
Global pressures, specific responses: effects of nutrient enrichment in streams from different...
14
Global pressures, specific responses: effects of nutrient enrichment in streams from different biomes This article has been downloaded from IOPscience. Please scroll down to see the full text article. 2013 Environ. Res. Lett. 8 014002 (http://iopscience.iop.org/1748-9326/8/1/014002) Download details: IP Address: 186.141.67.198 The article was downloaded on 11/01/2013 at 08:25 Please note that terms and conditions apply. View the table of contents for this issue, or go to the journal homepage for more Home Search Collections Journals About Contact us My IOPscience
Transcript of Global pressures, specific responses: effects of nutrient enrichment in streams from different...

Global pressures specific responses effects of nutrient enrichment in streams from different
biomes
This article has been downloaded from IOPscience Please scroll down to see the full text article
2013 Environ Res Lett 8 014002
(httpiopscienceioporg1748-932681014002)
Download details
IP Address 18614167198
The article was downloaded on 11012013 at 0825
Please note that terms and conditions apply
View the table of contents for this issue or go to the journal homepage for more
Home Search Collections Journals About Contact us My IOPscience
IOP PUBLISHING ENVIRONMENTAL RESEARCH LETTERS
Environ Res Lett 8 (2013) 014002 (13pp) doi1010881748-932681014002
Global pressures specific responseseffects of nutrient enrichment in streamsfrom different biomesJoan Artigas12 Emili Garcıa-Berthou3 Delia E Bauer4 Maria I Castro5Joaquın Cochero4 Darıo C Colautti6 Agustina Cortelezzi4John C Donato5 Arturo Elosegi7 Claudia Feijoo8 Adonis Giorgi8Nora Gomez3 Leonardo Leggieri8 Isabel Munoz9Alberto Rodrigues-Capıtulo4 Anna M Romanı3 and Sergi Sabater310
1 Clermont Universite Universite Blaise Pascal Laboratoire Microorganismes Genome etEnvironnement BP 10448 F-63000 Clermont-Ferrand France2 CNRS UMR 6023 LMGE F-63177 Aubiere France3 Institute of Aquatic Ecology University of Girona E-17071 Girona Spain4 Institute of Limnology Dr R A RingueletmdashILPLA Florencio Varela Buenos Aires Argentina5 Department of Biology National University of Colombia Bogota DC Colombia6 Biotechnology Research Institute National University of San Martin Buenos Aires Argentina7 Faculty of Science and Technology The University of the Basque Country Bilbao E-48080 Spain8 INEDES and Department of Basic Sciences National University of Lujan 6700 Lujan Argentina9 Department of Ecology University of Barcelona Barcelona E-08028 Spain10 Catalan Institute for Water Research (ICRA) Scientific and Technological Park of the University ofGirona Girona E-17003 Spain
E-mail sergisabaterudgedu
Received 7 August 2012Accepted for publication 10 December 2012Published 9 January 2013Online at stacksioporgERL8014002
AbstractWe assessed the effects of nutrient enrichment on three stream ecosystems running throughdistinct biomes (Mediterranean Pampean and Andean) We increased the concentrations of Nand P in the stream water 16ndash4-fold following a beforendashafter controlndashimpact paired series(BACIPS) design in each stream and evaluated changes in the biomass of bacteria primaryproducers invertebrates and fish in the enriched (E) versus control (C) reaches after nutrientaddition through a predictive-BACIPS approach The treatment produced variable biomassresponses (2ndash77 of explained variance) among biological communities and streams Thegreatest biomass response was observed for algae in the Andean stream (77 of the variance)although fish also showed important biomass responses (about 9ndash48) The strongest biomassresponse to enrichment (77 in all biological compartments) was found in the Andean streamThe magnitude and seasonality of biomass responses to enrichment were highly site specificoften depending on the basal nutrient concentration and on windows of ecological opportunity(periods when environmental constraints other than nutrients do not limit biomass growth)The Pampean stream with high basal nutrient concentrations showed a weak response toenrichment (except for invertebrates) whereas the greater responses of Andean streamcommunities were presumably favored by wider windows of ecological opportunity incomparison to those from the Mediterranean stream Despite variation among sitesenrichment globally stimulated the algal-based food webs (algae and invertebrate grazers) butnot the detritus-based food webs (bacteria and invertebrate shredders) This study shows that
Content from this work may be used under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 30 licence Anyfurther distribution of this work must maintain attribution to the author(s) and the title of the work journal citation and DOI
11748-932613014002+13$3300 ccopy 2013 IOP Publishing Ltd Printed in the UK
Environ Res Lett 8 (2013) 014002 J Artigas et al
nutrient enrichment tends to globally enhance the biomass of stream biological assemblagesbut that its magnitude and extent within the food web are complex and are strongly determinedby environmental factors and ecosystem structure
Keywords biomes fish invertebrates microbial biomass nutrient enrichment streamecosystems windows of opportunity
S Online supplementary data available from stacksioporgERL8014002mmedia
1 Introduction
Nutrient enrichment threatens aquatic ecosystems globally(UNEP 2007) causing harmful algal blooms alteringbiological communities and affecting the functioning offreshwater and coastal marine ecosystems (Smith 2003Sutton et al 2011) The effects of enrichment are particularlystrong in streams and estuaries where both biodiversityand efficiency of ecosystem processes are severely impaired(Sobczak et al 2005 Vorosmarty et al 2010) Eutrophicationdirectly affects primary producers (algae macrophytes) andmicrobial heterotrophs (bacteria fungi) as they can usedissolved nutrients (Stelzer et al 2003) but the effects can thenspread bottom-up to other trophic levels (Hart and Robinson1990) Nutrients can stimulate decomposition (Pascoal et al2005) as well as primary production (Mulholland et al2001) two key processes with implications on the streamfood webs (Tank et al 2010) However mixed bottom-upand top-down regulations are common in streams theinteraction between nutrient availability and grazing intensityregulating the biomass of primary producers and consumers(Holomuzki et al 2010) Furthermore the response ofdifferent groups of organisms to modified trophic resourcescan be quite specific and may also depend on partitioningand compartmentalization of food webs (Huxel et al 2002Stouffer and Bascompte 2011)
The overall response of autotrophs and heterotrophs tonutrient enrichment can be higher when the backgroundnutrient concentrations are low and other factors such aslight availability temperature and hydrological stability donot limit primary production (Hill et al 2009) The occurrenceof favorable periods for primary production and invertebratedevelopment largely depends on the phenology and climatecharacteristics of the site which mark the lsquowindows ofecological opportunityrsquo (Tockner et al 2010) For instancethe growth of autotrophs in Mediterranean forested streamsis mainly limited to a narrow window of opportunity in latewinter and early spring a period before the canopy closesand that usually has minimal flood disturbance whereasheterotrophs tend to peak in late summerndashautumn (eg Artigaset al 2009) In contrast many tropical streams are lessseasonal and feature small variability in temperature andlight availability throughout the year while hydrologicalpeaks tend to control algal biomass (Zapata and Donato2008) Therefore although nutrient enrichment is becomingincreasingly widespread the response of river ecosystems canbe site specific depending on local characteristics like climateor type of dominant organisms Thus it is important to gain an
understanding of the factors governing the response of riverecosystems to nutrient enrichment under different climaticsettings This objective can be best attained by means ofcontrolled experiments
We analyzed the biological responses of streamecosystems to experimental nutrient enrichment in threebioclimatic regions (Mediterranean Pampean and Andean)The systems differed in environmental features (hydrologytemperature light availability biological communities) aswell as in the extent of their respective nutrient limitationOur main hypothesis was that the three systems wouldrespond in specific ways to a similar nutrient enrichment Wehypothesized the increase in biomass to be proportional tobasal nutrient limitation and the duration of the response tobe limited by the windows of opportunity
2 Methods
21 Characteristics of the study sites
The experimental manipulations were conducted in threeheadwater streams located in the Mediterranean (FuirososSpain) Pampean (La Choza Argentina) and tropical Andean(Tota Colombia) regions All streams were of similar orderbut had specific climatic and ecoregional characteristicsdiffering in temperature patterns background nutrient levelslight and discharge as well as in the canopy cover
Fuirosos is a Mediterranean forested stream located at analtitude of 150 m in the mountains of NE Spain It drainsa siliceous 16 km2 basin covered by pine (Pinus halepensisMill) and cork oak (Quercus suber L) forests with less than5 covered by arable fields The climate is Mediterraneanwith hot dry summers and wet winters The average annualrainfall is 600ndash800 mm and the average water temperaturesrange from 4 C (January) to 20 C (August) The experimentswere performed in a third order section (41 41primeN 2 34primeE)with a well developed deciduous riparian forest composed ofhybrid plane (Platanus acerifolia (Aiton) Willd) and blackpoplar (Populus nigra L) which cast a strong shade fromlate spring (April) to autumn (November) The stream runsover boulders cobbles and sand but the seasonal input ofleaf detritus in autumn can completely cover the streambed (Artigas et al 2009) The main primary producers aremicroscopic algae growing on rocks and in sand which tendto be most abundant in early spring before canopy closure(Tornes and Sabater 2010)
La Choza is a Pampean stream located in the lowlandprairies of the province of Buenos Aires It drains a basin
2
Environ Res Lett 8 (2013) 014002 J Artigas et al
of 48 km2 on deep loess deposits mainly covered bytemperate grassland grazed by cattle although a quarter ofthe land is devoted to agriculture The climate is temperatecontinental resulting in wet springs and autumns The averageannual rainfall is 600ndash1200 mm and the average watertemperatures range from 10 C (June) to 24 C (January) Theexperiments were conducted in a third order section (34 42primeS59 04primeO) that was naturally devoid of riparian trees Thestreambed was composed of carbonate calcium precipitatesand deposits of silt and clay that re-suspend during stormepisodes (Rodrigues Capıtulo et al 2010) Primary producersare diverse and include suspended cells epipelic microalgaeand macrophytes
Tota is an Andean stream located at an altitude of2560 m in the Eastern Mountains of Colombia in BoyacaState The stream drains a 140 km2 basin on shale and clayApproximately 40 of the surface area of the basin is usedfor agriculture and pasture the rest is covered by secondaryforest and sparse vegetation The climate is cool and dry witha bimodal precipitation pattern temperatures showing littleseasonal variation The average annual rainfall is 730 mmand average water temperatures range from 11 to 13 CExperiments were conducted in a third order section wherethe riparian vegetation is composed of open forest dominatedby Alnus acuminata Kunth and Salix humboldtiana Willdwith a dense layer of Pennisetum clandestinum Hochst exChiov grass The streambed is composed of rocks cobblesand boulders and few deposits of sand and detritus Thoughlitter fall is relatively constant through the year detritusaccumulates during periods of low flow Macrophytes are rareor absent and epilithic and epipsammic algae are the mainprimary producers
22 Experimental manipulations
The experiment followed a BACIPS (beforendashafter controlndashimpact paired series) design in each stream we experimen-tally enhanced nutrient concentrations 2ndash4 fold over a 50 mreach (E enriched) whereas an upstream reach of similarmorphological and hydrological characteristics was kept asthe control (C) All reaches (C and E) were studied for 9ndash15months before the enrichment E reaches were then enrichedand all reaches studied for one year Nutrient amounts toadd to E reaches were calculated bi-weekly considering thebasal stream water nutrient concentrations (total ammonianitrate and phosphate) and discharge Nutrient solutions wereprepared with ammonium nitrate (NH4NO3) and ammoniumphosphate (NH4H2PO4) stored in 100 l tanks beside thestream bank and added continuously to the E reach Given thelow water velocity bags of fertilizer (12 N and 12 P) wereused to produce the necessary enrichment in the E reach ofthe Pampean stream The enrichment started in January 2004in the Mediterranean January 2007 in the Pampean and May2008 in the Andean stream
Ammonium nitrate and phosphate concentrations wereanalyzed in each of the stream reaches before and afterthe enrichment Water samples taken in triplicate werefiltered through 07 microm GFF filters (Whatman England)
and placed into 50 ml flasks for further analysis in thelaboratory Colorimetric methods were used for total ammonia(Solorzano 1969) phosphate (Murphy and Riley 1962) andnitrate (APHA 1989) determination
23 Biological analyses
The sampling was adapted to study the main compartmentspresent in each of the streams fine sediment and rockswere sampled in the Mediterranean and Andean streams finesediment the water column and macrophytes in the Pampeanstream All reaches were sampled monthly On each occasionwater temperature conductivity pH oxygen irradiance anddischarge were measured with portable meters and samplesto analyze nutrient concentrations and benthic organic matteraccumulation were collected
231 Rocky habitat Bacterial and algal biomass in theepilithon was measured from 1 cm2 ceramic tiles gluedonto rock slabs that were kept at a depth of 10ndash20 cm inriffle zones The samples were allowed to colonize for atleast two months before sampling On each occasion threetiles were taken for bacterial density estimates and preservedwith 4 formaldehyde and three more tiles were taken forchlorophyll-a concentration and kept frozen until analysisBacterial density was determined following Porter and Feig(1980) and chlorophyll-a concentration determined using themethod of Jeffrey and Humphrey (1975) Transformation intomicrobial biomass (microg carbon per cmminus2) was achieved byusing conversion factors of Frost et al (2005) for chlorophyll-aand Bratbak and Dundas (1984) for bacterial densities
The taxonomic composition of the diatom communitywas determined under the microscope Diatom communitieswere detached by sonication from glass tiles cleaned withsulfuric acid and hydrogen peroxide and mounted ontopermanent slides using Naphrax (refractive index of 174Brunel Microscopes Ltd England) About 400 frustules werecounted per sample and diatoms were identified to the lowesttaxonomic level possible using standard references (Krammerand Lange-Bertalot 1986ndash1991) and recent nomenclatureupdates
Invertebrates in the Andean stream were sampled directlyon cobbles using a Surber sampler with a 900 cm2 surfacearea and a 500 microm mesh size The samples (at least 5replicates) collected were fixed with 4 formaldehyde andinvertebrates were sorted under a dissecting microscope at400times Identification of invertebrates reached the genera leveland the species level when possible Biomass was calculatedas dry mass (70 C for 48 h) per square meter In theMediterranean stream the grazer Ancylus fluviatilis Mullerwas the most abundant invertebrate upon rocks ThereforeAncylus body mass was estimated as the average dry mass(70 C 48 h) of 40ndash100 individuals (corresponding to 10ndash20cobbles sampled) in C and E reaches during enrichmentexperiments There was no rocky substratum in the Pampeanstream
3
Environ Res Lett 8 (2013) 014002 J Artigas et al
232 Sediment habitat Epipsammic algae and bacteriawere sampled with plastic cylinders (43 cm in diameter2ndash4 cm depth) used as corers Three replicates were collectedat random at C and E reaches placed in plastic containers andsubsampled using an untapped syringe (12 cm in diameter)and equivalent to 1 g of sediment Samples were theneither preserved with 4 formaldehyde for bacterial countingor kept frozen to quantify chlorophyll-a The protocolsfor bacterial density and chlorophyll-a determination weresimilar to those used for the rocks Biomass results wereexpressed in microg carbon per cmminus2 of sediment
In the Mediterranean and Andean streams invertebratesin fine sediments were collected with plastic cores (5 cmin diameter 7ndash8 cm depth) Three core replicates werecollected per reach (C and E) and stream type In the Pampeanstream an Ekman dredge sampler (100 cm2 surface area) wasused to sample sediments (also triplicate samples) Sampleswere immediately filtered through 500 microm sieves and fixedwith formaldehyde Invertebrate identification and biomassdetermination procedures were similar to those used for therock assemblages
233 Water column In the Pampean stream suspendedchlorophyll-a was analyzed from 1 l water samples (3replicates per reach and occasion) Water was filtered throughGFF filters (07 microm Whatman England) and the filterswere analyzed for chlorophyll as the extracts from rocksBacterial density was estimated in 10 ml water subsamples(3 replicates) following the methodology described in Porterand Feig (1980) Algal and bacterial biomass in thewater column was estimated through the conversion factorsdescribed before and expressed by square meter of streambed
234 Macrophytes Macrophytes were surveyed bimonthlyduring the cold season and monthly during the warm seasonat the Pampean stream Macrophyte composition and coverwere determined from regularly spaced transects Macrophytesamples (3ndash10 replicates) were obtained at random in the Cand E reaches with a 649 cm2 square metal frame and stored in500 ml plastic bags Plants were rinsed with tap water sortedinto species and invertebrates removed Plants were dried(70 C to a constant dry weight) to obtain dry mass which wasexpressed in g mminus2 Invertebrates removed from macrophyteswere filtered through 500microm sieves fixed with formaldehydeidentified and counted Identification of invertebrates reachedthe genera level and the species level when possible
235 Fish Fish abundance and biomass in the Pampean andAndean streams were estimated bimonthly using multiple-pass sampling (at least four passes per occasion) afterblocking the C and E reaches with nets Fish density wastoo low in the Mediterranean stream (Mas-Martı et al 2010)to allow comparative sampling between reaches Samplingwas performed with electrofishing (EN ISO 140112003standard) In the Pampean stream sampling was also donewith seines when high water conductivity precluded the useof electrofishing All fish sampled were counted identified
measured (standard length) and then returned to the waterOnly a small subsample of about 20 individuals wasused to determine fish condition (body weight and lengthGarcıa-Berthou and Moreno-Amich 1993) Fish abundanceand biomass were expressed as catch per unit of effort(CPUE = fish numberarea sampled) and biomass per unit ofeffort (BPUE = fish biomassarea sampled) respectively
24 Data analysis
The results of the three enrichment experiments wereanalyzed following the predictive-BACIPS approach (Benceet al 1996) This method compares the relationship betweenthe values in the C and E reaches before and after the start ofthe experiment Although beforendashafter controlndashimpact pairedseries (BACIPS) designs are often analyzed with ANOVA theuse of statistical techniques with control sites as covariatesas in the predictive-BACIPS has the advantage of allowingthe effect size (ie the enrichment effects) to vary withthe magnitude of the control value and with environmentalconditions ie does not require the standard assumption ofadditivity (Bence et al 1996 Stewart-Oaten and Bence 2001)
For the predictive-BACIPS approach we used analysis ofcovariance (ANCOVA) of the values of the response variable(eg bacterial biomass) in the E reaches using values inC (control) as a covariate and the periods beforeafter thestart of the treatment (BA) as a binary factor ANCOVA wasfirst used to test interactions between the covariate (ie Cvalues) and the binary factor (BA) Significant interactionsindicate that slopes between both variables (correspondingto C and E values) were not equal before and after thestart of the treatment (Garcıa-Berthou and Moreno-Amich1993) In that case the treatment effect would depend onthe magnitude of the control values When the C times BAinteraction was not significant (P gt 010) it was removedfrom the model to improve the statistical power and astandard ANCOVA design was used (ie homogeneous slopeswere assumed Garcıa-Berthou and Moreno-Amich 1993)The proportion of variance (often termed eta squared η2)explained by the BA factor plus the C times BA interactionwas computed as a measure of the effect size of enrichmentBence et al (1996) recommend that emphasis in BACIPSdesigns is mainly placed on effect sizes rather than on Pvalues Average η2 values in the biological compartments(bacteria algae macrophytes water column invertebratesand fish) in each stream were calculated as an estimateof the magnitude of transferral of the nutrient enrichmentthrough the food web All data were analyzed with the Rsoftware (R Development Core Team 2012) Note that thedefault packages in R use sequential (ie type I) sum ofsquares (Hector et al 2010) and that the lsquocontrolrsquo covariatewas always tested first in the models Quantitative variables(response variables and covariates) were log10-transformedbefore statistical analyses in order to improve assumptions ofthe linear models (normality homoscedasticity and linearity)
An important assumption of BACIPS designs andregression of time-related variables in general is the lackof serial correlation (Bence et al 1996 Stewart-Oaten and
4
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 1 Environmental characteristics and organic matter availability in the Mediterranean Pampean and Andean streams Values ofwater temperature (C) light (micromol photons mminus2 sminus1) and discharge (l sminus1) during enrichment experiments are represented Windows ofecological opportunity (marked in gray right plot) were determined after considering favorable conditions for primary producers microbialheterotrophs and consumer development
Bence 2001) We used the BreuschndashGodfrey test to testwhether serial correlation was present in our data withserial correlation up to four order using the lsquobgtestrsquo functionof package lsquolmtestrsquo (Zeileis and Hothorn 2002) in R TheBreuschndashGodfrey test is more applicable and often morepowerful than the more widely used DurbinndashWatson testand other methods (Godfrey and Tremayne 2005) Only 1of the 23 analyses (combinations of variables and streams)showed significant serial correlation (P lt 005) Similarlyonly one case of the 23 raw variables showed serial correlation(P lt 005) in the enriched zones probably because samplingfrequency was low (about monthly) and equally spaced(Stewart-Oaten and Bence 2001) Since our time series wererelatively short (sample size lt 50) and serial correlationwas almost always non-significant we did not correct forautocorrelation as recommended by Bence (1995)
The effect of enrichment on biomass of the differenttrophic levels was expressed as the per cent ratio of biomasschange in the E reach with respect to the C reach The ratio ofbiomass change was therefore calculated as 100 (enriched minuscontrol)control This ratio was also used to calculateenrichment effects on diatom and invertebrate abundancesThe ratiorsquos coefficient of variation (CV) during the enrichmentwas useful to determine the potential fluctuations in biomassresponse caused by the treatment Windows of opportunitywere defined as time periods when no factor beyondnutrients exerts significant limitation on primary productionand consumers in the stream These periods were identifiedin the Mediterranean Pampean and Andean streams afteranalyzing the best combinations of water temperature lightand discharge
3 Results
31 Physical and chemical characteristics of the study sites
The Mediterranean and Pampean streams had large seasonalvariations in water temperature and light availability whilein contrast the seasonal variation in the Andean stream wasmuch lower (figure 1) Water flow also followed differentpatterns in the three systems Variability was high in theAndean stream (from 68 to 1800 l sminus1) and Mediterraneanstream (from 0 to 1400 l sminus1) and very low in the Pampean(from 0 to 40 l sminus1) Low flow values included up to twomonths of summer drought in the Mediterranean stream butwater remained in the channel in the Pampean because of itsvery low slope
Basal nutrient concentrations before the enrichment(table 1) were the highest in the Pampean stream (140 microg lminus1
SRP and 700 microg lminus1 DIN) characterized by low phosphoruscontent in the Mediterranean stream (14 microg lminus1 SRP430 microg lminus1 DIN) and with low DIN concentrations in theAndean stream (26 microg lminus1 SRP and 110 microg Lminus1 DIN) Theenrichment caused SRP to increasetimes2ndash4 and DIN to increasetimes16ndash28 (table 1) The enrichment caused the decrease of theNP ratios in the Mediterranean and Pampean streams but didnot greatly alter those of the Andean (table 1)
32 Windows of opportunity
Windows of ecological opportunity for primary producersmicrobial heterotrophs and consumers differed between
5
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 1 Water nutrient concentrations of soluble reactive phosphorus (SRP) and dissolved inorganic nitrogen (DIN sum ofammonia+ nitrate+ nitrite forms) in the C and E reaches of the Mediterranean Pampean and Andean streams Values are means andstandard errors of nutrients measured during the pre-enrichment (before) and enrichment (after) periods and are accompanied by NP molarratio calculations The increment (1) nutrient concentration during enrichment is also shown for each of the streams
Control reach Enriched reach 1 nutrient concentration
Mediterranean Before SRP (microg lminus1) 14plusmn 18 16plusmn 21DIN (microg lminus1) 432plusmn 438 523plusmn 491NP 68 71
After SRP (microg lminus1) 10plusmn 9 30plusmn 13 times3DIN (microg lminus1) 389plusmn 495 765plusmn 736 times2NP 84 56
Pampean Before SRP (microg lminus1) 143plusmn 17 173plusmn 31DIN (microg lminus1) 683plusmn 39 1093plusmn 228NP 11 14
After SRP (microg lminus1) 93plusmn 3 361plusmn 29 times4DIN (microg lminus1) 528plusmn 136 828plusmn 197 times16NP 13 5
Andean Before SRP (microg lminus1) 26plusmn 5 28plusmn 5DIN (microg lminus1) 55plusmn 21 78plusmn 23NP 5 6
After SRP (microg lminus1) 53plusmn 4 105plusmn 13 times2DIN (microg lminus1) 114plusmn 59 323plusmn 85 times28NP 5 7
streams in terms of duration and frequency (figure 1)Windows of opportunity in the Mediterranean stream werenarrow and limited to late springndashearly summer (moderatetemperature and light availability) and late summerndashearlyautumn periods (moderate temperatures and abundant al-lochthonous OM inputs) The greatest overlap of favor-able environmental conditions for biological developmentoccurred in the Pampean stream In this system higher lightelevated temperatures and low discharge values coincidedfrom winter to summer (SeptemberndashMarch) The absenceof riparian vegetation reduced allochthonous OM inputsin this stream but authocthonous OM was produced byalgae and macrophytes The Andean stream also had longfavorable periods of temperature light and allochthonous OMinputs only interrupted by high discharge periods betweenAprilndashMay and OctoberndashNovember The high flow periodsin this stream caused a momentary biomass decrease of allbiological groups The predictive-BACIPS approach (table 2)suggested that 10 out of 23 models had significant treatmentseffects (or interactions) and that the explained variance wasoften high (figure 6)
33 Bacterial biomass
Bacterial biomass before the enrichment ranged from 2to 16 microg C cmminus2 on rocks and fine sediment in theMediterranean and Andean streams from 5 to 700microg C cmminus2
on sediments 004ndash15 microg C cmminus2 on macrophytes and1ndash36microg C cmminus2 in the water column for the Pampean streamThe nutrient enrichment resulted in an increase in bacterialbiomass in the Mediterranean (rocks and fine sediment) andPampean (fine sediment) streams but not in the Andeanstream (figure 2 table 2) The largest increase in bacterial
biomass after enrichment (calculated as the biomass changeratio) occurred in the Mediterranean stream (65ndash80 overallincrease) In the Pampean stream bacterial biomass increasedbetween 20 and 30 in fine sediment and macrophytesbut decreased in the water column (10ndash20) Water flowconditions temperature and light availability covaried withbacterial biomass in the E reaches of the Mediterranean andPampean streams The temporal variability of the biomasschange ratio was the highest in the Mediterranean stream(coefficient of variation CV = 24) due to the episodichigher biomass between late spring and early summer in theenrichment period
34 Algal and macrophytic biomass and composition
Algal biomass before the enrichment averaged 205 plusmn31 microg C cmminus2 on rocks and 69plusmn11 microg C cmminus2 on sedimentsat the Mediterranean stream (figure 3) Algal biomassaccounted for 137 plusmn 18 microg C cmminus2 on rocky habitat in theAndean stream while reached 162 plusmn 29 microg C cmminus2 onfine sediment and 23 plusmn 3 microg C cmminus2 for suspended algaein the Pampean stream In this stream macrophytes biomassaccounted for 58plusmn 27 g C mminus2
Nutrient enrichment produced an increase in algalbiomass in the three streams (figure 3 table 2) althoughthe responses varied in magnitude and differed betweencompartments The largest increase occurred on rocks inthe Mediterranean stream (163 on average peak of 273during spring) In the Andean stream algal biomass on rocksrapidly increased (62) and remained steady thereafter Thesmallest biomass increase was registered in the Pampeanstream particularly in the water column and for macrophytesbut biomass increased up to 49 on fine sediments The
6
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 2 Predictive-BACIPS of the effects of nutrient enrichment on the biomass of biological compartments in the three streams Thevalues shown correspond to the estimated coefficients of the linear models (ANCOVAs) When the control times BA was not significant(P gt 010) it was deleted from the model see section 2 for further details ns P gt 010 a P le 010 P le 005 P le 001 P le 0001
Variable n Control BA Control times BA
MediterraneanBacteria rocks 18 0404 0068 nsBacteria fine sediment 23 0590 0078 nsAlgae rocks 19 0495 0202a nsAlgae fine sediment 23 1082 0315 minus0732Invertebrates fine sediment 7 minus1011 minus0072 1660a
PampeanBacteria fine sediment 11 0751 0510a nsBacteria suspended 11 0954 minus0018 nsBact macrophytes 8 0396 0097 nsAlgae fine sediment 10 1106 0206 nsAlgae suspended 5 0274 minus0079 nsAlgae macrophyte 10 0575 minus0150 nsMacrophytes 11 1093 0249 nsInvertebrates fine sediment 10 minus0130 0787 nsInvertebrates macrophytes 6 minus0251 minus0521a nsFish CPUE 4 1082 8221 nsFish BPUE 4 0745 29552 ns
AndeanBacteria rocks 7 0666a 0299 nsBacteria fine sediment 6 0150 minus2631a nsAlgae rocks 14 0158 minus0101 0728Invertebrates rocks 13 0469 minus0269 nsInvertebrates fine sediment 13 0432 minus0388 nsFish density 10 1012 minus0442 nsFish biomass 10 1020 minus0464a ns
algal biomass responses to nutrients in the three streamswere strongly related with favorable conditions of lighttemperature and water flow (figure 3) The difference betweenthe extension of the respective windows of opportunitydetermined the steadiness of the response The Mediterraneanand Andean streams represented the maximum and minimumvariability in favorable conditions for primary producersrsquogrowth accordingly the coefficients of variation of thebiomass ratiorsquos change were the highest in the Mediterranean(CV = 16) and the lowest in the Andean (CV = 08)stream
Nutrient enrichment affected the composition of benthicalgal assemblages Some diatom taxa decreased (egAchnanthes biasolettiana Grunow (minus57) Achnanthesminutissima Kutzing (minus432) in the MediterraneanFragilaria ulna (Nitzsch) Lange-Bertalot var ulna (minus68)in the Pampean and Cocconeis placentula Ehrenberg(minus40) in the Andean stream) whereas others increased(eg Rhoicosphenia abbreviata (Ag) Lange-Bertalot (94)and Cocconeis placentula Ehrenberg (594) in theMediterranean Nitzschia frustulum (Kutzing) Grunow varfrustulum (3528) in the Pampean and Reimeria sinuata(Gregory) Kociolek and Stoermer (646) in the Andeanstream) Enrichment in the Pampean stream had no effect onthe macrophyte composition Ludwigia sp and Bacopa spdominated both the C and E reaches and were later replacedby the macroalga Spirogyra sp
35 Invertebrate biomass and composition
Invertebrate biomass before the enrichment ranged from5 to 750 microg DM cmminus2 in the Mediterranean and Andeanstreams and reached 1093 plusmn 311 microg DM cmminus2 in thesediments of the Pampean stream The effects of nutrientenrichment were generally non-significant but often larger(figure 4 table 2) and more pronounced in some habitatsthan others In the Pampean stream effects were not apparenton invertebrates occurring on macrophytes and were minor(3) on fine sediments Invertebrate biomass increased upto 155 in the fine sediments of the Mediterranean streamwhile no increase was observed in the Andean stream Theeffects on invertebrate biomass were the least variable inthe Mediterranean stream (per cent increase ratio CV = 09CV = 12 in the Andean and 27 in the Pampean)
Although the general enrichment effects on invertebratebiomass were weak some specific invertebrate taxa weredistinctly affected The grazer Ancylus fluviatilis increasedby 17 in the Mediterranean stream and the grazerCamelobaetidus increased by 927 in the Andean streamDensities of the scrapers Heleobia parchappei Orbigny(in fine sediments) and Hyalella curvispina Shoemaker(on macrophytes) increased in the Pampean stream (48and 762 respectively) Potamopyrgus antipodarum Gray(scraper) and Corbicula fluminea Muller (filterer) occurredonly in the respective fertilized reaches of the Mediterraneanand Pampean streams The Tubificidae (minus38) and theTanytarsini chironomidae (minus29) significantly decreased in
7
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

IOP PUBLISHING ENVIRONMENTAL RESEARCH LETTERS
Environ Res Lett 8 (2013) 014002 (13pp) doi1010881748-932681014002
Global pressures specific responseseffects of nutrient enrichment in streamsfrom different biomesJoan Artigas12 Emili Garcıa-Berthou3 Delia E Bauer4 Maria I Castro5Joaquın Cochero4 Darıo C Colautti6 Agustina Cortelezzi4John C Donato5 Arturo Elosegi7 Claudia Feijoo8 Adonis Giorgi8Nora Gomez3 Leonardo Leggieri8 Isabel Munoz9Alberto Rodrigues-Capıtulo4 Anna M Romanı3 and Sergi Sabater310
1 Clermont Universite Universite Blaise Pascal Laboratoire Microorganismes Genome etEnvironnement BP 10448 F-63000 Clermont-Ferrand France2 CNRS UMR 6023 LMGE F-63177 Aubiere France3 Institute of Aquatic Ecology University of Girona E-17071 Girona Spain4 Institute of Limnology Dr R A RingueletmdashILPLA Florencio Varela Buenos Aires Argentina5 Department of Biology National University of Colombia Bogota DC Colombia6 Biotechnology Research Institute National University of San Martin Buenos Aires Argentina7 Faculty of Science and Technology The University of the Basque Country Bilbao E-48080 Spain8 INEDES and Department of Basic Sciences National University of Lujan 6700 Lujan Argentina9 Department of Ecology University of Barcelona Barcelona E-08028 Spain10 Catalan Institute for Water Research (ICRA) Scientific and Technological Park of the University ofGirona Girona E-17003 Spain
E-mail sergisabaterudgedu
Received 7 August 2012Accepted for publication 10 December 2012Published 9 January 2013Online at stacksioporgERL8014002
AbstractWe assessed the effects of nutrient enrichment on three stream ecosystems running throughdistinct biomes (Mediterranean Pampean and Andean) We increased the concentrations of Nand P in the stream water 16ndash4-fold following a beforendashafter controlndashimpact paired series(BACIPS) design in each stream and evaluated changes in the biomass of bacteria primaryproducers invertebrates and fish in the enriched (E) versus control (C) reaches after nutrientaddition through a predictive-BACIPS approach The treatment produced variable biomassresponses (2ndash77 of explained variance) among biological communities and streams Thegreatest biomass response was observed for algae in the Andean stream (77 of the variance)although fish also showed important biomass responses (about 9ndash48) The strongest biomassresponse to enrichment (77 in all biological compartments) was found in the Andean streamThe magnitude and seasonality of biomass responses to enrichment were highly site specificoften depending on the basal nutrient concentration and on windows of ecological opportunity(periods when environmental constraints other than nutrients do not limit biomass growth)The Pampean stream with high basal nutrient concentrations showed a weak response toenrichment (except for invertebrates) whereas the greater responses of Andean streamcommunities were presumably favored by wider windows of ecological opportunity incomparison to those from the Mediterranean stream Despite variation among sitesenrichment globally stimulated the algal-based food webs (algae and invertebrate grazers) butnot the detritus-based food webs (bacteria and invertebrate shredders) This study shows that
Content from this work may be used under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 30 licence Anyfurther distribution of this work must maintain attribution to the author(s) and the title of the work journal citation and DOI
11748-932613014002+13$3300 ccopy 2013 IOP Publishing Ltd Printed in the UK
Environ Res Lett 8 (2013) 014002 J Artigas et al
nutrient enrichment tends to globally enhance the biomass of stream biological assemblagesbut that its magnitude and extent within the food web are complex and are strongly determinedby environmental factors and ecosystem structure
Keywords biomes fish invertebrates microbial biomass nutrient enrichment streamecosystems windows of opportunity
S Online supplementary data available from stacksioporgERL8014002mmedia
1 Introduction
Nutrient enrichment threatens aquatic ecosystems globally(UNEP 2007) causing harmful algal blooms alteringbiological communities and affecting the functioning offreshwater and coastal marine ecosystems (Smith 2003Sutton et al 2011) The effects of enrichment are particularlystrong in streams and estuaries where both biodiversityand efficiency of ecosystem processes are severely impaired(Sobczak et al 2005 Vorosmarty et al 2010) Eutrophicationdirectly affects primary producers (algae macrophytes) andmicrobial heterotrophs (bacteria fungi) as they can usedissolved nutrients (Stelzer et al 2003) but the effects can thenspread bottom-up to other trophic levels (Hart and Robinson1990) Nutrients can stimulate decomposition (Pascoal et al2005) as well as primary production (Mulholland et al2001) two key processes with implications on the streamfood webs (Tank et al 2010) However mixed bottom-upand top-down regulations are common in streams theinteraction between nutrient availability and grazing intensityregulating the biomass of primary producers and consumers(Holomuzki et al 2010) Furthermore the response ofdifferent groups of organisms to modified trophic resourcescan be quite specific and may also depend on partitioningand compartmentalization of food webs (Huxel et al 2002Stouffer and Bascompte 2011)
The overall response of autotrophs and heterotrophs tonutrient enrichment can be higher when the backgroundnutrient concentrations are low and other factors such aslight availability temperature and hydrological stability donot limit primary production (Hill et al 2009) The occurrenceof favorable periods for primary production and invertebratedevelopment largely depends on the phenology and climatecharacteristics of the site which mark the lsquowindows ofecological opportunityrsquo (Tockner et al 2010) For instancethe growth of autotrophs in Mediterranean forested streamsis mainly limited to a narrow window of opportunity in latewinter and early spring a period before the canopy closesand that usually has minimal flood disturbance whereasheterotrophs tend to peak in late summerndashautumn (eg Artigaset al 2009) In contrast many tropical streams are lessseasonal and feature small variability in temperature andlight availability throughout the year while hydrologicalpeaks tend to control algal biomass (Zapata and Donato2008) Therefore although nutrient enrichment is becomingincreasingly widespread the response of river ecosystems canbe site specific depending on local characteristics like climateor type of dominant organisms Thus it is important to gain an
understanding of the factors governing the response of riverecosystems to nutrient enrichment under different climaticsettings This objective can be best attained by means ofcontrolled experiments
We analyzed the biological responses of streamecosystems to experimental nutrient enrichment in threebioclimatic regions (Mediterranean Pampean and Andean)The systems differed in environmental features (hydrologytemperature light availability biological communities) aswell as in the extent of their respective nutrient limitationOur main hypothesis was that the three systems wouldrespond in specific ways to a similar nutrient enrichment Wehypothesized the increase in biomass to be proportional tobasal nutrient limitation and the duration of the response tobe limited by the windows of opportunity
2 Methods
21 Characteristics of the study sites
The experimental manipulations were conducted in threeheadwater streams located in the Mediterranean (FuirososSpain) Pampean (La Choza Argentina) and tropical Andean(Tota Colombia) regions All streams were of similar orderbut had specific climatic and ecoregional characteristicsdiffering in temperature patterns background nutrient levelslight and discharge as well as in the canopy cover
Fuirosos is a Mediterranean forested stream located at analtitude of 150 m in the mountains of NE Spain It drainsa siliceous 16 km2 basin covered by pine (Pinus halepensisMill) and cork oak (Quercus suber L) forests with less than5 covered by arable fields The climate is Mediterraneanwith hot dry summers and wet winters The average annualrainfall is 600ndash800 mm and the average water temperaturesrange from 4 C (January) to 20 C (August) The experimentswere performed in a third order section (41 41primeN 2 34primeE)with a well developed deciduous riparian forest composed ofhybrid plane (Platanus acerifolia (Aiton) Willd) and blackpoplar (Populus nigra L) which cast a strong shade fromlate spring (April) to autumn (November) The stream runsover boulders cobbles and sand but the seasonal input ofleaf detritus in autumn can completely cover the streambed (Artigas et al 2009) The main primary producers aremicroscopic algae growing on rocks and in sand which tendto be most abundant in early spring before canopy closure(Tornes and Sabater 2010)
La Choza is a Pampean stream located in the lowlandprairies of the province of Buenos Aires It drains a basin
2
Environ Res Lett 8 (2013) 014002 J Artigas et al
of 48 km2 on deep loess deposits mainly covered bytemperate grassland grazed by cattle although a quarter ofthe land is devoted to agriculture The climate is temperatecontinental resulting in wet springs and autumns The averageannual rainfall is 600ndash1200 mm and the average watertemperatures range from 10 C (June) to 24 C (January) Theexperiments were conducted in a third order section (34 42primeS59 04primeO) that was naturally devoid of riparian trees Thestreambed was composed of carbonate calcium precipitatesand deposits of silt and clay that re-suspend during stormepisodes (Rodrigues Capıtulo et al 2010) Primary producersare diverse and include suspended cells epipelic microalgaeand macrophytes
Tota is an Andean stream located at an altitude of2560 m in the Eastern Mountains of Colombia in BoyacaState The stream drains a 140 km2 basin on shale and clayApproximately 40 of the surface area of the basin is usedfor agriculture and pasture the rest is covered by secondaryforest and sparse vegetation The climate is cool and dry witha bimodal precipitation pattern temperatures showing littleseasonal variation The average annual rainfall is 730 mmand average water temperatures range from 11 to 13 CExperiments were conducted in a third order section wherethe riparian vegetation is composed of open forest dominatedby Alnus acuminata Kunth and Salix humboldtiana Willdwith a dense layer of Pennisetum clandestinum Hochst exChiov grass The streambed is composed of rocks cobblesand boulders and few deposits of sand and detritus Thoughlitter fall is relatively constant through the year detritusaccumulates during periods of low flow Macrophytes are rareor absent and epilithic and epipsammic algae are the mainprimary producers
22 Experimental manipulations
The experiment followed a BACIPS (beforendashafter controlndashimpact paired series) design in each stream we experimen-tally enhanced nutrient concentrations 2ndash4 fold over a 50 mreach (E enriched) whereas an upstream reach of similarmorphological and hydrological characteristics was kept asthe control (C) All reaches (C and E) were studied for 9ndash15months before the enrichment E reaches were then enrichedand all reaches studied for one year Nutrient amounts toadd to E reaches were calculated bi-weekly considering thebasal stream water nutrient concentrations (total ammonianitrate and phosphate) and discharge Nutrient solutions wereprepared with ammonium nitrate (NH4NO3) and ammoniumphosphate (NH4H2PO4) stored in 100 l tanks beside thestream bank and added continuously to the E reach Given thelow water velocity bags of fertilizer (12 N and 12 P) wereused to produce the necessary enrichment in the E reach ofthe Pampean stream The enrichment started in January 2004in the Mediterranean January 2007 in the Pampean and May2008 in the Andean stream
Ammonium nitrate and phosphate concentrations wereanalyzed in each of the stream reaches before and afterthe enrichment Water samples taken in triplicate werefiltered through 07 microm GFF filters (Whatman England)
and placed into 50 ml flasks for further analysis in thelaboratory Colorimetric methods were used for total ammonia(Solorzano 1969) phosphate (Murphy and Riley 1962) andnitrate (APHA 1989) determination
23 Biological analyses
The sampling was adapted to study the main compartmentspresent in each of the streams fine sediment and rockswere sampled in the Mediterranean and Andean streams finesediment the water column and macrophytes in the Pampeanstream All reaches were sampled monthly On each occasionwater temperature conductivity pH oxygen irradiance anddischarge were measured with portable meters and samplesto analyze nutrient concentrations and benthic organic matteraccumulation were collected
231 Rocky habitat Bacterial and algal biomass in theepilithon was measured from 1 cm2 ceramic tiles gluedonto rock slabs that were kept at a depth of 10ndash20 cm inriffle zones The samples were allowed to colonize for atleast two months before sampling On each occasion threetiles were taken for bacterial density estimates and preservedwith 4 formaldehyde and three more tiles were taken forchlorophyll-a concentration and kept frozen until analysisBacterial density was determined following Porter and Feig(1980) and chlorophyll-a concentration determined using themethod of Jeffrey and Humphrey (1975) Transformation intomicrobial biomass (microg carbon per cmminus2) was achieved byusing conversion factors of Frost et al (2005) for chlorophyll-aand Bratbak and Dundas (1984) for bacterial densities
The taxonomic composition of the diatom communitywas determined under the microscope Diatom communitieswere detached by sonication from glass tiles cleaned withsulfuric acid and hydrogen peroxide and mounted ontopermanent slides using Naphrax (refractive index of 174Brunel Microscopes Ltd England) About 400 frustules werecounted per sample and diatoms were identified to the lowesttaxonomic level possible using standard references (Krammerand Lange-Bertalot 1986ndash1991) and recent nomenclatureupdates
Invertebrates in the Andean stream were sampled directlyon cobbles using a Surber sampler with a 900 cm2 surfacearea and a 500 microm mesh size The samples (at least 5replicates) collected were fixed with 4 formaldehyde andinvertebrates were sorted under a dissecting microscope at400times Identification of invertebrates reached the genera leveland the species level when possible Biomass was calculatedas dry mass (70 C for 48 h) per square meter In theMediterranean stream the grazer Ancylus fluviatilis Mullerwas the most abundant invertebrate upon rocks ThereforeAncylus body mass was estimated as the average dry mass(70 C 48 h) of 40ndash100 individuals (corresponding to 10ndash20cobbles sampled) in C and E reaches during enrichmentexperiments There was no rocky substratum in the Pampeanstream
3
Environ Res Lett 8 (2013) 014002 J Artigas et al
232 Sediment habitat Epipsammic algae and bacteriawere sampled with plastic cylinders (43 cm in diameter2ndash4 cm depth) used as corers Three replicates were collectedat random at C and E reaches placed in plastic containers andsubsampled using an untapped syringe (12 cm in diameter)and equivalent to 1 g of sediment Samples were theneither preserved with 4 formaldehyde for bacterial countingor kept frozen to quantify chlorophyll-a The protocolsfor bacterial density and chlorophyll-a determination weresimilar to those used for the rocks Biomass results wereexpressed in microg carbon per cmminus2 of sediment
In the Mediterranean and Andean streams invertebratesin fine sediments were collected with plastic cores (5 cmin diameter 7ndash8 cm depth) Three core replicates werecollected per reach (C and E) and stream type In the Pampeanstream an Ekman dredge sampler (100 cm2 surface area) wasused to sample sediments (also triplicate samples) Sampleswere immediately filtered through 500 microm sieves and fixedwith formaldehyde Invertebrate identification and biomassdetermination procedures were similar to those used for therock assemblages
233 Water column In the Pampean stream suspendedchlorophyll-a was analyzed from 1 l water samples (3replicates per reach and occasion) Water was filtered throughGFF filters (07 microm Whatman England) and the filterswere analyzed for chlorophyll as the extracts from rocksBacterial density was estimated in 10 ml water subsamples(3 replicates) following the methodology described in Porterand Feig (1980) Algal and bacterial biomass in thewater column was estimated through the conversion factorsdescribed before and expressed by square meter of streambed
234 Macrophytes Macrophytes were surveyed bimonthlyduring the cold season and monthly during the warm seasonat the Pampean stream Macrophyte composition and coverwere determined from regularly spaced transects Macrophytesamples (3ndash10 replicates) were obtained at random in the Cand E reaches with a 649 cm2 square metal frame and stored in500 ml plastic bags Plants were rinsed with tap water sortedinto species and invertebrates removed Plants were dried(70 C to a constant dry weight) to obtain dry mass which wasexpressed in g mminus2 Invertebrates removed from macrophyteswere filtered through 500microm sieves fixed with formaldehydeidentified and counted Identification of invertebrates reachedthe genera level and the species level when possible
235 Fish Fish abundance and biomass in the Pampean andAndean streams were estimated bimonthly using multiple-pass sampling (at least four passes per occasion) afterblocking the C and E reaches with nets Fish density wastoo low in the Mediterranean stream (Mas-Martı et al 2010)to allow comparative sampling between reaches Samplingwas performed with electrofishing (EN ISO 140112003standard) In the Pampean stream sampling was also donewith seines when high water conductivity precluded the useof electrofishing All fish sampled were counted identified
measured (standard length) and then returned to the waterOnly a small subsample of about 20 individuals wasused to determine fish condition (body weight and lengthGarcıa-Berthou and Moreno-Amich 1993) Fish abundanceand biomass were expressed as catch per unit of effort(CPUE = fish numberarea sampled) and biomass per unit ofeffort (BPUE = fish biomassarea sampled) respectively
24 Data analysis
The results of the three enrichment experiments wereanalyzed following the predictive-BACIPS approach (Benceet al 1996) This method compares the relationship betweenthe values in the C and E reaches before and after the start ofthe experiment Although beforendashafter controlndashimpact pairedseries (BACIPS) designs are often analyzed with ANOVA theuse of statistical techniques with control sites as covariatesas in the predictive-BACIPS has the advantage of allowingthe effect size (ie the enrichment effects) to vary withthe magnitude of the control value and with environmentalconditions ie does not require the standard assumption ofadditivity (Bence et al 1996 Stewart-Oaten and Bence 2001)
For the predictive-BACIPS approach we used analysis ofcovariance (ANCOVA) of the values of the response variable(eg bacterial biomass) in the E reaches using values inC (control) as a covariate and the periods beforeafter thestart of the treatment (BA) as a binary factor ANCOVA wasfirst used to test interactions between the covariate (ie Cvalues) and the binary factor (BA) Significant interactionsindicate that slopes between both variables (correspondingto C and E values) were not equal before and after thestart of the treatment (Garcıa-Berthou and Moreno-Amich1993) In that case the treatment effect would depend onthe magnitude of the control values When the C times BAinteraction was not significant (P gt 010) it was removedfrom the model to improve the statistical power and astandard ANCOVA design was used (ie homogeneous slopeswere assumed Garcıa-Berthou and Moreno-Amich 1993)The proportion of variance (often termed eta squared η2)explained by the BA factor plus the C times BA interactionwas computed as a measure of the effect size of enrichmentBence et al (1996) recommend that emphasis in BACIPSdesigns is mainly placed on effect sizes rather than on Pvalues Average η2 values in the biological compartments(bacteria algae macrophytes water column invertebratesand fish) in each stream were calculated as an estimateof the magnitude of transferral of the nutrient enrichmentthrough the food web All data were analyzed with the Rsoftware (R Development Core Team 2012) Note that thedefault packages in R use sequential (ie type I) sum ofsquares (Hector et al 2010) and that the lsquocontrolrsquo covariatewas always tested first in the models Quantitative variables(response variables and covariates) were log10-transformedbefore statistical analyses in order to improve assumptions ofthe linear models (normality homoscedasticity and linearity)
An important assumption of BACIPS designs andregression of time-related variables in general is the lackof serial correlation (Bence et al 1996 Stewart-Oaten and
4
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 1 Environmental characteristics and organic matter availability in the Mediterranean Pampean and Andean streams Values ofwater temperature (C) light (micromol photons mminus2 sminus1) and discharge (l sminus1) during enrichment experiments are represented Windows ofecological opportunity (marked in gray right plot) were determined after considering favorable conditions for primary producers microbialheterotrophs and consumer development
Bence 2001) We used the BreuschndashGodfrey test to testwhether serial correlation was present in our data withserial correlation up to four order using the lsquobgtestrsquo functionof package lsquolmtestrsquo (Zeileis and Hothorn 2002) in R TheBreuschndashGodfrey test is more applicable and often morepowerful than the more widely used DurbinndashWatson testand other methods (Godfrey and Tremayne 2005) Only 1of the 23 analyses (combinations of variables and streams)showed significant serial correlation (P lt 005) Similarlyonly one case of the 23 raw variables showed serial correlation(P lt 005) in the enriched zones probably because samplingfrequency was low (about monthly) and equally spaced(Stewart-Oaten and Bence 2001) Since our time series wererelatively short (sample size lt 50) and serial correlationwas almost always non-significant we did not correct forautocorrelation as recommended by Bence (1995)
The effect of enrichment on biomass of the differenttrophic levels was expressed as the per cent ratio of biomasschange in the E reach with respect to the C reach The ratio ofbiomass change was therefore calculated as 100 (enriched minuscontrol)control This ratio was also used to calculateenrichment effects on diatom and invertebrate abundancesThe ratiorsquos coefficient of variation (CV) during the enrichmentwas useful to determine the potential fluctuations in biomassresponse caused by the treatment Windows of opportunitywere defined as time periods when no factor beyondnutrients exerts significant limitation on primary productionand consumers in the stream These periods were identifiedin the Mediterranean Pampean and Andean streams afteranalyzing the best combinations of water temperature lightand discharge
3 Results
31 Physical and chemical characteristics of the study sites
The Mediterranean and Pampean streams had large seasonalvariations in water temperature and light availability whilein contrast the seasonal variation in the Andean stream wasmuch lower (figure 1) Water flow also followed differentpatterns in the three systems Variability was high in theAndean stream (from 68 to 1800 l sminus1) and Mediterraneanstream (from 0 to 1400 l sminus1) and very low in the Pampean(from 0 to 40 l sminus1) Low flow values included up to twomonths of summer drought in the Mediterranean stream butwater remained in the channel in the Pampean because of itsvery low slope
Basal nutrient concentrations before the enrichment(table 1) were the highest in the Pampean stream (140 microg lminus1
SRP and 700 microg lminus1 DIN) characterized by low phosphoruscontent in the Mediterranean stream (14 microg lminus1 SRP430 microg lminus1 DIN) and with low DIN concentrations in theAndean stream (26 microg lminus1 SRP and 110 microg Lminus1 DIN) Theenrichment caused SRP to increasetimes2ndash4 and DIN to increasetimes16ndash28 (table 1) The enrichment caused the decrease of theNP ratios in the Mediterranean and Pampean streams but didnot greatly alter those of the Andean (table 1)
32 Windows of opportunity
Windows of ecological opportunity for primary producersmicrobial heterotrophs and consumers differed between
5
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 1 Water nutrient concentrations of soluble reactive phosphorus (SRP) and dissolved inorganic nitrogen (DIN sum ofammonia+ nitrate+ nitrite forms) in the C and E reaches of the Mediterranean Pampean and Andean streams Values are means andstandard errors of nutrients measured during the pre-enrichment (before) and enrichment (after) periods and are accompanied by NP molarratio calculations The increment (1) nutrient concentration during enrichment is also shown for each of the streams
Control reach Enriched reach 1 nutrient concentration
Mediterranean Before SRP (microg lminus1) 14plusmn 18 16plusmn 21DIN (microg lminus1) 432plusmn 438 523plusmn 491NP 68 71
After SRP (microg lminus1) 10plusmn 9 30plusmn 13 times3DIN (microg lminus1) 389plusmn 495 765plusmn 736 times2NP 84 56
Pampean Before SRP (microg lminus1) 143plusmn 17 173plusmn 31DIN (microg lminus1) 683plusmn 39 1093plusmn 228NP 11 14
After SRP (microg lminus1) 93plusmn 3 361plusmn 29 times4DIN (microg lminus1) 528plusmn 136 828plusmn 197 times16NP 13 5
Andean Before SRP (microg lminus1) 26plusmn 5 28plusmn 5DIN (microg lminus1) 55plusmn 21 78plusmn 23NP 5 6
After SRP (microg lminus1) 53plusmn 4 105plusmn 13 times2DIN (microg lminus1) 114plusmn 59 323plusmn 85 times28NP 5 7
streams in terms of duration and frequency (figure 1)Windows of opportunity in the Mediterranean stream werenarrow and limited to late springndashearly summer (moderatetemperature and light availability) and late summerndashearlyautumn periods (moderate temperatures and abundant al-lochthonous OM inputs) The greatest overlap of favor-able environmental conditions for biological developmentoccurred in the Pampean stream In this system higher lightelevated temperatures and low discharge values coincidedfrom winter to summer (SeptemberndashMarch) The absenceof riparian vegetation reduced allochthonous OM inputsin this stream but authocthonous OM was produced byalgae and macrophytes The Andean stream also had longfavorable periods of temperature light and allochthonous OMinputs only interrupted by high discharge periods betweenAprilndashMay and OctoberndashNovember The high flow periodsin this stream caused a momentary biomass decrease of allbiological groups The predictive-BACIPS approach (table 2)suggested that 10 out of 23 models had significant treatmentseffects (or interactions) and that the explained variance wasoften high (figure 6)
33 Bacterial biomass
Bacterial biomass before the enrichment ranged from 2to 16 microg C cmminus2 on rocks and fine sediment in theMediterranean and Andean streams from 5 to 700microg C cmminus2
on sediments 004ndash15 microg C cmminus2 on macrophytes and1ndash36microg C cmminus2 in the water column for the Pampean streamThe nutrient enrichment resulted in an increase in bacterialbiomass in the Mediterranean (rocks and fine sediment) andPampean (fine sediment) streams but not in the Andeanstream (figure 2 table 2) The largest increase in bacterial
biomass after enrichment (calculated as the biomass changeratio) occurred in the Mediterranean stream (65ndash80 overallincrease) In the Pampean stream bacterial biomass increasedbetween 20 and 30 in fine sediment and macrophytesbut decreased in the water column (10ndash20) Water flowconditions temperature and light availability covaried withbacterial biomass in the E reaches of the Mediterranean andPampean streams The temporal variability of the biomasschange ratio was the highest in the Mediterranean stream(coefficient of variation CV = 24) due to the episodichigher biomass between late spring and early summer in theenrichment period
34 Algal and macrophytic biomass and composition
Algal biomass before the enrichment averaged 205 plusmn31 microg C cmminus2 on rocks and 69plusmn11 microg C cmminus2 on sedimentsat the Mediterranean stream (figure 3) Algal biomassaccounted for 137 plusmn 18 microg C cmminus2 on rocky habitat in theAndean stream while reached 162 plusmn 29 microg C cmminus2 onfine sediment and 23 plusmn 3 microg C cmminus2 for suspended algaein the Pampean stream In this stream macrophytes biomassaccounted for 58plusmn 27 g C mminus2
Nutrient enrichment produced an increase in algalbiomass in the three streams (figure 3 table 2) althoughthe responses varied in magnitude and differed betweencompartments The largest increase occurred on rocks inthe Mediterranean stream (163 on average peak of 273during spring) In the Andean stream algal biomass on rocksrapidly increased (62) and remained steady thereafter Thesmallest biomass increase was registered in the Pampeanstream particularly in the water column and for macrophytesbut biomass increased up to 49 on fine sediments The
6
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 2 Predictive-BACIPS of the effects of nutrient enrichment on the biomass of biological compartments in the three streams Thevalues shown correspond to the estimated coefficients of the linear models (ANCOVAs) When the control times BA was not significant(P gt 010) it was deleted from the model see section 2 for further details ns P gt 010 a P le 010 P le 005 P le 001 P le 0001
Variable n Control BA Control times BA
MediterraneanBacteria rocks 18 0404 0068 nsBacteria fine sediment 23 0590 0078 nsAlgae rocks 19 0495 0202a nsAlgae fine sediment 23 1082 0315 minus0732Invertebrates fine sediment 7 minus1011 minus0072 1660a
PampeanBacteria fine sediment 11 0751 0510a nsBacteria suspended 11 0954 minus0018 nsBact macrophytes 8 0396 0097 nsAlgae fine sediment 10 1106 0206 nsAlgae suspended 5 0274 minus0079 nsAlgae macrophyte 10 0575 minus0150 nsMacrophytes 11 1093 0249 nsInvertebrates fine sediment 10 minus0130 0787 nsInvertebrates macrophytes 6 minus0251 minus0521a nsFish CPUE 4 1082 8221 nsFish BPUE 4 0745 29552 ns
AndeanBacteria rocks 7 0666a 0299 nsBacteria fine sediment 6 0150 minus2631a nsAlgae rocks 14 0158 minus0101 0728Invertebrates rocks 13 0469 minus0269 nsInvertebrates fine sediment 13 0432 minus0388 nsFish density 10 1012 minus0442 nsFish biomass 10 1020 minus0464a ns
algal biomass responses to nutrients in the three streamswere strongly related with favorable conditions of lighttemperature and water flow (figure 3) The difference betweenthe extension of the respective windows of opportunitydetermined the steadiness of the response The Mediterraneanand Andean streams represented the maximum and minimumvariability in favorable conditions for primary producersrsquogrowth accordingly the coefficients of variation of thebiomass ratiorsquos change were the highest in the Mediterranean(CV = 16) and the lowest in the Andean (CV = 08)stream
Nutrient enrichment affected the composition of benthicalgal assemblages Some diatom taxa decreased (egAchnanthes biasolettiana Grunow (minus57) Achnanthesminutissima Kutzing (minus432) in the MediterraneanFragilaria ulna (Nitzsch) Lange-Bertalot var ulna (minus68)in the Pampean and Cocconeis placentula Ehrenberg(minus40) in the Andean stream) whereas others increased(eg Rhoicosphenia abbreviata (Ag) Lange-Bertalot (94)and Cocconeis placentula Ehrenberg (594) in theMediterranean Nitzschia frustulum (Kutzing) Grunow varfrustulum (3528) in the Pampean and Reimeria sinuata(Gregory) Kociolek and Stoermer (646) in the Andeanstream) Enrichment in the Pampean stream had no effect onthe macrophyte composition Ludwigia sp and Bacopa spdominated both the C and E reaches and were later replacedby the macroalga Spirogyra sp
35 Invertebrate biomass and composition
Invertebrate biomass before the enrichment ranged from5 to 750 microg DM cmminus2 in the Mediterranean and Andeanstreams and reached 1093 plusmn 311 microg DM cmminus2 in thesediments of the Pampean stream The effects of nutrientenrichment were generally non-significant but often larger(figure 4 table 2) and more pronounced in some habitatsthan others In the Pampean stream effects were not apparenton invertebrates occurring on macrophytes and were minor(3) on fine sediments Invertebrate biomass increased upto 155 in the fine sediments of the Mediterranean streamwhile no increase was observed in the Andean stream Theeffects on invertebrate biomass were the least variable inthe Mediterranean stream (per cent increase ratio CV = 09CV = 12 in the Andean and 27 in the Pampean)
Although the general enrichment effects on invertebratebiomass were weak some specific invertebrate taxa weredistinctly affected The grazer Ancylus fluviatilis increasedby 17 in the Mediterranean stream and the grazerCamelobaetidus increased by 927 in the Andean streamDensities of the scrapers Heleobia parchappei Orbigny(in fine sediments) and Hyalella curvispina Shoemaker(on macrophytes) increased in the Pampean stream (48and 762 respectively) Potamopyrgus antipodarum Gray(scraper) and Corbicula fluminea Muller (filterer) occurredonly in the respective fertilized reaches of the Mediterraneanand Pampean streams The Tubificidae (minus38) and theTanytarsini chironomidae (minus29) significantly decreased in
7
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
nutrient enrichment tends to globally enhance the biomass of stream biological assemblagesbut that its magnitude and extent within the food web are complex and are strongly determinedby environmental factors and ecosystem structure
Keywords biomes fish invertebrates microbial biomass nutrient enrichment streamecosystems windows of opportunity
S Online supplementary data available from stacksioporgERL8014002mmedia
1 Introduction
Nutrient enrichment threatens aquatic ecosystems globally(UNEP 2007) causing harmful algal blooms alteringbiological communities and affecting the functioning offreshwater and coastal marine ecosystems (Smith 2003Sutton et al 2011) The effects of enrichment are particularlystrong in streams and estuaries where both biodiversityand efficiency of ecosystem processes are severely impaired(Sobczak et al 2005 Vorosmarty et al 2010) Eutrophicationdirectly affects primary producers (algae macrophytes) andmicrobial heterotrophs (bacteria fungi) as they can usedissolved nutrients (Stelzer et al 2003) but the effects can thenspread bottom-up to other trophic levels (Hart and Robinson1990) Nutrients can stimulate decomposition (Pascoal et al2005) as well as primary production (Mulholland et al2001) two key processes with implications on the streamfood webs (Tank et al 2010) However mixed bottom-upand top-down regulations are common in streams theinteraction between nutrient availability and grazing intensityregulating the biomass of primary producers and consumers(Holomuzki et al 2010) Furthermore the response ofdifferent groups of organisms to modified trophic resourcescan be quite specific and may also depend on partitioningand compartmentalization of food webs (Huxel et al 2002Stouffer and Bascompte 2011)
The overall response of autotrophs and heterotrophs tonutrient enrichment can be higher when the backgroundnutrient concentrations are low and other factors such aslight availability temperature and hydrological stability donot limit primary production (Hill et al 2009) The occurrenceof favorable periods for primary production and invertebratedevelopment largely depends on the phenology and climatecharacteristics of the site which mark the lsquowindows ofecological opportunityrsquo (Tockner et al 2010) For instancethe growth of autotrophs in Mediterranean forested streamsis mainly limited to a narrow window of opportunity in latewinter and early spring a period before the canopy closesand that usually has minimal flood disturbance whereasheterotrophs tend to peak in late summerndashautumn (eg Artigaset al 2009) In contrast many tropical streams are lessseasonal and feature small variability in temperature andlight availability throughout the year while hydrologicalpeaks tend to control algal biomass (Zapata and Donato2008) Therefore although nutrient enrichment is becomingincreasingly widespread the response of river ecosystems canbe site specific depending on local characteristics like climateor type of dominant organisms Thus it is important to gain an
understanding of the factors governing the response of riverecosystems to nutrient enrichment under different climaticsettings This objective can be best attained by means ofcontrolled experiments
We analyzed the biological responses of streamecosystems to experimental nutrient enrichment in threebioclimatic regions (Mediterranean Pampean and Andean)The systems differed in environmental features (hydrologytemperature light availability biological communities) aswell as in the extent of their respective nutrient limitationOur main hypothesis was that the three systems wouldrespond in specific ways to a similar nutrient enrichment Wehypothesized the increase in biomass to be proportional tobasal nutrient limitation and the duration of the response tobe limited by the windows of opportunity
2 Methods
21 Characteristics of the study sites
The experimental manipulations were conducted in threeheadwater streams located in the Mediterranean (FuirososSpain) Pampean (La Choza Argentina) and tropical Andean(Tota Colombia) regions All streams were of similar orderbut had specific climatic and ecoregional characteristicsdiffering in temperature patterns background nutrient levelslight and discharge as well as in the canopy cover
Fuirosos is a Mediterranean forested stream located at analtitude of 150 m in the mountains of NE Spain It drainsa siliceous 16 km2 basin covered by pine (Pinus halepensisMill) and cork oak (Quercus suber L) forests with less than5 covered by arable fields The climate is Mediterraneanwith hot dry summers and wet winters The average annualrainfall is 600ndash800 mm and the average water temperaturesrange from 4 C (January) to 20 C (August) The experimentswere performed in a third order section (41 41primeN 2 34primeE)with a well developed deciduous riparian forest composed ofhybrid plane (Platanus acerifolia (Aiton) Willd) and blackpoplar (Populus nigra L) which cast a strong shade fromlate spring (April) to autumn (November) The stream runsover boulders cobbles and sand but the seasonal input ofleaf detritus in autumn can completely cover the streambed (Artigas et al 2009) The main primary producers aremicroscopic algae growing on rocks and in sand which tendto be most abundant in early spring before canopy closure(Tornes and Sabater 2010)
La Choza is a Pampean stream located in the lowlandprairies of the province of Buenos Aires It drains a basin
2
Environ Res Lett 8 (2013) 014002 J Artigas et al
of 48 km2 on deep loess deposits mainly covered bytemperate grassland grazed by cattle although a quarter ofthe land is devoted to agriculture The climate is temperatecontinental resulting in wet springs and autumns The averageannual rainfall is 600ndash1200 mm and the average watertemperatures range from 10 C (June) to 24 C (January) Theexperiments were conducted in a third order section (34 42primeS59 04primeO) that was naturally devoid of riparian trees Thestreambed was composed of carbonate calcium precipitatesand deposits of silt and clay that re-suspend during stormepisodes (Rodrigues Capıtulo et al 2010) Primary producersare diverse and include suspended cells epipelic microalgaeand macrophytes
Tota is an Andean stream located at an altitude of2560 m in the Eastern Mountains of Colombia in BoyacaState The stream drains a 140 km2 basin on shale and clayApproximately 40 of the surface area of the basin is usedfor agriculture and pasture the rest is covered by secondaryforest and sparse vegetation The climate is cool and dry witha bimodal precipitation pattern temperatures showing littleseasonal variation The average annual rainfall is 730 mmand average water temperatures range from 11 to 13 CExperiments were conducted in a third order section wherethe riparian vegetation is composed of open forest dominatedby Alnus acuminata Kunth and Salix humboldtiana Willdwith a dense layer of Pennisetum clandestinum Hochst exChiov grass The streambed is composed of rocks cobblesand boulders and few deposits of sand and detritus Thoughlitter fall is relatively constant through the year detritusaccumulates during periods of low flow Macrophytes are rareor absent and epilithic and epipsammic algae are the mainprimary producers
22 Experimental manipulations
The experiment followed a BACIPS (beforendashafter controlndashimpact paired series) design in each stream we experimen-tally enhanced nutrient concentrations 2ndash4 fold over a 50 mreach (E enriched) whereas an upstream reach of similarmorphological and hydrological characteristics was kept asthe control (C) All reaches (C and E) were studied for 9ndash15months before the enrichment E reaches were then enrichedand all reaches studied for one year Nutrient amounts toadd to E reaches were calculated bi-weekly considering thebasal stream water nutrient concentrations (total ammonianitrate and phosphate) and discharge Nutrient solutions wereprepared with ammonium nitrate (NH4NO3) and ammoniumphosphate (NH4H2PO4) stored in 100 l tanks beside thestream bank and added continuously to the E reach Given thelow water velocity bags of fertilizer (12 N and 12 P) wereused to produce the necessary enrichment in the E reach ofthe Pampean stream The enrichment started in January 2004in the Mediterranean January 2007 in the Pampean and May2008 in the Andean stream
Ammonium nitrate and phosphate concentrations wereanalyzed in each of the stream reaches before and afterthe enrichment Water samples taken in triplicate werefiltered through 07 microm GFF filters (Whatman England)
and placed into 50 ml flasks for further analysis in thelaboratory Colorimetric methods were used for total ammonia(Solorzano 1969) phosphate (Murphy and Riley 1962) andnitrate (APHA 1989) determination
23 Biological analyses
The sampling was adapted to study the main compartmentspresent in each of the streams fine sediment and rockswere sampled in the Mediterranean and Andean streams finesediment the water column and macrophytes in the Pampeanstream All reaches were sampled monthly On each occasionwater temperature conductivity pH oxygen irradiance anddischarge were measured with portable meters and samplesto analyze nutrient concentrations and benthic organic matteraccumulation were collected
231 Rocky habitat Bacterial and algal biomass in theepilithon was measured from 1 cm2 ceramic tiles gluedonto rock slabs that were kept at a depth of 10ndash20 cm inriffle zones The samples were allowed to colonize for atleast two months before sampling On each occasion threetiles were taken for bacterial density estimates and preservedwith 4 formaldehyde and three more tiles were taken forchlorophyll-a concentration and kept frozen until analysisBacterial density was determined following Porter and Feig(1980) and chlorophyll-a concentration determined using themethod of Jeffrey and Humphrey (1975) Transformation intomicrobial biomass (microg carbon per cmminus2) was achieved byusing conversion factors of Frost et al (2005) for chlorophyll-aand Bratbak and Dundas (1984) for bacterial densities
The taxonomic composition of the diatom communitywas determined under the microscope Diatom communitieswere detached by sonication from glass tiles cleaned withsulfuric acid and hydrogen peroxide and mounted ontopermanent slides using Naphrax (refractive index of 174Brunel Microscopes Ltd England) About 400 frustules werecounted per sample and diatoms were identified to the lowesttaxonomic level possible using standard references (Krammerand Lange-Bertalot 1986ndash1991) and recent nomenclatureupdates
Invertebrates in the Andean stream were sampled directlyon cobbles using a Surber sampler with a 900 cm2 surfacearea and a 500 microm mesh size The samples (at least 5replicates) collected were fixed with 4 formaldehyde andinvertebrates were sorted under a dissecting microscope at400times Identification of invertebrates reached the genera leveland the species level when possible Biomass was calculatedas dry mass (70 C for 48 h) per square meter In theMediterranean stream the grazer Ancylus fluviatilis Mullerwas the most abundant invertebrate upon rocks ThereforeAncylus body mass was estimated as the average dry mass(70 C 48 h) of 40ndash100 individuals (corresponding to 10ndash20cobbles sampled) in C and E reaches during enrichmentexperiments There was no rocky substratum in the Pampeanstream
3
Environ Res Lett 8 (2013) 014002 J Artigas et al
232 Sediment habitat Epipsammic algae and bacteriawere sampled with plastic cylinders (43 cm in diameter2ndash4 cm depth) used as corers Three replicates were collectedat random at C and E reaches placed in plastic containers andsubsampled using an untapped syringe (12 cm in diameter)and equivalent to 1 g of sediment Samples were theneither preserved with 4 formaldehyde for bacterial countingor kept frozen to quantify chlorophyll-a The protocolsfor bacterial density and chlorophyll-a determination weresimilar to those used for the rocks Biomass results wereexpressed in microg carbon per cmminus2 of sediment
In the Mediterranean and Andean streams invertebratesin fine sediments were collected with plastic cores (5 cmin diameter 7ndash8 cm depth) Three core replicates werecollected per reach (C and E) and stream type In the Pampeanstream an Ekman dredge sampler (100 cm2 surface area) wasused to sample sediments (also triplicate samples) Sampleswere immediately filtered through 500 microm sieves and fixedwith formaldehyde Invertebrate identification and biomassdetermination procedures were similar to those used for therock assemblages
233 Water column In the Pampean stream suspendedchlorophyll-a was analyzed from 1 l water samples (3replicates per reach and occasion) Water was filtered throughGFF filters (07 microm Whatman England) and the filterswere analyzed for chlorophyll as the extracts from rocksBacterial density was estimated in 10 ml water subsamples(3 replicates) following the methodology described in Porterand Feig (1980) Algal and bacterial biomass in thewater column was estimated through the conversion factorsdescribed before and expressed by square meter of streambed
234 Macrophytes Macrophytes were surveyed bimonthlyduring the cold season and monthly during the warm seasonat the Pampean stream Macrophyte composition and coverwere determined from regularly spaced transects Macrophytesamples (3ndash10 replicates) were obtained at random in the Cand E reaches with a 649 cm2 square metal frame and stored in500 ml plastic bags Plants were rinsed with tap water sortedinto species and invertebrates removed Plants were dried(70 C to a constant dry weight) to obtain dry mass which wasexpressed in g mminus2 Invertebrates removed from macrophyteswere filtered through 500microm sieves fixed with formaldehydeidentified and counted Identification of invertebrates reachedthe genera level and the species level when possible
235 Fish Fish abundance and biomass in the Pampean andAndean streams were estimated bimonthly using multiple-pass sampling (at least four passes per occasion) afterblocking the C and E reaches with nets Fish density wastoo low in the Mediterranean stream (Mas-Martı et al 2010)to allow comparative sampling between reaches Samplingwas performed with electrofishing (EN ISO 140112003standard) In the Pampean stream sampling was also donewith seines when high water conductivity precluded the useof electrofishing All fish sampled were counted identified
measured (standard length) and then returned to the waterOnly a small subsample of about 20 individuals wasused to determine fish condition (body weight and lengthGarcıa-Berthou and Moreno-Amich 1993) Fish abundanceand biomass were expressed as catch per unit of effort(CPUE = fish numberarea sampled) and biomass per unit ofeffort (BPUE = fish biomassarea sampled) respectively
24 Data analysis
The results of the three enrichment experiments wereanalyzed following the predictive-BACIPS approach (Benceet al 1996) This method compares the relationship betweenthe values in the C and E reaches before and after the start ofthe experiment Although beforendashafter controlndashimpact pairedseries (BACIPS) designs are often analyzed with ANOVA theuse of statistical techniques with control sites as covariatesas in the predictive-BACIPS has the advantage of allowingthe effect size (ie the enrichment effects) to vary withthe magnitude of the control value and with environmentalconditions ie does not require the standard assumption ofadditivity (Bence et al 1996 Stewart-Oaten and Bence 2001)
For the predictive-BACIPS approach we used analysis ofcovariance (ANCOVA) of the values of the response variable(eg bacterial biomass) in the E reaches using values inC (control) as a covariate and the periods beforeafter thestart of the treatment (BA) as a binary factor ANCOVA wasfirst used to test interactions between the covariate (ie Cvalues) and the binary factor (BA) Significant interactionsindicate that slopes between both variables (correspondingto C and E values) were not equal before and after thestart of the treatment (Garcıa-Berthou and Moreno-Amich1993) In that case the treatment effect would depend onthe magnitude of the control values When the C times BAinteraction was not significant (P gt 010) it was removedfrom the model to improve the statistical power and astandard ANCOVA design was used (ie homogeneous slopeswere assumed Garcıa-Berthou and Moreno-Amich 1993)The proportion of variance (often termed eta squared η2)explained by the BA factor plus the C times BA interactionwas computed as a measure of the effect size of enrichmentBence et al (1996) recommend that emphasis in BACIPSdesigns is mainly placed on effect sizes rather than on Pvalues Average η2 values in the biological compartments(bacteria algae macrophytes water column invertebratesand fish) in each stream were calculated as an estimateof the magnitude of transferral of the nutrient enrichmentthrough the food web All data were analyzed with the Rsoftware (R Development Core Team 2012) Note that thedefault packages in R use sequential (ie type I) sum ofsquares (Hector et al 2010) and that the lsquocontrolrsquo covariatewas always tested first in the models Quantitative variables(response variables and covariates) were log10-transformedbefore statistical analyses in order to improve assumptions ofthe linear models (normality homoscedasticity and linearity)
An important assumption of BACIPS designs andregression of time-related variables in general is the lackof serial correlation (Bence et al 1996 Stewart-Oaten and
4
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 1 Environmental characteristics and organic matter availability in the Mediterranean Pampean and Andean streams Values ofwater temperature (C) light (micromol photons mminus2 sminus1) and discharge (l sminus1) during enrichment experiments are represented Windows ofecological opportunity (marked in gray right plot) were determined after considering favorable conditions for primary producers microbialheterotrophs and consumer development
Bence 2001) We used the BreuschndashGodfrey test to testwhether serial correlation was present in our data withserial correlation up to four order using the lsquobgtestrsquo functionof package lsquolmtestrsquo (Zeileis and Hothorn 2002) in R TheBreuschndashGodfrey test is more applicable and often morepowerful than the more widely used DurbinndashWatson testand other methods (Godfrey and Tremayne 2005) Only 1of the 23 analyses (combinations of variables and streams)showed significant serial correlation (P lt 005) Similarlyonly one case of the 23 raw variables showed serial correlation(P lt 005) in the enriched zones probably because samplingfrequency was low (about monthly) and equally spaced(Stewart-Oaten and Bence 2001) Since our time series wererelatively short (sample size lt 50) and serial correlationwas almost always non-significant we did not correct forautocorrelation as recommended by Bence (1995)
The effect of enrichment on biomass of the differenttrophic levels was expressed as the per cent ratio of biomasschange in the E reach with respect to the C reach The ratio ofbiomass change was therefore calculated as 100 (enriched minuscontrol)control This ratio was also used to calculateenrichment effects on diatom and invertebrate abundancesThe ratiorsquos coefficient of variation (CV) during the enrichmentwas useful to determine the potential fluctuations in biomassresponse caused by the treatment Windows of opportunitywere defined as time periods when no factor beyondnutrients exerts significant limitation on primary productionand consumers in the stream These periods were identifiedin the Mediterranean Pampean and Andean streams afteranalyzing the best combinations of water temperature lightand discharge
3 Results
31 Physical and chemical characteristics of the study sites
The Mediterranean and Pampean streams had large seasonalvariations in water temperature and light availability whilein contrast the seasonal variation in the Andean stream wasmuch lower (figure 1) Water flow also followed differentpatterns in the three systems Variability was high in theAndean stream (from 68 to 1800 l sminus1) and Mediterraneanstream (from 0 to 1400 l sminus1) and very low in the Pampean(from 0 to 40 l sminus1) Low flow values included up to twomonths of summer drought in the Mediterranean stream butwater remained in the channel in the Pampean because of itsvery low slope
Basal nutrient concentrations before the enrichment(table 1) were the highest in the Pampean stream (140 microg lminus1
SRP and 700 microg lminus1 DIN) characterized by low phosphoruscontent in the Mediterranean stream (14 microg lminus1 SRP430 microg lminus1 DIN) and with low DIN concentrations in theAndean stream (26 microg lminus1 SRP and 110 microg Lminus1 DIN) Theenrichment caused SRP to increasetimes2ndash4 and DIN to increasetimes16ndash28 (table 1) The enrichment caused the decrease of theNP ratios in the Mediterranean and Pampean streams but didnot greatly alter those of the Andean (table 1)
32 Windows of opportunity
Windows of ecological opportunity for primary producersmicrobial heterotrophs and consumers differed between
5
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 1 Water nutrient concentrations of soluble reactive phosphorus (SRP) and dissolved inorganic nitrogen (DIN sum ofammonia+ nitrate+ nitrite forms) in the C and E reaches of the Mediterranean Pampean and Andean streams Values are means andstandard errors of nutrients measured during the pre-enrichment (before) and enrichment (after) periods and are accompanied by NP molarratio calculations The increment (1) nutrient concentration during enrichment is also shown for each of the streams
Control reach Enriched reach 1 nutrient concentration
Mediterranean Before SRP (microg lminus1) 14plusmn 18 16plusmn 21DIN (microg lminus1) 432plusmn 438 523plusmn 491NP 68 71
After SRP (microg lminus1) 10plusmn 9 30plusmn 13 times3DIN (microg lminus1) 389plusmn 495 765plusmn 736 times2NP 84 56
Pampean Before SRP (microg lminus1) 143plusmn 17 173plusmn 31DIN (microg lminus1) 683plusmn 39 1093plusmn 228NP 11 14
After SRP (microg lminus1) 93plusmn 3 361plusmn 29 times4DIN (microg lminus1) 528plusmn 136 828plusmn 197 times16NP 13 5
Andean Before SRP (microg lminus1) 26plusmn 5 28plusmn 5DIN (microg lminus1) 55plusmn 21 78plusmn 23NP 5 6
After SRP (microg lminus1) 53plusmn 4 105plusmn 13 times2DIN (microg lminus1) 114plusmn 59 323plusmn 85 times28NP 5 7
streams in terms of duration and frequency (figure 1)Windows of opportunity in the Mediterranean stream werenarrow and limited to late springndashearly summer (moderatetemperature and light availability) and late summerndashearlyautumn periods (moderate temperatures and abundant al-lochthonous OM inputs) The greatest overlap of favor-able environmental conditions for biological developmentoccurred in the Pampean stream In this system higher lightelevated temperatures and low discharge values coincidedfrom winter to summer (SeptemberndashMarch) The absenceof riparian vegetation reduced allochthonous OM inputsin this stream but authocthonous OM was produced byalgae and macrophytes The Andean stream also had longfavorable periods of temperature light and allochthonous OMinputs only interrupted by high discharge periods betweenAprilndashMay and OctoberndashNovember The high flow periodsin this stream caused a momentary biomass decrease of allbiological groups The predictive-BACIPS approach (table 2)suggested that 10 out of 23 models had significant treatmentseffects (or interactions) and that the explained variance wasoften high (figure 6)
33 Bacterial biomass
Bacterial biomass before the enrichment ranged from 2to 16 microg C cmminus2 on rocks and fine sediment in theMediterranean and Andean streams from 5 to 700microg C cmminus2
on sediments 004ndash15 microg C cmminus2 on macrophytes and1ndash36microg C cmminus2 in the water column for the Pampean streamThe nutrient enrichment resulted in an increase in bacterialbiomass in the Mediterranean (rocks and fine sediment) andPampean (fine sediment) streams but not in the Andeanstream (figure 2 table 2) The largest increase in bacterial
biomass after enrichment (calculated as the biomass changeratio) occurred in the Mediterranean stream (65ndash80 overallincrease) In the Pampean stream bacterial biomass increasedbetween 20 and 30 in fine sediment and macrophytesbut decreased in the water column (10ndash20) Water flowconditions temperature and light availability covaried withbacterial biomass in the E reaches of the Mediterranean andPampean streams The temporal variability of the biomasschange ratio was the highest in the Mediterranean stream(coefficient of variation CV = 24) due to the episodichigher biomass between late spring and early summer in theenrichment period
34 Algal and macrophytic biomass and composition
Algal biomass before the enrichment averaged 205 plusmn31 microg C cmminus2 on rocks and 69plusmn11 microg C cmminus2 on sedimentsat the Mediterranean stream (figure 3) Algal biomassaccounted for 137 plusmn 18 microg C cmminus2 on rocky habitat in theAndean stream while reached 162 plusmn 29 microg C cmminus2 onfine sediment and 23 plusmn 3 microg C cmminus2 for suspended algaein the Pampean stream In this stream macrophytes biomassaccounted for 58plusmn 27 g C mminus2
Nutrient enrichment produced an increase in algalbiomass in the three streams (figure 3 table 2) althoughthe responses varied in magnitude and differed betweencompartments The largest increase occurred on rocks inthe Mediterranean stream (163 on average peak of 273during spring) In the Andean stream algal biomass on rocksrapidly increased (62) and remained steady thereafter Thesmallest biomass increase was registered in the Pampeanstream particularly in the water column and for macrophytesbut biomass increased up to 49 on fine sediments The
6
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 2 Predictive-BACIPS of the effects of nutrient enrichment on the biomass of biological compartments in the three streams Thevalues shown correspond to the estimated coefficients of the linear models (ANCOVAs) When the control times BA was not significant(P gt 010) it was deleted from the model see section 2 for further details ns P gt 010 a P le 010 P le 005 P le 001 P le 0001
Variable n Control BA Control times BA
MediterraneanBacteria rocks 18 0404 0068 nsBacteria fine sediment 23 0590 0078 nsAlgae rocks 19 0495 0202a nsAlgae fine sediment 23 1082 0315 minus0732Invertebrates fine sediment 7 minus1011 minus0072 1660a
PampeanBacteria fine sediment 11 0751 0510a nsBacteria suspended 11 0954 minus0018 nsBact macrophytes 8 0396 0097 nsAlgae fine sediment 10 1106 0206 nsAlgae suspended 5 0274 minus0079 nsAlgae macrophyte 10 0575 minus0150 nsMacrophytes 11 1093 0249 nsInvertebrates fine sediment 10 minus0130 0787 nsInvertebrates macrophytes 6 minus0251 minus0521a nsFish CPUE 4 1082 8221 nsFish BPUE 4 0745 29552 ns
AndeanBacteria rocks 7 0666a 0299 nsBacteria fine sediment 6 0150 minus2631a nsAlgae rocks 14 0158 minus0101 0728Invertebrates rocks 13 0469 minus0269 nsInvertebrates fine sediment 13 0432 minus0388 nsFish density 10 1012 minus0442 nsFish biomass 10 1020 minus0464a ns
algal biomass responses to nutrients in the three streamswere strongly related with favorable conditions of lighttemperature and water flow (figure 3) The difference betweenthe extension of the respective windows of opportunitydetermined the steadiness of the response The Mediterraneanand Andean streams represented the maximum and minimumvariability in favorable conditions for primary producersrsquogrowth accordingly the coefficients of variation of thebiomass ratiorsquos change were the highest in the Mediterranean(CV = 16) and the lowest in the Andean (CV = 08)stream
Nutrient enrichment affected the composition of benthicalgal assemblages Some diatom taxa decreased (egAchnanthes biasolettiana Grunow (minus57) Achnanthesminutissima Kutzing (minus432) in the MediterraneanFragilaria ulna (Nitzsch) Lange-Bertalot var ulna (minus68)in the Pampean and Cocconeis placentula Ehrenberg(minus40) in the Andean stream) whereas others increased(eg Rhoicosphenia abbreviata (Ag) Lange-Bertalot (94)and Cocconeis placentula Ehrenberg (594) in theMediterranean Nitzschia frustulum (Kutzing) Grunow varfrustulum (3528) in the Pampean and Reimeria sinuata(Gregory) Kociolek and Stoermer (646) in the Andeanstream) Enrichment in the Pampean stream had no effect onthe macrophyte composition Ludwigia sp and Bacopa spdominated both the C and E reaches and were later replacedby the macroalga Spirogyra sp
35 Invertebrate biomass and composition
Invertebrate biomass before the enrichment ranged from5 to 750 microg DM cmminus2 in the Mediterranean and Andeanstreams and reached 1093 plusmn 311 microg DM cmminus2 in thesediments of the Pampean stream The effects of nutrientenrichment were generally non-significant but often larger(figure 4 table 2) and more pronounced in some habitatsthan others In the Pampean stream effects were not apparenton invertebrates occurring on macrophytes and were minor(3) on fine sediments Invertebrate biomass increased upto 155 in the fine sediments of the Mediterranean streamwhile no increase was observed in the Andean stream Theeffects on invertebrate biomass were the least variable inthe Mediterranean stream (per cent increase ratio CV = 09CV = 12 in the Andean and 27 in the Pampean)
Although the general enrichment effects on invertebratebiomass were weak some specific invertebrate taxa weredistinctly affected The grazer Ancylus fluviatilis increasedby 17 in the Mediterranean stream and the grazerCamelobaetidus increased by 927 in the Andean streamDensities of the scrapers Heleobia parchappei Orbigny(in fine sediments) and Hyalella curvispina Shoemaker(on macrophytes) increased in the Pampean stream (48and 762 respectively) Potamopyrgus antipodarum Gray(scraper) and Corbicula fluminea Muller (filterer) occurredonly in the respective fertilized reaches of the Mediterraneanand Pampean streams The Tubificidae (minus38) and theTanytarsini chironomidae (minus29) significantly decreased in
7
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
of 48 km2 on deep loess deposits mainly covered bytemperate grassland grazed by cattle although a quarter ofthe land is devoted to agriculture The climate is temperatecontinental resulting in wet springs and autumns The averageannual rainfall is 600ndash1200 mm and the average watertemperatures range from 10 C (June) to 24 C (January) Theexperiments were conducted in a third order section (34 42primeS59 04primeO) that was naturally devoid of riparian trees Thestreambed was composed of carbonate calcium precipitatesand deposits of silt and clay that re-suspend during stormepisodes (Rodrigues Capıtulo et al 2010) Primary producersare diverse and include suspended cells epipelic microalgaeand macrophytes
Tota is an Andean stream located at an altitude of2560 m in the Eastern Mountains of Colombia in BoyacaState The stream drains a 140 km2 basin on shale and clayApproximately 40 of the surface area of the basin is usedfor agriculture and pasture the rest is covered by secondaryforest and sparse vegetation The climate is cool and dry witha bimodal precipitation pattern temperatures showing littleseasonal variation The average annual rainfall is 730 mmand average water temperatures range from 11 to 13 CExperiments were conducted in a third order section wherethe riparian vegetation is composed of open forest dominatedby Alnus acuminata Kunth and Salix humboldtiana Willdwith a dense layer of Pennisetum clandestinum Hochst exChiov grass The streambed is composed of rocks cobblesand boulders and few deposits of sand and detritus Thoughlitter fall is relatively constant through the year detritusaccumulates during periods of low flow Macrophytes are rareor absent and epilithic and epipsammic algae are the mainprimary producers
22 Experimental manipulations
The experiment followed a BACIPS (beforendashafter controlndashimpact paired series) design in each stream we experimen-tally enhanced nutrient concentrations 2ndash4 fold over a 50 mreach (E enriched) whereas an upstream reach of similarmorphological and hydrological characteristics was kept asthe control (C) All reaches (C and E) were studied for 9ndash15months before the enrichment E reaches were then enrichedand all reaches studied for one year Nutrient amounts toadd to E reaches were calculated bi-weekly considering thebasal stream water nutrient concentrations (total ammonianitrate and phosphate) and discharge Nutrient solutions wereprepared with ammonium nitrate (NH4NO3) and ammoniumphosphate (NH4H2PO4) stored in 100 l tanks beside thestream bank and added continuously to the E reach Given thelow water velocity bags of fertilizer (12 N and 12 P) wereused to produce the necessary enrichment in the E reach ofthe Pampean stream The enrichment started in January 2004in the Mediterranean January 2007 in the Pampean and May2008 in the Andean stream
Ammonium nitrate and phosphate concentrations wereanalyzed in each of the stream reaches before and afterthe enrichment Water samples taken in triplicate werefiltered through 07 microm GFF filters (Whatman England)
and placed into 50 ml flasks for further analysis in thelaboratory Colorimetric methods were used for total ammonia(Solorzano 1969) phosphate (Murphy and Riley 1962) andnitrate (APHA 1989) determination
23 Biological analyses
The sampling was adapted to study the main compartmentspresent in each of the streams fine sediment and rockswere sampled in the Mediterranean and Andean streams finesediment the water column and macrophytes in the Pampeanstream All reaches were sampled monthly On each occasionwater temperature conductivity pH oxygen irradiance anddischarge were measured with portable meters and samplesto analyze nutrient concentrations and benthic organic matteraccumulation were collected
231 Rocky habitat Bacterial and algal biomass in theepilithon was measured from 1 cm2 ceramic tiles gluedonto rock slabs that were kept at a depth of 10ndash20 cm inriffle zones The samples were allowed to colonize for atleast two months before sampling On each occasion threetiles were taken for bacterial density estimates and preservedwith 4 formaldehyde and three more tiles were taken forchlorophyll-a concentration and kept frozen until analysisBacterial density was determined following Porter and Feig(1980) and chlorophyll-a concentration determined using themethod of Jeffrey and Humphrey (1975) Transformation intomicrobial biomass (microg carbon per cmminus2) was achieved byusing conversion factors of Frost et al (2005) for chlorophyll-aand Bratbak and Dundas (1984) for bacterial densities
The taxonomic composition of the diatom communitywas determined under the microscope Diatom communitieswere detached by sonication from glass tiles cleaned withsulfuric acid and hydrogen peroxide and mounted ontopermanent slides using Naphrax (refractive index of 174Brunel Microscopes Ltd England) About 400 frustules werecounted per sample and diatoms were identified to the lowesttaxonomic level possible using standard references (Krammerand Lange-Bertalot 1986ndash1991) and recent nomenclatureupdates
Invertebrates in the Andean stream were sampled directlyon cobbles using a Surber sampler with a 900 cm2 surfacearea and a 500 microm mesh size The samples (at least 5replicates) collected were fixed with 4 formaldehyde andinvertebrates were sorted under a dissecting microscope at400times Identification of invertebrates reached the genera leveland the species level when possible Biomass was calculatedas dry mass (70 C for 48 h) per square meter In theMediterranean stream the grazer Ancylus fluviatilis Mullerwas the most abundant invertebrate upon rocks ThereforeAncylus body mass was estimated as the average dry mass(70 C 48 h) of 40ndash100 individuals (corresponding to 10ndash20cobbles sampled) in C and E reaches during enrichmentexperiments There was no rocky substratum in the Pampeanstream
3
Environ Res Lett 8 (2013) 014002 J Artigas et al
232 Sediment habitat Epipsammic algae and bacteriawere sampled with plastic cylinders (43 cm in diameter2ndash4 cm depth) used as corers Three replicates were collectedat random at C and E reaches placed in plastic containers andsubsampled using an untapped syringe (12 cm in diameter)and equivalent to 1 g of sediment Samples were theneither preserved with 4 formaldehyde for bacterial countingor kept frozen to quantify chlorophyll-a The protocolsfor bacterial density and chlorophyll-a determination weresimilar to those used for the rocks Biomass results wereexpressed in microg carbon per cmminus2 of sediment
In the Mediterranean and Andean streams invertebratesin fine sediments were collected with plastic cores (5 cmin diameter 7ndash8 cm depth) Three core replicates werecollected per reach (C and E) and stream type In the Pampeanstream an Ekman dredge sampler (100 cm2 surface area) wasused to sample sediments (also triplicate samples) Sampleswere immediately filtered through 500 microm sieves and fixedwith formaldehyde Invertebrate identification and biomassdetermination procedures were similar to those used for therock assemblages
233 Water column In the Pampean stream suspendedchlorophyll-a was analyzed from 1 l water samples (3replicates per reach and occasion) Water was filtered throughGFF filters (07 microm Whatman England) and the filterswere analyzed for chlorophyll as the extracts from rocksBacterial density was estimated in 10 ml water subsamples(3 replicates) following the methodology described in Porterand Feig (1980) Algal and bacterial biomass in thewater column was estimated through the conversion factorsdescribed before and expressed by square meter of streambed
234 Macrophytes Macrophytes were surveyed bimonthlyduring the cold season and monthly during the warm seasonat the Pampean stream Macrophyte composition and coverwere determined from regularly spaced transects Macrophytesamples (3ndash10 replicates) were obtained at random in the Cand E reaches with a 649 cm2 square metal frame and stored in500 ml plastic bags Plants were rinsed with tap water sortedinto species and invertebrates removed Plants were dried(70 C to a constant dry weight) to obtain dry mass which wasexpressed in g mminus2 Invertebrates removed from macrophyteswere filtered through 500microm sieves fixed with formaldehydeidentified and counted Identification of invertebrates reachedthe genera level and the species level when possible
235 Fish Fish abundance and biomass in the Pampean andAndean streams were estimated bimonthly using multiple-pass sampling (at least four passes per occasion) afterblocking the C and E reaches with nets Fish density wastoo low in the Mediterranean stream (Mas-Martı et al 2010)to allow comparative sampling between reaches Samplingwas performed with electrofishing (EN ISO 140112003standard) In the Pampean stream sampling was also donewith seines when high water conductivity precluded the useof electrofishing All fish sampled were counted identified
measured (standard length) and then returned to the waterOnly a small subsample of about 20 individuals wasused to determine fish condition (body weight and lengthGarcıa-Berthou and Moreno-Amich 1993) Fish abundanceand biomass were expressed as catch per unit of effort(CPUE = fish numberarea sampled) and biomass per unit ofeffort (BPUE = fish biomassarea sampled) respectively
24 Data analysis
The results of the three enrichment experiments wereanalyzed following the predictive-BACIPS approach (Benceet al 1996) This method compares the relationship betweenthe values in the C and E reaches before and after the start ofthe experiment Although beforendashafter controlndashimpact pairedseries (BACIPS) designs are often analyzed with ANOVA theuse of statistical techniques with control sites as covariatesas in the predictive-BACIPS has the advantage of allowingthe effect size (ie the enrichment effects) to vary withthe magnitude of the control value and with environmentalconditions ie does not require the standard assumption ofadditivity (Bence et al 1996 Stewart-Oaten and Bence 2001)
For the predictive-BACIPS approach we used analysis ofcovariance (ANCOVA) of the values of the response variable(eg bacterial biomass) in the E reaches using values inC (control) as a covariate and the periods beforeafter thestart of the treatment (BA) as a binary factor ANCOVA wasfirst used to test interactions between the covariate (ie Cvalues) and the binary factor (BA) Significant interactionsindicate that slopes between both variables (correspondingto C and E values) were not equal before and after thestart of the treatment (Garcıa-Berthou and Moreno-Amich1993) In that case the treatment effect would depend onthe magnitude of the control values When the C times BAinteraction was not significant (P gt 010) it was removedfrom the model to improve the statistical power and astandard ANCOVA design was used (ie homogeneous slopeswere assumed Garcıa-Berthou and Moreno-Amich 1993)The proportion of variance (often termed eta squared η2)explained by the BA factor plus the C times BA interactionwas computed as a measure of the effect size of enrichmentBence et al (1996) recommend that emphasis in BACIPSdesigns is mainly placed on effect sizes rather than on Pvalues Average η2 values in the biological compartments(bacteria algae macrophytes water column invertebratesand fish) in each stream were calculated as an estimateof the magnitude of transferral of the nutrient enrichmentthrough the food web All data were analyzed with the Rsoftware (R Development Core Team 2012) Note that thedefault packages in R use sequential (ie type I) sum ofsquares (Hector et al 2010) and that the lsquocontrolrsquo covariatewas always tested first in the models Quantitative variables(response variables and covariates) were log10-transformedbefore statistical analyses in order to improve assumptions ofthe linear models (normality homoscedasticity and linearity)
An important assumption of BACIPS designs andregression of time-related variables in general is the lackof serial correlation (Bence et al 1996 Stewart-Oaten and
4
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 1 Environmental characteristics and organic matter availability in the Mediterranean Pampean and Andean streams Values ofwater temperature (C) light (micromol photons mminus2 sminus1) and discharge (l sminus1) during enrichment experiments are represented Windows ofecological opportunity (marked in gray right plot) were determined after considering favorable conditions for primary producers microbialheterotrophs and consumer development
Bence 2001) We used the BreuschndashGodfrey test to testwhether serial correlation was present in our data withserial correlation up to four order using the lsquobgtestrsquo functionof package lsquolmtestrsquo (Zeileis and Hothorn 2002) in R TheBreuschndashGodfrey test is more applicable and often morepowerful than the more widely used DurbinndashWatson testand other methods (Godfrey and Tremayne 2005) Only 1of the 23 analyses (combinations of variables and streams)showed significant serial correlation (P lt 005) Similarlyonly one case of the 23 raw variables showed serial correlation(P lt 005) in the enriched zones probably because samplingfrequency was low (about monthly) and equally spaced(Stewart-Oaten and Bence 2001) Since our time series wererelatively short (sample size lt 50) and serial correlationwas almost always non-significant we did not correct forautocorrelation as recommended by Bence (1995)
The effect of enrichment on biomass of the differenttrophic levels was expressed as the per cent ratio of biomasschange in the E reach with respect to the C reach The ratio ofbiomass change was therefore calculated as 100 (enriched minuscontrol)control This ratio was also used to calculateenrichment effects on diatom and invertebrate abundancesThe ratiorsquos coefficient of variation (CV) during the enrichmentwas useful to determine the potential fluctuations in biomassresponse caused by the treatment Windows of opportunitywere defined as time periods when no factor beyondnutrients exerts significant limitation on primary productionand consumers in the stream These periods were identifiedin the Mediterranean Pampean and Andean streams afteranalyzing the best combinations of water temperature lightand discharge
3 Results
31 Physical and chemical characteristics of the study sites
The Mediterranean and Pampean streams had large seasonalvariations in water temperature and light availability whilein contrast the seasonal variation in the Andean stream wasmuch lower (figure 1) Water flow also followed differentpatterns in the three systems Variability was high in theAndean stream (from 68 to 1800 l sminus1) and Mediterraneanstream (from 0 to 1400 l sminus1) and very low in the Pampean(from 0 to 40 l sminus1) Low flow values included up to twomonths of summer drought in the Mediterranean stream butwater remained in the channel in the Pampean because of itsvery low slope
Basal nutrient concentrations before the enrichment(table 1) were the highest in the Pampean stream (140 microg lminus1
SRP and 700 microg lminus1 DIN) characterized by low phosphoruscontent in the Mediterranean stream (14 microg lminus1 SRP430 microg lminus1 DIN) and with low DIN concentrations in theAndean stream (26 microg lminus1 SRP and 110 microg Lminus1 DIN) Theenrichment caused SRP to increasetimes2ndash4 and DIN to increasetimes16ndash28 (table 1) The enrichment caused the decrease of theNP ratios in the Mediterranean and Pampean streams but didnot greatly alter those of the Andean (table 1)
32 Windows of opportunity
Windows of ecological opportunity for primary producersmicrobial heterotrophs and consumers differed between
5
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 1 Water nutrient concentrations of soluble reactive phosphorus (SRP) and dissolved inorganic nitrogen (DIN sum ofammonia+ nitrate+ nitrite forms) in the C and E reaches of the Mediterranean Pampean and Andean streams Values are means andstandard errors of nutrients measured during the pre-enrichment (before) and enrichment (after) periods and are accompanied by NP molarratio calculations The increment (1) nutrient concentration during enrichment is also shown for each of the streams
Control reach Enriched reach 1 nutrient concentration
Mediterranean Before SRP (microg lminus1) 14plusmn 18 16plusmn 21DIN (microg lminus1) 432plusmn 438 523plusmn 491NP 68 71
After SRP (microg lminus1) 10plusmn 9 30plusmn 13 times3DIN (microg lminus1) 389plusmn 495 765plusmn 736 times2NP 84 56
Pampean Before SRP (microg lminus1) 143plusmn 17 173plusmn 31DIN (microg lminus1) 683plusmn 39 1093plusmn 228NP 11 14
After SRP (microg lminus1) 93plusmn 3 361plusmn 29 times4DIN (microg lminus1) 528plusmn 136 828plusmn 197 times16NP 13 5
Andean Before SRP (microg lminus1) 26plusmn 5 28plusmn 5DIN (microg lminus1) 55plusmn 21 78plusmn 23NP 5 6
After SRP (microg lminus1) 53plusmn 4 105plusmn 13 times2DIN (microg lminus1) 114plusmn 59 323plusmn 85 times28NP 5 7
streams in terms of duration and frequency (figure 1)Windows of opportunity in the Mediterranean stream werenarrow and limited to late springndashearly summer (moderatetemperature and light availability) and late summerndashearlyautumn periods (moderate temperatures and abundant al-lochthonous OM inputs) The greatest overlap of favor-able environmental conditions for biological developmentoccurred in the Pampean stream In this system higher lightelevated temperatures and low discharge values coincidedfrom winter to summer (SeptemberndashMarch) The absenceof riparian vegetation reduced allochthonous OM inputsin this stream but authocthonous OM was produced byalgae and macrophytes The Andean stream also had longfavorable periods of temperature light and allochthonous OMinputs only interrupted by high discharge periods betweenAprilndashMay and OctoberndashNovember The high flow periodsin this stream caused a momentary biomass decrease of allbiological groups The predictive-BACIPS approach (table 2)suggested that 10 out of 23 models had significant treatmentseffects (or interactions) and that the explained variance wasoften high (figure 6)
33 Bacterial biomass
Bacterial biomass before the enrichment ranged from 2to 16 microg C cmminus2 on rocks and fine sediment in theMediterranean and Andean streams from 5 to 700microg C cmminus2
on sediments 004ndash15 microg C cmminus2 on macrophytes and1ndash36microg C cmminus2 in the water column for the Pampean streamThe nutrient enrichment resulted in an increase in bacterialbiomass in the Mediterranean (rocks and fine sediment) andPampean (fine sediment) streams but not in the Andeanstream (figure 2 table 2) The largest increase in bacterial
biomass after enrichment (calculated as the biomass changeratio) occurred in the Mediterranean stream (65ndash80 overallincrease) In the Pampean stream bacterial biomass increasedbetween 20 and 30 in fine sediment and macrophytesbut decreased in the water column (10ndash20) Water flowconditions temperature and light availability covaried withbacterial biomass in the E reaches of the Mediterranean andPampean streams The temporal variability of the biomasschange ratio was the highest in the Mediterranean stream(coefficient of variation CV = 24) due to the episodichigher biomass between late spring and early summer in theenrichment period
34 Algal and macrophytic biomass and composition
Algal biomass before the enrichment averaged 205 plusmn31 microg C cmminus2 on rocks and 69plusmn11 microg C cmminus2 on sedimentsat the Mediterranean stream (figure 3) Algal biomassaccounted for 137 plusmn 18 microg C cmminus2 on rocky habitat in theAndean stream while reached 162 plusmn 29 microg C cmminus2 onfine sediment and 23 plusmn 3 microg C cmminus2 for suspended algaein the Pampean stream In this stream macrophytes biomassaccounted for 58plusmn 27 g C mminus2
Nutrient enrichment produced an increase in algalbiomass in the three streams (figure 3 table 2) althoughthe responses varied in magnitude and differed betweencompartments The largest increase occurred on rocks inthe Mediterranean stream (163 on average peak of 273during spring) In the Andean stream algal biomass on rocksrapidly increased (62) and remained steady thereafter Thesmallest biomass increase was registered in the Pampeanstream particularly in the water column and for macrophytesbut biomass increased up to 49 on fine sediments The
6
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 2 Predictive-BACIPS of the effects of nutrient enrichment on the biomass of biological compartments in the three streams Thevalues shown correspond to the estimated coefficients of the linear models (ANCOVAs) When the control times BA was not significant(P gt 010) it was deleted from the model see section 2 for further details ns P gt 010 a P le 010 P le 005 P le 001 P le 0001
Variable n Control BA Control times BA
MediterraneanBacteria rocks 18 0404 0068 nsBacteria fine sediment 23 0590 0078 nsAlgae rocks 19 0495 0202a nsAlgae fine sediment 23 1082 0315 minus0732Invertebrates fine sediment 7 minus1011 minus0072 1660a
PampeanBacteria fine sediment 11 0751 0510a nsBacteria suspended 11 0954 minus0018 nsBact macrophytes 8 0396 0097 nsAlgae fine sediment 10 1106 0206 nsAlgae suspended 5 0274 minus0079 nsAlgae macrophyte 10 0575 minus0150 nsMacrophytes 11 1093 0249 nsInvertebrates fine sediment 10 minus0130 0787 nsInvertebrates macrophytes 6 minus0251 minus0521a nsFish CPUE 4 1082 8221 nsFish BPUE 4 0745 29552 ns
AndeanBacteria rocks 7 0666a 0299 nsBacteria fine sediment 6 0150 minus2631a nsAlgae rocks 14 0158 minus0101 0728Invertebrates rocks 13 0469 minus0269 nsInvertebrates fine sediment 13 0432 minus0388 nsFish density 10 1012 minus0442 nsFish biomass 10 1020 minus0464a ns
algal biomass responses to nutrients in the three streamswere strongly related with favorable conditions of lighttemperature and water flow (figure 3) The difference betweenthe extension of the respective windows of opportunitydetermined the steadiness of the response The Mediterraneanand Andean streams represented the maximum and minimumvariability in favorable conditions for primary producersrsquogrowth accordingly the coefficients of variation of thebiomass ratiorsquos change were the highest in the Mediterranean(CV = 16) and the lowest in the Andean (CV = 08)stream
Nutrient enrichment affected the composition of benthicalgal assemblages Some diatom taxa decreased (egAchnanthes biasolettiana Grunow (minus57) Achnanthesminutissima Kutzing (minus432) in the MediterraneanFragilaria ulna (Nitzsch) Lange-Bertalot var ulna (minus68)in the Pampean and Cocconeis placentula Ehrenberg(minus40) in the Andean stream) whereas others increased(eg Rhoicosphenia abbreviata (Ag) Lange-Bertalot (94)and Cocconeis placentula Ehrenberg (594) in theMediterranean Nitzschia frustulum (Kutzing) Grunow varfrustulum (3528) in the Pampean and Reimeria sinuata(Gregory) Kociolek and Stoermer (646) in the Andeanstream) Enrichment in the Pampean stream had no effect onthe macrophyte composition Ludwigia sp and Bacopa spdominated both the C and E reaches and were later replacedby the macroalga Spirogyra sp
35 Invertebrate biomass and composition
Invertebrate biomass before the enrichment ranged from5 to 750 microg DM cmminus2 in the Mediterranean and Andeanstreams and reached 1093 plusmn 311 microg DM cmminus2 in thesediments of the Pampean stream The effects of nutrientenrichment were generally non-significant but often larger(figure 4 table 2) and more pronounced in some habitatsthan others In the Pampean stream effects were not apparenton invertebrates occurring on macrophytes and were minor(3) on fine sediments Invertebrate biomass increased upto 155 in the fine sediments of the Mediterranean streamwhile no increase was observed in the Andean stream Theeffects on invertebrate biomass were the least variable inthe Mediterranean stream (per cent increase ratio CV = 09CV = 12 in the Andean and 27 in the Pampean)
Although the general enrichment effects on invertebratebiomass were weak some specific invertebrate taxa weredistinctly affected The grazer Ancylus fluviatilis increasedby 17 in the Mediterranean stream and the grazerCamelobaetidus increased by 927 in the Andean streamDensities of the scrapers Heleobia parchappei Orbigny(in fine sediments) and Hyalella curvispina Shoemaker(on macrophytes) increased in the Pampean stream (48and 762 respectively) Potamopyrgus antipodarum Gray(scraper) and Corbicula fluminea Muller (filterer) occurredonly in the respective fertilized reaches of the Mediterraneanand Pampean streams The Tubificidae (minus38) and theTanytarsini chironomidae (minus29) significantly decreased in
7
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
232 Sediment habitat Epipsammic algae and bacteriawere sampled with plastic cylinders (43 cm in diameter2ndash4 cm depth) used as corers Three replicates were collectedat random at C and E reaches placed in plastic containers andsubsampled using an untapped syringe (12 cm in diameter)and equivalent to 1 g of sediment Samples were theneither preserved with 4 formaldehyde for bacterial countingor kept frozen to quantify chlorophyll-a The protocolsfor bacterial density and chlorophyll-a determination weresimilar to those used for the rocks Biomass results wereexpressed in microg carbon per cmminus2 of sediment
In the Mediterranean and Andean streams invertebratesin fine sediments were collected with plastic cores (5 cmin diameter 7ndash8 cm depth) Three core replicates werecollected per reach (C and E) and stream type In the Pampeanstream an Ekman dredge sampler (100 cm2 surface area) wasused to sample sediments (also triplicate samples) Sampleswere immediately filtered through 500 microm sieves and fixedwith formaldehyde Invertebrate identification and biomassdetermination procedures were similar to those used for therock assemblages
233 Water column In the Pampean stream suspendedchlorophyll-a was analyzed from 1 l water samples (3replicates per reach and occasion) Water was filtered throughGFF filters (07 microm Whatman England) and the filterswere analyzed for chlorophyll as the extracts from rocksBacterial density was estimated in 10 ml water subsamples(3 replicates) following the methodology described in Porterand Feig (1980) Algal and bacterial biomass in thewater column was estimated through the conversion factorsdescribed before and expressed by square meter of streambed
234 Macrophytes Macrophytes were surveyed bimonthlyduring the cold season and monthly during the warm seasonat the Pampean stream Macrophyte composition and coverwere determined from regularly spaced transects Macrophytesamples (3ndash10 replicates) were obtained at random in the Cand E reaches with a 649 cm2 square metal frame and stored in500 ml plastic bags Plants were rinsed with tap water sortedinto species and invertebrates removed Plants were dried(70 C to a constant dry weight) to obtain dry mass which wasexpressed in g mminus2 Invertebrates removed from macrophyteswere filtered through 500microm sieves fixed with formaldehydeidentified and counted Identification of invertebrates reachedthe genera level and the species level when possible
235 Fish Fish abundance and biomass in the Pampean andAndean streams were estimated bimonthly using multiple-pass sampling (at least four passes per occasion) afterblocking the C and E reaches with nets Fish density wastoo low in the Mediterranean stream (Mas-Martı et al 2010)to allow comparative sampling between reaches Samplingwas performed with electrofishing (EN ISO 140112003standard) In the Pampean stream sampling was also donewith seines when high water conductivity precluded the useof electrofishing All fish sampled were counted identified
measured (standard length) and then returned to the waterOnly a small subsample of about 20 individuals wasused to determine fish condition (body weight and lengthGarcıa-Berthou and Moreno-Amich 1993) Fish abundanceand biomass were expressed as catch per unit of effort(CPUE = fish numberarea sampled) and biomass per unit ofeffort (BPUE = fish biomassarea sampled) respectively
24 Data analysis
The results of the three enrichment experiments wereanalyzed following the predictive-BACIPS approach (Benceet al 1996) This method compares the relationship betweenthe values in the C and E reaches before and after the start ofthe experiment Although beforendashafter controlndashimpact pairedseries (BACIPS) designs are often analyzed with ANOVA theuse of statistical techniques with control sites as covariatesas in the predictive-BACIPS has the advantage of allowingthe effect size (ie the enrichment effects) to vary withthe magnitude of the control value and with environmentalconditions ie does not require the standard assumption ofadditivity (Bence et al 1996 Stewart-Oaten and Bence 2001)
For the predictive-BACIPS approach we used analysis ofcovariance (ANCOVA) of the values of the response variable(eg bacterial biomass) in the E reaches using values inC (control) as a covariate and the periods beforeafter thestart of the treatment (BA) as a binary factor ANCOVA wasfirst used to test interactions between the covariate (ie Cvalues) and the binary factor (BA) Significant interactionsindicate that slopes between both variables (correspondingto C and E values) were not equal before and after thestart of the treatment (Garcıa-Berthou and Moreno-Amich1993) In that case the treatment effect would depend onthe magnitude of the control values When the C times BAinteraction was not significant (P gt 010) it was removedfrom the model to improve the statistical power and astandard ANCOVA design was used (ie homogeneous slopeswere assumed Garcıa-Berthou and Moreno-Amich 1993)The proportion of variance (often termed eta squared η2)explained by the BA factor plus the C times BA interactionwas computed as a measure of the effect size of enrichmentBence et al (1996) recommend that emphasis in BACIPSdesigns is mainly placed on effect sizes rather than on Pvalues Average η2 values in the biological compartments(bacteria algae macrophytes water column invertebratesand fish) in each stream were calculated as an estimateof the magnitude of transferral of the nutrient enrichmentthrough the food web All data were analyzed with the Rsoftware (R Development Core Team 2012) Note that thedefault packages in R use sequential (ie type I) sum ofsquares (Hector et al 2010) and that the lsquocontrolrsquo covariatewas always tested first in the models Quantitative variables(response variables and covariates) were log10-transformedbefore statistical analyses in order to improve assumptions ofthe linear models (normality homoscedasticity and linearity)
An important assumption of BACIPS designs andregression of time-related variables in general is the lackof serial correlation (Bence et al 1996 Stewart-Oaten and
4
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 1 Environmental characteristics and organic matter availability in the Mediterranean Pampean and Andean streams Values ofwater temperature (C) light (micromol photons mminus2 sminus1) and discharge (l sminus1) during enrichment experiments are represented Windows ofecological opportunity (marked in gray right plot) were determined after considering favorable conditions for primary producers microbialheterotrophs and consumer development
Bence 2001) We used the BreuschndashGodfrey test to testwhether serial correlation was present in our data withserial correlation up to four order using the lsquobgtestrsquo functionof package lsquolmtestrsquo (Zeileis and Hothorn 2002) in R TheBreuschndashGodfrey test is more applicable and often morepowerful than the more widely used DurbinndashWatson testand other methods (Godfrey and Tremayne 2005) Only 1of the 23 analyses (combinations of variables and streams)showed significant serial correlation (P lt 005) Similarlyonly one case of the 23 raw variables showed serial correlation(P lt 005) in the enriched zones probably because samplingfrequency was low (about monthly) and equally spaced(Stewart-Oaten and Bence 2001) Since our time series wererelatively short (sample size lt 50) and serial correlationwas almost always non-significant we did not correct forautocorrelation as recommended by Bence (1995)
The effect of enrichment on biomass of the differenttrophic levels was expressed as the per cent ratio of biomasschange in the E reach with respect to the C reach The ratio ofbiomass change was therefore calculated as 100 (enriched minuscontrol)control This ratio was also used to calculateenrichment effects on diatom and invertebrate abundancesThe ratiorsquos coefficient of variation (CV) during the enrichmentwas useful to determine the potential fluctuations in biomassresponse caused by the treatment Windows of opportunitywere defined as time periods when no factor beyondnutrients exerts significant limitation on primary productionand consumers in the stream These periods were identifiedin the Mediterranean Pampean and Andean streams afteranalyzing the best combinations of water temperature lightand discharge
3 Results
31 Physical and chemical characteristics of the study sites
The Mediterranean and Pampean streams had large seasonalvariations in water temperature and light availability whilein contrast the seasonal variation in the Andean stream wasmuch lower (figure 1) Water flow also followed differentpatterns in the three systems Variability was high in theAndean stream (from 68 to 1800 l sminus1) and Mediterraneanstream (from 0 to 1400 l sminus1) and very low in the Pampean(from 0 to 40 l sminus1) Low flow values included up to twomonths of summer drought in the Mediterranean stream butwater remained in the channel in the Pampean because of itsvery low slope
Basal nutrient concentrations before the enrichment(table 1) were the highest in the Pampean stream (140 microg lminus1
SRP and 700 microg lminus1 DIN) characterized by low phosphoruscontent in the Mediterranean stream (14 microg lminus1 SRP430 microg lminus1 DIN) and with low DIN concentrations in theAndean stream (26 microg lminus1 SRP and 110 microg Lminus1 DIN) Theenrichment caused SRP to increasetimes2ndash4 and DIN to increasetimes16ndash28 (table 1) The enrichment caused the decrease of theNP ratios in the Mediterranean and Pampean streams but didnot greatly alter those of the Andean (table 1)
32 Windows of opportunity
Windows of ecological opportunity for primary producersmicrobial heterotrophs and consumers differed between
5
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 1 Water nutrient concentrations of soluble reactive phosphorus (SRP) and dissolved inorganic nitrogen (DIN sum ofammonia+ nitrate+ nitrite forms) in the C and E reaches of the Mediterranean Pampean and Andean streams Values are means andstandard errors of nutrients measured during the pre-enrichment (before) and enrichment (after) periods and are accompanied by NP molarratio calculations The increment (1) nutrient concentration during enrichment is also shown for each of the streams
Control reach Enriched reach 1 nutrient concentration
Mediterranean Before SRP (microg lminus1) 14plusmn 18 16plusmn 21DIN (microg lminus1) 432plusmn 438 523plusmn 491NP 68 71
After SRP (microg lminus1) 10plusmn 9 30plusmn 13 times3DIN (microg lminus1) 389plusmn 495 765plusmn 736 times2NP 84 56
Pampean Before SRP (microg lminus1) 143plusmn 17 173plusmn 31DIN (microg lminus1) 683plusmn 39 1093plusmn 228NP 11 14
After SRP (microg lminus1) 93plusmn 3 361plusmn 29 times4DIN (microg lminus1) 528plusmn 136 828plusmn 197 times16NP 13 5
Andean Before SRP (microg lminus1) 26plusmn 5 28plusmn 5DIN (microg lminus1) 55plusmn 21 78plusmn 23NP 5 6
After SRP (microg lminus1) 53plusmn 4 105plusmn 13 times2DIN (microg lminus1) 114plusmn 59 323plusmn 85 times28NP 5 7
streams in terms of duration and frequency (figure 1)Windows of opportunity in the Mediterranean stream werenarrow and limited to late springndashearly summer (moderatetemperature and light availability) and late summerndashearlyautumn periods (moderate temperatures and abundant al-lochthonous OM inputs) The greatest overlap of favor-able environmental conditions for biological developmentoccurred in the Pampean stream In this system higher lightelevated temperatures and low discharge values coincidedfrom winter to summer (SeptemberndashMarch) The absenceof riparian vegetation reduced allochthonous OM inputsin this stream but authocthonous OM was produced byalgae and macrophytes The Andean stream also had longfavorable periods of temperature light and allochthonous OMinputs only interrupted by high discharge periods betweenAprilndashMay and OctoberndashNovember The high flow periodsin this stream caused a momentary biomass decrease of allbiological groups The predictive-BACIPS approach (table 2)suggested that 10 out of 23 models had significant treatmentseffects (or interactions) and that the explained variance wasoften high (figure 6)
33 Bacterial biomass
Bacterial biomass before the enrichment ranged from 2to 16 microg C cmminus2 on rocks and fine sediment in theMediterranean and Andean streams from 5 to 700microg C cmminus2
on sediments 004ndash15 microg C cmminus2 on macrophytes and1ndash36microg C cmminus2 in the water column for the Pampean streamThe nutrient enrichment resulted in an increase in bacterialbiomass in the Mediterranean (rocks and fine sediment) andPampean (fine sediment) streams but not in the Andeanstream (figure 2 table 2) The largest increase in bacterial
biomass after enrichment (calculated as the biomass changeratio) occurred in the Mediterranean stream (65ndash80 overallincrease) In the Pampean stream bacterial biomass increasedbetween 20 and 30 in fine sediment and macrophytesbut decreased in the water column (10ndash20) Water flowconditions temperature and light availability covaried withbacterial biomass in the E reaches of the Mediterranean andPampean streams The temporal variability of the biomasschange ratio was the highest in the Mediterranean stream(coefficient of variation CV = 24) due to the episodichigher biomass between late spring and early summer in theenrichment period
34 Algal and macrophytic biomass and composition
Algal biomass before the enrichment averaged 205 plusmn31 microg C cmminus2 on rocks and 69plusmn11 microg C cmminus2 on sedimentsat the Mediterranean stream (figure 3) Algal biomassaccounted for 137 plusmn 18 microg C cmminus2 on rocky habitat in theAndean stream while reached 162 plusmn 29 microg C cmminus2 onfine sediment and 23 plusmn 3 microg C cmminus2 for suspended algaein the Pampean stream In this stream macrophytes biomassaccounted for 58plusmn 27 g C mminus2
Nutrient enrichment produced an increase in algalbiomass in the three streams (figure 3 table 2) althoughthe responses varied in magnitude and differed betweencompartments The largest increase occurred on rocks inthe Mediterranean stream (163 on average peak of 273during spring) In the Andean stream algal biomass on rocksrapidly increased (62) and remained steady thereafter Thesmallest biomass increase was registered in the Pampeanstream particularly in the water column and for macrophytesbut biomass increased up to 49 on fine sediments The
6
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 2 Predictive-BACIPS of the effects of nutrient enrichment on the biomass of biological compartments in the three streams Thevalues shown correspond to the estimated coefficients of the linear models (ANCOVAs) When the control times BA was not significant(P gt 010) it was deleted from the model see section 2 for further details ns P gt 010 a P le 010 P le 005 P le 001 P le 0001
Variable n Control BA Control times BA
MediterraneanBacteria rocks 18 0404 0068 nsBacteria fine sediment 23 0590 0078 nsAlgae rocks 19 0495 0202a nsAlgae fine sediment 23 1082 0315 minus0732Invertebrates fine sediment 7 minus1011 minus0072 1660a
PampeanBacteria fine sediment 11 0751 0510a nsBacteria suspended 11 0954 minus0018 nsBact macrophytes 8 0396 0097 nsAlgae fine sediment 10 1106 0206 nsAlgae suspended 5 0274 minus0079 nsAlgae macrophyte 10 0575 minus0150 nsMacrophytes 11 1093 0249 nsInvertebrates fine sediment 10 minus0130 0787 nsInvertebrates macrophytes 6 minus0251 minus0521a nsFish CPUE 4 1082 8221 nsFish BPUE 4 0745 29552 ns
AndeanBacteria rocks 7 0666a 0299 nsBacteria fine sediment 6 0150 minus2631a nsAlgae rocks 14 0158 minus0101 0728Invertebrates rocks 13 0469 minus0269 nsInvertebrates fine sediment 13 0432 minus0388 nsFish density 10 1012 minus0442 nsFish biomass 10 1020 minus0464a ns
algal biomass responses to nutrients in the three streamswere strongly related with favorable conditions of lighttemperature and water flow (figure 3) The difference betweenthe extension of the respective windows of opportunitydetermined the steadiness of the response The Mediterraneanand Andean streams represented the maximum and minimumvariability in favorable conditions for primary producersrsquogrowth accordingly the coefficients of variation of thebiomass ratiorsquos change were the highest in the Mediterranean(CV = 16) and the lowest in the Andean (CV = 08)stream
Nutrient enrichment affected the composition of benthicalgal assemblages Some diatom taxa decreased (egAchnanthes biasolettiana Grunow (minus57) Achnanthesminutissima Kutzing (minus432) in the MediterraneanFragilaria ulna (Nitzsch) Lange-Bertalot var ulna (minus68)in the Pampean and Cocconeis placentula Ehrenberg(minus40) in the Andean stream) whereas others increased(eg Rhoicosphenia abbreviata (Ag) Lange-Bertalot (94)and Cocconeis placentula Ehrenberg (594) in theMediterranean Nitzschia frustulum (Kutzing) Grunow varfrustulum (3528) in the Pampean and Reimeria sinuata(Gregory) Kociolek and Stoermer (646) in the Andeanstream) Enrichment in the Pampean stream had no effect onthe macrophyte composition Ludwigia sp and Bacopa spdominated both the C and E reaches and were later replacedby the macroalga Spirogyra sp
35 Invertebrate biomass and composition
Invertebrate biomass before the enrichment ranged from5 to 750 microg DM cmminus2 in the Mediterranean and Andeanstreams and reached 1093 plusmn 311 microg DM cmminus2 in thesediments of the Pampean stream The effects of nutrientenrichment were generally non-significant but often larger(figure 4 table 2) and more pronounced in some habitatsthan others In the Pampean stream effects were not apparenton invertebrates occurring on macrophytes and were minor(3) on fine sediments Invertebrate biomass increased upto 155 in the fine sediments of the Mediterranean streamwhile no increase was observed in the Andean stream Theeffects on invertebrate biomass were the least variable inthe Mediterranean stream (per cent increase ratio CV = 09CV = 12 in the Andean and 27 in the Pampean)
Although the general enrichment effects on invertebratebiomass were weak some specific invertebrate taxa weredistinctly affected The grazer Ancylus fluviatilis increasedby 17 in the Mediterranean stream and the grazerCamelobaetidus increased by 927 in the Andean streamDensities of the scrapers Heleobia parchappei Orbigny(in fine sediments) and Hyalella curvispina Shoemaker(on macrophytes) increased in the Pampean stream (48and 762 respectively) Potamopyrgus antipodarum Gray(scraper) and Corbicula fluminea Muller (filterer) occurredonly in the respective fertilized reaches of the Mediterraneanand Pampean streams The Tubificidae (minus38) and theTanytarsini chironomidae (minus29) significantly decreased in
7
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 1 Environmental characteristics and organic matter availability in the Mediterranean Pampean and Andean streams Values ofwater temperature (C) light (micromol photons mminus2 sminus1) and discharge (l sminus1) during enrichment experiments are represented Windows ofecological opportunity (marked in gray right plot) were determined after considering favorable conditions for primary producers microbialheterotrophs and consumer development
Bence 2001) We used the BreuschndashGodfrey test to testwhether serial correlation was present in our data withserial correlation up to four order using the lsquobgtestrsquo functionof package lsquolmtestrsquo (Zeileis and Hothorn 2002) in R TheBreuschndashGodfrey test is more applicable and often morepowerful than the more widely used DurbinndashWatson testand other methods (Godfrey and Tremayne 2005) Only 1of the 23 analyses (combinations of variables and streams)showed significant serial correlation (P lt 005) Similarlyonly one case of the 23 raw variables showed serial correlation(P lt 005) in the enriched zones probably because samplingfrequency was low (about monthly) and equally spaced(Stewart-Oaten and Bence 2001) Since our time series wererelatively short (sample size lt 50) and serial correlationwas almost always non-significant we did not correct forautocorrelation as recommended by Bence (1995)
The effect of enrichment on biomass of the differenttrophic levels was expressed as the per cent ratio of biomasschange in the E reach with respect to the C reach The ratio ofbiomass change was therefore calculated as 100 (enriched minuscontrol)control This ratio was also used to calculateenrichment effects on diatom and invertebrate abundancesThe ratiorsquos coefficient of variation (CV) during the enrichmentwas useful to determine the potential fluctuations in biomassresponse caused by the treatment Windows of opportunitywere defined as time periods when no factor beyondnutrients exerts significant limitation on primary productionand consumers in the stream These periods were identifiedin the Mediterranean Pampean and Andean streams afteranalyzing the best combinations of water temperature lightand discharge
3 Results
31 Physical and chemical characteristics of the study sites
The Mediterranean and Pampean streams had large seasonalvariations in water temperature and light availability whilein contrast the seasonal variation in the Andean stream wasmuch lower (figure 1) Water flow also followed differentpatterns in the three systems Variability was high in theAndean stream (from 68 to 1800 l sminus1) and Mediterraneanstream (from 0 to 1400 l sminus1) and very low in the Pampean(from 0 to 40 l sminus1) Low flow values included up to twomonths of summer drought in the Mediterranean stream butwater remained in the channel in the Pampean because of itsvery low slope
Basal nutrient concentrations before the enrichment(table 1) were the highest in the Pampean stream (140 microg lminus1
SRP and 700 microg lminus1 DIN) characterized by low phosphoruscontent in the Mediterranean stream (14 microg lminus1 SRP430 microg lminus1 DIN) and with low DIN concentrations in theAndean stream (26 microg lminus1 SRP and 110 microg Lminus1 DIN) Theenrichment caused SRP to increasetimes2ndash4 and DIN to increasetimes16ndash28 (table 1) The enrichment caused the decrease of theNP ratios in the Mediterranean and Pampean streams but didnot greatly alter those of the Andean (table 1)
32 Windows of opportunity
Windows of ecological opportunity for primary producersmicrobial heterotrophs and consumers differed between
5
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 1 Water nutrient concentrations of soluble reactive phosphorus (SRP) and dissolved inorganic nitrogen (DIN sum ofammonia+ nitrate+ nitrite forms) in the C and E reaches of the Mediterranean Pampean and Andean streams Values are means andstandard errors of nutrients measured during the pre-enrichment (before) and enrichment (after) periods and are accompanied by NP molarratio calculations The increment (1) nutrient concentration during enrichment is also shown for each of the streams
Control reach Enriched reach 1 nutrient concentration
Mediterranean Before SRP (microg lminus1) 14plusmn 18 16plusmn 21DIN (microg lminus1) 432plusmn 438 523plusmn 491NP 68 71
After SRP (microg lminus1) 10plusmn 9 30plusmn 13 times3DIN (microg lminus1) 389plusmn 495 765plusmn 736 times2NP 84 56
Pampean Before SRP (microg lminus1) 143plusmn 17 173plusmn 31DIN (microg lminus1) 683plusmn 39 1093plusmn 228NP 11 14
After SRP (microg lminus1) 93plusmn 3 361plusmn 29 times4DIN (microg lminus1) 528plusmn 136 828plusmn 197 times16NP 13 5
Andean Before SRP (microg lminus1) 26plusmn 5 28plusmn 5DIN (microg lminus1) 55plusmn 21 78plusmn 23NP 5 6
After SRP (microg lminus1) 53plusmn 4 105plusmn 13 times2DIN (microg lminus1) 114plusmn 59 323plusmn 85 times28NP 5 7
streams in terms of duration and frequency (figure 1)Windows of opportunity in the Mediterranean stream werenarrow and limited to late springndashearly summer (moderatetemperature and light availability) and late summerndashearlyautumn periods (moderate temperatures and abundant al-lochthonous OM inputs) The greatest overlap of favor-able environmental conditions for biological developmentoccurred in the Pampean stream In this system higher lightelevated temperatures and low discharge values coincidedfrom winter to summer (SeptemberndashMarch) The absenceof riparian vegetation reduced allochthonous OM inputsin this stream but authocthonous OM was produced byalgae and macrophytes The Andean stream also had longfavorable periods of temperature light and allochthonous OMinputs only interrupted by high discharge periods betweenAprilndashMay and OctoberndashNovember The high flow periodsin this stream caused a momentary biomass decrease of allbiological groups The predictive-BACIPS approach (table 2)suggested that 10 out of 23 models had significant treatmentseffects (or interactions) and that the explained variance wasoften high (figure 6)
33 Bacterial biomass
Bacterial biomass before the enrichment ranged from 2to 16 microg C cmminus2 on rocks and fine sediment in theMediterranean and Andean streams from 5 to 700microg C cmminus2
on sediments 004ndash15 microg C cmminus2 on macrophytes and1ndash36microg C cmminus2 in the water column for the Pampean streamThe nutrient enrichment resulted in an increase in bacterialbiomass in the Mediterranean (rocks and fine sediment) andPampean (fine sediment) streams but not in the Andeanstream (figure 2 table 2) The largest increase in bacterial
biomass after enrichment (calculated as the biomass changeratio) occurred in the Mediterranean stream (65ndash80 overallincrease) In the Pampean stream bacterial biomass increasedbetween 20 and 30 in fine sediment and macrophytesbut decreased in the water column (10ndash20) Water flowconditions temperature and light availability covaried withbacterial biomass in the E reaches of the Mediterranean andPampean streams The temporal variability of the biomasschange ratio was the highest in the Mediterranean stream(coefficient of variation CV = 24) due to the episodichigher biomass between late spring and early summer in theenrichment period
34 Algal and macrophytic biomass and composition
Algal biomass before the enrichment averaged 205 plusmn31 microg C cmminus2 on rocks and 69plusmn11 microg C cmminus2 on sedimentsat the Mediterranean stream (figure 3) Algal biomassaccounted for 137 plusmn 18 microg C cmminus2 on rocky habitat in theAndean stream while reached 162 plusmn 29 microg C cmminus2 onfine sediment and 23 plusmn 3 microg C cmminus2 for suspended algaein the Pampean stream In this stream macrophytes biomassaccounted for 58plusmn 27 g C mminus2
Nutrient enrichment produced an increase in algalbiomass in the three streams (figure 3 table 2) althoughthe responses varied in magnitude and differed betweencompartments The largest increase occurred on rocks inthe Mediterranean stream (163 on average peak of 273during spring) In the Andean stream algal biomass on rocksrapidly increased (62) and remained steady thereafter Thesmallest biomass increase was registered in the Pampeanstream particularly in the water column and for macrophytesbut biomass increased up to 49 on fine sediments The
6
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 2 Predictive-BACIPS of the effects of nutrient enrichment on the biomass of biological compartments in the three streams Thevalues shown correspond to the estimated coefficients of the linear models (ANCOVAs) When the control times BA was not significant(P gt 010) it was deleted from the model see section 2 for further details ns P gt 010 a P le 010 P le 005 P le 001 P le 0001
Variable n Control BA Control times BA
MediterraneanBacteria rocks 18 0404 0068 nsBacteria fine sediment 23 0590 0078 nsAlgae rocks 19 0495 0202a nsAlgae fine sediment 23 1082 0315 minus0732Invertebrates fine sediment 7 minus1011 minus0072 1660a
PampeanBacteria fine sediment 11 0751 0510a nsBacteria suspended 11 0954 minus0018 nsBact macrophytes 8 0396 0097 nsAlgae fine sediment 10 1106 0206 nsAlgae suspended 5 0274 minus0079 nsAlgae macrophyte 10 0575 minus0150 nsMacrophytes 11 1093 0249 nsInvertebrates fine sediment 10 minus0130 0787 nsInvertebrates macrophytes 6 minus0251 minus0521a nsFish CPUE 4 1082 8221 nsFish BPUE 4 0745 29552 ns
AndeanBacteria rocks 7 0666a 0299 nsBacteria fine sediment 6 0150 minus2631a nsAlgae rocks 14 0158 minus0101 0728Invertebrates rocks 13 0469 minus0269 nsInvertebrates fine sediment 13 0432 minus0388 nsFish density 10 1012 minus0442 nsFish biomass 10 1020 minus0464a ns
algal biomass responses to nutrients in the three streamswere strongly related with favorable conditions of lighttemperature and water flow (figure 3) The difference betweenthe extension of the respective windows of opportunitydetermined the steadiness of the response The Mediterraneanand Andean streams represented the maximum and minimumvariability in favorable conditions for primary producersrsquogrowth accordingly the coefficients of variation of thebiomass ratiorsquos change were the highest in the Mediterranean(CV = 16) and the lowest in the Andean (CV = 08)stream
Nutrient enrichment affected the composition of benthicalgal assemblages Some diatom taxa decreased (egAchnanthes biasolettiana Grunow (minus57) Achnanthesminutissima Kutzing (minus432) in the MediterraneanFragilaria ulna (Nitzsch) Lange-Bertalot var ulna (minus68)in the Pampean and Cocconeis placentula Ehrenberg(minus40) in the Andean stream) whereas others increased(eg Rhoicosphenia abbreviata (Ag) Lange-Bertalot (94)and Cocconeis placentula Ehrenberg (594) in theMediterranean Nitzschia frustulum (Kutzing) Grunow varfrustulum (3528) in the Pampean and Reimeria sinuata(Gregory) Kociolek and Stoermer (646) in the Andeanstream) Enrichment in the Pampean stream had no effect onthe macrophyte composition Ludwigia sp and Bacopa spdominated both the C and E reaches and were later replacedby the macroalga Spirogyra sp
35 Invertebrate biomass and composition
Invertebrate biomass before the enrichment ranged from5 to 750 microg DM cmminus2 in the Mediterranean and Andeanstreams and reached 1093 plusmn 311 microg DM cmminus2 in thesediments of the Pampean stream The effects of nutrientenrichment were generally non-significant but often larger(figure 4 table 2) and more pronounced in some habitatsthan others In the Pampean stream effects were not apparenton invertebrates occurring on macrophytes and were minor(3) on fine sediments Invertebrate biomass increased upto 155 in the fine sediments of the Mediterranean streamwhile no increase was observed in the Andean stream Theeffects on invertebrate biomass were the least variable inthe Mediterranean stream (per cent increase ratio CV = 09CV = 12 in the Andean and 27 in the Pampean)
Although the general enrichment effects on invertebratebiomass were weak some specific invertebrate taxa weredistinctly affected The grazer Ancylus fluviatilis increasedby 17 in the Mediterranean stream and the grazerCamelobaetidus increased by 927 in the Andean streamDensities of the scrapers Heleobia parchappei Orbigny(in fine sediments) and Hyalella curvispina Shoemaker(on macrophytes) increased in the Pampean stream (48and 762 respectively) Potamopyrgus antipodarum Gray(scraper) and Corbicula fluminea Muller (filterer) occurredonly in the respective fertilized reaches of the Mediterraneanand Pampean streams The Tubificidae (minus38) and theTanytarsini chironomidae (minus29) significantly decreased in
7
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 1 Water nutrient concentrations of soluble reactive phosphorus (SRP) and dissolved inorganic nitrogen (DIN sum ofammonia+ nitrate+ nitrite forms) in the C and E reaches of the Mediterranean Pampean and Andean streams Values are means andstandard errors of nutrients measured during the pre-enrichment (before) and enrichment (after) periods and are accompanied by NP molarratio calculations The increment (1) nutrient concentration during enrichment is also shown for each of the streams
Control reach Enriched reach 1 nutrient concentration
Mediterranean Before SRP (microg lminus1) 14plusmn 18 16plusmn 21DIN (microg lminus1) 432plusmn 438 523plusmn 491NP 68 71
After SRP (microg lminus1) 10plusmn 9 30plusmn 13 times3DIN (microg lminus1) 389plusmn 495 765plusmn 736 times2NP 84 56
Pampean Before SRP (microg lminus1) 143plusmn 17 173plusmn 31DIN (microg lminus1) 683plusmn 39 1093plusmn 228NP 11 14
After SRP (microg lminus1) 93plusmn 3 361plusmn 29 times4DIN (microg lminus1) 528plusmn 136 828plusmn 197 times16NP 13 5
Andean Before SRP (microg lminus1) 26plusmn 5 28plusmn 5DIN (microg lminus1) 55plusmn 21 78plusmn 23NP 5 6
After SRP (microg lminus1) 53plusmn 4 105plusmn 13 times2DIN (microg lminus1) 114plusmn 59 323plusmn 85 times28NP 5 7
streams in terms of duration and frequency (figure 1)Windows of opportunity in the Mediterranean stream werenarrow and limited to late springndashearly summer (moderatetemperature and light availability) and late summerndashearlyautumn periods (moderate temperatures and abundant al-lochthonous OM inputs) The greatest overlap of favor-able environmental conditions for biological developmentoccurred in the Pampean stream In this system higher lightelevated temperatures and low discharge values coincidedfrom winter to summer (SeptemberndashMarch) The absenceof riparian vegetation reduced allochthonous OM inputsin this stream but authocthonous OM was produced byalgae and macrophytes The Andean stream also had longfavorable periods of temperature light and allochthonous OMinputs only interrupted by high discharge periods betweenAprilndashMay and OctoberndashNovember The high flow periodsin this stream caused a momentary biomass decrease of allbiological groups The predictive-BACIPS approach (table 2)suggested that 10 out of 23 models had significant treatmentseffects (or interactions) and that the explained variance wasoften high (figure 6)
33 Bacterial biomass
Bacterial biomass before the enrichment ranged from 2to 16 microg C cmminus2 on rocks and fine sediment in theMediterranean and Andean streams from 5 to 700microg C cmminus2
on sediments 004ndash15 microg C cmminus2 on macrophytes and1ndash36microg C cmminus2 in the water column for the Pampean streamThe nutrient enrichment resulted in an increase in bacterialbiomass in the Mediterranean (rocks and fine sediment) andPampean (fine sediment) streams but not in the Andeanstream (figure 2 table 2) The largest increase in bacterial
biomass after enrichment (calculated as the biomass changeratio) occurred in the Mediterranean stream (65ndash80 overallincrease) In the Pampean stream bacterial biomass increasedbetween 20 and 30 in fine sediment and macrophytesbut decreased in the water column (10ndash20) Water flowconditions temperature and light availability covaried withbacterial biomass in the E reaches of the Mediterranean andPampean streams The temporal variability of the biomasschange ratio was the highest in the Mediterranean stream(coefficient of variation CV = 24) due to the episodichigher biomass between late spring and early summer in theenrichment period
34 Algal and macrophytic biomass and composition
Algal biomass before the enrichment averaged 205 plusmn31 microg C cmminus2 on rocks and 69plusmn11 microg C cmminus2 on sedimentsat the Mediterranean stream (figure 3) Algal biomassaccounted for 137 plusmn 18 microg C cmminus2 on rocky habitat in theAndean stream while reached 162 plusmn 29 microg C cmminus2 onfine sediment and 23 plusmn 3 microg C cmminus2 for suspended algaein the Pampean stream In this stream macrophytes biomassaccounted for 58plusmn 27 g C mminus2
Nutrient enrichment produced an increase in algalbiomass in the three streams (figure 3 table 2) althoughthe responses varied in magnitude and differed betweencompartments The largest increase occurred on rocks inthe Mediterranean stream (163 on average peak of 273during spring) In the Andean stream algal biomass on rocksrapidly increased (62) and remained steady thereafter Thesmallest biomass increase was registered in the Pampeanstream particularly in the water column and for macrophytesbut biomass increased up to 49 on fine sediments The
6
Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 2 Predictive-BACIPS of the effects of nutrient enrichment on the biomass of biological compartments in the three streams Thevalues shown correspond to the estimated coefficients of the linear models (ANCOVAs) When the control times BA was not significant(P gt 010) it was deleted from the model see section 2 for further details ns P gt 010 a P le 010 P le 005 P le 001 P le 0001
Variable n Control BA Control times BA
MediterraneanBacteria rocks 18 0404 0068 nsBacteria fine sediment 23 0590 0078 nsAlgae rocks 19 0495 0202a nsAlgae fine sediment 23 1082 0315 minus0732Invertebrates fine sediment 7 minus1011 minus0072 1660a
PampeanBacteria fine sediment 11 0751 0510a nsBacteria suspended 11 0954 minus0018 nsBact macrophytes 8 0396 0097 nsAlgae fine sediment 10 1106 0206 nsAlgae suspended 5 0274 minus0079 nsAlgae macrophyte 10 0575 minus0150 nsMacrophytes 11 1093 0249 nsInvertebrates fine sediment 10 minus0130 0787 nsInvertebrates macrophytes 6 minus0251 minus0521a nsFish CPUE 4 1082 8221 nsFish BPUE 4 0745 29552 ns
AndeanBacteria rocks 7 0666a 0299 nsBacteria fine sediment 6 0150 minus2631a nsAlgae rocks 14 0158 minus0101 0728Invertebrates rocks 13 0469 minus0269 nsInvertebrates fine sediment 13 0432 minus0388 nsFish density 10 1012 minus0442 nsFish biomass 10 1020 minus0464a ns
algal biomass responses to nutrients in the three streamswere strongly related with favorable conditions of lighttemperature and water flow (figure 3) The difference betweenthe extension of the respective windows of opportunitydetermined the steadiness of the response The Mediterraneanand Andean streams represented the maximum and minimumvariability in favorable conditions for primary producersrsquogrowth accordingly the coefficients of variation of thebiomass ratiorsquos change were the highest in the Mediterranean(CV = 16) and the lowest in the Andean (CV = 08)stream
Nutrient enrichment affected the composition of benthicalgal assemblages Some diatom taxa decreased (egAchnanthes biasolettiana Grunow (minus57) Achnanthesminutissima Kutzing (minus432) in the MediterraneanFragilaria ulna (Nitzsch) Lange-Bertalot var ulna (minus68)in the Pampean and Cocconeis placentula Ehrenberg(minus40) in the Andean stream) whereas others increased(eg Rhoicosphenia abbreviata (Ag) Lange-Bertalot (94)and Cocconeis placentula Ehrenberg (594) in theMediterranean Nitzschia frustulum (Kutzing) Grunow varfrustulum (3528) in the Pampean and Reimeria sinuata(Gregory) Kociolek and Stoermer (646) in the Andeanstream) Enrichment in the Pampean stream had no effect onthe macrophyte composition Ludwigia sp and Bacopa spdominated both the C and E reaches and were later replacedby the macroalga Spirogyra sp
35 Invertebrate biomass and composition
Invertebrate biomass before the enrichment ranged from5 to 750 microg DM cmminus2 in the Mediterranean and Andeanstreams and reached 1093 plusmn 311 microg DM cmminus2 in thesediments of the Pampean stream The effects of nutrientenrichment were generally non-significant but often larger(figure 4 table 2) and more pronounced in some habitatsthan others In the Pampean stream effects were not apparenton invertebrates occurring on macrophytes and were minor(3) on fine sediments Invertebrate biomass increased upto 155 in the fine sediments of the Mediterranean streamwhile no increase was observed in the Andean stream Theeffects on invertebrate biomass were the least variable inthe Mediterranean stream (per cent increase ratio CV = 09CV = 12 in the Andean and 27 in the Pampean)
Although the general enrichment effects on invertebratebiomass were weak some specific invertebrate taxa weredistinctly affected The grazer Ancylus fluviatilis increasedby 17 in the Mediterranean stream and the grazerCamelobaetidus increased by 927 in the Andean streamDensities of the scrapers Heleobia parchappei Orbigny(in fine sediments) and Hyalella curvispina Shoemaker(on macrophytes) increased in the Pampean stream (48and 762 respectively) Potamopyrgus antipodarum Gray(scraper) and Corbicula fluminea Muller (filterer) occurredonly in the respective fertilized reaches of the Mediterraneanand Pampean streams The Tubificidae (minus38) and theTanytarsini chironomidae (minus29) significantly decreased in
7
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
Table 2 Predictive-BACIPS of the effects of nutrient enrichment on the biomass of biological compartments in the three streams Thevalues shown correspond to the estimated coefficients of the linear models (ANCOVAs) When the control times BA was not significant(P gt 010) it was deleted from the model see section 2 for further details ns P gt 010 a P le 010 P le 005 P le 001 P le 0001
Variable n Control BA Control times BA
MediterraneanBacteria rocks 18 0404 0068 nsBacteria fine sediment 23 0590 0078 nsAlgae rocks 19 0495 0202a nsAlgae fine sediment 23 1082 0315 minus0732Invertebrates fine sediment 7 minus1011 minus0072 1660a
PampeanBacteria fine sediment 11 0751 0510a nsBacteria suspended 11 0954 minus0018 nsBact macrophytes 8 0396 0097 nsAlgae fine sediment 10 1106 0206 nsAlgae suspended 5 0274 minus0079 nsAlgae macrophyte 10 0575 minus0150 nsMacrophytes 11 1093 0249 nsInvertebrates fine sediment 10 minus0130 0787 nsInvertebrates macrophytes 6 minus0251 minus0521a nsFish CPUE 4 1082 8221 nsFish BPUE 4 0745 29552 ns
AndeanBacteria rocks 7 0666a 0299 nsBacteria fine sediment 6 0150 minus2631a nsAlgae rocks 14 0158 minus0101 0728Invertebrates rocks 13 0469 minus0269 nsInvertebrates fine sediment 13 0432 minus0388 nsFish density 10 1012 minus0442 nsFish biomass 10 1020 minus0464a ns
algal biomass responses to nutrients in the three streamswere strongly related with favorable conditions of lighttemperature and water flow (figure 3) The difference betweenthe extension of the respective windows of opportunitydetermined the steadiness of the response The Mediterraneanand Andean streams represented the maximum and minimumvariability in favorable conditions for primary producersrsquogrowth accordingly the coefficients of variation of thebiomass ratiorsquos change were the highest in the Mediterranean(CV = 16) and the lowest in the Andean (CV = 08)stream
Nutrient enrichment affected the composition of benthicalgal assemblages Some diatom taxa decreased (egAchnanthes biasolettiana Grunow (minus57) Achnanthesminutissima Kutzing (minus432) in the MediterraneanFragilaria ulna (Nitzsch) Lange-Bertalot var ulna (minus68)in the Pampean and Cocconeis placentula Ehrenberg(minus40) in the Andean stream) whereas others increased(eg Rhoicosphenia abbreviata (Ag) Lange-Bertalot (94)and Cocconeis placentula Ehrenberg (594) in theMediterranean Nitzschia frustulum (Kutzing) Grunow varfrustulum (3528) in the Pampean and Reimeria sinuata(Gregory) Kociolek and Stoermer (646) in the Andeanstream) Enrichment in the Pampean stream had no effect onthe macrophyte composition Ludwigia sp and Bacopa spdominated both the C and E reaches and were later replacedby the macroalga Spirogyra sp
35 Invertebrate biomass and composition
Invertebrate biomass before the enrichment ranged from5 to 750 microg DM cmminus2 in the Mediterranean and Andeanstreams and reached 1093 plusmn 311 microg DM cmminus2 in thesediments of the Pampean stream The effects of nutrientenrichment were generally non-significant but often larger(figure 4 table 2) and more pronounced in some habitatsthan others In the Pampean stream effects were not apparenton invertebrates occurring on macrophytes and were minor(3) on fine sediments Invertebrate biomass increased upto 155 in the fine sediments of the Mediterranean streamwhile no increase was observed in the Andean stream Theeffects on invertebrate biomass were the least variable inthe Mediterranean stream (per cent increase ratio CV = 09CV = 12 in the Andean and 27 in the Pampean)
Although the general enrichment effects on invertebratebiomass were weak some specific invertebrate taxa weredistinctly affected The grazer Ancylus fluviatilis increasedby 17 in the Mediterranean stream and the grazerCamelobaetidus increased by 927 in the Andean streamDensities of the scrapers Heleobia parchappei Orbigny(in fine sediments) and Hyalella curvispina Shoemaker(on macrophytes) increased in the Pampean stream (48and 762 respectively) Potamopyrgus antipodarum Gray(scraper) and Corbicula fluminea Muller (filterer) occurredonly in the respective fertilized reaches of the Mediterraneanand Pampean streams The Tubificidae (minus38) and theTanytarsini chironomidae (minus29) significantly decreased in
7
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13
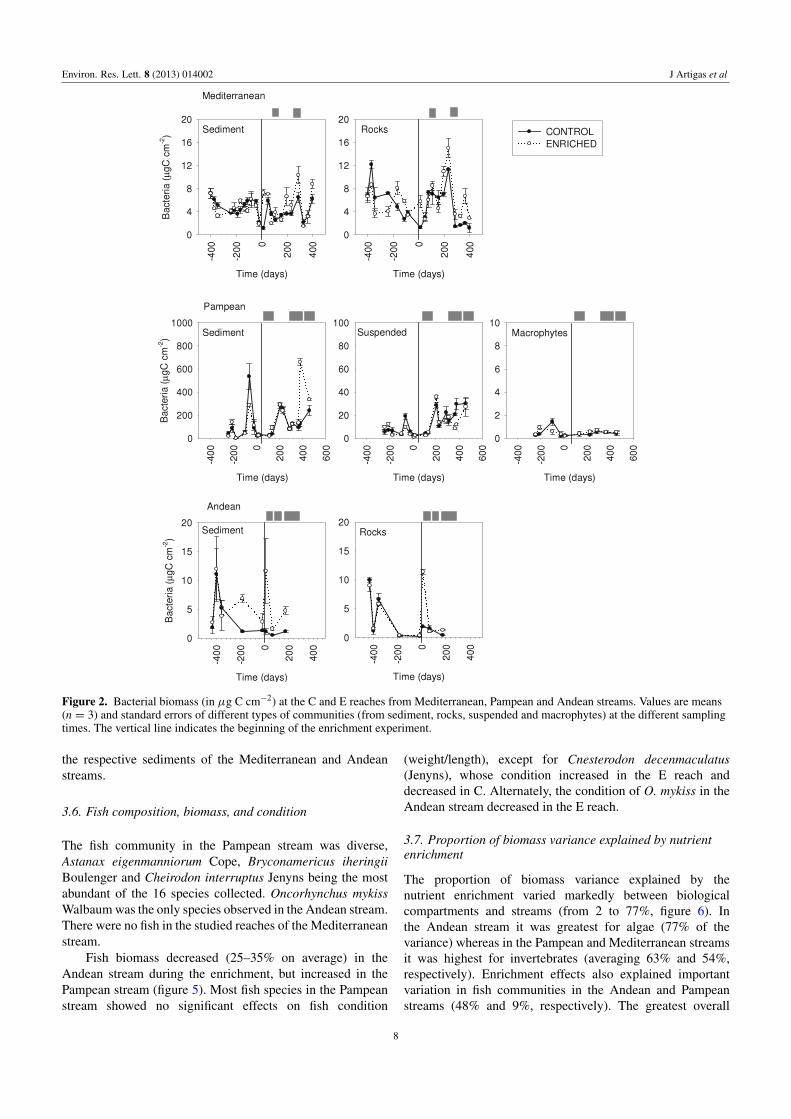
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 2 Bacterial biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
the respective sediments of the Mediterranean and Andeanstreams
36 Fish composition biomass and condition
The fish community in the Pampean stream was diverseAstanax eigenmanniorum Cope Bryconamericus iheringiiBoulenger and Cheirodon interruptus Jenyns being the mostabundant of the 16 species collected Oncorhynchus mykissWalbaum was the only species observed in the Andean streamThere were no fish in the studied reaches of the Mediterraneanstream
Fish biomass decreased (25ndash35 on average) in theAndean stream during the enrichment but increased in thePampean stream (figure 5) Most fish species in the Pampeanstream showed no significant effects on fish condition
(weightlength) except for Cnesterodon decenmaculatus(Jenyns) whose condition increased in the E reach anddecreased in C Alternately the condition of O mykiss in theAndean stream decreased in the E reach
37 Proportion of biomass variance explained by nutrientenrichment
The proportion of biomass variance explained by thenutrient enrichment varied markedly between biologicalcompartments and streams (from 2 to 77 figure 6) Inthe Andean stream it was greatest for algae (77 of thevariance) whereas in the Pampean and Mediterranean streamsit was highest for invertebrates (averaging 63 and 54respectively) Enrichment effects also explained importantvariation in fish communities in the Andean and Pampeanstreams (48 and 9 respectively) The greatest overall
8
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13
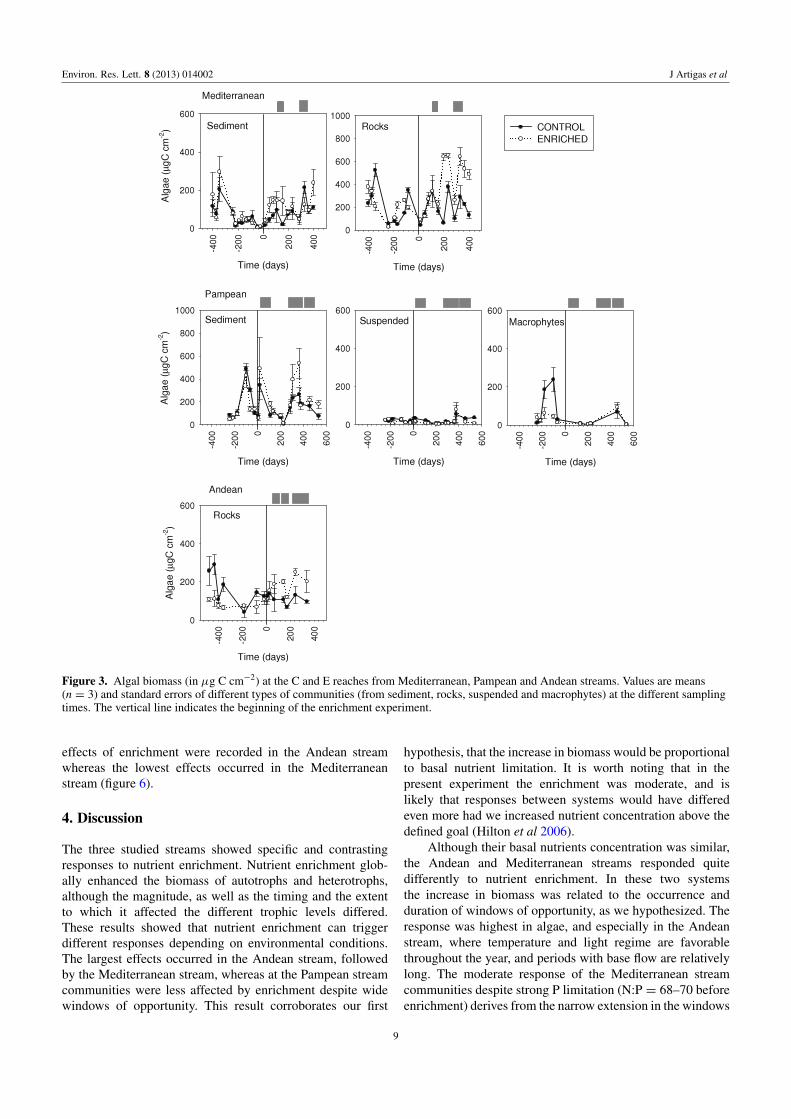
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 3 Algal biomass (in microg C cmminus2) at the C and E reaches from Mediterranean Pampean and Andean streams Values are means(n = 3) and standard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different samplingtimes The vertical line indicates the beginning of the enrichment experiment
effects of enrichment were recorded in the Andean streamwhereas the lowest effects occurred in the Mediterraneanstream (figure 6)
4 Discussion
The three studied streams showed specific and contrastingresponses to nutrient enrichment Nutrient enrichment glob-ally enhanced the biomass of autotrophs and heterotrophsalthough the magnitude as well as the timing and the extentto which it affected the different trophic levels differedThese results showed that nutrient enrichment can triggerdifferent responses depending on environmental conditionsThe largest effects occurred in the Andean stream followedby the Mediterranean stream whereas at the Pampean streamcommunities were less affected by enrichment despite widewindows of opportunity This result corroborates our first
hypothesis that the increase in biomass would be proportionalto basal nutrient limitation It is worth noting that in thepresent experiment the enrichment was moderate and islikely that responses between systems would have differedeven more had we increased nutrient concentration above thedefined goal (Hilton et al 2006)
Although their basal nutrients concentration was similarthe Andean and Mediterranean streams responded quitedifferently to nutrient enrichment In these two systemsthe increase in biomass was related to the occurrence andduration of windows of opportunity as we hypothesized Theresponse was highest in algae and especially in the Andeanstream where temperature and light regime are favorablethroughout the year and periods with base flow are relativelylong The moderate response of the Mediterranean streamcommunities despite strong P limitation (NP = 68ndash70 beforeenrichment) derives from the narrow extension in the windows
9
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13
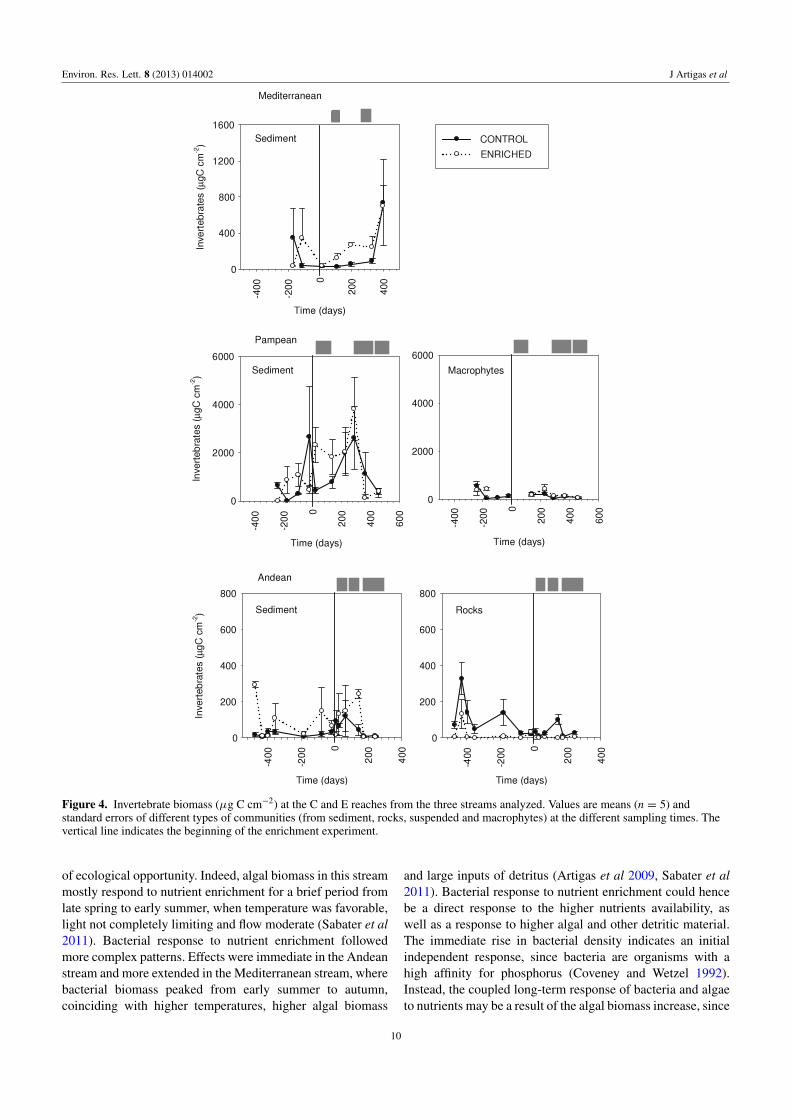
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 4 Invertebrate biomass (microg C cmminus2) at the C and E reaches from the three streams analyzed Values are means (n = 5) andstandard errors of different types of communities (from sediment rocks suspended and macrophytes) at the different sampling times Thevertical line indicates the beginning of the enrichment experiment
of ecological opportunity Indeed algal biomass in this streammostly respond to nutrient enrichment for a brief period fromlate spring to early summer when temperature was favorablelight not completely limiting and flow moderate (Sabater et al2011) Bacterial response to nutrient enrichment followedmore complex patterns Effects were immediate in the Andeanstream and more extended in the Mediterranean stream wherebacterial biomass peaked from early summer to autumncoinciding with higher temperatures higher algal biomass
and large inputs of detritus (Artigas et al 2009 Sabater et al2011) Bacterial response to nutrient enrichment could hencebe a direct response to the higher nutrients availability aswell as a response to higher algal and other detritic materialThe immediate rise in bacterial density indicates an initialindependent response since bacteria are organisms with ahigh affinity for phosphorus (Coveney and Wetzel 1992)Instead the coupled long-term response of bacteria and algaeto nutrients may be a result of the algal biomass increase since
10
Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
Figure 5 Fish biomass (fresh weight FW) per area sampled(BPUE) in the C and E reaches of the Pampean and Andean streamsat different sampling times The C communities are represented bysolid line while the dotted line is for E communities The verticalline indicates the beginning of the enrichment experiment
algae and bacteria maintain mutualistic relationships (Rier andStevenson 2002) expressed in higher bacterial biomass whenmore algal material is available
The effect of nutrient enrichment on consumer biomassand hence energy flow from producers to higher trophiclevels was in general moderate The magnitude observed inthe experimental response was related to the magnitude andduration of effects in basal trophic levels (Van de Koppelet al 2006) In the Mediterranean and Andean stream thesustained increase in algal biomass promoted a moderateincrease of consumer biomass (eg the grazers Ancylus orCamelobaetidus) as observed in other studies (Hairston andHairston 1993) In the Mediterranean stream the effect onalgal biomass was high but fluctuant and thus only someinvertebrate taxa with short life cycles and the capacity touse autochthonous organic matter (eg the grazer Ancylusfluviatilis) responded The large response in the Pampeanstream could be related with the microbial increase in thesediments a preferential habitat for invertebrates in thatsystem The fact that stream grazers were the ones showingthe highest response among invertebrates suggests that
Figure 6 Proportion of variance explained by nutrient enrichmenton the biomass of biological communities (bacteria algaemacrophytes invertebrates and fish) in the three streams The valuesrepresent the η2 values calculated with the beforeafter effect (BA)plus the controlenriched times time interaction (Ctimes BA) in theANCOVAs (see section 2 for further details) The values are theaverages of the different variables in table 2 averaged per biologicalcompartment and stream Bars indicate the standard error Pleasenote that standard errors could only be computed when there wasmore than one variable per compartment
algal-based food webs responded more than detritus-basedfood webs to the experimental manipulation It could happenthat enrichment was not sufficiently high to cause theresponse of decomposers observed in other experimentswhich used more substantial nutrient increases (Suberkroppet al 2010) Our biomass-based estimations could only yieldlimited information on trophic interactions and alternativeapproaches like stable isotopes (eg Bergfur et al 2009)might be necessary to disentangle the transference of effectsthrough the food webs in the streams In addition flowirregularities occurring in the Mediterranean stream (eg thelengthy drought during summer) could have impaired theresponse of consumers to bottom-up effects
The responses of the different trophic levels couldnot be separated from the complexity of the trophic foodweb The multiplicity of stream habitats as well as thecomplexity of the food web may cause disparity of biologicalresponses to nutrient enrichment (Davis et al 2010) Hiltonet al (2006) predicted two different types of response ofrivers to eutrophication in slow-flowing streams and riversthe response would be similar to that found in lakes andresult mainly in increased phytoplankton biomass whereasin fast-flowing streams the response would affect mainlybenthic organisms In our case the Pampean stream wasslow moving and very productive (Acuna et al 2011)but the response of incidental plankton was weak and themacrophytes were not affected At the top of the trophiccascade fish did not show an increase in biomass although thecondition of some species improved In the Pampean streamthe fish species Cnesterodon decenmaculatus improved theircondition during the enrichment This response was explained
11
Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
by greater amounts of autochthonous organic matter (algaemacrophytes) and higher quality relative to allochthonousdetritus that allows fish to survive unfavorable periods inbetter condition (see Wang et al 2006) In the Andeanstream where enrichment clearly enhanced autochthonousorganic matter resources fish condition was not improvedand biomass decreased However the greater mobility offish species as well as their feeding preferences (mostlysalmonids which are insectivorous) could be linked to thelarge response to enrichment observed by trout in the Andeanstream
Nutrient enrichment also had some effects on thestructure of biological assemblages Although we did notdetect reduced species richness the enrichment favored taxamore tolerant to the new conditions and therefore increasedin their relative abundances This was the case for severaldiatoms (Rhoicosphenia or Nitzschia Leira and Sabater2005) and several generalist invertebrates (Potamopgyrgusantipodarum or Corbicula fluminea) Increased dominanceof generalist taxa is a common outcome of nutrientenrichment (Stevenson and Pan 1999) However the fact thatsensitive species did not disappear shows that enrichmentlevels were moderate but alternatively may also indicativethat the enrichment was not sufficiently long Moderatenutrient addition tends to have smaller effects on communitycomposition than disturbances such as floods or severedroughts (Stevenson and Sabater 2010) The shift in speciesproportions might result in decreased efficiency of resourceuse but probably not in the impairment of the functioning ofthe whole ecosystem (Rapport and Whitford 1999)
The present research showed that stream ecosystemsrespond to moderate nutrient enrichment in highly specificways depending on environmental constraints and specificnutrient limitation In general the enrichment reinforcedautotrophic pathways (algaendashinvertebrate grazers) rather thanthe heterotrophic ones (bacteriandashinvertebrate shredders) thatcould have important consequences in cases of long-lastingenrichment Our results also suggested that the ecologicalconsequences can be highly site specific but that somepatterns can be related to existing climatic differences Weshowed that the tropical stream had the strongest responsebecause its windows of ecological opportunity were thewidest Windows of opportunity might be wide in tropicalstreams given their lesser seasonality and this makes themmore sensitive to eutrophication than other systems Thesesystems might be sensitive to additional disturbances suchas the elimination of riparian cover (which could favor lightentrance and warmer water) Other stream types (mostly inthe temperate region) may also increase their susceptibility toeutrophication because of ongoing scenarios of global changeIn many headwater streams of the temperate zone nutrientloads will increase and river discharge decrease (especiallyas a consequence of water abstraction Sabater and Tockner2010) Finally the effect of nutrient enrichment may beless predictable in those systems having high basal nutrientconcentrations such as the naturally enriched Pampeanstreams which could be close to saturation This stream showsthe most complex and more compartmentalized food web
of the three and this complexity can buffer the potentialtransference of bottom-up effects after nutrient addition
Overall these results indicate that a range of measurescould be taken to reduce the effects of eutrophication onstreams such as preserving the riparian vegetation to regulatelight and temperature or preserving of hydrological variabilityto guarantee nutrient dilution and biomass control
For site photographs please see the supplementarymaterial (available at stacksioporgERL8014002mmedia)
Acknowledgments
This study was funded by the project GLOBRIO of the BancoBilbao Vizcaya Argentaria (BBVA) Foundation Additionalfunding was obtained from the projects SCARCE (Con-solider Ingenio 2010 CSD2009-00065) and CARBONET(CGL2011-30474-C02-01) of the Spanish Ministry of Scienceand Innovation
References
Acuna V Vilches C and Giorgi A 2011 As productive and slow as astream can be-the metabolism of a Pampean stream J NorthAm Benthol Soc 30 71ndash83
APHA 1989 Standard Methods For the Examination of Water andWastewater 17th edn (Washington DC American PublicHealth Association)
Artigas J Romanı A M Gaudes A Munoz I and Sabater S 2009Organic matter availability structures microbial biomass andactivity in a Mediterranean stream Freshwater Biol54 2025ndash36
Bence J R 1995 Analysis of short time series correcting forautocorrelation Ecology 76 628ndash39
Bence J R Stewart-Oaten A and Schroeter S C 1996 Estimating thesize of an effect from a beforendashafter-controlndashimpact pairedseries design the predictive approach applied to a power plantstudy Detecting Ecological Impacts Concepts andApplications in Coastal Habitats ed R J Schmitt andC W Osenberg (San Diego CA Academic) pp 133ndash49
Bergfur J Johnson R K Sandin L and Goedkoop W 2009 Effects ofnutrient enrichment on C and N stable isotope ratios ofinvertebrates fish and their food resources in boreal streamsHydrobiologia 628 67ndash79
Bratbak G and Dundas I 1984 Bacterial dry matter content andbiomass estimations Appl Environ Microbiol 48 755ndash7
Coveney M F and Wetzel R G 1992 Effects of nutrients on specificgrowth rate of bacterioplankton in oligotrophic lake watercultures Appl Environ Microbiol 58 150ndash6
Davis J M Rosemond A D Eggert S L Cross W F andWallace J B 2010 Long-term nutrient enrichment decouplespredator and prey production Proc Natl Acad Sci 107 121ndash6
Frost P C Hillebrand H and Kahlert M 2005 Low algal carboncontent and its effects on the CP stoichiometry of periphytonFreshwater Biol 50 1800ndash7
Garcıa-Berthou E and Moreno-Amich R 1993 Multivariate analysisof covariance in morphometric studies of the reproductivecycle Can J Fish Aquat Sci 50 1394ndash9
Godfrey L and Tremayne A R 2005 The wild bootstrap andheteroskedasticity robust tests for serial correlation in dynamicregression models Comput Stat Data Anal 49 377ndash95
Hairston N G Jr and Hairston N G Sr 1993 Cause-effectrelationships in energy flow trophic structure and interspecificinteractions Am Natur 142 379ndash411
12
Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13

Environ Res Lett 8 (2013) 014002 J Artigas et al
Hart D D and Robinson C T 1990 Resource limitation on a streamcommunity phosphorus enrichment effects on periphyton andgrazers Ecology 71 1494ndash502
Hector A von Felten S and Schmid B 2010 Analysis of variancewith unbalanced data an update for ecology and evolutionJ Anim Ecol 79 308ndash16
Hill W R Fanta S E and Roberts B J 2009 Quantifying phosphorusand light effects in stream algae Limnol Oceanogr 54 368ndash80
Hilton J OrsquoHare M Bowes M J and Jones J I 2006 How green ismy river A new paradigm of eutrophication in rivers Sci TotalEnviron 365 66ndash83
Holomuzki J R Feminella J W and Power M E 2010 Bioticinteractions in freshwater benthic habitats J North AmBenthol Soc 29 220ndash44
Huxel G R McCann K and Polis G A 2002 Effects of partitioningallochthonous and autochthonous resources on food webstability Ecol Res 17 419ndash32
Jeffrey S and Humphrey G 1975 New spectrophotometric methodequation for determining chlorophyll a b c1 and c2 in higherplants algae and natural phytoplankton Biochim PhysiolPflanzen 167 194ndash204
Krammer K and Lange-Bertalot H 1986ndash1991 Bacillariophyceae 1Teil Naviculaceae 2 Teil Bacillariaceae EpithemiaceaeSurirellaceae 3 Teil Centrales Fragilariaceae Eunotiaceae4 Teil Achnanthaceae Kritische Erganzungen zu Navicula(Lineolatae) und Gomphonema (Stuttgart G Fischer)
Leira M and Sabater S 2005 Diatom assemblages distribution inCatalan rivers NE Spain in relation to chemical andphysiographical factors Water Res 39 73ndash82
Mas-Martı E Garcıa-Berthou E Sabater S Tomanova S andMunoz I 2010 Comparing fish assemblages and trophicecology of permanent and intermittent reaches in aMediterranean stream Hydrobiologia 657 167ndash80
Mulholland P J et al 2001 Inter-biome comparison of factorscontrolling stream metabolism Freshwater Biol 46 1503ndash17
Murphy J and Riley J P 1962 A modified single solution method forthe determination of phosphate in natural waters Ann ChimActa 27 31ndash6
Pascoal C Cassio F Marcotegui A Sanz B and Gomes P 2005 Roleof fungi bacteria and invertebrates in leaf litter breakdown ina polluted river J North Am Benthol Soc 24 784ndash97
Porter K G and Feig Y S 1980 The use of DAPI for identifying andcounting aquatic microflora Limnol Oceanogr 25 943ndash8
Rapport D J and Whitford W G 1999 How ecosystems respond tostress BioScience 49 193ndash203
R Development Core Team 2012 R A Language and Environmentfor Statistical Computing (Vienna R Foundation for StatisticalComputing) (wwwR-projectorg)
Rier S T and Stevenson R J 2002 Effects of light dissolved organiccarbon and inorganic nutrients on the relationship betweenalgae and heterotrophic bacteria in stream periphytonHydrobiologia 489 179ndash84
Rodrigues Capıtulo A Gomez N Giorgi A and Feijoo N 2010Global changes in pampean lowland streams (Argentina)implications for biodiversity and functioning Hydrobiologia657 53ndash70
Sabater S Artigas J Gaudes A Munoz I Urrea G andRomanı A M 2011 Moderate long-term nutrient inputenhances autotrophy in a forested Mediterranean streamFreshwater Biol 56 1266ndash80
Sabater S and Tockner K 2010 Effects of hydrologic alterations onthe ecological quality of river ecosystems Water Scarcity in theMediterranean Perspectives Under Global Change (TheHandbook of Environmental Chemistry) ed S Sabater andD Barcelo (Berlin Springer) (doi101007698-2009-24)
Smith V H 2003 Eutrophication of freshwater and coastal marineecosystems A global problem Environ Sci Pollut Res10 126ndash39
Sobczak W V Cloern J E Jassby A D Cole B E Schraga T S andArnsberg A 2005 Detritus fuels ecosystem metabolism but notmetazoan food webs in San Francisco estuaryrsquos freshwaterdelta Estuaries 28 124ndash37
Solorzano L 1969 Determination of ammonia in natural waters bythe phenolhypochlorite method Limnol Oceanogr 14 799ndash801
Stelzer R S Heffernan J and Likens G E 2003 The influence ofdissolved nutrients and particulate organic matter quality onmicrobial respiration and biomass in a forest streamFreshwater Biol 48 1925ndash37
Stevenson R J and Pan Y 1999 Assessing environmental conditionsin rivers and streams using diatoms The Diatoms Applicationfor the Environment and Earth Sciences ed E F Stoermer andJ P Smol (Cambridge Cambridge University Press) pp 11ndash40
Stevenson R J and Sabater S 2010 Understanding effects of a globalchange on river ecosystems science to support policy in achanging world Hydrobiologia 657 3ndash18
Stewart-Oaten A and Bence J R 2001 Temporal and spatial variationin environmental impact assessment Ecol Monograph71 305ndash39
Stouffer D B and Bascompte J 2011 Compartmentalizationincreases food-web persistence Proc Natl Acad Sci USA108 3648ndash52
Suberkropp K Gulis V Rosemond A D and Benstead J P 2010Ecosystem and physiological scales of microbial responses tonutrients in a detritus-based stream results of a five-yearcontinous enrichment Limnol Oceanogr 55 149ndash60
Sutton M A Oenema O Erisman J W Leip A van Grinsven H andWiniwarter W 2011 Too much of a good thing Nature472 159ndash61
Tank J L Rosi-Marshall E J Griffiths N A Entrekin S A andStephen M L 2010 A review of allochthonous organic matterdynamics and metabolism in streams J North Am BentholSoc 29 118ndash46
Tockner K Pusch M Borchardt D and Lorang M S 2010 Multiplestressors in coupled river-floodplain ecosystems FreshwaterBiol 55 135ndash51
Tornes E and Sabater S 2010 Variable discharge alters habitatsuitability for benthic algae and cyanobacteria in a forestedMediterranean stream Mar Freshwater Res 61 441ndash50
UNEP 2007 Global Environmental Outlook 4 Environment forDevelopment (Valletta United Nations EnvironmentProgramme)
Van de Koppel J Altieri A H Silliman B R Bruno J F andBertness M D 2006 Scale-dependent interactions andcommunity structure on cobble beaches Ecol Lett 9 45ndash50
Vorosmarty C J et al 2010 Global threats to human water securityand river biodiversity Nature 467 555ndash61
Wang L Robertson D M and Garrison P J 2006 Linkages betweennutrients and assemblages of macroinvertebrates and fish inwadeable streams implication to nutrient criteria developmentEnviron Manag 39 194ndash212
Zapata A M and Donato J C 2008 Regulacion hidrologica de labiomasa algal bentica Ecologıa de un rıo de montana de losAndes Colombianos (Rıo Tota Boyaca) ed J C Donato(Bogota Universidad Nacional de Colombia) pp 103ndash27
Zeileis A and Hothorn T 2002 Diagnostic checking in regressionrelationships R News 2 7ndash10 (httpCRANR-projectorgdocRnews)
13




















