
Spatial and temporal variations of microinvertebrates across temporary floodplains of the lower...
13
This article was downloaded by: [University of Botswana Library] On: 15 October 2012, At: 01:18 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK African Journal of Aquatic Science Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/taas20 Spatial and temporal variations in densities of small fishes across different temporary floodplain types of the lower Okavango Delta, Botswana N Siziba a , M J Chimbari a , K Mosepele a & H Masundire b a Okavango Research Institute, University of Botswana, Private Bag 285, Maun, Botswana b Department of Biological Sciences, University of Botswana, Private Bag 0022, Gaborone, Botswana Version of record first published: 20 Dec 2011. To cite this article: N Siziba, M J Chimbari, K Mosepele & H Masundire (2011): Spatial and temporal variations in densities of small fishes across different temporary floodplain types of the lower Okavango Delta, Botswana, African Journal of Aquatic Science, 36:3, 309-320 To link to this article: http://dx.doi.org/10.2989/16085914.2011.636912 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of Spatial and temporal variations of microinvertebrates across temporary floodplains of the lower...

This article was downloaded by: [University of Botswana Library]On: 15 October 2012, At: 01:18Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
African Journal of Aquatic SciencePublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/taas20
Spatial and temporal variations in densities of smallfishes across different temporary floodplain types ofthe lower Okavango Delta, BotswanaN Siziba a , M J Chimbari a , K Mosepele a & H Masundire ba Okavango Research Institute, University of Botswana, Private Bag 285, Maun, Botswanab Department of Biological Sciences, University of Botswana, Private Bag 0022,Gaborone, Botswana
Version of record first published: 20 Dec 2011.
To cite this article: N Siziba, M J Chimbari, K Mosepele & H Masundire (2011): Spatial and temporal variations in densitiesof small fishes across different temporary floodplain types of the lower Okavango Delta, Botswana, African Journal ofAquatic Science, 36:3, 309-320
To link to this article: http://dx.doi.org/10.2989/16085914.2011.636912
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss, actions,claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

African Journal of Aquatic Science 2011, 36(3): 309–320Printed in South Africa — All rights reserved
African Journal of Aquatic Science is co-published by NISC (Pty) Ltd and Taylor & Francis
Copyright © NISC (Pty) LtdAFRICAN JOURNAL OF
AQUATIC SCIENCEISSN 1608–5914 EISSN 1727–9364doi: 10.2989/16085914.2011.636912
The Okavango Delta is one of the largest inland tropical wetlands in Africa. Research on the fishes of Okavango Delta (hereafter referred to as the Delta) has mainly focused on adults of large-sized fish species (Merron et al. 1990, Merron 1991, 1993a, 1993b, Mosepele 2000, Mosepele and Kolding 2003, Mosepele and Mosepele 2005, Mosepele et al. 2005) and little work has been done on small-sized fish species and juveniles of larger fishes. There is evidence that crucial processes of fish stock regulation occur at the juvenile stages prior to their recruitment into the adult populations (Hjort 1926, Cushing and Harris 1973, Shepherd and Cushing 1980, Cushing 1990, Shepherd et al. 1990). Furthermore, small fishes are preyed upon by carnivorous fish species up the food chain. It has been suggested that temporary floodplains in the Delta provide important nursery habitats for juvenile fishes (Merron 1991, Høberg et al. 2002, Ramberg et al. 2006, Lindholm et al. 2007) and these systems also support large populations of small-sized fish species.
The magnitude of the annual floods that the Delta receives from the Okavango River influences the extent of flooding of the temporary floodplains. Depending on the history of flooding frequencies, the temporary floodplains of the Delta have been categorised into three main classes: (1) primary, (2) secondary and (3) more rarely flooded floodplains (Wolski and Murray-Hudson 2006, Murray-
Hudson et al. 2011). The least-elevated primary floodplains, located close to the distributory channels, flood almost every year, whereas secondary floodplains get inundated by water spilling from the primary floodplains. The more rarely flooded floodplains receive water only during high flood years. Normally, temporary floodplains desiccate during non-flooding months and support terrestrial processes such as growth of terrestrial flora and foraging by wildlife (Bonyongo 2004, Ramberg et al. 2006). These terrestrial processes, together with dead macrophytes of the previous flooding season, contribute to the accumulation of organic matter and nutrients that are important contributors to aquatic productivity during the next flooding season (Junk et al. 1989). High aquatic productivity in temporary floodplains is critical in these shallow systems for supporting improved production of small fishes.
Globally, there is increasing isolation of rivers from their important peripheral temporary floodplains with potential negative impacts on fish biodiversity in river-floodplain systems (Gehrke et al. 1995, Jurajda 1995, Schmitten 1999, Humphries and Lake 2000, Aarts et al. 2004). The magnitude of the annual flooding cycle of the Delta is threatened by increasing human water demands and climate change (Mbaiwa 2004, Murray-Hudson et al. 2006). These threats are likely to reduce the extent of flooding of the system, which might lead to loss of some temporary
Spatial and temporal variations in densities of small fishes across different temporary floodplain types of the lower Okavango Delta, Botswana
N Siziba1, MJ Chimbari1*, K Mosepele1 and H Masundire2
1 Okavango Research Institute, University of Botswana, Private Bag 285, Maun, Botswana2 Department of Biological Sciences, University of Botswana, Private Bag 0022, Gaborone, Botswana* Corresponding author, e-mail: [email protected]
Received 19 December 2010, accepted 25 July 2011
Small-sized and juvenile fishes as well as physicochemical water parameters in various primary, secondary and rarely flooded temporary floodplains of the Okavango Delta were assessed during the different hydrological phases of the 2009–2010 flooding season. Small fishes were sampled in the marginal zone of the floodplains using a throw-trap net and a suite of physicochemical properties were measured. Both physicochemical water quality parameters and densities of small fishes showed spatiotemporal variations across the temporary floodplain types. Turbidity, dissolved oxygen, conductivity, woody debris and chlorophyll a were all highest in rarely flooded floodplains compared to both primary and secondary floodplains (ANOVA, p < 0.05). Fish from 38 fish taxa belonging to 11 families were identified and classified during the study. The fish families Cichlidae (dominated by tilapias), Poeciliidae (Aplocheilichthys spp.) and Cyprinidae (Barbus spp.), were the most abundant across the study floodplains. The poeciliids were most abundant in frequently flooded primary and secondary floodplains, whereas juvenile cichlids dominated in rarely flooded floodplains. During high floods the rarely flooded portions of the delta function as important nursery habitats for juvenile cichlids, implying that a significant reduction in flooding may have negative effects on the delta’s tilapia stocks due to reduced recruitment success.
Keywords: Cichlidae, Poeciliidae, rarely flooded floodplains, wetlands
Introduction
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

Siziba, Chimbari, Mosepele and Masundire310
floodplains. The significance of both secondary and rarely flooded floodplains to small fishes in the Delta is poorly understood, since the only available preliminary surveys have been restricted to primary floodplains (Høberg et al. 2002, Lindholm et al. 2007).
Temporary floodplains are highly productive and often support large populations of small-sized fish species and juvenile fish, both here referred to as small fishes. Production of small fishes in these ephemeral systems has a regulatory effect on fish stocks. This study assessed the spatial and temporal variations in species richness, diversity and abundance of small fishes across the different temporary floodplain types of the lower Okavango Delta. Two hypotheses were tested: (1) species richness and diversity of small fishes is highest in the primary floodplains and lowest in rarely flooded floodplains, and (2) rarely flooded floodplains support higher densities of small fishes than both primary and secondary floodplains.
Materials and methods
Study areaThe study floodplains were randomly selected from the primary, secondary and rarely flooded floodplains in the lower Okavango Delta. The flooding frequency map of Wolski and Murray-Hudson (2006) showing different classes of temporary floodplains (primary, secondary and rarely flooded floodplains) was used in the selection of study floodplains. Since the study area lies largely in a wildlife area with dangerous animals, the accessibility of floodplains by either car or boat was a consideration during the selection of the study sites. Four primary floodplains, two secondary floodplains (Water lily and Wildebeest) and two rarely flooded floodplains (Daunara and Buffalo fence) were sampled (Figure 1). A summary of vegetation and habitat complexity in the different classes of sampled temporary floodplains is presented in Table 1.
Field samplingFloodplain habitat characteristicsEach study floodplain was sampled in May 2009 during the rising flood phase, in July 2009 at flood peak, and in October 2009 and February and March 2010 during the long flood recession phase. The rising flood phase was charac-terised by encroachment of floods into the dry savannah grasslands, the peak flooding phase was defined as the maximum water levels reached within the study floodplains, and the receding phase was characterised by gradual drying up of floodplains. The floodplain habitat variables considered included percentage cover by aquatic plants, woody debris (mainly comprising floating leaf litter, root masses and any other woody materials) and substrates. Aquatic vegetation abundance was expressed as percentage cover and the proportion (%) of woody debris at each study floodplain was determined following procedures described by Gorman and Karr (1978), Schlosser (1982) and Medeiros (2004). The dominant substrate of each study floodplain was also visually characterised as mud, silt-sand, gravel or clay. These variables were assessed from 10 (1 m2) quadrats that were randomly selected fromthe study floodplains.
Water sampling and physicochemical analysesPhysical water parameters were measured in situ. These included temperature and dissolved oxygen (%), measured using an Oxi 330i/set meter, turbidity (Eutech TN-100 turbidimeter), conductivity (Hanna Hi 9033 multi-range conductivity meter) and pH (330i/set). Water samples for total dissolved phosphorus and chlorophyll a were collected following the tube sampling method (Graves and Morrow 1998). Tubes of 6 cm diameter and different lengths depending on the depth of a particular sampling point were used. Sampling tubes were carefully pushed through the water column or sedges to 5 cm above the sediments so as to avoid disturbing the bottom sediments. Samples that were contaminated with bottom sediments were discarded. Sampling points at each study floodplain were randomly selected to cover deeper and more pelagic-open water zone (with a depth range of 1–1.18 m), sedges (0.6–1 m) and inundated grasses (less than 40 cm). On average 34–40 water samples were collected from each study floodplain to constitute a composite sample.
Water samples for determining chlorophyll a and total dissolved phosphorus were collected from these composite samples for analysis in the laboratory using standard procedures (APHA 1998). Water samples for chlorophyll a were filtered through GF/C filters (47 mm) and the filtrates were immediately stored in the dark, at low temperature (<4 °C), until further processing was possible. Chlorophyll a was determined spectrophotometrically at wavelength 663 nm in acetone extracts, using a Perkin Elmer, Lambda 20 UV/VS spectrometer.
Sampling for small fishes At each study floodplain a 3.77 m2 throw-trap net, modified from Pelicice et al. (2005), a gear used for sampling fish associated with macrophytes in Rosana, Brazil, was used to sample for small fishes. The throw-trap consisted of two ring-frames of 1.2 m in diameter (Figure 2). The light plastic ring-frame was fitted at the top of the net and a heavy metal ring was used to support the base. The two frames were joined to each other by a net of 1 mm mesh diameter. The tubular throw-trap net with both ends open had a height of 1.5 m. This sampling gear was considered the most suitable sampling approach because low conductivity and the dense emergent vegetation present at many sampling stations would have reduced the efficiency of electrofishing and seine netting, respectively.
At each sampling site, the net was deployed from the edges of the floodplains and the heavy bottom-metal frame was quickly pressed to the bottom, at the same time suspending the top frame with the aid of a pole to prevent the trapped fish from escaping. Using a 2 m stadia rod, several depth measurements were taken from the enclosed area, and the measurements were pooled to calculate mean depth. Vegetation was removed from inside the trap before fish were scooped out using an aluminium dip net (0.20 m × 0.45 m frame and 1 mm mesh size). Scooping was done inside the trap for at least 10 successive sweeps until no fish were captured. Due to dense sedges and the occurrence of crocodiles, sampling of the inundated shallow grass zones was constrained to a depth of usually less than 40 cm.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

African Journal of Aquatic Science 2011, 36(3): 309–320 311
Preliminary trials with the throw-trap net indicated that five deployments of the trap consistently represented the composition of the fish families within the study floodplains. As a result, five throw-trap samples per site were consid-ered sufficient. All fish specimens were preserved in 75% ethanol for laboratory analysis. In the laboratory, identifi-cation of sampled fish to species level was mainly based on bigger fishes (total length [TL] >20 mm) and it was only possible to identify the small juveniles (TL <20 mm) of
the most challenging families such as cichlids to family level. Taxonomic keys by Skelton (2001) were used during identification. The volume of sampled water was estimated by multiplying the surface area of enclosed area by the average depth of the same area. Subsequently, fish densities were standardised to individuals per m3 of sampled water and the relative abundance of each taxon was calculated as the percentage contributionto the density.
22°30' E
22°30' E22° E 23° E 23°30' E 24° E
20°0' S
19°0' S
19°30' S
18°30' S
0 40 80 km
Sampling sitesMain channel systems
OkavangoDelta
OKAVANGODELTA
BOTSWANA
AFRICA
Botswana
MOHEMBO
MAUN
Okavango
Nqoga
Boro
Xudum
Thao
ge
Chief’s Island
1 2
3
4
56
78
Figure 1: Map of the Okavango Delta showing the locations of the study floodplains. 1 = Lechwe, 2 = Floodplain C, 3 = Hippo floodplain, 4 = Aldrovanda, 5 = Water lily, 6 = Wildebeest, 7 = Daunara, 8 = Buffalo fence
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

Siziba, Chimbari, Mosepele and Masundire312
In addition, Lundgren multi-mesh nylon monofilament gillnets were used to obtain a more complete qualitative list of fish species present within the study floodplains. Lundgren nets, consisting of 12 × 3 m long panels with stretched mesh sizes of 10, 12.5, 16, 20, 25, 31, 39, 48, 58, 70, 86 and 110 mm were set within the study floodplains for approximately 12 h. These were set in deeper waters of the study floodplains, including the sedge-dominated zone and open waters, in July and October 2009 and February 2010. Fish collected were classified using taxonomic keys by Skelton (2001).
Statistical analysesHierarchical cluster analysis (Blackith and Reyment 1971) based on the Ward’s method with Euclidean distance was used to ascertain differences between study floodplains based on physicochemical water parameters. Before clustering, the data were standardised into z-scores since the considered variables were measured on different scales. Cluster analysis was employed to justify pooling data collected from different study floodplains by floodplain classes. Hierarchical cluster analysis was conducted using SPSS Release 17.0 software (SPSS, Inc., Chicago).
Data on environmental variables, including tempera-ture, pH, dissolved oxygen, oxygen percentage satura-tion, conductivity, turbidity, total dissolved phosphorus and chlorophyll a, aquatic vegetation cover and woody debris, were first tested for normality and homogeneity of variance using Kolmogorov-Smirnov and Levene’s tests, respectively. The data conformed to the assumptions of parametric tests. Data were then grouped by different floodplain types and different hydrological phases of flooding and were subsequently compared using two-way analysis of variance (ANOVA). Scheffe tests were conducted on significant ANOVAs to determine the source of variations. The significance levels for ANOVAs and Scheffe tests were set at p < 0.05.
Diversity indices based on data from the throw-trap net were calculated to describe the assemblages for small fishes across the different temporary floodplains. The Shannon-Wiener’s diversity (H’), Margalef’s diversity and Pielou’s evenness indices were calculated and a nonparametric test, Kruskal-Wallis (H) at p < 0.05, was used to compare these indices across the floodplain types. These indices were calculated and compared using PAST (PAleontological STatistics) software (Hammer et al. 2001). Density of small fishes was log-transformed (log[x + 1]) to improve on the assumptions of normality and homogeneity of variance before a two-way ANOVA was employed to assess the statistical differences across different floodplain types and by different hydrological phases of flooding. Statistical signif-icance was set at p < 0.05. Spearman rank correlations were calculated to assess the level of significance of relationships (if any) between the dominant fish families and floodplain habitat variables in the temporary floodplains of the Delta.
Characteristic Primary floodplains Secondary floodplains Rarely flooded floodplainsTerrestrial forests Absent Absent Dominated by Acacia spp. and Colopho-
spermum mopane forestsAquatic vegetation Dominated by sedges (Cyperus
articulatus and Schoenoplectus corymbosus) and submerged macrophytes
Same as primary floodplains Macrophytes were absent during the early phases of flooding and developed during the flood recession phase
Microhabitats Three distinct microhabitats: open water, sedges and inundated grasses
Same as primary floodplains Two microhabitats: open water and inundated grasses, at the onset of flooding and macrophytes appeared during the flood recession phase
Inundated grasses Dominated by Sporobolus spicatus, Cynodon dactylon, Vetiveria nigra-tana and Panicum repens
Same as primary floodplains Dominated by grasses belonging to the Poaceae
Table 1: Some characteristics of temporary floodplains of the lower Okavango Delta
Wooden pole forsuspending the net
Light PVC frame
Net (1 mm mesh)
Heavy metal frame
1.5
m
1.2 m
Figure 2: Sampling small fishes at one of the study floodplains, and schematic diagram of throw-trap net
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

African Journal of Aquatic Science 2011, 36(3): 309–320 313
Results
A dendrogram of the hierarchical cluster analysis of all eight study floodplains based on coefficients of similarity measured by Euclidean distance is shown in Figure 3. At a Euclidean distance of approximately 40, three major clusters were formed and these clusters separated floodplains into primary, secondary and rarely flooded floodplains (Figure 3). This confirmation of similarity among floodplains of the same type justified the pooling of data by floodplain types in the presentation of results.
Floodplain habitat characteristicsFloodplains selected from the primary and secondary floodplains had a higher percentage cover by aquatic vegeta-tion (Figure 4), whereas those from rarely flooded floodplains had lower macrophyte cover but higher proportions of woody debris (Figure 4). The variations in aquatic vegetation cover and woody debris across the floodplain types were signif-icant (ANOVA, p < 0.05). In rarely flooded floodplains, macrophytes were absent during the rising flood phase, but they established over time to record a high percentage cover during the late phases of flooding. During the late phases of flooding, Ludwigia stolonifera and Lagarosiphon spp. were dominant within the rarely flooded floodplains, whereas the sedges Cyperus articulatus and Schoenoplectus corymbosus were dominant throughout in primary and secondary floodplains. The substrates of primary and secondary floodplains were classified as being silt/sandy, whereas the rarely flooded floodplains had muddy–clay substrates.
Physicochemical water parametersSpatial variations of the physicochemical water parameters in primary, secondary and rarely flooded floodplains in different phases of flooding are shown in Table 2. While temperature did not show a clear pattern across the different floodplain types, it was significantly higher (ANOVA, p < 0.05) in all the floodplain types during the flood recession phase. Lowest water temperatures were recorded during peak flooding, coinciding with the cold winter months (Table 2). Dissolved oxygen (%) was highest in rarely flooded floodplains (ANOVA, p < 0.05), and a significant decline in dissolved oxygen across all the floodplains was also noted during the flood recession phase (ANOVA, p < 0.05).
The water of the floodplains was slightly neutral to acidic, and pH ranged between 6.0 and 7.7. The pH was significantly correlated to dissolved oxygen (Pearson r = 0.51, p < 0.05). Turbidity did not vary significantly (ANOVA, p > 0.05) across the different hydrological phases but was considerably higher in rarely flooded floodplains. The highest mean of 38.6 nephelometric turbidity units (NTU) was recorded in rarely flooded floodplains compared to the highest mean of 2.8 NTU in primary floodplains. Significant correlations (Pearson r = 0.69, p < 0.05) between turbidity with woody debris and chlorophyll a were recorded. Conductivity was significantly (ANOVA, p < 0.05) highest in rarely flooded floodplains.
Total dissolved phosphorus and chlorophyll aBoth total dissolved phosphorus and chlorophyll a were significantly highest during the flood recession phase (ANOVA, p < 0.05) and were at their lowest during the
rising flood phase. Except during the flood recession phase, chlorophyll a was generally highest in the rarely flooded floodplains while a noteworthy rise in chlorophyll a (ANOVA, p < 0.05) was observed in secondary floodplains during the flood recession phase (Table 2). Chlorophyll a was signifi-cantly correlated to total dissolved phosphorus (r = 0.26, p < 0.05) and water temperature (r = 0.51, p < 0.05).
Small fishesA total of 38 fish species from 11 families were identi-fied within the study floodplains (Table 3). Fish species richness was highest in primary floodplains and lowest in rarely flooded floodplains (Table 3). The Shannon-Wiener diversity, Margalef’s diversity and Pielou’s evenness indices were highest within the primary floodplains (Figure 5) and lowest in rarely flooded floodplains. The families
64 56EUCLIDEAN DISTANCE
Daunara – RF
Buffalo fence – RF
Water lily – SF
Wildebeest – SF
Floodplain C – PF
Aldrovanda – PF
Hippo – PF
Lechwe – PF
48 40 32 24 16 8
Figure 3: Hierarchical cluster analysis of the study floodplains based on floodplain habitat characteristics and physicochemical water parameters data. PF = primary floodplains, SF = secondary floodplains, RF = rarely flooded floodplains
10
20
30
40
50
60
70
80
Primary Secondary Rarely flooded
PR
OP
OR
TIO
N (%
)
FLOODPLAIN TYPES
Woody debrisVegetation cover
Figure 4: The proportions (%) of aquatic vegetation and woody debris in different floodplain types in the lower Okavango Delta in 2009–2010. Error bars represent the SE of the mean; n = 20 for primary floodplains, n = 10 for secondary floodplains, n = 10 for rarely flooded floodplains
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

Siziba, Chimbari, Mosepele and Masundire314
Distichodontidae and Anabantidae were sampled both in primary and secondary floodplains but not in rarely flooded floodplains. These families accounted for 4.9 % of the total catches of the throw-trap net (Table 4).
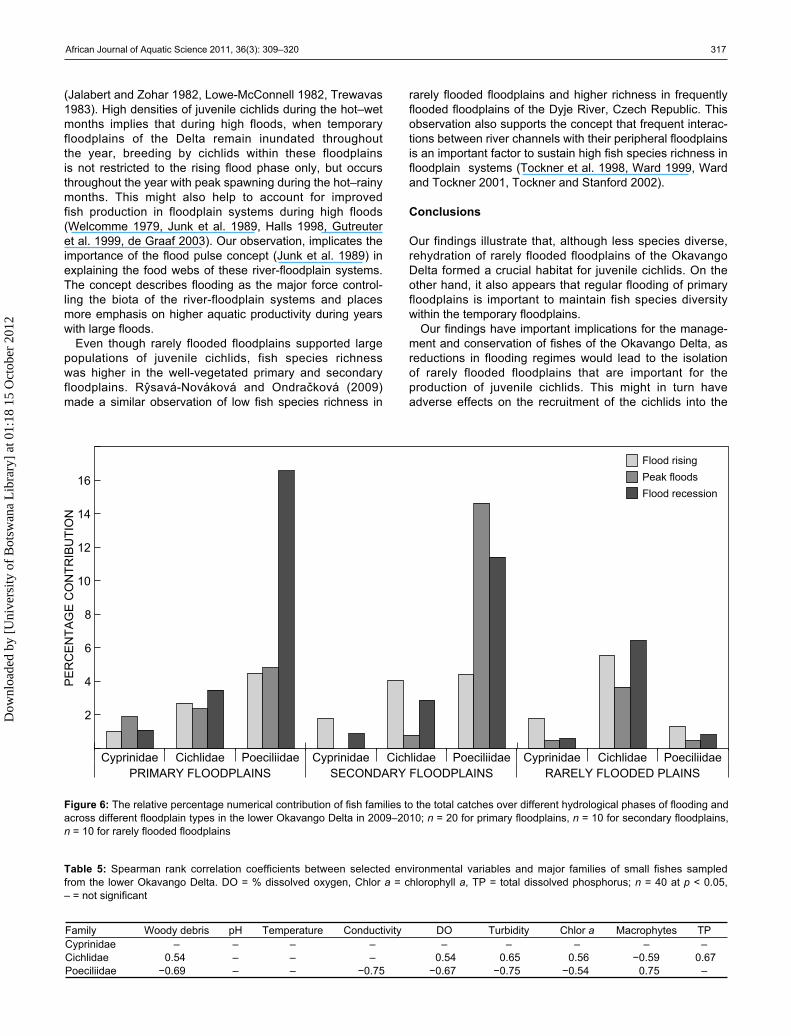
Cichlidae, dominated by juveniles of Tilapia sparrmanii, Pseudocrenilabrus philander and Tilapia rendalli, Poeciliidae (Aplocheilichthys hutereaui) and Cyprinidae (Barbus spp.) were the most abundant across the floodplain types (Table 4). The three dominant families accounted for 93.2% to the total catches of small fishes caught during this study. Although fish densities across different floodplain types were not significantly different (ANOVA, p < 0.05), spatial variations in species composition was observed. Primary and secondary flooded floodplains were dominated by poeciliids, whereas those from the rarely flooded floodplains were dominated by juvenile cichlids (Table 4). Juvenile cichlids constituted 72% of the catch from rarely flooded floodplains and only 21.7% of the catch in frequently flooded floodplains (Table 4). The percentage numerical contribu-tion of the most abundant fish families in different floodplain types to the total catches at different hydrological phases of flooding is illustrated in Figure 6. Rarely flooded floodplains supported the highest abundance (% number) of juvenile cichlids during all phases of flooding.
Densities of small fishes varied between phases of flooding (ANOVA, p < 0.05) with increased density of poeciliids in primary and secondary floodplains during the flood recession
phase (Figure 6). Densities of juvenile cichlids declined during peak flooding (Figure 6). High densities of juvenile cichlids was significantly (Spearman, p < 0.05) correlated with woody debris (r = 0.54), high dissolved oxygen (r = 0.54), high chlorophyll a (r = 0.56) and turbidity (r = 0.65) (Table 5). The highest density of poeciliids both in primary and secondary floodplains was observed at the time when the levels of dissolved oxygen were at their lowest (Spearman r = 0.67, p < 0.05). The density of poeciliids was also associated with macrophytes (Spearman r = 0.75, p < 0.05).
Discussion
The present work illustrates that the rarely flooded floodplains are important habitats for juvenile cichlids, whereas the primary and secondary floodplains supported mainly poeciliids. These observations are similar to those made by Høberg et al. (2002), who also reported high occurrence of juvenile cichlids, poeciliids and cyprinids within the temporary floodplains of the Delta. During the present study, 22 fish species were identified in rarely flooded floodplains compared to 36 sampled from primary floodplains. The relatively high number of fish species identi-fied in rarely flooded floodplains illustrates that a consider-able number of fish species utilise rarely flooded floodplains. Floodplain fishes are known to respond to annual flooding by migrating from dry season refuges to more productive
Variable Primary floodplains (n = 4) Secondary floodplains (n = 2) Rarely flooded floodplains (n = 2)Rising flood phase
Temperature (°C) 23.9 ± 0.6 23.4 ± 0.9 26.2 ± 0.2Oxygen saturation (%) 29.6 ± 4.6 51.2 ± 16.2 72.0 ± 8.3pH 6.5 ± 0.1 6.8 ± 0.6 7.0 ± 0.4Turbidity (NTU) 1.7 ± 0.4 1.9 ± 1.1 38.6 ± 13.9Conductivity (μS cm−1) 93.4 ± 1.4 105.8 ± 6.6 133.6 ± 4.6Total dissolved phosphorus (μg l−1) 27.1 ± 7.1 44.3 ± 6.9 34.8 ± 6.6Chlorophyll a (μg l−1) 14.5 ± 1.2 13.2 ± 1.0 32.6 ± 2.2Woody debris 33.8 ± 2.4 47.5 ± 2.5 82.5 ± 2.5Vegetation cover (%) 60.0 ± 5.0 40.0 ± 3.0 15.0 ± 1.0
Peak flooding phaseTemperature (°C) 20.7 ± 0.4 16.2 ± 1.2 19.9 ± 0.3Oxygen saturation (%) 32.1 ± 3.6 28.5 ± 0.1 60.3 ± 9.8pH 6.8 ± 0.1 6.7 ± 0.1 6.8 ± 0.1Turbidity (NTU) 1.9 ± 0.3 3.4 ± 0.5 9.0 ± 7.0Conductivity (μS cm−1) 104.1 ± 9.1 96.5 ± 3.7 144.8 ± 1.8Total dissolved phosphorus (μg l−1) 47.8 ± 12.5 20.3 ± 6.4 39.5 ± 19.6Chlorophyll a (μg l−1) 25.3 ± 3.8 24.6 ± 6.2 28.1 ± 4.3Woody debris (%) 37.5 ± 2.5 47.5 ± 2.5 75.0 ± 5.0Vegetation cover (%) 76.3 ± 1.3 50.0 ± 2.1 25.0 ± 5.0
Flood recession phaseTemperature (°C) 28.7 ± 0.4 35.6 ± 0.9 28.2 ± 0.7Oxygen saturation (%) 13.4 ± 3.2 14.9 ± 4.5 39.0 ± 5.2pH 5.9 ± 0.3 6.4 ± 0.5 6.3 ± 0.3Turbidity (NTU) 2.8 ± 0.4 7.7 ± 2.0 19.7 ± 3.8Conductivity (μS cm−1) 105.4 ± 6.3 129.8 ± 11.5 109.1 ± 5.3Total dissolved phosphorus (μg l−1) 48.1 ± 5.1 70.8 ± 20.1 73.2 ± 13.3Chlorophyll a (μg l−1) 27.8 ± 3.4 44.4 ± 9.7 29.9 ± 4.1Woody debris (%) 25.0 ± 3.3 40.0 ± 8.2 70.0 ± 1.8Vegetation cover (%) 94.3 ± 3.7 66.3 ± 5.2 65.0 ± 5.0
Table 2: Mean values of selected floodplain variables during different hydrological phases of flooding in the lower Okavango Delta (source: Siziba et al. 2011)
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

African Journal of Aquatic Science 2011, 36(3): 309–320 315
habitats favourable for feeding and spawning (Goulding 1980, Kok 1980, Welcomme 1979, Junk et al. 1989).
Cichlids are known to utilise diverse habitats, feeding mainly on detritus (Lowe-McConnell 1959, Merron 1991, Winemiller and Kelso-Winemiller 2003). Winemiller and Kelso-Winemiller (2003) observed that the detritus that was eaten by tilapiines of the Upper Zambezi was of multiple origins including terrestrial vegetation, periphyton,
phytoplankton and animal droppings. Thus, large quanti-ties of woody debris together with usually higher chloro-phyll a levels within the rarely flooded floodplains might have attracted high populations of cichlids. Furthermore, high densities of invertebrate fauna are known to develop soon after the flooding of habitats rich in woody debris (Pusey and Arthington 2003) and of these zooplankton form an important source of food for juvenile fishes (Brooks
SpeciesPrimary floodplains Secondary floodplains Rarely flooded floodplains
TTN LG TTN LG TTN LGAnabantidae
Microctenopoma intermedium √ – – – – –Cichlidae
Oreochromis andersonii √ √ √ √ √ √Oreochromis macrochir √ √ √ √ √ √Pharyngochromis acuticeps – √ – √ – – Pseudocrenilabrus philander √ – √ – √ –Sargochromis carlottae √ √ – √ – √Sargochromis codringtonii √ √ √ √ √ √Sargochromis greenwoodi – – – – √ √Serranochromis altus – √ – √ – –Serranochromis angusticeps – √ – √ – –Serranochromis robustus – √ – √ – – Serranochromis thumbergi – √ – √ – √Tilapia rendalli √ √ √ √ √ √Tilapia ruweti – – – – √ √Tilapia sparrmanii √ √ √ √ √ √
Characidae Brycinus lateralis √ √ – √ – √
Clariidae Clarias gariepinus √ √ √ √ √ √C. ngamensis – √ – √ √ –
Cyprinidae Barbus afrovernayi √ – – – – – Barbus barnardi √ √ √ – √ –Barbus bifrenatus √ – – – – –Barbus fasciolatus √ – – – – –Barbus haasianus √ – √ – √ –Barbus multilineatus √ – √ – – –Barbus paludinosus √ √ √ √ √ √Barbus poechii √ √ √ √ √ √Barbus radiatus √ – √ – √ –
Distichodontidae Hemigrammocharax multifasciatus √ – √ – – –
Hepsetidae Hepsetus odoe – √ – √ – –
MochokidaeSynodontis spp. – √ - √ –
Mormyridae Marcusenius macrolepidotus √ √ √ √ – √Mormyrus lacerda – √ – – – –Petrocephalus catostoma √ √ √ √ – √
Poeciliidae Aplocheilichthys hutereaui √ – √ – √ √Aplocheilichthys johnstoni √ – √ – – –Aplocheilichthys katangae √ – √ – – –Aplocheilichthys spp. √ – √ – – –
Schilbeidae Schilbe intermedius √ √ √ √ – √
Number of identified species 36 31 22
Table 3: Qualitative list of fish taxa collected from the study floodplains during the 2009–2010 flooding season. √ = present, – absent, TTN = throw-trap net, LG = Lundgren gillnets
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

Siziba, Chimbari, Mosepele and Masundire316
and Dodson 1965, May 1974, Cushing 1990), whereas macroinvertebrates (both terrestrial and aquatic) are also an important food source for adult fish.
Habitats that are rich in woody debris are among the most productive ecosystems on earth (Pusey and Arthington 2003) and are richer in invertebrates than those without woody debris (Jackson and Resh 1989). Woody debris is known to be decomposed to release nutrients (Harmon et al. 1986, Bragg and Kershner 1999, Roth et al. 2007), enhancing invertebrate production (Benke et al. 1984), providing shelter for fishes (Kamdem and Teugels 1997) and some fish directly eat partly decomposed woody debris (Kennard et al. 2001). Due to its ecological importance, spatial variation of fish abundance and assemblages has been related to woody debris abundance (Angermeier and Karr 1984, Everett and Ruiz 1993, Kennard 1995, Sindilariu et al. 2006). Hence, the positive correlation between juvenile cichlids and woody debris provides further evidence for the ecological importance of the inundation of woody debris rich habitats for fish abundance.
Some fish species are also sensitive to high turbidity (Matthews 1987) and this might explain the low populations of poeciliids in rarely flooded floodplains. Although there is a dearth of information on the ecology of A. hutereaui, it is known to inhabit well-vegetated and clear habitats (Wildekamp et al. 1986). An increase in some poeciliid
species has also been associated with periphyton availa-bility and decomposing macrophytes (Chapman et al. 1991). Thus, high relative abundances of poeciliids within primary and secondary floodplains may also be attributed to high presence of aquatic vegetation.
Variations in the composition of small fishes in the Delta’s temporary floodplains may be reflecting the influence of environmental gradients on fish species, as described in other studies (Wright 1986, Yao et al. 2005). Dissolved oxygen is one of the most critical water quality parameters that can affect fish survival and their distribution in aquatic systems (Rendón-Von et al. 2006, Agostinho et al. 2007). However, poeciliids have been observed to survive in habitats with low dissolved oxygen by accessing the water surface for respiration and spending most of their time at the water surface (Kramer and Mehegan 1981). This might help to account for the negative correlation between the abundance of poeciliids and dissolved oxygen observed during this study.
All the study floodplains went through two different phases. The first phase occurred during the rising flood phase until peak flooding and was characterised by comparably high levels of dissolved oxygen, low nutrients and reduced chlorophyll a levels. The second phase occurred when floods were receding and was characterised by high water temperatures, low levels of dissolved oxygen, and high concentrations of both total phosphorus and chlorophyll a. Our results on deteriorating water quality (low dissolved oxygen, increase in total phosphorus and chloro-phyll a) during the flood recession phase confirms similar observations made elsewhere (Chapman and Kramer 1991, Chapman et al. 2000). Ecologically, low levels of dissolved oxygen that coincided with an increase in both total phosphorus and chlorophyll a in these detritus-rich environ-ments can be attributed to high decomposition (Thomaz and de Assis Esteves 1997, Battle and Mihuc 2000, Carlvalho et al. 2005). Furthermore, deteriorating water quality during the flood recession phase can be attributed to the ‘dilution and concentration effect’ that has been described before for other floodplain systems (Welcomme 2001).
In the Okavango Delta, it has been suggested that spawning by most fish species is triggered by the arrival of the floods (Merron 1991). However, our results show that, in addition to small fishes that were observed during the rising flood phase, the density of fishes increased during the flood recession phase, especially during the hot–wet months of February and March 2010. Generally, high breeding activity of tilapias is known to be associated with hot–wet seasons
Shannon-Wiener diversityPielou’s evenness
Margalef's diversity
SPA
TIA
L VA
RIA
TIO
N
2
3
4
1
Primary Secondary Rarely floodedFLOODPLAIN TYPES
Family Primary floodplains Secondary floodplains Rarely flooded floodplainsPoeciliidae 52.5 60.7 12Cichlidae 21.7 20.0 72.0Distichodontidae 4.3 0.5 0Cyprinidae 12.1 11.5 15.0Mormyridae 2.8 0 0Clariidae 2.8 1 1Anabantidae 3.8 6.7 0
Table 4: Percentage numeric contribution of small fishes by families sampled (using throw-trap net) across the study floodplain types; n = 20 for primary floodplains, n = 10 for secondary floodplains, n = 10 for rarely flooded floodplains
Figure 5: Spatial variations in the diversity of small fishes of temporary floodplains in the lower Okavango Delta in 2009–2010. Error bars represent the SE of the mean; n = 20 for primary floodplains, n = 10 for secondary floodplains, n = 10 for rarely flooded floodplains
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012
African Journal of Aquatic Science 2011, 36(3): 309–320 317
(Jalabert and Zohar 1982, Lowe-McConnell 1982, Trewavas 1983). High densities of juvenile cichlids during the hot–wet months implies that during high floods, when temporary floodplains of the Delta remain inundated throughout the year, breeding by cichlids within these floodplains is not restricted to the rising flood phase only, but occurs throughout the year with peak spawning during the hot–rainy months. This might also help to account for improved fish production in floodplain systems during high floods (Welcomme 1979, Junk et al. 1989, Halls 1998, Gutreuter et al. 1999, de Graaf 2003). Our observation, implicates the importance of the flood pulse concept (Junk et al. 1989) in explaining the food webs of these river-floodplain systems. The concept describes flooding as the major force control-ling the biota of the river-floodplain systems and places more emphasis on higher aquatic productivity during years with large floods.
Even though rarely flooded floodplains supported large populations of juvenile cichlids, fish species richness was higher in the well-vegetated primary and secondary floodplains. Rŷsavá-Nováková and Ondračková (2009) made a similar observation of low fish species richness in
rarely flooded floodplains and higher richness in frequently flooded floodplains of the Dyje River, Czech Republic. This observation also supports the concept that frequent interac-tions between river channels with their peripheral floodplains is an important factor to sustain high fish species richness in floodplain systems (Tockner et al. 1998, Ward 1999, Ward and Tockner 2001, Tockner and Stanford 2002).
Conclusions
Our findings illustrate that, although less species diverse, rehydration of rarely flooded floodplains of the Okavango Delta formed a crucial habitat for juvenile cichlids. On the other hand, it also appears that regular flooding of primary floodplains is important to maintain fish species diversity within the temporary floodplains.
Our findings have important implications for the manage-ment and conservation of fishes of the Okavango Delta, as reductions in flooding regimes would lead to the isolation of rarely flooded floodplains that are important for the production of juvenile cichlids. This might in turn have adverse effects on the recruitment of the cichlids into the
Family Woody debris pH Temperature Conductivity DO Turbidity Chlor a Macrophytes TPCyprinidae – – – – – – – – –Cichlidae 0.54 – – – 0.54 0.65 0.56 −0.59 0.67Poeciliidae −0.69 – – −0.75 −0.67 −0.75 −0.54 0.75 –
Table 5: Spearman rank correlation coefficients between selected environmental variables and major families of small fishes sampled from the lower Okavango Delta. DO = % dissolved oxygen, Chlor a = chlorophyll a, TP = total dissolved phosphorus; n = 40 at p < 0.05,– = not significant
Flood risingPeak floodsFlood recession
2
4
6
8
10
12
14
16
Cyprinidae Cichlidae Poeciliidae Cyprinidae Cichlidae Poeciliidae Cyprinidae Cichlidae PoeciliidaePRIMARY FLOODPLAINS SECONDARY FLOODPLAINS RARELY FLOODED PLAINS
PE
RC
EN
TAG
E C
ON
TRIB
UTI
ON
Figure 6: The relative percentage numerical contribution of fish families to the total catches over different hydrological phases of flooding and across different floodplain types in the lower Okavango Delta in 2009–2010; n = 20 for primary floodplains, n = 10 for secondary floodplains, n = 10 for rarely flooded floodplains
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

Siziba, Chimbari, Mosepele and Masundire318
fishery. Furthermore, significant reductions in the inundation frequency of the regularly flooded floodplains may negatively affect fish species richness in these systems, resulting in the loss of some fish species.
Acknowledgements — The study was sponsored by the Carnegie Regional Initiative for Science Education (RISE). We thank Prof. Lars Ramberg, formerly of the Okavango Research Institute (ORI), Maun, for his inputs during conceptualisation of this study. Kemosedile Thebe and Kaelo Makati, ORI, Maun, assisted during field data collection. Selection of the study sites was done under My Community, Our Earth (MyCOE), an initiative aimed at improving the application of GIS and remote sensing in Africa, and the involve-ment of Dr Michael Murray-Hudson, ORI, Maun, in that project is appreciated. We are also grateful to two anonymous reviewers who commented extensively on this manuscript.
References
Aarts BGW, van den Brink FWB, Nienhuis PH. 2004. Habitat loss as the main cause of the slow recovery of fish faunas of regulated large rivers in Europe: the transversal floodplain gradient. River Research and Applications 20: 3–23.
Agostinho A, Pelicice F, Petry A, Gomes L, Júlio H. 2007. Fish diversity in the upper Paraná River basin: habitats, fisheries, management and conservation. Aquatic Ecosystem Health and Management 10: 174–186.
Angermeier PL, Karr JR. 1984. Relationships between woody debris and fish habitat in a small warm-water stream. Transactions of the American Fish Society 113: 716–726.
APHA (American Public Health Association). 1998. Standard methods for the examination of water and wastewater. Washington, DC: American Public Health Association.
Battle JM, Mihuc TB. 2000. Decomposition dynamics of aquatic macrophytes in the lower Atchafalaya, a large floodplain river. Hydrobiologia 418: 123–136.
Benke AC, Vanarsdall TC, Gillespie DM. 1984. Invertebrate productivity in a subtropical blackwater river: the importance of habitat and life history. Ecological Monographs 54: 25–63.
Blackith RE, Reyment RA. 1971. Multivariate morphometrics. London: Academic Press.
Bonyongo MC. 2004. The ecology of large herbivores in the Okavango Delta, Botswana. PhD thesis, University of Bristol, UK.
Bragg DC, Kershner JL. 1999. Coarse woody debris in riparian zones: opportunity for interdisciplinary interaction. Journal of Forestry 97: 30–35.
Brooks, JL, Dodson SI. 1965. Predation, body size and the composition of plankton. Science 150: 26–35.
Carlvalho P, Thomaz SM, Bini LM. 2005. The effect of temperature on decomposition of a potential nuisance species: the submerged aquatic macrophyte Egeria najas planchon (Hydrocharitaceae). Brazilian Journal of Biology 65: 51–60.
Chapman LJ, Chapman CA, Crisman TL, Prenger J. 2000. Predictors of seasonal oxygen levels in a Ugandan swamp/river system: a 3-year profile. Verhandlungen Internationale Vereinigung Limnologie 27: 3048–3053.
Chapman LJ, Kramer DL. 1991. Limnological observations of an intermittent dry tropical forest stream. Hydrobiologia 226: 153–166.
Chapman LJ, Kramer LD, Chapman AC. 1991. Population dynamics of the fish Poecilia gillii (Poeciliidae) in pools of an intermittent tropical stream. Journal of Animal Ecology 60: 441-453.
Cushing D. 1990. Plankton production and year-class strength in fish populations: an update of the match/mismatch hypothesis. Advances in Marine Biology 26: 249–293.
Cushing D, Harris J. 1973. Stock and recruitment and the problem of density dependence. Rapport et Proces Verbaux 164: 142–155.
de Graaf G. 2003. Dynamics of floodplain fisheries in Bangladesh, results of 8 years fisheries monitoring in the compartmentalization pilot project. Fisheries Management and Ecology 10: 191–199.
Everett RA, Ruiz GM. 1993. Coarse woody debris as a refuge from predation in aquatic communities. Oecologia 93: 475–486.
Gehrke PC, Brown P, Schiller CB, Moffatt DB, Bruce AM. 1995. River regulation and fish communities in the Murray-Darling River system, Australia. Regulated Rivers: Research and Management 11: 363–375.
Gorman OT, Karr JR. 1978. Habitat structure and stream fish communities. Ecology 59: 507–515.
Goulding, M. 1980. The fishes and the forest: explorations in Amazonian natural history. Berkeley: University of California Press.
Graves KG, Morrow JC. 1998. Tube sampler for zooplankton. Progressive Fish Culturist 50: 182–183.
Gutreuter S, Bartels AD, Irons K, Sandheinrich MB. 1999. Evaluation of the flood-pulse concept based on statistical models of growth of selected fishes of the upper Mississippi River system. Canadian Journal of Fisheries and Aquatic Sciences 56: 2282–2291.
Halls AS. 1998. An assessment of the impact of hydraulic engineering on floodplain fisheries and species assemblages in Bangladesh. PhD thesis, University of London, UK.
Hammer Ø, Harper DAT, Paul D. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9.
Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory SV, Lattin JD, Anderson NH, Cline SP, Aumen NG, Sedell JR et al. 1986. Ecology of coarse woody debris in temperate ecosystems. Advances in Ecological Research 15: 133–302.
Hjort J. 1926. Fluctuations in the year classes of important food fishes. ICES Journal of Marine Science 1: 5–38.
Høberg P, Lindholm M, Ramberg L, Hessen D. 2002. Aquatic food web dynamics on a floodplain in the Okavango Delta, Botswana. Hydrobiologia 470: 23–30.
Humphries P, Lake PS. 2000. Fish larvae and the management of regulated rivers. Regulated Rivers: Research and Management 16: 421–432.
Jackson JK, Resh VH. 1989. Distribution and abundance of adult aquatic insects in forest adjacent to a northern California stream. Environmental Entomology 18: 278–283.
Jalabert B, Zohar Y. 1982. Reproductive physiology in cichlid fishes, with particular reference to Tilapia and Sarotherodon. In: Pullin RSV, Lowe-McConnell RH (eds), The biology and culture of Tilapias. Manila: International Center for Living Aquatic Resources Management. pp 129–140.
Junk WJ, Bayley PB, Sparks RE. 1989. The flood pulse concept in river-floodplain system. Canadian Journal of Fisheries and Aquatic Sciences, Special Publication 106: 110–127.
Jurajda P. 1995. Effect of channelisation and regulation on fish recruitment in a flood-plain river. Regulated Rivers: Research and Management 10: 207–215.
Kamdem TA, Teugels GG. 1997. Patterns of microclimate use among fourteen fishes of the lower Ntem River basin (Cameroon). Aquatic Living Resources 10: 289–298.
Kennard MJ. 1995. Factors influencing freshwater fish assemblages in floodplain lagoons of the Normanby River, Cape York Peninsula: a large tropical Australian River. MSc thesis, Griffith University, Australia.
Kennard M, Pusey B, Arthington A. 2001. Trophic ecology of freshwater fishes in Australia. Summary report for freshwater ecology scoping study. Brisbane: Griffith University.
Kok HM. 1980. Ecological studies of some important fish species of the Pongolo floodplain, KwaZulu, South Africa. PhD thesis, University of Natal, Pietermaritzburg, South Africa.
Kramer DL, Mehegan JP. 1981. Aquatic surface respiration, an adaptive response to hypoxia in the guppy, Poecilia reticulata (Pisces, Poeciliidae). Environmental Biology of Fishes 6: 299–313.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

African Journal of Aquatic Science 2011, 36(3): 309–320 319
Lindholm M, Hessen D, Mosepele K, Wolski P. 2007. Food webs and energy fluxes on a seasonal floodplain: the influence of flood size. Wetlands 27: 775–784.
Lowe-McConnell RH. 1959. Breeding behaviour patterns and ecological differences between Tilapia species and their signifi-cance for evolution within the genus Tilapia (Pisces: Cichlidae). Proceedings of the Zoological Society of London 132: 1–30.
Lowe-McConnell RH. 1982. Tilapias in fish communities. In: Pullin RSV, Lowe-McConnell RH (eds), The biology and culture of tilapias. Manila: International Center for Living Aquatic Resources Management. pp 83–113.
Matthews WJ. 1987. Physicochemical tolerance and selectivity of stream fishes as related to their geographic ranges and local distributions. In: Matthews WJ, Heins DC (eds), Community and evolutionary ecology of North American stream fishes. Norman, Oklahoma: University of Oklahoma Press. pp 111–120.
May RM. 1974. Larval mortality in marine fishes and the critical period concept. In: Blaxter JH (ed.), The early history of fish. New York: Springer-Verlag. pp 3–19.
Mbaiwa J. (2004). Causes and possible solutions to water resource conflicts in the Okavango River Basin: the case of Angola, Namibia and Botswana. Physics and Chemistry of the Earth 29: 1319–1326.
Medeiros E. 2004. Trophic ecology and energy sources for fish on the floodplain of a regulated dry land river: Macintyre River, Australia. PhD thesis, Griffith University, Australia.
Merron GS. 1991. The ecology and management of the fishes of the Okavango Delta, Botswana, with particular reference to the role of the seasonal flood. PhD thesis, Rhodes University, South Africa.
Merron GS, 1993a. The diversity, distribution and abundance of the fishes in the Moremi Wildlife Reserve, Okavango Delta, Botswana. South African Journal of Wildlife Research 23: 115–122.
Merron GS. 1993b. Pack-hunting in two species of catfish, Clarias gariepinus and C. ngamensis, in the Okavango Delta, Botswana. Journal of Fish Biology 43: 575–584.
Merron GS, Holden KA, Bruton MN. 1990. The reproductive biology and early development of the African pike, Hepsetus odoe, in the Okavango Delta, Botswana. Environmental Biology of Fishes 28: 215–235.
Mosepele K. 2000. Preliminary length-based stock assessment of the main exploited stocks of the Okavango Delta fishery. MPhil thesis, University of Bergen, Norway.
Mosepele K, Basimane O, Mosepele B, Thethela B. 2005. Using population parameters to separate fish stocks in the Okavango Delta fishery, Botswana: a preliminary assessment. Botswana Notes and Records 37: 292–305.
Mosepele K, Kolding J. 2003. Fish stock assessment in the Okavango Delta: preliminary results from a length-based analysis. In: Bernard T, Mosepele K, Ramberg L (eds), Environmental monitoring of tropical and subtropical wetlands. Maun: Harry Oppenheimer Okavango Research Centre. pp 363–390.
Mosepele K, Mosepele B. 2005. Spatial and temporal variability in fishery and fish community structure in the Okavango Delta, Botswana: implications towards fisheries management. Botswana Notes and Records 37: 280–291.
Murray-Hudson M, Combs F, Wolski P, Brown MT. 2011. A vegetation-based hierarchical classification for seasonally pulsed floodplains in the Okavango Delta, Botswana. African Journal of Aquatic Science 36: 223–234.
Murray-Hudson M, Wolski P, Ringrose S. 2006. Scenarios of the impact of local and upstream changes in climate and water use on hydro-ecology in the Okavango Delta, Botswana. Journal of Hydrology 331: 73–84.
Pelicice FM, Agostinho AA, Thomaz SM. 2005. Fish assemblages associated with Egeria in a tropical reservoir: investigating the effects of plant biomass and diel period. Acta Oecologica 27: 9–16.
Pusey BJ, Arthington AH. 2003. Importance of the riparian zone to the conservation and management of freshwater fish: a review. Marine and Freshwater Research 54: 1–6.
Ramberg L, Hancock P, Lindholm M, Meyer T, Ringrose S, Sliva J, van As J, Vanderpost C. 2006. Species diversity of the Okavango Delta, Botswana. Aquatic Sciences 68: 310–337.
Rendón-Von OJ, Memije M, Ortiz A, Soares A, Guilhermino L. 2006. An integrated approach to assess water quality and environmental contamination in the fluvial-lagoon system of the Palizada River, Mexico. Environmental Toxicology and Chemistry 25: 3024–3034.
Roth BM, Kaplan IC, Sass GG, Johnson PT, Marburg IE, Yannarell AC, Havlicek DT, Willis TV, Turner MG, Carpenter SR. 2007. Linking terrestrial and aquatic ecosystems: the role of woody habitat in lake food webs. Ecological Modelling 203: 439–452.
Rŷsavá-Nováková M, Ondračková M. 2009. The importance of surrogate habitats in lowland river floodplains for fish community composition. Fisheries Management and Ecology 16: 468–477.
Schlosser IJ. 1982. Fish community structure and function along two habitat gradients in a headwater stream. Ecological Monographs 52: 395–414.
Schmitten RA. 1999. Essential fish habitat: opportunities and challenges for the next millennium. In: Benaka LR (ed.), Fish habitat: essential fish habitat and rehabilitation. Bethesda: American Fisheries Society. pp 3–10.
Shepherd J, Cushing D. 1980. A mechanism for density-dependent survival of larval fish as the basis of a stock-recruitment relationship. ICES Journal of Marine Science 39: 160–167.
Shepherd J, Cushing D, Beaverton R. 1990. Regulation in fish populations: myth or mirage? Philosophical Transactions: Biological Sciences 330: 151–164.
Sindilariu P, Freyhof J, Wolter C. 2001. Habitat use of juvenile fish in the lower Danube and the Danube Delta: implications for ecotone connectivity. Hydrobiologia 571: 51–61.
Siziba N, Chimbari MJ, Masundire H, Mosepele K. 2011. Spatial and temporal variations of microinvertebrates across temporary floodplains of the lower Okavango Delta, Botswana. Physics and Chemistry of the Earth 36: 939-948.
Skelton P. 2001. A complete guide to the freshwater fishes of southern Africa. Cape Town: Struik.
Thomaz S, de Assis Esteves F. 1997. Secondary productivity (3H-Leucine and 3H-Thymidine incorporation), abundance and biomass of the epiphytic bacteria attached to detritus of Typha domingensis pers. in a tropical coastal lagoon. Hydrobiologia 357: 17–26.
Tockner K, Schiemer F, Ward JV. 1998. Conservation by restoration: the management concept for a river floodplain system on the Danube River in Austria. Aquatic Conservation 8: 71–86.
Tockner K, Stanford JA. 2002. River flood plains: present state and future trends. Environmental Conservation 29: 308–330.
Trewavas E. 1983. Tilapiine fishes of the genus Sarotherodon, Oreochromis and Danakilia. London: British Museum (Natural History).
Ward JV. 1999. Riverine–wetland interactions. In: Sharitz RR, Gibbons JW (eds), Freshwater wetlands and wildlife. Aiken: Savannah River Ecology Laboratory. pp 385–400.
Ward JV, Tockner K. 2001. Biodiversity: towards a unifying theme for river ecology. Freshwater Biology 46: 807–819.
Welcomme RL. 1979. Fisheries ecology of floodplain rivers. London: Longman.
Welcomme RL. 2001. Inland fisheries: ecology and management. Oxford: Blackwell Science.
Wildekamp RH, Romand R, Scheel JJ. 1986. Cyprinodontidae. In: Daget J, Gosse JP, Thys van den Audenaerde DFE (eds), Check-list of the freshwater fishes of Africa. Bruxelles: ISNB; Tervuren: MRAC; Paris: ORSTOM. pp 165–276.
Winemiller KO, Kelso-Winemiller LC. 2003. Food habits of tilapiine
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012

Siziba, Chimbari, Mosepele and Masundire320
cichlids of the Upper Zambezi River and floodplain during the descending phase of the hydrologic cycle. Journal of Fish Biology 63: 120–128.
Wolski P, Murray-Hudson M. 2006. Reconstruction of 1989–2005 inundation history in the Okavango Delta from archival LandSat TM imagery. Poster presented at GLOBWETLANDS Symposium, ESA-ESRIN, Frascati, Italy, 19–20 October 2006.
Wright JM. 1986. The ecology of fish occurring in shallow water creeks of a Nigerian mangrove swamp. Journal of Fish Biology 29: 431–441.
Yao SS, Kouamélan PE, Koné T, N’Douba V, Bi G, Ollevier F, Thys van den Audenaerde DFE. 2005. Fish communities along environmental gradients within the Comoé River basin, Côte d’Ivoire. African Journal of Aquatic Science 30: 185–194.
Dow
nloa
ded
by [
Uni
vers
ity o
f B
otsw
ana
Lib
rary
] at
01:
18 1
5 O
ctob
er 2
012




















