
Geomorphic dynamics of floodplains: ecological implications and a potential modelling strategy
21
Geomorphic dynamics of floodplains: ecological implications and a potential modelling strategy KEITH RICHARDS, JAMES BRASINGTON and FRANCINE HUGHES Department of Geography, University of Cambridge, Cambridge, U.K. SUMMARY 1. The dynamics of channel migration and floodplain renewal constitute an important control of the ecological diversity of river corridors. Restoration initiatives should therefore assess whether these dynamics must be reinstated in order to address the cause rather than the symptoms of floodplain biodiversity decline. 2. Restoration of reach-scale dynamism in rivers where this is a natural behavioural process will restore smaller-scale geomorphological and sedimentological processes that encourage vegetation regeneration, but may require catchment-scale management of material flows. 3. Channel dynamics depend on the style of river–floodplain interaction, and this may be summarised in qualitative, classificatory, sedimentological models of floodplain architec- ture that have been somewhat neglected in the ecological literature. 4. One approach to the assessment of floodplain biodiversity and its restoration would be through the development of simulation models based on specified channel styles, and involving simplified hydrodynamics and successional changes. Such models, currently the subject of research as a spin-off from modelling studies of landscape evolution, would permit evaluation of the consequences for ecological diversity of implementing various management options that may affect the dynamics of channel migration. Keywords: braiding, channel dynamics, meandering, modelling, reach scale, succession Introduction: ecological concepts and management Interest in integrated restoration of rivers and their floodplains has developed in response to declining floodplain biodiversity, in spite of the naturally high level of ecological diversity in riparian corridors (Naiman, De ´camps & Pollock, 1993). This decline reflects the influences of flood control, flow manage- ment, channelisation and land use changes in these environments, all of which have separated the river from its riparian zone and floodplain. Responses have included both attempts to restore connectivity across the river–floodplain, and direct attempts to restore ecosystem state, for example of floodplain woodland (Peterken & Hughes, 1995). The practices of such restoration have often depended less on the applica- tion of theoretical principles than on empirical experience, which may risk failure through lack of understanding of differences between reaches and sites. This paper outlines an approach that addresses some of the theoretical underpinnings required by the restoration objective. It begins with an evaluation of diversity concepts, which concludes that the channel dynamics often constitute an essential control of floodplain ecological diversity. These dynamics are then shown to reflect the channel–floodplain associ- ation, as classified in sedimentological models, which have been neglected in ecology. Finally, an approach is discussed, based on simulation modelling (cf. Malanson, 1993; pp. 218–219) of the relationships between channel dynamics and biodiversity for par- ticular channel types. The form of modelling envis- aged, which is the subject of our ongoing research, is a Correspondence: Keith Richards, Department of Geography, University of Cambridge, Cambridge CB2 3EN, U.K. E-mail: [email protected] Freshwater Biology (2002) 47, 559–579 Ó 2002 Blackwell Science Ltd 559
Transcript of Geomorphic dynamics of floodplains: ecological implications and a potential modelling strategy

Geomorphic dynamics of floodplains: ecologicalimplications and a potential modelling strategy
KEITH RICHARDS, JAMES BRASINGTON and FRANCINE HUGHES
Department of Geography, University of Cambridge, Cambridge, U.K.
SUMMARY
1. The dynamics of channel migration and floodplain renewal constitute an important
control of the ecological diversity of river corridors. Restoration initiatives should therefore
assess whether these dynamics must be reinstated in order to address the cause rather than
the symptoms of floodplain biodiversity decline.
2. Restoration of reach-scale dynamism in rivers where this is a natural behavioural
process will restore smaller-scale geomorphological and sedimentological processes that
encourage vegetation regeneration, but may require catchment-scale management of
material flows.
3. Channel dynamics depend on the style of river–floodplain interaction, and this may be
summarised in qualitative, classificatory, sedimentological models of floodplain architec-
ture that have been somewhat neglected in the ecological literature.
4. One approach to the assessment of floodplain biodiversity and its restoration would be
through the development of simulation models based on specified channel styles, and
involving simplified hydrodynamics and successional changes. Such models, currently the
subject of research as a spin-off from modelling studies of landscape evolution, would
permit evaluation of the consequences for ecological diversity of implementing various
management options that may affect the dynamics of channel migration.
Keywords: braiding, channel dynamics, meandering, modelling, reach scale, succession
Introduction: ecological concepts
and management
Interest in integrated restoration of rivers and their
floodplains has developed in response to declining
floodplain biodiversity, in spite of the naturally high
level of ecological diversity in riparian corridors
(Naiman, Decamps & Pollock, 1993). This decline
reflects the influences of flood control, flow manage-
ment, channelisation and land use changes in these
environments, all of which have separated the river
from its riparian zone and floodplain. Responses have
included both attempts to restore connectivity across
the river–floodplain, and direct attempts to restore
ecosystem state, for example of floodplain woodland
(Peterken & Hughes, 1995). The practices of such
restoration have often depended less on the applica-
tion of theoretical principles than on empirical
experience, which may risk failure through lack of
understanding of differences between reaches and
sites. This paper outlines an approach that addresses
some of the theoretical underpinnings required by the
restoration objective. It begins with an evaluation of
diversity concepts, which concludes that the channel
dynamics often constitute an essential control of
floodplain ecological diversity. These dynamics are
then shown to reflect the channel–floodplain associ-
ation, as classified in sedimentological models, which
have been neglected in ecology. Finally, an approach
is discussed, based on simulation modelling (cf.
Malanson, 1993; pp. 218–219) of the relationships
between channel dynamics and biodiversity for par-
ticular channel types. The form of modelling envis-
aged, which is the subject of our ongoing research, is a
Correspondence: Keith Richards, Department of Geography,
University of Cambridge, Cambridge CB2 3EN, U.K.
E-mail: [email protected]
Freshwater Biology (2002) 47, 559–579
Ó 2002 Blackwell Science Ltd 559

simplified, coupled hydrodynamic-ecological model
which captures the dynamics of channel pattern
evolution and relevant plant succession trends, and
which could eventually be used to explore the impacts
of river management practices on floodplain biodi-
versity.
An initial conceptual issue to consider is the role of
channel dynamics in relation to biodiversity, which
itself, as Table 1 indicates, depends on several related
factors including species richness (SR) and relative
abundance (the latter also is needed because rare
species may be missed when sampling to estimate an
SR index). Species richness reflects the number of
species per habitat (the a-diversity), and the number
of habitats (N) in a region (Whittaker, 1960). If the
‘region’ is a floodplain reach, the number of habitats
reflects the geomorphological dynamics and the sedi-
mentological diversity. Total SR is, however, not
simply a sum of values of a across the N habitats, as
some species have wide tolerance, and appear in
several habitats; an ‘inverse’ b-diversity is thus also
required. There are then other aspects of within-
species diversity that these measures do not consider.
One is the diversity of age structure. In geomorpho-
logically dynamic environments, erosion and depos-
ition processes will locally destroy older vegetation,
and create new surfaces for colonisation by pioneer
species. Finally, diversity reflects the gene pool in a
particular environment, and the number of distinct
clones of each species. Despite the evident multifactor
definition required for ‘biodiversity’, there is strong
evidence from river environments that physical hab-
itat heterogeneity plays a crucial role (Ward &
Tockner, 2001). This heterogeneity depends on the
channel dynamics; and these dynamics in turn
encourage age structure diversity (Salo et al., 1986;
Nilsson et al., 1991). Thus, in developing restoration
proposals for floodplain environments, a valuable tool
would be one allowing assessment of the consequence
for habitat diversity of rates of river migration and
sedimentary turnover. Such a tool would permit
evaluation of the consequence of restoring channel
dynamics for the diversity of habitats and vegetation
communities, while also indicating the sustainability
of this diversity over time, and revealing the cases
where lateral channel stability inhibits turnover and
suppresses diversity as floodplain vegetation becomes
more uniform over time.
A second issue is that river ecology has spawned
several qualitative concepts to account for aspects of
spatial variation in ecology, but that do not provide a
rigorous framework for the analysis of restoration
options. The longitudinal ‘river continuum concept’
(Vannote et al., 1980) assumes longitudinal resource
gradients, with maximum biodiversity in the middle
reaches of rivers. Headwater streams have low ther-
mal heterogeneity, light levels and nutrient status,
while lowland reaches have high turbidity and low
oxygen levels; optimal conditions for biodiversity
occur in middle reaches. These, however, are also
characterised, at least in medium-sized rivers, by high
stream power (Lawler, 1992), implying greater pro-
pensity for disturbance, and contradicting another
qualitative model, the ‘intermediate disturbance
hypothesis’ (Connell, 1978). The intermediate distur-
bance hypothesis also involves a trade-off, between
competitive dominant species which monopolise sta-
ble habitats, and the few fugitive species that survive
high levels of instability. Maximum diversity is thus
expected in locations with intermediate levels of
disturbance. A third model emphasises lateral
connectivity, particularly through the ‘flood pulse
concept’ (Junk, Bayley & Sparks, 1989; Middleton,
1999), in which seasonal variation in river water level
controls the recruitment of obligate riparian species.
Without this flow variation, there is progressive
change to terrestrial (dry) species and communities.
Less extreme flow variations also influence the lateral
distribution of moisture, nutrients and sediments
(Tockner, Malard & Ward, 2000). The flood pulse is
also linked to the dynamics of lateral channel migra-
tion, which renew habitat for colonisation and regen-
eration, and are themselves flood-related. Channel
Table 1 Various measures of ecological diversity
Measure Definition
a-Diversity Species numbers in each
habitat
b-Diversity Proportion of habitats in
which a given species is
present
c-Diversity Total number of species in a
region
Habitat diversity Number of habitats in an area
Age diversity Frequency distribution of ages
of a species in an area or
habitat
Genetic diversity A measure of clonal variation in
a species
560 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

dynamics generate diverse habitat patch sizes, max-
imising the niches for colonisation by both edge and
interior species. Laterally, stable rivers tend to have
larger habitat patches, and low diversity because of
the limited extent of edge habitat. Highly disturbed
river environments exclude interior species (Ward,
Tockner & Schiemer, 1999). Thus the intermediate
disturbance hypothesis and the flood pulse concept
are closely interrelated, through their mutual depend-
ence on the channel dynamics.
These qualitative concepts provide useful organis-
ing principles for the diverse data from ecological case
studies. However, they lack the quantitative reliability
needed to provide a theoretical framework for prac-
tical management. Empirical data from field sites are
influenced by numerous causal factors, the controls of
observed longitudinal and lateral spatial patterns are
often unclear, and it is therefore risky drawing
management conclusions. However, the channel
dynamics are identified in the foregoing discussion
as a key control both of biodiversity and of spatial
pattern. This is because (i) biodiversity may be taken
at the river reach scale to be strongly related to habitat
diversity; (ii) habitat diversity reflects channel migra-
tion processes and (iii) the channel dynamics integrate
flood pulse and disturbance influences on the renewal
of succession and the regeneration of species. Thus,
one strategy to improve the capacity for successful
restoration initiatives is to develop simulation models
of the interdependent dynamics of channels and
floodplain vegetation that permit scenario evaluation.
This paper provides a review of the background to
recently initiated research by the authors to develop
such models, and to examine the practical potential
for such a modelling approach. In discussing this
background, the paper also seeks to identify know-
ledge gaps, and to encourage the closer integration of
the geomorphological and vegetation sciences in the
development of integrative modelling approaches to
riverine landscapes and river management.
Scales, processes and ecological management
As noted above, there are different approaches to
restoration, a particular issue being whether to focus
on restoring ‘products’ (species or ecosystems)
directly, or on ‘processes’, which then generate the
desired products in a quasi-natural manner. The latter
may represent a more sustainable approach requiring
less cumulative intervention. However, processes
vary in their significance with time and space scales,
and this places a premium on identification of the
significant processes. In Fig. 1, a range of linked,
scale-dependent fluvial and ecological processes is
suggested across a hierarchy of fluvial landscape units
(cf. Brierley & Fryirs, 2000). The ecological processes
identified are those relevant to the riparian ecology,
although variants may be equally applicable to the
aquatic ecology. The diagram implies feedback
amongst scales, with the larger spatial scales both
determining and being determined by processes at
smaller scales. A critical scale in this figure, however,
is the ‘reach’ scale, where the channel pattern
dynamics take place. When these dynamics operate
naturally, they are associated with sediment transport
processes that at smaller scales create bar-forms,
bedforms and particle motions that sustain ecological
dynamics at equivalent scales. At the reach scale, the
dominant fluvial and ecological processes seem to be
fundamentally congruent – involving both the
dynamic changes of meandering or braiding channel
patterns, and the patch dynamics of ecosystem turn-
over and maintenance (Pickett & White, 1985; Poole,
2002; Ward et al., 2002). These processes contribute to
biodiversity as measured at various levels – habitat
diversity, species diversity within patches, diverse age
structure between patches. Natural processes of flu-
vial disturbance thus create a mosaic of alluvial
surfaces with vegetation stands in different succes-
sional stages.
The underlying determinant of these processes is
the (intermediate) disturbance created by the flood
pulse. Usually in the ecological literature, the flood
pulse is concerned with water level variation, and
with moisture, nutrient and sediment supply factor,
which together provide regeneration opportunities
influential at a scale similar to that of an individual
plant. However, the flood pulse is also responsible for
erosion and sedimentation process, which provide
such opportunities at the scale of habitat patches.
Erosion and sedimentation may, however, occur at
particular locations in different events (Hughes, 1997);
thus the extension of the ‘flood pulse’ to the ‘flow
pulse’ concept (Tockner et al., 2000) is also a natural
concern for the fluvial geomorphologist. Furthermore,
a flood of a particular magnitude has different effects
on channel dynamics depending on the initial condi-
tion of the channel pattern prior to the event; for
Geomorphic dynamics of floodplains 561
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

example, such an event may fail to cut off a bend until
bank erosion has created sufficient sinuosity. Critic-
ally, however, the channel dynamics lead to new
surfaces for colonisation and regeneration; and mod-
elling tools are required to enhance understanding of
the ecological consequences of reach-scale channel
dynamics, and of their alteration by river manage-
ment practices.
Attempts have been made to convert the flood pulse
concept into a practical flow management tool, for
example in the ‘recruitment box’ method of Mahoney
& Rood (1998). This method is based on the flood as a
control over species regeneration, rather than as a
control either of ecosystem or of channel dynamics.
The recruitment box defines a seasonal period for
viable seedling establishment, when the timing of
seed release coincides with an appropriate range of
variation in stage and floodplain water table range,
and a rate of decline of the water table that seedlings
are capable of surviving (Fig. 2a). Such a scheme
clearly works best in rivers with a predictable spring
flood related to a measurable winter snow-pack. The
scientific basis for the definition of the ‘recruitment
box’ can, of course, be further improved through
experimental investigations of the tolerance of indi-
vidual species. Understanding is needed of such
phenomena as the optimal timing relationships
between seed production and seasonal flooding, the
critical soil water potentials for seed germination, and
the critical rates for seedling performance of soil water
decline following flooding (Hughes et al., 2000;
Karrenberg, Edwards & Kollmann, 2002). In addition,
there is room for elaborating the concept in relation to
plant communities or habitats rather than to individ-
ual species, as well as for improving the capacity to
simulate channel, habitat and ecosystem dynamics in
response to less predictable flood histories than those
fed by snowmelt.
Nevertheless, the ‘recruitment box’ idea, and its
scientific basis, offer the possibility of a predictive
model comparable with the aquatic habitat model
PHABSIM (Milhous, Updike & Schneider, 1989), but
applicable to the germination and survival of riparian
seedlings. This could be used to underpin flow
Fig. 1 A hierarchical representation of the correlated fluvial (bold text) and ecological (normal text) processes at different scales
in the fluvial environment; the highlighted reach scale processes are hypothesised as those whose manipulation is likely to be most
successful in conservation or restoration projects, because the processes are most strongly linked.
562 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

management practices that are ecologically beneficial
for riparian ecosystems, as is increasingly required
where river and flow management acknowledge the
needs of ecosystem maintenance (Petts, 1996). In
South Africa, for example, a recent Water Act makes
provision for defining and reserving the water
requirements for ecosystem conservation (Milton,
Gilvear & Hooper, 1995; Moon et al., 1997). It remains
a challenge to establish practical principles for
multi-objective ecological flow management, and to
implement these. As Fig. 2b illustrates various flow
requirements exist for different elements of a land-
scape consisting of a range of ecosystems (Hill &
Platts, 1991). However, a key point which is recog-
nised in flow allocation methods such as the Building
Block Methodology (King & Louwe, 1998) is that
different ecosystems require managed flows at differ-
ent times, and with different frequencies. Thus, the
flood pulse necessary for successful establishment
of riparian seedlings is only required intermittently,
and flow management in other years can be selected
to optimise other ecological processes. Where
multi-objective management broadens to include res-
toration of channel dynamism, additional assessment
tools will be required. Simulation models capable of
providing insights into the rates and patterns of
renewal of habitat as a result of actively managed
lateral channel migration would therefore add to the
battery of techniques available to those concerned
with riverine habitat conservation or restoration.
Channel dynamics, habitats
and floodplain ecology
The habitat diversity of floodplain environments is
partly dependent on dynamic characteristics of the
river system which express themselves at the reach
scale (Fig. 1). These include the type of channel and
the suite of sedimentary environments created by its
evolution; the processes of erosion and deposition that
drive the channel evolution; the rates of bank erosion,
channel migration and floodplain sediment turnover;
local patterns of erosion and sedimentation and their
effects on plant recruitment; and the effects of flow
control and channel management. There is often a
close interdependence between the channel dynamics
that renew habitat and succession processes, and the
ecological dynamics – as when the succession is partly
dependent on sedimentation accelerated by the pres-
ence of vegetation, or tree-throws cause erosion and
channel avulsion. Hence, in Fig. 1, the suggestion
emerges that the reach scale is that at which the most
intimate mutual association occurs between channel
and vegetation (patch) dynamics, and the greatest
potential for biogeomorphological management
exists; and that although other processes may interact
at both larger and smaller scales, they are favoured by
particular reach-scale circumstances.
Fluvial styles and habitat diversity
The balance of processes of channel pattern change,
and their rates, depend on channel pattern type, and
classification is a precursor to analysis of the relation-
ship between channel and ecological dynamics. A
useful classification in the habitat context is provided
by the facies models of fluvial architectural style
developed by Miall (1985). These models relate closely
to the classification of floodplains generated by
Nanson & Croke (1992) (Table 2), with the advantage
of explicit representation of the sedimentary elements
Fig. 2 (a) The ‘recruitment box’ concept; seedling establishment
occurs when seed release coincides with a favourable stage
range and rate of stage decline (after Mahoney & Rood, 1998).
(b) Minimum monthly flow requirements for fisheries, channel
maintenance, riparian habitat and valley maintenance, in the
Salmon River, Idaho (after Hill & Platts, 1991).
Geomorphic dynamics of floodplains 563
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

that define the range of habitats found in different
river environments. Each fluvial style has a set of
characteristic elements (Table 3), each of which is
composed of a range of lithofacies at a smaller scale.
This is ecologically significant, as a generally gravel-
based element can contain locally abundant fine
sediment in patches, and these may be important for
moisture retention and seed germination (Petts et al.,
2000). The lithofacies are various forms of deposit of
gravel, sand and fines, with various small-scale
sedimentary structures (bedforms) such as imbrica-
tion, cross-bedding and lamination (Miall, 1996, p. 79).
In common with the Nanson & Croke (1992)
classification, the fluvial styles in Miall’s (1985) clas-
sification vary with stream power and sediment
calibre. Miall (1985) identifies 16 distinctive fluvial
styles, and these range from low-power, mud-sand
dominated anastomosing rivers (class C2b in the
Nanson & Croke classification; Table 2), through
sandy, single-thread, high-sinuosity meandering riv-
ers (class B3b) to high power, multithread, braided
rivers (B1). Fig. 3 illustrates this spectrum of fluvial
styles, extracting from Miall’s continuum a set of
styles representing the range of basic geometries and
sedimentary facies. Fig. 3a shows the general spatial
structure of sedimentary environments characteristic
of an anastomosing river (Miall’s Model 8 in Table 3),
with extensive areas of overbank floodplain fines (FF),
Table 2 A classification of floodplains based on Nanson &
Croke (1992)
Class Fluvial style
A High energy, non-cohesive floodplains
A1 Confined, coarse-textured floodplains
A2 Confined, vertical-accretion floodplains
A3 Unconfined, vertical-accretion floodplains
A4 Cut-and-fill floodplains
B Medium-energy, non-cohesive floodplains
B1 Braided river floodplains
B2 Wandering gravel-bed river floodplains
B3 Meandering river, lateral-migration floodplains
B3a Lateral-migration, non-scrolled floodplains
B3b Lateral-migration, scrolled floodplains
B3c Lateral-migration/backswamp floodplains
B3d Lateral-migration, counterpoint floodplains
C Low-energy, cohesive floodplains
C1 Laterally stable, single-channel floodplains
C2 Anastomosing river floodplains
C2a Anastomosing river, organic-rich floodplains
C2b Anastomosing river, inorganic floodplains
Table 3 Common fluvial styles and their main architectural elements (after Miall, 1985, 1996)
Classification Architectural
no. Fluvial style Sedimentary facies element*
Gravel-dominated rivers
1 Gravel braided with sediment Gravel, minor sand SG (GB, SB)
gravity flows
2 Shallow gravel braided Gravel, minor sand GB (SB)
3 Deep gravel braided Gravel, minor sand, fines GB, SB, DA (FF)
Gravel wandering Gravel, minor sand, fines GB, DA, LA (FF)
4 Gravel meandering Gravel, minor sand, fines GB, DA, LA (FF)
Sand-dominated, high sinuosity rivers
5 Gravel-sand meandering Sand, pebbly SB, LA, FF (GB)
6 Sandy meandering Sand, minor fines LA, SB, FF, LV, CR, CS, FF (CH)
Ephemeral sandy meandering Sand, minor fines LA, SB (FF)
7 Fine-grained meandering Fine sand, silt, mud LA, SB, FF, LV, CR, CS, FF (CH)
8 Anastomosing Sand, fines SB, CH (LA), FF, LV, CR, CS
Sand-dominated, low-sinuosity rivers
Low sinuosity braided- Sand DA, LA, SB, FF
meandering with alternate bars
9 Shallow perennial braided rivers Sand SB (FF)
10 Deep perennial braided rivers Sand, minor fines DA, LA, SB (FF)
High energy sand-bed braided Sand, minor fines DA, SB, HO (FF)
11 Sheetflood distal braided Sand, minor fines SB (FF)
12 Flashy ephemeral sheetflood Sand, minor fines LS (FF)
*CH: channels, LS: laminated sand sheet, GB: gravel bars and bedforms, FF: overbank fines, SB: sandy bedforms, LV: levee, DA:
downstream-accretion macroform, CR: crevasse channel, LA: lateral-accretion macrofrom, CS: crevasse splay, HO: scour hollows,
CH(FF): abandoned channel, SG: sediment gravity flows.
564 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

coarser-grained areally distributed crevasse splay
deposits (CS), linear channel fill deposits (CS) and
occasional areas of lateral accretion deposit (LA).
Higher-energy meandering rivers with mainly sandy
bed material have the element structure suggested by
Fig. 3b (Miall’s Model 6), with similar sedimentary
elements in different proportions and spatial associ-
ations to those in Model 8, and with more extensive
lateral accretion deposits. Furthermore, they may
have additional features such as levees (LV) and
abandoned channel fills [CH(FF)], depending on the
sediment load character and the rates of bank erosion,
bend migration and bend cut-off. Fig. 3c shows
Miall’s Model 3, the deep, gravel-bed river with
well-defined topographic levels which are flooded
with different frequency. The dominant sedimentary
elements in this case are gravel bars and bedforms
(GB), sandy bedforms (SB), and overbank fines (FF).
Again, some other elements may occur (channel fills,
lateral and downstream accretion macro-form depos-
its) in specific examples.
Channel dynamics and biodiversity
Within each of these channel types, the dominant
processes of sediment transport, erosion and depos-
ition result in a particular spatial organisation of
sedimentary units, and thus create a habitat mosaic. In
anastomosing channels (Fig. 3a), strong interactions
may occur between fluvial and ecological processes,
because when an anastomosing floodplain is forested
(for example, the Gearagh on the River Lee in County
Cork, Ireland; Brown, Stone & Harwood, 1995), tree-
throws may play an important role in creating
crevasse splays, encouraging avulsion, causing the
abandonment of channels which subsequently infill,
and providing patch-regeneration opportunities. In a
single-thread meandering river, the dominant fluvial
processes are those of bend migration (which is
Fig. 3 A spectrum of fluvial styles, based on Miall (1985, 1996).
(a) An anastomosing river. (b) A sand-bed meandering river.
(c) A deep, gravel-bed braided river with well-defined topo-
graphic levels (printed with permission from Springer-Verlag).
Fig. 4 The vegetation mosaic of the lower Ain River, France
(from a map created by Amoros et al., 1986). The figure
illustrates the habitat complexity and diversity created by
the dynamics of river meander migration. The horizontal line at
the bottom is 1 km in length (printed with permission from
the authors).
Geomorphic dynamics of floodplains 565
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

associated with scroll bar deposition), bend cutoff
(associated with sedimentation in cut-off bends), and
the progressive in-fill of ox-bow lakes. The habitat
mosaic resulting from these processes includes levees,
back-swamps and crevasse splays. This results in high
levels of habitat diversity because of the geomorpho-
logical, topographic, sedimentological and edaphic
diversity, and also because of the diversity of ages of
(particular examples of) a given physical feature
within a reach. The processes of bank erosion, scroll
bar deposition and point bar growth (Nanson &
Beach, 1977; Salo et al., 1986; Shankman, 1993; Ward,
1998) have received most attention as processes of
habitat renewal and diversification. In particular,
scroll bars (see Fig. 3b) result in significant topo-
graphic differentiation between the resulting ridges
and swales on the surface of point bar complexes,
with a strong age structure amongst the trees on
successive ridges (Hickin, 1974). However, channel
avulsion, channel abandonment and abandoned chan-
nel in-fill are also significant factors in the generation
of a habitat mosaic. Fig. 4, an extract from a map
created by Amoros et al. (1986), is an excellent
example of this mosaic from the lower Ain River
close to its confluence with the Rhone in France
(Marston et al., 1995). This map was based on meti-
culous field surveys and remote sensing using aerial
photography. It shows the rich detail of a dynamically
evolving meandering riverine landscape, in which the
vegetation records the geomorphological history, and
that history defines the habitat diversity of the
landscape.
In a braided river, gravel and sand-bar and bedform
migration are the dominant sedimentary processes,
often resulting in channel avulsion because of the
water-level variations associated with bar migration
(Werritty & Ferguson, 1980). If there is a general
tendency for downcutting as channels change position
through avulsion (often into palaeochannels that have
not been infilled because of a lack of fines), then there
will be a pattern of abandoned bar surfaces of
differing age and elevation, on which vegetation
recruitment may be encouraged when thin spreads
of fines have occurred during flood events. The
potential for succession on these surfaces will, how-
ever, reflect the rate of migration and avulsion. In
braided rivers occupying mountain valleys, the loca-
tions of braided reaches may be intermittent, occur-
ring where the valley is wider than average or where
tributary or alluvial fan junctions locally enhance
sediment supply. These ‘sedimentation zones’ are
separated by narrow, single-thread, transport reaches
such as bedrock gorges (Church & Jones, 1982;
Church, 2002). Feedback between fluvial and ecolog-
ical processes in braided rivers manifest in vegetation
patterns on individual gravel-sand bar units and at
the reach scale, when sedimentation and plant suc-
cession processes influence one another. Sedimenta-
tion and bar formation, for example, may be
influenced by large woody debris accumulation
(Gurnell et al., 2000; Gurnell & Petts, 2002). Such
interactions can have an expression at a scale at which
it is difficult to unravel the roles of fluvial and
ecological processes. For example, over a 50-km reach
of the channelised River Isere, the area of vegetated
gravel bars between the dykes has expanded in the
last 50 years, beginning at three or four sites and
spreading upstream and downstream from these
(Fig. 5; Vautier, 2000). This pattern of development
could reflect the existence of gravel sedimentation
zones within the reach, where the bar forms encour-
age vegetation encroachment; or it could be that the
initial sites were locations of seed dispersal. A mixture
of the two processes is likely, reflecting the biophys-
ical nature of the river system.
Floodplain turnover rate and biodiversity
A further way in which the fluvial processes influence
biodiversity is in the general turnover rate of the
Fig. 5 The expansion of the percentage of bar surfaces along a
50 km reach of the Isere which were vegetated at different dates
between 1948 and 1996 (after Vautier, 2000; printed with
permission from the author).
566 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

floodplain alluvium, which depends on the rate at
which meanders sweep across the floodplain surface.
If this occurs either very slowly or very rapidly, low
biodiversity is likely, whereas intermediate rates of
turnover would maximise biodiversity (this illustrates
how the intermediate disturbance hypothesis is itself
a reflection of the channel dynamics). The rate of
turnover of the floodplain sediment store affects the
overall structure of the riparian vegetation. Rapid
turnover associated with channel migration causes a
higher proportion of the riparian area to be occupied
by vegetation in the pioneer stages, while a slow
turnover rate results in a higher percentage of mature
vegetation. One measure of turnover is the minimum
age of the oldest parts of the floodplain sediment
store, and Hughes (1997) quotes values of 100–
600 years for this, with the Amazon being an outlier
at 2000 years. In some cases, the floodplain has taken
much of the postglacial time period to evolve and turn
over, whereas the forest turnover time may be only of
the order of 500 years. In such cases, the evolution of
the fluvial landscape exercises little influence on the
ecosystem structure. However, when landscape and
ecosystem turnover rates converge, changes in the
landforms can have large impacts on the ecosystem
characteristics.
Local processes of sedimentation
and vegetation recruitment
Fluvial processes that control sedimentation, bar
growth and dissection, and bedform construction
associated with hydraulic roughness, all occur at
spatial scales below those of the reach-scale channel
dynamics (see Fig. 1). However, their spatial struc-
ture is determined by the larger-scale dynamics, and
within the sedimentary elements defined in Fig. 3,
smaller-scale patterns of sedimentation and sediment
size occur, which reflect the range of grain sizes in
transport, and flow patterns across the topography of
larger-scale forms. For example, Piegay et al. (2000)
demonstrate that sedimentation rates in abandoned
channels (perifluvial aquatic zones) differ systemat-
ically between braided and meandering reaches. Fine
sediment patches on gravel bar surfaces are import-
ant for the recruitment of individuals in the renewal
of plant succession, as seeds germinate and seedlings
survive in the patches of finer sediment that retain
moisture and provide an accessible nutrient store. As
Girel & Pautou (1997) demonstrate, these early
successional species may then trap more sediment,
and generate a microtopography which encourages
further spatial segregation of sediment sizes. In the
middle of a bar, canary grass (Phalaris arundinacea L.)
communities establish, encourage finer sediment
patches around them, and these sustain Calamagrostis
littorea (Schrad.) Beauv. and Festuca arundinacea
Schreb. In between, surface depressions accumulate
fines and attract species such as Ranunculus repens L.
and Deschampsia caespitosa (L.) Beauv. Gradually
these stabilise the surface for the invasion of woody
species (Salix, Populus). In addition, of course, there
are processes that favour vegetative reproduction.
Sedimentation buries woody debris and existing
seedlings, and encourages subsequent sprouting of
adventitious shoots. This suggests that the balance
between seed-based regeneration and vegetative
reproduction, and therefore the genetic diversity of
a riparian plant community, might be dependent on
the balance of erosive and depositional processes.
This hypothesis can be difficult to test, however. In a
comparative study of vegetation on bar surfaces in a
channelised and freely braiding reach of the River
Drome, M. Winfield and N. Barsoum (personal
communication) sampled leaf material from closely
spaced sites along eroding banks and at mid-bar
locations, avoiding sampling shoots which were
clearly from the same clone. The results of cluster
analysis of amplified fragment length polymorphism
(AFLP) bands show that the sampled individuals
were genetically very similar, but not sufficiently to
be regarded as clonal, with no differences between
the two river reaches. This finding suggests that, in
spite of the local and visible evidence of vegetative
regeneration, the bar-surface vegetation may be
dominated by seed-based recruitment.
Channel management and biodiversity
River regulation (by channelisation or flood control)
results in terrestrialisation of the vegetation, associ-
ated with a reduced rate of turnover of the fluvial
landscape, reduced rates of ecosystem change, reduc-
tions of channel and ecosystem dynamics and of
mosaic detail, reduced flood frequency, and loss
of habitat and age diversity. These consequences of
regulation on Alpine rivers such as the Rhone and the
Isere have been recorded by Girel & Pautou (1997).
Geomorphic dynamics of floodplains 567
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

The Isere has experienced a complex history of river
and flow management (Vautier, 2000), including
recent incision because of gravel extraction, and
reduction of flows because of reservoirs impounded
for hydropower generation. Today, there are stable
islands (both medial and lateral dyke-attached
islands), with a gravel substrate and silty sand at the
surface, anchored by vegetation growth. These veget-
ated islands are developing relatively uniform veget-
ation, whose flow resistance causes an increase of
flood water levels of about 1–1.5 m for a given
discharge. There is accordingly a need to destabilise
these islands to inhibit vegetation growth, reduce flow
resistance and maintain the integrity of the flood
protection afforded by the dykes. One means of
achieving this is to cut the vegetation, but this is an
expensive option because of the necessity for repetit-
ive cutting after rapid re-growth. An alternative
autogenic, self-maintaining approach is to enable the
re-introduction of sediment supply in order to restore
the dynamics of river scour and fill, with its potential
to create dynamic behaviour in the bar-forms (Dykaar
& Wigington, 2000). A similar set of ideas about
restoration of river bed dynamics has been considered
at the catchment scale by Bravard et al. (1999); renewal
of sediment supply restores channel dynamics, and
thus restores the dynamics of riparian vegetation. It
emerges that the biodiversity of islands is actually
enhanced by a management practice which encour-
ages sediment movement (J. Girel & F. Vautier,
personal communication); both at community and
species levels, islands on which vegetation is felled,
and surface sediment and roots are removed, appear
to be more diverse than islands where the vegetation
is simply cut. Although this is an artificial example, it
suggests a clear link to the conservation of ecological
diversity from the interdependence and maintenance
of fluvial and ecosystem dynamics.
Empirical investigation of channel
and vegetation dynamics
One means of assessing the relationship between
channel dynamics and biodiversity is an empirical
approach using remote sensing methods [cf. Vautier’s
(2000) work on the Isere shown in Fig. 5]. Sequential
multi-temporal remotely sensed imagery is a well-
established method for the analysis of channel
dynamics (Mertes, 2002). This may be based on
manual analysis of high-resolution aerial photogra-
phy, providing a synoptic perspective on channel
changes over wide areas (Lewin & Weir, 1977; Dykaar
& Wigington, 2000). However, developments in
spaceborne multi- and hyper-spectral imaging sys-
tems offer the potential to map fluvial environments
semi-automatically from surface reflectances (Muller,
Decamps & Dobson, 1993; Mertes, 2002). At first, this
was hindered by the mismatch between the scale of
fluvial landforms and the resolution of early satellite-
borne sensors (e.g. object space resolution of Landsat
MSS ¼ 80 m; Landsat TM ¼ 30 m). Subsequently, air-
borne imaging systems such as the Daedalus ATM
and the CASI, with high spatial and spectral resolu-
tions, have been used for geomorphological and
ecological studies on smaller rivers (Winterbottom &
Gilvear, 1997). However, by the late 1990s, spaceborne
sensors with appropriate spectral and spatial resolu-
tions have become available (Gilvear, Bryant & Hardy,
1999). These technical applications are set within a
theoretical debate about the potential of remote
sensing for vegetation mapping. Single classes may
not reflect the floristic composition of a multi-level
plant community, and as a result, mapping the habitat
may not be equivalent to mapping the vegetation.
However, in floodplain environments, the morphody-
namic and hydrological processes play a primary role
in structuring the vegetation, and there is a close
equivalence of habitat and plant community. Muller
(1997), thus, has shown how vegetation classifications
derived from aerial photography can be draped onto a
DTM, and then compared with flood extents mapped
through the use of inundation algorithms.
Additional technical developments, such as the
spatial database tools of Geographical Information
Systems (GIS), have greatly assisted the analysis of
channel dynamics using remotely sensed data. The
GIS facilitates the combination of information on
channel form from a variety of sources and scales,
with convenient intercomparison using simple over-
lay functions to analyse channel changes (Gurnell,
1997; Johnson, 2000). Moreover, GIS software allows
direct quantification of errors, and thus the means of
assessing the accuracy of detection of change. The
capacity to employ different sources of imagery
allows re-examination of archived aerial photographs
to gain insight into channel changes over time scales
relevant to plant and other ecological processes and
succession. For example, Rumsby, Brasington &
568 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

McVey (2001) have studied the plan-form dynamics of
the gravel-bedded River Feshie, U.K. [a wandering
gravel-bed river according to Miall’s (1985) classifica-
tion], analysing a multiscale (1 : 10 000–1 : 27 000)
combination of cartographic and aerial photographic
data, covering similar summer low flow periods of a
small 900–400 m reach between 1899 and 1997. Each
data source was rasterised and rectified to the
ordnance survey (OS) National Grid, and then resam-
pled to image files with a common object space pixel
resolution. After registration, geomorphological fea-
tures on the images were interpreted and a classifi-
cation scheme separating vegetated and unvegetated
zones of the currently active and the palaeochannel
network was manually digitised ‘on-screen’ with the
GIS software ArcView. Fig. 6 shows the geomorpho-
logical maps derived from the 1989 and 1997 sources.
These digital spatial databases can be analysed to
calculate changes in the areal proportion of channel
environments and habitats over time, using overlay
functions to determine the directions and types of
change between images. Validation tests on the
registration and digitising errors suggested that dif-
ferences greater than 4.5 m may be treated as signi-
ficant at the 95% confidence interval. A useful method
of representing the pattern and rate of typical channel
and floodplain changes is through the production of
tables which compare the surface cover properties of
areas digitised from one epoch to those at another;
Table 4 shows percentage changes between 1989 and
1997. Care is needed in interpreting these data, which
are sensitive to errors and uncertainty in the classifi-
cation. However, some changes are clearly evident.
For example, 36.4% of the active exposed gravel was
re-incorporated into the active wetted channel
between 1989 and 1997, while 2.4% of this same
surface type had been colonised and vegetated by
1997, and 19.3% of the active wetted channel in 1989
had become disconnected from the active channel and
converted to vegetated palaeochannel gravel.
While such analysis of changing morphology can
provide insights into the interplay between fluvial
and ecological dynamics, it is based on static views of
the channel at more or less arbitrary intervals. Any
direct link to particular floods is complicated by the
uneven distribution of such events over time, and the
non-linear relationship between flood magnitude and
channel change. Uncertainty about these relationships
can be reduced by reference to accurate river
discharge data and historical evidence, but such
Fig. 6 Maps of the geomorphology and vegetation of the River Feshie study reach based on 1989 and 1997 aerial photography
(after Rumsby et al., 2001).
Geomorphic dynamics of floodplains 569
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

empirical investigation of the direction and mechanism
of channel dynamics will always remain susceptible to
sampling errors. Thus, simulation modelling offers an
additional tool to interrogate the processes of channel
and vegetation dynamics in floodplain environments.
Modelling channel dynamics and biodiversity
Empirical approaches to the assessment of time
variation in the spatial structure of habitats and
communities in floodplain environments provide
valuable data on the interaction between channel
dynamics and biodiversity, but it is difficult to
extrapolate their results to other sites, and to use
them for management guidance in different locations.
An alternative whose potential has yet to be fully
explored, but which has been emphasised throughout
this paper, is the use of coupled models of channel
dynamics and plant community succession. The
output of a channel dynamics model is the spatial
mosaic of physical units (or habitats), and the ages
since the last destruction and renewal of the surface at
particular sites. A successional model could be linked
to this channel dynamics model, and would show
how the plant succession interacts with the channel
dynamics in generating different levels of species or
community diversity. In an uncoupled form, the
succession would simply follow the variation in
surface stability, but a coupled form could involve a
feedback in which the erodibility or rate of sedimen-
tation is a function of the vegetation properties at
different stages in the succession. Whichever mode is
employed, this simulation approach will permit
exploration of the ways in which key parameters of
both component sub-models affect the biodiversity of
the simulated reach; and will also assist in providing
quantitative tests of some qualitative hypotheses, such
as the intermediate disturbance hypothesis. It should
also allow simulations of the consequences of changes
arising through flow and river control, and therefore
provide a useful management tool.
Such a modelling approach requires a combination
of sub-models of (i) the channel dynamics and (ii) the
succession process. The nature of the first of these sub-
models will depend on the channel type (fluvial style),
and the two classes of channel dynamics model
discussed here are for single-thread meandering
rivers (Fig. 3b) and braided rivers (Fig. 3c). The
succession process is represented by an empirical
relationship based on observed successional path-
ways on floodplains in particular climatic regions.
These components are discussed in the sections
below.
The dynamics of single-thread meandering rivers
A simulation model of meandering river dynamics,
including a representation of the evolution of the
floodplain topography, is that developed by Howard
(1992). It has three main components. The first is a
model of flow, bed topography and sediment trans-
port in a meandering river. This model is based on the
Table 4 Percentage changes in the areas of various surface types in the reach of the River Feshie shown in Fig. 6, between 1989 and
1997. For example, 45.7% of the area occupied by the active wetted channel in 1989 was also active wetted channel in 1997, while
30.1% had changed to active exposed gravel
Surface type in 1989
Active zone Inactive zone
Active Active Palaeo- Palaeo-
Surface type wetted exposed Vegetated Palaeo- vegetated unvegetated
in 1997 channel gravel gravel channel Gravel Gravel Unclassified
Active wetted channel 45.7 36.4 7.6 0.7 1.0 0.1 3.0
Active exposed gravel 30.1 34.6 10.3 0.6 0.4 1.5 5.9
Vegetated gravel 0.3 2.4 0.0 0.0 0.0 0.0 0.0
Palaeo-channel 0.2 0.8 0.0 59.9 5.2 17.7 0.7
Palaeo-channel, 19.3 13.3 69.2 27.8 73.9 38.1 1.3
vegetated gravel
Palaeo-channel, 1.9 3.0 12.9 10.4 6.1 42.6 11.7
unvegetated gravel
Unclassified 2.6 9.5 0.0 0.6 13.4 0.0 77.4
570 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

work by Johannesson & Parker (1989), which although
one-dimensional, implicitly (if approximately) deals
with the effects of channel curvature on bed topog-
raphy (such as the point bar lateral slope), and on flow
and sediment transport. The second component is a
relationship of near-bank depth and velocity to the
rates of bank erosion and lateral migration. This
defines the bank erosion rate as a relationship
between the increasing local deviations of the near-
bed velocity, depth and shear stress from the average,
and the changing curvature around a bend. As the
river migrates and bends evolve as a result of spatially
differentiated bank erosion rates, circumstances arise
in which acute bends approach a neck cut-off position.
The model thus has a threshold distance at which
approaching channel segments are forced to cut off
the bend between them, and the bend is abandoned
and separated from the channel. The third component
is a model of floodplain sedimentation. This involves
two processes, those of point bar deposition and
overbank sediment diffusion. The deposition rate is
modelled as a function of relative floodplain height,
and distance from the channel, providing a crude
approximation of deposition close to the bank (form-
ing levees) and more distant overbank sedimentation.
When the channel sweeps through a model cell, the
elevation is set equal to the mean bed elevation, but as
the channel moves away, the elevation is adjusted to
represent the lateral depth perturbation on the point
bar side of a channel bend.
This model produces realistic simulations of sinu-
ous meander trains from initially straight channels
with random perturbations. After a few thousand
iterations, the simulated channel has developed
enough cut-off bends to create a path that wanders
laterally as well as displaying bends of different
amplitude, shape and ‘age’. Fig. 7 illustrates a short
section from a longer reach simulated by this model,
showing the properties the model generates that are
critical for the question of vegetation succession and
the age and elevation of the floodplain surface. With
relatively low deposition rates, the abandoned
channels do not fill-in with sediment, and remain
low-lying habitats conducive to the maintenance of
wetlands (these are marked in Fig. 7b as ‘sloughs’).
Point bars have an internal age variation consistent
with point bar growth, but after bend cut-off, adjacent
‘fossil’ point bars have different ages. The top left-
hand corner of Fig. 7a illustrates this clearly, because
a series of adjacent abandoned point bars have
maximum ages (in numbers of iterations) of 500,
600, 800, 1300 and 1500. The physical meaning of an
iteration has to be determined through comparison of
the simulation with a suitable real-world (prototype)
case of a dynamically evolving meandering river. The
elevation diagram (Fig. 7b) shows no levee develop-
ment, but Howard (1992) reviews the need to refine
the deposition model to allow the changing rates of
deposition with distance from the channel also to
reflect variation in grain size, and for an improved
Fig. 7 An illustration of the Howard (1992) simulation model
for a freely meandering river, showing (a) contours of floodplain
age in hundreds of iterations (areas older than 2100 iterations are
bordered by dashed lines and are uncontoured), and (b)
contours of floodplain elevation for a relatively low deposition
rate (low elevation sloughs shown by ‘S’) (from Howard, 1992;
printed with permission from John Wiley & Sons Ltd, New
York).
Geomorphic dynamics of floodplains 571
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

representation of discharge variation. However, the
model clearly has considerable potential for evaluating
the relationship between meandering channel dynam-
ics, habitat diversity and diversity of plant communi-
ties (as illustrated in Fig. 4), as it captures the main
features of the elevation and age structure of the
floodplain. Coupling its predictions of floodplain
properties with simple succession models such as
that shown in Fig. 9 (see below) would provide an
excellent basis for testing in a more quantitative
manner such concepts as the intermediate disturbance
hypothesis, simply by sensitivity analyses of the
ecological consequences of simulations with different
parameter values controlling the rates of bank erosion
and floodplain deposition.
The dynamics of braided rivers
A model which may be used to simulate the dynamics
of braided river systems for the purpose of investi-
gating the habitat diversity of their floodplains is that
of Murray & Paola (1997). This is a cellular automata,
or coupled map lattice model, based on the routing of
water and sediment across a regular grid topography.
It proceeds by routing an input of water at the top
edge of a reach consisting of nx by ny cells, with initial
topography such as a random elevation field added to
a linear plane whose slope represents the valley
gradient. A predefined discharge, Q, is routed from
each input cell (denoted by subscript i) to those cells
in the next row downstream that are lower in
elevation (denoted by the subscript j), in proportion
to the gradients to these cells (slope, S), and according
to the rule
Qi ¼ ½Sni =ð1=RSn
j Þ�Qn
where the normalisation by the sum of slopes ensures
that all water entering a cell leaves it within an
iteration of the model, and the exponent n is normally
0.5. The water transports sediment from cell to cell
according to a transport relationship in which the
transport rate is a function of discharge and gradient
(i.e. stream power). A variety of forms of transport
law may be employed, with or without a threshold of
erosion. A simple form is
Qsi ¼ K½QiSi þ eRQujSuj�m
where the second term adjusts the sediment flux into
each cell by the sum of the stream powers from the
three adjacent upstream cells (here identified by the
subscript uj), to represent the effect of flow momen-
tum from the upstream cells driving sediment against
adverse gradients. The parameter e is a fraction set by
Murray & Paola (1997) at 0.35. The exponent m is
normally taken to be about 2.5, in common with many
published sediment transport relationships for fluvial
environments (see, for example, Kirkby, 1971; Rich-
ards, 1982, p. 113).
During each iteration of the model, the cell eleva-
tions are adjusted according to a simple sediment
continuity relationship. This lowers the elevation by
an amount equivalent to net volumetric export of
sediment from the cell (that is, output less input)
divided by the cell area, or is aggraded by an
elevation change in which net import of sediment is
divided by the cell area and is also adjusted to reflect
the higher porosity of the deposited sediment. The
topography is rapidly self-organised by the transport
law until, depending on the initial topography and
the various parameter values selected, a braided
pattern is created (Fig. 8). As continuing erosion and
deposition takes place, and occasional avulsions
occur, the details of the geometry change, for given
sets of parameter values, an ‘average’ and reasonably
stable morphological structure may be maintained
over a relatively large number of model iterations. In
the example in Fig. 8, the model included addition of
a sediment flux into the upper end of the simulated
reach, and this was varied sinusoidally with a period
of 10 000 iterations. During the high sediment feed
stages of the cycle, aggradation occurred at the upper
end of the simulated reach. The evolution of the
channel is illustrated by the map of discharge. It is
clear that an increased area of bar surface which is
inundated by shallow flow develops in the upper part
of the reach as the accumulating sediment progrades
downstream. In addition, although the periods of low
sediment feed result in incision of the channel, the
channel does not become locked into a stable position,
and even in the downstream reaches there are major
shifts of the channel. With this model, it is a relatively
straightforward procedure to count the number of
iterations for which each cell has been stable, and to
generate a map at selected time steps of the ‘ages’ (in
numbers of iterations) of every cell. This means that it
is possible to generate a history of the intermittent
renewal of vegetation succession within the simulated
reach.
572 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

To use this model in ecological simulation, further
developments are required. For example, proper
scaling of the model to real time (rather than iteration
counts) is necessary so that succession processes
lasting several decades can be represented. Murray
& Paola (1997) provide some insights into the scaling,
which requires selection of a prototype real stream,
the assumption that the model scales like the proto-
type in some respects (for example, that length scales
are similar), and then a calculation of other scale ratios
(such as the time–scale ratio of model to protoype)
using the routing equations built into the model,
which involve some non-linearities. One simple first
approximation is, however provided by considering
the iteration period. In a single model iteration, the
discharge that enters at the top of the reach is routed
to the bottom. If a length scale is assumed that
matches the model to a real prototype, then an
approximate estimate can be made of the velocity,
and hence of the time period implied by an iteration.
In Fig. 8, a cell size of 10 m implies an iteration
equivalent to about 12 h, and the evolution from the
pattern in the left-hand diagram to that in the right
represents about 500 years. During this period, signi-
ficant channel shifts and bar developments has
occurred in the model, with obvious implications for
the distribution of habitats, the progress of succession
and the biodiversity.
A successional sub-model to coupleto the fluvial dynamics sub-model
Whether the simulated channel is meandering or
braided, the objectives of the simulation will only be
satisfied if a succession sub-model can be linked to
the sub-model of channel dynamics. There are no
deductive or theoretical grounds on which a general,
quantitative succession model can be based, so it will
inevitably rely on empirical data. Furthermore, the
particular sequence of species and communities in a
riparian plant succession will be specific to the
regional climatic and biophysical conditions for the
simulated case, and the model of plant succession
will therefore be case-dependent. An empirically
generated set of rules defining the successional
process will, of course, run the risk of being affected
by factors other than those related to channel
dynamics (which include surface age, elevation,
sediment type and moisture availability). For exam-
Fig. 8 A run of the Murray & Paola (1997) braided river model
in a reach of 22 · 400 cells with a sinusoidally varying sediment
feed having a period of 10 000 iterations. Darker shades in the
topography map are lower elevations, light shades are higher
elevations. Darker shades in the discharge map are higher cell
discharges. The flow (and channel) pattern has changed mark-
edly between 135 000 and 475 000 iterations (from Murray &
Paola, 1997; printed with permission from John Wiley & Sons
Ltd, New York).
Geomorphic dynamics of floodplains 573
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

ple, if the set of field sites investigated in the
construction of the succession model straddles cli-
matic zones, the empirical evidence of transitions
from one community to another may be distorted by
this effect, as will be the simulated diversity arising
from linking this model to that of the channel
dynamics. Even more insidiously, if the floodplains
sampled in developing an empirical succession
model are associated with a river whose dynamics
are non-equilibrium in their style and rate – for
example, because of historic flow and channel man-
agement practices – this too will distort the succes-
sion simulated in response to the channel dynamics
defined by one the models discussed above. Thus, a
premium is placed on the manner in which fieldwork
is conducted to define a succession model.
Cordes, Hughes & Getty (1997) provide an example
of the kind of field investigation required to construct
a successional model. This involved mapping at
representative sites, using field survey and aerial
photography to identify in an iterative manner the
range of riparian plant communities. Communities
were identified in relation to both their topographic
position, in terms of landform unit and elevation
above low flow level, and their age as measured using
dendrochronology. The communities were then
organised into a successional model using space-time
substitution based on examining transects from the
active channel margins to the distal parts of the
floodplain. Two models were constructed. The first
was for the upper Red Deer River in Alberta, Canada,
a gravel-bed river with narrow, locally incised chan-
nels with limited floodplain development. Early
successional stages here involve sparsely vegetated
gravel bars, which are then colonised by willows
which trap sediment and are followed by poplars or
larger, more mature willows, and eventually spruce.
Later succession depends on the density of poplar
establishment in the earlier stages, and this is partly
dependent on the elevation, landform and substratum
type. The model for the lower Red Deer River reflects
the different biophysical and climatic zone of its
lower reaches, and the sandier bed material and
distinctive channel morphology in this meandering
channel, with many alluvial islands and extensive
point bar areas with large stands of riparian wood-
land. The lower Red Deer River has many of the
characteristics of Miall’s (1985) Model 6 channel style
(see Fig. 3b).
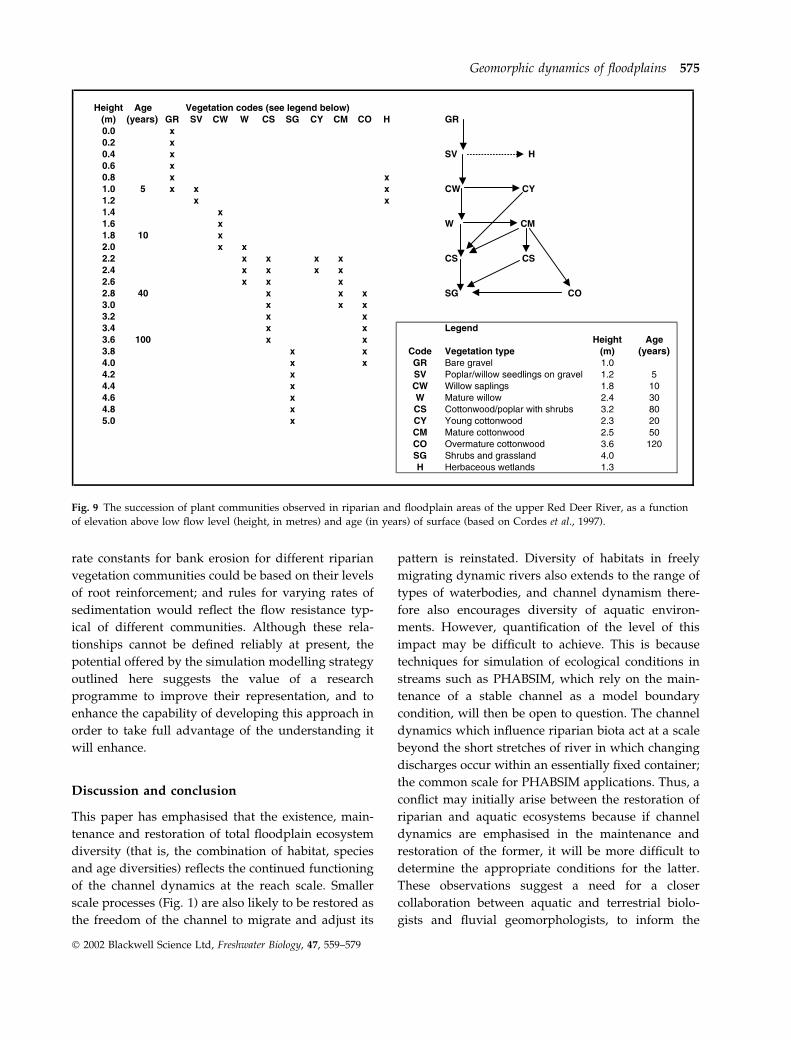
Fig. 9 illustrates, in two different but simplified
forms, a succession model for this reach. The succes-
sion here is towards a cottonwood-dominated com-
munity similar to that described by Dykaar &
Wigington (2000) for the Willamette River in Oregon,
U.S.A. One successional path follows a relatively
direct route from seedlings on sandy, gravelly bars
through various forms of cottonwood or cottonwood-
poplar mixtures, to a shrub-grass community. Some
locations, however, experience various deviations
from this path, reflecting the topography and soils of
particular sites. For example, the ‘young cottonwood’
community (CY) is confined to narrow linear bands on
sandy levees, while the CO community (see Fig. 9) is
mainly found on scroll-bar ridges. There is also a
‘herbaceous wetland’ community (H) which is initi-
ated at low elevations, and essentially occupies the
habitat of abandoned channels after bend cutoffs, and
is a distinctive community reflecting another kind of
topographic element created by the channel dynamics.
The implications of this complexity of successional
trajectories are, of course, that a more sophisticated set
of simulation rules is required than simply a model in
which the correlated variables of age and height
determine the vegetation community. There is an
interaction between age, height, and the habitat
‘patch’ characteristics, partly defined by patch shape
and spatial arrangement, and partly by the patch
sedimentology, soil type and moisture status. There is
also likely to be a stochastic element, in which there
are differential probabilities of taking one or the other
route at a particular junction (as in Fig. 9), depending
on the type of habitat patch. None of these would
constitute insurmountably difficult additional ele-
ments of a successional sub-model. Indeed, the rich-
ness of behaviour that they would represent would
give interesting possibilities for an exploration of the
sensitivity of habitat, community and age diversity to
different values of the critical age, height and patch
properties at which vegetation communities change,
and the different probabilities that control the timing
of these changes. A further area for added sophisti-
cation lies in the addition of routines to allow full
coupling of the successional sub-model to the channel
dynamic sub-model. These would be required to
effect a feedback between the evolving vegetation
community and the parameters in the meandering or
braiding models that control rates of bank erosion and
overbank sedimentation. Rules that define different
574 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579
rate constants for bank erosion for different riparian
vegetation communities could be based on their levels
of root reinforcement; and rules for varying rates of
sedimentation would reflect the flow resistance typ-
ical of different communities. Although these rela-
tionships cannot be defined reliably at present, the
potential offered by the simulation modelling strategy
outlined here suggests the value of a research
programme to improve their representation, and to
enhance the capability of developing this approach in
order to take full advantage of the understanding it
will enhance.
Discussion and conclusion
This paper has emphasised that the existence, main-
tenance and restoration of total floodplain ecosystem
diversity (that is, the combination of habitat, species
and age diversities) reflects the continued functioning
of the channel dynamics at the reach scale. Smaller
scale processes (Fig. 1) are also likely to be restored as
the freedom of the channel to migrate and adjust its
pattern is reinstated. Diversity of habitats in freely
migrating dynamic rivers also extends to the range of
types of waterbodies, and channel dynamism there-
fore also encourages diversity of aquatic environ-
ments. However, quantification of the level of this
impact may be difficult to achieve. This is because
techniques for simulation of ecological conditions in
streams such as PHABSIM, which rely on the main-
tenance of a stable channel as a model boundary
condition, will then be open to question. The channel
dynamics which influence riparian biota act at a scale
beyond the short stretches of river in which changing
discharges occur within an essentially fixed container;
the common scale for PHABSIM applications. Thus, a
conflict may initially arise between the restoration of
riparian and aquatic ecosystems because if channel
dynamics are emphasised in the maintenance and
restoration of the former, it will be more difficult to
determine the appropriate conditions for the latter.
These observations suggest a need for a closer
collaboration between aquatic and terrestrial biolo-
gists and fluvial geomorphologists, to inform the
Fig. 9 The succession of plant communities observed in riparian and floodplain areas of the upper Red Deer River, as a function
of elevation above low flow level (height, in metres) and age (in years) of surface (based on Cordes et al., 1997).
Geomorphic dynamics of floodplains 575
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

choice of restoration aims, policy and practice, and to
ensure that the research and data needs are met for
restoration of the appropriate dynamics at the appro-
priate scale, in cases where this becomes economically
and politically viable (Dykaar & Wigington, 2000).
These research needs focus on dynamic channels and
ecosystems at a coarser spatial scale than the present
emphasis at the grain, bedform and microhabitat
scale, and this presents a challenge for fluvial geo-
morphologists, ecologists and river managers, who
will face new questions and a lack of reliable answers
until the gaps in current knowledge have been filled.
Nevertheless, a particular advantage of a modelling
strategy is that it draws attention to such existing
deficiencies, and the focus of this paper on simulation
modelling of the interaction between channel and
ecosystem dynamics has encouraged identification of
some key research needs. These require interdisci-
plinary research at the interface between hydrology,
geomorphology and ecology. For example, successful
manipulation of the flood pulse for ecological restor-
ation requires more information on the optimal timing
of relationships between seed production and flood-
ing; critical soil water potentials for the germination of
seeds; critical rates of soil water decline to optimise
seedling performance and more information on the
response of community dynamics to hydrological
processes to supplement that on the behaviour of
individual species. Restoration of geomorphological
dynamics will require more reliable data on, descrip-
tions of, and parameters to represent: the relationship
between rates of channel change, floodplain turnover
and habitat diversity for different channel patterns;
the balance of seed-based regeneration and vegetative
reproduction in relation to erosion and sedimentation
and their effects on genetic diversity; the effects of
different riparian vegetation on bank erosion rates;
and the rates and patterns of sedimentation within
riparian vegetation during overbank flow. Investiga-
tions into these questions will inform the develop-
ment and calibration of simulation models of the kind
discussed in this paper, so that these models may be
used to address a variety of practical questions.
Topics of practical relevance are likely to include
establishing the magnitude and frequency of the
controlled flood pulse required to maintain an appro-
priate floodplain turnover rate and pattern-related
habitat diversity in order to sustain ecosystem
dynamics. As most restoration initiatives will be
relatively local in scale, eco-geomorphological mod-
elling will help to determine which rivers, and which
reaches within them, are most suited to conservation
or restoration of natural channel dynamics. It will also
help to answer questions about the desirability of
permitting channel dynamics in a succession of
reaches in order to encourage larger-scale river
corridor ecological processes, for example at meta-
population levels. Of course, there will also be many
issues to address which are more dependent on policy
and politics. Restoring channel dynamics may require
that flood embankments are retired further from the
river or abandoned, but this could jeopardise the
integrity of adjacent land uses, and create conflict
between the restoration objective and economic activ-
ities, requiring cost-benefit analyses to resolve the
conflict. The feasibility of restoration of the dynamic
behaviour of river channels in their floodplains may
also depend on legal challenges to land ownership, as
bank erosion, channel migration and avulsion occur.
However, the practice of river and ecosystem man-
agement can only be assisted by the development of
simulation models that provide the capacity to exam-
ine a range of scenarios, to establish the available
choices, and to underpin review of the costs and
benefits of conservation and restoration of the eco-
system diversity of dynamic floodplain environments.
Acknowledgments
This paper is based on research conducted for two
EC-funded research projects: Floodplain Biodiversity
and Restoration (FLOBAR): Hydrological and geo-
morphological mechanisms influencing floodplain
diversity and their application to the restoration of
European floodplains (1996–99; EC Contract No.
ENV4-CT96-0317); and ‘Floodplain Biodiversity and
Restoration (FLOBAR 2): Integrated natural science
and socio-economic approaches to catchment flow
management’ (2000–2003; EC Contract No. ENK1-CT-
1999-00031 FLOBAR2). We are endebted to colleagues
and partners in these projects, in Cambridge, Gre-
noble, Clermont-Ferrand, Toulouse, and Umea; Bill
Adams, Adrian Hayes, Nadia Barsoum, Mark Win-
field, Jean-Luc Peiry, Guy Pautou, Jacky Girel, Phil-
lippe Belleudy, Remi Foussadier, Franck Vautier,
Gerard Marigo, Etienne Muller, Henri Decamps, Luc
Lambs, Helene Guilloy, Christer Nilsson and Mats
Johansson.
576 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

References
Amoros C., Bravard J.P., Castella C. et al. (1986)
Recherches Interdisciplinaires sur les ecosystemes de la
Basse-Plaine de l’Ain (France): potentialites evolutives et
gestion. Documents de Cartographie Ecologique 29.
Universite Scientifique, Technologique et Medicale de
Grenoble, CNRS, Grenoble, 166 pp.
Bravard J.P., Landon N., Peiry J.-L. & Piegay H. (1999)
Principles of engineering geomorphology for man-
aging channel erosion and bedload transport, exam-
ples from French rivers. Geomorphology, 31, 291–311.
Brierley G.J. & Fryirs K. (2000) River styles, a geomorphic
approach to catchment characterization: implications
for river rehabilitation in Bega Catchment, New South
Wales, Australia. Environmental Management, 25, 661–
679.
Brown A.G., Stone P. & Harwood K. (1995) The Biogeo-
morphology of a Wooded Anastomosing River: The Gearagh
on the River Lee in County Cork, Ireland. Occasional Paper
32. Department of Geography, University of Leicester,
Leicester. 76.
Church M.A. (2002) Geomorphic thresholds in riverine
landscapes. Freshwater Biology, 47, 541–557.
Church M.A. & Jones D. (1982) Channel bars in gravel-
bed rivers. In: Gravel-Bed Rivers (Eds R.D. Hey, J.C.
Bathurst & C.R. Thorne), pp. 291–338. John Wiley &
Sons, Chichester.
Connell J.H. (1978) Diversity in tropical rain forests and
coral reefs. Science, 199, 1302–1310.
Cordes L.D., Hughes F.M.R. & Getty M. (1997) Factors
affecting the regeneration and distribution of riparian
woodlands along a northern prairie river: the Red Deer
River, Alberta, Canada. Journal of Biogeography, 24, 675–
695.
Dykaar B.B. & Wigington P.J. Jr (2000) Floodplain
formation and cottonwood colonization patterns on
the Willamette River, Oregon, USA. Environmental
Management, 25, 87–104.
Gilvear D.J., Bryant R. & Hardy T. (1999) Remote
sensing of channel morphology and in-stream fluvial
processes. Progress in Environmental Science, 1, 257–
284.
Girel J. & Pautou G. (1997) The influence of sediment on
vegetation structure. In: Buffer Zones: Their Processes and
Potential in Water Protection (Eds N.E. Haycock, T.P.
Burt, K.H.T. Goulding & G. Pinay), pp. 93–112. Quest
Environmental, Harpenden.
Gurnell A.M. (1997) Channel change on the River
Dee meanders, 1946–1992 from the analysis of air
photographs. Regulated Rivers: Research and Manage-
ment, 12, 13–26.
Gurnell A.M. & Petts G.E. (2002) Island-dominated
landscapes of large floodplain rivers, a European
perspective. Freshwater Biology, 47, 581–600.
Gurnell A.M., Petts G.E., Harris N.M., Ward J.V., Tockner
K., Edwards P.J. & Kollman J. (2000) Large wood
retention in river channels: the case of the Fiume
Tagliamento, Italy. Earth Surface Processes and Land-
forms, 25, 255–275.
Hickin E.J. (1974) The development of meanders in
natural river channels. American Journal of Science, 274,
414–442.
Hill M. & Platts W.S. (1991) Ecological and geomorpho-
logical concepts for instream and out-of-channel flow
requirements. Rivers, 2, 319–343.
Howard A.D. (1992) Modelling channel migration and
floodplain sedimentation in meandering streams. In:
Lowland Rivers: Geomorphological Perspectives (Eds P.A.
Carling & G.E. Petts), pp. 1–41. John Wiley & Sons Ltd,
Chichester.
Hughes F.M.R. (1997) Floodplain biogeomorphology.
Progress in Physical Geography, 21, 501–529.
Hughes F.M.R., Barsoum N., Richards K.S., Winfield M.
& Hayes A. (2000) The response of male and female
black poplar (Populus nigra L. subsp. betulifolia (Pursh)
W. Wettst.) cuttings to different water table depths and
sediment types: implications for flow management and
river corridor biodiversity. Hydrological Processes, 14,
3075–3098.
Johannesson J. & Parker G. (1989) Linear theory of river
meanders. In: River Meandering (Eds S. Ikeda &
G. Parker), pp. 181–204. Water Resources Monograph
12, American Geophysical Union, Washington, DC
Johnson W.C. (2000) Tree recruitment and survival in
rivers: influence of hydrological processes. Hydrological
Processes, 14, 3051–3074.
Junk W.J., Bayley P.B. & Sparks R.E. (1989) The flood
pulse concept in river-floodplain systems. Canadian
Special Publication of Fisheries and Aquatic Sciences, 106,
110–127.
Karrenberg S., Edwards P.J. & Kollmann J. (2002) The life
history of Salicacaeae living in the active zone of flood-
plains. Freshwater Biology, 47, 733–748.
King J. & Louwe D. (1998) Instream flow requirements
for regulated rivers in South Africa using the building
block methodology. Aquatic Ecosystem Health and Man-
agement, 1, 109–124.
Kirkby M.J. (1971) Hillslope process-response models
based on the continuity equation. In: Slopes: Form and
Process (Ed. D. Brunsden), pp. 15–30. Institute of British
Geographers, London.
Lawler D.M. (1992) Process dominance in bank erosion
systems. In: Lowland Rivers: Geomorphological Perspec-
Geomorphic dynamics of floodplains 577
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

tives (Eds P.A. Carling & G.E. Petts), pp. 117–143. John
Wiley & Sons Ltd, Chichester.
Lewin J. & Weir M.J.C. (1977) Morphology and recent
history on the lower Spey. Scottish Geographical Maga-
zine, 93, 45–51.
Mahoney J.M. & Rood S.B. (1998) Streamflow require-
ments for cottonwood seedling recruitment – an
integrative model. Wetlands, 18, 634–645.
Malanson C.P. (1993) Riparian Landscapes. Cambridge
University Press, Cambridge.
Marston R.A., Girel J., Pautou G., Piegay H., Bravard J.P.
& Arneson C. (1995) Channel metamorphosis, flood-
plain disturbance, and vegetation development – Ain
River. Geomorphology, 13, 121–131.
Mertes L.A.K. (2002) Remote sensing of riverine land-
scapes. Freshwater Biology, 47, 799–816.
Miall A.D. (1985) Architectural element analysis: a new
method of facies analysis applied to fluvial deposits.
Earth Science Reviews, 22, 261–308.
Miall A.D. (1996) The Geology of Fluvial Deposits: Sedimen-
tary Facies, Basin Analysis, and Petroleum Geology.
Springer, Berlin.
Middleton B. (1999) Flood Pulsing and Disturbance
Dynamics. John Wiley & Sons Ltd, New York.
Milhous R.T., Updike M.A. & Schneider D.M. (1989)
Physical Habitat Simulation system reference manual –
Version II, Instream Flow Information Paper No. 26,
Vol. 89, U.S. Fish and Wildlife Service, Biological
Report, Washington, DC.
Milton E.J., Gilvear D.J. & Hooper I.D. (1995) Investi-
gating river channel changes using remotely sensed
data. In: Changing River Channels (Eds A.M. Gurnell &
G.E. Petts), pp. 277–301. John Wiley & Sons, Chiches-
ter.
Moon B.P., van Niekerk A.W., Heritage G.L., Roger K.H.
& James C.S. (1997) A geomorphological approach to
the ecological management of rivers in the Kruger
National Park: the case of the Sabie River. Transactions,
Institute of British Geographers, New Series, 22, 31–48.
Muller E. (1997) Mapping riparian vegetation along
rivers: old concepts and new methods. Aquatic Botany,
58, 411–437.
Muller E., Decamps H. & Dobson M.K. (1993) Contribu-
tion of space remote sensing to river studies. Freshwater
Biology, 29, 301–312.
Murray A.B. & Paola C. (1997) Properties of a cellular
braided-stream model. Earth Surface Processes and
Landforms, 22, 1001–1025.
Naiman R.T., Decamps H. & Pollock M. (1993) The role of
riparian corridors in maintaining regional biodiversity.
Ecological Applications, 3, 209–212.
Nanson G.C. & Beach H.F. (1977) Forest succession and
sedimentation on a meandering river floodplain,
Northeast British Columbia, Canada. Journal of Bio-
geography, 4, 229–251.
Nanson G.C. & Croke J.C. (1992) A genetic classification
of floodplains. Geomorphology, 4, 459–486.
Nilsson C., Grelsson G., Johansson M. & Sperens U.
(1991) Small rivers behave like large rivers: effects of
postglacial history on plant species richness along river
banks. Journal of Biogeography, 18, 533–541.
Peterken G.F. & Hughes F.M.R. (1995) Restoration of
floodplain forests in Britain. Forestry, 68, 187–202.
Petts G.E. (1996) Water allocation to protect river
ecosystems. Regulated Rivers: Research and Management,
12, 353–365.
Petts G.E., Gurnell A.M., Gerrard A.J. et al. (2000)
Longitudinal variations in exposed riverine sediments;
a context for the ecology of the Fiume Tagliamento,
Italy. Aquatic Conservation: Marine and Freshwater Eco-
systems, 10, 249–266.
Pickett S.T.A. & White P.S. (Eds) (1985) The Ecology of
Natural Disturbance and Patch Dynamics. Academic
Press, San Diego.
Piegay H., Bornette G., Citterio A., Herouin E., Moilin B.
& Statiotis C. (2000) Channel instability as a control on
silting dynamics and vegetation patterns within peri-
fluvial zones. Hydrological Processes, 14, 3011–3029.
Poole G.C. (2002) Fluvial landscape ecology: addressing
uniqueness within the river discontinuum. Freshwtaer
Biology, 47, 641–660.
Richards K.S. (1982) Rivers: Form and Process in Alluvial
Channels. Methuen, London. 361 pp.
Rumsby B.T., Brasington J. & McVey R. (2001) The
potential for high resolution fluvial archives in braided
rivers: quantifying historic reach-scale channel and
floodplain development in the River Feshie, Scotland.
In: River Basin Sediment Systems: Archives of Environ-
mental Change (Eds D. Maddy, M.G. Macklin &
J. Woodward). Balkema, Rotterdam.
Salo J., Kalliola R., Hakkinen I., Makinen Y., Niemala P.,
Puhakka M. & Coley P.D. (1986) River dynamics and
the diversity of Amazon lowland forests. Nature, 322,
254–258.
Shankman D. (1993) Channel migration and vegetation
patterns in the southeastern coastal plain. Conservation
Biology, 7, 176–183.
Tockner K., Malard F. & Ward J.V. (2000) An extension of
the Flood Pulse Concept. Hydrological Processes, 14,
2861–2883.
Vannote R.L., Minshall G.W., Cummins K.W., Sedell J.R.
& Cushing C.E. (1980) The river continuum concept.
Canadian Journal of Fisheries and Aquatic Sciences, 37,
130–137.
Vautier F. (2000) Dynamique geomorphologique et vegetali-
sation des cours d’eau endigues: l’exemple de l’Isere dans le
578 K. Richards et al.
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579

Gresivaudan. PhD Thesis, l’Universite Joseph Fourier,
Grenoble, France.
Ward J.V. (1998) Riverine landscapes: biodiversity pat-
terns, diversity regimes and aquatic conservation.
Biological Conservation, 83, 269–278.
Ward J.V. & Tockner K. (2001) Biodiversity: towards a
unifying theme for river ecology. Freshwater Biology, 46,
807–819.
Ward J.V., Tockner K., Arscott B.D. & Claret C. (2002)
Riverine landscape diversity. Freshwater Biology, 47,
517–539.
Ward J.V., Tockner K. & Schiemer F. (1999) Biodiversity
of floodplain ecosystems: ecotones and connectivity.
Regulated Rivers: Research and Management, 15, 125–139.
Werritty A. & Ferguson R.I. (1980) Pattern changes in a
Scottish braided river over 1, 30 and 200 years. In:
Timescales in Geomorphology (Eds R.A. Cullingford,
D.A. Davidson & J. Lewin), pp. 53–68. John Wiley &
Sons, Chichester.
Whittaker R.H. (1960) Vegetation of the Siskyou Moun-
tains, Oregon and California. Ecological Monographs, 30,
279–338.
Winterbottom S. & Gilvear D.J. (1997) Quantification of
channel bed morphology in gravel-bed rivers using
airborne multispectral imagery and aerial photography.
Regulated Rivers: Research and Management, 13, 489–499.
(Manuscript accepted 11 November 2001)
Geomorphic dynamics of floodplains 579
Ó 2002 Blackwell Science Ltd, Freshwater Biology, 47, 559–579




















