
Germanium as a tool to dissect boron toxicity effects in barley and wheat
10
Germanium as a tool to dissect boron toxicity effects in barley and wheat Julie E. Hayes A,C , Margaret Pallotta A , Ute Baumann A , Bettina Berger B , Peter Langridge A and Tim Sutton A A Australian Centre for Plant Functional Genomics, School of Agriculture, Food and Wine, The University of Adelaide, Urrbrae, SA 5064, Australia. B The Plant Accelerator, The University of Adelaide, Waite Campus, Urrbrae, SA 5064, Australia. C Corresponding author. Email: [email protected] Abstract. Tolerance to boron (B) toxicity in barley (Hordeum vulgare L.) is partially attributable to HvNIP2;1, an aquaporin with permeability to B, as well as to silicon, arsenic and germanium (Ge). In this study, we mapped leaf symptoms of Ge toxicity in a doubled-haploid barley population (Clipper Sahara 3771). Two quantitative trait loci (QTL) associated with Ge toxicity symptoms were identified, located on Chromosomes 6H and 2H. These QTL co-located with two of four B toxicity tolerance loci previously mapped in the same population. The B toxicity tolerance gene underlying the 6H locus encodes HvNIP2;1, whereas the gene(s) and mechanisms underlying the 2H locus are as yet unknown. We provide examples of the application of Ge in studying specific aspects of B toxicity tolerance in plants, including screening of wheat (Triticum aestivum L.) and barley populations for altered function of HvNIP2;1 and related proteins. In particular, Ge may facilitate elucidation of the mechanism and gene(s) underlying the barley Chromosome 2H B tolerance locus. Additional keywords: aquaporins, HvNIP2;1, LemnaTec digital imaging and analysis. Received 2 November 2012, accepted 12 February 2013, published online 2 April 2013 Introduction The elements boron (B), silicon (Si), arsenic (As) and germanium (Ge) are classified in the periodic table as metalloids and they share several similar properties. All form weak acids in solution with high dissociation constants (pK a values of 8.6–9.8), so that in physiological pH ranges, they occur predominantly as free acid molecules. The free acids of each metalloid are small molecules (atomic radii of 3.43–4.48 Å), and all have recently been identified as substrates for transport through certain members of the nodulin-26-like intrinsic protein (NIP) family of aquaporins (water channel proteins; Takano et al. 2006; Bienert et al. 2008; reviewed in Bhattacharjee et al. 2008). The chemical similarities between Si and Ge, and the implications of this for plant biology have been recognised for some time. For example, Takahashi et al.(1976) found that the uptake of Ge and Si appeared to be similar, and that Si-accumulating plant species were particularly sensitive to Ge toxicity. This enabled Ge toxicity screens to be used to identify rice (Oryza sativa L.) mutants defective in Si accumulation (Ma et al. 2002), and was also useful in later identifying and characterising the genes underlying the mutations (Ma et al. 2006; Ma et al. 2007). Ge competitively inhibits the uptake of Si in wheat (Triticum aestivum L.) (Rains et al. 2006) and, recently, the radioactive 68 Ge isotope was validated as a suitable tracer for use in studies of Si transport in higher plants (Nikolic et al. 2007). There have also been a small number of studies looking at the ability of Ge to substitute for B in plant metabolism (Loomis and Durst 1991; Cakmak et al. 1995; Ishii et al. 2002). These studies focussed on the ability of Ge to overcome symptoms of B deficiency, and it was found that Ge is unable to substitute directly for B in its primary role in crosslinking cell wall pectins (Ishii et al. 2002). The relationship between Ge and B toxicities in plants has not been investigated, although barley (Hordeum vulgare L.) is reportedly much more sensitive to Ge (toxicity symptoms observed in the mM range; Halperin et al. 1995) compared with B (1–5 mM; Reid et al. 2004) when grown in nutrient solution. B toxicity has been recognised as a significant problem affecting cereal crop production for nearly 30 years (Cartwright et al. 1984). Considerable intraspecific variation for B toxicity tolerance exists, however, and wheat and barley breeding populations have been developed and exploited to identify the chromosomal regions associated with tolerance (Paull et al. 1995; Jefferies et al. 1999, 2000; Schnurbusch et al. 2007). In barley, four quantitative trait loci (QTL) associated with B toxicity tolerance were identified, using a doubled-haploid (DH) population developed from a cross between the malt variety Clipper and an Algerian landrace Sahara 3771 (Jefferies et al. 1999; Karakousis et al. 2003). A major QTL on Chromosome 4H is associated with shoot B concentration, leaf symptom expression, root growth CSIRO PUBLISHING Functional Plant Biology, 2013, 40, 618–627 http://dx.doi.org/10.1071/FP12329 Journal compilation Ó CSIRO 2013 www.publish.csiro.au/journals/fpb
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Germanium as a tool to dissect boron toxicity effects in barley and wheat

Germanium as a tool to dissect boron toxicity effectsin barley and wheat
Julie E. HayesA,C, Margaret PallottaA, Ute BaumannA, Bettina BergerB, Peter LangridgeA
and Tim SuttonA
AAustralian Centre for Plant Functional Genomics, School of Agriculture, Food and Wine,The University of Adelaide, Urrbrae, SA 5064, Australia.
BThe Plant Accelerator, The University of Adelaide, Waite Campus, Urrbrae, SA 5064, Australia.CCorresponding author. Email: [email protected]
Abstract. Tolerance to boron (B) toxicity in barley (Hordeum vulgare L.) is partially attributable to HvNIP2;1, anaquaporinwith permeability to B, aswell as to silicon, arsenic and germanium (Ge). In this study,wemapped leaf symptomsof Ge toxicity in a doubled-haploid barley population (Clipper�Sahara 3771). Two quantitative trait loci (QTL) associatedwith Ge toxicity symptoms were identified, located on Chromosomes 6H and 2H. These QTL co-located with two of four Btoxicity tolerance loci previously mapped in the same population. The B toxicity tolerance gene underlying the 6H locusencodes HvNIP2;1, whereas the gene(s) and mechanisms underlying the 2H locus are as yet unknown. We provideexamples of the application of Ge in studying specific aspects of B toxicity tolerance in plants, including screening ofwheat (Triticum aestivum L.) and barley populations for altered function of HvNIP2;1 and related proteins. In particular,Ge may facilitate elucidation of the mechanism and gene(s) underlying the barley Chromosome 2H B tolerance locus.
Additional keywords: aquaporins, HvNIP2;1, LemnaTec digital imaging and analysis.
Received 2 November 2012, accepted 12 February 2013, published online 2 April 2013
Introduction
The elements boron (B), silicon (Si), arsenic (As) and germanium(Ge) are classified in the periodic table as metalloids and theyshare several similar properties. All form weak acids in solutionwith high dissociation constants (pKa values of 8.6–9.8), so thatin physiological pH ranges, they occur predominantly as freeacid molecules. The free acids of each metalloid are smallmolecules (atomic radii of 3.43–4.48 Å), and all have recentlybeen identified as substrates for transport through certainmembers of the nodulin-26-like intrinsic protein (NIP) familyof aquaporins (water channel proteins; Takano et al. 2006;Bienert et al. 2008; reviewed in Bhattacharjee et al. 2008).
The chemical similarities between Si and Ge, and theimplications of this for plant biology have been recognised forsome time. For example, Takahashi et al. (1976) found thatthe uptake of Ge and Si appeared to be similar, and thatSi-accumulating plant species were particularly sensitive to Getoxicity. This enabled Ge toxicity screens to be used to identifyrice (Oryza sativa L.) mutants defective in Si accumulation(Ma et al. 2002), and was also useful in later identifying andcharacterising the genes underlying the mutations (Ma et al.2006; Ma et al. 2007). Ge competitively inhibits the uptakeof Si in wheat (Triticum aestivum L.) (Rains et al. 2006) and,recently, the radioactive 68Ge isotope was validated as asuitable tracer for use in studies of Si transport in higher plants(Nikolic et al. 2007).
There have also been a small number of studies looking atthe ability of Ge to substitute for B in plant metabolism (Loomisand Durst 1991; Cakmak et al. 1995; Ishii et al. 2002). Thesestudies focussed on the ability of Ge to overcome symptoms ofB deficiency, and it was found that Ge is unable to substitutedirectly for B in its primary role in crosslinking cell wall pectins(Ishii et al. 2002). The relationship between Ge and B toxicitiesin plants has not been investigated, although barley (Hordeumvulgare L.) is reportedly much more sensitive to Ge (toxicitysymptoms observed in the mM range; Halperin et al. 1995)compared with B (1–5mM; Reid et al. 2004) when grown innutrient solution.
B toxicity has been recognised as a significant problemaffecting cereal crop production for nearly 30 years(Cartwright et al. 1984). Considerable intraspecific variationfor B toxicity tolerance exists, however, and wheat and barleybreeding populations have been developed and exploited toidentify the chromosomal regions associated with tolerance(Paull et al. 1995; Jefferies et al. 1999, 2000; Schnurbuschet al. 2007). In barley, four quantitative trait loci (QTL)associated with B toxicity tolerance were identified, using adoubled-haploid (DH) population developed from a crossbetween the malt variety Clipper and an Algerian landraceSahara 3771 (Jefferies et al. 1999; Karakousis et al. 2003). Amajor QTL on Chromosome 4H is associated with shoot Bconcentration, leaf symptom expression, root growth
CSIRO PUBLISHING
Functional Plant Biology, 2013, 40, 618–627http://dx.doi.org/10.1071/FP12329
Journal compilation � CSIRO 2013 www.publish.csiro.au/journals/fpb

suppression and vegetative growth (Jefferies et al. 1999). Theunderlying gene at this locus is HvBot1 (Sutton et al. 2007),encoding an anion-permeable channel with similar propertiesto the B transporter originally identified in Arabidopsis thaliana(L.) Heynh. (AtBor1; Takano et al. 2002).
A second important QTL associated with shoot Bconcentration in barley is located on Chromosome 6H(Jefferies et al. 1999), where HvNIP2;1, a different type oftransporter, was identified as the gene underlying this locus(Schnurbusch et al. 2010). HvNIP2;1 encodes an aquaporinwith permeability to boric acid. In the B-tolerant barleygenotype, Sahara 3771, reduced expression of HvNIP2;1limits the entry of B into the plant (Schnurbusch et al. 2010).HvNIP2;1 and its direct orthologue in rice, OsNIP2;1 (Lsi1),were also independently identified as transporters for both Si(Ma et al. 2006; Chiba et al. 2009) andAs (Ma et al. 2008).Whenexpressed in yeast, HvNIP2;1 increases sensitivity to B, As andGe (Schnurbusch et al. 2010), suggesting that the aquaporinis permeable to these metalloids. Genetic studies with theClipper� Sahara 3771 DH population have shown that theHvNIP2;1 gene at the 6H B tolerance locus may account fora reduction in leaf B accumulation of up to 40%, depending onalleles at the other known loci (Schnurbusch et al. 2010).
A further two significant QTL associated with B toxicitytolerance were identified on Chromosomes 2H and 3H(Jefferies et al. 1999). A region on barley Chromosome 2His associated with leaf symptom expression and a locus onChromosome 3H is associated with a reduction in the effect ofB toxicity on root length. The identities of the genes underlyingthese QTL are not yet known, but genetic improvement in Btoxicity tolerance in barley may require selection for all four loci.The favourable alleles for each of the four loci are derived fromSahara 3771 (Jefferies et al. 1999).
In this work, we use a whole-plant approach to investigatethe potential for Ge to be used as an analogue for B in plantresearch, using Ge toxicity tolerance screening in cereals as anapplication. We propose that Ge may be particularly useful forstudying specific aspects of B toxicity tolerance in plants,including the role of HvNIP2;1 and related aquaporins.
Materials and methodsPlant culture
Seeds of wheat (36 genotypes of Triticum aestivum L. and eightgenotypes of T. durum L., listed in Table S1, available asSupplementary Material to this paper) and of the barley(Hordeum vulgare L.) DH population, Clipper� Sahara 3771(146 lines), were imbibed for 2–3 days at 4�C on moist filterpaper in Petri dishes. Germinated seeds were then transferredto containers of nutrient solution containing 5000mMNH4NO3,5mM KNO3, 2000mM Ca(NO3)2, 2000mM MgSO4, 100mMKH2PO4, 50mMNaFe(III) EDTA, 50mMB(OH)3, 5mMMnCl2,10mMZnSO4, 0.5mMCuSO4 and 0.1mMNa2MoO3. Seedlingswere cultured with aeration, either in a temperature-controlledglasshouse under ambient light (~23�C : 19�C day : night;barley Ge toxicity experiment) or in a growth room(12 h : 12 h day : night regime at 23�C : 15�C and 50% : 70%relative humidity and 115mmolm–2 s–1 light at plant height;barley B toxicity experiment and wheat Ge toxicity
experiment). At the emergence of the first leaf, solutions werereplaced with nutrient solution containing either 40mMGeO2 oran additional 5mM B(OH)3. Seedlings were grown for a further8–10 days before assessment of leaf symptoms of toxicity.Nutrient solutions were replaced at least twice weekly duringthe experiments.
Assessment of Ge and B toxicity symptoms
Two methods were used to assess leaf necrosis. Objectivemeasurements were made using a plant image capture andanalysis system (3D Scanalyzer, LemnaTec, Würselen,Germany). Harvested shoots of barley were placed on a flatblue surface and photographed from above. Then, using theLemnaGrid software for image analysis, the images wereseparated into foreground (plant) and background (blueimaging background) based on nearest neighbour coloursegmentation. All pixels of the image identified as the plantwere categorised into two colour classes: ‘healthy’ for green,and ‘necrotic’ for brown and white areas of the leaves. Thepercentage of necrotic area relative to the total plant size wasused as a measurement of toxicity symptoms. Symptoms of Band Ge toxicity were also assessed by visual comparison of theextent of necrosis in the first leaf to that in the parental barleyvarieties, Clipper and Sahara 3771. Scores were assigned on ascale of 0–5 (0–1 = Sahara 3771-like, with few or no symptoms;2 = necrosis at tip or base of first leaf; 3 = necrosis along firstleaf, but less extensive than for Clipper; 4–5 = severe necrosis,similar to or more extensive than Clipper). For both Ge and B,the reliability of the visual method of symptom assessment wasinvestigated by comparing scores to the objective measurementsof percentage necrosis.
QTL analyses were performed by simple interval mappingusing QGene ver. 4.3.6 (Joehanes and Nelson 2008). Wheatsymptom scores data were analysed by one-way ANOVA andTukey’s range testingusingGENSTAT software (www.vsni.co.uk).
HvNIP2;1 mutagenesis and functional assessment in yeast
Targeted mutations in the coding sequence of HvNIP2;1 weregenerated by in vitro site-directed mutagenesis (QuikChange IISite-Directed Mutagenesis Kit, Stratagene, La Jolla, CA, USA)of HvNIP2;1 in the Gateway entry vector pCR8 (Invitrogen,Carlsbad, CA, USA), using specific primer pairs detailed inTable S2. Mutations in the resulting plasmids were confirmedby sequencing. Mutated HvNIP2;1 clones were then transferredto a Gateway-enabled destination vector for yeast expression,pYES-DEST52 (Invitrogen).
Yeast (strain InVSc2, Invitrogen) was transformed using astandard lithium acetate method (Gietz and Woods 2002) andmaintained on minimal selection media, either with glucose orgalactose as a source of carbon. From each transformation, threeindependent yeast colonies were cultured in liquid galactosemedium at 30�C for 2 days. Cell densities were equalised forall samples and then 10-mL aliquots of undiluted and diluted(10�1, 10�2 and 10�3) samples were transferred to Petri dishes ofsolid media containing galactose and a range of concentrationsof B (5–20mM B(OH)3), Ge (100–200mgGemL–1 as GeO2) orAs (100–200mM As as As2O3). Plates were incubated at 30�Cfor 2–5 days. In all cases, growth of HvNIP2;1-expressing clones
Germanium and boron toxicities in cereals Functional Plant Biology 619

was compared with that of yeast transformed with a modifiedversion of the pYES-DEST52 vector (ccdB gene deleted) grownon the same plate.
Results
Identification of QTL associated with leaf symptomsof Ge toxicity in barley
The barley genotypes Clipper and Sahara 3771 differed greatlyin their tolerance to Ge based on assessment of leaf toxicitysymptoms (Fig. 1). The leaf blades and stem of theClipper seedlings developed necrotic lesions, beginning ataround 4 days after addition of Ge to the nutrient solution andbecoming more severe in the oldest leaf blades than in newlyemerging leaves (Fig. 1a). Sahara 3771 barley seedlings grownin the same solution remained healthy in appearance, with onlya small number of necrotic lesions appearing at the base of thestem after 8 days (Fig. 1b).
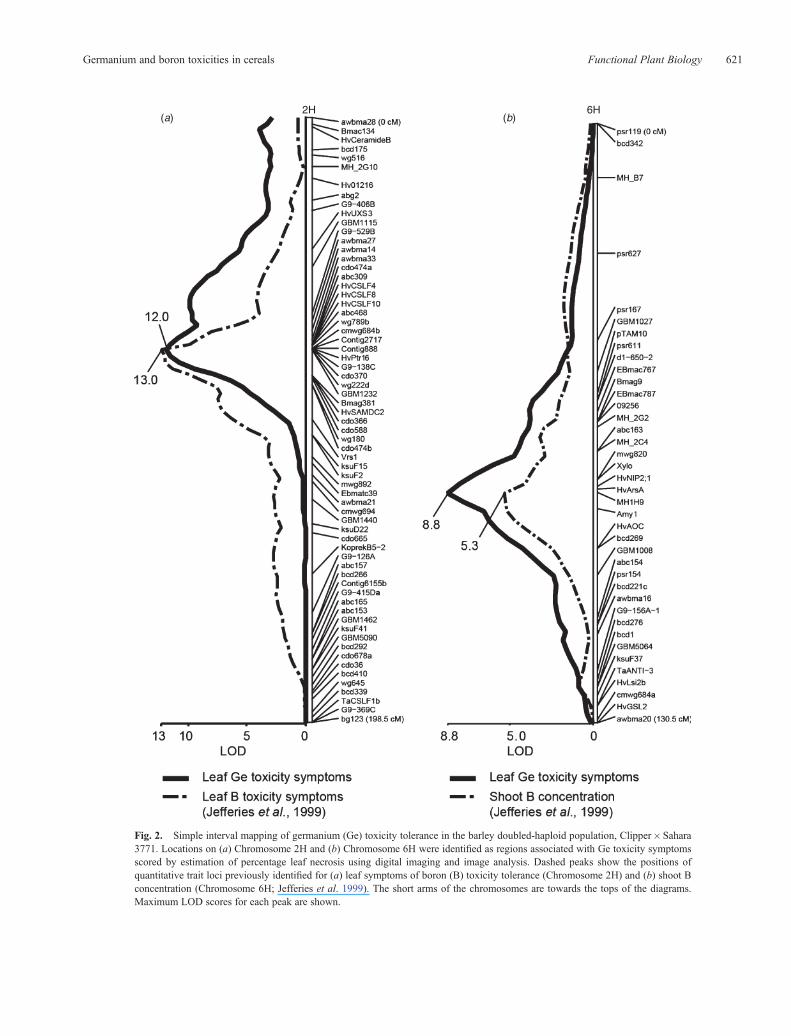
Leaf symptoms of Ge toxicity were assessed in 146 lines of abarleyDHpopulation developed from the cross Clipper� Sahara3771. Digital imaging and image analysis using LemnaTecScanalyser technology measured necrosis of seedling leafblades and stems ranging between 11% and 63%.Clipper seedlings developed an average of 55% necrosis after8 days’ growth at 40mM GeO2, but Sahara 3771 seedlingsshowed, on average, 11% necrosis. Simple interval mappingof percentage necrosis identified two significant QTL, onChromosomes 2H (logarithm (base 10) of odds (LOD) = 13.0)and 6H (LOD= 8.8; Fig. 2). The QTL on Chromosome 2H
co-locates with a QTL previously identified for B toxicitytolerance based on leaf symptom expression (Fig. 2a andJefferies et al. 1999). The most closely associated markers arethe same for both QTL in this region. The QTL on Chromosome6H co-locates with a second QTL identified for B toxicitytolerance, using shoot B concentration as a trait (Fig. 2b andJefferies et al. 1999). The markers most closely associated withboth Ge and B toxicity tolerance at this locus include HvNIP2;1,the gene at the 6H B toxicity tolerance locus. For both the 6Hand 2H loci, the favourable alleles for Ge toxicity tolerancebased on the severity of the leaf symptoms are derived fromSahara 3771. Critically, there was no association of regions onChromosome 4H with Ge toxicity tolerance.
To verify the genetic location of the previously identified Btolerance QTL, leaf symptoms of B toxicity were re-assessed inthe Clipper�Sahara 3771 DH population using a LemnaTecScanalyser for image capture and analysis. The populationwas grown hydroponically and treated with 5mM B(OH)3 for8 days. Simple interval mapping of percentage necrosis asdetermined by image analysis confirmed the presence of twomajor QTL, located on Chromosomes 2H and 4H (Fig. 3). Thepositions of the QTL peaks were the same in this study asdescribed previously, although the sizes of the peaks andassociated LOD scores were different (Fig. 3 and Jefferieset al. 1999).
The co-location of Ge toxicity tolerance QTL with Btoxicity tolerance QTL on Chromosomes 2H and 6H, but notChromosome 4H, suggested that Ge toxicity could be used tolook specifically at functional aspects of HvNIP2;1-derived B
(a) (b)
Fig. 1. Symptoms of germanium toxicity on leaves of the barley (Hordeum vulgare L.) genotypes(a) Clipper (boron toxicity-intolerant) and (b) Sahara 3771 (boron toxicity-tolerant). Seedlings weregrown in nutrient solution supplemented with 40mM GeO2 for 8 days.
620 Functional Plant Biology J. E. Hayes et al.
2H(a) (b)
6H
Fig. 2. Simple interval mapping of germanium (Ge) toxicity tolerance in the barley doubled-haploid population, Clipper�Sahara3771. Locations on (a) Chromosome 2H and (b) Chromosome 6H were identified as regions associated with Ge toxicity symptomsscored by estimation of percentage leaf necrosis using digital imaging and image analysis. Dashed peaks show the positions ofquantitative trait loci previously identified for (a) leaf symptoms of boron (B) toxicity tolerance (Chromosome 2H) and (b) shoot Bconcentration (Chromosome 6H; Jefferies et al. 1999). The short arms of the chromosomes are towards the tops of the diagrams.Maximum LOD scores for each peak are shown.
Germanium and boron toxicities in cereals Functional Plant Biology 621

2H(a) (b)
4H
Fig. 3. Simple interval mapping of boron (B) toxicity tolerance in the barley doubled-haploid population, Clipper�Sahara 3771. Locations on (a)Chromosome 2H and (b) Chromosome 4H were confirmed as regions associated with B toxicity symptoms scored by estimation of percentage leafnecrosis using digital imaging and image analysis. Dashed peaks show the positions of quantitative trait loci on these chromosomes previouslyidentified for leaf symptoms of B toxicity tolerance (Jefferies et al. 1999). The short arms of the chromosomes are towards the tops of the diagrams.Maximum LOD scores for each peak are shown.
622 Functional Plant Biology J. E. Hayes et al.

tolerance on Chromosome 6H, and also as a tool to facilitatethe identification of the unknown gene(s) underlying the 2Hlocus. Two small screening experiments were thereforeundertaken, looking at natural variation relating to these Btoxicity tolerance loci and induced mutations in wheat andbarley.
Validation of visual assessment of leaf symptomsof Ge and B toxicities
We first investigated the validity of using visual scoring for thescreening experiments as a more rapid method of assessingleaf necrosis caused by Ge and B toxicities. Leaf symptomsof Ge and B toxicities in the Clipper� Sahara 3771 DHpopulation were estimated visually on the same seedlingsused for image analysis, and the two datasets were compared.There were significant correlations between percentagenecrosis determined by image analysis and visual scoring (R2
values of 0.85 and 0.79 for Ge and B, respectively; Fig. 4).Interval mapping using visual symptom scores also identifiedthe same QTL as shown in Figs 2 and 3, with similar peaks andLOD scores. Thus, visual scoring of Ge symptoms is suitablefor screening.
Variation for Ge toxicity tolerance in wheat
A set of bread wheat and durum genotypes (Table S1), chosen torepresent a range of reported levels of tolerance to both B toxicityand deficiency, were assessed visually for Ge toxicity tolerance.There was considerable variation observed in Ge toxicitytolerance between the genotypes, with two bread wheatvarieties, Condor and Kenya Farmer, showing significantlyfewer leaf symptoms (P < 0.05; Fig. 5) than the other breadwheat genotypes, whereas the variety Lin Calel wasparticularly intolerant to Ge toxicity. Notably, however, theparents of populations used to study the genetics of B toxicitytolerance in wheat (Halberd�Cranbrook, Halberd�Wl*MMC,G61450�Kenya Farmer; Paull et al. 1995; Jefferies et al. 2000)did not show similar differences in Ge toxicity tolerance,suggesting that the B toxicity tolerance traits in these geneticresources are not related to sequences orthologous to HvNIP2;1or the gene(s) underlying barley Chromosome 2H. None of thedurum or bread wheat genotypes were as tolerant to Ge asSahara 3371 barley (Fig. 5; P< 0.001), which carries both ofthe 2H and 6H Ge and B toxicity tolerance alleles.
Screening for Ge toxicity-tolerant mutants in barley
Approximately 4500 mutated generation 3 (M3) individualsfrom an ethane methyl sulphonate (EMS)-mutagenised barleytilling population (H. vulgare cv. Flagship) were phenotypicallyscreened for Ge toxicity tolerance to identify mutants relating toeitherHvNIP2;1 or the gene(s) at the Chromosome 2H QTL.Weidentified 44 putative mutants in the initial screen. Ten of themutants were confirmed as Ge-tolerant in the M4 generation.Sequence analysis of the HvNIP2;1 open reading frame fromthese individuals failed to reveal any mutations. However, twoconfirmed mutants also showed reduced leaf symptoms in asubsequent hydroponics-based B toxicity screen, suggestingpossible linkage of the mutation(s) to the known B toxicitytolerance loci in chromosome regions 2H and 6H.
Targeted mutation analysis of HvNIP2;1 in yeast
HvNIP2;1 is permeable to a range of metalloids, which couldhave implications for crops where arsenic accumulation posessignificant health risks (Meharg 2004; Ma et al. 2008). Wetherefore developed a set of induced HvNIP2;1 mutants tomanipulate the transport selectivity of the protein, targetingresidues on Helix 2 (H2) and Helix 5 (H5) (Wallace andRoberts 2005; Table S2). At position H2, glycine wassubstituted for residues with increasingly larger, neutral sidechains (alanine, cysteine, valine). At position H5, serine wassubstituted for valine, which is the amino acid at this positionin GmNOD26, a member of the NIP family that is notpermeable to B (Schnurbusch et al. 2010). An additionalmutation (Ala132Thr) was generated to re-create a mutant ofOsNIP2;1 which is reported to have disrupted Si (Ge)transport (Ma et al. 2006).
Mutants were characterised by heterologous expression inyeast and comparison to yeast expressing wild-type HvNIP2;1when grown on high concentrations of B, Ge and As (Fig. 6).Yeast expressing HvNIP2;1 grew poorly on all three metalloids,suggesting that the expression of the aquaporin increasedpermeability of the yeast cell membranes to B, Ge and As.
100
80
R2 = 0.85
R2 = 0.79
(a)
(b)
60
40
20
0
80
60
% n
ecro
sis
(imag
e an
alys
is)
Symptom score
40
20
00 1 2 3 4 5
Fig. 4. Correlations between leaf symptom scores assigned by visualassessment and percentage necrosis estimated by digital image analysis,for (a) germanium and (b) boron toxicities applied to lines of the barleydoubled-haploid population, Clipper� Sahara 3771. Linear trend lines werefitted to the data and R2 values for best fits are shown.
Germanium and boron toxicities in cereals Functional Plant Biology 623

However, yeast that expressed versions of HvNIP2;1 carryingthe mutations showed restored growth on high concentrationsof both B and Ge, which suggested reduced permeability tothese species. Two of the mutations (Gly88Ala and Gly88Cys)showed a specific growth restoration in B and Ge, but theinhibition of growth on As was retained (Fig. 6). Thissuggests that these mutations altered the substrate specificityof HvNIP2;1. None of the mutations resulted in a distinctionbetween B and Ge as substrates for HvNIP2;1.
DiscussionThe barley landrace Sahara 3771 is widely reported to be tolerantto B toxicity, both in the field and in controlled growthenvironments. B toxicity tolerance in this variety is due, inpart, to both high expression of the HvBot1 B transporter(Sutton et al. 2007) and reduced expression of HvNIP2;1, anaquaporin with permeability to B (Schnurbusch et al. 2010). Inthis study, we report that Sahara 3771 barley is also tolerant totoxic concentrations of Ge (Figs 1 and 5). Sahara 3771 seedlings
5
4
3
2
Leaf
sym
ptom
sco
re
1
0
Fig. 5. Variation in germanium (Ge) toxicity tolerance for a set of bread wheat (Triticum aestivum; solid bars) anddurum (T. durum; open bars) genotypes, assessed by visual scoring of leaf symptoms of Ge toxicity (0 = no symptoms;5 = extensive necrosis). Scores are also shown for the barley genotypes Clipper and Sahara 3771. Measurements are meansof eight replicates and error bars indicate s.d.
Empty vector
10 mM B
(a) (b) (c) (d )
200 µg Ge mL–1 100 µM As (III) No treatment
HvNIP2;1
NIP G88A
NIP G88C
NIP G88V
NIP G88A/S207V
NIP A132T
Fig. 6. Permeability to (a) boron (B), (b) germanium (Ge) and (c) arsenic (As) of nonmutatedHvNIP2;1 and versions with mutations in key amino acid residues, as assessed by toxicity toleranceassays in yeast. Spots are three independent yeast clones for each transformation, plated at equivalentinitial cell densities (~5000 cells in 10mL) and incubated at 30�C on minimal galactose media for2 (Ge, As) or 5 (B) days. (d) Growth of each clone is also shown on unamended media (2 days) todemonstrate equivalent cell densities.
624 Functional Plant Biology J. E. Hayes et al.

showed few symptoms of Ge toxicity but, by comparison, leafblades and stems of the Clipper barley variety developed manynecrotic lesions when grown in the presence of Ge (Fig. 1).Necrosis first developed on the oldest leaves in a similar mannerto the development of leaf symptoms of B toxicity (Reid et al.2004). However, the distribution of necrotic lesions moreuniformly along the leaf blade in plants grown in toxic Ge,together with a much lower threshold of toxicity for Ge(Halperin et al. 1995), suggests that not all aspects of B andGe toxicities are the same. We mapped leaf symptoms of Getoxicity in a DH population derived from the crossClipper� Sahara 3771 and two loci for Ge toxicity tolerancewere identified (Fig. 2). A locus on Chromosome 6Hcorresponded to a QTL previously identified in this populationfor B toxicity tolerance (Jefferies et al. 1999), under whichHvNIP2;1 lies (Schnurbusch et al. 2010). HvNIP2;1 is anaquaporin which may be multifunctional, having reportedpermeability to both B and Si, as well as to As and Ge (Chibaet al. 2009; Schnurbusch et al. 2010). The reduced expressionof HvNIP2;1 in Sahara 3771 likely contributes to both a B anda Ge toxicity tolerance phenotype. A second QTL associatedwith Ge toxicity tolerance was identified on barley Chromosome2H. This peak co-located with another QTL for B toxicitytolerance. The B toxicity tolerance QTL on Chromosome 2His associated with reduced leaf symptom expression (Jefferieset al. 1999; Fig. 3), but the gene(s) underlying this locus are notyet known. The identification of the same QTL peak using Getolerance screening indicates that the underlying mechanismcould well involve an aquaporin related to HvNIP2;1. Analternative explanation may be that a more general stresstolerance mechanism is derived from this locus that regulatesthe initiation and development of necrosis. This could be testedby mapping leaf symptom development in response to otherstress elicitors such as plant disease, light stress or herbicides(e.g. Casano et al. 2001; Wu and von Tiedemann 2004).Importantly, identification of the 2H B tolerance locus usingGe indicates that Ge will be a particularly useful tool foridentifying the mechanism and gene(s) underlying this locusthat are responsible for B toxicity tolerance. A secondimportant finding was that there was no association betweenGe toxicity tolerance and the 4H B toxicity tolerance QTLpreviously identified in barley (HvBot1; Sutton et al. 2007).This establishes Ge as a means to dissect specific aspects ofB toxicity in the absence of the confounding effects of HvBot1.
To validate the location of the 2H B toxicity toleranceQTL identified by Jefferies et al. (1999), we re-screened theClipper� Sahara 3771 DH population in hydroponics at toxicconcentrations of B, and used digital imaging and imageanalysis (LemnaTec Scanalyser) to assess leaf necrosisobjectively. Previous identification of the QTL associated withleaf symptoms of B toxicity relied on visual assessment ofseedlings grown in boxes of field soil (Jefferies et al. 1999). Itis possible that the position of the QTL was affected by theheterogeneous nature of the soil or by interactions with soilsalinity (Rivandi et al. 2011). However, our re-analysisconfirmed the location of the 2H QTL as well as that of the4H B tolerance locus (HvBot1; Sutton et al. 2007; Fig. 3).Additionally, comparisons between visual scoring ofsymptoms and image analyses of necrosis for both B and Ge
toxicities determined that visual scoring can be a reliableassessment method for both traits (Fig. 4). In hydroponics, the2H B toxicity tolerance QTL peak for leaf symptom expression(LOD 5.7) was smaller than that obtained by Jefferies et al.(1999) (LOD 13.0), whereas the 4H QTL peak as determinedby leaf symptom expression in this study (LOD 9.2) was largerthan previously observed (LOD 3.7; Fig. 3). This informationsuggests that HvBot1 contributes more strongly to leaf symptomexpression of B toxicity in hydroponics than in soil-basedsystems, possibly due to the increased exposure andsubsequent movement of B into the root or shoot in plantsgrown in hydroponics compared with soil.
We undertook a phenotypic mutant screen to search for Getoxicity-tolerant mutants in an EMS-mutagenised population ofbarley (cv. Flagship). At least two of the 10 Ge-tolerant mutantlines also showed reduced leaf symptoms of B toxicity. Thesewill be assessed for linkage of the mutant phenotypes to eitherthe 2H or 6H B toxicity tolerance loci. Although none of themutants so far identified were found to have altered sequencesin the open reading frame of HvNIP2;1, Ge screening remainsa valid approach for identifying variants in this gene. Ge hasbeen used successfully to screen a rice mutant population formutations in OsNIP2;1 (Lsi1; Ma et al. 2006), suggesting thatmutations in HvNIP2;1 may also not be lethal.
We also used Ge toxicity to test for the possibility that wheatcarries NIP2;1- or barley 2H locus-related B toxicity tolerance.Considerable variation in Ge toxicity tolerance was observedbetween genotypes, but this did not obviously relate to reporteddifferences in either B toxicity or B deficiency tolerance inwheat.For example, Halberd (B toxicity-tolerant) and Cranbrook(B toxicity-intolerant; Jefferies et al. 2000) had similar levelsof leaf necrosis when treated with toxic levels of Ge (Fig. 5).None of the wheat genotypes were as tolerant to Ge toxicity asSahara 3771 barley, suggesting that no genotype in this screenpossessed both aquaporin- and barley Chromosome 2H-relatedB tolerance traits.
Finally, we used Ge to test the effect of specific mutationsin HvNIP2;1 on transport selectivity, using yeast phenotypingassays in a similar manner to Bienert et al. (2008). It is predictedthat residues at positions H2 and H5 in the aromatic/arginine(ar/R) selectivity filter are primarily responsible for transportselectivity of aquaporins belonging to the NIP family (Wallaceand Roberts 2005). Therefore, we substituted the amino acidsat these positions for residues with larger side chains to try toconstrict this filter. Replacement of glycine at position H2with alanine (Gly88Ala) or cysteine (Gly88Cys) disrupted theapparent ability of HvNIP2;1 to transport both Ge and B,whereas selectivity for As was maintained. A further threemutations (Gly88Val, Gly88Ala /Ser207Val and Ala132Thr)disrupted permeability to all three metalloids tested. Althoughwe did not test these mutations for effects on HvNIP2;1expression, our results are consistent with a recent study byMitani-Ueno et al. (2011), who characterised mutants ofOsNIP2;1 and AtNIP5;1 in Xenopus laevis oocytes and foundthat As permeability was less affected by changes in the ar/Rfilter than permeability to the other tested metalloids. In ourexperiments, the mutation Ala132Thr was generated to re-create a mutant of OsNIP2;1 that has disrupted Si and Ge (Maet al. 2006), and As (Ma et al. 2008) transport, and it served as
Germanium and boron toxicities in cereals Functional Plant Biology 625

a useful control. Although gene translation and protein functionmay be different in yeast compared with plants, the effect of themutation Ala132Thr was as predicted, resulting in disruptedpermeability to B, Ge and As. Importantly, we found that noneof the mutant HvNIP2;1 proteins discriminated between B andGe, supporting the idea that Ge is indeed a suitable analogue forB in studies relating to HvNIP2;1.
In this report, we have shown that the metalloid element Geis a particularly useful tool for targeting specific aspects of Btoxicity tolerance in plants, including that related to aquaporinsand also to reduced leaf symptom expression. Ge has potentialapplications for identification of natural and induced variationin HvNIP2;1 and related aquaporins, in isolation fromconfounding effects of Bot1-like transporters. We believe thatGe may also be critical for identifying the mechanism andgene(s) responsible for reduced leaf symptoms of borontoxicity, which underlie the B toxicity tolerance locus onbarley Chromosome 2H.
Acknowledgements
Research at the Australian Centre for Plant Functional Genomics is jointlyfunded by the Australian Research Council, the Grains Research andDevelopment Corporation, the University of Adelaide and the SouthAustralian Government. The authors gratefully acknowledge technicalassistance from Ms Alison Hay, instrument and software support fromLemnaTec GmbH (Würselen, Germany; http://www.lemnatec.com/), andcritical reading of the manuscript by Joanne Tilbrook.
References
Bhattacharjee H, Mukhopadhyay R, Thiyagarajan S, Rosen BP (2008)Aquaglyceroporins: ancient channels for metalloids. Journal ofBiology 7, 33. doi:10.1186/jbiol91
Bienert GP, Thorsen M, Schüssler MD, Nilsson HR, Wagner A, Tamás MJ,JahnTP (2008)A subgroupof plant aquaporins facilitate thebi-directionaldiffusion of As(OH)3 and Sb(OH)3 across membranes. BMC Biology 6,26. doi:10.1186/1741-7007-6-26
Cakmak I, Kurz H, Marschner H (1995) Short-term effects of boron,germanium and high light intensity on membrane permeability inboron deficient leaves of sunflower. Physiologia Plantarum 95, 11–18.doi:10.1111/j.1399-3054.1995.tb00801.x
Cartwright B, Zarcinas BA, Mayfield AH (1984) Toxic concentrations ofboron in a red-brown earth at Gladstone, South Australia. AustralianJournal of Soil Research 22, 261–272. doi:10.1071/SR9840261
Casano LM, Martìn M, Sabater B (2001) Hydrogen peroxide mediates theinduction of chloroplastic Ndh complex under photooxidative stress inbarley. Plant Physiology 125, 1450–1458. doi:10.1104/pp.125.3.1450
Chiba Y, Mitani N, Yamaji N, Ma JF (2009) HvLsi1 is a silicon influxtransporter in barley.ThePlant Journal57, 810–818. doi:10.1111/j.1365-313X.2008.03728.x
Gietz RD, Woods RA (2002) Transformation of yeast by the Liac/SS carrierDNA/PEG method. Methods in Enzymology 350, 87–96. doi:10.1016/S0076-6879(02)50957-5
Halperin SJ, Barzilay A, CarsonM, Roberts C, Lynch J, Komarneni S (1995)Germanium accumulation and toxicity in barley. Journal of PlantNutrition 18, 1417–1426. doi:10.1080/01904169509364991
Ishii T,MatsunagaT, Iwai H, Satoh S, Taoshita J (2002)Germaniumdoes notsubstitute for boron in cross-linking of rhamnogalacturonan II in pumpkincell walls. Plant Physiology 130, 1967–1973. doi:10.1104/pp.009514
Jefferies SP, Barr AR, Karakousis A, Kretshmer JM, Manning S, ChalmersKJ, Nelson JC, Islam AKMR, Langridge P (1999) Mapping ofchromosome regions conferring boron toxicity tolerance in barley
(Hordeum vulgare L.). Theoretical and Applied Genetics 98,1293–1303. doi:10.1007/s001220051195
Jefferies SP, Pallotta MA, Paull JG, Karakousis A, Kretshmer JM, ManningS, Islam AKMR, Langridge P, Chalmers KJ (2000) Mapping andvalidation of chromosome regions conferring boron toxicity tolerancein wheat (Triticum aestivum L.). Theoretical and Applied Genetics 101,767–777. doi:10.1007/s001220051542
Joehanes R, Nelson JC (2008) QGene 4.0, an extensible Java QTL-analysisplatform. Bioinformatics 24, 2788–2789. doi:10.1093/bioinformatics/btn523
Karakousis A, Barr AR, Kretschmer JM, Manning S, Jefferies SP, ChalmersKJ, Islam AKM, Langridge P (2003) Mapping and QTL analysis of thebarley population Clipper�Sahara. Australian Journal of AgriculturalResearch 54, 1137–1140. doi:10.1071/AR02180
Loomis WD, Durst RW (1991) Boron and cell walls. In: ‘Current topics inplant biochemistry and physiology, Vol. 10’. (Eds DD Randall, DGBlevins, CD Miles) pp. 149–178. (University of Missouri: Columbia,MO)
Ma JF, Goto S, Tamai K, Ichii M, Wu GF (2002) A rice mutant defective inSi uptake. Plant Physiology 130, 2111–2117. doi:10.1104/pp.010348
Ma JF, Tamai K, Yamaji N, Mitani N, Konishi S, Katsuhara M, Ishiguro M,Murata Y, Yano M (2006) A silicon transporter in rice. Nature 440,688–691. doi:10.1038/nature04590
Ma JF, Yamaji N, Mitani N, Tamai K, Konishi S, Fujiwara T, Katsuhara M,Yano M (2007) An efflux transporter of silicon in rice. Nature 448,209–212. doi:10.1038/nature05964
Ma JF, Yamaji N, Mitani N, Xu XY, Su YH, McGrath SP, Zhao FJ (2008)Transporters of arsenite in rice and their role in arsenic accumulation inrice grain.Proceedings of the National Academy of Sciences of the UnitedStates of America 105, 9931–9935. doi:10.1073/pnas.0802361105
Meharg AA (2004) Arsenic in rice – understanding a new disaster forSouth-east Asia. Trends in Plant Science 9, 415–417. doi:10.1016/j.tplants.2004.07.002
Mitani-Ueno N, Yamaji N, Zhao FJ, Ma JF (2011) The aromatic/arginineselectivity filter of NIP aquaporins plays a critical role in substrateselectivity for silicon, boron and arsenic. Journal of ExperimentalBotany 62, 4391–4398. doi:10.1093/jxb/err158
NikolicM,NikolicN,LiangYC,KirkbyEA,RömheldV (2007)Germanium-68 as an adequate tracer for silicon transport in plants. Characterization ofsilicon uptake in different crop species. Plant Physiology 143, 495–503.doi:10.1104/pp.106.090845
Paull JG, Rathjen AJ, Langridge P (1995) Location of genes controllingboron tolerance of wheat. In ‘Proceedings of the 8th International WheatGenetics Symposium, Beijing, 19–24 July 1993’. (Eds ZS Li, ZY Xin)pp. 1065–1069. (China Agricultural Scientech Press)
Rains DW, Epstein E, Zasoski RJ, Aslam M (2006) Active siliconuptake by wheat. Plant and Soil 280, 223–228. doi:10.1007/s11104-005-3082-x
Reid RJ, Hayes JE, Post A, Stangoulis JCR, Graham RD (2004) A criticalanalysis of the causes of boron toxicity in plants. Plant, Cell &Environment 27, 1405–1414. doi:10.1111/j.1365-3040.2004.01243.x
Rivandi J, Miyazaki J, Hrmova M, Pallotta M, Tester M, Collins NC (2011)A SOS3 homologue maps to HvNax4, a barley locus controlling anenvironmentally sensitive Na+ exclusion trait. Journal of ExperimentalBotany 62, 1201–1216. doi:10.1093/jxb/erq346
SchnurbuschT,CollinsNC,EastwoodRF,SuttonT, JefferiesSP,LangridgeP(2007) Fine mapping and targeted SNP survey using rice–wheat genecolinearity in the region of the Bo1 boron toxicity tolerance locus ofbread wheat. Theoretical and Applied Genetics 115, 451–461.doi:10.1007/s00122-007-0579-0
Schnurbusch T, Hayes J, HrmovaM, BaumannU, Ramesh SA, Tyerman SD,Langridge P, Sutton T (2010) Boron toxicity tolerance in barley throughreduced expression of the multifunctional aquaporin HvNIP2;1. PlantPhysiology 153, 1706–1715. doi:10.1104/pp.110.158832
626 Functional Plant Biology J. E. Hayes et al.

Sutton T, Baumann U, Hayes J, Collins NC, Shi BJ, Schnurbusch T, Hay A,Mayo G, Pallotta M, Tester M, Langridge P (2007) Boron-toxicitytolerance in barley arising from efflux transporter amplification.Science 318, 1446–1449. doi:10.1126/science.1146853
Takahashi E, Syo S, Miyake Y (1976) Effect of germanium on the growthof plants with special reference to the silicon nutrition (parts 1–3).Journal of the Science of Soil and Manure, Japan 47, 183–190.
Takano J,NoguchiK,YasunoriM,KobayashiM,GajdosZ,MiwaK,HayashiH, Yoneyama T, Fujiwara T (2002) Arabidopsis boron transporter forxylem loading. Nature 420, 337–340. doi:10.1038/nature01139
Takano J, Wada M, Ludewig U, Schaaf G, von Wirén N, Fujiwara T (2006)The Arabidopsis major intrinsic protein NIP5;1 is essential for efficientboron uptake and plant development under boron limitation. The PlantCell 18, 1498–1509. doi:10.1105/tpc.106.041640
Wallace IS, Roberts DM (2005) Distinct transport selectivity of twostructural subclasses of the nodulin-like intrinsic protein family ofplant aquaglyceroporin channels. Biochemistry 44, 16826–16834.doi:10.1021/bi0511888
Wu YX, von Tiedemann A (2004) Light-dependent oxidative stressdetermines physiological leaf spot formation in barley. Phytopathology94, 584–592. doi:10.1094/PHYTO.2004.94.6.584
Germanium and boron toxicities in cereals Functional Plant Biology 627
www.publish.csiro.au/journals/fpb




















