
Geomicrobiology of Iron Layers in the Sediment of Lake Superior
18
ORIGINAL PAPER Geomicrobiology of Iron Layers in the Sediment of Lake Superior M. Dittrich 1 • L. Moreau 1 • J. Gordon 1 • S. Quazi 1 • C. Palermo 1 • R. Fulthorpe 1 • S. Katsev 2 • J. Bollmann 3 • A. Chesnyuk 1 Received: 18 August 2014 / Accepted: 27 February 2015 Ó Springer Science+Business Media Dordrecht 2015 Abstract Previous studies of deeply oxygenated (8–12 cm) sediments in Lake Superior revealed prominent iron (Fe)-rich layers, up to 2 cm thick. Whereas metal enrichments are common near the oxic–anoxic boundary in aquatic sediments, several geomicrobiological aspects of their formation remain unknown. In this study, we analyzed geochemical and microbiological signatures associated with the Fe-rich layer in Lake Superior at two locations (230 and 310-m water depth) in the Eastern Basin of the lake. We measured oxygen and metal contents in the sediment solid phase and imaged the Fe-rich layer with transmission and scanning electron microscope (TEM and SEM). Bacterial communities were characterized using tag-encoded FLX small subunit ribosomal gene amplicon pyrosequencing. The SEM images reveal filamentous structures encrusted with spheres of ca. 20 nm in diameter. TEM and energy dispersive X-ray spectroscopy observations of thin sections indicate that bacteria cells served as nucleation surfaces for Fe-oxide formation. The Fe-rich layers, in contrast to the layers directly above them, were dominated by members of the NC10 phylum, known for anaerobic methane oxidation, and also harbored a high proportion of Nitrospira species that were most similar to members of a Fe-rich seep. Taken together the results suggests that microorganisms, whose cell surfaces are serving as templates for iron oxide precipitates that might also be mediating the iron oxidation, support formation of the Fe-rich layers. Keywords Iron-rich layer Pyrosequencing Nitrospira Sediments Lake Superior Electronic supplementary material The online version of this article (doi:10.1007/s10498-015-9258-y) contains supplementary material, which is available to authorized users. & M. Dittrich [email protected] 1 Department of Physical and Environmental Sciences, University of Toronto Scarborough, Toronto, ON, Canada 2 Large Lakes Observatory and Department of Physics, University of Minnesota Duluth, Duluth, MN, USA 3 Department of Earth Sciences, University of Toronto, Toronto, ON, Canada 123 Aquat Geochem DOI 10.1007/s10498-015-9258-y
Transcript of Geomicrobiology of Iron Layers in the Sediment of Lake Superior

ORI GIN AL PA PER
Geomicrobiology of Iron Layers in the Sedimentof Lake Superior
M. Dittrich1• L. Moreau1
• J. Gordon1•
S. Quazi1 • C. Palermo1• R. Fulthorpe1
•
S. Katsev2• J. Bollmann3
• A. Chesnyuk1
Received: 18 August 2014 / Accepted: 27 February 2015� Springer Science+Business Media Dordrecht 2015
Abstract Previous studies of deeply oxygenated (8–12 cm) sediments in Lake Superior
revealed prominent iron (Fe)-rich layers, up to 2 cm thick. Whereas metal enrichments are
common near the oxic–anoxic boundary in aquatic sediments, several geomicrobiological
aspects of their formation remain unknown. In this study, we analyzed geochemical and
microbiological signatures associated with the Fe-rich layer in Lake Superior at two locations
(230 and 310-m water depth) in the Eastern Basin of the lake. We measured oxygen and metal
contents in the sediment solid phase and imaged the Fe-rich layer with transmission and
scanning electron microscope (TEM and SEM). Bacterial communities were characterized
using tag-encoded FLX small subunit ribosomal gene amplicon pyrosequencing. The SEM
images reveal filamentous structures encrusted with spheres of ca. 20 nm in diameter. TEM
and energy dispersive X-ray spectroscopy observations of thin sections indicate that bacteria
cells served as nucleation surfaces for Fe-oxide formation. The Fe-rich layers, in contrast to
the layers directly above them, were dominated by members of the NC10 phylum, known for
anaerobic methane oxidation, and also harbored a high proportion of Nitrospira species that
were most similar to members of a Fe-rich seep. Taken together the results suggests that
microorganisms, whose cell surfaces are serving as templates for iron oxide precipitates that
might also be mediating the iron oxidation, support formation of the Fe-rich layers.
Keywords Iron-rich layer � Pyrosequencing � Nitrospira � Sediments � Lake Superior
Electronic supplementary material The online version of this article (doi:10.1007/s10498-015-9258-y)contains supplementary material, which is available to authorized users.
& M. [email protected]
1 Department of Physical and Environmental Sciences, University of Toronto Scarborough, Toronto,ON, Canada
2 Large Lakes Observatory and Department of Physics, University of Minnesota Duluth, Duluth,MN, USA
3 Department of Earth Sciences, University of Toronto, Toronto, ON, Canada
123
Aquat GeochemDOI 10.1007/s10498-015-9258-y

1 Introduction
Sediments of several freshwater bodies across the planet contain oxidized iron (Fe) and
manganese (Mn) in the form of nodules or thin metal-rich layers, which have recently
become a topic of emerging interest and speculation (Och et al. 2012; Torres et al. 2014).
Metal-rich layers are well known in marine sediments, where their formation has been
linked either to direct sedimentation from the water column or to diagenetic processes in
the sediment, such as mineralization of organic matter, reductive mineral dissolution,
diffusion of dissolved metals, and mineral re-precipitation (Granina et al. 2004; Kasten
et al. 1998; Richardson 1974). At the oxic–anoxic interface, Fe and Mn are particularly
important due to their redox chemistry: Their reduced forms are highly soluble, while the
solubility of the oxidized species is very low (Stumm and Morgan 1995). The oxidized
phases of Fe and Mn are used as electron acceptors in organic matter mineralization after
the more potent electron acceptors, oxygen and nitrate, are depleted (Berner 1980; Froelich
et al. 1979). The reduced Fe and Mn can be preserved in sediment solids mainly as pyrite,
iron sulfides, or carbonates (Schaller and Wehrli 1997).
Whereas the redistribution of Fe and Mn in marine sediments during early diagenesis has
been extensively recorded and modeled for many years (Robbins and Callender 1975;
Froelich et al. 1979; Aller 1980; Kasten et al. 1998), detailed studies of Fe- and Mn-rich
layers in freshwater sediments are still scarce (Och et al. 2012; Torres et al. 2014; Granina
et al. 2004; Zakharova et al. 2010). Particularly rare are investigations that go beyond simple
geochemistry and include geomicrobiological aspects of the formation of metal-rich layers.
Past research focused mainly on the geochemical processes, such as reduction–oxidation
reactions and redox-sensitive metal redistribution, as well as formation of multiple layers
under non-steady-state conditions. The composition of the associated microbial communities
and their distribution in the sediment are rarely studied, despite the recognized importance of
microorganisms (Posth et al. 2014). In this study, we investigate the composition of the
microbial community associated with Fe-rich sediment layers and visualize the association
of Fe-mineral precipitates and microbes. Our study uses the sediment recovered from Lake
Superior, a large and relatively unproductive lake where sediment is characterized by low
organic matter content and deep penetration of oxygen (Li et al. 2012).
In oligotrophic freshwater lakes, Fe-rich layers occur at neutral pH and puzzlingly, often in
both anoxic and oxic sediment depths. In Lake Constance, a mesotrophic lake in Europe, a Fe-
rich layer was found in sediments 4 cm below the sediment–water interface, in the zone of
oxygen and nitrogen reduction where Fe oxidation can occur (Hauck et al. 2001). An attempt to
identify the group of microorganisms associated with iron oxidation, such as those that may
couple metal oxidation to nitrate reduction, was not successful (Hauck et al. 2001). The distri-
bution of all metabolic groups, such as chemotrophic, mixotrophic, and lithotrophic denitrifiers,
as well as denitrifiers capable of aerobic iron oxidation throughout the upper sediment layers was
relatively uniform (Hauck et al. 2001), leading to the conclusion that no specific microbial
metabolism could be linked to Fe-rich layers in Lake Constance. In oligotrophic Lake Vermilion
(USA), ferromanganese nodules were found near the sediment surface, similar to nodules found
in other lakes around the globe (see references in Sommers et al. 2002). The nodules consisted of
laminated layers enriched in Mn and Fe, and differed in size and morphology. The hypothesis of
a biogenic origin of Fe–Mn nodules has been supported by electron microscopy images of
microbial cells from cultured bacteria able to precipitate Fe and Mn oxides, which in turn served
as substrates for sulfate reducers (Sommers et al. 2002). However, the geochemical data on pore
water and sediment particles were not available in that study.
Aquat Geochem
123

Similar Fe-rich layers and nodules were found in Lake Baikal (Russia), along with high
concentrations of manganese (Baturin et al. 2009). The Fe enrichments occurred as either
irregular nodules or crusts 1 to 4 cm thick (Baturin et al. 2009; Granina et al. 2011). Some
Fe-rich layers were found in the oxidized upper sedimentary layer, whereas others were
found in the reduced sediment below (Baturin et al. 2009). Chemical analyses revealed that
these nodules and crusts consisted of 18.3–84.4 % Fe2O3, 0.57–27.3 % MnO2,
7.94–47.38 % SiO2, and 2.1–14.00 % Al2O3 (Baturin et al. 2009). Several other elements
were found in these formations in trace amounts (Muller et al. 2002), but the main com-
ponents were clearly oxidized iron and manganese, as well as silicon from diatom frustules.
Baturin et al. 2009 proposed that ferromanganese nodules and Fe- and Mn-rich layers
found in the anoxic sediments could be attributed to abiotic processes in Lake Baikal. At
the oxic sediment surface, Fe(II) is oxidized, whereas in anoxic sediment below, iron
precipitates dissolve. The Fe-rich layers are subsequently covered by sediment deposited
above them (Baturin et al. 2009), forming multiple Fe-rich layers in sediments. However,
this explanation does not account for the existence of a well-defined layer, as opposed to
continuous burial of oxidized iron. Taking into consideration possible secondary diagenetic
pathways, Och et al. (2012) hypothesized that Fe and Mn layers could have resulted from
recycling of Fe and Mn oxides, which consist of reductive dissolution of Fe and Mn oxides
below the O2 penetration depth, followed by Fe and Mn oxides precipitation and accu-
mulation. Indeed, in a recent study in Lake Baikal, Torres et al. (2014) found that Fe and
Mn layers can be formed by dissolution of particulate Fe and Mn coupled to the anaerobic
oxidation of methane. Dissolved Fe(II) diffuses upward to reduce particulate Mn(IV) and
thus forming a sharp mineral boundary. It has been also shown that particulate Mn can be
dissolved; hence, an anaerobic nitrification of NH4? has been observed at locations with
Mn(IV).
Most of the studies conducted to date in both marine and freshwater sediments have
focused on abiotic processes in Fe- and Mn-rich layer formation. Arguments exist, how-
ever, that support both abiotic and biotic roles for their origin. Most recent studies in Lake
Baikal show high abundances of Fe and Mn oxidizers close to the metal-enriched layers
(Granina et al. 2010; Zakharova et al. 2010). The biogenic origin of ferromanganese
nodules in freshwater systems is not a new idea. For example, Butkevich (1928) demon-
strated the development of layering in brown water-rich top sediments from the Pechora
Lake and White Seas, and isolated Fe-depositing microorganisms from metal-rich layers.
Direct precipitation of Fe- and Mn-oxides by bacteria has also been postulated (Zakharova
et al. 2010).
Similar to Lake Baikal, Lake Superior has a large surface area, a high abundance of Fe-
rich minerals in its watershed, permanently cold bottom waters, and relatively low bio-
logical productivity (Sterner 2010). Sediments found across the Lake Superior basin
contain prominent Fe-rich crusts, sometimes as multiple layers (Li et al. 2012). Well-
defined Mn and Fe layers in the oxidized upper sediment have been reported at multiple
locations (Sly and Thomas 1974; Li et al. 2012). Though typically located near the depth of
oxygen penetration or nitrogen penetration (Li and Katsev 2014), these layers are also
sometimes found in the presently anoxic or in the presently oxic zones (Richardson and
Nealson 1998). The organic carbon mineralization in Lake Superior is dominated by
aerobic respiration: At the offshore low-sedimentation locations, more than 95 % of carbon
is mineralized using oxygen as electron acceptor (Carlton et al. 1989; Li et al. 2012),
denitrification accounts for less than 2 % (Li and Katsev 2014), while metal reduction and
sulfate reduction account for even smaller fractions (Li and Katsev 2014; Li 2014). At the
Eastern Basin locations investigated here, oxygen penetrates 10–12 cm into the sediment
Aquat Geochem
123

(Li et al. 2012), and in low-sedimentation offshore areas exceeds 6–12 cm (Li et al. 2012;
Li and Katsev 2014). Nitrate is found to penetrate even deeper (Carlton et al. 1989; Li and
Katsev 2014). The overlap of the geochemical profiles of nitrate and ferrous iron provides
evidence for the coupling of iron oxidation to nitrate reduction (Li and Katsev 2014), a
phenomenon known from previous studies (Kasten et al. 2003; Wilson et al. 1985; Berger
1983; Froelich et al. 1979).
The involvement of microorganisms in the oxidation of metals and their roles in the
coupling of Fe and N geochemical cycles remain speculative. Bacteria are primarily in-
volved in reducing Fe (Straub and Buchholz-Cleven, 1998). Their involvement in Fe
oxidation was demonstrated for chemotrophic, denitrifying bacteria (Hauck et al. 2001)
isolated from freshwater ditches, streams and ponds (Straub and Buchholz-Cleven 1998),
as well as Lake Vermilion (Sommers et al. 2002) and Lake Constance. In the latter case,
bacterial strains such as Acidovorax sp. strain BoFeN1 have been shown to oxidize iron
through the reduction of nitrate (Pantke et al. 2012). Verification of the anaerobic oxidation
of ferrous iron by denitrifying bacteria would imply the closing of the iron cycle within the
anoxic zone of the sediment, with implications for both modern geochemical budgets and
the processes that might have shaped the sediment geochemistry during the Earth’s geo-
logical history (e.g., Crowe et al. 2008). Thus, more information is needed on the microbial
populations, functions, and activities.
In this study, we tested the hypothesis that Fe-oxidizing microbes participate in the
formation of Fe-rich layers in Lake Superior. We used electron microscopy to image
sediment sections and analyzed the microbial populations within the iron-rich layers. These
microbial communities have been compared to the communities in overlying layers, in-
cluding the layers enriched in manganese, to understand the origin and diagenesis of the
Fe-rich layers.
2 Materials and Methods
2.1 Sample Collection
Sediment cores were collected from Lake Superior with an Ocean Instruments multi-corer in
September 2010 and July 2012 aboard the R/V Blue Heron. Samples were collected at two
locations in the Eastern Basin of Lake Superior: Eastern (EM, 229 m) Sep 2010, 47� 33.370
86� 35.680 and Eastern Deep (ED, 310 m) Sep 2010, 47� 31.530 87� 07.490 (Fig. 1). Sediment
cores of 94 mm inner diameter ranged 30–35 cm in length. The cores were kept at the in situ
temperature of 4 �C and sectioned in anaerobic conditions in a glove bag to prevent
oxidation. Oxygen vertical distributions were determined within 40 min of sampling in
subsampled cores with an internal diameter of 5 cm and were from the same multicorer in
2010. A Unisense (Clark-type) microelectrode (Revsbech 1989) calibrated in air-saturated
and N2-purged water saturated with bi-dithionite was used for O2 measurements.
2.2 Sediment Chemical Analysis
Sedimentary iron content was determined using a modified approach of that described by
Tessier et al. (1979), Chester and Hughes (1967), (Mehra and Jackson 1960), as well as in
Poulton and Canfield (2005) (see details in Supplementary Material). Here, we present the
results on reducible Fe and Mn oxides sediment depth profiles.
Aquat Geochem
123

2.3 Electron Microscopy and Electron Dispersive X-Ray Spectroscopy
2.3.1 Transmission Electron Microscopy
Transmission electron microscopy (TEM) was employed to study sediment layers for
evidence of microbial cells in Fe-rich layers. Samples were homogenized and then incu-
bated in 2 % glutaraldehyde solution in Sorenson’s phosphate buffer (0.1 M, pH 7.4) to
allow for protein fixation and then washed with ethanol and propylene oxide. Samples were
then embedded in Spurr low-viscosity epoxy, cut into 70 nm sections onto copper grids
and stained with a uranyl acetate and lead citrate mixture. These samples were then
analyzed by transmission electron micrograph using the Hitachi H7500 with the energy of
10 kV at the Centre for the Neurobiology of Stress (CNS) at the University of Toronto,
Scarborough Campus. The Hitachi S-5200 in conjunction with the OXFORD INCA energy
dispersive X-ray spectrometer (SEM–EDS) from the Center for Nanostructure Imaging
(CNI) at the University of Toronto was also used in elemental mapping of samples.
2.3.2 Scanning Electron Microscopy
Scanning electron microscope (SEM) photographs and electron dispersive X-ray (EDX)
mapping were carried out to determine whether bacteria were visible on the iron crust.
Samples were prepared with a gold coating to provide electrical conductivity and achieve
higher image resolution. The images were then taken with a Zeiss Supra VP55 SEM, at the
Geology Department of the University of Toronto. Images were taken at the energy of
10 kV and magnification levels ranging from 5.56–192.74 kx.
Fig. 1 Map of Lake Superior, on the inlet is an image of Fe crust in the sediment sample from the stationEM in 2010
Aquat Geochem
123

2.4 Microbial Community Composition
2.4.1 Enumeration of Bacteria Cell Abundances
Sediment samples from EM station, 6–7 cm, and 9–12 cm, enriched in Fe, collected in 2012
and as well as sediments sampled from the 8–10 cm (Fe-rich layer) and 10–12 cm depths of
cores ED stations collected in 2010, were preserved in Life Guard1 2868-10 soil preservation,
were thawed at room temperature, and were shaken vigorously to break up large particles. The
total cell count of prokaryotes was determined using 4060-diamidin-2-phenylindole (DAPI), a
fluorescent blue dye, which binds to DNA. Cells were separated from the sediment particles
using a procedure modified from Epstein and Rossel (1995). For each sample depth, 0.5 g
preserved sample was combined with 2.0 ml filter-sterilized water, vortexed, and sonicated
for 90 s at 30 W using a Bronwill Scientific Biosonick III ultrasonic cell disrupter, at an
intensity setting of 40/100. The sample was then diluted 10009 and combined with DAPI to
produce a final concentration of 10 lg/ml. After 30 min of incubation in the dark, the solution
was filtered onto a 0.2-lm-pore-size polycarbonate membrane. The filters were placed on
slides in mounting oil and frozen. When counted, samples were thawed for at least 15 min and
viewed using a Nikon Eclipse E600 microscope 1009 magnification oil emersion objective
lens set. Slides were viewed using a DAPI filter of excitation wavelength 340–380 nm. At
12509 magnification, the number of cells per viewing field were counted and recorded, to a
minimum of 400 cells. Concentration factors were used to back calculate to the original
density of bacteria/g of dry sediment (DW) using porosity values.
2.4.2 Community Composition
2.4.2.1 DNA Extraction DNA was extracted and amplified from the sediment samples at
the following sites and depths: ED 7–8 cm, ED 10–12 cm, ED 18–20 cm, EM 7–8 cm, EM
10–12 cm, and EM 18–20 cm. DNA was extracted from the sediment following the pro-
tocol of a PowerSoil DNA isolation kit. The 16S ribosomal DNA (rDNA) region was
amplified in a polymerase chain reaction (PCR) using 341F-clamp forward primer and
907R reverse primer (Supplementary Material Table 1). Amplified DNA was then ana-
lyzed in denaturing gradient gel electrophoresis (DGGE), as described by Straub and
Buchholz-Cleven (1998). The gel was run from 20 h at 70 volts and was stained with
ethidium bromide for 30 min. Two ladders were used: one consisting of known microbes,
and the other of combined samples of 16S extracted from Lake Superior’s sediment.
Using GelCompar II, DGGE fingerprints were analyzed to compare communities at the
iron layers of sediment (10–12 and 18–20 cm), with those above. The comparison was
performed using Pearson’s correlation similarity coefficient, with 2 % optimization, and
UPGMA (unweighted pair group method with arithmetic mean) cluster analysis.
Using a PowerSoil DNA Isolation Kit (MO BIO Laboratories, Inc), approximately
350 lg of each sediment sample was used to extract bacterial genomic DNA. To con-
centrate DNA yield prior to purification, sediment samples were dried in a Savant
SpeedVac centrifugal evaporator (Thermo Fisher Scientific, USA). The DNA samples
were purified to remove amplification inhibitors using Healthcare illustra MicroSpin S-300
HR Columns (General Electric, USA) and following the manufacturer’s protocol. DNA
concentrations were determined using a NanoDrop ND-1000 v.3.7.0 UV–Vis spec-
trophotometer (NanoDrop Technologies, USA) according to the manufacturer’s guidelines.
Extracted and purified DNA samples were stored at -80 �C.
Aquat Geochem
123

2.4.2.2 Pyrosequencing DNA samples from 8 to 10 cm and 10 to 12 cm of cores ED
and EM collected in 2010 were sent to the Research and Testing Lab (Texas) for
pyrosequencing of 16S rRNA gene amplicons generated using barcoded universal pri-
mers 27f-519r. Sequences obtained were initially processed in MOTHUR for trimming
and chimera checking (http://www.mothur.org/wiki/Sequence_processing). Sequences
were aligned, clustered, and dereplicated using tools in the RDP II pyrosequencing
pipeline. Sequences were clustered using various percent similarities as cutoffs to define
OTU’s. Representative sequences from clusters were compared to those in GenBank
using BLASTn and submitted to RDP II classifier in order to obtain a phylogenetic
assignment if possible.
DNA samples from Mn and Fe layers (6–7 cm and 9–12 cm sediment depths, respec-
tively) from EM station collected in 2012 were sent to the MrDNA Molecular Research LP
for pyrosequencing of 16S rRNA gene amplicons. In a modified version of amplicon
pyrosequencing (bTEFAP) initially described by Dowd et al. (2008), a single-step 30 cycle
PCR was performed using HotStarTaq Plus Master Mix Kit (Qiagen, USA) and the same
primers as above.
The PCR was performed under the following conditions: 94 �C for 3 min, then 28
cycles of 94 �C for 30 s; 53 �C for 40 s and 72 �C for 1 min; followed by a final elon-
gation step at 72 �C for 5 min. Amplicon products from different samples were combined
in equivalent concentrations and purified using Agencourt Ampure beads (Agencourt
Bioscience Corporation, USA). Roche 454 FLX titanium instruments and reagents were
used to obtain sequences the samples following the manufacturer’s protocol.
A proprietary analysis pipeline (www.mrdnalab.com, MR DNA, Shallowater, TX) was
used to process the Q25 sequence data derived from the sequencing process. Barcodes and
primers were removed; then, sequences containing fewer than 200 base pairs, sequences
with ambiguous base assignments, and sequences with homopolymer runs greater than six
base pairs were eliminated. Sequences were then deionized, and chimeras were removed.
After removing singleton sequences, the sequences were clustered into OTUs and defined
based on relatedness at 3 % divergence (97 % similarity). Taxonomic classifications of
OTUs were achieved using BLASTn against a curated database derived from Greengenes
(http://greengenes.lbl.gov; DeSantis et al. 2006), RDPII (http://rdp.cme.msu.edu) and
NCBI (www.ncbi.nlm.nih.gov). Sequences were initially processed chimera for trimming
and checking. Sequences were divided into those from the iron layer and those from the
layer directly above using a perl script. Sequences were aligned, clustered, and derepli-
cated using tools in the RDP II pyrosequencing pipeline. Sequences were clustered using
various percent similarities as cut offs to define community microbial relationships.
Representative sequences from clusters were compared to those in GenBank using
BLASTn and submitted to RDP II classifier in order to obtain a phylogenetic assignment if
possible.
3 Results
3.1 Iron and Manganese Layers
The sequential iron extractions from Lake Superior sediments show high amounts of iron
from the dithionite extraction within the 8–10 cm band for site ED, while the EM site has a
broader layer at a depth of 9–12 cm as shown in Fig. 2. The EM also displayed an overall
Aquat Geochem
123
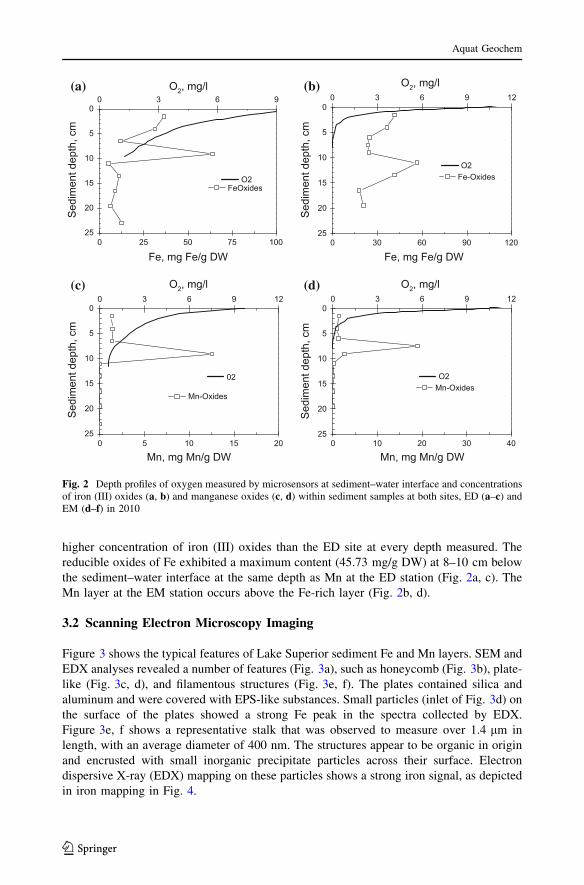
higher concentration of iron (III) oxides than the ED site at every depth measured. The
reducible oxides of Fe exhibited a maximum content (45.73 mg/g DW) at 8–10 cm below
the sediment–water interface at the same depth as Mn at the ED station (Fig. 2a, c). The
Mn layer at the EM station occurs above the Fe-rich layer (Fig. 2b, d).
3.2 Scanning Electron Microscopy Imaging
Figure 3 shows the typical features of Lake Superior sediment Fe and Mn layers. SEM and
EDX analyses revealed a number of features (Fig. 3a), such as honeycomb (Fig. 3b), plate-
like (Fig. 3c, d), and filamentous structures (Fig. 3e, f). The plates contained silica and
aluminum and were covered with EPS-like substances. Small particles (inlet of Fig. 3d) on
the surface of the plates showed a strong Fe peak in the spectra collected by EDX.
Figure 3e, f shows a representative stalk that was observed to measure over 1.4 lm in
length, with an average diameter of 400 nm. The structures appear to be organic in origin
and encrusted with small inorganic precipitate particles across their surface. Electron
dispersive X-ray (EDX) mapping on these particles shows a strong iron signal, as depicted
in iron mapping in Fig. 4.
25
20
15
10
5
0
0 10 20 30 40
0 3 6 9 12
Mn-OxidesO2
25
20
15
10
5
0
0 30 60 90 120
0 3 6 9 12
Sed
imen
t dep
th, c
mS
edim
ent d
epth
, cm
Sed
imen
t dep
th, c
mS
edim
ent d
epth
, cm
Fe-OxidesO2
25
20
15
10
5
0
0 5 10 15 20
0 3 6 9 12
Mn, mg Mn/g DW Mn, mg Mn/g DW
Mn-Oxides
02
25
20
15
10
5
0
0 25 50 75 100
0 3 6 9
Fe, mg Fe/g DW Fe, mg Fe/g DW
FeOxidesO2
O2, mg/l O2, mg/l
O2, mg/lO2, mg/l
(a) (b)
(c) (d)
Fig. 2 Depth profiles of oxygen measured by microsensors at sediment–water interface and concentrationsof iron (III) oxides (a, b) and manganese oxides (c, d) within sediment samples at both sites, ED (a–c) andEM (d–f) in 2010
Aquat Geochem
123

3.3 Transmission Electron Spectroscopy
TEM images revealed small fibril structures roughly 1 lm in length after staining with
uranyl acetate and lead citrate (Fig. 5). Our observations of unstained samples showed
little to none of these structures, indicating that they may be of organic nature. Heavily
encrusted ring structures were also identified with and without staining, and are similar in
diameter to the previously observed stalks from the SEM images. EDX of these ring
Fig. 3 SEM images of iron-layer samples a represents an overview over the sample, b represents a honey-home structures, c–d show a thin Al–Si plates covers with Fe particles and extracellular polymericsubstances. Filamentous structure similar to sheaths of iron-oxidizing bacteria Leptothrix ochracea(e) covered with spherical to irregular shapes particles attached to sheaths/thin filamentous (f)
Aquat Geochem
123

structures revealed them to be encrusted with iron particles, consistent with the previous
observations of stalks. Other elements identified within the encrustation include P, C, Si,
and O as shown in Fig. 6.
3.4 Microbial Community Composition
The prokaryotic cell density in the sediment samples decreases with distance from the
sediment surface, once below the uppermost 0.5 cm of the sediment (Fig. 1 Supplementary
Material). At both the ED and EM sites, the sediment in the first 0–0.5 cm contains a much
lower concentration of bacteria than that in the layer from 0.5 to 1 cm of sediment. At the
ED site, the bacterial concentration was only (7.97 ± 3.30)*108 cells/g dry weight in the
uppermost 0.5 cm of sediment, as opposed to (2.30 ± 2.11)*109 cells/g dry weight found
in the sediment directly below. At the EM site, there were (7.07 ± 4.1)*108 cells/g dry
weight in the uppermost 0.5 cm of sediment, versus (1.09 ± 2.9)*108 cells/g dry weight
living directly below. This strong increase in bacterial abundance at the ED station occurs
within uppermost 0.5 cm (Fig. 1 Supplementary Material). Although bacterial abundance
is a nearly twice as great at the ED site as the EM site, nearly identical trends in abundance
occur. The bacterial abundance increases strongly in the layer 0.5–1 cm beneath the sur-
face. The populations then show exponential decline, rapidly dropping to concentrations of
1.5–2*108 cells/g of dry sediment (ED) and 3–5*108 cells/g of dry sediment (EM). The
density then plateaus, remaining relatively constant within these ranges.
6507 sequence reads were obtained for sample ED 8–10 cm, 4131 for sample ED
10–12cm, 9455 reads for sample EM 8–10 cm, and 5969 reads for sample EM 10–12 cm.
The iron layers, ED 8–10 cm and EM 10–12 cm, both differed from the sediments directly
above them in that they contained far fewer gram-positive organisms, far more pro-
teobacteria, and a high proportion of Nitrospira species within the community as shown in
Supplementary Materials Table 1 and 2. Both sediment layers contain very high propor-
tions of unclassifiable sequences 121 of 189 clusters from the no Fe layer, and 163 of 234
clusters from the iron layers. This is reflected in the data shown in Supplementary Ma-
terials Table 1, where representative sequences of the dominant clusters (those containing
Fig. 4 SEM image and EDX iron mapping of the particle attached to Al–Si plate (a). The gray square onthe left image indicates the boundaries of the mapping shown on the (b)
Aquat Geochem
123

high percentages of the total number of sequences) shown with their closest GenBank
matches. We show here the data for representatives of clusters defined at 20 % similarity of
sequence. This represents a groups of bacteria defined as being roughly in the same family
or order. The same exercise performed with sequences representing clusters defined at 5 %
similarity gives much the same information. Both sediment layers contain very high
proportions of unclassifiable sequences—121 of 189 clusters from the no Fe layer, and 163
Fig. 5 TEM images from the iron-layer sediment samples obtained from site ED (a–d) and site EM (e,f) stained with uranyl acetate and lead citrate, a, c show an overview of the samples with cross sections ofsheaths encrusted by an amorphous to poorly crystalline precipitates appear as circular or elongatedstructures, which are little or heavily encrusted (b, d) showing different level of biomineralization; e,f represent EM site with small little encrusted structures, similar to viruses
Aquat Geochem
123

of 234 clusters from the iron layers. The most dominant cluster (20 %) in Fe-rich layer
contains unclassified bacteria showing the best matches to organisms from Lake Wash-
ington sediments and from groundwater (Table 1 Supplementary Material). The next most
common organism appears to be Nitrospira, with relatively high similarity to organisms
found in an iron II-rich seep (Table 1 Supplementary Material). Other relatively common
organisms (i.e., found in the top ten most abundant clusters) are related to those found in
limestone systems, groundwater systems, soils, or sediments.
4 Discussion
4.1 Origin of Iron and Manganese Layers
Sedimentary Fe and Mn transformations have been studied extensively in marine systems
(Kasten et al. 1998; Maerz et al. 2012) and less intensively in freshwater environments. It is
widely recognized that mineralization of organic matter, which drives the redox zonation in
sediment, leads to the reduction of Mn and Fe (oxy) hydroxide after the oxygen and nitrate
supplies are depleted. The reduced Mn and Fe diffusing from the deeper sediment are
oxidized by oxygen and/or nitrate. As a result, Mn and Fe layers can form at the oxic–anoxic
boundary in the sediment. The position of metal-rich layers within the sediment can be
strongly affected by non-steady-state conditions, which may shift the layers vertically: up on
the reduction time scale or down on the burial time scale (e.g., Katsev et al. 2006). While the
abiotic controls of the involved reactions have been investigated in detail, studies of the
microbial impacts on Fe- and Mn-rich layers in freshwater sediments are still scarce.
Whereas previous studies focused on isolation of microorganisms and mineral pre-
cipitation, our microscopic investigations show microbial cells embedded in Fe oxides,
providing direct evidence of microbial involvement in Fe-rich layer formation (Posth et al.
2014). Our microscopic and spectroscopic images suggest that Fe oxides precipitated on
the surface of cells, so that cells became completely encrusted in Fe particles. The
Fig. 6 TEM-EDX line scans of a sheaths demonstrated a homogenous distribution of mostly Fe, significantamounts of Si, and minor P. Number at the left corner reflects the amplitude of the EDAX-signal
Aquat Geochem
123

microscopic images do not offer details on the mechanism of Fe precipitation, but they do
demonstrate that cell surfaces are able to act as templates for the growth of Fe particles.
The observations stress the importance of microbial-induced Fe oxides formations in
sediments; in addition, any abiotic pathway and or the Fe-oxide precipitation known to be
carried out by microaerophilic or anoxygenic phototrophic Fe oxidizers (Posth et al. 2014).
The images of the encrusted cells from Lake Superior sediments look similar to cells found
in Fe-rich hydrothermal vents (Fortin et al. 1998).
4.2 Microbial Communities in Sediments of Lake Superior
Lake Superior has a low bacterial abundance in comparison with other large lakes. For
example, the microbial cell density measured in different freshwater sediments is typically
between 1.6*109 cells/g DW and 1.7*1010 cells/g DW (Straub and Buchholz-Cleven
1998). Lake Superior’s maximum bacterial density estimate of 2.30 ± 2.11*109 cells/g
DW was only found at the ED site between 0.5 and 1.0 cm depths. Most of the other cell
densities found in the sediment cores from Lake Superior were on the order of 108. In
upper sediment layers, this disparity in bacterial density may be related to sampling in mid-
September, after the growing season has peaked. June bacterial density measurements
observed in a series of temperate freshwater reservoir sediments of different trophic levels
were invariably higher than those taken in the fall (Wobus et al. 2003). However, the low
bacterial density measurements are unsurprising given the lake’s low productivity and low
sedimentary concentration of organic matter (3–5 wt %; Li 2014). In oligotrophic Lakes,
there is less organic matter to act as a nutrient and energy source for bacterial populations.
Surprisingly, Wobus et al. (2003) found no statistically significant differences in total
bacterial population densities between eutrophic and oligotrophic reservoirs. The bacterial
densities measured through DAPI staining for all reservoirs ranged from 1*1010 to
10*1010 cells/g DW, regardless of trophic state (Wobus et al. 2003). However, the study
included only shallow water bodies, so organic matter content even in oligotrophic systems
would have been higher than those in the sediments of Lake Superior.
In their profile of a 23-cm sediment core from Lake Erken, Sweden, Haglund et al.
(2003) found that bacterial abundances decreased with depth, after peaking 0.5 cm below
the sediment interface. The cell density in Lake Erken’s sediment drops from 4*1010 cells/g
DW at 0.5-cm depth to 1.8*1010 cells/g DW at the bottom of sediment. This decrease in
bacterial abundance with depth is a common trend (Haglund et al. 2003). Deep sediment
does not contain a higher percentage of dead cells; the number of viable bacteria at any
depth is approximately 57–63 %, which suggests that neither grazing nor viruses are
primarily responsible for the decrease in abundance (Haglund et al. 2003). Since porosity
decreases with depth, and diffusion from the surface becomes more difficult with increased
distance, reduced organic matter and nutrient availability may be responsible for the de-
cline in Lake Superior’s sediment population with depth.
At most sediment depths, bacteria appear to be twice as abundant in the sediment at the
deep site (ED) than at the shallower one (EM). This may be due to different nutrient
concentration in the pore water at the two sites: the concentrations of ammonium and
phosphate were considerably higher at EM (Li and Katsev 2014; Li 2014). Higher levels of
oxygen and organic matter both allow increased cellular respiration and can support
bacterial communities at greater densities. At the ED site, oxygen levels were higher than
at the EM site, allowing increased respiration. Varying nutritional levels for microorgan-
isms, such as OM content, could also affect growth patterns. These parameters vary most
in the surface sediments, where the largest difference in bacterial populations between the
Aquat Geochem
123

two sites has been found (Fig. 1 Supplementary Material). The difference is less between
the EM and ED site’s bacterial abundance at the lower layers, regions with low metabolic
activities.
The pyrosequencing data show that the microbial community of the non-iron layer
above the iron crust is strongly dominated (46.5 % of reads) by gram-positive bacteria
represented by the Paenisporosarcina species. All other clusters containing relatively high
numbers of sequences are unclassifiable—but do show high similarity to sequences derived
from soils, groundwater areas, deep sediments and/or limestone environments. The Fe-rich
layer community is highly dissimilar; proteobacteria dominate rather than Firmicutes
(Supplementary Materials Table 2, Fig. 7). The dominant strains, representing 20 % of
sequences, are related to candidatus Methylomirabilis, a member of the NC10 phylum that
oxidizes methane via denitrification through an unusual pathway that produces oxygen
from nitrate (Wu et al. 2012; Ettwig et al. 2010). Also of interest is the Nitrospira cluster
representing [5 % of the community, for it the sequences are most similar to an unclas-
sified organism found within Lake Washington and to a microbe found within a Fe-rich
seep (Supplementary Materials Table 2). Iron precipitating microbes have been observed
0
5
10
15
% o
f Seq
uenc
es
ED 8-10 cmEM 8-10 cmEM 10-12 cmED 10-12 cm
0
10
20
40
50
60
% o
f Seq
uenc
es
ED 8-10 cmEM 8-10 cmEM 10-12 cmED 10-12 cm
(a)
(b)
Fig. 7 Percentage of sequences of each phylum for four sediment layers a represents 0–60 % of thesequences, while 0–15 % sequences are shown at (b)
Aquat Geochem
123

in Lake Washington, described as twisted tubules and irregular particulates (Shapiro et al.
1971). The Fe-seep microbes were isolated from a Fe-rich pond in which Fe oxidation was
occurring (Bruun et al. 2010). Sequence matches strongly support the presence of iron
oxidizers in these layers. We postulate these are the species that might precipitate Fe
oxides at the cell surfaces by serving as the templates as shown by TEM experiments.
Pyrosequencing of Fe-rich and above-lying sediment layers from the EM station was
carried out in 2012 and showed agreement with the previous results. Indeed, phyla of
proteobacteria, together with actinobacteria, chloroflexi and nitrospirae, dominated in both
above-lying and Fe-rich layers. Recently, neutrophilic proteobacteria were found in the
environment where microbes actively drive Fe and Mn cycles (Supplementary Materials
Table 2, Baskar et al. 2012), confirming the presence of the similar microbial communities
in Fe-rich habitats. The 2011 data show the presence of Methylomirabilis, a nitrate re-
ducing methane oxidizer that was thought to be NC10.
4.3 Cell-Mineral Interfaces: Biomineralization Phenomenon
In many studies, the identification of iron biomineralization has relied on the observation
of characteristic iron-encrusted structures such as stalks and sheaths (Hanert 2002; James
et al. 2002). Iron-encrusted, filamentous structures observed under SEM in this study
indicate the presence of an iron-oxidizing microorganism.
Studies on Gallionella ferruginea, iron-oxidizing chemolithotrophic bacteria that live in
low-oxygen conditions (Koschinsky et al. 2002) show similarities to organisms within
Lake Superior. G. ferruginea oxidizes dissolved iron, therefore removing it from the water
and producing an insoluble precipitate of ferric hydroxide onto the outside of long, stalk-
like structures.
Studies on BoFeN1, another iron-oxidizing microbe, also find similar encrustation pat-
terns of iron. However, Fe precipitates directly onto the outer membranes of these cells
(Kappler and Straub 2005). Experiments, such as those by Clarke et al. (1997) using TEM
techniques on other iron-biomineralizing microbes, reveal a common theme in the circular
encrustation of microbes with iron precipitates. These structures were observed within the
Lake Superior iron-layer samples, with inorganic precipitates on the outer surface of cells.
The presence of nodules on the outer surface of these structures, as well as the approximate
size correlates with the observed encrustation of the filamentous structures seen under SEM.
SEM-EDX mapping analysis of the observed nodules shows a high concentration of
iron within the nodule structure, leading to the conclusion that the observed particles are
iron precipitates (Fig. 5). Using TEM-EDX, the observed circular structures were analyzed
to determine the nature of the precipitates on their outer edge. Iron concentrations were
found to be higher on these structures than in the background sediment. Carbon was also
found on the ring structures at a much higher reading than the background-embedding
plastic. These results suggest that these structures are organic in nature and may be re-
sponsible for the concentration of iron within the iron layer.
5 Conclusions
We investigated oxidized Fe formations from sediments of Lake Superior. Microorganisms
identified in the Fe-rich layers included many that are capable of iron oxidation and iron
oxide precipitation at their cell surfaces. The identification of DNA sequences of organisms
Aquat Geochem
123

closely related to neutrophilic proteobacteria supports this hypothesis. Sequence matches
strongly support the presence of iron oxidizers in Fe-rich layers. We postulate that these
are the species that may precipitate Fe oxides besides the cell surfaces that serve as the
templates as shown by TEM experiments. The Fe-rich layers, in contrast to the layers
directly above them, were dominated by members of the NC10 phylum, known for
anaerobic methane oxidation, and also harbored a high proportion of Nitrospira species
that were most similar to members of a Fe-rich seep. Electron microscopy reveals
filamentous microbial structures, which were covered with Fe precipitates. Finally, TEM
sections demonstrated that iron oxides as well as silica and phosphorus are mineralized
directly on cell surfaces, supporting the contribution of the biomineralization process on
the formation and preservation of iron and manganese crusts.
Acknowledgments We acknowledge the support from the National Sciences and Engineering ResearchCouncil of Canada (Discovery grant to M. Dittrich), Canada Foundation for Innovation and Ontario Re-search (Fund for Leaders Opportunity Fund, Grant Nr 22404), and the US National Science Foundation(Grant OCE 0961720 to SK). M.D. was supported by start-up funding from the University of TorontoScarborough. We are grateful to Basirath Raoof for help with the laboratory experiments, B. Temkin(UTSC) and D. Holmyard (Mount Sinai Hospital, Toronto) for their assistance with SEM and TEMimaging and Jiying Li for sharing the data and help during the cruises 2010 and 2012. We thank BernhardWehrli and the anonymous reviewer whose comments and suggestions helped us to improve the manuscriptsignificantly.
References
Aller RC (1980) Diagenetic processes near the sediment-water interface of Long Island Sound. II. Fe andMn. In: Saltzman B (ed) Esfuarine physics and chemistry: studies in Long Island Sound. Advances ingeophysics, vol 22. Academic Press, London, pp 35l–415
Baskar S, Baskar R, Thorseth IH, Ovreas L, Pedersen RB (2012) Microbially induced Iron precipitationassociated with a neutrophilic spring at Borra Caves, Vishakhapatnnam, India. Astrobiology12:327–346
Baturin G, Yushina I, Zolotykh E (2009) Variations in the elemental composition of ferromanganesestructures from Lake Baikal. Oceanology 49:505–514
Berger WH, Finkel RC, Killingley JS, Marchig V (1983) Glacial-Holocene transition in deep-sea sediments:manganese-spike in the east-eqautorial pacific. Nature 303:231–233
Berner RA (1980) Early diagenesis. A theoretical approach. Princeton University Press, PrincetonBruun A-M, Finster K, Gunnlaugsson HP, Nørnberg P, Friedrich MW (2010) A comprehensive investigation
on iron cycling in a freshwater seep including microscopy, cultivation and molecular communityanalysis. Geomicrobiol J 27:15–34
Butkevich VS (1928) The marine iron-manganese deposits formation and the participation of micro-orga-nizms in it. Trudy Morskogo Nauchnogo Instituta 3:5–80
Carlton R, Walker G, Klug M, Wetzel R (1989) Relative values of oxygen, nitrate, and sulfate to terminalmicrobial processes in the sediments of Lake Superior. J Great Lakes Res 15:133–140
Chester R, Hughes MJ (1967) A chemical technique for the separation of ferro-manganese minerals,carbonate minerals and adsorbed trace elements from pelagic sediments. Chem Geol 2:249–262
Clarke WA, Konhauser KO, Thomas JC, Bottrell SH (1997) Ferric hydroxide and ferric hydroxysulfateprecipitation by bacteria in an acid mine drainage lagoon. FEMS Microbiol Rev 20:351–361
Crowe SA, Jones C, Katsev S, Magen C, O’Neill AH, Sturm A, Canfield DE, Haffner GD, Mucci A, SundbyB, Fowle DA (2008) Photoferrotrophs thrive in an Archean Ocean analogue. Proc Natl Acad Sci USA105:15938–15943
DeSantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K, Huber T, Dalevi D, Hu P, AndersenGL (2006) Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible withARB. Appl Environ Microbiol 72:141–146
Dowd SE, Sun Y, Wolcott RD, Domingo A, Carroll JA (2008) Bacterial tag-encoded FLX ampliconpyrosequencing (bTEFAP) for microbiome studies: bacterial diversity in the ileum of newly weanedSalmonella-infected pigs. Foodborne Pathog Dis 5:459–472
Aquat Geochem
123

Epstein S, Rossel J (1995) Enumeration of sandy sediment bacteria: search for optimal protocol. Mar EcolProg Ser 117:289–298
Ettwig KF, Butler MK, Le Paslier D, Pelletier E, Mangenot S, Kuypers MMM, Schreiber F et al (2010)Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature 464:543–548
Fortin D, Ferris FG, Scott SD (1998) Formation of Fe-silicates and Fe-oxides on bacterial surfaces insamples collected near hydrothermal vents on the Southern Explorer Ridge in the northeast PacificOcean. Am Miner 83:1399–1408
Froelich PN, Klinkhammer GP, Bender ML, Luedke NA, Heath GR, Gullen D, Dauphin P, Hammond D,Hartman B (1979) Early oxidation of organic matter in pelagic sediments of the eastern equatorialAtlantic: suboxic diagenesis. Geochim Cosmochim Acta 43:1075–1090
Granina L, Mueller B, Wehrli B (2004) Origin and dynamics of Fe and Mn sedimentary layers in LakeBaikal. Chem Geol 205:55–72
Granina L, Mats VD, Phedorin MA (2010) Iron-manganese formations in the Baikal region. Russ GeolGeophys 51:650–660
Granina LZ, Zakharova YP, Parfenova VV (2011) Biogenic Fe and Mn accumulation in the bottom sedi-ments of Lake Baikal. Geochem Int 49:1154–1160
Haglund A, Lantz P, Tornblom E, Tranvik L (2003) Depth distribution of active bacteria and bacterialactivity in lake sediment. FEMS Microbiol Ecol 46:31–38
Hanert HH (2002) Bacterial and chemical iron oxide deposition in a shallow bay on Palaea Kameni,Santorini, Greece: microscopy, electron probe microanalysis, and photometry of in situ experiments.Geomicrobiol J 19:317–342
Hauck S, Benz M, Brune A, Schink B (2001) Ferrous iron oxidation by denitrifying bacteria in profundalsediments of a deep lake (Lake Constance). FEMS Microbiol Ecol 37:127–134
James RE, Scott Sd, Fortin D, Clark ID, Ferris FG (2002) Regulation of Fe3?-oxide formation among Fe2?-oxidizing bacteria. Geomicrobiol J 6:537–543
Kappler A, Straub KL (2005) Geomicrobiological cycling of iron. Rev Miner Geochem 59:85–108Kasten S, Freudenthal T, Gingele FX, Schulz HD (1998) Simultaneous formation of iron-rich layers at
different redox boundaries in sediments of the Amazon deep-sea fan. Geochim Cosmochim Acta62:2253–2264
Kasten S, Zabel M, Heuer V, Hensen C (2003) Processes and Signals of non-steady-state diagenesis inDeep-Sea sediments and their pore waters. In: Wefer G, Mulitza S, Ratmeyer S (eds) The SouthAtlantic in the Late Quaternary. Springer, pp 431–459
Katsev S, Sundby B, Mucci A (2006) Modeling vertical excursions of the redox boundary in sediments:application to deep basins of the Arctic Ocean. Limnol Oceanogr 51:1581–1593
Koschinsky A, Gaye-Haake B, Arndt C, Maue G, Spitzy A, Winkler A, Halbach P (2002) Experiments onthe influence of sediment disturbances on the biogeochemistry of the deep-sea environment. Deep SeaRes Part II 48:3629–3651
Li J (2014) Sediment diagenesis in large lakes Superior and Malawi, geochemical cycles and budgets andcomparisons to marine sediments. Dissertation, University of Minnesota
Li J, Katsev S (2014) Nitrogen cycling in deeply oxygenated sediments: results in Lake Superior andimplications for marine sediments. Limnol Oceanogr 59:465–481
Li J, Crowe SA, Miklesh D, Kistner M, Canfield DE, Katsev S (2012) Carbon mineralization and oxygendynamics in sediments with deep oxygen penetration, Lake Superior. Limnol Oceanogr 56:1634–1650
Maerz C, Stratmann A, Matthiessen J, Meinhardt AK, Eckert S, Schnetger B, Vogt C, Stein R, Brumsack HJ(2012) Manganese-rich brown layers in Arctic Ocean sediments: composition, formation mechanisms,and diagenetic overprint. Geochim Cosmochim Acta 75:7668–7687
Mehra OP, Jackson ML (1960) Iron oxide removal from soils and clays by a dithionite–citrate systembuffered with sodium bicarbonate. In: 7th National conference clays clay minerals, pp 317–327
Muller B, Granina L, Schaller T, Ulrich A, Wehrli B (2002) P, As, Sb, Mo, and other elements insedimentary Fe/Mn of Lake Baikal. Environ Sci Technol 36:411–420
Och LM, Mueller B, Voegelin A, Ulrich A, Goettliche J, Steiniger R, Mangold S, Vologina E, Sturm M(2012) New insights into the formation and burial of Fe/Mn accumulations in Lake Baikal sediments.Chem Geol 330–331:244–259
Pantke C, Obst M, Benzerara K, Morin G, Ona-Nguema G, Dippon U, Kappler A (2012) Green rustformation during Fe(II) oxidation by the nitrate-reducing acidovorax sp. strain BoFeN1. Environ SciTechnol 46:1439–1446
Posth NR, Canfield DE, Kappler A (2014) Biogenic Fe(III) minerals: from formation to diagenesis andpreservation in the rock record. Earth Sci Rev 135:103–121
Poulton SW, Canfield DE (2005) Development of a sequential extraction procedure for iron: implicationsfor iron partitioning in continentally derived particulates. Chem Geol 214:209–221
Aquat Geochem
123

Revsbech NP (1989) An oxygen microelectrode with a guard cathode. Limnol Oceanogr 34:472–476Richardson DS (1974) The origin of iron-rich layers in sediments of the western equatorial Atlantic Ocean.
Dissertation. Columbia UniversityRichardson LL, Nealson KH (1998) Distributions of manganese, iron, and manganese-oxidizing bacteria in
Lake Superior sediments of different organic carbon content. J Great Lakes Res 15:123–132Robbins JA, Callender E (1975) Diagenesis of manganese in Lake Michigan sediments. Am J Sci
275:512–533Schaller T, Wehrli B (1997) Geochemical-focusing of manganese in lake sediments—an indicator of deep-
water oxygen conditions. Aquat Geochem 2:359–378Shapiro J, Edmondson WT, Alison DE (1971) Changes in the chemical composition of sediments of Lake
Washington 1958–1970. Limnol Oceanogr 16:437–452Sly PG, Thomas RL (1974) Review of geological research as it relates to an understanding of Great Lakes
limnology. J Fish Res Board Can 31:795–825Sommers M, Dollhopf M, Douglas S (2002) Freshwater ferromanganese stromatolites from Lake Vermilion,
Minnesota: microbial culturing and environmental scanning electron microscopy investigations.Geomicrobiol J 19:407–427
Sterner RW (2010) C:N:P stoichiometry in Lake Superior: freshwater sea as end member. Inland Waters1:29–46
Straub K, Buchholz-Cleven B (1998) Enumeration and detection of anaerobic ferrous iron-oxidizing, ni-trate-reducing bacteria from diverse European sediments. Appl Environ Microbiol 64:4846–5633
Stumm W, Morgan JJ (1995) Aquatic chemistry: chemical equilibria and rates in natural waters. John Wiley& Sons, New York
Tessier A, Campbell PGC, Bolsson M (1979) Sequential extraction procedure for the speciation of par-ticulate trace metals. Anal Chem 51:844–851
Torres NT, Och LM, Hauser PC, Furrer G, Brandl H, Vologina E, Sturm M, Burgmann H, Muller B (2014)Early diagenetic processes generate iron and manganese oxide layers in the sediments of Lake Baikal,Siberia. Environ Sci Process Impacts 16:879–889
Wilson TRS, Thomson J, Colley S, Hydes DJ, Higgs NC, Sørensen J (1985) Early organic diagenesis: thesignificance of progressive subsurface oxidation fronts in pelagic sediments. Geochim CosmochimActa 49:811–822
Wobus A, Bleul C, Maassen S, Scheerer C, Schuppler M, Jacobs E, Roske I (2003) Microbial diversity andfunctional characterisation of sediments from reservoirs of different trophic state. Microbiol Ecol46:331–347
Wu ML, Van Teeseling MCF, Willems MJR, Van Donselaar EG, Klingl A, Rachel R, Geerts WJC, JettenMSM, Strous M, Van Niftrik L (2012) Ultrastructure of the denitrifying methanotroph ‘‘Candidatusmethylomirabilis oxyfera,’’ a novel polygon-shaped bacterium. J Bacteriol 194:284–291
Zakharova YR, Parfenova VV, Granina LZ, Kravchenko OS, Zemskaya TI (2010) Distribution of iron andmanganese oxidizing bacteria in the bottom sediments of Lake Baikal. Inland Water Biol 3:313–321
Aquat Geochem
123




















