
Prenatal, Perinatal, and Postnatal Risk Factors in Obsessive–Compulsive Disorder

ARCHIVAL REPORT
Global Resting-State Functional Magnetic ResonanceImaging Analysis Identifies Frontal Cortex, Striatal,and Cerebellar Dysconnectivity in Obsessive-Compulsive Disorder
Alan Anticevic, Sien Hu, Sheng Zhang, Aleksandar Savic, Eileen Billingslea, Suzanne Wasylink,Grega Repovs, Michael W. Cole, Sarah Bednarski, John H. Krystal, Michael H. Bloch,Chiang-Shan R. Li, and Christopher PittengerBackground: Obsessive-compulsive disorder (OCD) is associated with regional hyperactivity in cortico-striatal circuits. However, thelarge-scale patterns of abnormal neural connectivity remain uncharacterized. Resting-state functional connectivity studies have shownaltered connectivity within the implicated circuitry, but they have used seed-driven approaches wherein a circuit of interest is defined apriori. This limits their ability to identify network abnormalities beyond the prevailing framework. This limitation is particularlyproblematic within the prefrontal cortex (PFC), which is large and heterogeneous and where a priori specification of seeds is thereforedifficult. A hypothesis-neutral, data-driven approach to the analysis of connectivity is vital.
Methods: We analyzed resting-state functional connectivity data collected at 3T in 27 OCD patients and 66 matched control subjectswith a recently developed data-driven global brain connectivity (GBC) method, both within the PFC and across the whole brain.
Results: We found clusters of decreased connectivity in the left lateral PFC in both whole-brain and PFC-restricted analyses. IncreasedGBC was found in the right putamen and left cerebellar cortex. Within regions of interest in the basal ganglia and thalamus, we identifiedincreased GBC in dorsal striatum and anterior thalamus, which was reduced in patients on medication. The ventral striatum/nucleusaccumbens exhibited decreased global connectivity but increased connectivity specifically with the ventral anterior cingulate cortex insubjects with OCD.
Conclusions: These findings identify previously uncharacterized PFC and basal ganglia dysconnectivity in OCD and reveal differentiallyaltered GBC in dorsal and ventral striatum. Results highlight complex disturbances in PFC networks, which could contribute to disruptedcortical-striatal-cerebellar circuits in OCD.
Key Words: Basal ganglia, functional connectivity, globalconnectivity, obsessive-compulsive disorder, prefrontal cortex,resting-state fMRI
Obsessive-compulsive disorder (OCD) is characterized byintrusive, anxiety-producing thoughts and repetitive, com-pulsive behaviors, which cause enormous distress (1,2).
Obsessive-compulsive disorder affects 2.7% of the population atsome point during their lives (3,4). Of these, up to 65% report thattheir illness produces major functional impairment (4).
OCD was among the first neuropsychiatric conditions in whichpathological activity in a defined brain circuit was identified.
From the Departments of Psychiatry (AA, SH, SZ, AS, EB, SW, SB, JHK, MHB,C-SRL, CP), Psychology (CP), and Neurobiology (C-SRL), Child StudyCenter (MHB, CP), Abraham Ribicoff Research Facilities (AA, CP), NIAAACenter for the Translational Neuroscience of Alcoholism (AA, JHK), andInterdepartmental Neuroscience Program (C-SRL, CP), Yale University;University Psychiatric Hospital Vrapce (AS), University of Zagreb,Zagreb, Croatia; Department of Psychology (GR), University of Ljubl-jana, Ljubljana, Slovenia; and Washington University (MWC), St. Louis,Missouri.
Address correspondence to Christopher Pittenger, M.D., Ph.D., YaleUniversity, Department of Psychiatry, 34 Park Street, New Haven, CT06519; E-mail: [email protected].
Authors C-SRL and CP contributed equally to this work.
Received May 16, 2013; revised Oct 21, 2013; accepted Oct 22, 2013.
0006-3223/$36.00http://dx.doi.org/10.1016/j.biopsych.2013.10.021
Positron emission tomography studies of glucose use (5–7),positron emission tomography and single photon emissioncomputed tomography studies of brain perfusion (8,9), andfunctional/structural magnetic resonance imaging (MRI) investi-gations (10–14) suggest increased cerebral metabolism in cortico-striato-thalamo-cortical (CSTC) circuits, particularly in the caudateand putamen, the anterior thalamus, the orbitofrontal cortex(OFC), and (more variably) the anterior cingulate cortex (ACC)(11,15,16). This circuitry is activated after symptom provocation inpatients (17–19); the hyperactivity is reduced in parallel withsymptomatic improvement after either pharmacotherapy orpsychotherapy (20–22).
The evolutionarily well-preserved CSTC circuitry is involved indiverse computations, including reward processing, action selec-tion, habit formation, and motor control (23–26). To a firstapproximation, it consists of parallel loops (24,27,28). This parallelorganization is complicated by heterogeneity within the striatumitself (29), by afferents from other structures (24), and by recentlyappreciated di-synaptic interactions with the cerebellum (30,31).Despite these complexities, the notion of parallel, functionallyspecialized information-processing loops remains of heuristicvalue for conceptualizing circuit-level disturbances.
Abnormal activity in a ventral striatum-OFC loop is wellreplicated in the OCD functional neuroimaging literature (11)and has been linked with symptom severity (32,33). Dysregulationin this “affective loop” might be associated with abnormal rewardprocessing (34). Abnormal activity in the more dorsal striatum,especially the head of the right caudate and the putamen,
BIOL PSYCHIATRY 2013;]:]]]–]]]& 2013 Society of Biological Psychiatry

2 BIOL PSYCHIATRY 2013;]:]]]–]]] A. Anticevic et al.
implicates the “cognitive loop,” communicating with the dorso-lateral prefrontal cortex (PFC) (35), although functional imaginganalyses of the dorsal PFC have been inconsistent (11).
Traditional functional magnetic resonance imaging (fMRI)studies analyze the blood oxygen level–dependent (BOLD) signalafter stimulation or during a task. However, BOLD signal fluctua-tions at rest contain information about functional networkarchitecture (36–39). Resting-state functional connectivity,defined as correlations between these spontaneous BOLD fluctu-ations, has been increasingly used to analyze neural circuitdynamics both in normal populations (28,40,41) and in patho-logical states (42). In particular, several recent studies have usedseed-based analyses of resting-state data to probe functionalconnectivity within CSTC networks in both adult and pediatricOCD (32,33,43–50). These studies have provided confirmatoryevidence of dysconnectivity within the networks previouslyidentified with baseline or provocation-related perfusion meas-ures (11,15,35). Differential patterns of dysconnectivity have beenobserved in dorsal and ventral striatal networks (32,33).
Most resting-state functional connectivity analyses in OCDhave used seed-based approaches, in which specific regions ofinterest (ROIs) are identified on the basis of a priori anatomicalconsiderations [e.g., (32,33)] or on task-based activation [e.g.,(46)], and the activity of all other brain regions is correlatedagainst those seeds. This approach is useful for testing regionallyspecific hypotheses of altered functional connectivity, but it haslimited power to detect dysconnectivity not predicted a priori.This is particularly problematic when searching for dysconnectiv-ity within large and heterogeneous regions such as basal gangliaor PFC, in which identifying appropriate seeds for connectivityanalysis is challenging. Individual differences in anatomy furthercomplicate seed definition. Independent component analysesrepresent an alternative to the a priori specification of a seedbut have not been extensively applied in OCD (50).
An alternative, data-driven approach, successfully used incomplex neuropsychiatric illness (51–53), has the potential toidentify novel patterns of brain dysconnectivity independent of apriori seeds. This permits both independent support of estab-lished hypotheses and the discovery of new patterns of con-nectivity alterations that might be missed by seed-drivenapproaches. Such a “bottom-up” approach is particularly well-suited to characterization of abnormalities in PFC connectivity.Here we have applied an approach, derived from graph theory, inwhich the global connectivity of all voxels in the brain isdetermined, relative to all other voxels, and then comparedbetween diagnostic groups (54–56). This global brain connectivity(GBC) analysis has proven powerful for detecting “hubs” ofconnectivity in healthy populations (57) and has also beensuccessfully applied to studies of PFC dysconnectivity in schizo-phrenia and bipolar disorder (51–53) as well as followingpharmacological challenge (58). A single recent study hasexamined global connectivity in OCD and identified predictedabnormalities in the OFC and the ventral striatum (59).
GBC addresses qualitatively different questions about brainconnectivity than seed-based analyses. Areas of high GBC aremaximally functionally connected with other areas and mightplay a role in coordinating large-scale patterns of brain activity(54,55). A significant between-group difference in GBC thusindicates areas and networks in which the large-scale coordina-tion of information processing might be altered in a disease state.Decreased GBC in a disease state might suggest decreasedparticipation of a particular brain region in broader networks,whereas increased GBC might suggest a pathological broadening
www.sobp.org/journal
or synchronization of functional networks. Importantly, suchalterations in connectivity need not parallel the metabolic activityof the implicated brain areas. We examined alterations in networkconnectivity in 27 OCD patients, compared with 66 matchedhealthy comparison subjects (HCS), within the PFC and across thewhole brain (to provide a fully unbiased, data-driven, and partiallyindependent search). We predicted alterations both in prefrontalnetwork connectivity and in the basal ganglia.
Methods and Materials
ParticipantsAll procedures were approved by the Yale Human Investigations
Committee. Adult OCD patients, 18–65 years of age, were recruitedthrough the Yale OCD Research Clinic through referrals andadvertising. Diagnoses were confirmed by clinical interview by apsychiatrist and the Structured Clinical Interview for DSM-IV (60). Allpatients met DSM-IV criteria for OCD. Comorbid DSM-IV depressionand other anxiety disorders were permitted, to capture a representa-tive sample. Patients with any psychotic illness, autism, history ofsubstance abuse, major head trauma or neurological disease,unstable medical illness, or pregnancy were excluded. High-qualityneuroimaging data that passed quality control criteria (Supplement 1)were acquired from 27 of 32 patients (84.6%) and are included inanalysis (Table 1 and Supplement 1). Of these patients, 13 weremedication-free for at least 8 weeks at the time of fMRI scanning;14 were treated with a selective serotonin reuptake inhibitorantidepressant at a stable dose for $ 8 weeks at the time ofscanning (Table 1). We examined the effect of medication and ofcomorbid depression in secondary analyses. The HCS, matched asa group for age and gender, were recruited through advertising;high-quality data were acquired from 66 of 80 subjects (83.5%)and included in analysis. The OCD symptom severity wasevaluated with the Yale-Brown Obsessive Compulsive Scale (Y-BOCS) (61,62); comorbid symptomatology was evaluated with theHamilton Depression Rating Scale (HAM-D, 17-item version) (63)and the Hamilton Anxiety Rating Scale (HAM-A) (64). Ratings wereperformed on the same day as the scan in most cases, and alwayswithin 3 days of the day of scanning.
Data AcquisitionImaging data were acquired on a Siemens Trio 3T scanner
(Siemens, Munich, Germany). A standard 12-channel head coilwas used, with foam padding to minimize head motion. Partic-ipants were instructed to rest with their eyes closed duringscanning but were monitored to ensure they stayed awake.Images sensitive to BOLD signal were acquired with a T2*-weighted gradient-echo planar imaging sequence sensitive toBOLD contrast (repetition time/echo time ¼ 2000/25 msec, flipangle ¼ 851, field of view ¼ 220 � 220 mm, acquisition matrix ¼64 � 64). Thirty-two axial slices (4 mm) were collected withoutgap; acquisition lasted 10 min and produced 300 volumetricimages/subject with 3.4375 � 3.4375 mm voxels. For spatialnormalization and structural segmentation, high-resolutionimages were acquired with a T1-weighted, three-dimensional fastspoiled gradient-echo sequence (repetition time/echo time ¼2530/3.66 msec, flip angle ¼ 71, field of view ¼ 256 � 256 mm,matrix ¼ 256 � 256, 176 slices without gap, voxel size ¼ 1 mm3).
Global Brain ConnectivityAll analyses followed our prior published approaches (51); see
Supplement 1 for details. Briefly, GBC (55) was computed (51) as

Table 1. Demographic Data and Clinical Characteristics of Study Groups
Characteristic
HCS(n ¼ 66)
Patients(n ¼ 27) Significance
Mean SD Mean SD t/χ2 p (2-tailed)
Age (yrs) 33.00 10.36 36.37 13.59 .58 .562Gender (% male) 62 56 1.30 .198Participants’ Education (yrs) 15.85 3.08 15.41 2.21 .67 .504Fam Hx OCD (%) — — 56Age of Onset of Minor Symptoms — — 10.52 6.23Age of Onset of Major OCD Symptoms — — 19.00 13.46Age at Dx — — 25.22 12.82Comorbid Depression (lifetime) (%) — — 70Receiving Psychotherapy (%) — — 15Receiving Stable SSRI Pharmacotherapy (%) 52HAM-D 17 — — 12.65 6.31HAM-A — — 13.60 7.85YBOCS-Total 27.15 5.74YBOCS-O — — 13.93 2.84YBOCS-C — — 13.22 3.19
SNR Ratio 231.61 75.94 237.77 84.60 .34 .73% of Frames Removed with Scrubbing 10.37 10.99 11.39 14.86 .37 .71
Age, education levels, parental education, and age at diagnosis are expressed in years; comorbid depression andcurrent therapy are expressed as percentage. Dx, diagnosis; Fam Hx, family history; HAM-A, Hamilton Anxiety RatingScale; HAM-D, Hamilton Depression Rating Scale; HCS, healthy control subjects; OCD, patients diagnosed withobsessive-compulsive disorder; SNR, signal-to-noise; SSRI, selective serotonin reuptake inhibitor; YBOCS-C, Yale-BrownObsessive Compulsive Scale-compulsions; YBOCS-O, Yale-Brown Obsessive Compulsive Scale-obsession.
A. Anticevic et al. BIOL PSYCHIATRY 2013;]:]]]–]]] 3
the average of the BOLD signal time series correlation of eachvoxel with every other gray matter voxel in the prefrontal lobes,as identified by Freesurfer automated segmentation (Figure 1)(65), or in the whole brain (Figures 2 and 3). For specificexamination of striatal and thalamic ROIs (Figure 4), a probabilisticROI mask for each region was derived on the basis of Freesurfersegmentation for each subject; average GBC across all voxels inthis ROI was extracted for each subject. Seed-based connectivityanalysis (Figure 5; Figure S1 in Supplement 1) was performed aspreviously (51).
Results
OCD Is Associated with Reduced Prefrontal ConnectivityTo test for alterations in intrinsic PFC networks in OCD, we
performed a GBC analysis restricted to PFC gray matter voxels,anatomically defined for each subject (restricted global brain
Restricted Prefrontal rGBC Differences in
right lateral surfac
Z-value0-5
left lateral surfaceaxial view
z=-15
RL
Figure 1. Reduced prefrontal cortex (PFC) restricted global brain connectivitypatients showed significantly decreased PFC-rGBC connectivity relative to healtblack border marks the approximate PFC boundary. (B) PFC rGBC reductions foall voxels in identified clusters of dysconnectivity (see Table 2 for individuahistograms of voxel distribution within regions of between-group difference a
connectivity [rGBC]) (see Supplement 1 for details). This analysis isbased on the hypothesis that there is dysconnectivity intrinsic tothe PFC in OCD but is otherwise data-driven. It revealed severalclusters of reduced PFC rGBC in OCD relative to HCS (Figure 1;Table 2). We did not find any clusters of increased PFC rGBC inOCD patients. Reduced GBC was seen bilaterally in the lateralOFC; we did not find any evidence of reduced functionalconnectivity in ACC.
OCD Is Associated with Reduced Prefrontal and IncreasedStriatal and Cerebellar Whole-Brain GBC
We next extended this approach to the entire brain, in awholly data-driven analysis. Between-group t tests revealed fiveclusters that differed significantly between OCD patients andcontrol subjects (Table 2): three clusters of decreased GBCcentered within the PFC, and clusters of increased GBC centeredin the right putamen and the left cerebellum (Figure 2). All of
PFC rGBC Connectivity
Average connectivity strength [Fz]
# of
vox
els
(acr
oss
subj
ects
)
0
5
10
15
20
-0.05 0.00 0.05 0.10
Group
OCD
HCS
Obsessive-Compulsive Disorder
eOCD vs. HCS (d = 1.77)
(rGBC) in obsessive-compulsive disorder (OCD). (A) Clusters where OCDhy comparison subjects (HCS) (see Table 2 for all cluster coordinates). Ther OCD patients (red distribution) relative to HCS (blue distribution), acrossl cluster statistics). Reported effect sizes are computed across subjects;re provided for illustrative purposes. L, left; R, right.
www.sobp.org/journal

GBC Under-Connectivity
Average connectivity strength [Fz]#
of v
oxel
s (a
cros
s su
bjec
ts)
0
20
40
60
80
100
120
-0.02 -0.01 0.00 0.01 0.02 0.03
GroupOCD
HCS
GBC Over-Connectivity
Average connectivity strength [Fz]
# of
vox
els
(acr
oss
subj
ects
)
0
5
10
15
20
-0.02 -0.01 0.00 0.01 0.02 0.03
GroupOCD
HCSy=3z=5
y = -61x=0
Whole-brain GBC Differences in Obsessive-Compulsive Disorder
left lateral surface
Z-value0-5
reduced GBCin OCD
coronal viewaxial view
sagittal view coronal view
Z-value50
puta
men
cere
bellu
m
RL RL
RL
Figure 2. Reduced prefrontal and increased striatal-cerebellar whole-brain GBC in OCD. (A) Clusters whereOCD patients showed significantly decreased whole-brain GBC connectivity relative to HCS. This patternwas centered on the left middle frontal gyrus, inferiorfrontal gyrus, and precentral gyrus (see Table 2 forcluster coordinates). (B) GBC reductions for OCD patients(red histogram) relative to HCS (blue histogram), aver-aged across all identified clusters (see Table 2 forindividual cluster statistics). (C) Clusters where OCDpatients showed significantly increased whole-brainGBC connectivity relative to HCS. This pattern wascentered on the right putamen and left cerebellum(see Table 2 for cluster coordinates). (D) GBC increasesfor OCD patients (red histogram) relative to HCS (bluehistogram) across both clusters (see Table 2 for indivi-dual cluster statistics). Blue vertical dashed line marksthe mean for the HCS group. Abbreviations as inFigure 1.
4 BIOL PSYCHIATRY 2013;]:]]]–]]] A. Anticevic et al.
these findings remained significant when we explicitly covariedfor effects of age and gender; within the OCD group, there wereno differences in GBC between subjects with and without ahistory of comorbid depression. Effect sizes were robust acrossPFC regions of reduced GBC (Cohen’s d ¼ 1.39) and across theputamen/cerebellar regions of increased GBC (Cohen’s d ¼ .99).
Overlapping Loci of Reduced Prefrontal rGBC Identified byWhole-Brain GBC and Prefrontal rGBC
To examine the convergence of whole-brain and PFC-restricted analyses, we computed a formal conjunction of allPFC voxels passing type I error correction in each analysis. Twoclusters of overlap between the analyses (left inferior and superiorfrontal gyri) emerged (Figure 3A, Table 2). The OCD subjectsshowed reduced GBC across the two clusters (Figure 3B) (Cohen’s
Conj
left lateral surf
whole-brain GBC effect
GBC effect
Figure 3. Conjunction of PFC and whole-brain GBCeffects in OCD. (A) Red clusters mark regions whereOCD patients showed significantly decreased whole-brainGBC connectivity relative to HCS (Figure 2). Yellowclusters show regions where OCD patients significantlydecreased PFC rGBC connectivity relative to HCS(Figure 1). Blue foci mark the conjunction. The PFC andwhole-brain effects are partially independent (becausethe whole-brain analysis involves many more voxels). (B)Reductions for OCD patients (red histogram) relative toHCS (blue histogram) averaged across the two clusterssurviving the conjunction (see Table 2 for individualcluster statistics). Abbreviations as in Figure 1.
www.sobp.org/journal
d ¼ 1.26). Collectively, the PFC-focused analyses and theconjunction with whole-brain effects support the hypothesis thatOCD is associated with alterations in PFC network connectivity.
The middle frontal gyrus area of reduced GBC emerged only inthe whole-brain analysis, not in the PFC-restricted analysis. Thissuggests that its connectivity and its dysregulation in OCD mightdiffer qualitatively from that of the other two PFC regions. Toassess this, we performed follow-up exploratory seed-basedconnectivity analyses on the three PFC clusters identified by thewhole-brain GBC analysis (Figure 2). The middle frontal gyruscluster showed reduced connectivity to lateral PFC but strongerconnectivity with the medial frontal cortex in OCD patients thanin HCS. This pattern was qualitatively distinct from the other PFCclusters (Figure S1 in Supplement 1), indicating a complex patternof PFC alterations in OCD.
unction of PFC and Whole-brain GBC Effects
Whole-brain GBC Connectivity
Average connectivity strength [Fz]
# of
vox
els
(acr
oss
subj
ects
)
0
10
20
30
40
-0.01 0.00 0.01 0.02 0.03
GroupOCD
HCS
ace
Z-value0-5
Putamen GBC Over-Connectivity
Average connectivity strength [Fz]
# of
vox
els
(acr
oss
subj
ects
)
0
20
40
60
80
100
120
-0.01 0.00 0.01 0.02 0.03
Thalamus GBC Over-Connectivity
Average connectivity strength [Fz]
# of
vox
els
(acr
oss
subj
ects
)
0
20
40
60
80
100
120
140
-0.01 0.00 0.01 0.02 0.03
Caudate GBC Over-Connectivity
Average connectivity strength [Fz]#
of v
oxel
s (a
cros
s su
bjec
ts)
0
20
40
60
80
-0.01 0.00 0.01 0.02 0.03
Accumbens GBC Under-Connectivity
Average connectivity strength [Fz]
# of
vox
els
(acr
oss
subj
ects
)
0
2
4
6
8
10
-0.01 0.00 0.01 0.02 0.03 GroupOCD
HCS
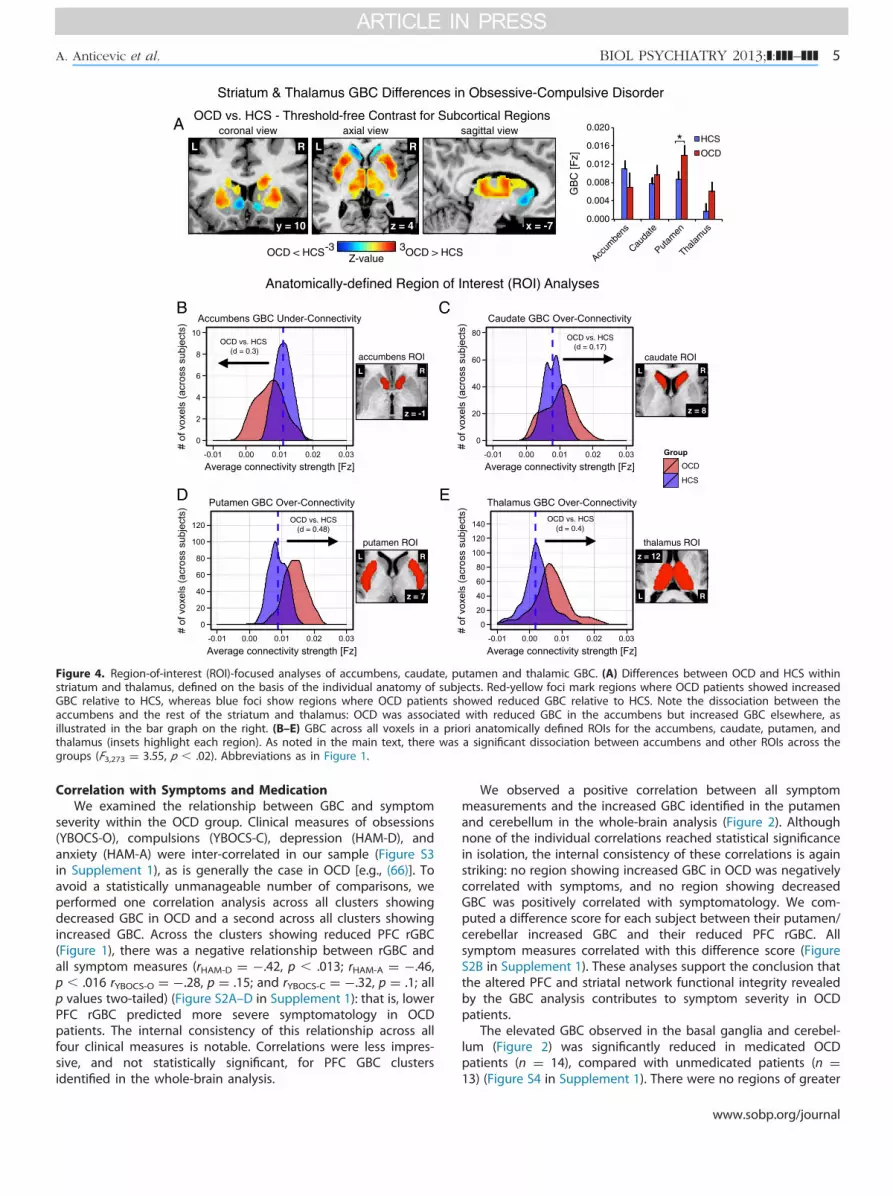
Striatum & Thalamus GBC Differences in Obsessive-Compulsive Disorder
z = 8z = -1
z = 12
z = 7
B
D
C
E
accumbens ROI caudate ROI
putamen ROI thalamus ROI
OCD vs. HCS (d = 0.48)
OCD vs. HCS (d = 0.17)
OCD vs. HCS (d = 0.3)
RL
RL
RL
RL
x = -7y = 10 z = 4
A coronal view axial view sagittal view
Z-value3-3
OCD vs. HCS - Threshold-free Contrast for Subcortical Regions
OCD < HCS OCD > HCS
RL RL
0.000
0.004
0.008
0.012
0.016
0.020
Cauda
te
Putam
en
Thalam
us
GB
C [F
z]
HCS
OCD
OCD vs. HCS (d = 0.4)
Accum
bens
Figure 4. Region-of-interest (ROI)-focused analyses of accumbens, caudate, putamen and thalamic GBC. (A) Differences between OCD and HCS withinstriatum and thalamus, defined on the basis of the individual anatomy of subjects. Red-yellow foci mark regions where OCD patients showed increasedGBC relative to HCS, whereas blue foci show regions where OCD patients showed reduced GBC relative to HCS. Note the dissociation between theaccumbens and the rest of the striatum and thalamus: OCD was associated with reduced GBC in the accumbens but increased GBC elsewhere, asillustrated in the bar graph on the right. (B–E) GBC across all voxels in a priori anatomically defined ROIs for the accumbens, caudate, putamen, andthalamus (insets highlight each region). As noted in the main text, there was a significant dissociation between accumbens and other ROIs across thegroups (F3,273 ¼ 3.55, p � .02). Abbreviations as in Figure 1.
A. Anticevic et al. BIOL PSYCHIATRY 2013;]:]]]–]]] 5
Correlation with Symptoms and MedicationWe examined the relationship between GBC and symptom
severity within the OCD group. Clinical measures of obsessions(YBOCS-O), compulsions (YBOCS-C), depression (HAM-D), andanxiety (HAM-A) were inter-correlated in our sample (Figure S3in Supplement 1), as is generally the case in OCD [e.g., (66)]. Toavoid a statistically unmanageable number of comparisons, weperformed one correlation analysis across all clusters showingdecreased GBC in OCD and a second across all clusters showingincreased GBC. Across the clusters showing reduced PFC rGBC(Figure 1), there was a negative relationship between rGBC andall symptom measures (rHAM-D ¼ �.42, p � .013; rHAM-A ¼ �.46,p � .016 rYBOCS-O ¼ �.28, p ¼ .15; and rYBOCS-C ¼ �.32, p ¼ .1; allp values two-tailed) (Figure S2A–D in Supplement 1): that is, lowerPFC rGBC predicted more severe symptomatology in OCDpatients. The internal consistency of this relationship across allfour clinical measures is notable. Correlations were less impres-sive, and not statistically significant, for PFC GBC clustersidentified in the whole-brain analysis.
We observed a positive correlation between all symptommeasurements and the increased GBC identified in the putamenand cerebellum in the whole-brain analysis (Figure 2). Althoughnone of the individual correlations reached statistical significancein isolation, the internal consistency of these correlations is againstriking: no region showing increased GBC in OCD was negativelycorrelated with symptoms, and no region showing decreasedGBC was positively correlated with symptomatology. We com-puted a difference score for each subject between their putamen/cerebellar increased GBC and their reduced PFC rGBC. Allsymptom measures correlated with this difference score (FigureS2B in Supplement 1). These analyses support the conclusion thatthe altered PFC and striatal network functional integrity revealedby the GBC analysis contributes to symptom severity in OCDpatients.
The elevated GBC observed in the basal ganglia and cerebel-lum (Figure 2) was significantly reduced in medicated OCDpatients (n ¼ 14), compared with unmedicated patients (n ¼13) (Figure S4 in Supplement 1). There were no regions of greater
www.sobp.org/journal

Over-Connectivity w/Accumb.
Average connectivity strength [Fz]
# of
vox
els
(acr
oss
subj
ects
)
0
2
4
6
8
10
-0.05 0.00 0.05 0.10 0.15
Increased Accumbens Seed Connectivity for OCD vs. HCS
x = -13 z = 19y = 28
Z-value50
sagittal view coronal view axial view
L R RL
z = -1
L
accumbens seed
R
OCD vs. HCS (d = 0.78)
GroupOCD
HCS
HAM-A Symptom Severity
Mea
n Fz
(Acc
umbe
ns-A
CC
)
-0.2
-0.1
0.0
0.1
0.2
0.3
0.4
0 5 10 15 20 25
Anxiety Symptoms &Accumbens-ACC Connectivity
r= 0.55, p<.0035
Figure 5. Seed-based anatomically defined nucleus accumbens group difference in connectivity. Prior seed-based connectivity analyses of striatalstructures in OCD have reported an increase in coupling between the nucleus accumbens and ventral anterior cingulate cortex (ACC) (32,33).We performed a similar analysis to test whether such increased seed-based connectivity coexisted with the reduced GBC seen in the accumbens(Figure 4). (A) Increased connectivity with the anatomically defined accumbens seed was seen in the ventral ACC (Talairach coordinates: x ¼ �16, y ¼ 27, z¼ 14; 864 mm3 in size; mean t statistic ¼ 3.19). (B) Increased accumbens-ACC connectivity, across all voxels in the area of significant between-groupdifference. Given a priori evidence for increased connectivity between ACC and accumbens in OCD (32), to increase power, we employed a small-volumetype I error correction for PFC voxels identical to that used in our restricted GBC approach reported in the main text. (C) Accumbens-medial PFCconnectivity positively correlated with symptom severity across all four measures reported in the main text (Hamilton Depression Rating Scale ¼ .31;Hamilton Anxiety Rating Scale [HAM-A] ¼ .55; Yale-Brown Obsessive Compulsive Scale-obsession ¼ .17; Yale-Brown Obsessive Compulsive Scale-compulsions ¼ .24), particularly prominently for HAM-A (r ¼ .55, p � .0035, surviving Bonferroni correction across four comparisons). Abbreviations as inFigure 1.
6 BIOL PSYCHIATRY 2013;]:]]]–]]] A. Anticevic et al.
GBC in medicated than unmedicated patients; there were nostatistically significant effects of medication in the PFC clusters ofhypoconnectivity (Figures 1 and 2).
Differential Dysconnectivity in Dorsal and Ventral StriatumIn a more hypothesis-guided approach, we next examined
global connectivity within anatomically defined ROIs for theaccumbens, putamen, caudate, and thalamus. Although it isrestricted to a priori defined ROIs, this analysis is conceptuallyand computationally distinct from previously reported seed-based analyses (32,33) in that it quantitatively captures theconnectivity of a ROI with the entire rest of the brain ratherthan identifying specific voxels correlated with a definedseed. Figure 4A shows the between-group GBC contrast(OCD vs. HCS) across the anatomically defined mask, at arelaxed statistical threshold to facilitate visual inspection. TheROI-specific analyses revealed a striking dissociation: theaccumbens was associated with decreased whole-brain GBCin OCD, relative to HCS (Figure 4B), but all other ROIs wereassociated with increased whole-brain GBC for OCD pati-ents relative to HCS (Figure 4C–E). A two-way analysisof variance with a between-group factor of diagnosis(OCD vs. HCS) and a within-subject factor of region con-firmed a significant diagnosis � region interaction (F3,273 ¼
www.sobp.org/journal
3.55, p � .02). This dissociation reveals a qualitatively differentpattern of network dysfunction in ventral versus dorsalstriatal loops in OCD.
Previous seed-based connectivity analyses (32,33) reportedincreased functional connectivity between a voxel in the ventralstriatum and the ventromedial and orbitofrontal PFC in subjectswith OCD; this contrasts with our finding of reduced GBC in theaccumbens. We performed a similar seed-based analysis toidentify specific regions of dysconnectivity to the accumbensbetween OCD patients and HCS, although here we explicitly usedthe entire anatomically defined accumbens region as the seedrather than arbitrarily chosen voxels within it. This analysisrevealed a cluster in the ventral ACC in which connectivity tothe accumbens was increased (Figure 5A). This increased con-nectivity (Figure 5B) (Cohen’s d ¼ .78) positively correlated withsymptom severity (HAM-D ¼ .31; HAM-A ¼ .55; YBOCS-O ¼ .17;YBOCS-C ¼ .24); the correlation was most impressive with HAM-A(p � .0035, surviving Bonferroni correction across four compar-isons) (Figure 5C).
Discussion
Corticostriatal circuits are central to reward processing, actionselection, habit formation, and motor control (23–26).

Table 2. Region Coordinates and Effect Size Estimates
X Y Z Hemisphere Anatomical LandmarkEffect Sizes (Cohen’s d)—
OCD vs. HCS Mean tCluster Size
(mm3) p
Whole-Brain GBC Findings�34 �61 �32 left Cerebellum 1.06 4.64 2268 .00001�44 41 �3 left Inferior frontal gyrus (BA 47) �1.41 5.15 5535 .0000014 �1 8 right Putamen .71 3.06 1269 .00288
�46 23 19 left Middle frontal gyrus (BA 46) �1.18 4.54 4536 .00002�36 1 37 left Precentral gyrus (BA 6) �.95 3.68 1674 .00040
Prefrontal rGBC Findings�28 15 �18 left Inferior frontal gyrus (BA 47) �1.01 4.25 729 .0000530 12 �19 right Inferior frontal gyrus (BA 47) �.90 3.96 729 .00015
�45 39 �1 left Inferior frontal gyrus (BA 47) �1.08 4.49 3267 .00002�16 61 10 left Medial frontal gyrus (BA 10) �1.10 4.87 2565 .00000�36 4 37 left Precentral gyrus (BA 6) �.99 4.00 2430 .0001310 22 63 right Superior frontal gyrus (BA 6) �.94 4.10 729 .00009
Conjunction (Whole-Brain and Prefrontal Findings)�46 40 �1 left Inferior frontal gyrus (BA 47) �1.23 4.66 2187 .00001�37 2 38 left Precentral gyrus (BA 6) �.91 3.44 1026 .00089
Effect sizes reflect standard Cohen’s d estimates for obsessive-compulsive disorder (OCD) relative to the healthy comparison subjects (HCS) group (81).Negative Cohen’s d value denotes a reduction for OCD vs. HCS.
BA, Brodmann area; rGBC, restricted global brain connectivity.
A. Anticevic et al. BIOL PSYCHIATRY 2013;]:]]]–]]] 7
Dysregulation of this circuitry contributes to numerous forms ofpsychopathology. In particular, metabolic hyperactivity of dorsaland ventral striatum and regions of the PFC has been implicatedin OCD pathophysiology (11,15,16), a view supported by recentseed-based functional connectivity analyses (32,33). Intrinsic PFCfunctional connectivity has been less examined in OCD. Weidentify specific patterns of cortical and subcortical dysconnec-tivity in OCD, with a recently developed global connectivitymetric. Results revealed dysregulated connectivity in unexpectedregions of lateral PFC, differential abnormalities in dorsaland ventral striatal circuits, and new evidence for cerebellarabnormalities.
Abnormalities in CSTC Connectivity Identified with a Data-Driven Approach
The functional heterogeneity and anatomical complexity ofthe CSTC and PFC make circuitry analysis challenging, especiallyin the context of psychopathology. Traditional resting-state fMRIprovides tools for testing functional network abnormalities butrequire a priori specification of seed ROIs. The specification ofmultiple seeds allows broader exploration but increases the riskof false positives due to multiple comparisons. Therefore, weused a data-driven GBC approach to identify disrupted connec-tivity in OCD in a hypothesis-independent way (59), circum-venting the need for a priori seed specification (54–56). Weprovide both independent support for existing models of CSTCdysregulation in OCD and important new findings, especiallywithin the PFC.
Prefrontal Hypo-Connectivity in OCDAbnormalities in PFC function are implicated in many neuro-
psychiatric conditions. The PFC is large and heterogeneous.Data-driven approaches have been particularly useful in thiscontext (51–53). We analyzed connectivity by computing GBC intwo ways: restricted to the PFC (Figure 1), and across the entirebrain (Figure 2). These analyses converged on two nodes ofdysconnectivity in the left lateral PFC, centered on thesupplementary motor area and on the ventrolateral prefrontalcortex (VLPFC). Convergence across analyses provides added
confidence in the observed connectivity alterations. Connectivityin these regions correlated negatively with symptom severity(Figure S2 in Supplement 1).
Both PFC regions have interesting implications for OCDpathophysiology. Increased somatosensory evoked potentialsand reduced sensory gating of event-related potentials werereported in the precentral gyrus in OCD (67); repetitive low-frequency supplementary motor area stimulation has beenreported to improve symptoms in treatment-resistant patients(68). The anterior VLPFC has been implicated in cognitive control.Damage to the left VLPFC has been associated with impairmentsin task switching, possibly reflecting a deficiency in top-downregulation of established procedures (69); a related deficiencymight contribute to the perseverative behaviors seen in OCD andto difficulty in controlling habitual behavior (70). The homologousregion on the right has been associated with response inhibition(69,71).
A third PFC region, centered on the left middle prefrontalgyrus, was found to be hypo-connected only in the whole-brainanalysis (Figure 3). This implies that connectivity abnormalities inthis PFC region differ qualitatively from the other two. Anexploratory between-group analysis, using each of these threeregions as a seed, confirmed this (Figure S1 in Supplement 1). Themiddle PFC region exhibited decreased connectivity with thelateral cortical surface but increased connectivity with the medialPFC. The qualitative distinction between the connectivity patternsof the three PFC clusters emphasizes the heterogeneous ways inwhich psychopathology can influence connectivity and theimportance of well-powered data-driven efforts to identify keynodes of dysregulation.
Pathology of the OFC has frequently been associated withOCD (11). We find decreased GBC in the lateral OFC in OCDpatients, only in the PFC-restricted analysis (Figure 1). Thiscontrasts with the findings of a recent connectivity analysis (59).That study applied a threshold to the connectivity measure andtherefore counts only the strongest connections, whereas ourmetric incorporates information from both strong and weakconnections. Imaging in this area is particularly challenging,because artifactual signal dropout can compromise analysis.
www.sobp.org/journal

8 BIOL PSYCHIATRY 2013;]:]]]–]]] A. Anticevic et al.
Analysis of larger samples will be critical to clarify these OFCconnectivity abnormalities.
Interestingly, dysconnectivity in these prefrontal regions cor-related most significantly with anxiety symptoms (Figure S1 inSupplement 1) rather than with Y-BOCS. PFC pathology is well-established in anxiety disorders more generally [e.g., (72)]; itmight be that lateral prefrontal hypoconnectivity contributes toanxiety symptoms in these OCD patients rather than to OCDsymptoms per se. Anxiety is prominent in many cases of OCD.Comparison with an anxious control group in a future replicationstudy would clarify whether the pattern of dysconnectivity wedescribe is specific to OCD or is a more general signature ofanxiety symptoms. This is an important area for future study.
Striatal and Cerebellar Hyperconnectivity in OCDOur whole-brain GBC analysis also revealed increased func-
tional connectivity in the right putamen and the left cerebellum(Figure 2). The right caudate and putamen have emerged as lociof pathology in many previous studies, using a variety of func-tional and structural imaging methods (11,13,15,73). The emer-gence of this striatal GBC abnormality increases confidence thatreported alterations are consistent with previous studies usingother methods.
The cerebellum has not received major attention in studies ofOCD. A recently reported morphological analysis in a pediatriccohort described reduced gray matter that correlated with ticseverity and relative enlargement that correlated with OCDseverity, suggesting a potentially important role (74). Abnormal-ities in cerebellar perfusion have also been reported (11). There isincreasing appreciation that the interconnectivity between asso-ciation neocortex, limbic cortex, and the cerebellar hemispheresmight play a key role in the processing of multimodal informa-tion. In turn, disruption of this circuitry might play an importantand underappreciated role in neuropsychiatric disease (75).
Cerebellar and striatal hyperconnectivity was modulated bymedication status. Within the OCD subjects, hyperconnectivitywas reduced in caudate, putamen, and cerebellum in the subsetwho were on stable selective serotonin reuptake inhibitormedication, relative to those who were unmedicated (Figure S4in Supplement 1). The two subgroups were indistinguishable interms of age, sex, and symptom severity. A medication effect onstriatal connectivity was also reported in a recent study ofconnectivity in OCD, although the effect was reported in theventral striatum rather than the caudate and putamen (59). Theseobservations indicate that pharmacotherapy can modulate thecircuitry that is altered in OCD. It remains possible that medicatedand unmedicated patients differ in some aspect of their under-lying disease; comparison of connectivity before and after treat-ment in a longitudinal study would be required to definitivelyclarify this question.
Global Connectivity Dissociation Within the StriatumAnalysis focused on striatal and thalamic ROIs revealed a
striking dissociation between the nucleus accumbens and thedorsal striatum for OCD patients: GBC was increased in thecaudate and putamen, as initially revealed by whole-brainanalysis (Figure 2), but was decreased in the anatomically definednucleus accumbens (Figure 4). Our results emphasize that striatalnetwork pathology in OCD is not unitary but rather differs acrossanatomically discrete information processing loops (32,33,76).
The finding of reduced GBC in the ventral striatum contrastswith the reports of increased seed-based functional connectivitywith OFC and ventromedial PFC (32,33). However, when we
www.sobp.org/journal
performed a similar seed-based analysis, we also found increasedconnectivity between the accumbens, anatomically defined ineach subject, and a region of the ventral ACC in OCD patients(Figure 5) (32,33). This co-occurrence of reduced GBC butincreased focal accumbens-cingulate functional connectivity sug-gests that nucleus accumbens might be integrated into anarrowed functional circuitry, with greater connectivity withventromedial PFC but decreased interactions with other brainregions.
The nucleus accumbens (77) and adjacent white matter (78)have been targeted for deep brain stimulation in OCD. A recentreport found that accumbens deep brain stimulation normalizedconnectivity with the medial PFC in OCD (79), further supportingthe functional importance of the network dysregulationcharacterized here.
Study LimitationsThe GBC measure seems to be quite robust in samples of
this size [e.g., (54)]; nevertheless, our single sample is alimitation, especially given the exploratory nature of ouranalysis, and replication will be critical. We have been rigorousin the elimination of images potentially compromised bymovement artifact; replication in larger samples with state-of-the-art movement correction will help solidify and refine theresults. We did not distinguish between dimensions of OCDsymptomatology (80), which might influence network dysregu-lation (33). Larger cohorts will be required to adequatelyaccount for the symptomatic heterogeneity among OCDpatients. Half of our patients were medicated, which mighthave increased heterogeneity within the OCD cohort. Connec-tivity abnormalities were associated with disease severity andcorrelated with all symptom measures but most robustly withmeasures of anxiety, rather than of OCD specifically. Identificationof abnormalities in the basal ganglia circuitry, which have notgenerally been implicated in other anxiety disorders, suggests thatthese abnormalities are indeed characteristic of OCD. Nonetheless,comparison with an anxious control group would be valuable toestablish the specificity of this effect. Lastly, these connectivityanalyses are correlational and therefore cannot establish whetherobserved abnormalities are causes or consequences of OCDsymptomatology.
ConclusionsData-driven connectivity analyses hold great promise for the
identification of dysregulated network patterns beyond thosepredicted by current theories. Our analyses reveal new patterns ofdisrupted functional connectivity in OCD. We find reducedconnectivity across three lateral PFC areas. Although disruptionin PFC was expected and motivated our analysis, these specificareas were not defined a priori and might not have emerged fromseed-driven analyses. We also observe a fascinating bidirectionaldysregulation of connectivity within the striatum; GBC isincreased for dorsal striatum and thalamus, whereas the accum-bens exhibits increased connectivity with a region of the ventralACC but decreased connectivity overall, suggesting a potentialnarrowing of the information processing networks in which itparticipates. Finally, our analysis identified increased GBC in thecerebellar cortex, which has not been a major focus of interest inOCD but merits further study.
This work was supported by DP5OD012109 (AA), 2P50AA012879(Principal Investigator: JHK; AA), The Yale Center for ClinicalInvestigation (UL1RR024139), The Brain and Behavior Research

A. Anticevic et al. BIOL PSYCHIATRY 2013;]:]]]–]]] 9
Foundation (C-SRL, AA), The Fullbright Foundation (AS),K99MH0968010 (MWC), K08MH081190 (CP), R01MH095790 (CP),The Doris Duke Charitable Foundation (CP), The Allison FamilyFoundation (CP), The Nancy Taylor Foundation for Chronic Disease(CP), and the State of Connecticut through support of the AbrahamRibicoff Research Facilities at the Connecticut Mental Health Center.
John Krystal consults for several pharmaceutical and biotechnol-ogy companies, with compensation �$10,000/year; these compa-nies include AbbVie, Amgen, AstraZeneca Pharmaceuticals, Bristol-Myers Squibb, Eli Lilly and Company, Janssen Research andDevelopment, Lundbeck Research USA, Otsuka Pharmaceutical,Development and Commercialization, Sage Therapeutics, ShirePharmaceuticals, Sunovion Pharmaceuticals, Takeda Industries,and Teva Pharmaceutical Industries. Dr. Krystal is a member ofthe following scientific advisory boards: CHDI Foundation; LohoclaResearch Corporation, Mnemosyne Pharmaceuticals; Naurex; andPfizer Pharmaceuticals. In addition, Dr. Krystal is a board member ofthe Coalition for Translational Research in Alcohol and SubstanceUse Disorders, past president of the American College of Neuro-psychopharmacology, editor of Biological Psychiatry, and anemployee of the Yale University School of Medicine and the VeteransAdministration Connecticut Health System. He is an originator onthe following patent: Seibyl JP, Krystal JH, and Charney DS;Dopamine and noradrenergic reuptake inhibitors in treatment ofschizophrenia; Patent #: 5 447 948; September 5, 1995. In addition,he is an originator of the following relevant pending patents:1) Vladimir, Coric; Krystal, John H, Sanacora, Gerard—GlutamateAgents in the Treatment of Mental Disorders No 11/399 188; April 5,2006 (Pending); and 2) Intranasal Administration of Ketamine toTreat Depression (Pending). S. Bednarski is currently an employee ofBristol Myers-Squibb, although she was not at the time of herparticipation in this work. The remaining authors report nobiomedical financial interests or potential conflicts of interest.
Supplementary material cited in this article is available online athttp://dx.doi.org/10.1016/j.biopsych.2013.10.021.
1. American Psychiatric Association (2007): Practice Guideline for theTreatment of Patients with Obsessive-Compulsive Disorder. Arlington,VA: American Psychiatric Association.
2. Jenike MA (2004): Clinical practice. Obsessive-compulsive disorder. NEngl J Med 350:259–265.
3. Kessler RC, Petukhova M, Sampson NA, Zaslavsky AM, Wittchen HU(2012): Twelve-month and lifetime prevalence and lifetime morbid riskof anxiety and mood disorders in the United States. Int J MethodsPsychiatr Res 21:169–184.
4. Ruscio AM, Stein DJ, Chiu WT, Kessler RC (2010): The epidemiology ofobsessive-compulsive disorder in the National Comorbidity SurveyReplication. Mol Psychiatry 15:53–63.
5. Baxter LR Jr, Schwartz JM, Mazziotta JC, Phelps ME, Pahl JJ, Guze BH,et al. (1988): Cerebral glucose metabolic rates in nondepressed patientswith obsessive-compulsive disorder. Am J Psychiatry 145:1560–1563.
6. Baxter LR Jr, Phelps ME, Mazziotta JC, Guze BH, Schwartz JM, Selin CE(1987): Local cerebral glucose metabolic rates in obsessive-compulsivedisorder. A comparison with rates in unipolar depression and innormal controls. Arch Gen Psychiatry 44:211–218.
7. Swedo SE, Schapiro MB, Grady CL, Cheslow DL, Leonard HL, Kumar A,et al. (1989): Cerebral glucose metabolism in childhood-onset obses-sive-compulsive disorder. Arch Gen Psychiatry 46:518–523.
8. Machlin SR, Harris GJ, Pearlson GD, Hoehn-Saric R, Jeffery P, CamargoEE (1991): Elevated medial-frontal cerebral blood flow in obsessive-compulsive patients: A SPECT study. Am J Psychiatry 148:1240–1242.
9. Rauch SL, Jenike MA, Alpert NM, Baer L, Breiter HC, Savage CR, et al.(1994): Regional cerebral blood flow measured during symptomprovocation in obsessive-compulsive disorder using oxygen 15-labeled carbon dioxide and positron emission tomography. ArchGen Psychiatry 51:62–70.
10. Whiteside SP, Port JD, Abramowitz JS (2004): A meta-analysis offunctional neuroimaging in obsessive-compulsive disorder. PsychiatryRes 132:69–79.
11. Menzies L, Chamberlain SR, Laird AR, Thelen SM, Sahakian BJ,Bullmore ET (2008): Integrating evidence from neuroimaging andneuropsychological studies of obsessive-compulsive disorder: Theorbitofronto-striatal model revisited. Neurosci Biobehav Rev 32:525–549.
12. Rotge JY, Guehl D, Dilharreguy B, Tignol J, Bioulac B, Allard M, et al.(2009): Meta-analysis of brain volume changes in obsessive-compulsive disorder. Biol Psychiatry 65:75–83.
13. Radua J, van den Heuvel OA, Surguladze S, Mataix-Cols D (2010):Meta-analytical comparison of voxel-based morphometry studies inobsessive-compulsive disorder vs other anxiety disorders. Arch GenPsychiatry 67:701–711.
14. Radua J, Mataix-Cols D (2009): Voxel-wise meta-analysis of grey matterchanges in obsessive-compulsive disorder. Br J Psychiatry 195:393–402.
15. Maia TV, Cooney RE, Peterson BS (2008): The neural bases ofobsessive-compulsive disorder in children and adults. Dev Psychopa-thol 20:1251–1283.
16. Saxena S, Bota RG, Brody AL (2001): Brain-behavior relationships inobsessive-compulsive disorder. Semin Clin Neuropsychiatry 6:82–101.
17. Breiter HC, Rauch SL, Kwong KK, Baker JR, Weisskoff RM, Kennedy DN,et al. (1996): Functional magnetic resonance imaging of symptomprovocation in obsessive-compulsive disorder. Arch Gen Psychiatry 53:595–606.
18. Adler CM, McDonough-Ryan P, Sax KW, Holland SK, Arndt S,Strakowski SM (2000): fMRI of neuronal activation with symptomprovocation in unmedicated patients with obsessive compulsivedisorder. J Psychiatr Res 34:317–324.
19. Rotge JY, Langbour N, Jaafari N, Guehl D, Bioulac B, Aouizerate B, et al.(2010): Anatomical alterations and symptom-related functional activityin obsessive-compulsive disorder are correlated in the lateral orbito-frontal cortex. Biol Psychiatry 67:e37–e38.
20. Baxter LR Jr, Schwartz JM, Bergman KS, Szuba MP, Guze BH, MazziottaJC, et al. (1992): Caudate glucose metabolic rate changes with bothdrug and behavior therapy for obsessive-compulsive disorder. ArchGen Psychiatry 49:681–689.
21. Swedo SE, Pietrini P, Leonard HL, Schapiro MB, Rettew DC, GoldbergerEL, et al. (1992): Cerebral glucose metabolism in childhood-onsetobsessive-compulsive disorder. Revisualization during pharmacother-apy. Arch Gen Psychiatry 49:690–694.
22. Saxena S, Brody AL, Ho ML, Alborzian S, Maidment KM, Zohrabi N,et al. (2002): Differential cerebral metabolic changes with paroxetinetreatment of obsessive-compulsive disorder vs major depression. ArchGen Psychiatry 59:250–261.
23. Graybiel AM (2008): Habits, rituals, and the evaluative brain. Annu RevNeurosci 31:359–387.
24. Haber SN, Knutson B (2010): The reward circuit: Linking primateanatomy and human imaging. Neuropsychopharmacology 35:4–26.
25. Grillner S, Robertson B, Stephenson-Jones M (2013): The evolutionaryorigin of the vertebrate basal ganglia and its role in action-selection[published online ahead of print February 25]. J Physiol.
26. Pennartz CM, Berke JD, Graybiel AM, Ito R, Lansink CS, van der Meer M,et al. (2009): Corticostriatal interactions during learning, memoryprocessing, and decision making. J Neurosci 29:12831–12838.
27. Alexander GE, DeLong MR, Strick PL (1986): Parallel organization offunctionally segregated circuits linking basal ganglia and cortex. AnnuRev Neurosci 9:357–381.
28. Choi EY, Yeo BT, Buckner RL (2012): The organization of the humanstriatum estimated by intrinsic functional connectivity. J Neurophysiol108:2242–2263.
29. Crittenden JR, Graybiel AM (2011): Basal Ganglia disorders associatedwith imbalances in the striatal striosome and matrix compartments.Front Neuroanat 5:59.
30. Bostan AC, Dum RP, Strick PL (2010): The basal ganglia communicatewith the cerebellum. Proc Natl Acad Sci U S A 107:8452–8456.
31. Hoshi E, Tremblay L, Feger J, Carras PL, Strick PL (2005): Thecerebellum communicates with the basal ganglia. Nat Neurosci 8:1491–1493.
32. Harrison BJ, Soriano-Mas C, Pujol J, Ortiz H, Lopez-Sola M,Hernandez-Ribas R, et al. (2009): Altered corticostriatal functional
www.sobp.org/journal

10 BIOL PSYCHIATRY 2013;]:]]]–]]] A. Anticevic et al.
connectivity in obsessive-compulsive disorder. Arch Gen Psychiatry 66:1189–1200.
33. Harrison BJ, Pujol J, Cardoner N, Deus J, Alonso P, Lopez-Sola M, et al.(2013): Brain corticostriatal systems and the major clinical symptomdimensions of obsessive-compulsive disorder. Biol Psychiatry 73:321–328.
34. Figee M, Vink M, de Geus F, Vulink N, Veltman DJ, Westenberg H, et al.(2011): Dysfunctional reward circuitry in obsessive-compulsive disor-der. Biol Psychiatry 69:867–874.
35. Rotge JY, Guehl D, Dilharreguy B, Cuny E, Tignol J, Bioulac B, et al.(2008): Provocation of obsessive-compulsive symptoms: A quantitativevoxel-based meta-analysis of functional neuroimaging studies. JPsychiatry Neurosci 33:405–412.
36. Fox MD, Raichle ME (2007): Spontaneous fluctuations in brain activityobserved with functional magnetic resonance imaging. Nat RevNeurosci 8:700–711.
37. Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, Raichle ME(2005): The human brain is intrinsically organized into dynamic,anticorrelated functional networks. Proc Natl Acad Sci U S A 102:9673–9678.
38. Biswal BB (2012): Resting state fMRI: A personal history. Neuroimage62:938–944.
39. Biswal B, Yetkin FZ, Haughton VM, Hyde JS (1995): Functionalconnectivity in the motor cortex of resting human brain using echo-planar MRI. Magn Reson Med 34:537–541.
40. Biswal BB, Mennes M, Zuo X-N, Gohel S, Kelly C, Smith SM, et al. (2010):Toward discovery science of human brain function. Proc Natl Acad SciU S A 107:4734–4739.
41. Yeo BT, Krienen FM, Sepulcre J, Sabuncu MR, Lashkari D, HollinsheadM, et al. (2011): The organization of the human cerebral cortexestimated by intrinsic functional connectivity. J Neurophysiol 106:1125–1165.
42. Fox MD, Greicius M (2010): Clinical applications of resting statefunctional connectivity. Front Syst Neurosci 4:19.
43. Fitzgerald KD, Stern ER, Angstadt M, Nicholson-Muth KC, Maynor MR,Welsh RC, et al. (2010): Altered function and connectivity of the medialfrontal cortex in pediatric obsessive-compulsive disorder. Biol Psychia-try 68:1039–1047.
44. Fitzgerald KD, Welsh RC, Stern ER, Angstadt M, Hanna GL, Abelson JL,et al. (2011): Developmental alterations of frontal-striatal-thalamicconnectivity in obsessive-compulsive disorder. J Am Acad ChildAdolesc Psychiatry 50:938–948; e933.
45. Jang JH, Kim JH, Jung WH, Choi JS, Jung MH, Lee JM, et al. (2010):Functional connectivity in fronto-subcortical circuitry during the restingstate in obsessive-compulsive disorder. Neurosci Lett 474:158–162.
46. Kang DH, Jang JH, Han JY, Kim JH, Jung WH, Choi JS, et al. (2013):Neural correlates of altered response inhibition and dysfunctionalconnectivity at rest in obsessive-compulsive disorder. Prog Neuro-psychopharmacol Biol Psychiatry 40:340–346.
47. Li P, Li S, Dong Z, Luo J, Han H, Xiong H, et al. (2012): Altered restingstate functional connectivity patterns of the anterior prefrontal cortexin obsessive-compulsive disorder. Neuroreport 23:681–686.
48. Sakai Y, Narumoto J, Nishida S, Nakamae T, Yamada K, Nishimura T,et al. (2011): Corticostriatal functional connectivity in non-medicatedpatients with obsessive-compulsive disorder. Eur Psychiatry 26:463–469.
49. Stern ER, Fitzgerald KD, Welsh RC, Abelson JL, Taylor SF (2012): Resting-state functional connectivity between fronto-parietal and default modenetworks in obsessive-compulsive disorder. PLoS One 7:e36356.
50. Zhang T, Wang J, Yang Y, Wu Q, Li B, Chen L, et al. (2011): Abnormalsmall-world architecture of top-down control networks in obsessive-compulsive disorder. J Psychiatry Neurosci 36:23–31.
51. Anticevic A, Brumbaugh MS, Winkler AM, Lombardo LE, Barrett J,Corlett PR, et al. (2013): Global prefrontal and fronto-amygdaladysconnectivity in bipolar I disorder with psychosis history. BiolPsychiatry 73:565–573.
52. Anticevic A, Repovs G, Krystal JH, Barch DM (2012): A broken filter:Prefrontal functional connectivity abnormalities in schizophreniaduring working memory interference. Schizophr Res 141:8–14.
53. Cole MW, Anticevic A, Repovs G, Barch D (2011): Variable globaldysconnectivity and individual differences in schizophrenia. BiolPsychiatry 70:43–50.
www.sobp.org/journal
54. Buckner RL, Sepulcre J, Talukdar T, Krienen FM, Liu H, Hedden T, et al.(2009): Cortical hubs revealed by intrinsic functional connectivity:Mapping, assessment of stability, and relation to Alzheimer’s disease.J Neurosci 29:1860–1873.
55. Cole MW, Pathak S, Schneider W (2010): Identifying the brain’s mostglobally connected regions. Neuroimage 49:3132–3148.
56. Martuzzi R, Ramani R, Qiu M, Shen X, Papademetris X, Constable RT(2011): A whole-brain voxel based measure of intrinsic connectivitycontrast reveals local changes in tissue connectivity with anestheticwithout a priori assumptions on thresholds or regions of interest.Neuroimage 58:1044–1050.
57. Cole MW, Yarkoni T, Repovs G, Anticevic A, Braver TS (2012): Globalconnectivity of prefrontal cortex predicts cognitive control andintelligence. J Neurosci 32:8988–8999.
58. Driesen NR, McCarthy G, Bhagwagar Z, Bloch M, Calhoun V, D’SouzaDC, et al. (2013): Relationship of resting brain hyperconnectivity andschizophrenia-like symptoms produced by the NMDA receptor antag-onist ketamine in humans. Mol Psychiatry 18:1199–1204.
59. Beucke JC, Sepulcre J, Talukdar T, Linnman C, Zschenderlein K, EndrassT, et al. (2013): Abnormally high degree connectivity of the orbito-frontal cortex in obsessive-compulsive disorder. JAMA Psychiatry 70:619–629.
60. First MB, Spitzer RL, Gibbon M, Williams JBW (2001): Structured ClinicalInterview for the DSM-IV-TR Axis I Disorders. Washington, DC: AmericanPsychiatric Press.
61. Goodman WK, Price LH, Rasmussen SA, Mazure C, Fleischmann RL, HillCL, et al. (1989): The Yale-Brown Obsessive Compulsive Scale. I.Development, use, and reliability. Arch Gen Psychiatry 46:1006–1011.
62. Goodman WK, Price LH, Rasmussen SA, Mazure C, Delgado P,Heninger GR, et al. (1989): The Yale-Brown Obsessive CompulsiveScale. II. Validity. Arch Gen Psychiatry 46:1012–1016.
63. Hamilton M (1960): A rating scale for depression. J Neurol NeurosurgPsychiatry 23:56–62.
64. Hamilton M (1959): The assessment of anxiety states by rating. Br JMed Psychol 32:50–55.
65. Fischl B, Salat DH, Busa E, Albert M, Dieterich M, Haselgrove C, et al.(2002): Whole brain segmentation: Automated labeling of neuro-anatomical structures in the human brain. Neuron 33:341–355.
66. Pittenger C, Kelmendi B, Wasylink S, Bloch MH, Coric V (2008): Riluzoleaugmentation in treatment-refractory obsessive-compulsive disorder:A series of 13 cases, with long-term follow-up. J Clin Psychopharmacol28:363–367.
67. Rossi S, Bartalini S, Ulivelli M, Mantovani A, Di Muro A, Goracci A, et al.(2005): Hypofunctioning of sensory gating mechanisms in patientswith obsessive-compulsive disorder. Biol Psychiatry 57:16–20.
68. Mantovani A, Simpson HB, Fallon BA, Rossi S, Lisanby SH (2010):Randomized sham-controlled trial of repetitive transcranial magneticstimulation in treatment-resistant obsessive-compulsive disorder. Int JNeuropsychopharmacol 13:217–227.
69. Aron AR, Monsell S, Sahakian BJ, Robbins TW (2004): A componentialanalysis of task-switching deficits associated with lesions of left andright frontal cortex. Brain 127:1561–1573.
70. Gillan CM, Papmeyer M, Morein-Zamir S, Sahakian BJ, Fineberg NA,Robbins TW, et al. (2011): Disruption in the balance between goal-directed behavior and habit learning in obsessive-compulsive disor-der. Am J Psychiatry 168:718–726.
71. Aron AR, Robbins TW, Poldrack RA (2004): Inhibition and the rightinferior frontal cortex. Trends Cogn Sci 8:170–177.
72. Ochsner KN, Ray RR, Hughes B, McRae K, Cooper JC, Weber J, et al.(2009): Bottom-up and top-down processes in emotion gene-ration: Common and distinct neural mechanisms. Psychol Sci 20:1322–1331.
73. Rotge JY, Langbour N, Guehl D, Bioulac B, Jaafari N, Allard M, et al.(2010): Gray matter alterations in obsessive-compulsive disorder: Ananatomic likelihood estimation meta-analysis. Neuropsychopharmacol-ogy 35:686–691.
74. Tobe RH, Bansal R, Xu D, Hao X, Liu J, Sanchez J, et al. (2010):Cerebellar morphology in Tourette syndrome and obsessive-compulsive disorder. Ann Neurol 67:479–487.
75. Buckner RL, Krienen FM, Castellanos A, Diaz JC, Yeo BT (2011): Theorganization of the human cerebellum estimated by intrinsic func-tional connectivity. J Neurophysiol 106:2322–2345.

A. Anticevic et al. BIOL PSYCHIATRY 2013;]:]]]–]]] 11
76. Alexander GE, Crutcher MD, DeLong MR (1990): Basal ganglia-thalamocortical circuits: Parallel substrates for motor, oculomotor,“prefrontal” and “limbic” functions. Prog Brain Res 85:119–146.
77. Denys D, Mantione M, Figee M, van den Munckhof P, Koerselman F,Westenberg H, et al. (2010): Deep brain stimulation of the nucleusaccumbens for treatment-refractory obsessive-compulsive disorder.Arch Gen Psychiatry 67:1061–1068.
78. Goodman WK, Foote KD, Greenberg BD, Ricciuti N, Bauer R, Ward H,et al. (2010): Deep brain stimulation for intractable obsessive
compulsive disorder: pilot study using a blinded, staggered-onsetdesign. Biol Psychiatry 67:535–542.
79. Figee M, Luigjes J, Smolders R, Valencia-Alfonso CE, van Wingen G, deKwaasteniet B, et al. (2013): Deep brain stimulation restores frontostriatalnetwork activity in obsessive-compulsive disorder. Nat Neurosci 16:386–387.
80. Bloch MH, Landeros-Weisenberger A, Rosario MC, Pittenger C, Leck-man JF (2008): Meta-analysis of the symptom structure of obsessive-compulsive disorder. Am J Psychiatry 165:1532–1542.
81. Cohen J (1992): A power primer. Psychol Bull 112:155–159.
www.sobp.org/journal
Copyright © 2022 FDOKUMEN




















