
Vistas desde el globo aerostático: El Sindbart aeronáutico de HC Artmann
Upload
independentCategory
view
0download
0
lnt. J. Cancer: 58,108-115 (1994) 0 1994 Wiley-Liss, Inc.
Publication of the International Union Against Cancer Publication de I’Union Internationale Contre le Cancel
GLYCOLIPIDS OF GERM CELL TUMORS: EXTENDED GLOBO-SERIES GLYCOLIPIDS ARE A HALLMARK OF HUMAN EMBRYONAL CARCINOMA CELLS Jonathan WENK’, Peter w. ANDREWS~, Jochen cASPER3, Jun-ichi HATA4, Martin F. PERA5, Alexander VON KEITZ6, Ivan DAMJANOV~ and Bruce A. FENDERSON’S’ IDepartment of Pathology and Cell Biology, Thomas Jefferson University, 1020 Locust Street, Philadelphia, PA 19107, USA; 2Department of Biomedical Science, The University of Shefield, Western Bank, Shefield, S10 2TN, UK; 3Department of Hematology and Oncology, Medizinische Hochschule Hannover, Postfach 61 01 SO, 0-3000 Hannover 61, Germany; 4Department of Pathology, Keio University School of Medicine, 3.5 Shinanomachi, Shinjuku-ku Tokyo 160, Japan; T R C Growth Factors, Department of Zoology, University of Oxford, South Parks Road, Oxford OX1 3PS, UK; 6Department of Urology, University of Marburg, Baldinger St., 3.5043 Marburg, Germany.
Glycolipids of human germ cell tumor lines were analyzed to define the most common immunohistochemical profiles of embryonal carcinoma (EC), differentiated derivatives of EC, yolk sac carcinoma (YC) and choriocarcinoma (CC). Glycolipid composition was examined by high-performance thin-layer chromatography (HPTLC) combined with immunostaining with a panel of anti-carbohydrate monoclonal antibodies (MAbs). All EC cell lines were found to contain high levels of globo-series glycolipids, including globotriosylceramide (Gb3), globoside (Gb4), Gb5 (GalpI - 3Gb4) and GL7 (sialyl GalPI + 3Gb4). Somatic differentiated derivatives (e.g., EC cells treated with retinoic acid) contained decreased levels of globo-series glyco- lipids and increased levels of lacto- and ganglio-series glycolip- ids, including GD3, GT3 and GDZ. CC cell lines contained relatively large amounts of Gb3 but did not contain extended globo-series glycolipids Gb5 and GL7. CC cell lines also con- tained a macroglycolipid reactive with the antibody to SSEA- I (Le”). Glycolipids were not detected in two YC cell lines, while other YC cell lines contained globo-series core glycolipids (Gb3 and Gb4) and gangliosides. We conclude that EC, YC and CC have distinct patterns of membrane glycolipid expression that can be identified by HPTLC and immunostaining. Our results indicate that globo-series glycolipids Gb5 and GL7, which carry stage-specific embryonic antigens 3 and 4 (SSEA-3 and SSEA-4), are a hallmark of human EC cells. Cell lines derived from human germ cell tumors that do not express Gb5 and GL7 deserve to be re-evaluated, since they may represent different stem cells, most likely equivalent t o somatic cells and their developmen- tally committed precursors (e.g., neuroblasts).
< 1904 M ~ l l C V - L l S S , h1c.
Testicular germ cell tumors are among the most common groups of cancer in young adult males (reviewed by Damjanov, 1991). They are traditionally divided into seminomas (S) and non-seminomatous germ cell tumors (NSGCT). The latter group includes (i) teratocarcinomas composed of embryonal carcinoma (EC) and various somatic and extraembryonic derivatives of these stem cells, (ii) pure EC composed of stem cells that cannot differentiate, and (iii) choriocarcinoma (CC) and yolk sac carcinoma (YC), which may be associated with teratocarcinomas or grow as distinct tumors.
Our approach to the study of germ cell tumors has been based on monoclonal antibodies (MAbs) that react with cell surface antigens expressed on tumor stem cells (reviewed by Fenderson and Andrews, 1992). Detailed biochemical studies have shown that many of these antigens are oligosaccharides carried on membrane glycolipids and glycoproteins. For techni- cal reasons, the carbohydrate epitopes of glycolipids have proved easier to analyze than those of glycoproteins. Glycolip- ids are relatively easy to extract from whole cells, to separate on high-performance thin-layer chromatography (HPTLC) plates, to identify using anti-carbohydrate MAbs and to purify. This approach may overlook carbohydrate antigens that are carried exclusively on glycoproteins. However, our recent studies using an inhibitor of glycolipid biosynthesis (PDMP)
suggest that many carbohydrate differentiation antigens are carried either exclusively on glycolipids or on both glycolipids and glycoproteins (Fenderson et al., 1993). In either case, glycolipid analyses provide a reproducible means for character- izing oligosaccharide antigens and the cells that carry them.
Previously, we focused on the glycolipids of a pluripotent human EC cell line, NTERA-2 (Andrews et al., 1984), and asked the following questions: ( i ) What are the major glycolip- ids in these pluripotent EC cells and how does glycolipid composition change during chemically induced differentiation (Fenderson et al., 1987; Andrews et al., 1990)? (ii) What is the enzymatic basis of these differentiation-dependent glycolipid changes (Chen et al., 1989)? (iii) What is the effect of inhibition of glycolipid synthesis on surface antigen expression and cell differentiation (Fenderson et al., 1993)? Our results suggested that globo-series glycolipids of the P blood group system are the most prominent (immunodominant) markers of human EC cells and that differentiation of EC cells is accompanied by a shift in glycolipid assembly from globo-series to lacto- and ganglio-series (reviewed by Fenderson and Andrews, 1992).
In the present study, as part of a NATO Advanced Studies Workshop on “Phenotypic Markers of Human Germ Cell Tumors,” we examined the glycolipid composition of over 40 different human germ cell tumor lines to test the generality of our conclusions, based on the study of the NTERA-2 cell line. The results corroborate our previous findings and indicate that globo-series glycolipids carrying stage-specific embryonic anti- gens 3 and 4 (SSEA-3 and SSEA-4) are present on all human EC cells, but not on YC, CC or other stem cells derived from human germ cell tumors. A summary of the workshop findings, including detailed information on cell surface, cytoplasmic and genetic markers of human germ cell tumor lines, is in prepara- tion.
MATERIAL AND METHODS Established cell lines
Human germ cell tumor lines were received in Philadelphia as either lyophilized or frozen samples, representing 0.2-0.5 ml packed cells. A complete list of these cells and their providers is shown in Table I. A tentative phenotype was assigned to each cell line based primarily upon published data. However, in certain cases these designations were modified, prior to glycolipid analysis, based upon morphological and other obse‘r-
7To whom corresoondence should be addressed. at Deoartment of Pathology and Cell Biology, Thomas Jefferson University, 1020 Locust Street, Rm. 233 Jefferson Hall, Philadelphia, PA 19107 USA. Fax: (215) 955-8703.
Received: December 21, 1993 and in revised form February 22, 1994.

GLYCOLIPIDS OF HUMAN GERM CELL TUMORS 109
TABLE I - GLYCOLIPID EXPRESSION IN HUMAN GERM CELLTUMOR LINES
Cell line Ph Provider References Gb3 Gb4 Gb5 GLI Le‘ GD3 9-0-GD3 GT3 9-0-GT3 GD2
1. H12.1 2. H12.7 3. H32 4. NCR-G2 5. NCR-G3 6. GCT27C4 7. GCT27XG 8. GCT35 9. GCT44
10. GCT46 11. GCT48 12. GCT72 13. NT2/D1 14. 2102Ep (p l l ) 15. s2.2 16. 833KE 17. T84-5130 18. SUSA-scraped 19. SUSA-trypsin 20. 1218K _ _ ~~~~~~
21. UMTC-1 22. NCCIT 23. NOTHAmass 24. 1618K _ _ 25. 2102E RP met 26. 1777N RP met 27. NT2/D1 28. PC21 A1 mouse 29. 1156QE 30. TERA-1 31. 1777N RP met 32. 2102E~ ( ~ 2 4 ) 33. 577MC \L ’ 34. 2806B 35. NT2/D1 HB21 36. NT2/D1 RA23 37. NT2/DlRA7 38. NT2/D1 HB7 39. NT2IDl HB14, 40. NT2/D1 RA14 41. HAZ-1 42. HAZ-3 43. PA1-6928 44. PA1-CLlL59 45. PA1-9117T1 46. PA1-9115T1 47. PA1 Darental 48. 1411H 49. 1411H 50. 1411H RO met - 51. JAR 52. BEWO
EC EC EC EC EC EC EC EC YC YC EC YC1 EC EC S EC EC EC EC EC D EC EC EC EC EC EC EC1 EC EC D EC D D D D D D D D D D Dn Dn Dn Dn Dn YC YC YC cc cc
Casper Casper Casper Hata Hata Pera Pera Pera Pera Pera Pera Pera von Keitz Andrews von Keitz Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews Andrews
9 9 10 16 16 23,24 23,24 23 23 23 23 23 5,13 3,31 30 3,7, 31 12,19 18 18 3, 31 15 11,27 1 28 31,32 8 5,13 4 3,31 3, 13 8 3,31 3,31 17 6 2 2 6 6 2 20 20 25,26 25,26 25,26 25,26 14,33 29,32 29,32 29 22 21
+++ +++ +++ +++ +++ +++ +++ +++ + +++ ++ ++ +++ + +++ +++
+++ + +++ + ++ ++ +++ +++ +++ + +++ +++
++ ++ +++ ++ ++ +++ ++ ++ +++ + +++ +++ + + +++ ++ + +++ ++ +++ +++
++
++ ++ ++ ++ +++ ++ ++ ++ ++ ++ ++ + ++ ++ +++ ++ + ++ ++
++ ++ ++ + ++ ++
++ ++ + ++ +++ + ++
+++ +++
+++ +++ +++ ++ +++ +++ +++ + +++ +++ ++ +++ +++ +++ +++ +++ +++
++ + ++ +++ +
+ + ++ ++ ++
+++ ++
+
+ ++
+ ++
+ ++-I
+
t+ + ++ ++ ++ + +
+ +
+ + +
+ +
+
+
++ + +++ +++ +++ ++ + + ++
+++ ++ + + +++ + +++ + +++
+++ ++ +++ + +++
+
. . . . Human cell lines were derived from tumors reported to have testicular rimaries, except for 1618K, HAZ and NCCIT (all
extragonadal), PA1 (ovary) and BEWO and JAR (gestational choriocarcinomasy A tentative phenotype (Ph) was assigned to each cell line, based rimarily upon published data. Differentiation of NT2/D1 EC cells was induced by treating cells for 7, 14 or 21 days in the presence o?either 10 FM RA or 3 mM HMBA. GCT72 is known to exhibit characteristics of visceral endoderm (Pera et al., 1987) and was therefore given a classification of YC1 to distinguish it from the other YC cell lines which resemble parietal endoderm. Samples of 2102Ep EC cells from passages 11 and 24 ( p l l and p24) were analyzed. NT2/D1 EC cells were grown and provided by different laboratories. PC21A1 is a somatic cell hybrid between mouse PCC4aza EC cells and human 2102Ep EC cells, and in most respects, resembles the mouse parental cell line (Andrews et al., 1983). It was therefore given a classification of EC1 to distinguish it from the human EC cell lines. Results represent a synthesis of HPTLC orcinol and immunostaining data. The scale is negative (no symbol) to strong positive (+ + +). LeX antigen was in all cases found to be carried on a macroglycolipid. References: 1. Andrews (data not shown); 2. Andrews, 1984; 3. Andrews, et al., 1980; 4. Andrews et al., 1983; 5. Andrews et al., 1984; 6. Andrews eta[., 1990; 7. Bronson et al., 1980; 8. Bronson et al., 1983; 9. Casper et al., 1987; 10. Casper (data not shown); 11. Damjanov et al., 1993; 12. Fenderson et al., 1987; 13. Fogh and Trernpe, 1975; 14. Giovanella et al., 1974; 15. Grossman and Wedemeyer, 1986; 16. Hata et al., 1992; 17. Hiraoka et aL, 1988; 18. Hogan et al., 1977; 19. Oosterhuis (data not shown); 20. Oosterhuis et al., 1985; 21. Pattillo and Gey, 1968; 22. Pattillo et al., 1971; 23. Pera et al., 1987; 24. Pera et al., 1989; 25. Tainsky et al., 1984; 26. Tainsky et al., 1988; 27. Teshima et al., 1988; 28. Vogelzang et al., 1983; 29. Vogelzang et aL, 1985; 30. von Keitz et al., 1994; 31. Wang et al., 1980; 32. Wang et al., 1981; 33. Zeuthen et al., 1980.
vations made in the laboratories providing samples (data not shown, but see Andrews and Damjanov, 1994). Thus 2806B and UMTC-1 were both reported to be EC cell lines by their originators, but in the present study these cell lines did not
present the characteristic morphology and growth pattern of other EC cells, nor did they express characteristic EC cell- surface antigens (e.g., SSEA-3 and SSEA-4). They were therefore designated as having a “differentiated” phenotype.

110 WENK ETAL.
The ovarian cell line PA1, which has been described as an EC cell line (Zeuthen et al., 1980), did not in our hands resemble other EC cell lines in culture. Moreover, PA1 and its sublines derived by Tainsky et al. (1984, 1988) formed tumors that in our opinion were predominantly neuroepithelial. Also, one of us (I.D.) has reviewed the histology of the tumor from which PA1 was derived and concluded that it would best be described as an immature teratoma with neuroepithelial fea- tures rather than as a teratocarcinoma containing an EC element. Accordingly, for this study, the PA1 sublines were designated as having a “differentiated neuroepithelial” pheno- type.
SUSA EC cells exhibited striking differences in growth and morphology depending on how the cells were passaged. If cultures were passaged by scraping, which tends to keep cells in clumps, a typical EC morphology was maintained. However, if cultures were passaged using trypsin, which disperses the cells more, cultures became much more heterogeneous, with many cells exhibiting a non-EC (differentiated) cell appear- ance. Samples of both types of culture (i.e., SUSA-scraped and SUSA-trypsin) were analyzed. Differentiation of the NTERA-2 subline, designated NT2/D1, was achieved by culturing cells for 7, 14 or 21 days in the presence of either 10 )LM all-trans retinoic acid (RA) or 3 mM hexamethylene bisacetamide (HMBA), as described previously (Fenderson et al., 1987; Andrews et aL, 1990). RA was from Eastman Kodak (Roches- ter, NY); HMBA was from Sigma (St. Louis, MO).
Glycolipid extraction Glycolipids were extracted from cell pellets using isopropyl
alcoho1:hexane:water (55:25:20, v/v/v), as described previously (Fenderson et al., 1987). In brief, total lipid extracts were partitioned into an upper and lower phase. The upper phase was desalted using C18 reverse phase columns (Analytichem, Harbor City, CA). Phospholipids were not removed due to small sample sizes. For HPTLC immunostaining with anti- SSEA-3 antibody, upper phase glycolipids were first passed over DEAE-Sephadex A-25 to remove acidic glycolipids (gan- gliosides), which have been shown to mask antibody binding to neutral glycolipid antigens (Fenderson et al., 1987).
Glycolipid analysis Specific glycolipids were identified by co-migration with
pure glycolipid standards, and HPTLC immunostaining with specific anti-carbohydrate MAbs (Fenderson et al., 1987; Andrews et al., 1990). In brief, glycolipid samples were streaked onto Whatman HP-KF silica gel plates and subjected to ascending chromatography using a solvent system of chloro- form:methanol:water (50:40 10, v/v/v) containing 0.05% (w/v) CaC12. After drying, HPTLC plates were coated with 0.5% (w/v) polyisobutyl-methacrylate (Aldrich, Milwaukee, WI) in diethyl ether for 1 min, blocked for 2 hr with 5% BSA in phosphate-buffered saline (PBS) and then reacted with pri- mary antibody overnight. Bound antibody was detected using alkaline phosphatase-conjugated goat anti-mouse antibody (HyClone, Logan, UT) diluted 1:lOOO. Color reaction was obtained using bromochloroindolyl phosphate and nitroblue tetrazolium.
MAbs were obtained and used as described previously (Fenderson et al., 1987; Andrews et al., 1990). Gb3 was detected using antibody 1A4E10; Gb4 and Gb5 were detected using antibody to SSEA-3; GL7 was detected using antibody to SSEA-4; blood group A type 3 chain (globo-A) was detected using antibody HH5; LeX was detected using antibody to SSEA-1; GD3 was detected using antibody R24; 9-0-acetyl- GD3 was detected using antibody ME311; GT3 and 9-0-acetyl- GT3 were detected using antibody A2B5; GD2 was detected using antibody VIN-2PB-22. Glycolipids are designated accord- ing to the recommendations of the IUPAC-IUB Nomenclature Committee (1978).
RESULTS Phenotypic designation of cell lines
Prior to glycolipid analysis, each cell line was assigned a putative phenotype to assist in evaluating the results. Cells were designated as having an S, EC, YC, CC, undefined differentiated (D) or differentiated neuroepithelial (Dn) phe- notype, based upon published descriptions and our morphologi- cal observations (see “Material and Methods”).
HPTLC analysis The glycolipid composition of each cell line was analyzed by
HPTLC. The profile of lower-phase (non-polar) and upper- phase (polar) glycolipids is shown in Figures 1 and 2 respec- tively. Specific glycolipid antigens, identified by immunostain- ing with anti-glycolipid antibodies, are shown in Figures 3 and 4. Our results are summarized in Table I and glycolipid structures are shown in Table 11. Our major findings are summarized as follows.
EC cell lines. All EC cell lines tested (n = 25) contained high levels of globo-series glycolipids, including globo-triosylce- ramide (Gb3), globoside (Gb4), galactosyl globoside (Gb5) and sialyl galactosyl globoside (GL7). The glycolipids of early- and late-passage 2102Ep EC cells were nearly identical, suggesting stability of globo-series glycolipid expression in culture. However, subtle differences in glycolipid expression among the putative EC cell lines were evident. For example, a few EC cell lines (notably NCR-G2, GCT27C4, GCT27XG, SUSA-trypsin and 1777N RP met) expressed an abundance of GL7 but very little Gb5. Globo-series blood group A antigen was readily detected in only two EC cell lines, NT2/D1 and SUSA (data not shown).
Lacto- and ganglio-series glycolipids, such as GD3, GD2 and Lex, were detected in several EC cell lines (eg., NOTH A mass, NT2/D1, SUSA, NCR-G2 and GCT27XG). These glycolipids have been identified previously as markers of cell differentia- tion (Fenderson et al., 1987; Andrews et al., 1990). Thus the presence of lacto- and ganglio-series glycolipids in extracts of cloned EC cell lines could indicate spontaneous differentia- tion. This hypothesis is supported by analysis of glycolipid expression in SUSA cells passaged by scraping vs. trypsin. In keeping with the morphological evidence for spontaneous differentiation, SUSA cells passaged with trypsin exhibited a notable decrease in globo-series glycolipids (Gb5 and GL7) and an increase in lacto- and ganglio-series glycolipids (Le” and GD3).
Differentiated derivatives of EC stem cells. Two types of differentiated cell were analyzed in our study: (i) cell lines lacking an EC morphology and characteristic EC cell surface antigens (1777N RP met diff, 2806B, 577MF, HAZ-1 and HAZ-3) and (ii) derivatives of pluripotent NT2/D1 EC cells induced to differentiate for varying lengths of time by culture in the presence of RA or HMBA. In both types of differenti- ated cell (i.e., established cell lines and chemically induced terminal cultures), globo-series glycolipids were only weakly detected. The minor band observed at the position of Gb3 (Fig. 1, CTH) was not labeled with anti-Gb3 antibody, and its lack of color change upon orcinol-charring suggested that it was a phospholipid. The glycolipid band co-migrating with Gb4 standard was also not labeled with anti-SSEA-3 antibody, suggesting another tetraosylceramide, most likely paraglobo- side or asialo-GM1. RA- and HMBA-treated NT2/D1 EC cells both exhibited a significant decrease in globo-series glycolipid expression and an increase in ganglio-series glycolip- ids, including GD3,9-O-acetyl GD3, GT3,9-O-acetyl GT3 and GD2 (Fig. 4). Gangliosides were also detected in the estab- lished cell lines 2806B, 577MF and UMTC-1, supporting morphological evidence that they are not EC cell lines. However, lacto- and ganglio-series glycolipids were not de-

GLYCOLIPIDS OF HUMAN GERM CELL TUMORS 111
FIGURE 1 - Lower-phase glycolipids of human germ cell tumor lines. Each lane represents the glycolipid equivalent of approximately 2 pg cellular protein. Glycolipid standards (S) are included on the left side of each plate. HPTLC plates were develo ed using chloroform/methanol/water (50:40:10) containing 0.05% CaCI2 and stained for carbohydrate by the orcinol/sulfuric acixreaction. Samples are identified by number in Table I . Note the prominent expression of ceramide trihexoside (CTH/Gb3) in extracts of most EC cell lines.
tected in three other differentiated cell lines (1777N RP met diff, HAZ-1 and HAZ-3).
The PA1 sublines. As discussed above (“Material and Methods”), PA1 appears to represent a differentiated, neuro- epithelial stem cell line originating from an immature ovarian teratoma. PA1 and its 4 sublines contained the globo-series core glycolipid Gb3 but lacked the extended globo-series glycolipids (Gb5 and GL7) which characterize the whole panel of testicular EC cells that we have analyzed. Despite their apparent neuroepithelial character, the PA1 sublines did not contain gangliosides GD3, 9-0-acetyl GD3, GT3, 9-0-acetyl GT3 or GD2 (Fig. 4).
CC cell lines. Gestational CC cell lines BEWO and JAR contained an abundance of globo-triosylceramide (Gb3) but lacked extended globo-series glycolipids Gb5 and GL7, which are characteristic of human EC cells. BEWO and JAR also expressed a macroglycolipid reactive with antibody to SSEA-1 (Lex), further distinguishing them from the panel of EC cells.
YC cell lines. Glycolipids were not detected by either orcinol reaction or HPTLC immunostaining in 2 isolates of the YC cell line 1411H (testicular primary and metastatic nodule). In contrast, 2 other YC cell lines with distinct properties of parietal endoderm (GCT44 and GCT46) contained globo- series core glycolipids Gb3 and Gb4, as well as gangliosides. The profile of gangliosides in GCT44 was similar to that found in NT2/D1 EC cells induced to differentiate with RA or HMBA (Fig. 4). A fifth YC cell line with properties of visceral endoderm (GCT72) simply expressed the globo-series core glycolipid Gb3 (Fig. 1).
Other cell lines. The S2 cell line is reported to represent either seminoma or an intermediate cell type between semi- noma and EC (von Keitz et al., 1994). Like the panel of EC cells tested, S2 cells contained extended globo-series glycolip- ids Gb4 and GL7; however, Gb5 was not detected. Like SUSA
EC cells, S2 seminoma-like cells contained a macroglycolipid reactive with the antibody to SSEA-1 (LeX). However, unlike other EC cell lines tested, the S2 cell line contained a large amount of lactosylceramide (CDH, Fig. 1). The mouse-human hybrid cell line PC21A1 contained trace amounts of globo- tetraosylceramide (Gb4) and the disialyl ganglioside GD3.
DISCUSSION
In the present report, we examined the glycolipid profile of over 40 different human germ cell tumor lines to identify reliable immunohistochemical markers for clinical and basic research. Our results indicate that EC, YC and CC cell lines express distinct sets of glycolipid antigens that can be identi- fied by HPTLC and immunostaining with anti-glycolipid MAbs (Tables I, 11). Extended globo-series glycolipids Gb5 and GL7 were identified in all human EC cell lines tested, including cell lines derived from testicular and extra-gonadal primaries. These results confirm previous studies of glycolipid expression in 2102Ep (Kannagi et al., 1983a,b) and NT2/D1 (Fenderson et al., 1987) EC cells and complement the results of recent cell surface immunolabeling assays (data not shown).
Gb5 and GL7 were described initially as carriers of 2 developmentally regulated antigens, SSEA-3 and SSEA-4. Anti-SSEA-3 antibody was shown to react with Gb4, Gb5 and GL7 (Kannagi et al., 1983b), whereas anti-SSEA-4 antibody was shown to react exclusively with GL7 (Kannagi et al., 1983~). In addition to human EC cells, SSEA-3 and -4 are present on mouse oocytes, cleavage-stage embryos, and extra- embryonic endodermal cells (Shevinsky et al., 1982; Kannagi et al., 1983~; Fox et al., 1984). These antigens are also present on human erythrocytes and belong to the P blood group system: SSEA-3 correlates with the P antigen, whereas SSEA-4 is
112 WENK ETAL.
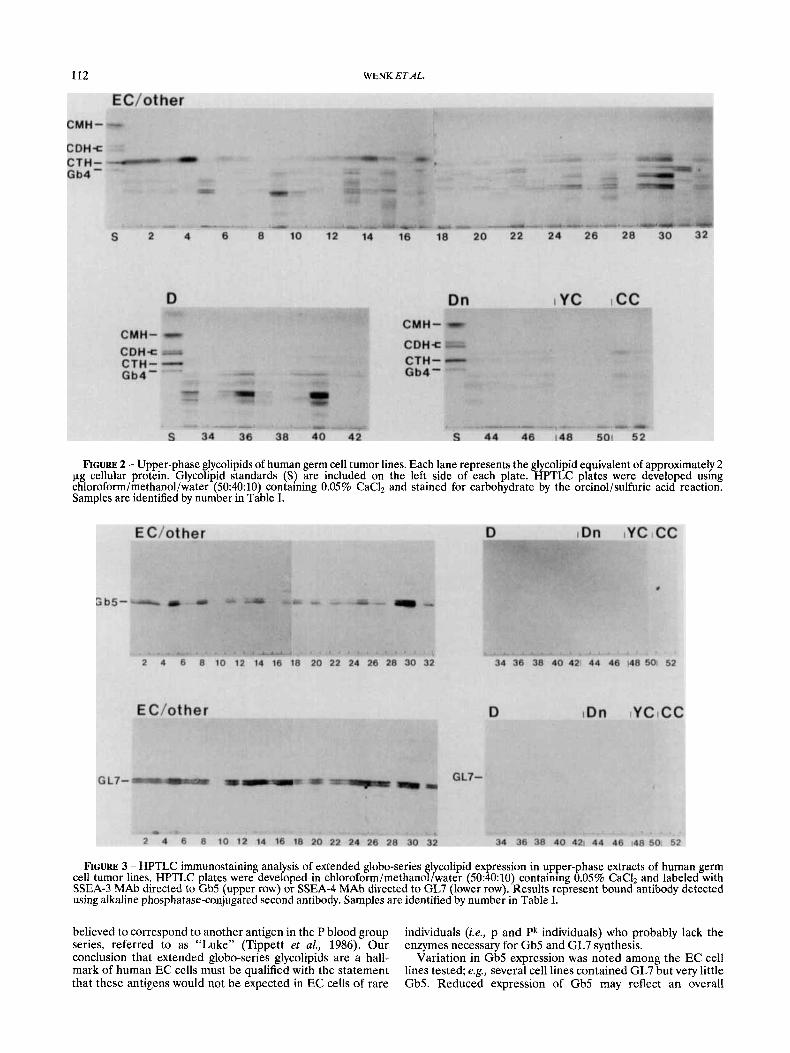
FIGURE 2 - Upper-phase glycolipids of human germ cell tumor lines. Each lane represents the glycolipid equivalent of approximately 2 pg cellular protein. Glycolipid standards (S) are included on the left side of each plate. HPTLC plates were developed using chloroform/methanol/water (50:40:10) containing 0.05% CaCI, and stained for carbohydrate by the orcinol/sulfuric acid reaction. Samples are identified by number in Table I.
FIGURE 3 - HPTLC immunostaining analysis of extended globo-series glycolipid expression in upper-phase extracts of human germ cell tumor lines. HPTLC plates were developed in chloroform/methanol/water (50:40:10) containing 0.05% CaC1, and labeled with SSEA-3 MAb directed to Gb5 (upper row) or SSEA-4 MAb directed to GL7 (lower row). Results represent bound antibody detected using alkaline phosphatase-conjugated second antibody. Samples are identified by number in Table I.
believed to correspond t o another antigen in the P blood group series, referred to as “Luke” (Tippett et al., 1986). Our conclusion that extended globo-series glycolipids are a hall- mark of human EC cells must be qualified with the statement that these antigens would not be expected in EC cells of rare
individuals (ie., p and Pk individuals) who probably lack the enzymes necessary for Gb5 and GL7 synthesis.
Variation in Gb5 expression was noted among the EC cell lines tested; eg., several cell lines contained GL7 but very little Gb5. Reduced expression of Gb5 may reflect a n overall
GLYCOLIPIDS OF HUMAN GERM CELL TUMORS 113
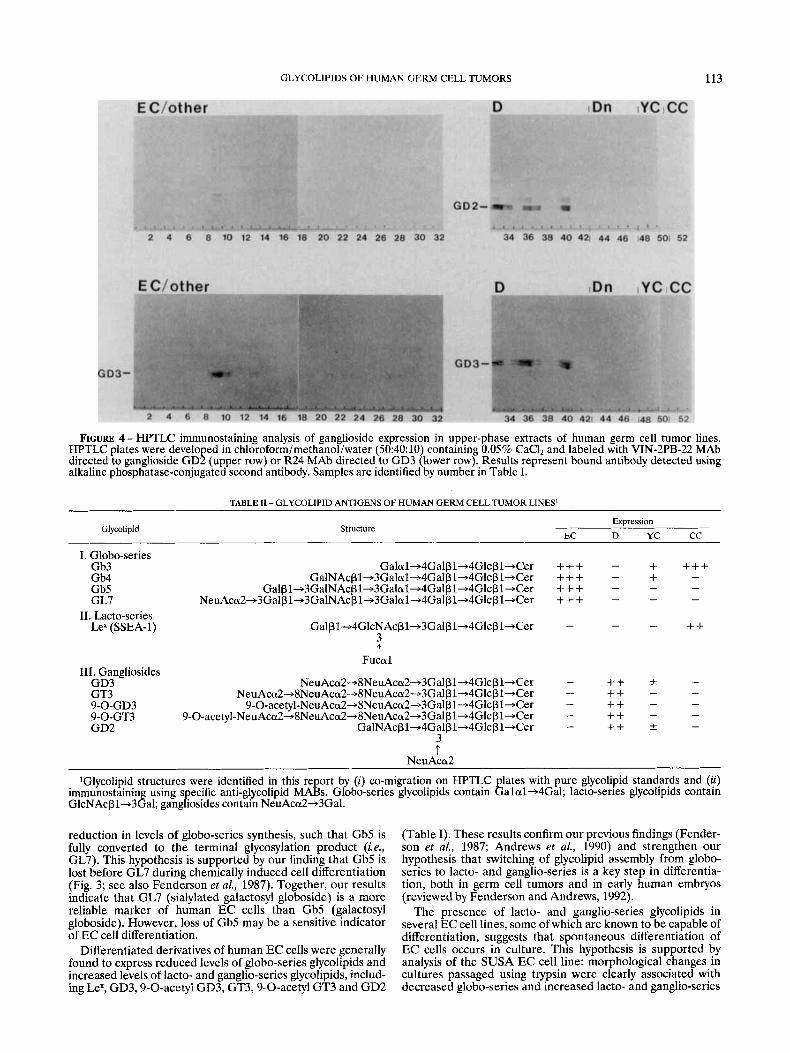
FIGURE 4 - HPTLC immunostaining analysis of gan lioside expression in upper-phase extracts of human germ cell tumor lines. HPTLC plates were developed in chloroforrn/methanof/water (50:40:10) containing 0.05% CaCl, and labeled with VIN-2PB-22 MAb directed to ganglioside GD2 (upper row) or R24 MAb directed to GD3 (lower row). Results represent bound antibody detected using alkaline phosphatase-conjugated second antibody. Samples are identified by number in Table I.
TABLE I1 - GLYCOLIPID ANTIGENS OF HUMAN GERM CELL TUMOR LINES'
Expression EC D YC CC
Glycolipid Structure
I. Globo-series Gb3 Gb4 Gb5 GL7
11. Lacto-series Lex (SSEA-1)
Galal-+4Galpl+4Glcpl+Cer + + + - + + + + - GalNAc~1+3Galal-+4Gal~l+4Glcpl+Cer + + + - +
Gal~1+3GalNAcpl+3Galal+4Galpl+4Glcpl-+Cer + + + - - - NeuAccu2+3Galp1+3GalNAc~1+3Galal+4Galpl+4Glcpl+Cer + + + - - -
- ++ - - Gal~l+4GlcNAcpl+3Galpl-+4Glcpl+Cer 3 t
Fucal 111. Gangliosides
- NeuAccu2-+8NeuAca2-+3Galp1+4Glcpl+Cer - ++ * NeuAcd-+8NeuAca2-+8NeuAca2-+3Galp1+4Glcpl+Cer - ++ -
GD2 GalNAc~1+4Galpl+4Glcpl+Cer - ++ &
- GD3 GT3
- ++ - ++ - - 9-O-acetyl-NeuAca2-+8NeuAca2-+3Galp1+4Glcpl+Cer
9-O-acetyl-NeuAca2+8NeuAca2~8NeuAca2+3Gal~l+4Glc~l +Cer - - 9-0-GD3 9-0-GT3
- 3 t
NeuAca2 ~
'Glycolipid structures were identified in this report by (i) co-migration on HPTLC plates with pure glycolipid standards and (ii) immunostaining using specific anti-glycolipid MAF3s. Globo-series glycolipids contain Galal+4Gal; lacto-series glycolipids contain GlcNAcpl+3Gal; gangliosides contain NeuAca2+3Gal.
reduction in levels of globo-series synthesis, such that Gb5 is fully converted to the terminal glycosylation product (i.e., GL7). This hypothesis is supported by our finding that Gb5 is lost before GL7 during chemically induced cell differentiation (Fig. 3; see also Fenderson et al., 1987). Together, our results indicate that GL7 (sialylated galactosyl globoside) is a more reliable marker of human EC cells than Gb5 (galactosyl globoside). However, loss of Gb5 may be a sensitive indicator of EC cell differentiation.
Differentiated derivatives of human EC cells were generally found to express reduced levels of globo-series glycolipids and increased levels of lacto- and ganglio-series glycolipids, includ- ing Lex, GD3,g-O-acetyl GD3, GT3,9-O-acetyl GT3 and GD2
(Table I). These results confirm our previous findings (Fender- son et al., 1987; Andrews et al., 1990) and strengthen our hypothesis that switching of glycolipid assembly from globo- series to lacto- and ganglio-series is a key step in differentia- tion, both in germ cell tumors and in early human embryos (reviewed by Fenderson and Andrews, 1992).
The presence of lacto- and ganglio-series glycolipids in several EC cell lines, some of which are known to be capable of differentiation, suggests that spontaneous differentiation of EC cells occurs in culture. This hypothesis is supported by analysis of the SUSA EC cell line: morphological changes in cultures passaged using trypsin were clearly associated with decreased globo-series and increased lacto- and ganglio-series

114 WENK ETAL.
glycolipid expression (Table I). The developmental status of non-EC cells appearing in mixed cultures remains unknown; however, gangliosides are known to be associated with neurons (Andrews, 1984; Fenderson et aL, 1987) and certain endoder- ma1 cells derived from germ cell tumors (eg., GCT44). HOW- ever, only 3 of 6 spontaneously differentiated germ cell tumor lines contained gangliosides, indicating that these glycolipids are not an invariant marker of somatic cell differentiation.
All PA1 sublines contained the globo-series core structure Gb3 but lacked extended globo-series glycolipids Gb5 and GL7, which are characteristic of human EC cells (Table I). Similar results were reported previously by Fukuda et aL, (1986), who also noted the presence of unusual mono- and disialyl glycolipids with blood group type 1 sequence in these cells. PA1 cells are clearly distinct from other human EC cell lines in culture. Based on their glycolipid analysis, Fukuda et al. (1986) speculated that these cells represent a developmen- tal stage different from other mouse and human EC cells (either an earlier or a later stage). However our results (Table I), as well as other morphological and biochemical data (Andrews and Damjanov, 1994), strongly suggest that PA1 is not an EC cell line. Indeed, as discussed above, these cells appear to represent neuroepithelial stem cells derived from an immature ovarian teratoma.
The CC cell lines BEWO and JAR contained an abundance of globo-series core structure Gb3 but lacked extended globo- series glycolipids Gb5 and GL7, which are characteristic of human EC cells. In this respect, BEWO and JAR display a glycolipid profile very similar to the PA-1 sublines. However, unlike most of the PA-1 sublines (PA1-6928 being an excep- tion), BEWO and JAR contained a macroglycolipid reactive with the MAb to SSEA-1 (Le"). This glycolipid was immobile in the solvent system used here and may be presumed to carry a large, fucosylated lactosaminoglycan. The presence of fuco- sylated glycans in human choriocarcinoma cell lines was reported previously by Blaineau et al. (1983).
HPTLC analysis did not reveal a consistent glycolipid pattern among the 5 YC cell lines examined (Table I). It will be important to examine a larger panel of YCs in order to draw firm conclusions regarding glycosylation in these germ cell tumors. Similarly, our HPTLC analysis did not clarify the differentiation status of the S2 seminoma-like cell line. One previous study of glycolipid expression in testicular tumors noted an accumulation of Gb3 in clinical samples of both S and EC (Ohyam et al., 1990). However, unlike EC cell lines, the S2 cell line contained reduced levels of Gb3 and increased levels of CDH (Fig. 1). At the present time, all we can say is that the glycolipids of S2 are similar to those of human EC cells.
Differences in glycolipid composition between discrete histo- logic types of cells are believed to reflect differences in glycosyltransferase activity and gene expression. This hypoth- esis is supported by the results of a detailed analysis of glycosyltransferase activity in NT2/Dl EC cells (Chen et aL, 1989). In this study, the activities of a1 -+ 4galactosyltransfer- ase (the key enzyme for synthesis of globotriosyl core struc- ture) and P l -+ 3 galactosyltransferase (the key enzyme for synthesis of Gb5 and GL7) were both reduced 4-fold during RA-induced differentiation. By contrast, the activities of p l -+
3N-acetyl-glucosaminyltransferase (the key enzyme for synthe- sis of lactotriosyl core structure) and a2 -+ 3sialytransferase (the key enzyme for synthesis of ganglio core structure) were both increased during RA-induced differentiation. Since some of these enzymes have been characterized and their respective genes cloned, it will be possible in the near future to relate changes in membrane glycolipids to changes in glycosyltransfer- ase gene expression.
GERM CELL TUMOR
(++) Embryonal Carcinoma
(++) Choriocarcinoma
(++) Differentiated Somatic Cell
FIGURE 5 - Scheme summarizing the use of glycolipid analysis for diagnosis of human germ cell cancer. Embryonal carcinomas are characterized by the expression of extended globo-series glycolipids (Gb5 and GL7) which carry stage-specific embryonic antigens 3 and 4 (SSEA-3 and SSEA-4). Choriocarcinomas are characterized by the expression of globo-triosylceramide (Gb3) and macroglycolipid reactive with antibody to SSEA-1 (Le"). Differentiated derivatives of pluripotent stem cells may express ganglio-series glycolipids including GD3, GT3 and GD2.
In summary, our results suggest that glycolipid composition analysis provides an improved basis for classifying germ cell tumors (Fig. 5; Table 11). Thus an immunohistochemical finding of SSEA-3 and SSEA-4 (Gb5 and GL7) indicates embryonal carcinoma, whereas SSEA-1 macroglycolipid and Gb3 indicates choriocarcinoma. Similarly, ganglioside expres- sion indicates neuroectodermal differentiation in a germ cell tumor. This classification scheme should be useful to surgical pathologists as an aid in tumor diagnosis. One qualification, however, is that tumor cells in vitro and in vivo may express different sets of membrane glycolipids. However, immunohis- tochemical studies have clearly demonstrated SSEA-3 and SSEA-4 in surgical specimens of EC (Damjanov et aL, 1982). Nonetheless, further studies of glycolipid expression in clinical specimens are warranted. Future studies on the glycolipids of gonocytes, carcinoma in situ and seminoma should provide further insights into the histogenetic progression of germ cell malignancies, as well as the role of aberrant glycosylation in the etiology of cancer.
ACKNOWLEDGEMENTS
This work was supported by Teaching and Research Train- ing grant CA48010 to Jefferson Medical College from the National Cancer Institute (J.W.), a Biomedical Research Support grant from Thomas Jefferson University (B.F.), USPHS grants CA29894 and A124943 from the National Institutes of Health (P.A.) and by funds from the UK Cancer Research Campaign (M.P.). The Advanced Studies Workshop held in Oxford, UK (August 1992), was supported by NATO and the Cancer Research Campaign. Antibody R24 was the generous gift of Dr. K. Lloyd (New York). Antibody HH5 was the generous gift of Dr. H. Clausen (Copenhagen). Antibody ME311 was the generous gift of Dr. M. Herlyn (Philadelphia).
REFERENCES
ANDREWS, P.W., Retinoic acid induces neuronal differentiation of a cloned human embryonal carcinoma cell line in vitro. Dev. Biol., 103, 285-293 (1984).
ANDREWS, P.W. and DAMJANOV, I., Cell lines from human germ cell tumors. In: R.J. Hay, J.-G. Park and A. Gazdar (eds.), Atlas of Human Tumor Cell Lines, pp. 443-476, Academic Press, San Diego (1994).

GLYCOLIPIDS OF HUMAN GERM CELL TUMORS 115
ANDREWS, P.W., BRONSON, D.L., BENHAM, F., STRICKLAND, S. and KNOWLES, B.B., A comparative study of eight cell lines derived from human testicular teratocarcinoma. Inf. J. Cancer, 26,269-280 (1980). ANDREWS, P.W., DAMJANOV, I., SIMON, D., BANTING, G., CARLIN, C., DRACOPOLI, N.C. and FOGH, J., Pluripotent embryonal carcinoma clones derived from the human teratocarcinoma cell line Tera-2: differentiation in vivo and in vitro. Lab. Invest., 50,147-162 (1984). ANDREWS, P.W., GOODFELLOW, P.N. and DAMJANOV, I., Human teratocarcinoma cells in culture. Cancer Suw., 2,41-73 (1983). ANDREWS, P.W., NUDELMAN, E., HAKOMORI, S. and FENDERSON, B.A., Different patterns of glycolipid antigens are expressed following differentiation of TERA-2 human embryonal carcinoma cells induced by retinoic acid, hexamethylene bisacetamide (HMBA) or bromodeoxy- uridine (BUdR). Differentiation, 43,131-138 (1990). BLAINEAU, C., LE PENDU, J., ARNAUD, D., CONNAN, F. and AVNER, P., The glycosidic antigen recognised by a novel monoclonal antibody, 75.12, is developmentally regulated on mouse embryonal carcinoma cells. EMBO J., 2,2217-2222 (1983). BRONSON, D.L., ANDREWS, P.W., SOLTER, D., CERVENKA, J., LANCE, P.H. and FRALEY, E.E., A cell line derived from a metastasis of a human testicular germ-cell tumor. Cancer Res., 40,2500-2506 (1980). BRONSON, D.L., ANDREWS, P.W., VESSELLA, R.L. and FRALEY, E.E., In vitro differentiation of human embryonal carcinoma cells. In: L.M. Silver. G.R. Martin and S. Strickland (eds.). Terafocarcinoma Stem Cells, ’Cold Spring Harbor Conferences bn Cell Proliferation, 10, pp. 597605, Cold Spring Harbor Press, New York (1983). CASPER, J., SCHMOLL, H.-J., SCHNAIDT, U. and FONATSCH, C., Cell lines of human germinal cancer. Int. J. Androl., 10,105-113 (1987). CHEN, C., FENDERSON, B.A., ANDREWS, P.W. and HAKOMORI, S., Glycolipid glycosyltransferases in human embryonal carcinoma cells durin retinoic acid induced differentiation. Biochemisfiy, 28, 2229- 2238 f1989). DAMJANOV, I., Pathobiology of human germ cell neoplasia. Recent Results CancerRes., 123,l-19 (1991). DAMJANOV, I., Fox, N., KNOWLES, B.B., SOLTER, D., LANGE, P.H. and FRALEY, E.E., Immunohistochemical localization of rnurine stage- specific embryonic antigen in human testicular germ cell tumors. Amer.
DAMJANOV, I., HORVAT, B. and GIBAS, Z., Retinoic acid-induced differentiation of the developmentally pluripotent human germ cell tumor-derived cell line, NCCIT. Lab. Invest., 68,220-232 (1993). FENDERSON, B.A. and ANDREWS, P.W., Carbohydrate antigens of embryonal carcinoma cells: changes upon differentiation. In: H. Clausen and E. Dabelsteen (eds.), Carbohydrate Pathology, Acta pathol. microbiol. immunol. Scand. 100, 109-1 18 (1992). FENDERSON, B.A., ANDREWS, P.W., NUDELMAN, E., CLAUSEN, H. and HAKOMORI, S., Glycolipid core structure switching from globo- to lacto- and ganglio-series during retinoic acid-induced differentiation of TERA-2-derived human embryonal carcinoma cells. Dev. Biol., 122, 21-34 (1987). FENDERSON, B.A., W I N , N. and ANDREWS, P.W., Differentiation antigens of human germ cell tumors: distribution of carbohydrate epitopes on glycolipids and glycoproteins analyzed using PDMP, an inhibitor of glycolipid synthesis. Europ. Urol., 23,30-37 (1993). FOGH, J . and TREMPE, G., New human tumor cell lines. In: J. Fogh ed.), Human Tumor Cells In Vitro, pp. 115-159, Plenum, New York
i1975). Fox, N.W., DAMJANOV, I., KNOWLES, B.B. and SOLTER, D., Stage- specific embryonic antigen 3 as a marker of visceral extraembryonic endoderm. Dev. Biol., 103,263-266 (1984). FUKUDA, N.M., BOTHNER, B., LLOYD, K.O., RETTIG, W.J., TILLER, P.R. and DELL, A., Structures of glycosphingolipids isolated from human embryonal carcinoma cells. The presence of mono- and disialosyl glycoli ids with blood group type 1 sequence. J. biol. Chem., 261,5145-5153 6986) . GIOVANELLA, B.C., STEHLIN, J.S. and WILLIAMS, L.J., Heterotransplan- tation of human malignant tumors in “nude” thymusless mice. 11. Malignant tumors induced by injection of cell cultures derived from human solid tumors. J. nut. Cancer Inst., 52,921-930 (1974). GROSSMAN, H.B. and WEDEMEYER, G., Um-TC-1, a new human testicular carcinoma cell line. CancerJ., 1,22-24 (1986). HATA, J., FUJUMOTO, J., ISHII, E., KOKAI, Y., MATSUBAYASHI, Y., ABE, H., KUSAKARI, S., YAMADA, T., UMEZAWA, A,, KIKUCHI, H. and MARUYAMA, T., Differentiation of human germ cell tumor cells in vivo and vifro. Acta Histochem. Cytochem., 25,563-576 (1992). HIRAOKA, A., VOGELZANG, N.J., ROSNER, M.C. and GOLOMB, H.M., Ultrastructure of four human germ cell tumor-derived cell lines: effect
J. Pathol., 108,225-230 (1982).
of 12-0-tetradecanoylphorbol-13-acetate. Cancer Invest., 6, 393-402 (1988). HOGAN, B., FELLOUS, M., AVNER, P. and JACOB, F., Isolation of a human teratoma cell line which expresses F9 antigen. Nature (Lond.),
IUPAC-IUB, Commission on Biochemical Nomenclature. Biochem. J.,
KANNAGI, R., COCHRAN, N.A., ISHIGAMI, F., HAKOMORI, S., ANDREWS, P.W.. KNOWLES, B.B. and SOLTER. D.. Stage-soecific embrvonic
270,515-518 (1977).
171,21-35 (1978).
antigens (SSEA13 and -4) are epitopes of a‘ uiique globo-series ganglioside isolated from human teratocarcinoma cells. EMBO J., 2, 2355-2361 (1983~) . KANNAGI, R., LEVERY, S.B., ISHIGAMI, F., HAKOMORI, S., SHEVINSKI, L.H., KNOWLES, B.B. and SOLTER, D., New globoseries glycosphingo- lipids in human teratocarcinoma reactive with the monoclonal anti- body directed to a develo mentally regulated anti en, stage-specific embryonic antigen 3. J. bio! Chem., 258,8934-8942 t1983b). OHYAMA, C., FUKUSHI, Y., SATOH, M., SAITOH, S., ORIKASA, S., NUDELMAN, E., STROUD, M. and HAKOMORI, S., Changes in glycolipid expression in human testicular tumor. Int. J. Cancer, 45, 1040-1044 (1990). OOSTERHUIS, J.W., DEJONG, B., VAN DALEN, I., VAN DER MEER, I., VISSER, M., DE LEU, L., MESANDER, G., COLLARD, J.G., SCHRAFFORD KOOPS, H. and SLEIJFER, D.M., Identical chromosome translocations involving the region of the c-myb oncogene in four metastases of a mediastinal teratocarcinoma. Cancer Genet. Cytogenef., 15, 99-107 (1985). PATTILLO, R.A. and GEY, G.O., The establishment of a cell line of human hormone-synthesizing trophoblastic cells in vitro. Cancer Res.,
PATTILLO, R.A., RUCKERT, A,, HUSSA, R. BERNSTEIN, R. and DELFS, E., The Jar cell line. Continuous human multihormone production and controls. In Vitro, 6,398-399 (1971). PERA, M.F., BLASCO-LAFITA, M.J. and MILLS, J., Cultured stem-cells from human testicular teratomas: the nature of human embryonal carcinoma, and its comparison with two types of yolk-sac carcinoma. Int. J. Cancer, 40,334343 (1987). PERA, M.F., COOPER, S., MILLS, J. and PARRINGTON, J.M., Isolation and characterization of a multipotent clone of human embryonal carcinoma cells. Differentiation, 42,lO-23 (1989). SHEVINSKY, L.H., KNOWLES, B.B., DAMJANOV, I. and SOLTER, D., Monoclonal antibody to murine embryos defines a stage-specific
28,1231-1236 (1968).
embryonic antigen on mouse embryos and human teratocarcinoma cells. Cell, 30,697-705 (1982). TAINSKY, M.A., COOPER, C.S., GIOVANELLA, B.C. and VAN DE WOUDE, G.F., An activated rasN gene: detected in late but not early passage human PA-1 teratocarcinoma cells. Science, 225,643-645 (1984). TAINSKY, M.A., KRIZMAN, D.B., CHIAO, P.J., YIN, S O . and GIOV- ANELLA, B.C., PA-1, a human cell model for multistage carcinogenesis: oncogenes and other factors. Anticancer Rex, 8,899-914 (1988). TESHIMA, S., SHIMOSATO, Y., HIROHASHI, S., TOME, Y., HAYASHI, I., KANAZAWA, H. and KAKIZOE, T., Four new human germ cell tumor cell lines. Lab. Invest., 59,328-336 (1988). TIPPETT, P., ANDREWS, P.W., KNOWLES, B.B., SOLTER, D. and GOOD- FELLOW, P.N., Red cell antigens P (globoside) and Luke: identification by monoclonal antibodies defining the murine stage-specific embry- onic antigens -3 and -4 (SSEA-3 and -4). Vox Sang., 51,53-56 (1986). VOGELZANG, N., ANDREWS, P.W. and BRONSON, D., An extragonadal human embryonal carcinoma cell line 1618K. Proc. Amer. Assoc. Cancer Res., 24,3 (1983). VOGELZANG, N.J., BRONSON, D.L., SAVINO, D., VESSELLA, R.L. and FRALEY, E.E., A human embryonal-yolk sac carcinoma model system in athymic mice. Cancer, 55,2584-2593 (1985). VON KEITZ, A.T., RIEDMILLER, H., NEUMANN, K., GUTSCHANK, W. and FONATSCH, C., Establishment and characterization of a seminoma cell-line (S2). Invest. Urol., (1994) (In press). WANG, N., PERKINS, K.L., BRONSON, D.L. and FRALEY, E.E., Cytoge- netic evidence for pre-meiotic transformation of human testicular cancers. Cancer Res., 41,2135-2140 (1981). WANG, N., TREND, B., BRONSON, D.L. and FRALEY, E.E., Nonrandom abnormalities in chromosome 1 in human testicular cancers. Cancer Res., 40,796-802 (1980). ZEUTHEN, J., NORGAARD, J.O.R., AVNER, P., FELLOUS, M., WAR- TIOVAARA, J., VAHERI, A., ROSEN, A. and GIOVANELLA, B.C., Charac- terization of a human ovarian teratocarcinoma-derived cell line. Int. J. Cancer, 25,19-32 (1980).
Copyright © 2022 FDOKUMEN




















