
Lipase-Catalyzed Acidolysis of Egg-Yolk Phosphatidylcholine ...
Upload
independentCategory
view
2download
0
Int. J. Cancer: 40, 334-343 (1987) 0 1987 Alan R. Liss, Inc.
Publication of the International Unlon Against Cancer Publication de I'Unlon Internationale Contre le Cancer
CULTURED STEM-CELLS FROM HUMAN TESTICULAR TERATOMAS: THE
WITH TWO TYPES OF YOLK-SAC CARCINOMA Martin F. PERA', M.J. BLASCO LAFITA and Judith MILLS Radiotherapy Research Laboratories, Section of Radiotherapy, Institute of Cancer Research, Clifton Avenue, Sutton, Surrey SM2 5PX UK.
NATURE OF HUMAN EMBRYONAL CARCINOMA, AND ITS COMPARISON
The understanding of differentiation in human teratomas requires a better definition of the phenotype and develop- mental potential of the stem cells in these tumours. We de- scribe the characterization of 6 new cell lines from human testicular teratomas which are representative of 3 distinct cell types. Cell lines GCT 27, GCT 35, and GCT 48, identified as embryonal Carcinoma, comprise epithelial cells which express cytokeratin intermediate filaments and desrnoplakins, as de- termined by indirect immunofluorescence microscopy. A mi- nority of the cells also express vimentin. Most cells in these cultures show surface staining with monoclonal antibodies (MAbs) to stage-specific embryonic antigens SSEA 3 and SSEA 4 but not SSEA I. Staining with MAb W6/32, which recognizes HLA A,B and C chains in the presence of 6-2 microglobulin, is not above background level. When injected into nude mice, GCT 27 cells form tumours consisting of embryonal carci- noma, somatic tissues, and cells positive in immunocytochem- ical assays for alphafoetoprotein (AFP) and human chorionic gonadotrophin (HCG); GCT 35 cells form embryonal carcino- mas with foci of AFP and HCG-positive cells; and GCT 48 cells form embryonal carcinoma only. A second type of cell (GCT 72) displays some properties of rodent visceral endoderm. GCT 72 cells contain cytokeratin intermediate filaments, but not vimentin, and show very strong staining at cell borders with anti-desmoplakin I + II antibody. A t the cell surface, GCT 72 cells express the epitopes recognized by antibodies to SSEA 3, SSEA 4, and SSEA I; staining with W6/32 i s negligible. Levels of AFP in supernatants from GCT 72 cultures are in excess of 500 KIU/ I. The tumours formed following inocula- tion of nude mice with GCT 72 cells are solid yolk-sac tu- mours, with all cells strongly positive for AFP. A third cell type (GCT 44 and GCT 46), resembles in some ways rodent parietal endoderm. These cells uniformly coexpress keratin and vimentin intermediate filaments, but not desmoplakins. The determinants recognized by MAbs to SSEA 3,4, or I are not detected on the majority of cells in these cultures. In striking contrast to the other teratoma lines, these cells can attach to untreated tissue culture plastic in serum-free me- dium and may be serially cultivated under these conditions. The tumours formed in nude mice by these 2 cell lines are yolk-sac carcinomas with endodermal sinus tumour histology. Thus, human testicular teratomas consist of epithelial cells which may be nullipotent, pluripotent or committed to ex- traembryonic endodermal differentiation. The phenotype of human embryonal carcinoma cells shares some common fea- tures with the progenitors of extraembryonic endoderm in the mouse.
Human testicular teratomas present a particularly striking illustration of the relationship between the processes of deter- mination and differentiation and expression of the malignant phenotype. These tumours consist of undifferentiated stem cells known as embryonal carcinoma cells. In addition, they may contain differentiated somatic tissues, which are usually benign, or cells resembling extraembryonic elements, such as trophoblast or yolk sac (Damjanov, 1983). In human testicular teratomas, the trophoblastic cells are usually terminally differ- entiated syncytiotrophoblasts; malignant choriocarcinoma, with proliferating cytotrophoblastic cells, occurs only rarely. In contrast, malignant yolk-sac elements occur in 40% of testicular germ-cell tumours (Talerman, 1975; Damjanov, 1983). One major limitation to the use of human teratoma cell
lines in the study of development is that the nature of the stem cells in human teratomas, and their relationship to the better differentiated somatic and extra-embryonic elements, remains poorly defined. In particular, although the existence of a com- mon stem cell, capable of giving rise to trophoblast, yolk-sac and somatic cells, has been postulated, such a cell has never been isolated in v i m . A cell with this developmental potential would correspond to a very early stem cell in the preimplan- tation embryo, if it is assumed that differentiation in human teratomas mimics normal processes, as is sometimes the case with mouse teratocarcinomas (Martin, 1980). Understanding differentiation in the human cells will require clear-cut defini- tion of the phenotype of the tumour stem cells, which are recognizable morphologically only as undifferentiated carci- noma. Distinction at the cellular level between embryonal carcinoma and yolk-sac carcinoma stem cells has long been a particular problem in the pathology of human tumours (Parkin- son and Beilby, 1977).
Although several groups have reported the isolation of cul- tured cell lines from human teratomas (Andrews et al. , 1980, 1983; Cotte er al., 1981, 1982; McIlhinney, 1983; Vozelzang et al., 1985; Sekiya et al . , 1985), the cells have not always been characterized in detail, and no consensus regarding the phenotype of embryonal carcinoma, its counterpart in normal embryology, and its differentiation capacity, has emerged. Recently, 2 groups reported the isolation of clones from the established line Tera 2, which had the ability to differentiate into neurons in vitro when treated with retinoic acid (Andrews, 1984; Thompson et al., 1984); one group also reported differ- entiation into other cell types in xenografts (Andrews et al., 1984). It is not clear whether the properties of these clones are generally representative of human embryonal carcinoma stem cells. Andrews et al. (1983) have suggested, on the basis of their cell-surface antigen expression and the presence of tro- phoblastic elements in human teratomas in vivo, that human embryonal carcinoma cells indeed do correspond to a very early, preblastocyst, pluripotent cell.
Here we report the isolation and characterization of new cell lines from human testicular teratomas. We have identified 3 distinct types of cultured stem cell, each with a characteristic pattern of cell-surface antigen expression, intermediate fila- ment and desmoplakin content, production of oncofoetal pro- teins, growth requirements in vitro, and histology of tumours formed in nude mice. The properties of the cultured cell types are consistent with their identification as embryonal carci- noma, visceral yolk-sac carcinoma, and parietal yolk-sac car- cinoma. We confirm and extend some of the previous observations regarding the phenotype of human embryonal carcinoma cells, but suggest a different interpretation of these findings. We report for the first time extensive characterization of 2 distinct human yolk-sac carcinoma cell variants in vitro,
'To whom reprint requests should be addressed.
Received: February 11, 1987 and in revised form March 27, 1987.

CELL BIOLOGY OF HUMAN EMBRYONAL AND YOLK-SAC CARCINOMAS 335
TABLE I - SPECIFICITY AND SOURCES OF IMMUNOLOGICAL REAGENTS USED IN THlS STUDY
Source Reference Antigen or epitope recognized Monoclonal antibodies
Anti-SSEA 1
Anti-SSEA 3
Anti-SSEA 4
W632
CAM5.2
Anti-vimentin MAb
Anti-desmoplakin I + 11 clone DP 1 + 2 2.15
Gal0 144GlcNAC t l , 3 Fuca
3GalNAcflI-+ 3Gal a1 +4Galfi1 -4GlcBI-Cer
NeuAca2 -3GalPI- 3GalNAcP1+ 3Gala 1 -+4Galfll+ 4Glcfll-tCer HLA-A,B,C with P2-
microglobulin Cytokeratins 8, 18
and 19 Vimentin 57kDa poly-
peptide
Desmoplakins 1 and 2 250Kd and 2 15KD
Dr. P. Goodfellow Imperial Cancer Research Fund London, UK
Dr. P. Goodfellow Imperial Cancer Research Fund London, UK
Dr. P. Andrews The Wistar Inst. Philadelphia
Serotec,
ICRF Oxford, UK
Euro-Diagnostics, Apeldoom, Holland
BCL, Lewes, UK
Solter and Knowles, 1978
Shevinsky et a l . , 1982
Kannagi et al . . 1983
Barnstable, et al., 1978
Mahn et al., 1984
Van Muijen et al . . 1985
Cowin et al., 1985
Antiserum Conjugate Source
Rabbit anti-human - alphafoetoprotein
Rabbit anti-human
Rabbit anti-mouse chorionic gonadotropin
immunoglobulins
-
FITC
Dakopatts, Copenhagen,
Dakopatts, Copenhagen,
Dakopatts, Copenhagen,
Denmark
Denmark
Denmark Sheep anti-rabbit Alkaline Sigma, Poole, UK
immunoglobulins phosphatase
and compare the properties of these committed cell lines to embryonal carcinoma.
Establishment of cell lines Biopsy specimens from primary testicular tumours, or from
para-aortic lymph-node or lung metastases, were placed in serum-free Dulbecco's modified Eagle's medium (DMEM) at 4°C immediately following removal from the patient, and were transported to the laboratory as soon as possible. The specimens were dissected with crossed scalpels. A portion was fixed for routine histology, and the remaining fragments (less than l m 3 ) were digested for 2-16 hr in complete culture medium (DMEM, 10% foetal calf serum, 2 m ~ glutamine, 1 pglml hydrocortisone, 50pg/ml streptomycin and 100 unit/ ml penicillin) containing 1 mg/ml collagenase. The tissue digest was passed through a 250-pm mesh nylon filter, then the filtrate was resuspended in fresh culture medium and placed in plastic tissue-culture flasks under a gas phase of 90% N2, 5% O2 and 5% COz, with Swiss 3T3 cells (4 X 104/cm2 culture surface area) which had previously received 200 Gy ionizing radiation. The cultures were fed fresh medium twice weekly. When they reached confluence, the cells were har- vested with trypsin:versene (0.05% trypsin and 0.02% ver- sene), and replated at a 1:4 split ratio. Once established, the cultures were routinely subcultured at a 1: lO split ratio.
MATERIAL AND METHODS
Indirect immunofluorescence microscopy The sources of the antibodies used in this study and the
references describing their characterization are shown in Table I. Cells for indirect immunofluorescence studies were grown on multi-well slides in the presence of feeder cells. When the cultures were two-thirds confluent, the slides were rinsed with phosphate-buffered saline (180 m~ NaCl, 3.4 mM KCl, 9.7 mM Na2HP04, 1.8 mM KH2P04, PBS), fixed in acetone at - 20°C for 5 min, and then air-dried. In humidified cham- bers, primary antibodies were applied for 1 hr, followed by appropriate fluorescein-conjugated second antibodies for 30 min. The slides were stained with 1 pglml Hoechst 33258 dye in PBS for 15 min to allow identification of mouse feeder cells by nuclear staining pattern (Moser et al . , 1975), and to con- firm the absence of Mycoplasma infection (Chen, 1977). The slides were mounted in Citifluor mountant, and examined on a Zeiss Standard 16 microscope using epifluorescence optics and 25X, 4 0 X or 63X objectives. Specimens were photo- graphed with Kodak TriX black and white ASA 400 film, uprated to ASA 800 in exposure and processing. Secretion of AFP and HCG into culture medium
Cultures grown under standard conditions received fresh medium when approximately 75 % confluent. Three days later, this medium was harvested, and alphafoetoprotein (AFP) and the beta subunit of human chorionic gonadotrophin (HCG)

336 PERA. BLASCO LAFITA AND MlLLS
were measured in the unconcentrated supernatant by radioim- munoassay (Kardana and Bagshawe, 1976). Medium from cultures of 3T3 feeder cells served as a control; the level of AFP was less than 10 kilo international units (KIU)/litre, and that of HCG less than 5 international units (IU)/litre, in the control supernatants.
Growth requirements in vitro To determine the feeder-cell requirement of the cultured
teratoma cell lines, near-confluent cultures were harvested and plated at either high density (1110 split ratio) or low density (<400 cells/cm2) in the presence or absence of feeder cells. Growth was assessed in high-density cultures by inspection, and in low-density cultures by counting the number of colonies containing more than 50 cells after fixing and staining the culture dishes at the end of a 14-day incubation period.
To assess the capacity of cell lines to grow in the absence of serum, near-confluent cultures were harvested with trypsin- versene, then resuspended in serum-free medium (Iscove mod- ified Dulbecco medium, supplemented with 35 pg/ml human transferrin, 5 pg/ml bovine insulin, 2.5 p g / d bovine serum albumin, 20pM ethanolamine, and 2 m ~ glutamine) containing 1 mg/ml soybean trypsin inhibitor. The cells were then washed in serum-free medium and plated out at a 1:4 split ratio in serum-free medium. Attachment and growth were assessed by inspection, and growing cultures which reached confluence were subcultured at least 3 times under serum-free conditions to confirm that cell attachment and growth were independent of serum factors.
Tumorigenicity in nude mice Healthy, growing monolayer cultures were harvested as de-
scribed above and resuspended in serum-free DMEM, then 0.2 ml of a cell suspension containing 2.5 x lo6-1.0 X lo7 cells was inoculated intramuscularly into the gastrocnemius muscle of 4-6 week old nu/nu male athymic mice (BALB/c strain). The animals were maintained in isolators and observed weekly for signs of tumour growth. When tumours reached a diameter of lcm, the animals were killed and the tumours removed, fixed in absolute ethanol, embedded in paraffin, sectioned, and stained with haematoxylin and eosin. In all cases, fixed tumour sections were also stained immunocyto- chemically for AFP and HCG, using rabbit antisera to human AFP and HCG, and goat anti-rabbit immunoglobulins conju- gated to alkaline phosphatase. Endogenous alkaline phospha- tase activity was destroyed by pretreatment of the slides with 15% glacial acetic acid for 5 min. Specific binding of the second antibody was visualized by using naphthol-AS-MX
TABLE I1 - ORIGIN OF CULTURED TERATOMA CELL LINES
Cell line Site of lesion Pathological diagnosis
GCT27 Testis Malignant teratoma intermediate
GCT35 Testis Malignant teratoma intermediate
GCT48 Testis Malignant teratoma undifferentiated1
GCT72 Scrotum Malignant teratoma (locally invasive undifferentiated testicular primary)
GCT44 Para-aortic lymph- Malignant node metastasis teratoma
seminoma
FIGURE 1 - Phase-contrast micrographs of cultured cell lines from human testicular teratomas. (a) GCT 27, showing morphology typical of embryonal carcinoma. (b) GCT 72, a yolk-sac carcinoma cell line with properties resembling visceral endoderm. Large, flattened cells (arrow) were commonly seen in growing colonies. (c) GCT44, a yolk- sac carcinoma cell line with some properties resembling those of pa- rietal endoderm; note the mesenchymal appearance of these cells. Scale bar = 1 0 0 ~ ~ .
undifferentiated, yolk-sac elements
GCT46 Lung metastasis Malignant teratoma phosphate-TR fast red substrate. The sections were photo- undifferentiated, graphed with a Wratten 44 filter, which made positive, red yolk-sac elements staining appear black.
CELL BIOLOGY OF H U M A N EMBRYONAL A N D YOLK-SAC CARCINOMAS 331
TABLE I11 - FEEDER-CELL AND SERUM DEPENDENCE OF CULTURED TERATOMA CELL LINES
Feeder-cell dependence Growth at hieh Colonv formation'
Serum dependence Attachment in Growth in
serum-free density with& serum-free 3T3 cells' + 3T3 -3T3 medium' medium4
GCT27 GCT35 GCT48
18.4 9.1 5 .8 1 .o 0. I < .01
- GCT72 - < .01 < .01 - GCT44 + 4 . 6 < .01 + t GCT46 + 5.1 2.4 + + ' +Serial cultivation of cells without feeder layer at a 1:lO split ratio achieved; -, no serial cultivation possible.-'Colonies formedi100 cells inoculated (%).-'+,
attachment o f >90% of cells 24 hr after subculture: -, attachment of < 10% of cells 24 hr after s~bculture.-~+Serial cultivation for at least 4 passages at a 1.3 split ratio in serum-free medium, with population doubling times > 50% of those in serum-containing medium.
TABLE IV - EXPRESSION OF CELL-SURFACE, STRUCTURAL, AND ONCOFOETAL MARKERS IN CULTURED TERATOMA CELL LINES
SSEAI ' SSEA3' SSEA4' HLA-A,B,C' Keratin' Vimetm A F P ~ HCG4 Demo-' Plakins
- - GCT27 + +++ + + + +/- +++ + + GCT35 + +++ +++ +/ - +++ + + + + + GCT48 - + + + +++ + I - +++ + ++ GCT72 +++ + + + +++ +/- + + + - + + + + + +
- - +/- +++ + + + GCT44 - GCT46 + - - +/ - + + + +++
+ - -
- - - - + -
I+++, >90% of cells positive; +, < 10% positive cells; + I - , low uniform level of staining equal to or slightly above second antibody only; -, no positive cells seen.-'+, light staining at most intercellular borders; + + , moderate staining at all intercellular borders: + + +, strong staining at all intercellular borders; -, no stainmg seen.-'.4 + , AFP, 10-50K IU/I in supernatant; or HCG, 10-50 IU/I in supernatant: + + +, AFP > 500K IU/I in supernatant: -, AFP < 10K lU/l in supernatant or HCG < 10 IU/I in supernatants. Results from 3 separate determinations.
RESULTS
Establishment of cell lines from human testicular gerrn-cell tumours
Cultures were initiated from biopsy specimens of 29 tu- mours, including 8 specimens of malignant teratoma, undiffer- entiated (embryonal carcinoma), 8 specimens of malignant teratoma, intermediate (teratocarcinoma), and 13 specimens of seminoma. Specimens consisting entirely of necrotic cells, stroma, or histologically normal tissue were excluded from analysis. The success rate for establishment of cell lines from specimens of malignant teratoma (undifferentiated) was 5 out of 8 attempts and that of specimens from malignant teratoma (intermediate), 6 out of 8. In contrast, seminoma cells could not be cultivated beyond primary culture under these condi- tions; numerous modifications to the culture system failed to support growth of seminoma cells. Thus, teratoma and semi- noma cells show entirely different growth requirements in vitro.
Details of the origin of the 6 cell lines discussed in this report are given in Table 11. Studies of the 5 remaining cell lines which we have established are still in progress, but it appears that 4 show characteristics similar to those of the embryonal carcinoma lines described below, one resembles the visceral yolk-sac-type carcinoma, and one is similar to the lines resembling parietal yolk sac. Embryonal carcinoma cell lines
Cell lines GCT 27, 35 and 48 all have certain features in common. As the predominant cell type in these cultures is the one which grew most frequently from testicular germ-cell tumours, and these lines showed the most impressive evidence for multipotential differentiation in xenograft, we have identi- fied them as embryonal carcinoma cells.
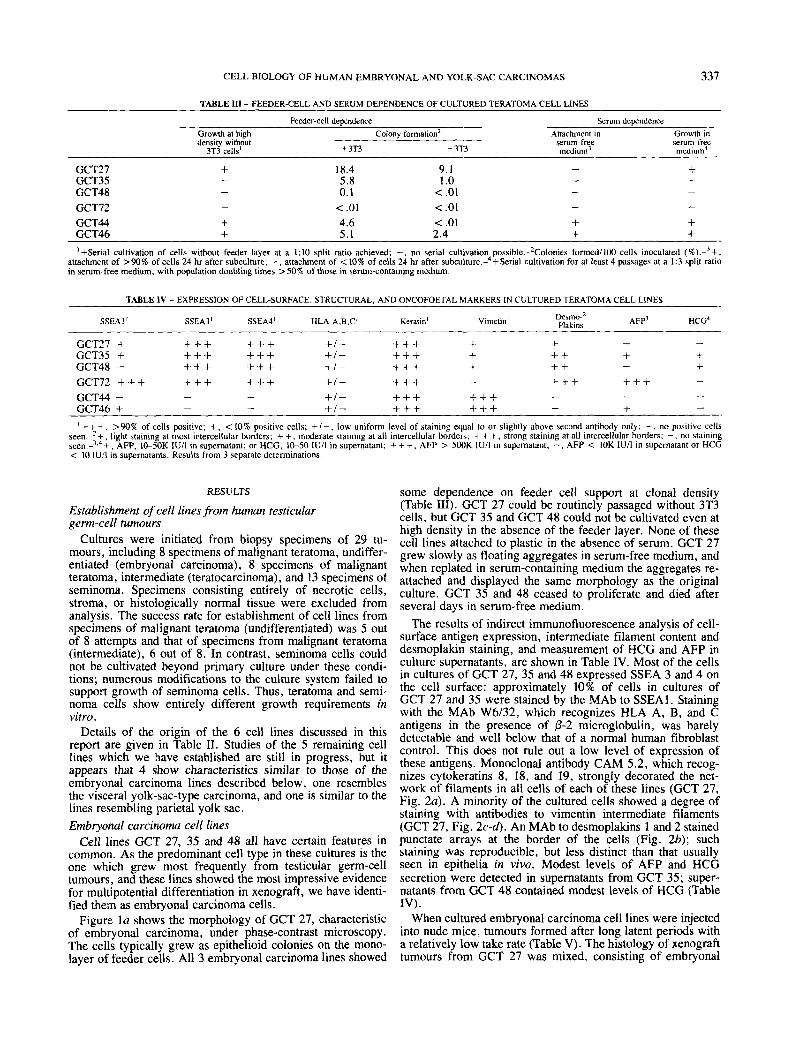
Figure la shows the morphology of GCT 27, characteristic of embryonal carcinoma, under phase-contrast microscopy. The cells typically grew as epithelioid colonies on the mono- layer of feeder cells. All 3 embryonal carcinoma lines showed
some dependence on feeder cell support at clonal density (Table 111). GCT 27 could be routinely passaged without 3T3 cells, but GCT 35 and GCT 48 could not be cultivated even at high density in the absence of the feeder layer. None of these cell lines attached to plastic in the absence of serum. GCT 27 grew slowly as floating aggregates in serum-free medium, and when replated in serum-containing medium the aggregates re- attached and displayed the same morphology as the original culture. GCT 35 and 48 ceased to proliferate and died after several days in serum-free medium.
The results of indirect immunofluorescence analysis of cell- surface antigen expression, intermediate filament content and desmoplakin staining, and measurement of HCG and AFP in culture supernatants, are shown in Table IV. Most of the cells in cultures of GCT 27, 35 and 48 expressed SSEA 3 and 4 on the cell surface: approximately 10% of cells in cultures of GCT 27 and 35 were stained by the MAb to SSEAI. Staining with the MAb W6l32, which recognizes HLA A, B, and C antigens in the presence of p-2 microglobulin, was barely detectable and well below that of a normal human fibroblast control. This does not rule out a low level of expression of these antigens. Monoclonal antibody CAM 5.2, which recog- nizes cytokeratins 8, 18, and 19, strongly decorated the net- work of filaments in all cells of each of these lines (GCT 27, Fig. 2a). A minority of the cultured cells showed a degree of staining with antibodies to vimentin intermediate filaments (GCT 27, Fig. 2 c - 4 . An MAb to desmoplakins 1 and 2 stained punctate arrays at the border of the cells (Fig. 2b); such staining was reproducible, but less distinct than that usually seen in epithelia in vivo. Modest levels of AFP and HCG secretion were detected in supernatants from GCT 35; super- natants from GCT 48 contained modest levels of HCG (Table IV) .
When cultured embryonal carcinoma cell lines were injected into nude mice, tumours formed after long latent periods with a relatively low take rate (Table V). The histology of xenograft tumours from GCT 27 was mixed. consisting of embryonal
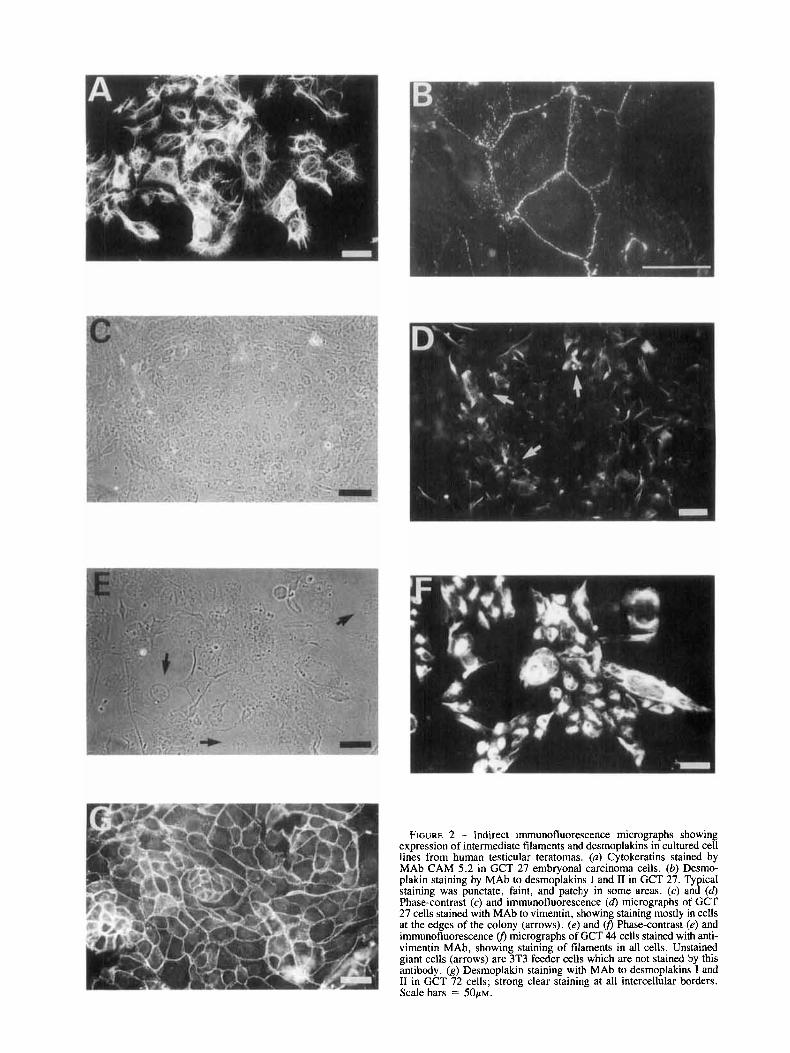
FIGURE 2 - Indirect immunofluorescence micrographs showing expression of intermediate filaments and desmoplakins in cultured cell lines from human testicular teratomas. (a) Cytokeratins stained by MA\, CAM 5.2 in GCT 27 embryonal carcinoma cells. (b) Desmo- plakin staining by MAb to desmoplakins I and I1 in GCT 27. Typical staining was punctate, faint, and patchy in some areas. (c) and (d) Phase-contrast (c) and immunofluorescence (6) micrographs of GCT 27 cells stained with MAb to vimentin, showing staining mostly in cells at the edges of the colony (arrows). (e) and U, Phase-contrast (e) and immunofluorescence U, micrographs of GCT 44 cells stained with anti- vimentin MAb, showing staining of filaments in all cells. Unstained giant cells (arrows) are 3T3 feeder cells which are not stained by this antibody. (g) Desmoplakin staining with MAb to desmoplakins I and I1 in GCT 72 cells; strong clear staining at all intercellular borders. Scale bars = 5 0 ~ ~ .

C E L L BIOLOGY OF H U M A N E M B R Y O N A L A N D YOLK-SAC CARCINOMAS 339
TABLE V - TUMOUR FORMATION FOLLOWING INOCULATION OF CULTURED TERATOMA CELLS INTO NUDE MICE
Differentiated Marker Expression % Tumours formedl Median
Inoculated’ (weeks) elements A F P I HCG’ animals latency period Histology
GCT27 30
GCT35 30
GCT48 20
GCT72 70
GCT44 60
GCT46 40
14 Malignant Glandular, teratoma, squamous and intermediate cuboidal
epithelium cartilage, bone
10 Malignant None teratoma, undifferentiated
teratoma, undifferentiated
teratoma, solid yolk sac undifferentiated
6 Malignant Yolk sac teratoma, (endodermal undifferentiated sinus tumour)
29 Malignant None
7 Malignant None
12 Malignant Yolk sac teratoma, (endodermal undifferentiated sinus tumour)
+ +
+
-
+++
+
- +
‘Data from groups of 10-20 mice. In cases of low take rates, fragments of primary tumours from injected cells were passaged in nude mice to confirm tumorigenicity, histology. and marker expression. +, Foci of cells stained positively for AFP in histological sections; + + +, all cells stained positively for AFP in histological sections; -, no cells stained positively for AFP in histological sections.-’+, Isolated multinucleate giant cells staining positively for HCG; -, no cells staining positively for HCG.
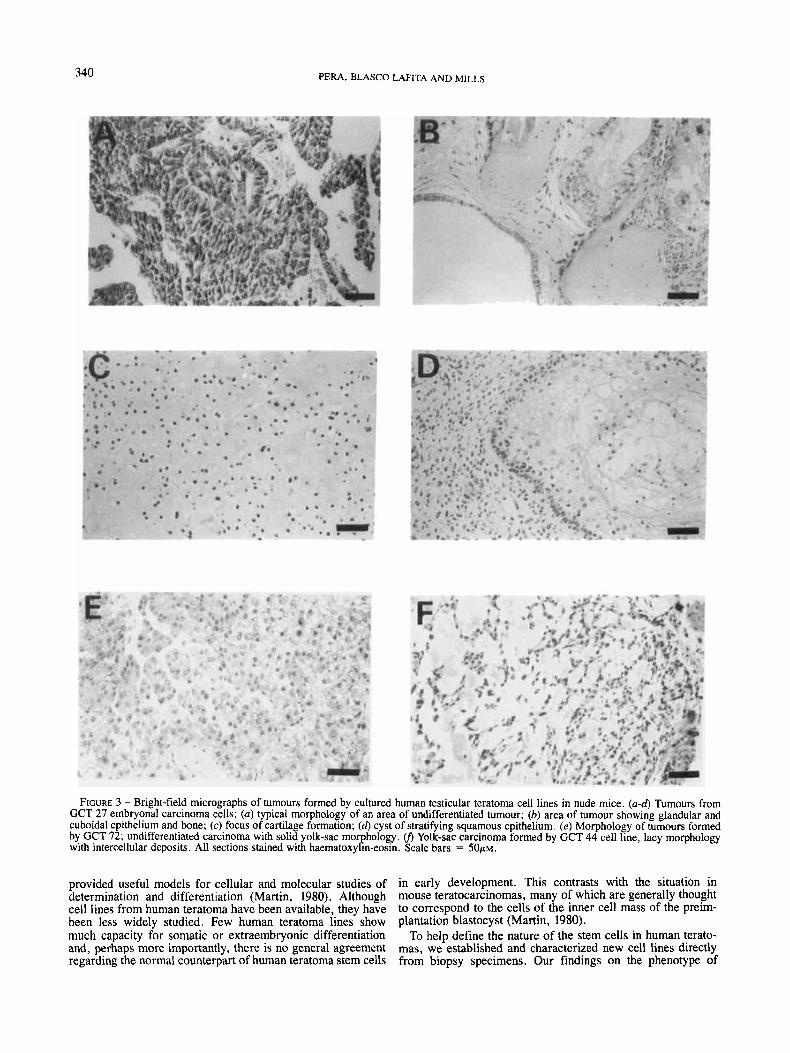
carcinoma, somatic elements including squamous, cuboidal and glandular epithelia, as well as bone and cartilage (Fig. 3a- d), foci of cells staining positively for AFP, and isolated giant multinucleated cells which were strongly positive for HCG (Fig. 4a-b). Xenografts from GCT 35 (Table V) contained embryonal carcinoma and cells staining positively for AFP, as well as giant multinucleated HCG positive cells. No somatic elements were seen. GCT 48 cells gave rise to tumours con- taining undifferentiated embryonal carcinoma only. No expres- sion of AFP or HCG was detected (Table V).
Malignant yolk-sac carcinoma cell line resembling rodent visceral endode rm
GCT 72 was established from a scrota1 biopsy of a locally invasive testicular teratoma (Fig. lb). These cells grew in epithelial colonies with a distinct morphology, clear-cut cell borders, some large flattened cells, and a vesiculated appear- ance. The cell line could not be subcultured even at high density in the absence of 3T3 cells (Table 111). In the absence of serum, GCT 72 cells did not attach to the culture surface, but remained as floating aggregates, and did not proliferate.
Immunofluorescence studies (Table IV) revealed expression of SSEA 1, SSEA 3 and SSEA 4 on the cell surface of this line. No significant staining above background with the MAb W6/32 was detected. GCT 72 cells were strongly keratin- positive, but no vimentin staining was seen. The anti-desmo- plakin antibody strongly stained intercellular junctions in cul- tures of GCT 72 (compare Fig. 2g and 2b). Levels of AFP in GCT 72 culture supernatant were in excess of 500 KIUlliter, a value at least 20-times higher than that measured in any other cultured cell supernatant. Levels of HCG were not above those found in control cultures of 3T3 feeder cells.
When GCT 72 cells were injected into nude mice, tumours formed in the majority of animals within 7 weeks of inocula- tion (Table V). The tumours all displayed solid yolk-sac mor- phology (Fig. 3e), and all cells in the xenograft were stained strongly with antisera to AFP (Fig. 4c). Yolk-sac carcinoma cell line resembling rodent parietal endoderm
Two cell lines, GCT 44 and GCT 46, were established from
metastatic teratomas. Under phase-contrast microscopy, these 2 cell lines were distinguished from those discussed above by their mesenchymal appearance, which somewhat resembled that of established rodent lines such as 3T3 fibroblasts (GCT 44, Fig. lc). GCT 44 and 46 cells could be subcultivated at high density in the absence of feeder cells, but showed some feeder dependence for clonal growth (Table 111). In striking contrast to the other teratoma cell lines described above, both GCT 44 and 46 could attach, grow, and be serially cultivated in serum-free medium containing only insulin, transferrin, albumin and ethanolamine (Table 111). The cellular phenotypes under the 2 growth conditions were similar (data not shown).
No cells staining with antibodies recognizing SSEA 3 or 4 were observed in cultures of GCT 44 or 46; a minority of GCT 46 cells were SSEAI-positive but this antigen was not expressed on GCT 44 cells (Table IV). Staining with W6/32 was negligible. Like the teratoma lines discussed above, GCT 44 and 46 cells contained cytokeratin intermediate filaments, but in cultures of these 2 cell lines, the entire population also showed well-defined staining of filaments with antibodies to vimentin (Fig. 2e-j). Another feature distinguishing these 2 cell lines was the absence of any expression of desmoplakins, even in cultures of closely apposed cells (Table IV). Modest levels of AFP were sometimes detected in the culture super- natant of GCT 46, but not GCT 44 (Table IV). Secretion of HCG into the culture supernatants of these 2 lines was not detected.
Injection of GCT 44 or GCT 46 cells into nude mice pro- duced tumours in animals within 2 to 3 months (Table V). The tumours displayed endodermal sinus yolk-sac morphology, of the reticulated, lacy, or vitelline type (Fig. 3j). Sheets of solid yolk-sac similar to that formed in nude mice by GCT 72 were not seen. Occasional areas of these 2 tumours were positive for AFP, but the reaction was weaker than that seen with GCT 72, and the majority of the tumour was negative. No somatic elements, and no cells staining with antisera to HCG, were found (Table V).
DISCUSSION
In the mouse, cultured embryonal carcinoma cells have
340 P E W , BLASCO LAFITA AND MILLS
FIGURE 3 - Bright-field micrographs of tumours formed by cultured human testicular teratoma cell lines in nude mice. (a-d) Tumours from GCT 27 embryonal carcinoma cells; (a) typical morphology of an area of undifferentiated tumour; (b) area of tumour showing glandular and cuboidal epithelium and bone; (c) focus of cartilage formation; (d) cyst of stratifying squamous epithelium. (e) Morphology of tumours formed by GCT 72; undifferentiated carcinoma with solid yolk-sac morphology. v) Yolk-sac carcinoma formed by GCT 44 cell line, lacy morphology with intercellular deposits. All sections stained with haematoxylin-eosin. Scale bars = 5 0 ~ ~ .
provided useful models for cellular and molecular studies of in early development. This contrasts with the situation in determination and differentiation (Martin, 1980). Although mouse teratocarcinomas, many of which are generally thought cell lines from human teratoma have been available, they have to correspond to the cells of the inner cell mass of the preim- been less widely studied. Few human teratoma lines show plantation blastocyst (Martin, 1980). much capacity for somatic or extraembryonic differentiation To help define the nature of the stem cells in human terato- and, perhaps more importantly, there is no general agreement mas, we established and characterized new cell lines directly regarding the normal counterpart of human teratoma stem cells from biopsy specimens. Our findings on the phenotype of

CELL BIOLOGY OF HUMAN EMBRYONAL AND YOLK-SAC CARCINOMAS 341
embryonal carcinoma cells in vitro broadly confirmed the observations of Andrews et al. (1983) on several long-estab- lished lines 2102EP and Tera 2. Most human embryonal carcinoma stem cells express the stage-specific embryonic antigens SSEA 3 and SSEA 4, and not SSEA 1. Although these cell-surface glycolipid determinants are neither cell-type- nor cell-lineage-specific (Feizi, 1985), and their expression can be modulated by their environment in vivo (Andrews et al., 1985), their patterns of expression on human embryonal carcinoma are relatively consistent, and differ from those seen in the yolk-sac variants which we studied. Thus, these cell- surface antigens appear to be informative differentiation mark- ers for human teratomas, within a narrowly defined context. We also found that expression of histocompatibility antigens was difficult to detect on embryonal carcinoma cells. Our immunofluorescence results do not, of course, rule out a low level of expression of histocompatibility antigens, as shown in other human teratoma lines (Andrews et al., 1983) and in early mouse embryos (Goldbard et al., 1985) using more sensitive techniques. Our data confirmed the expression of cytokeratin intermediate filaments in embryonal carcinoma, previously demonstrated in several cell lines (Damjanov er al., 1984) and in tumours directly removed from patients (Battifora et al., 1984). We have shown further that the human lines express desmoplakins, major protein components of desmosomes. Thus, human embryonal carcinoma cells are clearly epithelial in nature.
In their differentiation capacity, our 3 embryonal carcinoma cell lines varied, showing little capacity for differentiation (GCT 48), intermediate capacity for extraembryonic differen- tiation (GCT 35), or extensive capacity for somatic and ex- traembryonic differentiation (GCT 27). Studies of the differentiation capacity of clonal derivatives of GCT 27 indi- cate that this line is multipotential in vitro; this line is a promising candidate for a human cell line capable of trophob- lastic, yolk-sac and considerable somatic differentiation.
The question of which stem cells in normal embryonic de- velopment most closely correspond to human embryonal car- cinoma must be discussed with reference to mouse embryology, since extensive immunological, molecular and structural stud- ies of early human embryos are not available. However, ultra- structural comparisons of mouse embryos with early human embryos developing in vitro have shown many similarities (Mohr and Trounson, 1982). Observations on the expression of SSEA 3 and the trophoblastic differentiation seen in the human tumours have led to the hypothesis that the stem cells must resemble very early embryonic cells (Andrews et al., 1983). However, SSEA 3 expression is not confined to the early stages of mouse embryogenesis; the antigen reappears on primitive endoderm in the implanting blastocyst and later on visceral endoderm (Fox et al., 1984). The epithelial nature of human embryonal carcinoma cells is difficult to reconcile with the above hypothesis. Cells expressing cytokeratin inter- mediate filaments and desmosomes are seen in the preimplan- tation embryo, but only in those cells destined to give rise to trophoblast and not in pluripotent cells (Jackson ef al., 1980). It is conceivable that there are major differences between humans and rodents, as yet undiscovered, in the timing of appearance and developmental fate of epithelial cells in early embryos. On the other hand, the epithelial nature of the human cells, their surface marker expression, and the appearance of malignant yolk-sac elements in about 40% of the tumours, all suggest by analogy to mouse development (Fox et al., 1984; Jackson et al., 1981; Gardner, 1984) that these cells are phe- notypically similar in some ways to the early precursors of extraembryonic endoderm in mouse embryos. Such a pheno- type is not necessarily incompatible with differentiation into trophoblastic or somatic elements, because results with several
FIGURE 4 - Immunocytochemical staining for HCG and AFP in tu- mours formed by cultured human testicular teratoma cell lines in nude mice. (a) Tumour formed by GCT 27 showing 2 giant cells staining positively for HCG (arrows). (b) Tumour formed by GCT 27 with several foci of cells staining positively for AFP (arrows). (c) Tumour formed by GCT 72 stained for AFP; all cells show strong positive re- actions. Staining was visualized by alkaline phosphatase-conjugated second antibodies using napthol AS-MX fast red TR salt substrate. The sections were counterstained with haematoxylin and photographed un- der a Wratten 44 filter which makes red (positive) areas appear dark. Scale bar = 5 0 ~ ~ .
experimental rodent systems are strongly suggestive of trans- differentiation of extraembryonic endoderm under pathologi- cal circumstances (Sobis and Vandeputte, 1982; Van Hove et al., 1985; Damjanov et al., 1985). Differentiation in human

342 PERA, BLASCO LAFITA AND MILLS
teratomas may not be a perfect reflection of normal develop- mental processes.
The cell biology of human yolk-sac carcinoma has not been extensively described. Teilum (1959) was the first to recognize the resemblance between these tumours and the portion of the rodent placenta derived from the visceral and parietal yolk sacs; his observations have been widely confirmed and the comparison extended to human yolk sac at later stages of development (Nogales-Fernandez et al., 1977). Several yolk- sac xenografts have been reported in the literature. McIlhinney (1983) described a cell line, thought to be derived from a seminoma, which produced AFP as a xenograft. Recent ex- amination of the line in this laboratory, with some of the immunological markers used here, indicates that this tumour is not a seminoma but rather appears to be a solid yolk-sac carcinoma. Vozelzang et al. (1985) described a cultured cell line which gave rise to both embryonal carcinoma and yolk- sac elements in xenograft, but did not give an extensive de- scription of the cell phenotype in vitro.
We describe here 2 types of yolk-sac carcinoma cells that could be distinguished from embryonal carcinoma and from each other by their antigen profile, their growth requirements and the histology of the tumours they produce in mice. GCT 72, which showed well-defined desmoplakin staining in vitro, secreted high levels of AFP and formed tumours of solid yolk- sac morphology, may represent a human malignant form of visceral yolk sac, a well-differentiated secretory epithelium (Hogan et al . , 1983). GCT 44 and 46, which did not show desmoplakin staining, uniformly coexpressed both keratin and vimentin intermediate filaments, and could attach and grow in serum-free medium, resemble in some respects rodent parietal endoderm, a tissue consisting of epithelial cells which are individual and motile, and produce basement membrane ma- terial (Hogan et ul., 1983; Lane et d., 1983). Detailed analysis of the secretion of extracellular matrix molecules by these cell lines will be the subject of a separate report; basement mem- brane components are produced by these lines but there are some differences from the results with mouse extraembryonic endoderm. While the tumours formed by these 2 cell lines did not resemble mouse parietal yolk-sac neoplasms, the tumours do display the type of histological pattern which Teilum (1959) originally compared to the yolk-sac-derived portion of the placenta of rodents, the endodermal sinuses of Duval, formed by the incorporation of the junction between the visceral and parietal yolk sacs into the placenta. These cell lines also
secrete factors which stimulate attachment and colony forma- tion in vitro of embryonal carcinoma. It is likely that the yolk- sac elements are among the more aggressively malignant com- ponents of human teratomas (Parkinson and Beilby, 1977), and further study of growth control in these cells will be of inter- est. We have recently isolated a cell line from a primary testicular tumour with properties similar to those of GCT 44 and 46, both of which were derived from metastatic lesions. Therefore, the properties of these lines probably reflect inher- ent features of the cell type, rather than features particular to metastases.
Thus, cultured stem cells from human teratomas are epithe- lial cells which may be nullipotent, multipotent, or committed to extraembryonic endodermal differentiation. Recent studies with MAbs produced in this laboratory which recognize cyto- plasmic and nuclear components of human embryonal carci- noma are strongly suggestive of a close relationship between human embryonal carcinoma and the yolk-sac lines described here. We therefore suggest the possibility that human em- bryonal carcinoma is an epithelial tumour which may most closely resemble precursors of extraembryonic endoderm. The possibility that such cells in humans or rodents may display multipotential differentiation capacity raises interesting ques- tions regarding the nature of determination, and its reversibil- ity, in extraembryonic endodermal cell lineages.
ACKNOWLEDGEMENTS
We thank Profs. M. Peckham and A. Horwich, as well as Dr. G . Steel, for their support and encouragement. We are indebted to many consultant urologists, particularly Mr. W. Hendry (Royal Marsden Hospital, London) and Surgeon Com- mander I. Jenkins (Royal Naval Hospital, Portsmouth) and the tissue collectors Mrs. G. Jay and Mrs. J. Nicholls, for the provision of biopsy specimens. Prof. K. Bagshawe and Dr. H. Mitchell (Dept. of Medical Oncology, Charing Cross Hospital, London) kindly provided determinations of AFP and HCG. Dr. P. Andrews (Wistar Institute, Philadelphia, PA) and Drs. P. Goodfellow and C. Makin (Imperial Cancer Research Fund, London) donated antibodies used in this study. We are espe- cially grateful to Dr. S. Dilly (Dept. of Pathology, Royal Marsden Hospital) for advice on interpretation of tumour his- tology. Work in our laboratory is supported by grants from the Medical Research Council, the Cancer Research Campaign and the Bob Champion Cancer Trust.
REFERENCES
ANDREW, P.W., Retinoic acid induces neuronal differentiation of a cloned human embryonal carcinoma cell line in vitro. Dev. Biol., 103, 285-293 (1984). ANDREWS, P.W., BRONSON, D.L., BENHAM, F., STRICKLAND, S., and KNOWLES, B.B., A comparative study of eight cell lines derived from human testicular teratocarcinoma. Znt. J. Cancer, 26, 269-280 (1980). ANDREWS, P.W., DAMJANOV, I., SIMON, D.. BANTING, G.S., CARLIN, c . , DRACOPOLI, N.C., and FOGH, J . , Pluripotent embryonal carcinoma clones derived from the human teratocarcinoma cell line Tera-2. Lab. Invest.,
ANDREWS, P.W., DAMJANOV, I., SIMON, D. and DIGNAZIO, M., A pluri- potent human stem-cell clone isolated from the TERA-2 teratocarcinoma line lacks antigens SSEA-3 and SSEA-4 in vitro, but expresses these antigens when grown as a xenograft tumor. Differentiation, 29, 127-135 (1985). ANDREWS, P. W., GOODFELLQW, P.N., and DAMJANOV, I., Human terato- carcinoma cells in culture. Cancer Sum., 2 , 41-73 (1983). BARNSTABLE, C.J., BODMER, W.F., BROWN, G., GALFRE, G., MILSTEIN, C., WILLIAMS, A.F., and ZIEGLER, A,, Production of monoclonal antibod- ies to group A erythrocytes, HLA and other human cell surface antigens: new tools for genetic analysis. Cell, 14, 9-20 (1978). BATTIFORA, H., SHEIBANI, K., TUBBS, R.R., KIPINSKI, M.I., and SUN,
SO, 147-162 (1984).
T.-T., Antikeratin antibodies in tumor diagnosis. Cancer, 54, 843-848 ( 1984). CHEN, T.R., In situ detection of Mycoplasma contamination in cell cul- tures by fluorescent Hoechst 33258 stain. Exp. Cell Rex, 104, 255-256 (1977). COTTE, C.A., EASTY, G.C., and NEVILLE, A.M., Establishment and properties of human germ cell tumors in tissue culture. Cancer Res., 41, 1422-1427 (1981). COTTE, C., RAGHAVAN, D., MCILHINNEY, R.A.J., and MONAGHAN, P., Characterization of a new human cell line derived from a xenografted embryonal carcinoma. In Virro, 18, 739-749 (1982).
COWIN, P., KAPPRELL, H.-P., and FRANKE, W. W., The complement of desmosomal plaque proteins in different cell types. J . Cell Biol., 101, 1442-1444 (1985). DAMJANOV, I., The pathology of human teratomas. In: I. Damjanov, B. Knowles and D. Solter (eds.), The human feratomas, pp. 23-66, Humana Press, Clifton, NJ (1983). DAMJANOV, I., CLARK, R.K., and ANDREWS, P.W., Cytoskeleton of hu- man embryonal carcinoma cells. Cel Difl., 15, 133-139 (1984).
carcinoma: mouse teratocarcinoma-derived tumour stem cells differentiat- DAMJANOV, I., DAMIANOV, A., and ANDREWS, P.W., Trophectodermal

CELL BIOLOGY OF HUMAN EMBRYONAL AND YOLK-SAC CARCINOMAS 343
ing into trophoblastic and yolk sac elements. I. Embryo/. exp. Morph.. 86, 125-141 (1985). FEIZI, T., Demonstration by monoclonal antibodies that carbohydrate structures of glycoproteins and glycolipids are onco-developmental anti- gens. Nature (Lond.). 314, 53-57 (1985). Fox, N.W., DAMIANOV, I . , KNOWLES, B.B., and SOLTER, D., Stage- specific embryonic antigen 3 as a marker of visceral extraembryonic endoderm. Develop. Eiol., 103, 263-266 (1984). GARDNER, R.L., Origin and differentiation of extaembryonic tissues in the mouse. Int. Rev. exp. Pathol., 24,63-133 (1984). GOLDBARD, S.B., GOLLNICK, S.O., and WARNER, C.M., Synthesis of H- 2 antigens by preimplantation mouse embryos. Eiol. Reprod., 33, 30-36 (1985). HOGAN, B.L.M., BARLOW, D.P., and TILLY, R., F9 teratocarcinoma cells as a model for the differentiation of parietal and visceral endoderm in the mouse embryo. Cancer Surv., 2 , 115-140 (1983). JACKSON, B.W.. GRUND, c . , SCHMID, E., BURKI. K., FRANKE, W. W., and ILLMENSEE, K., Formation of cytoskeletal elements during mouse embryo- genesis-intermediate filaments of the cytokeratin type and desmosomes in preimplantation embryos. Differentiation, 17, 161- 179 (1980). JACKSON, B. W., GRUND, C . , WINTER, S. , FRANKE, W.W., and ILLMENSEE, K., Formation of cytoskeletal elements during mouse embryogenesis. 11. Epithelial differentiation and intermediate-sized filaments in early postim- plantation embryos. Differenriation, 20, 203-216 (1981).
P.W., KNOWLES, B.B., and SOLTER, D., Stage-specific embryonic antigens (SSEA-3 and -4) are epitopes of a unique globo-series ganglioside isolated from human teratocarcinoma cells. EMEO J . , 2, 2355-2361 (1983).
KANNAGI, R., COCHRAN, N.A., ISHIGAMI, F., HAKOMORI, s., ANDREWS,
KARDANA, A,, and BAGSHAWE, K.D., A rapid, sensitive and specific radioimmunoassay for human chorionic gonadotrophin. J. immunol. Merh., 9,297-305 (1976). LANE, E.B., HOGAN, B.L.M., KURKINEN, M., and Garrels, J.I., Co- expression of vimentin and cytokeratins in parietal endoderm cells of early mouse embryo. Nuture (Lond.). 303,701-704 (1983). MAKIN, C.A., BOBROW, L.G., and BODMER, W.F., Monoclonal antibody to cytokeratin for use in routine histopathology. J . clin. Path., 37,975-983 (1984). MARTIN, G.R., Teratocarcinomas and mammalian embryogenesis. Sci- ence. 209, 768-775 (1980). MCILHINNEY, R . A . J . . The biology of human germ cell tumours: experi- mental approaches. In: A. McLaren and C.C. Wylie (eds.), Current problems in germ cell differentiation, pp. 175- 197, Cambridge University Press, Cambridge (1983).
MOHR, L.R., and TROCNSON, A.O., Comparative ultrastructure of hatched human, mouse and bovine blastocysts. J . Reprod. Ferfil., 66, 499-504 (1982). MOSER, F.G., DORMAN. B.P., and RUDDLE, F.H., Mouse-human hetero- karyon analysis with a 33258 Hoechst-Giemsa technique. J . Cell Biol.,
NOGALES-FERNANDEZ, F., SILVERBERG, S.G. , BLOUSTEIN, P.A., MARTI- NEZ-HERNANDEZ, A., and PIERCE, G.B., Yolk sac carcinoma (endodermal sinus tumor)-ultrastructure and histogenesis of gonadal and extragonadal tumors in comparison with normal human yolk sac, Cancer, 39,1462-1474 (1977). PARKINSON, C., and BEILBY, J.O.W., Features of prognostic significance in testicular germ cell tumours. J . din. Path., 30, 113-119 (1977). SEKIYA, S., KAWATA, M., IWASAWA, H . , INABA, N., SUGITA, M., SUZUKI, N., MOTOYAMA, T., YAMAMOTO, T., and TAKAMIZAWA, H . , Characteriza- tion of human embryonal carcinoma cell lines derived from testicular germ-cell tumors. Differentiation, 29, 259-267 (1985).
Monoclonal antibody to murine embryos defines a stage specific embry-
66, 676-680 (1975).
SHEVINSKY, L.H. , KNOWLES, B.B., DAMJANOV, I., and SOLTER, D.,
onic antigen expressed on mouse embryos and human teratocarcinoma cells. Cell, 30, 697-705 (1982). SOBIS, H. , and VANDEPUTTE, M., Development of teratomas from yolk sac of genetically sterile embryos. Devel. Eiol., 92, 553-556 (1982). SOLTER, D., and KNOWLES, B.B., Monoclonal antibody defining a stage specific embryonic antigen (SSEA-1). Proc. nat. Acad. Sci. (wash.), 75,
TALERMAN, A., The incidence of yolk sac tumor (endodermal sinus tumor) elements in germ cell tumors of the testis in adults. Cancer, 36, 211-215 (1975). TEILUM, G . , Endodermal sinus tumors of the ovary and testis. Cancer,
THOMPSON, S . , STERN, P.L., WEBB, M., WALSH, F.S., ENGSTROM, W., EVANS, E.P., SHI, W.K., HOPKINS. B., and GRAHAM, C.F.. Cloned human teratoma cells differentiate into neuron-like cells and other cell types in retinoic acid. J . Cell Sci., 72 , 37-64 (1984). VAN HOVE, L., SOBIS, H . , Lu, Y.L., and VANDEPUTTE, M., Multipoten- tiality of yolk-sac carcinoma cell clones. Int. J. Cancer, 36,61-67 (1985). VAN MUIJEN, G.N.P., RUITER, D.J., and WARNAAR, S.O., Intermediate filaments in Merkel cell tumors. Hum. Path., 16, 590-595 (1985). VOZELZANG, N.J . , BRONSON, D., SAVINO, D., VESSELLA, R.L., and FRALEY, E.F.. A human embryonal-yolk sac carcinoma model system in athymic mice. Cancer, 55, 2584-2593 (1985).
5565-5569 (1978).
12, 1092-1105 (1959).
Copyright © 2022 FDOKUMEN




















