin neurokinin-1 system: mutational analy-sis of intracellular loop 3. Eur J Biochem 251:217–226
28. Schwartz TW, Holst B 2002 Textbook of receptor phar-macology. In: Forman JC, Johansen T, eds. 2nd ed. BocaRaton, FL: CRC; 81–110
29. Schwartz TW, Frimurer TM, Holst B, Rosenkilde MM,Elling CE 2006 Molecular mechanism of 7TM receptoractivation: a global toggle switch model. Annu Rev Phar-macol Toxicol 46:481–519
30. Moechars D, Depoortere I, Moreaux B, de Smet B, GorisI, Hoskens L, Daneels G, Kass S, Ver Donck L, Peeters T,Coulie B 2006 Altered gastrointestinal and metabolicfunction in the GPR39-obestatin receptor-knockoutmouse. Gastroenterology 131:1131–1141
31. Jackson VR, Nothacker HP, Civelli O 2006 GPR39 re-ceptor expression in the mouse brain. Neuroreport 17:813–816
32. Lavorgna G, Dahary D, Lehner B, Sorek R, SandersonCM, Casari G 2004 In search of antisense. Trends Bio-chem Sci 29:88–94
33. Lapidot M, Pilpel Y 2006 Genome-wide natural antisensetranscription: coupling its regulation to its different reg-ulatory mechanisms. EMBO Rep 7:1216–1222
34. Ploug M 2003 Structure-function relationships in the in-teraction between the urokinase-type plasminogen acti-vator and its receptor. Curr Pharm Des 9:1499–1528
35. Fajans SS, Bell GI, Polonsky KS 2001 Molecular mech-anisms and clinical pathophysiology of maturity-onsetdiabetes of the young. N Engl J Med 345:971–980
36. Frayling TM, Evans JC, Bulman MP, Pearson E, Allen L,Owen K, Bingham C, Hannemann M, Shepherd M, EllardS, Hattersley AT 2001 �-Cell genes and diabetes: mo-lecular and clinical characterization of mutations in tran-scription factors. Diabetes 50 (Suppl 1):S94–S100
37. Odom DT, Zizlsperger N, Gordon DB, Bell GW, RinaldiNJ, Murray HL, Volkert TL, Schreiber J, Rolfe PA, GiffordDK, Fraenkel E, Bell GI, Young RA 2004 Control of pan-creas and liver gene expression by HNF transcriptionfactors. Science 303:1378–1381
38. Horton RM, Hunt HD, Ho SN, Pullen JK, Pease LR 1989Engineering hybrid genes without the use of restrictionenzymes: gene splicing by overlap extension. Gene 77:61–68
39. Diaz Heijtz R, Castellanos FX 2006 Differential effects ofa selective dopamine D1-like receptor agonist on motoractivity and c-fos expression in the frontal-striatalcircuitry of SHR and Wistar-Kyoto rats. Behav BrainFunct 2:18
40. Zhang JV, Klein C, Ren P-G, Kass S, Ver Donck L,Moechars D, Hsueh AJW 2007 Response to comment on“Obestatin, a peptide encoded by the ghrelin gene, op-poses ghrelin’s effects on food intake.” Science 315:766
Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremostprofessional society serving the endocrine community.
1698 Mol Endocrinol, July 2007, 21(7):1685–1698 Egerod et al. • GPR39-1a: A Metabolic 7TM Receptor Target