
Genotyping of CYP2D6 Polymorphisms by MALDI-TOF Mass Spectrometry in Sardinian People
10
Hindawi Publishing Corporation ISRN Genetics Volume 2013, Article ID 609797, 10 pages http://dx.doi.org/10.5402/2013/609797 Research Article Genotyping of CYP2D6 Polymorphisms by MALDI-TOF Mass Spectrometry in Sardinian People Matteo Falzoi, 1,2,3 Luigi Pira, 1 Paolo Lazzari, 1 and Luca Pani 2 1 PharmaNess S.c.a r.l., Parco Scientifico e Tecnologico della Sardegna, Edificio 5, Localit` a Piscina Manna, 09010 Pula, Italy 2 Unit` a Operativa di Cagliari, Parco Scientifico e Tecnologico della Sardegna, Istituto di Farmacologia Traslazionale del Consiglio Nazionale delle Ricerche CNR–IFT, Edificio 5, Localit` a Piscina Manna, 09010 Pula, Italy 3 Dipartimento di Scienze della Vita e dell’Ambiente, Laboratori di Genetica, Universit` a degli Studi di Cagliari, Via T. Fiorelli 1, 09126 Cagliari, Italy Correspondence should be addressed to Luigi Pira; [email protected] Received 7 March 2013; Accepted 4 April 2013 Academic Editors: B. Chen, M. A. Chiurillo, G. Giovambattista, and A. Yamamoto Copyright © 2013 Matteo Falzoi et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. e CYP2D6 enzyme is involved in the metabolism of many commonly prescribed drugs. e presence of CYP2D6 gene SNPs can alter CYP2D6 enzymatic activity with effects ranging considerably within a population. Objectives. In this study, we have developed a genotyping platform able to determine the alleles related to interindividual variability in the CYP2D6 gene. Design and Methods. We used a long PCR strategy coupled to MALDI-TOF mass spectrometry (Sequenom) to develop a SNPs genotyping method. Furthermore, an amplification allele specific was carried out to infer the correct allelic phase. Results. We tested the multiplex platform in 250 DNA Sardinian samples and found it to be 100% concordant with the sequencing results of our previous work. Conclusions. e MALDI-TOF-based multiplexing system allowed simultaneous and efficient genotyping of a set of CYP2D6 SNPs, evidencing its potential use in diagnostic test development to predict drug responses and clinical outcomes. 1. Introduction e presence of polymorphisms in the cytochrome P450 2D6 (CYP2D6) gene may modulate enzyme levels affecting individual responses to pharmacological treatment in drug level, response, and adverse reactions and includes individu- als with ultrarapid (UM), extensive (EM), intermediate (IM), and poor (PM) metabolizer status. Furthermore, the presence of multiple functional gene copies or the deletion of the entire gene results in increased or absent drug metabolism, respectively [1, 2]. Genotypic analysis to identify individual polymorphisms has become increasingly important during drug development and for selection of individualized ther- apies. For this reason, high-throughput technology for rapid, accurate, and efficient genotyping is needed. Several strate- gies and methods for SNPs genotyping have been developed. Techniques commonly used, such as PCR-RFLP, real-time PCR, and the TaqMan allele-specific assays from Applied Biosystems (CA, USA), are oſten laborious, and a restricted number of alleles can be simultaneously detected by these techniques. Conversely, the high-throughput oligonucleotide microarray technology, such as the Affymetrix/Roche (CA, USA) AmpliChip CYP450 GeneChip test, has the disad- vantage over other assays in that it cannot be customized by the user, and the benefits of this technology are oſten not compensable due to unfavourable costs [3]. In our previous work [4], we tried to create a complete genotyping platform for the simultaneous analysis of CYP3A4, CYP3A5, CYP2C9, CYP2C19, and CYP2D6 SNPs. e genotyping of the CYP2D6 gene was difficult due to its polymorphic nature, the presence of two flanking pseudogenes and copy number variants. To avoid false genotyping, resulting from nonspe- cific coamplification of the highly homologous pseudogenes, we developed the analysis of this gene in another way by using long PCR protocol coupled with single allele analysis and followed by sequencing [5]. In this work, we aimed to create a CYP2D6 medium-throughput SNPs screening platform using the Sequenom (CA, USA) matrix-assisted
Transcript of Genotyping of CYP2D6 Polymorphisms by MALDI-TOF Mass Spectrometry in Sardinian People

Hindawi Publishing CorporationISRN GeneticsVolume 2013 Article ID 609797 10 pageshttpdxdoiorg1054022013609797
Research ArticleGenotyping of CYP2D6 Polymorphisms byMALDI-TOF Mass Spectrometry in Sardinian People
Matteo Falzoi123 Luigi Pira1 Paolo Lazzari1 and Luca Pani2
1 PharmaNess Sca rl Parco Scientifico e Tecnologico della Sardegna Edificio 5 Localita Piscina Manna 09010 Pula Italy2 Unita Operativa di Cagliari Parco Scientifico e Tecnologico della Sardegna Istituto di Farmacologia Traslazionale del ConsiglioNazionale delle Ricerche CNRndashIFT Edificio 5 Localita Piscina Manna 09010 Pula Italy
3 Dipartimento di Scienze della Vita e dellrsquoAmbiente Laboratori di Genetica Universita degli Studi di Cagliari Via T Fiorelli 109126 Cagliari Italy
Correspondence should be addressed to Luigi Pira luigipirapharmanesscom
Received 7 March 2013 Accepted 4 April 2013
Academic Editors B Chen M A Chiurillo G Giovambattista and A Yamamoto
Copyright copy 2013 Matteo Falzoi et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The CYP2D6 enzyme is involved in the metabolism of many commonly prescribed drugs The presence of CYP2D6 gene SNPs canalter CYP2D6 enzymatic activity with effects ranging considerably within a populationObjectives In this study we have developeda genotyping platform able to determine the alleles related to interindividual variability in the CYP2D6 gene Design and MethodsWe used a long PCR strategy coupled to MALDI-TOF mass spectrometry (Sequenom) to develop a SNPs genotyping methodFurthermore an amplification allele specific was carried out to infer the correct allelic phase Results We tested the multiplexplatform in 250 DNA Sardinian samples and found it to be 100 concordant with the sequencing results of our previous workConclusionsTheMALDI-TOF-basedmultiplexing system allowed simultaneous and efficient genotyping of a set of CYP2D6 SNPsevidencing its potential use in diagnostic test development to predict drug responses and clinical outcomes
1 Introduction
The presence of polymorphisms in the cytochrome P4502D6 (CYP2D6) gene may modulate enzyme levels affectingindividual responses to pharmacological treatment in druglevel response and adverse reactions and includes individu-als with ultrarapid (UM) extensive (EM) intermediate (IM)and poor (PM)metabolizer status Furthermore the presenceof multiple functional gene copies or the deletion of theentire gene results in increased or absent drug metabolismrespectively [1 2] Genotypic analysis to identify individualpolymorphisms has become increasingly important duringdrug development and for selection of individualized ther-apies For this reason high-throughput technology for rapidaccurate and efficient genotyping is needed Several strate-gies and methods for SNPs genotyping have been developedTechniques commonly used such as PCR-RFLP real-timePCR and the TaqMan allele-specific assays from AppliedBiosystems (CA USA) are often laborious and a restricted
number of alleles can be simultaneously detected by thesetechniques Conversely the high-throughput oligonucleotidemicroarray technology such as the AffymetrixRoche (CAUSA) AmpliChip CYP450 GeneChip test has the disad-vantage over other assays in that it cannot be customizedby the user and the benefits of this technology are oftennot compensable due to unfavourable costs [3] In ourprevious work [4] we tried to create a complete genotypingplatform for the simultaneous analysis of CYP3A4 CYP3A5CYP2C9 CYP2C19 and CYP2D6 SNPs The genotyping ofthe CYP2D6 gene was difficult due to its polymorphic naturethe presence of two flanking pseudogenes and copy numbervariants To avoid false genotyping resulting from nonspe-cific coamplification of the highly homologous pseudogeneswe developed the analysis of this gene in another way byusing long PCR protocol coupled with single allele analysisand followed by sequencing [5] In this work we aimedto create a CYP2D6 medium-throughput SNPs screeningplatform using the Sequenom (CA USA) matrix-assisted
2 ISRN Genetics
Table 1 List of 69 analyzed SNPs and correlation to aminoacidchanges or transcriptional modificafications
CYP2D6 SNPs Variations-1584CgtG mdash-1426CgtT mdash-1235AgtG mdash-1000GgtA mdash-948CgtA mdash-750 -749delGA mdash-740CgtT mdash-678GgtA mdash19GgtA V7M31GgtA V11M77GgtA R26H82CgtT mdash100CgtT P34S124GgtA G42R137 138insT Frameshift214GgtC (lowast) mdash310GgtT mdash746CgtG mdash843TgtG mdash883GgtC Splicing defect957CgtT A85V974CgtA L91M984AgtG H94R997CgtG mdash1039CgtT mdash1513CgtT mdash1659GgtA V136I1661GgtC mdash1704CgtG Q151E1707delT Frameshift1724CgtT mdash1749AgtG N166D1757CgtT mdash1758GgtA G169R1758GgtT G169X1846GgtA Splicing defect1863 1864insTTTCGCCCC 174 175insFRP1869TgtC mdash1943GgtA R201H1979TgtC mdash2291GgtA mdash2483GgtT A237S2539 2542delAACT Frameshift2549delA Frameshift2575CgtA mdash2587 2590delGACT Frameshift2615 2617delAAG K281del2850CgtT R296C
Table 1 Continued
CYP2D6 SNPs Variations2853AgtC I297L2935AgtC H324P2939GgtA mdash2988GgtA Splicing defect3176CgtT mdash3183GgtA V338M3198CgtG R343G3277TgtC I369T3288GgtA mdash3384AgtC mdash3582AgtG mdash3584GgtA mdash3790CgtT mdash3828GgtA mdash3853GgtA E410K3877GgtA E418K3948TgtG mdash4115CgtT mdash4155CgtT (∘) H478Y4180GgtC S486T4401CgtT mdash4481GgtA mdashIn MALDI-TOF MS analysis (lowast) 214GgtC SNP was used to discriminategene conversion to CYP2D7P in Intron 1 and (∘) 4155CgtT SNP was usedto discriminate gene conversion to CYP2D7P in Exon 9 A = alanine R =arginine N = asparagine D = aspartic acid C = cysteine E = glutamic acideQ = glutamine G = glycine H = histidine I = isoleucine L = leucine K= lysine M = methionine F = phenylalanine P = proline S = serine T =threonineW= tryptophan Y= tyrosineV= valine In bold characters newlydiscovered SNPs in [5 17 18 21]
laser desorptionionization (MALDI) time-of-flight (TOF)mass spectrometry (MS) [6] a widely used technology thatis proving to be a competitive analysis method in SNPsgenotyping Advantages of MALDI-TOF MS over previouslydescribed methods include the option for medium-high-throughput automated analysis of SNPs the relative ease ofsetup formultiplex assays and the reduced costs per genotype[3]
2 Methods and Materials
SNPs and Sequences Selection The 69 polymorphisms ana-lyzed in this study were selected using principal SNP webdatabases such as the Human CYP Allele NomenclatureCommittee [7] and the NCBI Single Nucleotide Polymor-phism dbSNP [8] websites Selection criteria dependedmainly on the pharmacogenetic effects described for everyallele in the Caucasian population [5 7 9ndash16] Not all selectedSNPs are involved in aminoacidic or transcriptional variation(Table 1) Some of these are silent or promoter leader trailerand intronic changes and inserted in our study because theyare essential for the reconstruction of haplotype phases TheSNPsrsquo recombination allowed the reconstruction of 66 among
ISRN Genetics 3
Table 2 List of primers used inCYP2D6 long PCRprotocolsThe 51015840 10-mer tagwas added to PCRprimers in order to improve PCR efficiency
Nucleotides position Name 51015840 10ndashmer tag 51015840-Sequence-31015840 DirectionFrom -1780 to -1758 P-1780 [5] ACGTTGGATG GTCCTCCTGTCCTCAGTGGAT ForwardFrom -1584 to -1559 P-1584 WT [5] ACGTTGGATG CAGCCTGGACAACTTGGAAGAAGCC ForwardFrom -1584 to -1559 P-1584 MUT [5] ACGTTGGATG CAGCCTGGACAACTTGGAAGAAGCG ForwardFrom 4706 to 4728 2D6ndashR [19] ACGTTGGATG ACTGAGCCCTGGGAGGTAGGTA Reverse
allelic variants and subvariants List of alleles is summarizedin Table 5
DNASamples In this study we reanalyzed the genomicDNAsstudied in [5] Sardinian DNA samples were gently furnishedby Professor Francesco Cucca INN-CNR Cagliari DirectorAll participating individuals provided informed consent togenetic test To genotype these samples we implemented aCYP2D6 Genotyping Platform based on MALDI-TOF MSThis way we would be able to compare our genotyping resultswith a previous sequencing analysis in [5]
21 CYP2D6 Genotyping Platform
211 Long Primary PCR Selective amplification of theCYP2D6 genewas carried outmodifying a long PCR protocolimplemented in our previous work [5] Forward primer (P-1780 Table 2) was designed in a highly nonhomologousCYP2D6CYP2D7PCYP2D8P 51015840 untranslated region [5]The reverse primer used was 2D6-R (Table 2) as previouslydescribed [19] A 51015840 10-mer tag (51015840-ACGTTGGATG-31015840) wasadded to both PCR primers in order to improve PCRefficiency PCR reactions were performed in a final volume of5 120583L using the QIAGEN (Hilden Germany) LongRange PCRKit protocol [20] with the following minor modifications20 ng genomicDNA 400 120583Mof each PCR primer (MetabionMartinsried Germany) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 500 120583M Invitrogen (CA USA) 21015840-
deoxynucleoside-51015840-triphosphate (dNTP) Set PCR GradeThe PCR conditions were as follows initial denaturation at93∘C for 3min 35 cycles at 93∘C for 30 s 61∘C for 30 s and68∘C for 6min
212 SAP Dephosphorylation To neutralize unincorporateddNTPs after amplification reactions 03U of ShrimpAlkalinePhosphatase (SAP) (Sequenom) [26 27] was used SAPcleaved a phosphate from the unincorporated dNTPs con-verting them to 21015840-deoxynucleoside-51015840-diphosphate (dNDP)and rendering them unavailable for following reactionsDephosphorylation conditions were as follows 37∘C for20min and 85∘C for 5min
213 Assay Design SNP-specific unextended minisequenc-ing primers (UEPs) and multiplexed UEPs assays weredesigned using both the SequenomMassARRAYassay designversion 31 and the RealSNP assay database [28] A sequenceof 400 base pairs (bp) flanking each selected SNP was down-loaded from the corresponding genomic sequence stored in
the public NCBI Single Nucleotide Polymorphism dbSNPdatabase [8] or the Ensembl Genome Browser [29] and wasanalyzed by Vector NTI Suite Software version 55 (InforMaxOxford UK) Combination of the UEPs into multiplex assayswas supported by these software applications to allow theoptimization of several different parameters for example toavoid the risk of primer-primer interactions and hairpin-loop formations GC content molecular mass range andannealing temperatures To achieve the highest possiblemultiplexing levels we tested many primer combinationsleading us to the final assay design consisting in a total of69 SNPs successfully assembled in five medium-plex assays(13- 14- 13- 14- and 15-plex) (Table 3) For some SNPsSequenom MassARRAY assay design and RealSNP assaydatabase could not design SNP-specific UEPs because of thepresence of proximal SNPsOther SNPswere excludedduringplatform validation because of UEPs cross-hybridization inhighly homologous PCR template regions or for the presenceof primer dimers which created false allele But the highnumber of SNPs inserted in our analysis has allowed us toinfer the correct alleles for each DNA analysis List of SNPsnot included is summarized in Table 4
214 iPLEX Reactions iPLEX reactions were carried outfollowing the Sequenom standard lowmedium-plex protocol[6 26 27] with minor modifications Because of the lengthof the primary PCR fragment and the high GC contentwe increased the denaturation time at 94∘C from 5 to30 sec and the annealing temperature from 52 to 56∘CAn iPLEX reaction cocktail was added to the amplificationproducts and thermocycled to process the iPLEX reac-tion which involved the enzymatic addition of one of thefour mass-modified nucleotides 2101584031015840-dideoxynucleoside-51015840-triphosphate (ddNTP) into the polymorphic site During theiPLEX reaction each primer was extended by one of theddNTPswhich terminated the extension of primers thus pro-ducing allele-specific extension products of different massesThe iPLEX reaction cocktail included 0222X iPLEX bufferplus 05X iPLEX termination mix 05X iPLEX enzyme andUEPs which were divided into four mass groups accordingto the position of their respectivemass peaks to use the wholespectrum (final concentrations of 08ndash20 120583M) In the iPLEXtermination mix all four ddNTPs are present at the sameconcentration The reactions were performed using a twocycling loop program initial denaturation at 94∘C for 30 sfollowed by 40 cycles of 94∘C for 30 s 56∘C for 5 s and 80∘Cfor 5 s This annealing and extension procedure was repeatedfour times (to give a total of 200 short cycles) followedby a final extension step of 3min at 72∘C After desalting
4 ISRN Genetics
Table3Sequ
encesa
ndmolecular
weightsof
unextend
ed(U
EP)a
ndextend
ed(EP)
iPLE
Xprim
ers
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(1)13-PL
EX3877GgtA
CCTT
CCGCT
TCCA
CCCC
F4978
CCTT
CCGCT
TCCA
CCCC
A5249
ACC
TTCC
GCT
TCCA
CCCC
G5265
G3183GgtA
CCGCA
CCTG
CCCT
ATCA
R5051
CCGCA
CCTG
CCCT
ATCA
C5299
GCC
GCA
CCTG
CCCT
ATCA
T5378
A2615
2617
delAAG
GGCA
GCC
ACTC
TCAC
CTR5091
GGCA
GCC
ACTC
TCAC
CTC
5339
del
GGCA
GCC
ACTC
TCAC
CTT
5418
AGA
2483GgtT
GCG
TAGGAC
CTTG
CCAG
R5211
GCG
TAGGAC
CTTG
CCAG
C5459
GGCG
TAGGAC
CTTG
CCAG
A5483
T3853GgtA
GAAG
CGGAAG
GGCT
TCT
R5275
GAAG
CGGAAG
GGCT
TCTC
5523
GGAAG
CGGAAG
GGCT
TCTT
5603
A3277Tgt
CTG
CAGCG
CTTT
GGGGAC
AF5556
TGCA
GCG
CTTT
GGGGAC
AC5803
CTG
CAGCG
CTTT
GGGGAC
AT5883
T4180GgtC
AGCT
CATA
GGGGGAT
GGG
R5645
AGCT
CATA
GGGGGAT
GGGC
5892
GAG
CTCA
TAGGGGGAT
GGGG
5932
C2939GgtA
TGGGCT
CACG
CTGCA
CATC
R5766
TGGGCT
CACG
CTGCA
CATC
C6013
GTG
GGCT
CACG
CTGCA
CATC
T6093
A3288GgtA
GAT
GTC
ATAT
GGGTC
ACAC
R5828
GAT
GTC
ATAT
GGGTC
ACAC
C6075
GGAT
GTC
ATAT
GGGTC
ACAC
T6155
A3176Cgt
TGGCC
GTG
TCCA
ACAG
GAG
ATF6167
GGCC
GTG
TCCA
ACAG
GAG
ATC
6414
CGGCC
GTG
TCCA
ACAG
GAG
ATT
6494
T2850Cgt
TAG
CTTC
AAT
GAT
GAG
AAC
CTG
F6454
AGCT
TCAAT
GAT
GAG
AAC
CTGC
6701
CAG
CTTC
AAT
GAT
GAG
AAC
CTGT
6781
T2935AgtC
AGTG
CTCA
CGCT
GCA
CATC
CGGA
R7010
AGTG
CTCA
CGCT
GCA
CATC
CGGAG
7297
CAG
TGCT
CACG
CTGCA
CATC
CGGAT
7337
A1757Cgt
TTT
GTG
CCGCC
TTCG
CCAAC
CACT
CF7201
TTGTG
CCGCC
TTCG
CCAAC
CACT
CC7448
CTT
GTG
CCGCC
TTCG
CCAAC
CACT
CT7528
T(2)14-PL
EX1661GgtC
CAGAG
GCG
CTTC
TCCG
TF5162
CAGAG
GCG
CTTC
TCCG
TC5410
CCA
GAG
GCG
CTTC
TCCG
TG5450
G1869Tgt
CAC
GGCT
TTGTC
CAAG
AGR5210
ACGGCT
TTGTC
CAAG
AGA
5482
TAC
GGCT
TTGTC
CAAG
AGG
5498
C1846
GgtA
GGGGCG
AAAG
GGGCG
TCR5342
GGGGCG
AAAG
GGGCG
TCC
5588
GGGGGCG
AAAG
GGGCG
TCT
5668
A214GgtC
TGGAG
GGCG
GCA
GAG
GT
F5357
TGGAG
GGCG
GCA
GAG
GTC
5604
CTG
GAG
GGCG
GCA
GAG
GTG
5644
G19GgtA
CTGGGGCT
AGAAG
CACT
GF5565
CTGGGGCT
AGAAG
CACT
GA
5836
ACT
GGGGCT
AGAAG
CACT
GG
5852
G1707delT
GCG
GCC
TCCT
CGGTC
ACCC
R5702
GCG
GCC
TCCT
CGGTC
ACCC
C5949
del
GCG
GCC
TCCT
CGGTC
ACCC
A5973
T746CgtG
GCG
ACCC
AGCC
TCCT
GAT
CR5710
GCG
ACCC
AGCC
TCCT
GAT
CC5957
GGCG
ACCC
AGCC
TCCT
GAT
CG5997
C997CgtG
GAAC
AGGCG
GGCG
GTC
GGC
R5920
GAAC
AGGCG
GGCG
GTC
GGCC
6167
GGAAC
AGGCG
GGCG
GTC
GGCG
6207
C100CgtT
AAT
GCT
GGGCT
GCA
CGCT
ACF6118
AAT
GCT
GGGCT
GCA
CGCT
ACC
6365
CAAT
GCT
GGGCT
GCA
CGCT
ACT
6445
T1979Tgt
CGGAC
AGCC
CGAC
TCCT
CCTT
CR6303
GGAC
AGCC
CGAC
TCCT
CCTT
CA6574
TGGAC
AGCC
CGAC
TCCT
CCTT
CG6590
C-948Cgt
ACA
GGCT
GGGGCA
AGGGCC
TTC
F6488
CAGGCT
GGGGCA
AGGGCC
TTCC
6735
CCA
GGCT
GGGGCA
AGGGCC
TTCA
6759
A1749AgtG
GCC
CATC
ACCC
ACCG
GAG
TGGT
R6681
GCC
CATC
ACCC
ACCG
GAG
TGGTC
6929
GGCC
CATC
ACCC
ACCG
GAG
TGGTT
7008
A77GgtA
GTA
GCG
TGCA
GCC
CAGCG
TTGG
R6792
GTA
GCG
TGCA
GCC
CAGCG
TTGGC7040
GGTA
GCG
TGCA
GCC
CAGCG
TTGGT
7120
A1513Cgt
TAG
CTGGAC
AGAG
CCAG
GGAC
TGF6835
AGCT
GGAC
AGAG
CCAG
GGAC
TGC7082
CAG
CTGGAC
AGAG
CCAG
GGAC
TGT7162
T(3)13-PL
EX984AgtG
GCG
AGTG
TCCT
CGCC
GR4874
GCG
AGTG
TCCT
CGCC
GC
5121
GGCG
AGTG
TCCT
CGCC
GT
5201
A1863
1864
ins
TTTC
GCC
CCGAC
GCC
CCTT
TCGCC
CCF5043
GAC
GCC
CCTT
TCGCC
CCA
5315
ndashGAC
GCC
CCTT
TCGCC
CCT
5370
TTTC
GCC
CC1039Cgt
TAC
CCAG
ATCC
TGGGTT
TF5161
ACCC
AGAT
CCTG
GGTT
TC5409
CAC
CCAG
ATCC
TGGGTT
TT5489
T1943GgtA
CCAG
CAGCC
TGAG
GAAG
R5229
CCAG
CAGCC
TGAG
GAAG
C5477
GCC
AGCA
GCC
TGAG
GAAG
T5557
A974CgtA
CGGCC
GTG
CGCG
AGGCG
F5253
CGGCC
GTG
CGCG
AGGCG
C5501
CCG
GCC
GTG
CGCG
AGGCG
A5525
A1704
CgtG
GGCA
AGAAG
TCGCT
GGAG
F5614
GGCA
AGAAG
TCGCT
GGAG
C5861
CGGCA
AGAAG
TCGCT
GGAG
G5901
G310GgtT
TAAAT
GCC
CTTC
TCCA
GGA
R5748
TAAAT
GCC
CTTC
TCCA
GGAC
5995
GTA
AAT
GCC
CTTC
TCCA
GGAA
6019
T957CgtT
GTC
GTG
CTCA
ATGGGCT
GG
F5876
GTC
GTG
CTCA
ATGGGCT
GGC
6123
CGTC
GTG
CTCA
ATGGGCT
GGT
6203
T883GgtC
TCCC
CGAAG
CGGCG
CCGCA
AR6073
TCCC
CGAAG
CGGCG
CCGCA
AC6320
GTC
CCCG
AAG
CGGCG
CCGCA
AG6360
C1659GgtA
CGCG
AGCA
GAG
GCG
CTTC
TCC
F6408
CGCG
AGCA
GAG
GCG
CTTC
TCCA
6679
ACG
CGAG
CAGAG
GCG
CTTC
TCCG
6695
G31GgtA
AGCA
GGAAG
ATGGCC
ACTA
TCA
R6777
AGCA
GGAAG
ATGGCC
ACTA
TCAC
7024
GAG
CAGGAAG
ATGGCC
ACTA
TCAT
7104
A1724Cgt
TGCG
AAG
GCG
GCA
CAAAG
GCA
GGC
R7158
GCG
AAG
GCG
GCA
CAAAG
GCA
GGCA7429
TGCG
AAG
GCG
GCA
CAAAG
GCA
GGCG7445
C124GgtA
TCAC
ATGCA
GCA
GGTT
GCC
CAGCC
R7299
TCAC
ATGCA
GCA
GGTT
GCC
CAGCC
C7546
GTC
ACAT
GCA
GCA
GGTT
GCC
CAGCC
T7626
A
ISRN Genetics 5
Table3Con
tinued
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(4)14-PL
EX3198Cgt
GAC
CCAT
CTCT
GGTC
GCC
R5082
ACCC
ATCT
CTGGTC
GCC
C5330
GAC
CCAT
CTCT
GGTC
GCC
G5370
C2549delA
TGGGTC
CCAG
GTC
ATCC
R5162
TGGGTC
CCAG
GTC
ATCC
G5450
del
TGGGTC
CCAG
GTC
ATCC
T5490
A3582AgtG
GAAT
GTT
GGAG
GAC
CCA
F5259
GAAT
GTT
GGAG
GAC
CCAA
5531
AGAAT
GTT
GGAG
GAC
CCAG
5547
G2539
2542
delAAC
TAC
AGCT
GGAT
GAG
CTGCT
F5540
ACAG
CTGGAT
GAG
CTGCT
A5811
AAC
TAC
AGCT
GGAT
GAG
CTGCT
G5827
del
3790Cgt
TCC
ACTC
TCAC
CCTG
CATC
TF5620
CCAC
TCTC
ACCC
TGCA
TCTC
5867
CCC
ACTC
TCAC
CCTG
CATC
TT5947
T4115Cgt
TAC
CTCC
CTGCT
GCA
GCA
CTT
F5989
ACCT
CCCT
GCT
GCA
GCA
CTTC
6236
CAC
CTCC
CTGCT
GCA
GCA
CTTT
6316
T44
81GgtA
GAAT
CTGAC
TGCC
CAGAT
TGF6117
GAAT
CTGAC
TGCC
CAGAT
TGA
6388
AGAAT
CTGAC
TGCC
CAGAT
TGG
6404
G3828GgtA
ACTC
ATCA
CCAAC
CTGTC
ATC
F6270
ACTC
ATCA
CCAAC
CTGTC
ATCA
6541
AAC
TCAT
CACC
AAC
CTGTC
ATCG
6557
G3384AgtC
CATG
CTGGGGCT
ATCA
CCAG
GR6447
CATG
CTGGGGCT
ATCA
CCAG
GG
6734
CCA
TGCT
GGGGCT
ATCA
CCAG
GT
6774
A2853AgtC
GAG
AAC
AGGTC
AGCC
ACCA
CTA
R6722
GAG
AAC
AGGTC
AGCC
ACCA
CTAG
7010
CGAG
AAC
AGGTC
AGCC
ACCA
CTAT
7050
A2988GgtA
CATG
TGCC
CCCG
CCTG
TACC
CTT
R6888
CATG
TGCC
CCCG
CCTG
TACC
CTTC
7135
GCA
TGTG
CCCC
CGCC
TGTA
CCCT
TT7215
A3584GgtA
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
F7098
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
A7369
AGTC
AGAAT
GTT
GGAG
GAC
CCAAC
G7385
G44
01Cgt
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
R7546
TAAC
TGAC
ATCT
GCT
CAGCC
TCAAC
A7817
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
G7833
C2587
2590
delGAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GA
F7846
ATCA
GCT
CAGCC
CCCC
CGAG
ACCT
GAC8093
GAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GAG8133
del
(5)15-PL
EX3948Tgt
GTC
TCAG
CAGGTG
CCTG
F4873
TCTC
AGCA
GGTG
CCTG
G5160
GTC
TCAG
CAGGTG
CCTT
5200
T-1235AgtG
GCA
CCAC
CCAG
CCTA
ATR5084
GCA
CCAC
CCAG
CCTA
ATC
5332
GGCA
CCAC
CCAG
CCTA
ATT
5411
A137138insT
GAAG
TCCA
CATG
CAGCA
R5188
GAAG
TCCA
CATG
CAGCA
A5460
TGAAG
TCCA
CATG
CAGCA
G5476
mdash4155Cgt
TCA
GCC
CCGGCC
CAGCC
ACF5376
CAGCC
CCGGCC
CAGCC
ACC
5623
CCA
GCC
CCGGCC
CAGCC
ACT
5703
T82Cgt
TGTG
TAGCG
TGCA
GCC
CAGC
R5830
GTG
TAGCG
TGCA
GCC
CAGCA
6101
TGTG
TAGCG
TGCA
GCC
CAGCG
6117
C-1584CgtG
CTGGAC
AAC
TTGGAAG
AAC
CF6135
CTGGAC
AAC
TTGGAAG
AAC
CC6382
CCT
GGAC
AAC
TTGGAAG
AAC
CG6422
G2575Cgt
AGAC
CTGGGAC
CCAG
CCCA
GCC
F6362
GAC
CTGGGAC
CCAG
CCCA
GCC
C6609
CGAC
CTGGGAC
CCAG
CCCA
GCC
A6633
A843TgtG
ACTA
GGAC
CTGTA
GTC
TGGGG
F6502
ACTA
GGAC
CTGTA
GTC
TGGGGG
6789
GAC
TAGGAC
CTGTA
GTC
TGGGGT
6829
T-740
CgtT
ACAG
ACTC
ACAC
TGAC
ACTT
AGR6672
ACAG
ACTC
ACAC
TGAC
ACTT
AGA
6944
TAC
AGAC
TCAC
ACTG
ACAC
TTAG
G6960
C-678GgtA
CTTT
GTG
TGGGTG
ATTT
TCTG
CF6769
CTTT
GTG
TGGGTG
ATTT
TCTG
CA7041
ACT
TTGTG
TGGGTG
ATTT
TCTG
CG7057
G-750
-749
delGA
TGTG
ACTG
GTG
TGTG
TGAG
AGA
F6902
TGTG
ACTG
GTG
TGTG
TGAG
AGAA
7173
del
TGTG
ACTG
GTG
TGTG
TGAG
AGAG
7189
GA
2291GgtA
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CR7204
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CC7451
GCA
CTCG
CCAAG
TGCC
AGCC
TCCA
CT7531
A-1426CgtT
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
R7286
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
A7533
TGTG
TGCC
ACCA
CGTC
TAGCT
TTTT
G7573
C
1758GgtTgt
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCF7490
TTGTG
CCGCC
TTCG
CCAAC
CACT
CCA7761
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCG7777
GTT
GTG
CCGCC
TTCG
CCAAC
CACT
CCT7817
T-100
0GgtA
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGT
R7635
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGTC7882
GAC
ATCC
TCCC
GGGCT
GCC
TGAG
GGTT
7962
AUDU
nextendedminindashsequ
encing
prim
erDire
ctionFFo
rwardUEP
directionR
ReverseU
EPdirectionUMU
nextendedminindashsequ
encing
prim
ersM
assEL
ME
xtendedprim
erLo
wer
MassEH
ME
xtended
prim
erHigherM
assEL
CEx
tend
edprim
erlower
Massn
ucleotideC
alledEH
CEx
tend
edprim
erhigh
erMassn
ucleotideC
alledAllmolecular
weightm
assesa
reexpressedin
Dalton(D
a)
6 ISRN Genetics
Table 4 List of SNPs excluded in assay design validation andcorrelation to transcriptional variations
SNPs excluded Variations Reason-1770GgtA mdash Cross-hybridization-1298GgtA mdash Primer dimers-1253AgtG mdash Primer dimers
-1237 -1236insAA mdash Proximal SNP -1235AgtGPrimer dimers
1973 1974insG Frameshift Proximal SNP 1979TgtC1976GgtA mdash Proximal SNP 1979TgtC1978CgtT mdash Proximal SNP 1979TgtC2097AgtG mdash Primer dimers2470TgtC mdash Cross hybridization4042GgtA mdash Primer dimers
by addition of 6mg clean resin (Sequenom) each 384-wellsample was diluted with 16 120583L of sterilized H
2Odd
Multiplex PCR reactions SAP dephosphorylation andiPLEX reactionswere performedusingThermo-Fast 384 PCRPlates (ABgene Epsom UK) and a DNA Engine Tetrad 2Peltier Thermal Cycler (Bio-Rad CA USA) All pipettingsteps were performed using the automatic station MatrixPlateMate 2 times 2 (Sequenom)
215 MALDI-TOF MS Measurement An aliquot rangingfrom 15 to 20 nL of each iPLEX reaction product was loadedin a 384-spot SpectroChip (Sequenom) using the MassAr-ray Nanodispenser (Samsung Seoul Repubic of Korea)SpectroChip analysis was performed by MassARRAY Com-pact System (Sequenom) After laser desorptionionizationautomated spectra acquisition analysis was performed andinterpreted using Sequenom MassARRAY RT version 33software Examples of multiplex mass spectrum and clusterplot distributions are shown in Figures 1 and 2
22 CYP2D6 Single Allele Genotyping Following directionof our previous work [5] we decided to apply the singleallele protocol creating a single allele genotyping methodMALDI-TOFMS based For each sample a double long PCRreaction was carried out using P-1584 WT or P-1584 MUT[5] (Table 2) as forward primers The reverse primer was2D6-R [19] for both PCR reactions In this way we wereable to directly determine a direct and correct chromosomephase in samples presenting with a heterozygous status forminus1584GgtC SNP PCR reactions were performed in a finalvolume of 5 120583L using 20 ng genomic DNA 400 120583M of eachPCR primer (Metabion) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 800 120583MInvitrogen dNTP set PCR grade
The PCR conditions were as follows initial denaturation at93∘C for 3min 10 cycles at 93∘C for 30 s 67∘C for 30 s and68∘C for 3min 25 cycles at 93∘C for 30 s 65∘C for 30 s and68∘C for 6min SAP dephosphorylation iPLEX ReactionsandMALDI-TOFMSmeasurement were performed without
Table 5CYP2D6 allele frequencies in 250 healthy Sardinian peopleTotal chromosomes number = 500 Human cytochrome P450 AlleleNomenclature Committee [7] served as reference for variant alleleand correlated enzymatic activity
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast1A 148 296 EMlowast1B 5 10 EMlowast1D 4 08 EMlowast1E mdash mdash EMlowast2A 75 150 EMlowast2B mdash mdash EMlowast2D mdash mdash EMlowast2E mdash mdash EMlowast2F mdash mdash EMlowast2G mdash mdash EMlowast2K mdash mdash EMlowast2L 11 22 EMlowast2M 8 16 EMSH3 7 14 Not knownSH4 1 02 Not knownlowast3A mdash mdash PMlowast3B 11 22 PMlowast4A 84 168 PMlowast4B mdash mdash PMlowast4D mdash mdash PMlowast4K mdash mdash PMlowast4L mdash mdash PMlowast4M mdash mdash PMlowast4N mdash mdash PMlowast5 5 10 PMlowast6A 1 02 PMlowast6C mdash mdash PMlowast6D mdash mdash PMlowast7 mdash mdash PMlowast8 mdash mdash PMlowast9 1 02 IMlowast10A mdash mdash IMlowast10B 27 54 IMlowast11 mdash mdash PMlowast12 mdash mdash PMlowast14A mdash mdash PMlowast14B mdash mdash IMlowast15 3 06 PMlowast17 mdash mdash IMlowast19 mdash mdash PMlowast20 1 02 PMlowast22 mdash mdash Not knownlowast23 mdash mdash Not known
ISRN Genetics 7
Table 5 Continued
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast24 mdash mdash Not knownlowast25 mdash mdash Not knownlowast26 mdash mdash Not knownlowast27 mdash mdash EMlowast28 4 08 Not knownlowast29 mdash mdash IMlowast30 mdash mdash Not knownlowast31 mdash mdash PMlowast32 mdash mdash Not knownlowast33 mdash mdash EMlowast35A 5 10 EMlowast36 mdash mdash IMlowast37 mdash mdash Not knownlowast38 mdash mdash PMlowast39 mdash mdash EMlowast41 46 92 IMSH1 41 82 Not knownSH2 2 04 Not knownlowast43 mdash mdash Not knownlowast58 mdash mdash Not knownlowast59 mdash mdash IMlowast64 mdash mdash Not knownlowast65 mdash mdash Not knownlowast1xN 4 08 UMlowast2xN 6 12 UMSH1234 = Sardinian haplotype 1234 [5 22ndash25] CYP2D6lowast5lowast1xN andlowast2xN alleles were evaluated byThe CYP2D6Applied Biosystems CNVAssayin [5]
modifications as indicated in paragraph 1 ldquoCYP2D6 Geno-typing Platformrdquo In multiplexed assay 5 minus1584GgtC UEP(Table 3) was excluded Examples of cluster distribution fornovel 3176CgtT and 3948TgtG SNPs [5 17 18] are shown inFigure 3
3 Results and Discussion
A CYP2D6 screening assay was developed using theSequenom MassARRAY platform to simultaneously identifythe most frequent and some rare CYP2D6 Caucasian allelesWe have modified the basic Sequenom iPLEX assay and useda new primary PCR strategy based on the amplification ofthe entire gene [5] coupled with multiplex primer extensionreactionsThis strategy avoids false genotyping which wouldresult in nonspecific coamplification of the homologouspseudogenes CYP2D7P and CYP2D8P and secondly itreduces the number of PCR primers used to select regionscontaining the targeted polymorphisms Multiplexing wasperformed for 69 SNPs which represents 66 of the mostfrequent and some rarer variants and subvariants reported
5000 5500 6000 6500 7000 7500 8000
0123456789
10
Mass
Inte
nsity
lowast lowast TCG A
Figure 1 An example of 15ndashplex mass spectrum is shown (Well 5)List of SNPs investigated in this plex is shown in Table 3 The figureshows two examples highlighted in different colours Unextendprimer (UEP) peak is marked by an asterisk and dotted arrow whilethe solid arrows indicate the presence of the two different allelesDotted vertical lines represent UEPs and extended primers (EPs)expected masses In blue an AG heterozygous genotype example isshown lowast minus1235AgtG UEP expected mass = 5084 Dalton (Da) Gminus1235AgtGEP expectedmass = 5332Da Aminus1235AgtGEP expectedmass = 5411 Da In red a CC wild type homozygous genotypeexample is shown lowast 82CgtT UEP expected mass = 5830Da T82CgtT not EP expected mass = 6101Da C 82CgtT EP expectedmass = 6117Da
to date in the Caucasian population [5 7 9ndash16] and knownto be responsible for absent reduced or extensive metabolicactivity (Tables 1 and 5) Due to the high possibility ofrecombination it was possible to insert African African-American (11986211988411987521198636lowast2119871 lowast2 119872 lowast4119873 lowast6 4 and lowast6 5) andAsian (11986211988411987521198636lowast3119860 lowast4119861 lowast4119871 lowast14 lowast36 and lowast39) variantsin the study A sample of 250 unrelated healthy Sardinianindividuals analyzed in [5] was submitted to MALDI-TOFMS genotyping for these 69 CYP2D6 SNPs An example ofmultiplex mass spectrum is shown in Figure 1 In Figure 2(a)an example of the cluster plot distribution for the 1661GgtCSNP is shown Spectrum peak intensities were not correctlybalanced in some heterozygous samples (Figures 2(c) and2(d)) which appeared as outliers in the cluster distribution(Figure 2(a) point 120575 and 120576) The CYP2D6 Applied BiosystemsCopy Number Variation (CNV) Assay used to analyze theseDNAs in our previous work [5] detected the presence ofduplications or multiplications in 100 of the analyzedsamples presenting this kind of distribution Furthermore insamples presenting a heterozygous status for minus1584GgtC SNPwe applied long PCR single allele analysis [5] to MALDI-TOF MS screening assays and inferred a direct haplotypephase To verify if our CYP2D6 platform works correctly wecompared genotyping results and haplotype phase elaboratedin the two screening platforms with our results found in [5]A consensus of 100 was found for all samples (Table 5)Moreover for all samples presenting the 214GgtC SNP inhomo- or heterozygous status in MALDI-TOF MS analysisit was possible to verify in our previous sequencing analysis
8 ISRN Genetics
120572
120573
120574
120577
120575
120576
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0010203040506070809
1
(a)
0
2
4
6
8
10
12
14
16
18
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(b)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(c)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(d)
Figure 2 In (a) a Cluster plot for 1661GgtC SNP is shown Homozygous wild type GG samples are displayed in (120572) homozygous mutate CCsamples in (120573) and heterozygous GC samples in (120574) in (120575) and (120576) are displayed outlier samples resulting in gene copy number variation [5](120577) negative control (H2Odd) In (b) aMass Spectra for the detection of 1661GgtC in a heterozygous GC sample displayed in (a) position (120574)In (c) a Mass Spectra for the detection of 1661GgtC in the outlier sample displayed in (a) position (120575) and resulting in 11986211988411987521198636lowast1 times 119873 allele[5] In (d) Mass Spectra for the detection of 1661G gt C in an outlier sample displayed in (a) position (120576) and resulting in 11986211988411987521198636lowast2 times 119873allele [5]
[5] the presence of all SNPs correlated to gene conversionto CYP2D7P in Intron 1 (Figure 4) which indicated thereliability of MALDI-TOF analysis for this variation
4 Conclusions
Differences in drug responses could be due to genetic fac-tors Knowledge of individual genetic profiling is clinicallyimportant and provides benefits for future medical care bypredicting the drug response or developing DNA-based testsSubstantial interindividual variability in response to specifictherapies might be caused by the presence of polymorphismsin genes encoding components of drugmetabolismpathwayssuch as theCYP450 family genes Polymorphisms inCYP2D6gene have been thoroughly investigated including their asso-ciations with the incidence of adverse reactions In this study
we have developed a reliablemedium-throughput genotypingplatformusing the SequenomMassARRAY system to providea simultaneous screening of the most relevant CYP2D6 genevariants in several hundreds of subjects MALDI-TOF MS-based analyses have the potential to become a useful approachin clinical diagnoses as they are very flexible and applicableto individualized genetic therapies The screening platformdeveloped in this study coupled with The CYP2D6 AppliedBiosystems CNVAssay provides a robust alternative tomanycurrently available CYP450-genotyping approaches and aimsto increase the number of responders and to limit theincidences of adverse events Moreover by modifying longPCR Forward primer we have implemented a rapid strategyto infer the phase by direct analysis using MALDI-TOS MSmultiplex assays Concordance between data found in thiswork and our direct genomic DNA sequencing analysis donein [5] reliably validated ourMALDI-TOFMS-based platform
ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml

2 ISRN Genetics
Table 1 List of 69 analyzed SNPs and correlation to aminoacidchanges or transcriptional modificafications
CYP2D6 SNPs Variations-1584CgtG mdash-1426CgtT mdash-1235AgtG mdash-1000GgtA mdash-948CgtA mdash-750 -749delGA mdash-740CgtT mdash-678GgtA mdash19GgtA V7M31GgtA V11M77GgtA R26H82CgtT mdash100CgtT P34S124GgtA G42R137 138insT Frameshift214GgtC (lowast) mdash310GgtT mdash746CgtG mdash843TgtG mdash883GgtC Splicing defect957CgtT A85V974CgtA L91M984AgtG H94R997CgtG mdash1039CgtT mdash1513CgtT mdash1659GgtA V136I1661GgtC mdash1704CgtG Q151E1707delT Frameshift1724CgtT mdash1749AgtG N166D1757CgtT mdash1758GgtA G169R1758GgtT G169X1846GgtA Splicing defect1863 1864insTTTCGCCCC 174 175insFRP1869TgtC mdash1943GgtA R201H1979TgtC mdash2291GgtA mdash2483GgtT A237S2539 2542delAACT Frameshift2549delA Frameshift2575CgtA mdash2587 2590delGACT Frameshift2615 2617delAAG K281del2850CgtT R296C
Table 1 Continued
CYP2D6 SNPs Variations2853AgtC I297L2935AgtC H324P2939GgtA mdash2988GgtA Splicing defect3176CgtT mdash3183GgtA V338M3198CgtG R343G3277TgtC I369T3288GgtA mdash3384AgtC mdash3582AgtG mdash3584GgtA mdash3790CgtT mdash3828GgtA mdash3853GgtA E410K3877GgtA E418K3948TgtG mdash4115CgtT mdash4155CgtT (∘) H478Y4180GgtC S486T4401CgtT mdash4481GgtA mdashIn MALDI-TOF MS analysis (lowast) 214GgtC SNP was used to discriminategene conversion to CYP2D7P in Intron 1 and (∘) 4155CgtT SNP was usedto discriminate gene conversion to CYP2D7P in Exon 9 A = alanine R =arginine N = asparagine D = aspartic acid C = cysteine E = glutamic acideQ = glutamine G = glycine H = histidine I = isoleucine L = leucine K= lysine M = methionine F = phenylalanine P = proline S = serine T =threonineW= tryptophan Y= tyrosineV= valine In bold characters newlydiscovered SNPs in [5 17 18 21]
laser desorptionionization (MALDI) time-of-flight (TOF)mass spectrometry (MS) [6] a widely used technology thatis proving to be a competitive analysis method in SNPsgenotyping Advantages of MALDI-TOF MS over previouslydescribed methods include the option for medium-high-throughput automated analysis of SNPs the relative ease ofsetup formultiplex assays and the reduced costs per genotype[3]
2 Methods and Materials
SNPs and Sequences Selection The 69 polymorphisms ana-lyzed in this study were selected using principal SNP webdatabases such as the Human CYP Allele NomenclatureCommittee [7] and the NCBI Single Nucleotide Polymor-phism dbSNP [8] websites Selection criteria dependedmainly on the pharmacogenetic effects described for everyallele in the Caucasian population [5 7 9ndash16] Not all selectedSNPs are involved in aminoacidic or transcriptional variation(Table 1) Some of these are silent or promoter leader trailerand intronic changes and inserted in our study because theyare essential for the reconstruction of haplotype phases TheSNPsrsquo recombination allowed the reconstruction of 66 among
ISRN Genetics 3
Table 2 List of primers used inCYP2D6 long PCRprotocolsThe 51015840 10-mer tagwas added to PCRprimers in order to improve PCR efficiency
Nucleotides position Name 51015840 10ndashmer tag 51015840-Sequence-31015840 DirectionFrom -1780 to -1758 P-1780 [5] ACGTTGGATG GTCCTCCTGTCCTCAGTGGAT ForwardFrom -1584 to -1559 P-1584 WT [5] ACGTTGGATG CAGCCTGGACAACTTGGAAGAAGCC ForwardFrom -1584 to -1559 P-1584 MUT [5] ACGTTGGATG CAGCCTGGACAACTTGGAAGAAGCG ForwardFrom 4706 to 4728 2D6ndashR [19] ACGTTGGATG ACTGAGCCCTGGGAGGTAGGTA Reverse
allelic variants and subvariants List of alleles is summarizedin Table 5
DNASamples In this study we reanalyzed the genomicDNAsstudied in [5] Sardinian DNA samples were gently furnishedby Professor Francesco Cucca INN-CNR Cagliari DirectorAll participating individuals provided informed consent togenetic test To genotype these samples we implemented aCYP2D6 Genotyping Platform based on MALDI-TOF MSThis way we would be able to compare our genotyping resultswith a previous sequencing analysis in [5]
21 CYP2D6 Genotyping Platform
211 Long Primary PCR Selective amplification of theCYP2D6 genewas carried outmodifying a long PCR protocolimplemented in our previous work [5] Forward primer (P-1780 Table 2) was designed in a highly nonhomologousCYP2D6CYP2D7PCYP2D8P 51015840 untranslated region [5]The reverse primer used was 2D6-R (Table 2) as previouslydescribed [19] A 51015840 10-mer tag (51015840-ACGTTGGATG-31015840) wasadded to both PCR primers in order to improve PCRefficiency PCR reactions were performed in a final volume of5 120583L using the QIAGEN (Hilden Germany) LongRange PCRKit protocol [20] with the following minor modifications20 ng genomicDNA 400 120583Mof each PCR primer (MetabionMartinsried Germany) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 500 120583M Invitrogen (CA USA) 21015840-
deoxynucleoside-51015840-triphosphate (dNTP) Set PCR GradeThe PCR conditions were as follows initial denaturation at93∘C for 3min 35 cycles at 93∘C for 30 s 61∘C for 30 s and68∘C for 6min
212 SAP Dephosphorylation To neutralize unincorporateddNTPs after amplification reactions 03U of ShrimpAlkalinePhosphatase (SAP) (Sequenom) [26 27] was used SAPcleaved a phosphate from the unincorporated dNTPs con-verting them to 21015840-deoxynucleoside-51015840-diphosphate (dNDP)and rendering them unavailable for following reactionsDephosphorylation conditions were as follows 37∘C for20min and 85∘C for 5min
213 Assay Design SNP-specific unextended minisequenc-ing primers (UEPs) and multiplexed UEPs assays weredesigned using both the SequenomMassARRAYassay designversion 31 and the RealSNP assay database [28] A sequenceof 400 base pairs (bp) flanking each selected SNP was down-loaded from the corresponding genomic sequence stored in
the public NCBI Single Nucleotide Polymorphism dbSNPdatabase [8] or the Ensembl Genome Browser [29] and wasanalyzed by Vector NTI Suite Software version 55 (InforMaxOxford UK) Combination of the UEPs into multiplex assayswas supported by these software applications to allow theoptimization of several different parameters for example toavoid the risk of primer-primer interactions and hairpin-loop formations GC content molecular mass range andannealing temperatures To achieve the highest possiblemultiplexing levels we tested many primer combinationsleading us to the final assay design consisting in a total of69 SNPs successfully assembled in five medium-plex assays(13- 14- 13- 14- and 15-plex) (Table 3) For some SNPsSequenom MassARRAY assay design and RealSNP assaydatabase could not design SNP-specific UEPs because of thepresence of proximal SNPsOther SNPswere excludedduringplatform validation because of UEPs cross-hybridization inhighly homologous PCR template regions or for the presenceof primer dimers which created false allele But the highnumber of SNPs inserted in our analysis has allowed us toinfer the correct alleles for each DNA analysis List of SNPsnot included is summarized in Table 4
214 iPLEX Reactions iPLEX reactions were carried outfollowing the Sequenom standard lowmedium-plex protocol[6 26 27] with minor modifications Because of the lengthof the primary PCR fragment and the high GC contentwe increased the denaturation time at 94∘C from 5 to30 sec and the annealing temperature from 52 to 56∘CAn iPLEX reaction cocktail was added to the amplificationproducts and thermocycled to process the iPLEX reac-tion which involved the enzymatic addition of one of thefour mass-modified nucleotides 2101584031015840-dideoxynucleoside-51015840-triphosphate (ddNTP) into the polymorphic site During theiPLEX reaction each primer was extended by one of theddNTPswhich terminated the extension of primers thus pro-ducing allele-specific extension products of different massesThe iPLEX reaction cocktail included 0222X iPLEX bufferplus 05X iPLEX termination mix 05X iPLEX enzyme andUEPs which were divided into four mass groups accordingto the position of their respectivemass peaks to use the wholespectrum (final concentrations of 08ndash20 120583M) In the iPLEXtermination mix all four ddNTPs are present at the sameconcentration The reactions were performed using a twocycling loop program initial denaturation at 94∘C for 30 sfollowed by 40 cycles of 94∘C for 30 s 56∘C for 5 s and 80∘Cfor 5 s This annealing and extension procedure was repeatedfour times (to give a total of 200 short cycles) followedby a final extension step of 3min at 72∘C After desalting
4 ISRN Genetics
Table3Sequ
encesa
ndmolecular
weightsof
unextend
ed(U
EP)a
ndextend
ed(EP)
iPLE
Xprim
ers
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(1)13-PL
EX3877GgtA
CCTT
CCGCT
TCCA
CCCC
F4978
CCTT
CCGCT
TCCA
CCCC
A5249
ACC
TTCC
GCT
TCCA
CCCC
G5265
G3183GgtA
CCGCA
CCTG
CCCT
ATCA
R5051
CCGCA
CCTG
CCCT
ATCA
C5299
GCC
GCA
CCTG
CCCT
ATCA
T5378
A2615
2617
delAAG
GGCA
GCC
ACTC
TCAC
CTR5091
GGCA
GCC
ACTC
TCAC
CTC
5339
del
GGCA
GCC
ACTC
TCAC
CTT
5418
AGA
2483GgtT
GCG
TAGGAC
CTTG
CCAG
R5211
GCG
TAGGAC
CTTG
CCAG
C5459
GGCG
TAGGAC
CTTG
CCAG
A5483
T3853GgtA
GAAG
CGGAAG
GGCT
TCT
R5275
GAAG
CGGAAG
GGCT
TCTC
5523
GGAAG
CGGAAG
GGCT
TCTT
5603
A3277Tgt
CTG
CAGCG
CTTT
GGGGAC
AF5556
TGCA
GCG
CTTT
GGGGAC
AC5803
CTG
CAGCG
CTTT
GGGGAC
AT5883
T4180GgtC
AGCT
CATA
GGGGGAT
GGG
R5645
AGCT
CATA
GGGGGAT
GGGC
5892
GAG
CTCA
TAGGGGGAT
GGGG
5932
C2939GgtA
TGGGCT
CACG
CTGCA
CATC
R5766
TGGGCT
CACG
CTGCA
CATC
C6013
GTG
GGCT
CACG
CTGCA
CATC
T6093
A3288GgtA
GAT
GTC
ATAT
GGGTC
ACAC
R5828
GAT
GTC
ATAT
GGGTC
ACAC
C6075
GGAT
GTC
ATAT
GGGTC
ACAC
T6155
A3176Cgt
TGGCC
GTG
TCCA
ACAG
GAG
ATF6167
GGCC
GTG
TCCA
ACAG
GAG
ATC
6414
CGGCC
GTG
TCCA
ACAG
GAG
ATT
6494
T2850Cgt
TAG
CTTC
AAT
GAT
GAG
AAC
CTG
F6454
AGCT
TCAAT
GAT
GAG
AAC
CTGC
6701
CAG
CTTC
AAT
GAT
GAG
AAC
CTGT
6781
T2935AgtC
AGTG
CTCA
CGCT
GCA
CATC
CGGA
R7010
AGTG
CTCA
CGCT
GCA
CATC
CGGAG
7297
CAG
TGCT
CACG
CTGCA
CATC
CGGAT
7337
A1757Cgt
TTT
GTG
CCGCC
TTCG
CCAAC
CACT
CF7201
TTGTG
CCGCC
TTCG
CCAAC
CACT
CC7448
CTT
GTG
CCGCC
TTCG
CCAAC
CACT
CT7528
T(2)14-PL
EX1661GgtC
CAGAG
GCG
CTTC
TCCG
TF5162
CAGAG
GCG
CTTC
TCCG
TC5410
CCA
GAG
GCG
CTTC
TCCG
TG5450
G1869Tgt
CAC
GGCT
TTGTC
CAAG
AGR5210
ACGGCT
TTGTC
CAAG
AGA
5482
TAC
GGCT
TTGTC
CAAG
AGG
5498
C1846
GgtA
GGGGCG
AAAG
GGGCG
TCR5342
GGGGCG
AAAG
GGGCG
TCC
5588
GGGGGCG
AAAG
GGGCG
TCT
5668
A214GgtC
TGGAG
GGCG
GCA
GAG
GT
F5357
TGGAG
GGCG
GCA
GAG
GTC
5604
CTG
GAG
GGCG
GCA
GAG
GTG
5644
G19GgtA
CTGGGGCT
AGAAG
CACT
GF5565
CTGGGGCT
AGAAG
CACT
GA
5836
ACT
GGGGCT
AGAAG
CACT
GG
5852
G1707delT
GCG
GCC
TCCT
CGGTC
ACCC
R5702
GCG
GCC
TCCT
CGGTC
ACCC
C5949
del
GCG
GCC
TCCT
CGGTC
ACCC
A5973
T746CgtG
GCG
ACCC
AGCC
TCCT
GAT
CR5710
GCG
ACCC
AGCC
TCCT
GAT
CC5957
GGCG
ACCC
AGCC
TCCT
GAT
CG5997
C997CgtG
GAAC
AGGCG
GGCG
GTC
GGC
R5920
GAAC
AGGCG
GGCG
GTC
GGCC
6167
GGAAC
AGGCG
GGCG
GTC
GGCG
6207
C100CgtT
AAT
GCT
GGGCT
GCA
CGCT
ACF6118
AAT
GCT
GGGCT
GCA
CGCT
ACC
6365
CAAT
GCT
GGGCT
GCA
CGCT
ACT
6445
T1979Tgt
CGGAC
AGCC
CGAC
TCCT
CCTT
CR6303
GGAC
AGCC
CGAC
TCCT
CCTT
CA6574
TGGAC
AGCC
CGAC
TCCT
CCTT
CG6590
C-948Cgt
ACA
GGCT
GGGGCA
AGGGCC
TTC
F6488
CAGGCT
GGGGCA
AGGGCC
TTCC
6735
CCA
GGCT
GGGGCA
AGGGCC
TTCA
6759
A1749AgtG
GCC
CATC
ACCC
ACCG
GAG
TGGT
R6681
GCC
CATC
ACCC
ACCG
GAG
TGGTC
6929
GGCC
CATC
ACCC
ACCG
GAG
TGGTT
7008
A77GgtA
GTA
GCG
TGCA
GCC
CAGCG
TTGG
R6792
GTA
GCG
TGCA
GCC
CAGCG
TTGGC7040
GGTA
GCG
TGCA
GCC
CAGCG
TTGGT
7120
A1513Cgt
TAG
CTGGAC
AGAG
CCAG
GGAC
TGF6835
AGCT
GGAC
AGAG
CCAG
GGAC
TGC7082
CAG
CTGGAC
AGAG
CCAG
GGAC
TGT7162
T(3)13-PL
EX984AgtG
GCG
AGTG
TCCT
CGCC
GR4874
GCG
AGTG
TCCT
CGCC
GC
5121
GGCG
AGTG
TCCT
CGCC
GT
5201
A1863
1864
ins
TTTC
GCC
CCGAC
GCC
CCTT
TCGCC
CCF5043
GAC
GCC
CCTT
TCGCC
CCA
5315
ndashGAC
GCC
CCTT
TCGCC
CCT
5370
TTTC
GCC
CC1039Cgt
TAC
CCAG
ATCC
TGGGTT
TF5161
ACCC
AGAT
CCTG
GGTT
TC5409
CAC
CCAG
ATCC
TGGGTT
TT5489
T1943GgtA
CCAG
CAGCC
TGAG
GAAG
R5229
CCAG
CAGCC
TGAG
GAAG
C5477
GCC
AGCA
GCC
TGAG
GAAG
T5557
A974CgtA
CGGCC
GTG
CGCG
AGGCG
F5253
CGGCC
GTG
CGCG
AGGCG
C5501
CCG
GCC
GTG
CGCG
AGGCG
A5525
A1704
CgtG
GGCA
AGAAG
TCGCT
GGAG
F5614
GGCA
AGAAG
TCGCT
GGAG
C5861
CGGCA
AGAAG
TCGCT
GGAG
G5901
G310GgtT
TAAAT
GCC
CTTC
TCCA
GGA
R5748
TAAAT
GCC
CTTC
TCCA
GGAC
5995
GTA
AAT
GCC
CTTC
TCCA
GGAA
6019
T957CgtT
GTC
GTG
CTCA
ATGGGCT
GG
F5876
GTC
GTG
CTCA
ATGGGCT
GGC
6123
CGTC
GTG
CTCA
ATGGGCT
GGT
6203
T883GgtC
TCCC
CGAAG
CGGCG
CCGCA
AR6073
TCCC
CGAAG
CGGCG
CCGCA
AC6320
GTC
CCCG
AAG
CGGCG
CCGCA
AG6360
C1659GgtA
CGCG
AGCA
GAG
GCG
CTTC
TCC
F6408
CGCG
AGCA
GAG
GCG
CTTC
TCCA
6679
ACG
CGAG
CAGAG
GCG
CTTC
TCCG
6695
G31GgtA
AGCA
GGAAG
ATGGCC
ACTA
TCA
R6777
AGCA
GGAAG
ATGGCC
ACTA
TCAC
7024
GAG
CAGGAAG
ATGGCC
ACTA
TCAT
7104
A1724Cgt
TGCG
AAG
GCG
GCA
CAAAG
GCA
GGC
R7158
GCG
AAG
GCG
GCA
CAAAG
GCA
GGCA7429
TGCG
AAG
GCG
GCA
CAAAG
GCA
GGCG7445
C124GgtA
TCAC
ATGCA
GCA
GGTT
GCC
CAGCC
R7299
TCAC
ATGCA
GCA
GGTT
GCC
CAGCC
C7546
GTC
ACAT
GCA
GCA
GGTT
GCC
CAGCC
T7626
A
ISRN Genetics 5
Table3Con
tinued
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(4)14-PL
EX3198Cgt
GAC
CCAT
CTCT
GGTC
GCC
R5082
ACCC
ATCT
CTGGTC
GCC
C5330
GAC
CCAT
CTCT
GGTC
GCC
G5370
C2549delA
TGGGTC
CCAG
GTC
ATCC
R5162
TGGGTC
CCAG
GTC
ATCC
G5450
del
TGGGTC
CCAG
GTC
ATCC
T5490
A3582AgtG
GAAT
GTT
GGAG
GAC
CCA
F5259
GAAT
GTT
GGAG
GAC
CCAA
5531
AGAAT
GTT
GGAG
GAC
CCAG
5547
G2539
2542
delAAC
TAC
AGCT
GGAT
GAG
CTGCT
F5540
ACAG
CTGGAT
GAG
CTGCT
A5811
AAC
TAC
AGCT
GGAT
GAG
CTGCT
G5827
del
3790Cgt
TCC
ACTC
TCAC
CCTG
CATC
TF5620
CCAC
TCTC
ACCC
TGCA
TCTC
5867
CCC
ACTC
TCAC
CCTG
CATC
TT5947
T4115Cgt
TAC
CTCC
CTGCT
GCA
GCA
CTT
F5989
ACCT
CCCT
GCT
GCA
GCA
CTTC
6236
CAC
CTCC
CTGCT
GCA
GCA
CTTT
6316
T44
81GgtA
GAAT
CTGAC
TGCC
CAGAT
TGF6117
GAAT
CTGAC
TGCC
CAGAT
TGA
6388
AGAAT
CTGAC
TGCC
CAGAT
TGG
6404
G3828GgtA
ACTC
ATCA
CCAAC
CTGTC
ATC
F6270
ACTC
ATCA
CCAAC
CTGTC
ATCA
6541
AAC
TCAT
CACC
AAC
CTGTC
ATCG
6557
G3384AgtC
CATG
CTGGGGCT
ATCA
CCAG
GR6447
CATG
CTGGGGCT
ATCA
CCAG
GG
6734
CCA
TGCT
GGGGCT
ATCA
CCAG
GT
6774
A2853AgtC
GAG
AAC
AGGTC
AGCC
ACCA
CTA
R6722
GAG
AAC
AGGTC
AGCC
ACCA
CTAG
7010
CGAG
AAC
AGGTC
AGCC
ACCA
CTAT
7050
A2988GgtA
CATG
TGCC
CCCG
CCTG
TACC
CTT
R6888
CATG
TGCC
CCCG
CCTG
TACC
CTTC
7135
GCA
TGTG
CCCC
CGCC
TGTA
CCCT
TT7215
A3584GgtA
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
F7098
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
A7369
AGTC
AGAAT
GTT
GGAG
GAC
CCAAC
G7385
G44
01Cgt
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
R7546
TAAC
TGAC
ATCT
GCT
CAGCC
TCAAC
A7817
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
G7833
C2587
2590
delGAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GA
F7846
ATCA
GCT
CAGCC
CCCC
CGAG
ACCT
GAC8093
GAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GAG8133
del
(5)15-PL
EX3948Tgt
GTC
TCAG
CAGGTG
CCTG
F4873
TCTC
AGCA
GGTG
CCTG
G5160
GTC
TCAG
CAGGTG
CCTT
5200
T-1235AgtG
GCA
CCAC
CCAG
CCTA
ATR5084
GCA
CCAC
CCAG
CCTA
ATC
5332
GGCA
CCAC
CCAG
CCTA
ATT
5411
A137138insT
GAAG
TCCA
CATG
CAGCA
R5188
GAAG
TCCA
CATG
CAGCA
A5460
TGAAG
TCCA
CATG
CAGCA
G5476
mdash4155Cgt
TCA
GCC
CCGGCC
CAGCC
ACF5376
CAGCC
CCGGCC
CAGCC
ACC
5623
CCA
GCC
CCGGCC
CAGCC
ACT
5703
T82Cgt
TGTG
TAGCG
TGCA
GCC
CAGC
R5830
GTG
TAGCG
TGCA
GCC
CAGCA
6101
TGTG
TAGCG
TGCA
GCC
CAGCG
6117
C-1584CgtG
CTGGAC
AAC
TTGGAAG
AAC
CF6135
CTGGAC
AAC
TTGGAAG
AAC
CC6382
CCT
GGAC
AAC
TTGGAAG
AAC
CG6422
G2575Cgt
AGAC
CTGGGAC
CCAG
CCCA
GCC
F6362
GAC
CTGGGAC
CCAG
CCCA
GCC
C6609
CGAC
CTGGGAC
CCAG
CCCA
GCC
A6633
A843TgtG
ACTA
GGAC
CTGTA
GTC
TGGGG
F6502
ACTA
GGAC
CTGTA
GTC
TGGGGG
6789
GAC
TAGGAC
CTGTA
GTC
TGGGGT
6829
T-740
CgtT
ACAG
ACTC
ACAC
TGAC
ACTT
AGR6672
ACAG
ACTC
ACAC
TGAC
ACTT
AGA
6944
TAC
AGAC
TCAC
ACTG
ACAC
TTAG
G6960
C-678GgtA
CTTT
GTG
TGGGTG
ATTT
TCTG
CF6769
CTTT
GTG
TGGGTG
ATTT
TCTG
CA7041
ACT
TTGTG
TGGGTG
ATTT
TCTG
CG7057
G-750
-749
delGA
TGTG
ACTG
GTG
TGTG
TGAG
AGA
F6902
TGTG
ACTG
GTG
TGTG
TGAG
AGAA
7173
del
TGTG
ACTG
GTG
TGTG
TGAG
AGAG
7189
GA
2291GgtA
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CR7204
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CC7451
GCA
CTCG
CCAAG
TGCC
AGCC
TCCA
CT7531
A-1426CgtT
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
R7286
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
A7533
TGTG
TGCC
ACCA
CGTC
TAGCT
TTTT
G7573
C
1758GgtTgt
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCF7490
TTGTG
CCGCC
TTCG
CCAAC
CACT
CCA7761
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCG7777
GTT
GTG
CCGCC
TTCG
CCAAC
CACT
CCT7817
T-100
0GgtA
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGT
R7635
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGTC7882
GAC
ATCC
TCCC
GGGCT
GCC
TGAG
GGTT
7962
AUDU
nextendedminindashsequ
encing
prim
erDire
ctionFFo
rwardUEP
directionR
ReverseU
EPdirectionUMU
nextendedminindashsequ
encing
prim
ersM
assEL
ME
xtendedprim
erLo
wer
MassEH
ME
xtended
prim
erHigherM
assEL
CEx
tend
edprim
erlower
Massn
ucleotideC
alledEH
CEx
tend
edprim
erhigh
erMassn
ucleotideC
alledAllmolecular
weightm
assesa
reexpressedin
Dalton(D
a)
6 ISRN Genetics
Table 4 List of SNPs excluded in assay design validation andcorrelation to transcriptional variations
SNPs excluded Variations Reason-1770GgtA mdash Cross-hybridization-1298GgtA mdash Primer dimers-1253AgtG mdash Primer dimers
-1237 -1236insAA mdash Proximal SNP -1235AgtGPrimer dimers
1973 1974insG Frameshift Proximal SNP 1979TgtC1976GgtA mdash Proximal SNP 1979TgtC1978CgtT mdash Proximal SNP 1979TgtC2097AgtG mdash Primer dimers2470TgtC mdash Cross hybridization4042GgtA mdash Primer dimers
by addition of 6mg clean resin (Sequenom) each 384-wellsample was diluted with 16 120583L of sterilized H
2Odd
Multiplex PCR reactions SAP dephosphorylation andiPLEX reactionswere performedusingThermo-Fast 384 PCRPlates (ABgene Epsom UK) and a DNA Engine Tetrad 2Peltier Thermal Cycler (Bio-Rad CA USA) All pipettingsteps were performed using the automatic station MatrixPlateMate 2 times 2 (Sequenom)
215 MALDI-TOF MS Measurement An aliquot rangingfrom 15 to 20 nL of each iPLEX reaction product was loadedin a 384-spot SpectroChip (Sequenom) using the MassAr-ray Nanodispenser (Samsung Seoul Repubic of Korea)SpectroChip analysis was performed by MassARRAY Com-pact System (Sequenom) After laser desorptionionizationautomated spectra acquisition analysis was performed andinterpreted using Sequenom MassARRAY RT version 33software Examples of multiplex mass spectrum and clusterplot distributions are shown in Figures 1 and 2
22 CYP2D6 Single Allele Genotyping Following directionof our previous work [5] we decided to apply the singleallele protocol creating a single allele genotyping methodMALDI-TOFMS based For each sample a double long PCRreaction was carried out using P-1584 WT or P-1584 MUT[5] (Table 2) as forward primers The reverse primer was2D6-R [19] for both PCR reactions In this way we wereable to directly determine a direct and correct chromosomephase in samples presenting with a heterozygous status forminus1584GgtC SNP PCR reactions were performed in a finalvolume of 5 120583L using 20 ng genomic DNA 400 120583M of eachPCR primer (Metabion) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 800 120583MInvitrogen dNTP set PCR grade
The PCR conditions were as follows initial denaturation at93∘C for 3min 10 cycles at 93∘C for 30 s 67∘C for 30 s and68∘C for 3min 25 cycles at 93∘C for 30 s 65∘C for 30 s and68∘C for 6min SAP dephosphorylation iPLEX ReactionsandMALDI-TOFMSmeasurement were performed without
Table 5CYP2D6 allele frequencies in 250 healthy Sardinian peopleTotal chromosomes number = 500 Human cytochrome P450 AlleleNomenclature Committee [7] served as reference for variant alleleand correlated enzymatic activity
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast1A 148 296 EMlowast1B 5 10 EMlowast1D 4 08 EMlowast1E mdash mdash EMlowast2A 75 150 EMlowast2B mdash mdash EMlowast2D mdash mdash EMlowast2E mdash mdash EMlowast2F mdash mdash EMlowast2G mdash mdash EMlowast2K mdash mdash EMlowast2L 11 22 EMlowast2M 8 16 EMSH3 7 14 Not knownSH4 1 02 Not knownlowast3A mdash mdash PMlowast3B 11 22 PMlowast4A 84 168 PMlowast4B mdash mdash PMlowast4D mdash mdash PMlowast4K mdash mdash PMlowast4L mdash mdash PMlowast4M mdash mdash PMlowast4N mdash mdash PMlowast5 5 10 PMlowast6A 1 02 PMlowast6C mdash mdash PMlowast6D mdash mdash PMlowast7 mdash mdash PMlowast8 mdash mdash PMlowast9 1 02 IMlowast10A mdash mdash IMlowast10B 27 54 IMlowast11 mdash mdash PMlowast12 mdash mdash PMlowast14A mdash mdash PMlowast14B mdash mdash IMlowast15 3 06 PMlowast17 mdash mdash IMlowast19 mdash mdash PMlowast20 1 02 PMlowast22 mdash mdash Not knownlowast23 mdash mdash Not known
ISRN Genetics 7
Table 5 Continued
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast24 mdash mdash Not knownlowast25 mdash mdash Not knownlowast26 mdash mdash Not knownlowast27 mdash mdash EMlowast28 4 08 Not knownlowast29 mdash mdash IMlowast30 mdash mdash Not knownlowast31 mdash mdash PMlowast32 mdash mdash Not knownlowast33 mdash mdash EMlowast35A 5 10 EMlowast36 mdash mdash IMlowast37 mdash mdash Not knownlowast38 mdash mdash PMlowast39 mdash mdash EMlowast41 46 92 IMSH1 41 82 Not knownSH2 2 04 Not knownlowast43 mdash mdash Not knownlowast58 mdash mdash Not knownlowast59 mdash mdash IMlowast64 mdash mdash Not knownlowast65 mdash mdash Not knownlowast1xN 4 08 UMlowast2xN 6 12 UMSH1234 = Sardinian haplotype 1234 [5 22ndash25] CYP2D6lowast5lowast1xN andlowast2xN alleles were evaluated byThe CYP2D6Applied Biosystems CNVAssayin [5]
modifications as indicated in paragraph 1 ldquoCYP2D6 Geno-typing Platformrdquo In multiplexed assay 5 minus1584GgtC UEP(Table 3) was excluded Examples of cluster distribution fornovel 3176CgtT and 3948TgtG SNPs [5 17 18] are shown inFigure 3
3 Results and Discussion
A CYP2D6 screening assay was developed using theSequenom MassARRAY platform to simultaneously identifythe most frequent and some rare CYP2D6 Caucasian allelesWe have modified the basic Sequenom iPLEX assay and useda new primary PCR strategy based on the amplification ofthe entire gene [5] coupled with multiplex primer extensionreactionsThis strategy avoids false genotyping which wouldresult in nonspecific coamplification of the homologouspseudogenes CYP2D7P and CYP2D8P and secondly itreduces the number of PCR primers used to select regionscontaining the targeted polymorphisms Multiplexing wasperformed for 69 SNPs which represents 66 of the mostfrequent and some rarer variants and subvariants reported
5000 5500 6000 6500 7000 7500 8000
0123456789
10
Mass
Inte
nsity
lowast lowast TCG A
Figure 1 An example of 15ndashplex mass spectrum is shown (Well 5)List of SNPs investigated in this plex is shown in Table 3 The figureshows two examples highlighted in different colours Unextendprimer (UEP) peak is marked by an asterisk and dotted arrow whilethe solid arrows indicate the presence of the two different allelesDotted vertical lines represent UEPs and extended primers (EPs)expected masses In blue an AG heterozygous genotype example isshown lowast minus1235AgtG UEP expected mass = 5084 Dalton (Da) Gminus1235AgtGEP expectedmass = 5332Da Aminus1235AgtGEP expectedmass = 5411 Da In red a CC wild type homozygous genotypeexample is shown lowast 82CgtT UEP expected mass = 5830Da T82CgtT not EP expected mass = 6101Da C 82CgtT EP expectedmass = 6117Da
to date in the Caucasian population [5 7 9ndash16] and knownto be responsible for absent reduced or extensive metabolicactivity (Tables 1 and 5) Due to the high possibility ofrecombination it was possible to insert African African-American (11986211988411987521198636lowast2119871 lowast2 119872 lowast4119873 lowast6 4 and lowast6 5) andAsian (11986211988411987521198636lowast3119860 lowast4119861 lowast4119871 lowast14 lowast36 and lowast39) variantsin the study A sample of 250 unrelated healthy Sardinianindividuals analyzed in [5] was submitted to MALDI-TOFMS genotyping for these 69 CYP2D6 SNPs An example ofmultiplex mass spectrum is shown in Figure 1 In Figure 2(a)an example of the cluster plot distribution for the 1661GgtCSNP is shown Spectrum peak intensities were not correctlybalanced in some heterozygous samples (Figures 2(c) and2(d)) which appeared as outliers in the cluster distribution(Figure 2(a) point 120575 and 120576) The CYP2D6 Applied BiosystemsCopy Number Variation (CNV) Assay used to analyze theseDNAs in our previous work [5] detected the presence ofduplications or multiplications in 100 of the analyzedsamples presenting this kind of distribution Furthermore insamples presenting a heterozygous status for minus1584GgtC SNPwe applied long PCR single allele analysis [5] to MALDI-TOF MS screening assays and inferred a direct haplotypephase To verify if our CYP2D6 platform works correctly wecompared genotyping results and haplotype phase elaboratedin the two screening platforms with our results found in [5]A consensus of 100 was found for all samples (Table 5)Moreover for all samples presenting the 214GgtC SNP inhomo- or heterozygous status in MALDI-TOF MS analysisit was possible to verify in our previous sequencing analysis
8 ISRN Genetics
120572
120573
120574
120577
120575
120576
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0010203040506070809
1
(a)
0
2
4
6
8
10
12
14
16
18
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(b)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(c)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(d)
Figure 2 In (a) a Cluster plot for 1661GgtC SNP is shown Homozygous wild type GG samples are displayed in (120572) homozygous mutate CCsamples in (120573) and heterozygous GC samples in (120574) in (120575) and (120576) are displayed outlier samples resulting in gene copy number variation [5](120577) negative control (H2Odd) In (b) aMass Spectra for the detection of 1661GgtC in a heterozygous GC sample displayed in (a) position (120574)In (c) a Mass Spectra for the detection of 1661GgtC in the outlier sample displayed in (a) position (120575) and resulting in 11986211988411987521198636lowast1 times 119873 allele[5] In (d) Mass Spectra for the detection of 1661G gt C in an outlier sample displayed in (a) position (120576) and resulting in 11986211988411987521198636lowast2 times 119873allele [5]
[5] the presence of all SNPs correlated to gene conversionto CYP2D7P in Intron 1 (Figure 4) which indicated thereliability of MALDI-TOF analysis for this variation
4 Conclusions
Differences in drug responses could be due to genetic fac-tors Knowledge of individual genetic profiling is clinicallyimportant and provides benefits for future medical care bypredicting the drug response or developing DNA-based testsSubstantial interindividual variability in response to specifictherapies might be caused by the presence of polymorphismsin genes encoding components of drugmetabolismpathwayssuch as theCYP450 family genes Polymorphisms inCYP2D6gene have been thoroughly investigated including their asso-ciations with the incidence of adverse reactions In this study
we have developed a reliablemedium-throughput genotypingplatformusing the SequenomMassARRAY system to providea simultaneous screening of the most relevant CYP2D6 genevariants in several hundreds of subjects MALDI-TOF MS-based analyses have the potential to become a useful approachin clinical diagnoses as they are very flexible and applicableto individualized genetic therapies The screening platformdeveloped in this study coupled with The CYP2D6 AppliedBiosystems CNVAssay provides a robust alternative tomanycurrently available CYP450-genotyping approaches and aimsto increase the number of responders and to limit theincidences of adverse events Moreover by modifying longPCR Forward primer we have implemented a rapid strategyto infer the phase by direct analysis using MALDI-TOS MSmultiplex assays Concordance between data found in thiswork and our direct genomic DNA sequencing analysis donein [5] reliably validated ourMALDI-TOFMS-based platform
ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml

ISRN Genetics 3
Table 2 List of primers used inCYP2D6 long PCRprotocolsThe 51015840 10-mer tagwas added to PCRprimers in order to improve PCR efficiency
Nucleotides position Name 51015840 10ndashmer tag 51015840-Sequence-31015840 DirectionFrom -1780 to -1758 P-1780 [5] ACGTTGGATG GTCCTCCTGTCCTCAGTGGAT ForwardFrom -1584 to -1559 P-1584 WT [5] ACGTTGGATG CAGCCTGGACAACTTGGAAGAAGCC ForwardFrom -1584 to -1559 P-1584 MUT [5] ACGTTGGATG CAGCCTGGACAACTTGGAAGAAGCG ForwardFrom 4706 to 4728 2D6ndashR [19] ACGTTGGATG ACTGAGCCCTGGGAGGTAGGTA Reverse
allelic variants and subvariants List of alleles is summarizedin Table 5
DNASamples In this study we reanalyzed the genomicDNAsstudied in [5] Sardinian DNA samples were gently furnishedby Professor Francesco Cucca INN-CNR Cagliari DirectorAll participating individuals provided informed consent togenetic test To genotype these samples we implemented aCYP2D6 Genotyping Platform based on MALDI-TOF MSThis way we would be able to compare our genotyping resultswith a previous sequencing analysis in [5]
21 CYP2D6 Genotyping Platform
211 Long Primary PCR Selective amplification of theCYP2D6 genewas carried outmodifying a long PCR protocolimplemented in our previous work [5] Forward primer (P-1780 Table 2) was designed in a highly nonhomologousCYP2D6CYP2D7PCYP2D8P 51015840 untranslated region [5]The reverse primer used was 2D6-R (Table 2) as previouslydescribed [19] A 51015840 10-mer tag (51015840-ACGTTGGATG-31015840) wasadded to both PCR primers in order to improve PCRefficiency PCR reactions were performed in a final volume of5 120583L using the QIAGEN (Hilden Germany) LongRange PCRKit protocol [20] with the following minor modifications20 ng genomicDNA 400 120583Mof each PCR primer (MetabionMartinsried Germany) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 500 120583M Invitrogen (CA USA) 21015840-
deoxynucleoside-51015840-triphosphate (dNTP) Set PCR GradeThe PCR conditions were as follows initial denaturation at93∘C for 3min 35 cycles at 93∘C for 30 s 61∘C for 30 s and68∘C for 6min
212 SAP Dephosphorylation To neutralize unincorporateddNTPs after amplification reactions 03U of ShrimpAlkalinePhosphatase (SAP) (Sequenom) [26 27] was used SAPcleaved a phosphate from the unincorporated dNTPs con-verting them to 21015840-deoxynucleoside-51015840-diphosphate (dNDP)and rendering them unavailable for following reactionsDephosphorylation conditions were as follows 37∘C for20min and 85∘C for 5min
213 Assay Design SNP-specific unextended minisequenc-ing primers (UEPs) and multiplexed UEPs assays weredesigned using both the SequenomMassARRAYassay designversion 31 and the RealSNP assay database [28] A sequenceof 400 base pairs (bp) flanking each selected SNP was down-loaded from the corresponding genomic sequence stored in
the public NCBI Single Nucleotide Polymorphism dbSNPdatabase [8] or the Ensembl Genome Browser [29] and wasanalyzed by Vector NTI Suite Software version 55 (InforMaxOxford UK) Combination of the UEPs into multiplex assayswas supported by these software applications to allow theoptimization of several different parameters for example toavoid the risk of primer-primer interactions and hairpin-loop formations GC content molecular mass range andannealing temperatures To achieve the highest possiblemultiplexing levels we tested many primer combinationsleading us to the final assay design consisting in a total of69 SNPs successfully assembled in five medium-plex assays(13- 14- 13- 14- and 15-plex) (Table 3) For some SNPsSequenom MassARRAY assay design and RealSNP assaydatabase could not design SNP-specific UEPs because of thepresence of proximal SNPsOther SNPswere excludedduringplatform validation because of UEPs cross-hybridization inhighly homologous PCR template regions or for the presenceof primer dimers which created false allele But the highnumber of SNPs inserted in our analysis has allowed us toinfer the correct alleles for each DNA analysis List of SNPsnot included is summarized in Table 4
214 iPLEX Reactions iPLEX reactions were carried outfollowing the Sequenom standard lowmedium-plex protocol[6 26 27] with minor modifications Because of the lengthof the primary PCR fragment and the high GC contentwe increased the denaturation time at 94∘C from 5 to30 sec and the annealing temperature from 52 to 56∘CAn iPLEX reaction cocktail was added to the amplificationproducts and thermocycled to process the iPLEX reac-tion which involved the enzymatic addition of one of thefour mass-modified nucleotides 2101584031015840-dideoxynucleoside-51015840-triphosphate (ddNTP) into the polymorphic site During theiPLEX reaction each primer was extended by one of theddNTPswhich terminated the extension of primers thus pro-ducing allele-specific extension products of different massesThe iPLEX reaction cocktail included 0222X iPLEX bufferplus 05X iPLEX termination mix 05X iPLEX enzyme andUEPs which were divided into four mass groups accordingto the position of their respectivemass peaks to use the wholespectrum (final concentrations of 08ndash20 120583M) In the iPLEXtermination mix all four ddNTPs are present at the sameconcentration The reactions were performed using a twocycling loop program initial denaturation at 94∘C for 30 sfollowed by 40 cycles of 94∘C for 30 s 56∘C for 5 s and 80∘Cfor 5 s This annealing and extension procedure was repeatedfour times (to give a total of 200 short cycles) followedby a final extension step of 3min at 72∘C After desalting
4 ISRN Genetics
Table3Sequ
encesa
ndmolecular
weightsof
unextend
ed(U
EP)a
ndextend
ed(EP)
iPLE
Xprim
ers
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(1)13-PL
EX3877GgtA
CCTT
CCGCT
TCCA
CCCC
F4978
CCTT
CCGCT
TCCA
CCCC
A5249
ACC
TTCC
GCT
TCCA
CCCC
G5265
G3183GgtA
CCGCA
CCTG
CCCT
ATCA
R5051
CCGCA
CCTG
CCCT
ATCA
C5299
GCC
GCA
CCTG
CCCT
ATCA
T5378
A2615
2617
delAAG
GGCA
GCC
ACTC
TCAC
CTR5091
GGCA
GCC
ACTC
TCAC
CTC
5339
del
GGCA
GCC
ACTC
TCAC
CTT
5418
AGA
2483GgtT
GCG
TAGGAC
CTTG
CCAG
R5211
GCG
TAGGAC
CTTG
CCAG
C5459
GGCG
TAGGAC
CTTG
CCAG
A5483
T3853GgtA
GAAG
CGGAAG
GGCT
TCT
R5275
GAAG
CGGAAG
GGCT
TCTC
5523
GGAAG
CGGAAG
GGCT
TCTT
5603
A3277Tgt
CTG
CAGCG
CTTT
GGGGAC
AF5556
TGCA
GCG
CTTT
GGGGAC
AC5803
CTG
CAGCG
CTTT
GGGGAC
AT5883
T4180GgtC
AGCT
CATA
GGGGGAT
GGG
R5645
AGCT
CATA
GGGGGAT
GGGC
5892
GAG
CTCA
TAGGGGGAT
GGGG
5932
C2939GgtA
TGGGCT
CACG
CTGCA
CATC
R5766
TGGGCT
CACG
CTGCA
CATC
C6013
GTG
GGCT
CACG
CTGCA
CATC
T6093
A3288GgtA
GAT
GTC
ATAT
GGGTC
ACAC
R5828
GAT
GTC
ATAT
GGGTC
ACAC
C6075
GGAT
GTC
ATAT
GGGTC
ACAC
T6155
A3176Cgt
TGGCC
GTG
TCCA
ACAG
GAG
ATF6167
GGCC
GTG
TCCA
ACAG
GAG
ATC
6414
CGGCC
GTG
TCCA
ACAG
GAG
ATT
6494
T2850Cgt
TAG
CTTC
AAT
GAT
GAG
AAC
CTG
F6454
AGCT
TCAAT
GAT
GAG
AAC
CTGC
6701
CAG
CTTC
AAT
GAT
GAG
AAC
CTGT
6781
T2935AgtC
AGTG
CTCA
CGCT
GCA
CATC
CGGA
R7010
AGTG
CTCA
CGCT
GCA
CATC
CGGAG
7297
CAG
TGCT
CACG
CTGCA
CATC
CGGAT
7337
A1757Cgt
TTT
GTG
CCGCC
TTCG
CCAAC
CACT
CF7201
TTGTG
CCGCC
TTCG
CCAAC
CACT
CC7448
CTT
GTG
CCGCC
TTCG
CCAAC
CACT
CT7528
T(2)14-PL
EX1661GgtC
CAGAG
GCG
CTTC
TCCG
TF5162
CAGAG
GCG
CTTC
TCCG
TC5410
CCA
GAG
GCG
CTTC
TCCG
TG5450
G1869Tgt
CAC
GGCT
TTGTC
CAAG
AGR5210
ACGGCT
TTGTC
CAAG
AGA
5482
TAC
GGCT
TTGTC
CAAG
AGG
5498
C1846
GgtA
GGGGCG
AAAG
GGGCG
TCR5342
GGGGCG
AAAG
GGGCG
TCC
5588
GGGGGCG
AAAG
GGGCG
TCT
5668
A214GgtC
TGGAG
GGCG
GCA
GAG
GT
F5357
TGGAG
GGCG
GCA
GAG
GTC
5604
CTG
GAG
GGCG
GCA
GAG
GTG
5644
G19GgtA
CTGGGGCT
AGAAG
CACT
GF5565
CTGGGGCT
AGAAG
CACT
GA
5836
ACT
GGGGCT
AGAAG
CACT
GG
5852
G1707delT
GCG
GCC
TCCT
CGGTC
ACCC
R5702
GCG
GCC
TCCT
CGGTC
ACCC
C5949
del
GCG
GCC
TCCT
CGGTC
ACCC
A5973
T746CgtG
GCG
ACCC
AGCC
TCCT
GAT
CR5710
GCG
ACCC
AGCC
TCCT
GAT
CC5957
GGCG
ACCC
AGCC
TCCT
GAT
CG5997
C997CgtG
GAAC
AGGCG
GGCG
GTC
GGC
R5920
GAAC
AGGCG
GGCG
GTC
GGCC
6167
GGAAC
AGGCG
GGCG
GTC
GGCG
6207
C100CgtT
AAT
GCT
GGGCT
GCA
CGCT
ACF6118
AAT
GCT
GGGCT
GCA
CGCT
ACC
6365
CAAT
GCT
GGGCT
GCA
CGCT
ACT
6445
T1979Tgt
CGGAC
AGCC
CGAC
TCCT
CCTT
CR6303
GGAC
AGCC
CGAC
TCCT
CCTT
CA6574
TGGAC
AGCC
CGAC
TCCT
CCTT
CG6590
C-948Cgt
ACA
GGCT
GGGGCA
AGGGCC
TTC
F6488
CAGGCT
GGGGCA
AGGGCC
TTCC
6735
CCA
GGCT
GGGGCA
AGGGCC
TTCA
6759
A1749AgtG
GCC
CATC
ACCC
ACCG
GAG
TGGT
R6681
GCC
CATC
ACCC
ACCG
GAG
TGGTC
6929
GGCC
CATC
ACCC
ACCG
GAG
TGGTT
7008
A77GgtA
GTA
GCG
TGCA
GCC
CAGCG
TTGG
R6792
GTA
GCG
TGCA
GCC
CAGCG
TTGGC7040
GGTA
GCG
TGCA
GCC
CAGCG
TTGGT
7120
A1513Cgt
TAG
CTGGAC
AGAG
CCAG
GGAC
TGF6835
AGCT
GGAC
AGAG
CCAG
GGAC
TGC7082
CAG
CTGGAC
AGAG
CCAG
GGAC
TGT7162
T(3)13-PL
EX984AgtG
GCG
AGTG
TCCT
CGCC
GR4874
GCG
AGTG
TCCT
CGCC
GC
5121
GGCG
AGTG
TCCT
CGCC
GT
5201
A1863
1864
ins
TTTC
GCC
CCGAC
GCC
CCTT
TCGCC
CCF5043
GAC
GCC
CCTT
TCGCC
CCA
5315
ndashGAC
GCC
CCTT
TCGCC
CCT
5370
TTTC
GCC
CC1039Cgt
TAC
CCAG
ATCC
TGGGTT
TF5161
ACCC
AGAT
CCTG
GGTT
TC5409
CAC
CCAG
ATCC
TGGGTT
TT5489
T1943GgtA
CCAG
CAGCC
TGAG
GAAG
R5229
CCAG
CAGCC
TGAG
GAAG
C5477
GCC
AGCA
GCC
TGAG
GAAG
T5557
A974CgtA
CGGCC
GTG
CGCG
AGGCG
F5253
CGGCC
GTG
CGCG
AGGCG
C5501
CCG
GCC
GTG
CGCG
AGGCG
A5525
A1704
CgtG
GGCA
AGAAG
TCGCT
GGAG
F5614
GGCA
AGAAG
TCGCT
GGAG
C5861
CGGCA
AGAAG
TCGCT
GGAG
G5901
G310GgtT
TAAAT
GCC
CTTC
TCCA
GGA
R5748
TAAAT
GCC
CTTC
TCCA
GGAC
5995
GTA
AAT
GCC
CTTC
TCCA
GGAA
6019
T957CgtT
GTC
GTG
CTCA
ATGGGCT
GG
F5876
GTC
GTG
CTCA
ATGGGCT
GGC
6123
CGTC
GTG
CTCA
ATGGGCT
GGT
6203
T883GgtC
TCCC
CGAAG
CGGCG
CCGCA
AR6073
TCCC
CGAAG
CGGCG
CCGCA
AC6320
GTC
CCCG
AAG
CGGCG
CCGCA
AG6360
C1659GgtA
CGCG
AGCA
GAG
GCG
CTTC
TCC
F6408
CGCG
AGCA
GAG
GCG
CTTC
TCCA
6679
ACG
CGAG
CAGAG
GCG
CTTC
TCCG
6695
G31GgtA
AGCA
GGAAG
ATGGCC
ACTA
TCA
R6777
AGCA
GGAAG
ATGGCC
ACTA
TCAC
7024
GAG
CAGGAAG
ATGGCC
ACTA
TCAT
7104
A1724Cgt
TGCG
AAG
GCG
GCA
CAAAG
GCA
GGC
R7158
GCG
AAG
GCG
GCA
CAAAG
GCA
GGCA7429
TGCG
AAG
GCG
GCA
CAAAG
GCA
GGCG7445
C124GgtA
TCAC
ATGCA
GCA
GGTT
GCC
CAGCC
R7299
TCAC
ATGCA
GCA
GGTT
GCC
CAGCC
C7546
GTC
ACAT
GCA
GCA
GGTT
GCC
CAGCC
T7626
A
ISRN Genetics 5
Table3Con
tinued
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(4)14-PL
EX3198Cgt
GAC
CCAT
CTCT
GGTC
GCC
R5082
ACCC
ATCT
CTGGTC
GCC
C5330
GAC
CCAT
CTCT
GGTC
GCC
G5370
C2549delA
TGGGTC
CCAG
GTC
ATCC
R5162
TGGGTC
CCAG
GTC
ATCC
G5450
del
TGGGTC
CCAG
GTC
ATCC
T5490
A3582AgtG
GAAT
GTT
GGAG
GAC
CCA
F5259
GAAT
GTT
GGAG
GAC
CCAA
5531
AGAAT
GTT
GGAG
GAC
CCAG
5547
G2539
2542
delAAC
TAC
AGCT
GGAT
GAG
CTGCT
F5540
ACAG
CTGGAT
GAG
CTGCT
A5811
AAC
TAC
AGCT
GGAT
GAG
CTGCT
G5827
del
3790Cgt
TCC
ACTC
TCAC
CCTG
CATC
TF5620
CCAC
TCTC
ACCC
TGCA
TCTC
5867
CCC
ACTC
TCAC
CCTG
CATC
TT5947
T4115Cgt
TAC
CTCC
CTGCT
GCA
GCA
CTT
F5989
ACCT
CCCT
GCT
GCA
GCA
CTTC
6236
CAC
CTCC
CTGCT
GCA
GCA
CTTT
6316
T44
81GgtA
GAAT
CTGAC
TGCC
CAGAT
TGF6117
GAAT
CTGAC
TGCC
CAGAT
TGA
6388
AGAAT
CTGAC
TGCC
CAGAT
TGG
6404
G3828GgtA
ACTC
ATCA
CCAAC
CTGTC
ATC
F6270
ACTC
ATCA
CCAAC
CTGTC
ATCA
6541
AAC
TCAT
CACC
AAC
CTGTC
ATCG
6557
G3384AgtC
CATG
CTGGGGCT
ATCA
CCAG
GR6447
CATG
CTGGGGCT
ATCA
CCAG
GG
6734
CCA
TGCT
GGGGCT
ATCA
CCAG
GT
6774
A2853AgtC
GAG
AAC
AGGTC
AGCC
ACCA
CTA
R6722
GAG
AAC
AGGTC
AGCC
ACCA
CTAG
7010
CGAG
AAC
AGGTC
AGCC
ACCA
CTAT
7050
A2988GgtA
CATG
TGCC
CCCG
CCTG
TACC
CTT
R6888
CATG
TGCC
CCCG
CCTG
TACC
CTTC
7135
GCA
TGTG
CCCC
CGCC
TGTA
CCCT
TT7215
A3584GgtA
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
F7098
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
A7369
AGTC
AGAAT
GTT
GGAG
GAC
CCAAC
G7385
G44
01Cgt
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
R7546
TAAC
TGAC
ATCT
GCT
CAGCC
TCAAC
A7817
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
G7833
C2587
2590
delGAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GA
F7846
ATCA
GCT
CAGCC
CCCC
CGAG
ACCT
GAC8093
GAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GAG8133
del
(5)15-PL
EX3948Tgt
GTC
TCAG
CAGGTG
CCTG
F4873
TCTC
AGCA
GGTG
CCTG
G5160
GTC
TCAG
CAGGTG
CCTT
5200
T-1235AgtG
GCA
CCAC
CCAG
CCTA
ATR5084
GCA
CCAC
CCAG
CCTA
ATC
5332
GGCA
CCAC
CCAG
CCTA
ATT
5411
A137138insT
GAAG
TCCA
CATG
CAGCA
R5188
GAAG
TCCA
CATG
CAGCA
A5460
TGAAG
TCCA
CATG
CAGCA
G5476
mdash4155Cgt
TCA
GCC
CCGGCC
CAGCC
ACF5376
CAGCC
CCGGCC
CAGCC
ACC
5623
CCA
GCC
CCGGCC
CAGCC
ACT
5703
T82Cgt
TGTG
TAGCG
TGCA
GCC
CAGC
R5830
GTG
TAGCG
TGCA
GCC
CAGCA
6101
TGTG
TAGCG
TGCA
GCC
CAGCG
6117
C-1584CgtG
CTGGAC
AAC
TTGGAAG
AAC
CF6135
CTGGAC
AAC
TTGGAAG
AAC
CC6382
CCT
GGAC
AAC
TTGGAAG
AAC
CG6422
G2575Cgt
AGAC
CTGGGAC
CCAG
CCCA
GCC
F6362
GAC
CTGGGAC
CCAG
CCCA
GCC
C6609
CGAC
CTGGGAC
CCAG
CCCA
GCC
A6633
A843TgtG
ACTA
GGAC
CTGTA
GTC
TGGGG
F6502
ACTA
GGAC
CTGTA
GTC
TGGGGG
6789
GAC
TAGGAC
CTGTA
GTC
TGGGGT
6829
T-740
CgtT
ACAG
ACTC
ACAC
TGAC
ACTT
AGR6672
ACAG
ACTC
ACAC
TGAC
ACTT
AGA
6944
TAC
AGAC
TCAC
ACTG
ACAC
TTAG
G6960
C-678GgtA
CTTT
GTG
TGGGTG
ATTT
TCTG
CF6769
CTTT
GTG
TGGGTG
ATTT
TCTG
CA7041
ACT
TTGTG
TGGGTG
ATTT
TCTG
CG7057
G-750
-749
delGA
TGTG
ACTG
GTG
TGTG
TGAG
AGA
F6902
TGTG
ACTG
GTG
TGTG
TGAG
AGAA
7173
del
TGTG
ACTG
GTG
TGTG
TGAG
AGAG
7189
GA
2291GgtA
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CR7204
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CC7451
GCA
CTCG
CCAAG
TGCC
AGCC
TCCA
CT7531
A-1426CgtT
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
R7286
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
A7533
TGTG
TGCC
ACCA
CGTC
TAGCT
TTTT
G7573
C
1758GgtTgt
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCF7490
TTGTG
CCGCC
TTCG
CCAAC
CACT
CCA7761
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCG7777
GTT
GTG
CCGCC
TTCG
CCAAC
CACT
CCT7817
T-100
0GgtA
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGT
R7635
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGTC7882
GAC
ATCC
TCCC
GGGCT
GCC
TGAG
GGTT
7962
AUDU
nextendedminindashsequ
encing
prim
erDire
ctionFFo
rwardUEP
directionR
ReverseU
EPdirectionUMU
nextendedminindashsequ
encing
prim
ersM
assEL
ME
xtendedprim
erLo
wer
MassEH
ME
xtended
prim
erHigherM
assEL
CEx
tend
edprim
erlower
Massn
ucleotideC
alledEH
CEx
tend
edprim
erhigh
erMassn
ucleotideC
alledAllmolecular
weightm
assesa
reexpressedin
Dalton(D
a)
6 ISRN Genetics
Table 4 List of SNPs excluded in assay design validation andcorrelation to transcriptional variations
SNPs excluded Variations Reason-1770GgtA mdash Cross-hybridization-1298GgtA mdash Primer dimers-1253AgtG mdash Primer dimers
-1237 -1236insAA mdash Proximal SNP -1235AgtGPrimer dimers
1973 1974insG Frameshift Proximal SNP 1979TgtC1976GgtA mdash Proximal SNP 1979TgtC1978CgtT mdash Proximal SNP 1979TgtC2097AgtG mdash Primer dimers2470TgtC mdash Cross hybridization4042GgtA mdash Primer dimers
by addition of 6mg clean resin (Sequenom) each 384-wellsample was diluted with 16 120583L of sterilized H
2Odd
Multiplex PCR reactions SAP dephosphorylation andiPLEX reactionswere performedusingThermo-Fast 384 PCRPlates (ABgene Epsom UK) and a DNA Engine Tetrad 2Peltier Thermal Cycler (Bio-Rad CA USA) All pipettingsteps were performed using the automatic station MatrixPlateMate 2 times 2 (Sequenom)
215 MALDI-TOF MS Measurement An aliquot rangingfrom 15 to 20 nL of each iPLEX reaction product was loadedin a 384-spot SpectroChip (Sequenom) using the MassAr-ray Nanodispenser (Samsung Seoul Repubic of Korea)SpectroChip analysis was performed by MassARRAY Com-pact System (Sequenom) After laser desorptionionizationautomated spectra acquisition analysis was performed andinterpreted using Sequenom MassARRAY RT version 33software Examples of multiplex mass spectrum and clusterplot distributions are shown in Figures 1 and 2
22 CYP2D6 Single Allele Genotyping Following directionof our previous work [5] we decided to apply the singleallele protocol creating a single allele genotyping methodMALDI-TOFMS based For each sample a double long PCRreaction was carried out using P-1584 WT or P-1584 MUT[5] (Table 2) as forward primers The reverse primer was2D6-R [19] for both PCR reactions In this way we wereable to directly determine a direct and correct chromosomephase in samples presenting with a heterozygous status forminus1584GgtC SNP PCR reactions were performed in a finalvolume of 5 120583L using 20 ng genomic DNA 400 120583M of eachPCR primer (Metabion) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 800 120583MInvitrogen dNTP set PCR grade
The PCR conditions were as follows initial denaturation at93∘C for 3min 10 cycles at 93∘C for 30 s 67∘C for 30 s and68∘C for 3min 25 cycles at 93∘C for 30 s 65∘C for 30 s and68∘C for 6min SAP dephosphorylation iPLEX ReactionsandMALDI-TOFMSmeasurement were performed without
Table 5CYP2D6 allele frequencies in 250 healthy Sardinian peopleTotal chromosomes number = 500 Human cytochrome P450 AlleleNomenclature Committee [7] served as reference for variant alleleand correlated enzymatic activity
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast1A 148 296 EMlowast1B 5 10 EMlowast1D 4 08 EMlowast1E mdash mdash EMlowast2A 75 150 EMlowast2B mdash mdash EMlowast2D mdash mdash EMlowast2E mdash mdash EMlowast2F mdash mdash EMlowast2G mdash mdash EMlowast2K mdash mdash EMlowast2L 11 22 EMlowast2M 8 16 EMSH3 7 14 Not knownSH4 1 02 Not knownlowast3A mdash mdash PMlowast3B 11 22 PMlowast4A 84 168 PMlowast4B mdash mdash PMlowast4D mdash mdash PMlowast4K mdash mdash PMlowast4L mdash mdash PMlowast4M mdash mdash PMlowast4N mdash mdash PMlowast5 5 10 PMlowast6A 1 02 PMlowast6C mdash mdash PMlowast6D mdash mdash PMlowast7 mdash mdash PMlowast8 mdash mdash PMlowast9 1 02 IMlowast10A mdash mdash IMlowast10B 27 54 IMlowast11 mdash mdash PMlowast12 mdash mdash PMlowast14A mdash mdash PMlowast14B mdash mdash IMlowast15 3 06 PMlowast17 mdash mdash IMlowast19 mdash mdash PMlowast20 1 02 PMlowast22 mdash mdash Not knownlowast23 mdash mdash Not known
ISRN Genetics 7
Table 5 Continued
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast24 mdash mdash Not knownlowast25 mdash mdash Not knownlowast26 mdash mdash Not knownlowast27 mdash mdash EMlowast28 4 08 Not knownlowast29 mdash mdash IMlowast30 mdash mdash Not knownlowast31 mdash mdash PMlowast32 mdash mdash Not knownlowast33 mdash mdash EMlowast35A 5 10 EMlowast36 mdash mdash IMlowast37 mdash mdash Not knownlowast38 mdash mdash PMlowast39 mdash mdash EMlowast41 46 92 IMSH1 41 82 Not knownSH2 2 04 Not knownlowast43 mdash mdash Not knownlowast58 mdash mdash Not knownlowast59 mdash mdash IMlowast64 mdash mdash Not knownlowast65 mdash mdash Not knownlowast1xN 4 08 UMlowast2xN 6 12 UMSH1234 = Sardinian haplotype 1234 [5 22ndash25] CYP2D6lowast5lowast1xN andlowast2xN alleles were evaluated byThe CYP2D6Applied Biosystems CNVAssayin [5]
modifications as indicated in paragraph 1 ldquoCYP2D6 Geno-typing Platformrdquo In multiplexed assay 5 minus1584GgtC UEP(Table 3) was excluded Examples of cluster distribution fornovel 3176CgtT and 3948TgtG SNPs [5 17 18] are shown inFigure 3
3 Results and Discussion
A CYP2D6 screening assay was developed using theSequenom MassARRAY platform to simultaneously identifythe most frequent and some rare CYP2D6 Caucasian allelesWe have modified the basic Sequenom iPLEX assay and useda new primary PCR strategy based on the amplification ofthe entire gene [5] coupled with multiplex primer extensionreactionsThis strategy avoids false genotyping which wouldresult in nonspecific coamplification of the homologouspseudogenes CYP2D7P and CYP2D8P and secondly itreduces the number of PCR primers used to select regionscontaining the targeted polymorphisms Multiplexing wasperformed for 69 SNPs which represents 66 of the mostfrequent and some rarer variants and subvariants reported
5000 5500 6000 6500 7000 7500 8000
0123456789
10
Mass
Inte
nsity
lowast lowast TCG A
Figure 1 An example of 15ndashplex mass spectrum is shown (Well 5)List of SNPs investigated in this plex is shown in Table 3 The figureshows two examples highlighted in different colours Unextendprimer (UEP) peak is marked by an asterisk and dotted arrow whilethe solid arrows indicate the presence of the two different allelesDotted vertical lines represent UEPs and extended primers (EPs)expected masses In blue an AG heterozygous genotype example isshown lowast minus1235AgtG UEP expected mass = 5084 Dalton (Da) Gminus1235AgtGEP expectedmass = 5332Da Aminus1235AgtGEP expectedmass = 5411 Da In red a CC wild type homozygous genotypeexample is shown lowast 82CgtT UEP expected mass = 5830Da T82CgtT not EP expected mass = 6101Da C 82CgtT EP expectedmass = 6117Da
to date in the Caucasian population [5 7 9ndash16] and knownto be responsible for absent reduced or extensive metabolicactivity (Tables 1 and 5) Due to the high possibility ofrecombination it was possible to insert African African-American (11986211988411987521198636lowast2119871 lowast2 119872 lowast4119873 lowast6 4 and lowast6 5) andAsian (11986211988411987521198636lowast3119860 lowast4119861 lowast4119871 lowast14 lowast36 and lowast39) variantsin the study A sample of 250 unrelated healthy Sardinianindividuals analyzed in [5] was submitted to MALDI-TOFMS genotyping for these 69 CYP2D6 SNPs An example ofmultiplex mass spectrum is shown in Figure 1 In Figure 2(a)an example of the cluster plot distribution for the 1661GgtCSNP is shown Spectrum peak intensities were not correctlybalanced in some heterozygous samples (Figures 2(c) and2(d)) which appeared as outliers in the cluster distribution(Figure 2(a) point 120575 and 120576) The CYP2D6 Applied BiosystemsCopy Number Variation (CNV) Assay used to analyze theseDNAs in our previous work [5] detected the presence ofduplications or multiplications in 100 of the analyzedsamples presenting this kind of distribution Furthermore insamples presenting a heterozygous status for minus1584GgtC SNPwe applied long PCR single allele analysis [5] to MALDI-TOF MS screening assays and inferred a direct haplotypephase To verify if our CYP2D6 platform works correctly wecompared genotyping results and haplotype phase elaboratedin the two screening platforms with our results found in [5]A consensus of 100 was found for all samples (Table 5)Moreover for all samples presenting the 214GgtC SNP inhomo- or heterozygous status in MALDI-TOF MS analysisit was possible to verify in our previous sequencing analysis
8 ISRN Genetics
120572
120573
120574
120577
120575
120576
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0010203040506070809
1
(a)
0
2
4
6
8
10
12
14
16
18
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(b)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(c)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(d)
Figure 2 In (a) a Cluster plot for 1661GgtC SNP is shown Homozygous wild type GG samples are displayed in (120572) homozygous mutate CCsamples in (120573) and heterozygous GC samples in (120574) in (120575) and (120576) are displayed outlier samples resulting in gene copy number variation [5](120577) negative control (H2Odd) In (b) aMass Spectra for the detection of 1661GgtC in a heterozygous GC sample displayed in (a) position (120574)In (c) a Mass Spectra for the detection of 1661GgtC in the outlier sample displayed in (a) position (120575) and resulting in 11986211988411987521198636lowast1 times 119873 allele[5] In (d) Mass Spectra for the detection of 1661G gt C in an outlier sample displayed in (a) position (120576) and resulting in 11986211988411987521198636lowast2 times 119873allele [5]
[5] the presence of all SNPs correlated to gene conversionto CYP2D7P in Intron 1 (Figure 4) which indicated thereliability of MALDI-TOF analysis for this variation
4 Conclusions
Differences in drug responses could be due to genetic fac-tors Knowledge of individual genetic profiling is clinicallyimportant and provides benefits for future medical care bypredicting the drug response or developing DNA-based testsSubstantial interindividual variability in response to specifictherapies might be caused by the presence of polymorphismsin genes encoding components of drugmetabolismpathwayssuch as theCYP450 family genes Polymorphisms inCYP2D6gene have been thoroughly investigated including their asso-ciations with the incidence of adverse reactions In this study
we have developed a reliablemedium-throughput genotypingplatformusing the SequenomMassARRAY system to providea simultaneous screening of the most relevant CYP2D6 genevariants in several hundreds of subjects MALDI-TOF MS-based analyses have the potential to become a useful approachin clinical diagnoses as they are very flexible and applicableto individualized genetic therapies The screening platformdeveloped in this study coupled with The CYP2D6 AppliedBiosystems CNVAssay provides a robust alternative tomanycurrently available CYP450-genotyping approaches and aimsto increase the number of responders and to limit theincidences of adverse events Moreover by modifying longPCR Forward primer we have implemented a rapid strategyto infer the phase by direct analysis using MALDI-TOS MSmultiplex assays Concordance between data found in thiswork and our direct genomic DNA sequencing analysis donein [5] reliably validated ourMALDI-TOFMS-based platform
ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml
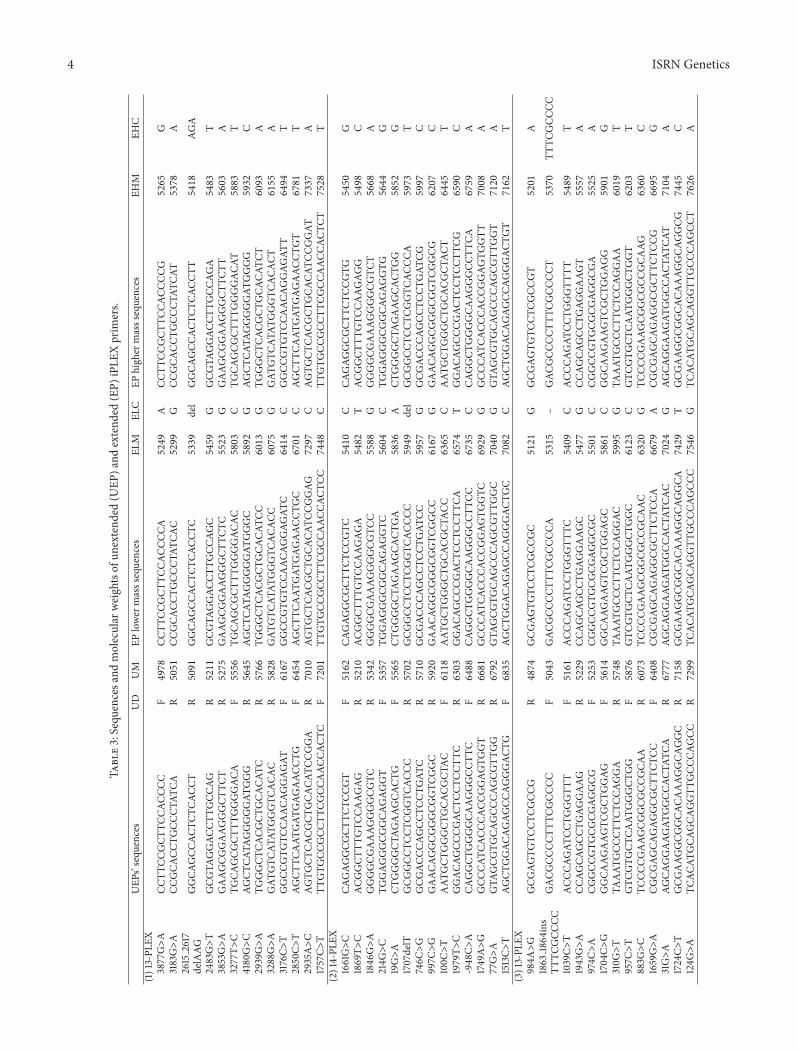
4 ISRN Genetics
Table3Sequ
encesa
ndmolecular
weightsof
unextend
ed(U
EP)a
ndextend
ed(EP)
iPLE
Xprim
ers
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(1)13-PL
EX3877GgtA
CCTT
CCGCT
TCCA
CCCC
F4978
CCTT
CCGCT
TCCA
CCCC
A5249
ACC
TTCC
GCT
TCCA
CCCC
G5265
G3183GgtA
CCGCA
CCTG
CCCT
ATCA
R5051
CCGCA
CCTG
CCCT
ATCA
C5299
GCC
GCA
CCTG
CCCT
ATCA
T5378
A2615
2617
delAAG
GGCA
GCC
ACTC
TCAC
CTR5091
GGCA
GCC
ACTC
TCAC
CTC
5339
del
GGCA
GCC
ACTC
TCAC
CTT
5418
AGA
2483GgtT
GCG
TAGGAC
CTTG
CCAG
R5211
GCG
TAGGAC
CTTG
CCAG
C5459
GGCG
TAGGAC
CTTG
CCAG
A5483
T3853GgtA
GAAG
CGGAAG
GGCT
TCT
R5275
GAAG
CGGAAG
GGCT
TCTC
5523
GGAAG
CGGAAG
GGCT
TCTT
5603
A3277Tgt
CTG
CAGCG
CTTT
GGGGAC
AF5556
TGCA
GCG
CTTT
GGGGAC
AC5803
CTG
CAGCG
CTTT
GGGGAC
AT5883
T4180GgtC
AGCT
CATA
GGGGGAT
GGG
R5645
AGCT
CATA
GGGGGAT
GGGC
5892
GAG
CTCA
TAGGGGGAT
GGGG
5932
C2939GgtA
TGGGCT
CACG
CTGCA
CATC
R5766
TGGGCT
CACG
CTGCA
CATC
C6013
GTG
GGCT
CACG
CTGCA
CATC
T6093
A3288GgtA
GAT
GTC
ATAT
GGGTC
ACAC
R5828
GAT
GTC
ATAT
GGGTC
ACAC
C6075
GGAT
GTC
ATAT
GGGTC
ACAC
T6155
A3176Cgt
TGGCC
GTG
TCCA
ACAG
GAG
ATF6167
GGCC
GTG
TCCA
ACAG
GAG
ATC
6414
CGGCC
GTG
TCCA
ACAG
GAG
ATT
6494
T2850Cgt
TAG
CTTC
AAT
GAT
GAG
AAC
CTG
F6454
AGCT
TCAAT
GAT
GAG
AAC
CTGC
6701
CAG
CTTC
AAT
GAT
GAG
AAC
CTGT
6781
T2935AgtC
AGTG
CTCA
CGCT
GCA
CATC
CGGA
R7010
AGTG
CTCA
CGCT
GCA
CATC
CGGAG
7297
CAG
TGCT
CACG
CTGCA
CATC
CGGAT
7337
A1757Cgt
TTT
GTG
CCGCC
TTCG
CCAAC
CACT
CF7201
TTGTG
CCGCC
TTCG
CCAAC
CACT
CC7448
CTT
GTG
CCGCC
TTCG
CCAAC
CACT
CT7528
T(2)14-PL
EX1661GgtC
CAGAG
GCG
CTTC
TCCG
TF5162
CAGAG
GCG
CTTC
TCCG
TC5410
CCA
GAG
GCG
CTTC
TCCG
TG5450
G1869Tgt
CAC
GGCT
TTGTC
CAAG
AGR5210
ACGGCT
TTGTC
CAAG
AGA
5482
TAC
GGCT
TTGTC
CAAG
AGG
5498
C1846
GgtA
GGGGCG
AAAG
GGGCG
TCR5342
GGGGCG
AAAG
GGGCG
TCC
5588
GGGGGCG
AAAG
GGGCG
TCT
5668
A214GgtC
TGGAG
GGCG
GCA
GAG
GT
F5357
TGGAG
GGCG
GCA
GAG
GTC
5604
CTG
GAG
GGCG
GCA
GAG
GTG
5644
G19GgtA
CTGGGGCT
AGAAG
CACT
GF5565
CTGGGGCT
AGAAG
CACT
GA
5836
ACT
GGGGCT
AGAAG
CACT
GG
5852
G1707delT
GCG
GCC
TCCT
CGGTC
ACCC
R5702
GCG
GCC
TCCT
CGGTC
ACCC
C5949
del
GCG
GCC
TCCT
CGGTC
ACCC
A5973
T746CgtG
GCG
ACCC
AGCC
TCCT
GAT
CR5710
GCG
ACCC
AGCC
TCCT
GAT
CC5957
GGCG
ACCC
AGCC
TCCT
GAT
CG5997
C997CgtG
GAAC
AGGCG
GGCG
GTC
GGC
R5920
GAAC
AGGCG
GGCG
GTC
GGCC
6167
GGAAC
AGGCG
GGCG
GTC
GGCG
6207
C100CgtT
AAT
GCT
GGGCT
GCA
CGCT
ACF6118
AAT
GCT
GGGCT
GCA
CGCT
ACC
6365
CAAT
GCT
GGGCT
GCA
CGCT
ACT
6445
T1979Tgt
CGGAC
AGCC
CGAC
TCCT
CCTT
CR6303
GGAC
AGCC
CGAC
TCCT
CCTT
CA6574
TGGAC
AGCC
CGAC
TCCT
CCTT
CG6590
C-948Cgt
ACA
GGCT
GGGGCA
AGGGCC
TTC
F6488
CAGGCT
GGGGCA
AGGGCC
TTCC
6735
CCA
GGCT
GGGGCA
AGGGCC
TTCA
6759
A1749AgtG
GCC
CATC
ACCC
ACCG
GAG
TGGT
R6681
GCC
CATC
ACCC
ACCG
GAG
TGGTC
6929
GGCC
CATC
ACCC
ACCG
GAG
TGGTT
7008
A77GgtA
GTA
GCG
TGCA
GCC
CAGCG
TTGG
R6792
GTA
GCG
TGCA
GCC
CAGCG
TTGGC7040
GGTA
GCG
TGCA
GCC
CAGCG
TTGGT
7120
A1513Cgt
TAG
CTGGAC
AGAG
CCAG
GGAC
TGF6835
AGCT
GGAC
AGAG
CCAG
GGAC
TGC7082
CAG
CTGGAC
AGAG
CCAG
GGAC
TGT7162
T(3)13-PL
EX984AgtG
GCG
AGTG
TCCT
CGCC
GR4874
GCG
AGTG
TCCT
CGCC
GC
5121
GGCG
AGTG
TCCT
CGCC
GT
5201
A1863
1864
ins
TTTC
GCC
CCGAC
GCC
CCTT
TCGCC
CCF5043
GAC
GCC
CCTT
TCGCC
CCA
5315
ndashGAC
GCC
CCTT
TCGCC
CCT
5370
TTTC
GCC
CC1039Cgt
TAC
CCAG
ATCC
TGGGTT
TF5161
ACCC
AGAT
CCTG
GGTT
TC5409
CAC
CCAG
ATCC
TGGGTT
TT5489
T1943GgtA
CCAG
CAGCC
TGAG
GAAG
R5229
CCAG
CAGCC
TGAG
GAAG
C5477
GCC
AGCA
GCC
TGAG
GAAG
T5557
A974CgtA
CGGCC
GTG
CGCG
AGGCG
F5253
CGGCC
GTG
CGCG
AGGCG
C5501
CCG
GCC
GTG
CGCG
AGGCG
A5525
A1704
CgtG
GGCA
AGAAG
TCGCT
GGAG
F5614
GGCA
AGAAG
TCGCT
GGAG
C5861
CGGCA
AGAAG
TCGCT
GGAG
G5901
G310GgtT
TAAAT
GCC
CTTC
TCCA
GGA
R5748
TAAAT
GCC
CTTC
TCCA
GGAC
5995
GTA
AAT
GCC
CTTC
TCCA
GGAA
6019
T957CgtT
GTC
GTG
CTCA
ATGGGCT
GG
F5876
GTC
GTG
CTCA
ATGGGCT
GGC
6123
CGTC
GTG
CTCA
ATGGGCT
GGT
6203
T883GgtC
TCCC
CGAAG
CGGCG
CCGCA
AR6073
TCCC
CGAAG
CGGCG
CCGCA
AC6320
GTC
CCCG
AAG
CGGCG
CCGCA
AG6360
C1659GgtA
CGCG
AGCA
GAG
GCG
CTTC
TCC
F6408
CGCG
AGCA
GAG
GCG
CTTC
TCCA
6679
ACG
CGAG
CAGAG
GCG
CTTC
TCCG
6695
G31GgtA
AGCA
GGAAG
ATGGCC
ACTA
TCA
R6777
AGCA
GGAAG
ATGGCC
ACTA
TCAC
7024
GAG
CAGGAAG
ATGGCC
ACTA
TCAT
7104
A1724Cgt
TGCG
AAG
GCG
GCA
CAAAG
GCA
GGC
R7158
GCG
AAG
GCG
GCA
CAAAG
GCA
GGCA7429
TGCG
AAG
GCG
GCA
CAAAG
GCA
GGCG7445
C124GgtA
TCAC
ATGCA
GCA
GGTT
GCC
CAGCC
R7299
TCAC
ATGCA
GCA
GGTT
GCC
CAGCC
C7546
GTC
ACAT
GCA
GCA
GGTT
GCC
CAGCC
T7626
A
ISRN Genetics 5
Table3Con
tinued
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(4)14-PL
EX3198Cgt
GAC
CCAT
CTCT
GGTC
GCC
R5082
ACCC
ATCT
CTGGTC
GCC
C5330
GAC
CCAT
CTCT
GGTC
GCC
G5370
C2549delA
TGGGTC
CCAG
GTC
ATCC
R5162
TGGGTC
CCAG
GTC
ATCC
G5450
del
TGGGTC
CCAG
GTC
ATCC
T5490
A3582AgtG
GAAT
GTT
GGAG
GAC
CCA
F5259
GAAT
GTT
GGAG
GAC
CCAA
5531
AGAAT
GTT
GGAG
GAC
CCAG
5547
G2539
2542
delAAC
TAC
AGCT
GGAT
GAG
CTGCT
F5540
ACAG
CTGGAT
GAG
CTGCT
A5811
AAC
TAC
AGCT
GGAT
GAG
CTGCT
G5827
del
3790Cgt
TCC
ACTC
TCAC
CCTG
CATC
TF5620
CCAC
TCTC
ACCC
TGCA
TCTC
5867
CCC
ACTC
TCAC
CCTG
CATC
TT5947
T4115Cgt
TAC
CTCC
CTGCT
GCA
GCA
CTT
F5989
ACCT
CCCT
GCT
GCA
GCA
CTTC
6236
CAC
CTCC
CTGCT
GCA
GCA
CTTT
6316
T44
81GgtA
GAAT
CTGAC
TGCC
CAGAT
TGF6117
GAAT
CTGAC
TGCC
CAGAT
TGA
6388
AGAAT
CTGAC
TGCC
CAGAT
TGG
6404
G3828GgtA
ACTC
ATCA
CCAAC
CTGTC
ATC
F6270
ACTC
ATCA
CCAAC
CTGTC
ATCA
6541
AAC
TCAT
CACC
AAC
CTGTC
ATCG
6557
G3384AgtC
CATG
CTGGGGCT
ATCA
CCAG
GR6447
CATG
CTGGGGCT
ATCA
CCAG
GG
6734
CCA
TGCT
GGGGCT
ATCA
CCAG
GT
6774
A2853AgtC
GAG
AAC
AGGTC
AGCC
ACCA
CTA
R6722
GAG
AAC
AGGTC
AGCC
ACCA
CTAG
7010
CGAG
AAC
AGGTC
AGCC
ACCA
CTAT
7050
A2988GgtA
CATG
TGCC
CCCG
CCTG
TACC
CTT
R6888
CATG
TGCC
CCCG
CCTG
TACC
CTTC
7135
GCA
TGTG
CCCC
CGCC
TGTA
CCCT
TT7215
A3584GgtA
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
F7098
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
A7369
AGTC
AGAAT
GTT
GGAG
GAC
CCAAC
G7385
G44
01Cgt
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
R7546
TAAC
TGAC
ATCT
GCT
CAGCC
TCAAC
A7817
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
G7833
C2587
2590
delGAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GA
F7846
ATCA
GCT
CAGCC
CCCC
CGAG
ACCT
GAC8093
GAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GAG8133
del
(5)15-PL
EX3948Tgt
GTC
TCAG
CAGGTG
CCTG
F4873
TCTC
AGCA
GGTG
CCTG
G5160
GTC
TCAG
CAGGTG
CCTT
5200
T-1235AgtG
GCA
CCAC
CCAG
CCTA
ATR5084
GCA
CCAC
CCAG
CCTA
ATC
5332
GGCA
CCAC
CCAG
CCTA
ATT
5411
A137138insT
GAAG
TCCA
CATG
CAGCA
R5188
GAAG
TCCA
CATG
CAGCA
A5460
TGAAG
TCCA
CATG
CAGCA
G5476
mdash4155Cgt
TCA
GCC
CCGGCC
CAGCC
ACF5376
CAGCC
CCGGCC
CAGCC
ACC
5623
CCA
GCC
CCGGCC
CAGCC
ACT
5703
T82Cgt
TGTG
TAGCG
TGCA
GCC
CAGC
R5830
GTG
TAGCG
TGCA
GCC
CAGCA
6101
TGTG
TAGCG
TGCA
GCC
CAGCG
6117
C-1584CgtG
CTGGAC
AAC
TTGGAAG
AAC
CF6135
CTGGAC
AAC
TTGGAAG
AAC
CC6382
CCT
GGAC
AAC
TTGGAAG
AAC
CG6422
G2575Cgt
AGAC
CTGGGAC
CCAG
CCCA
GCC
F6362
GAC
CTGGGAC
CCAG
CCCA
GCC
C6609
CGAC
CTGGGAC
CCAG
CCCA
GCC
A6633
A843TgtG
ACTA
GGAC
CTGTA
GTC
TGGGG
F6502
ACTA
GGAC
CTGTA
GTC
TGGGGG
6789
GAC
TAGGAC
CTGTA
GTC
TGGGGT
6829
T-740
CgtT
ACAG
ACTC
ACAC
TGAC
ACTT
AGR6672
ACAG
ACTC
ACAC
TGAC
ACTT
AGA
6944
TAC
AGAC
TCAC
ACTG
ACAC
TTAG
G6960
C-678GgtA
CTTT
GTG
TGGGTG
ATTT
TCTG
CF6769
CTTT
GTG
TGGGTG
ATTT
TCTG
CA7041
ACT
TTGTG
TGGGTG
ATTT
TCTG
CG7057
G-750
-749
delGA
TGTG
ACTG
GTG
TGTG
TGAG
AGA
F6902
TGTG
ACTG
GTG
TGTG
TGAG
AGAA
7173
del
TGTG
ACTG
GTG
TGTG
TGAG
AGAG
7189
GA
2291GgtA
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CR7204
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CC7451
GCA
CTCG
CCAAG
TGCC
AGCC
TCCA
CT7531
A-1426CgtT
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
R7286
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
A7533
TGTG
TGCC
ACCA
CGTC
TAGCT
TTTT
G7573
C
1758GgtTgt
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCF7490
TTGTG
CCGCC
TTCG
CCAAC
CACT
CCA7761
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCG7777
GTT
GTG
CCGCC
TTCG
CCAAC
CACT
CCT7817
T-100
0GgtA
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGT
R7635
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGTC7882
GAC
ATCC
TCCC
GGGCT
GCC
TGAG
GGTT
7962
AUDU
nextendedminindashsequ
encing
prim
erDire
ctionFFo
rwardUEP
directionR
ReverseU
EPdirectionUMU
nextendedminindashsequ
encing
prim
ersM
assEL
ME
xtendedprim
erLo
wer
MassEH
ME
xtended
prim
erHigherM
assEL
CEx
tend
edprim
erlower
Massn
ucleotideC
alledEH
CEx
tend
edprim
erhigh
erMassn
ucleotideC
alledAllmolecular
weightm
assesa
reexpressedin
Dalton(D
a)
6 ISRN Genetics
Table 4 List of SNPs excluded in assay design validation andcorrelation to transcriptional variations
SNPs excluded Variations Reason-1770GgtA mdash Cross-hybridization-1298GgtA mdash Primer dimers-1253AgtG mdash Primer dimers
-1237 -1236insAA mdash Proximal SNP -1235AgtGPrimer dimers
1973 1974insG Frameshift Proximal SNP 1979TgtC1976GgtA mdash Proximal SNP 1979TgtC1978CgtT mdash Proximal SNP 1979TgtC2097AgtG mdash Primer dimers2470TgtC mdash Cross hybridization4042GgtA mdash Primer dimers
by addition of 6mg clean resin (Sequenom) each 384-wellsample was diluted with 16 120583L of sterilized H
2Odd
Multiplex PCR reactions SAP dephosphorylation andiPLEX reactionswere performedusingThermo-Fast 384 PCRPlates (ABgene Epsom UK) and a DNA Engine Tetrad 2Peltier Thermal Cycler (Bio-Rad CA USA) All pipettingsteps were performed using the automatic station MatrixPlateMate 2 times 2 (Sequenom)
215 MALDI-TOF MS Measurement An aliquot rangingfrom 15 to 20 nL of each iPLEX reaction product was loadedin a 384-spot SpectroChip (Sequenom) using the MassAr-ray Nanodispenser (Samsung Seoul Repubic of Korea)SpectroChip analysis was performed by MassARRAY Com-pact System (Sequenom) After laser desorptionionizationautomated spectra acquisition analysis was performed andinterpreted using Sequenom MassARRAY RT version 33software Examples of multiplex mass spectrum and clusterplot distributions are shown in Figures 1 and 2
22 CYP2D6 Single Allele Genotyping Following directionof our previous work [5] we decided to apply the singleallele protocol creating a single allele genotyping methodMALDI-TOFMS based For each sample a double long PCRreaction was carried out using P-1584 WT or P-1584 MUT[5] (Table 2) as forward primers The reverse primer was2D6-R [19] for both PCR reactions In this way we wereable to directly determine a direct and correct chromosomephase in samples presenting with a heterozygous status forminus1584GgtC SNP PCR reactions were performed in a finalvolume of 5 120583L using 20 ng genomic DNA 400 120583M of eachPCR primer (Metabion) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 800 120583MInvitrogen dNTP set PCR grade
The PCR conditions were as follows initial denaturation at93∘C for 3min 10 cycles at 93∘C for 30 s 67∘C for 30 s and68∘C for 3min 25 cycles at 93∘C for 30 s 65∘C for 30 s and68∘C for 6min SAP dephosphorylation iPLEX ReactionsandMALDI-TOFMSmeasurement were performed without
Table 5CYP2D6 allele frequencies in 250 healthy Sardinian peopleTotal chromosomes number = 500 Human cytochrome P450 AlleleNomenclature Committee [7] served as reference for variant alleleand correlated enzymatic activity
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast1A 148 296 EMlowast1B 5 10 EMlowast1D 4 08 EMlowast1E mdash mdash EMlowast2A 75 150 EMlowast2B mdash mdash EMlowast2D mdash mdash EMlowast2E mdash mdash EMlowast2F mdash mdash EMlowast2G mdash mdash EMlowast2K mdash mdash EMlowast2L 11 22 EMlowast2M 8 16 EMSH3 7 14 Not knownSH4 1 02 Not knownlowast3A mdash mdash PMlowast3B 11 22 PMlowast4A 84 168 PMlowast4B mdash mdash PMlowast4D mdash mdash PMlowast4K mdash mdash PMlowast4L mdash mdash PMlowast4M mdash mdash PMlowast4N mdash mdash PMlowast5 5 10 PMlowast6A 1 02 PMlowast6C mdash mdash PMlowast6D mdash mdash PMlowast7 mdash mdash PMlowast8 mdash mdash PMlowast9 1 02 IMlowast10A mdash mdash IMlowast10B 27 54 IMlowast11 mdash mdash PMlowast12 mdash mdash PMlowast14A mdash mdash PMlowast14B mdash mdash IMlowast15 3 06 PMlowast17 mdash mdash IMlowast19 mdash mdash PMlowast20 1 02 PMlowast22 mdash mdash Not knownlowast23 mdash mdash Not known
ISRN Genetics 7
Table 5 Continued
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast24 mdash mdash Not knownlowast25 mdash mdash Not knownlowast26 mdash mdash Not knownlowast27 mdash mdash EMlowast28 4 08 Not knownlowast29 mdash mdash IMlowast30 mdash mdash Not knownlowast31 mdash mdash PMlowast32 mdash mdash Not knownlowast33 mdash mdash EMlowast35A 5 10 EMlowast36 mdash mdash IMlowast37 mdash mdash Not knownlowast38 mdash mdash PMlowast39 mdash mdash EMlowast41 46 92 IMSH1 41 82 Not knownSH2 2 04 Not knownlowast43 mdash mdash Not knownlowast58 mdash mdash Not knownlowast59 mdash mdash IMlowast64 mdash mdash Not knownlowast65 mdash mdash Not knownlowast1xN 4 08 UMlowast2xN 6 12 UMSH1234 = Sardinian haplotype 1234 [5 22ndash25] CYP2D6lowast5lowast1xN andlowast2xN alleles were evaluated byThe CYP2D6Applied Biosystems CNVAssayin [5]
modifications as indicated in paragraph 1 ldquoCYP2D6 Geno-typing Platformrdquo In multiplexed assay 5 minus1584GgtC UEP(Table 3) was excluded Examples of cluster distribution fornovel 3176CgtT and 3948TgtG SNPs [5 17 18] are shown inFigure 3
3 Results and Discussion
A CYP2D6 screening assay was developed using theSequenom MassARRAY platform to simultaneously identifythe most frequent and some rare CYP2D6 Caucasian allelesWe have modified the basic Sequenom iPLEX assay and useda new primary PCR strategy based on the amplification ofthe entire gene [5] coupled with multiplex primer extensionreactionsThis strategy avoids false genotyping which wouldresult in nonspecific coamplification of the homologouspseudogenes CYP2D7P and CYP2D8P and secondly itreduces the number of PCR primers used to select regionscontaining the targeted polymorphisms Multiplexing wasperformed for 69 SNPs which represents 66 of the mostfrequent and some rarer variants and subvariants reported
5000 5500 6000 6500 7000 7500 8000
0123456789
10
Mass
Inte
nsity
lowast lowast TCG A
Figure 1 An example of 15ndashplex mass spectrum is shown (Well 5)List of SNPs investigated in this plex is shown in Table 3 The figureshows two examples highlighted in different colours Unextendprimer (UEP) peak is marked by an asterisk and dotted arrow whilethe solid arrows indicate the presence of the two different allelesDotted vertical lines represent UEPs and extended primers (EPs)expected masses In blue an AG heterozygous genotype example isshown lowast minus1235AgtG UEP expected mass = 5084 Dalton (Da) Gminus1235AgtGEP expectedmass = 5332Da Aminus1235AgtGEP expectedmass = 5411 Da In red a CC wild type homozygous genotypeexample is shown lowast 82CgtT UEP expected mass = 5830Da T82CgtT not EP expected mass = 6101Da C 82CgtT EP expectedmass = 6117Da
to date in the Caucasian population [5 7 9ndash16] and knownto be responsible for absent reduced or extensive metabolicactivity (Tables 1 and 5) Due to the high possibility ofrecombination it was possible to insert African African-American (11986211988411987521198636lowast2119871 lowast2 119872 lowast4119873 lowast6 4 and lowast6 5) andAsian (11986211988411987521198636lowast3119860 lowast4119861 lowast4119871 lowast14 lowast36 and lowast39) variantsin the study A sample of 250 unrelated healthy Sardinianindividuals analyzed in [5] was submitted to MALDI-TOFMS genotyping for these 69 CYP2D6 SNPs An example ofmultiplex mass spectrum is shown in Figure 1 In Figure 2(a)an example of the cluster plot distribution for the 1661GgtCSNP is shown Spectrum peak intensities were not correctlybalanced in some heterozygous samples (Figures 2(c) and2(d)) which appeared as outliers in the cluster distribution(Figure 2(a) point 120575 and 120576) The CYP2D6 Applied BiosystemsCopy Number Variation (CNV) Assay used to analyze theseDNAs in our previous work [5] detected the presence ofduplications or multiplications in 100 of the analyzedsamples presenting this kind of distribution Furthermore insamples presenting a heterozygous status for minus1584GgtC SNPwe applied long PCR single allele analysis [5] to MALDI-TOF MS screening assays and inferred a direct haplotypephase To verify if our CYP2D6 platform works correctly wecompared genotyping results and haplotype phase elaboratedin the two screening platforms with our results found in [5]A consensus of 100 was found for all samples (Table 5)Moreover for all samples presenting the 214GgtC SNP inhomo- or heterozygous status in MALDI-TOF MS analysisit was possible to verify in our previous sequencing analysis
8 ISRN Genetics
120572
120573
120574
120577
120575
120576
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0010203040506070809
1
(a)
0
2
4
6
8
10
12
14
16
18
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(b)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(c)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(d)
Figure 2 In (a) a Cluster plot for 1661GgtC SNP is shown Homozygous wild type GG samples are displayed in (120572) homozygous mutate CCsamples in (120573) and heterozygous GC samples in (120574) in (120575) and (120576) are displayed outlier samples resulting in gene copy number variation [5](120577) negative control (H2Odd) In (b) aMass Spectra for the detection of 1661GgtC in a heterozygous GC sample displayed in (a) position (120574)In (c) a Mass Spectra for the detection of 1661GgtC in the outlier sample displayed in (a) position (120575) and resulting in 11986211988411987521198636lowast1 times 119873 allele[5] In (d) Mass Spectra for the detection of 1661G gt C in an outlier sample displayed in (a) position (120576) and resulting in 11986211988411987521198636lowast2 times 119873allele [5]
[5] the presence of all SNPs correlated to gene conversionto CYP2D7P in Intron 1 (Figure 4) which indicated thereliability of MALDI-TOF analysis for this variation
4 Conclusions
Differences in drug responses could be due to genetic fac-tors Knowledge of individual genetic profiling is clinicallyimportant and provides benefits for future medical care bypredicting the drug response or developing DNA-based testsSubstantial interindividual variability in response to specifictherapies might be caused by the presence of polymorphismsin genes encoding components of drugmetabolismpathwayssuch as theCYP450 family genes Polymorphisms inCYP2D6gene have been thoroughly investigated including their asso-ciations with the incidence of adverse reactions In this study
we have developed a reliablemedium-throughput genotypingplatformusing the SequenomMassARRAY system to providea simultaneous screening of the most relevant CYP2D6 genevariants in several hundreds of subjects MALDI-TOF MS-based analyses have the potential to become a useful approachin clinical diagnoses as they are very flexible and applicableto individualized genetic therapies The screening platformdeveloped in this study coupled with The CYP2D6 AppliedBiosystems CNVAssay provides a robust alternative tomanycurrently available CYP450-genotyping approaches and aimsto increase the number of responders and to limit theincidences of adverse events Moreover by modifying longPCR Forward primer we have implemented a rapid strategyto infer the phase by direct analysis using MALDI-TOS MSmultiplex assays Concordance between data found in thiswork and our direct genomic DNA sequencing analysis donein [5] reliably validated ourMALDI-TOFMS-based platform
ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml
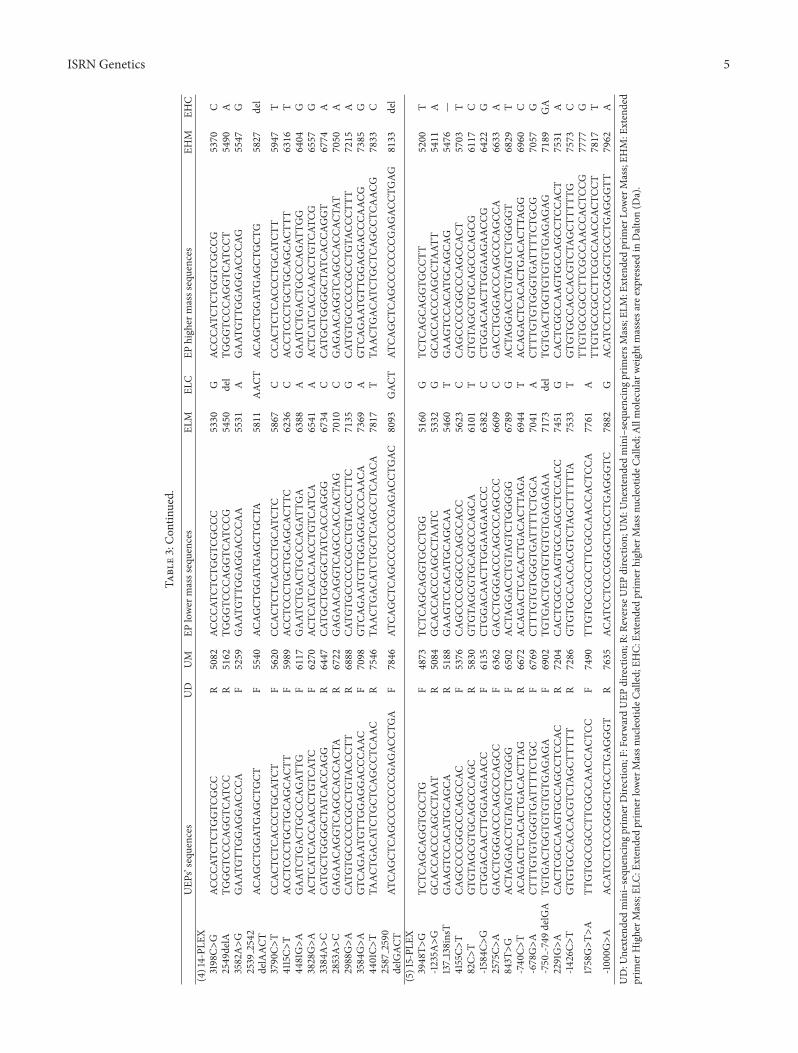
ISRN Genetics 5
Table3Con
tinued
UEP
srsquosequ
ences
UD
UM
EPlower
masssequences
ELM
ELC
EPhigh
ermasssequences
EHM
EHC
(4)14-PL
EX3198Cgt
GAC
CCAT
CTCT
GGTC
GCC
R5082
ACCC
ATCT
CTGGTC
GCC
C5330
GAC
CCAT
CTCT
GGTC
GCC
G5370
C2549delA
TGGGTC
CCAG
GTC
ATCC
R5162
TGGGTC
CCAG
GTC
ATCC
G5450
del
TGGGTC
CCAG
GTC
ATCC
T5490
A3582AgtG
GAAT
GTT
GGAG
GAC
CCA
F5259
GAAT
GTT
GGAG
GAC
CCAA
5531
AGAAT
GTT
GGAG
GAC
CCAG
5547
G2539
2542
delAAC
TAC
AGCT
GGAT
GAG
CTGCT
F5540
ACAG
CTGGAT
GAG
CTGCT
A5811
AAC
TAC
AGCT
GGAT
GAG
CTGCT
G5827
del
3790Cgt
TCC
ACTC
TCAC
CCTG
CATC
TF5620
CCAC
TCTC
ACCC
TGCA
TCTC
5867
CCC
ACTC
TCAC
CCTG
CATC
TT5947
T4115Cgt
TAC
CTCC
CTGCT
GCA
GCA
CTT
F5989
ACCT
CCCT
GCT
GCA
GCA
CTTC
6236
CAC
CTCC
CTGCT
GCA
GCA
CTTT
6316
T44
81GgtA
GAAT
CTGAC
TGCC
CAGAT
TGF6117
GAAT
CTGAC
TGCC
CAGAT
TGA
6388
AGAAT
CTGAC
TGCC
CAGAT
TGG
6404
G3828GgtA
ACTC
ATCA
CCAAC
CTGTC
ATC
F6270
ACTC
ATCA
CCAAC
CTGTC
ATCA
6541
AAC
TCAT
CACC
AAC
CTGTC
ATCG
6557
G3384AgtC
CATG
CTGGGGCT
ATCA
CCAG
GR6447
CATG
CTGGGGCT
ATCA
CCAG
GG
6734
CCA
TGCT
GGGGCT
ATCA
CCAG
GT
6774
A2853AgtC
GAG
AAC
AGGTC
AGCC
ACCA
CTA
R6722
GAG
AAC
AGGTC
AGCC
ACCA
CTAG
7010
CGAG
AAC
AGGTC
AGCC
ACCA
CTAT
7050
A2988GgtA
CATG
TGCC
CCCG
CCTG
TACC
CTT
R6888
CATG
TGCC
CCCG
CCTG
TACC
CTTC
7135
GCA
TGTG
CCCC
CGCC
TGTA
CCCT
TT7215
A3584GgtA
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
F7098
GTC
AGAAT
GTT
GGAG
GAC
CCAAC
A7369
AGTC
AGAAT
GTT
GGAG
GAC
CCAAC
G7385
G44
01Cgt
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
R7546
TAAC
TGAC
ATCT
GCT
CAGCC
TCAAC
A7817
TTA
ACTG
ACAT
CTGCT
CAGCC
TCAAC
G7833
C2587
2590
delGAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GA
F7846
ATCA
GCT
CAGCC
CCCC
CGAG
ACCT
GAC8093
GAC
TAT
CAGCT
CAGCC
CCCC
CGAG
ACCT
GAG8133
del
(5)15-PL
EX3948Tgt
GTC
TCAG
CAGGTG
CCTG
F4873
TCTC
AGCA
GGTG
CCTG
G5160
GTC
TCAG
CAGGTG
CCTT
5200
T-1235AgtG
GCA
CCAC
CCAG
CCTA
ATR5084
GCA
CCAC
CCAG
CCTA
ATC
5332
GGCA
CCAC
CCAG
CCTA
ATT
5411
A137138insT
GAAG
TCCA
CATG
CAGCA
R5188
GAAG
TCCA
CATG
CAGCA
A5460
TGAAG
TCCA
CATG
CAGCA
G5476
mdash4155Cgt
TCA
GCC
CCGGCC
CAGCC
ACF5376
CAGCC
CCGGCC
CAGCC
ACC
5623
CCA
GCC
CCGGCC
CAGCC
ACT
5703
T82Cgt
TGTG
TAGCG
TGCA
GCC
CAGC
R5830
GTG
TAGCG
TGCA
GCC
CAGCA
6101
TGTG
TAGCG
TGCA
GCC
CAGCG
6117
C-1584CgtG
CTGGAC
AAC
TTGGAAG
AAC
CF6135
CTGGAC
AAC
TTGGAAG
AAC
CC6382
CCT
GGAC
AAC
TTGGAAG
AAC
CG6422
G2575Cgt
AGAC
CTGGGAC
CCAG
CCCA
GCC
F6362
GAC
CTGGGAC
CCAG
CCCA
GCC
C6609
CGAC
CTGGGAC
CCAG
CCCA
GCC
A6633
A843TgtG
ACTA
GGAC
CTGTA
GTC
TGGGG
F6502
ACTA
GGAC
CTGTA
GTC
TGGGGG
6789
GAC
TAGGAC
CTGTA
GTC
TGGGGT
6829
T-740
CgtT
ACAG
ACTC
ACAC
TGAC
ACTT
AGR6672
ACAG
ACTC
ACAC
TGAC
ACTT
AGA
6944
TAC
AGAC
TCAC
ACTG
ACAC
TTAG
G6960
C-678GgtA
CTTT
GTG
TGGGTG
ATTT
TCTG
CF6769
CTTT
GTG
TGGGTG
ATTT
TCTG
CA7041
ACT
TTGTG
TGGGTG
ATTT
TCTG
CG7057
G-750
-749
delGA
TGTG
ACTG
GTG
TGTG
TGAG
AGA
F6902
TGTG
ACTG
GTG
TGTG
TGAG
AGAA
7173
del
TGTG
ACTG
GTG
TGTG
TGAG
AGAG
7189
GA
2291GgtA
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CR7204
CACT
CGCC
AAG
TGCC
AGCC
TCCA
CC7451
GCA
CTCG
CCAAG
TGCC
AGCC
TCCA
CT7531
A-1426CgtT
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
R7286
GTG
TGCC
ACCA
CGTC
TAGCT
TTTT
A7533
TGTG
TGCC
ACCA
CGTC
TAGCT
TTTT
G7573
C
1758GgtTgt
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCF7490
TTGTG
CCGCC
TTCG
CCAAC
CACT
CCA7761
ATT
GTG
CCGCC
TTCG
CCAAC
CACT
CCG7777
GTT
GTG
CCGCC
TTCG
CCAAC
CACT
CCT7817
T-100
0GgtA
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGT
R7635
ACAT
CCTC
CCGGGCT
GCC
TGAG
GGTC7882
GAC
ATCC
TCCC
GGGCT
GCC
TGAG
GGTT
7962
AUDU
nextendedminindashsequ
encing
prim
erDire
ctionFFo
rwardUEP
directionR
ReverseU
EPdirectionUMU
nextendedminindashsequ
encing
prim
ersM
assEL
ME
xtendedprim
erLo
wer
MassEH
ME
xtended
prim
erHigherM
assEL
CEx
tend
edprim
erlower
Massn
ucleotideC
alledEH
CEx
tend
edprim
erhigh
erMassn
ucleotideC
alledAllmolecular
weightm
assesa
reexpressedin
Dalton(D
a)
6 ISRN Genetics
Table 4 List of SNPs excluded in assay design validation andcorrelation to transcriptional variations
SNPs excluded Variations Reason-1770GgtA mdash Cross-hybridization-1298GgtA mdash Primer dimers-1253AgtG mdash Primer dimers
-1237 -1236insAA mdash Proximal SNP -1235AgtGPrimer dimers
1973 1974insG Frameshift Proximal SNP 1979TgtC1976GgtA mdash Proximal SNP 1979TgtC1978CgtT mdash Proximal SNP 1979TgtC2097AgtG mdash Primer dimers2470TgtC mdash Cross hybridization4042GgtA mdash Primer dimers
by addition of 6mg clean resin (Sequenom) each 384-wellsample was diluted with 16 120583L of sterilized H
2Odd
Multiplex PCR reactions SAP dephosphorylation andiPLEX reactionswere performedusingThermo-Fast 384 PCRPlates (ABgene Epsom UK) and a DNA Engine Tetrad 2Peltier Thermal Cycler (Bio-Rad CA USA) All pipettingsteps were performed using the automatic station MatrixPlateMate 2 times 2 (Sequenom)
215 MALDI-TOF MS Measurement An aliquot rangingfrom 15 to 20 nL of each iPLEX reaction product was loadedin a 384-spot SpectroChip (Sequenom) using the MassAr-ray Nanodispenser (Samsung Seoul Repubic of Korea)SpectroChip analysis was performed by MassARRAY Com-pact System (Sequenom) After laser desorptionionizationautomated spectra acquisition analysis was performed andinterpreted using Sequenom MassARRAY RT version 33software Examples of multiplex mass spectrum and clusterplot distributions are shown in Figures 1 and 2
22 CYP2D6 Single Allele Genotyping Following directionof our previous work [5] we decided to apply the singleallele protocol creating a single allele genotyping methodMALDI-TOFMS based For each sample a double long PCRreaction was carried out using P-1584 WT or P-1584 MUT[5] (Table 2) as forward primers The reverse primer was2D6-R [19] for both PCR reactions In this way we wereable to directly determine a direct and correct chromosomephase in samples presenting with a heterozygous status forminus1584GgtC SNP PCR reactions were performed in a finalvolume of 5 120583L using 20 ng genomic DNA 400 120583M of eachPCR primer (Metabion) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 800 120583MInvitrogen dNTP set PCR grade
The PCR conditions were as follows initial denaturation at93∘C for 3min 10 cycles at 93∘C for 30 s 67∘C for 30 s and68∘C for 3min 25 cycles at 93∘C for 30 s 65∘C for 30 s and68∘C for 6min SAP dephosphorylation iPLEX ReactionsandMALDI-TOFMSmeasurement were performed without
Table 5CYP2D6 allele frequencies in 250 healthy Sardinian peopleTotal chromosomes number = 500 Human cytochrome P450 AlleleNomenclature Committee [7] served as reference for variant alleleand correlated enzymatic activity
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast1A 148 296 EMlowast1B 5 10 EMlowast1D 4 08 EMlowast1E mdash mdash EMlowast2A 75 150 EMlowast2B mdash mdash EMlowast2D mdash mdash EMlowast2E mdash mdash EMlowast2F mdash mdash EMlowast2G mdash mdash EMlowast2K mdash mdash EMlowast2L 11 22 EMlowast2M 8 16 EMSH3 7 14 Not knownSH4 1 02 Not knownlowast3A mdash mdash PMlowast3B 11 22 PMlowast4A 84 168 PMlowast4B mdash mdash PMlowast4D mdash mdash PMlowast4K mdash mdash PMlowast4L mdash mdash PMlowast4M mdash mdash PMlowast4N mdash mdash PMlowast5 5 10 PMlowast6A 1 02 PMlowast6C mdash mdash PMlowast6D mdash mdash PMlowast7 mdash mdash PMlowast8 mdash mdash PMlowast9 1 02 IMlowast10A mdash mdash IMlowast10B 27 54 IMlowast11 mdash mdash PMlowast12 mdash mdash PMlowast14A mdash mdash PMlowast14B mdash mdash IMlowast15 3 06 PMlowast17 mdash mdash IMlowast19 mdash mdash PMlowast20 1 02 PMlowast22 mdash mdash Not knownlowast23 mdash mdash Not known
ISRN Genetics 7
Table 5 Continued
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast24 mdash mdash Not knownlowast25 mdash mdash Not knownlowast26 mdash mdash Not knownlowast27 mdash mdash EMlowast28 4 08 Not knownlowast29 mdash mdash IMlowast30 mdash mdash Not knownlowast31 mdash mdash PMlowast32 mdash mdash Not knownlowast33 mdash mdash EMlowast35A 5 10 EMlowast36 mdash mdash IMlowast37 mdash mdash Not knownlowast38 mdash mdash PMlowast39 mdash mdash EMlowast41 46 92 IMSH1 41 82 Not knownSH2 2 04 Not knownlowast43 mdash mdash Not knownlowast58 mdash mdash Not knownlowast59 mdash mdash IMlowast64 mdash mdash Not knownlowast65 mdash mdash Not knownlowast1xN 4 08 UMlowast2xN 6 12 UMSH1234 = Sardinian haplotype 1234 [5 22ndash25] CYP2D6lowast5lowast1xN andlowast2xN alleles were evaluated byThe CYP2D6Applied Biosystems CNVAssayin [5]
modifications as indicated in paragraph 1 ldquoCYP2D6 Geno-typing Platformrdquo In multiplexed assay 5 minus1584GgtC UEP(Table 3) was excluded Examples of cluster distribution fornovel 3176CgtT and 3948TgtG SNPs [5 17 18] are shown inFigure 3
3 Results and Discussion
A CYP2D6 screening assay was developed using theSequenom MassARRAY platform to simultaneously identifythe most frequent and some rare CYP2D6 Caucasian allelesWe have modified the basic Sequenom iPLEX assay and useda new primary PCR strategy based on the amplification ofthe entire gene [5] coupled with multiplex primer extensionreactionsThis strategy avoids false genotyping which wouldresult in nonspecific coamplification of the homologouspseudogenes CYP2D7P and CYP2D8P and secondly itreduces the number of PCR primers used to select regionscontaining the targeted polymorphisms Multiplexing wasperformed for 69 SNPs which represents 66 of the mostfrequent and some rarer variants and subvariants reported
5000 5500 6000 6500 7000 7500 8000
0123456789
10
Mass
Inte
nsity
lowast lowast TCG A
Figure 1 An example of 15ndashplex mass spectrum is shown (Well 5)List of SNPs investigated in this plex is shown in Table 3 The figureshows two examples highlighted in different colours Unextendprimer (UEP) peak is marked by an asterisk and dotted arrow whilethe solid arrows indicate the presence of the two different allelesDotted vertical lines represent UEPs and extended primers (EPs)expected masses In blue an AG heterozygous genotype example isshown lowast minus1235AgtG UEP expected mass = 5084 Dalton (Da) Gminus1235AgtGEP expectedmass = 5332Da Aminus1235AgtGEP expectedmass = 5411 Da In red a CC wild type homozygous genotypeexample is shown lowast 82CgtT UEP expected mass = 5830Da T82CgtT not EP expected mass = 6101Da C 82CgtT EP expectedmass = 6117Da
to date in the Caucasian population [5 7 9ndash16] and knownto be responsible for absent reduced or extensive metabolicactivity (Tables 1 and 5) Due to the high possibility ofrecombination it was possible to insert African African-American (11986211988411987521198636lowast2119871 lowast2 119872 lowast4119873 lowast6 4 and lowast6 5) andAsian (11986211988411987521198636lowast3119860 lowast4119861 lowast4119871 lowast14 lowast36 and lowast39) variantsin the study A sample of 250 unrelated healthy Sardinianindividuals analyzed in [5] was submitted to MALDI-TOFMS genotyping for these 69 CYP2D6 SNPs An example ofmultiplex mass spectrum is shown in Figure 1 In Figure 2(a)an example of the cluster plot distribution for the 1661GgtCSNP is shown Spectrum peak intensities were not correctlybalanced in some heterozygous samples (Figures 2(c) and2(d)) which appeared as outliers in the cluster distribution(Figure 2(a) point 120575 and 120576) The CYP2D6 Applied BiosystemsCopy Number Variation (CNV) Assay used to analyze theseDNAs in our previous work [5] detected the presence ofduplications or multiplications in 100 of the analyzedsamples presenting this kind of distribution Furthermore insamples presenting a heterozygous status for minus1584GgtC SNPwe applied long PCR single allele analysis [5] to MALDI-TOF MS screening assays and inferred a direct haplotypephase To verify if our CYP2D6 platform works correctly wecompared genotyping results and haplotype phase elaboratedin the two screening platforms with our results found in [5]A consensus of 100 was found for all samples (Table 5)Moreover for all samples presenting the 214GgtC SNP inhomo- or heterozygous status in MALDI-TOF MS analysisit was possible to verify in our previous sequencing analysis
8 ISRN Genetics
120572
120573
120574
120577
120575
120576
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0010203040506070809
1
(a)
0
2
4
6
8
10
12
14
16
18
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(b)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(c)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(d)
Figure 2 In (a) a Cluster plot for 1661GgtC SNP is shown Homozygous wild type GG samples are displayed in (120572) homozygous mutate CCsamples in (120573) and heterozygous GC samples in (120574) in (120575) and (120576) are displayed outlier samples resulting in gene copy number variation [5](120577) negative control (H2Odd) In (b) aMass Spectra for the detection of 1661GgtC in a heterozygous GC sample displayed in (a) position (120574)In (c) a Mass Spectra for the detection of 1661GgtC in the outlier sample displayed in (a) position (120575) and resulting in 11986211988411987521198636lowast1 times 119873 allele[5] In (d) Mass Spectra for the detection of 1661G gt C in an outlier sample displayed in (a) position (120576) and resulting in 11986211988411987521198636lowast2 times 119873allele [5]
[5] the presence of all SNPs correlated to gene conversionto CYP2D7P in Intron 1 (Figure 4) which indicated thereliability of MALDI-TOF analysis for this variation
4 Conclusions
Differences in drug responses could be due to genetic fac-tors Knowledge of individual genetic profiling is clinicallyimportant and provides benefits for future medical care bypredicting the drug response or developing DNA-based testsSubstantial interindividual variability in response to specifictherapies might be caused by the presence of polymorphismsin genes encoding components of drugmetabolismpathwayssuch as theCYP450 family genes Polymorphisms inCYP2D6gene have been thoroughly investigated including their asso-ciations with the incidence of adverse reactions In this study
we have developed a reliablemedium-throughput genotypingplatformusing the SequenomMassARRAY system to providea simultaneous screening of the most relevant CYP2D6 genevariants in several hundreds of subjects MALDI-TOF MS-based analyses have the potential to become a useful approachin clinical diagnoses as they are very flexible and applicableto individualized genetic therapies The screening platformdeveloped in this study coupled with The CYP2D6 AppliedBiosystems CNVAssay provides a robust alternative tomanycurrently available CYP450-genotyping approaches and aimsto increase the number of responders and to limit theincidences of adverse events Moreover by modifying longPCR Forward primer we have implemented a rapid strategyto infer the phase by direct analysis using MALDI-TOS MSmultiplex assays Concordance between data found in thiswork and our direct genomic DNA sequencing analysis donein [5] reliably validated ourMALDI-TOFMS-based platform
ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml

6 ISRN Genetics
Table 4 List of SNPs excluded in assay design validation andcorrelation to transcriptional variations
SNPs excluded Variations Reason-1770GgtA mdash Cross-hybridization-1298GgtA mdash Primer dimers-1253AgtG mdash Primer dimers
-1237 -1236insAA mdash Proximal SNP -1235AgtGPrimer dimers
1973 1974insG Frameshift Proximal SNP 1979TgtC1976GgtA mdash Proximal SNP 1979TgtC1978CgtT mdash Proximal SNP 1979TgtC2097AgtG mdash Primer dimers2470TgtC mdash Cross hybridization4042GgtA mdash Primer dimers
by addition of 6mg clean resin (Sequenom) each 384-wellsample was diluted with 16 120583L of sterilized H
2Odd
Multiplex PCR reactions SAP dephosphorylation andiPLEX reactionswere performedusingThermo-Fast 384 PCRPlates (ABgene Epsom UK) and a DNA Engine Tetrad 2Peltier Thermal Cycler (Bio-Rad CA USA) All pipettingsteps were performed using the automatic station MatrixPlateMate 2 times 2 (Sequenom)
215 MALDI-TOF MS Measurement An aliquot rangingfrom 15 to 20 nL of each iPLEX reaction product was loadedin a 384-spot SpectroChip (Sequenom) using the MassAr-ray Nanodispenser (Samsung Seoul Repubic of Korea)SpectroChip analysis was performed by MassARRAY Com-pact System (Sequenom) After laser desorptionionizationautomated spectra acquisition analysis was performed andinterpreted using Sequenom MassARRAY RT version 33software Examples of multiplex mass spectrum and clusterplot distributions are shown in Figures 1 and 2
22 CYP2D6 Single Allele Genotyping Following directionof our previous work [5] we decided to apply the singleallele protocol creating a single allele genotyping methodMALDI-TOFMS based For each sample a double long PCRreaction was carried out using P-1584 WT or P-1584 MUT[5] (Table 2) as forward primers The reverse primer was2D6-R [19] for both PCR reactions In this way we wereable to directly determine a direct and correct chromosomephase in samples presenting with a heterozygous status forminus1584GgtC SNP PCR reactions were performed in a finalvolume of 5 120583L using 20 ng genomic DNA 400 120583M of eachPCR primer (Metabion) 02U QIAGEN LongRange PCRenzyme 1X QIAGEN LongRange PCR buffer (containingMgCl
225mM) and 800 120583MInvitrogen dNTP set PCR grade
The PCR conditions were as follows initial denaturation at93∘C for 3min 10 cycles at 93∘C for 30 s 67∘C for 30 s and68∘C for 3min 25 cycles at 93∘C for 30 s 65∘C for 30 s and68∘C for 6min SAP dephosphorylation iPLEX ReactionsandMALDI-TOFMSmeasurement were performed without
Table 5CYP2D6 allele frequencies in 250 healthy Sardinian peopleTotal chromosomes number = 500 Human cytochrome P450 AlleleNomenclature Committee [7] served as reference for variant alleleand correlated enzymatic activity
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast1A 148 296 EMlowast1B 5 10 EMlowast1D 4 08 EMlowast1E mdash mdash EMlowast2A 75 150 EMlowast2B mdash mdash EMlowast2D mdash mdash EMlowast2E mdash mdash EMlowast2F mdash mdash EMlowast2G mdash mdash EMlowast2K mdash mdash EMlowast2L 11 22 EMlowast2M 8 16 EMSH3 7 14 Not knownSH4 1 02 Not knownlowast3A mdash mdash PMlowast3B 11 22 PMlowast4A 84 168 PMlowast4B mdash mdash PMlowast4D mdash mdash PMlowast4K mdash mdash PMlowast4L mdash mdash PMlowast4M mdash mdash PMlowast4N mdash mdash PMlowast5 5 10 PMlowast6A 1 02 PMlowast6C mdash mdash PMlowast6D mdash mdash PMlowast7 mdash mdash PMlowast8 mdash mdash PMlowast9 1 02 IMlowast10A mdash mdash IMlowast10B 27 54 IMlowast11 mdash mdash PMlowast12 mdash mdash PMlowast14A mdash mdash PMlowast14B mdash mdash IMlowast15 3 06 PMlowast17 mdash mdash IMlowast19 mdash mdash PMlowast20 1 02 PMlowast22 mdash mdash Not knownlowast23 mdash mdash Not known
ISRN Genetics 7
Table 5 Continued
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast24 mdash mdash Not knownlowast25 mdash mdash Not knownlowast26 mdash mdash Not knownlowast27 mdash mdash EMlowast28 4 08 Not knownlowast29 mdash mdash IMlowast30 mdash mdash Not knownlowast31 mdash mdash PMlowast32 mdash mdash Not knownlowast33 mdash mdash EMlowast35A 5 10 EMlowast36 mdash mdash IMlowast37 mdash mdash Not knownlowast38 mdash mdash PMlowast39 mdash mdash EMlowast41 46 92 IMSH1 41 82 Not knownSH2 2 04 Not knownlowast43 mdash mdash Not knownlowast58 mdash mdash Not knownlowast59 mdash mdash IMlowast64 mdash mdash Not knownlowast65 mdash mdash Not knownlowast1xN 4 08 UMlowast2xN 6 12 UMSH1234 = Sardinian haplotype 1234 [5 22ndash25] CYP2D6lowast5lowast1xN andlowast2xN alleles were evaluated byThe CYP2D6Applied Biosystems CNVAssayin [5]
modifications as indicated in paragraph 1 ldquoCYP2D6 Geno-typing Platformrdquo In multiplexed assay 5 minus1584GgtC UEP(Table 3) was excluded Examples of cluster distribution fornovel 3176CgtT and 3948TgtG SNPs [5 17 18] are shown inFigure 3
3 Results and Discussion
A CYP2D6 screening assay was developed using theSequenom MassARRAY platform to simultaneously identifythe most frequent and some rare CYP2D6 Caucasian allelesWe have modified the basic Sequenom iPLEX assay and useda new primary PCR strategy based on the amplification ofthe entire gene [5] coupled with multiplex primer extensionreactionsThis strategy avoids false genotyping which wouldresult in nonspecific coamplification of the homologouspseudogenes CYP2D7P and CYP2D8P and secondly itreduces the number of PCR primers used to select regionscontaining the targeted polymorphisms Multiplexing wasperformed for 69 SNPs which represents 66 of the mostfrequent and some rarer variants and subvariants reported
5000 5500 6000 6500 7000 7500 8000
0123456789
10
Mass
Inte
nsity
lowast lowast TCG A
Figure 1 An example of 15ndashplex mass spectrum is shown (Well 5)List of SNPs investigated in this plex is shown in Table 3 The figureshows two examples highlighted in different colours Unextendprimer (UEP) peak is marked by an asterisk and dotted arrow whilethe solid arrows indicate the presence of the two different allelesDotted vertical lines represent UEPs and extended primers (EPs)expected masses In blue an AG heterozygous genotype example isshown lowast minus1235AgtG UEP expected mass = 5084 Dalton (Da) Gminus1235AgtGEP expectedmass = 5332Da Aminus1235AgtGEP expectedmass = 5411 Da In red a CC wild type homozygous genotypeexample is shown lowast 82CgtT UEP expected mass = 5830Da T82CgtT not EP expected mass = 6101Da C 82CgtT EP expectedmass = 6117Da
to date in the Caucasian population [5 7 9ndash16] and knownto be responsible for absent reduced or extensive metabolicactivity (Tables 1 and 5) Due to the high possibility ofrecombination it was possible to insert African African-American (11986211988411987521198636lowast2119871 lowast2 119872 lowast4119873 lowast6 4 and lowast6 5) andAsian (11986211988411987521198636lowast3119860 lowast4119861 lowast4119871 lowast14 lowast36 and lowast39) variantsin the study A sample of 250 unrelated healthy Sardinianindividuals analyzed in [5] was submitted to MALDI-TOFMS genotyping for these 69 CYP2D6 SNPs An example ofmultiplex mass spectrum is shown in Figure 1 In Figure 2(a)an example of the cluster plot distribution for the 1661GgtCSNP is shown Spectrum peak intensities were not correctlybalanced in some heterozygous samples (Figures 2(c) and2(d)) which appeared as outliers in the cluster distribution(Figure 2(a) point 120575 and 120576) The CYP2D6 Applied BiosystemsCopy Number Variation (CNV) Assay used to analyze theseDNAs in our previous work [5] detected the presence ofduplications or multiplications in 100 of the analyzedsamples presenting this kind of distribution Furthermore insamples presenting a heterozygous status for minus1584GgtC SNPwe applied long PCR single allele analysis [5] to MALDI-TOF MS screening assays and inferred a direct haplotypephase To verify if our CYP2D6 platform works correctly wecompared genotyping results and haplotype phase elaboratedin the two screening platforms with our results found in [5]A consensus of 100 was found for all samples (Table 5)Moreover for all samples presenting the 214GgtC SNP inhomo- or heterozygous status in MALDI-TOF MS analysisit was possible to verify in our previous sequencing analysis
8 ISRN Genetics
120572
120573
120574
120577
120575
120576
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0010203040506070809
1
(a)
0
2
4
6
8
10
12
14
16
18
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(b)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(c)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(d)
Figure 2 In (a) a Cluster plot for 1661GgtC SNP is shown Homozygous wild type GG samples are displayed in (120572) homozygous mutate CCsamples in (120573) and heterozygous GC samples in (120574) in (120575) and (120576) are displayed outlier samples resulting in gene copy number variation [5](120577) negative control (H2Odd) In (b) aMass Spectra for the detection of 1661GgtC in a heterozygous GC sample displayed in (a) position (120574)In (c) a Mass Spectra for the detection of 1661GgtC in the outlier sample displayed in (a) position (120575) and resulting in 11986211988411987521198636lowast1 times 119873 allele[5] In (d) Mass Spectra for the detection of 1661G gt C in an outlier sample displayed in (a) position (120576) and resulting in 11986211988411987521198636lowast2 times 119873allele [5]
[5] the presence of all SNPs correlated to gene conversionto CYP2D7P in Intron 1 (Figure 4) which indicated thereliability of MALDI-TOF analysis for this variation
4 Conclusions
Differences in drug responses could be due to genetic fac-tors Knowledge of individual genetic profiling is clinicallyimportant and provides benefits for future medical care bypredicting the drug response or developing DNA-based testsSubstantial interindividual variability in response to specifictherapies might be caused by the presence of polymorphismsin genes encoding components of drugmetabolismpathwayssuch as theCYP450 family genes Polymorphisms inCYP2D6gene have been thoroughly investigated including their asso-ciations with the incidence of adverse reactions In this study
we have developed a reliablemedium-throughput genotypingplatformusing the SequenomMassARRAY system to providea simultaneous screening of the most relevant CYP2D6 genevariants in several hundreds of subjects MALDI-TOF MS-based analyses have the potential to become a useful approachin clinical diagnoses as they are very flexible and applicableto individualized genetic therapies The screening platformdeveloped in this study coupled with The CYP2D6 AppliedBiosystems CNVAssay provides a robust alternative tomanycurrently available CYP450-genotyping approaches and aimsto increase the number of responders and to limit theincidences of adverse events Moreover by modifying longPCR Forward primer we have implemented a rapid strategyto infer the phase by direct analysis using MALDI-TOS MSmultiplex assays Concordance between data found in thiswork and our direct genomic DNA sequencing analysis donein [5] reliably validated ourMALDI-TOFMS-based platform
ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml
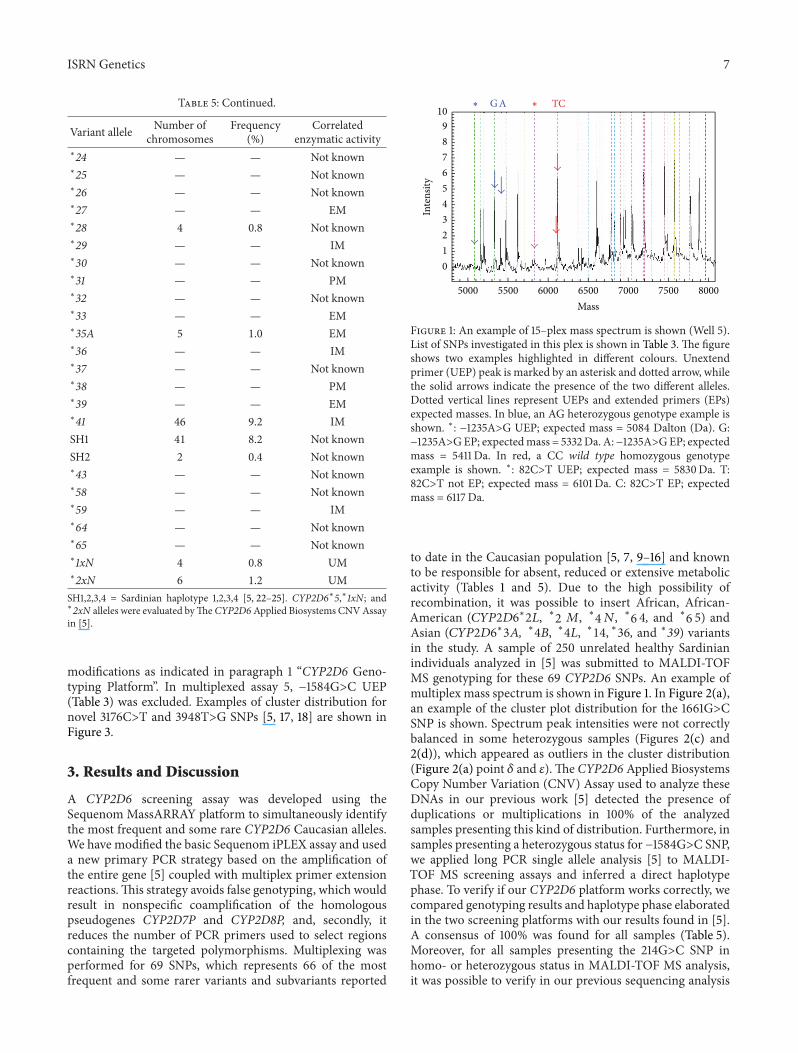
ISRN Genetics 7
Table 5 Continued
Variant allele Number ofchromosomes
Frequency()
Correlatedenzymatic activity
lowast24 mdash mdash Not knownlowast25 mdash mdash Not knownlowast26 mdash mdash Not knownlowast27 mdash mdash EMlowast28 4 08 Not knownlowast29 mdash mdash IMlowast30 mdash mdash Not knownlowast31 mdash mdash PMlowast32 mdash mdash Not knownlowast33 mdash mdash EMlowast35A 5 10 EMlowast36 mdash mdash IMlowast37 mdash mdash Not knownlowast38 mdash mdash PMlowast39 mdash mdash EMlowast41 46 92 IMSH1 41 82 Not knownSH2 2 04 Not knownlowast43 mdash mdash Not knownlowast58 mdash mdash Not knownlowast59 mdash mdash IMlowast64 mdash mdash Not knownlowast65 mdash mdash Not knownlowast1xN 4 08 UMlowast2xN 6 12 UMSH1234 = Sardinian haplotype 1234 [5 22ndash25] CYP2D6lowast5lowast1xN andlowast2xN alleles were evaluated byThe CYP2D6Applied Biosystems CNVAssayin [5]
modifications as indicated in paragraph 1 ldquoCYP2D6 Geno-typing Platformrdquo In multiplexed assay 5 minus1584GgtC UEP(Table 3) was excluded Examples of cluster distribution fornovel 3176CgtT and 3948TgtG SNPs [5 17 18] are shown inFigure 3
3 Results and Discussion
A CYP2D6 screening assay was developed using theSequenom MassARRAY platform to simultaneously identifythe most frequent and some rare CYP2D6 Caucasian allelesWe have modified the basic Sequenom iPLEX assay and useda new primary PCR strategy based on the amplification ofthe entire gene [5] coupled with multiplex primer extensionreactionsThis strategy avoids false genotyping which wouldresult in nonspecific coamplification of the homologouspseudogenes CYP2D7P and CYP2D8P and secondly itreduces the number of PCR primers used to select regionscontaining the targeted polymorphisms Multiplexing wasperformed for 69 SNPs which represents 66 of the mostfrequent and some rarer variants and subvariants reported
5000 5500 6000 6500 7000 7500 8000
0123456789
10
Mass
Inte
nsity
lowast lowast TCG A
Figure 1 An example of 15ndashplex mass spectrum is shown (Well 5)List of SNPs investigated in this plex is shown in Table 3 The figureshows two examples highlighted in different colours Unextendprimer (UEP) peak is marked by an asterisk and dotted arrow whilethe solid arrows indicate the presence of the two different allelesDotted vertical lines represent UEPs and extended primers (EPs)expected masses In blue an AG heterozygous genotype example isshown lowast minus1235AgtG UEP expected mass = 5084 Dalton (Da) Gminus1235AgtGEP expectedmass = 5332Da Aminus1235AgtGEP expectedmass = 5411 Da In red a CC wild type homozygous genotypeexample is shown lowast 82CgtT UEP expected mass = 5830Da T82CgtT not EP expected mass = 6101Da C 82CgtT EP expectedmass = 6117Da
to date in the Caucasian population [5 7 9ndash16] and knownto be responsible for absent reduced or extensive metabolicactivity (Tables 1 and 5) Due to the high possibility ofrecombination it was possible to insert African African-American (11986211988411987521198636lowast2119871 lowast2 119872 lowast4119873 lowast6 4 and lowast6 5) andAsian (11986211988411987521198636lowast3119860 lowast4119861 lowast4119871 lowast14 lowast36 and lowast39) variantsin the study A sample of 250 unrelated healthy Sardinianindividuals analyzed in [5] was submitted to MALDI-TOFMS genotyping for these 69 CYP2D6 SNPs An example ofmultiplex mass spectrum is shown in Figure 1 In Figure 2(a)an example of the cluster plot distribution for the 1661GgtCSNP is shown Spectrum peak intensities were not correctlybalanced in some heterozygous samples (Figures 2(c) and2(d)) which appeared as outliers in the cluster distribution(Figure 2(a) point 120575 and 120576) The CYP2D6 Applied BiosystemsCopy Number Variation (CNV) Assay used to analyze theseDNAs in our previous work [5] detected the presence ofduplications or multiplications in 100 of the analyzedsamples presenting this kind of distribution Furthermore insamples presenting a heterozygous status for minus1584GgtC SNPwe applied long PCR single allele analysis [5] to MALDI-TOF MS screening assays and inferred a direct haplotypephase To verify if our CYP2D6 platform works correctly wecompared genotyping results and haplotype phase elaboratedin the two screening platforms with our results found in [5]A consensus of 100 was found for all samples (Table 5)Moreover for all samples presenting the 214GgtC SNP inhomo- or heterozygous status in MALDI-TOF MS analysisit was possible to verify in our previous sequencing analysis
8 ISRN Genetics
120572
120573
120574
120577
120575
120576
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0010203040506070809
1
(a)
0
2
4
6
8
10
12
14
16
18
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(b)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(c)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(d)
Figure 2 In (a) a Cluster plot for 1661GgtC SNP is shown Homozygous wild type GG samples are displayed in (120572) homozygous mutate CCsamples in (120573) and heterozygous GC samples in (120574) in (120575) and (120576) are displayed outlier samples resulting in gene copy number variation [5](120577) negative control (H2Odd) In (b) aMass Spectra for the detection of 1661GgtC in a heterozygous GC sample displayed in (a) position (120574)In (c) a Mass Spectra for the detection of 1661GgtC in the outlier sample displayed in (a) position (120575) and resulting in 11986211988411987521198636lowast1 times 119873 allele[5] In (d) Mass Spectra for the detection of 1661G gt C in an outlier sample displayed in (a) position (120576) and resulting in 11986211988411987521198636lowast2 times 119873allele [5]
[5] the presence of all SNPs correlated to gene conversionto CYP2D7P in Intron 1 (Figure 4) which indicated thereliability of MALDI-TOF analysis for this variation
4 Conclusions
Differences in drug responses could be due to genetic fac-tors Knowledge of individual genetic profiling is clinicallyimportant and provides benefits for future medical care bypredicting the drug response or developing DNA-based testsSubstantial interindividual variability in response to specifictherapies might be caused by the presence of polymorphismsin genes encoding components of drugmetabolismpathwayssuch as theCYP450 family genes Polymorphisms inCYP2D6gene have been thoroughly investigated including their asso-ciations with the incidence of adverse reactions In this study
we have developed a reliablemedium-throughput genotypingplatformusing the SequenomMassARRAY system to providea simultaneous screening of the most relevant CYP2D6 genevariants in several hundreds of subjects MALDI-TOF MS-based analyses have the potential to become a useful approachin clinical diagnoses as they are very flexible and applicableto individualized genetic therapies The screening platformdeveloped in this study coupled with The CYP2D6 AppliedBiosystems CNVAssay provides a robust alternative tomanycurrently available CYP450-genotyping approaches and aimsto increase the number of responders and to limit theincidences of adverse events Moreover by modifying longPCR Forward primer we have implemented a rapid strategyto infer the phase by direct analysis using MALDI-TOS MSmultiplex assays Concordance between data found in thiswork and our direct genomic DNA sequencing analysis donein [5] reliably validated ourMALDI-TOFMS-based platform
ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml
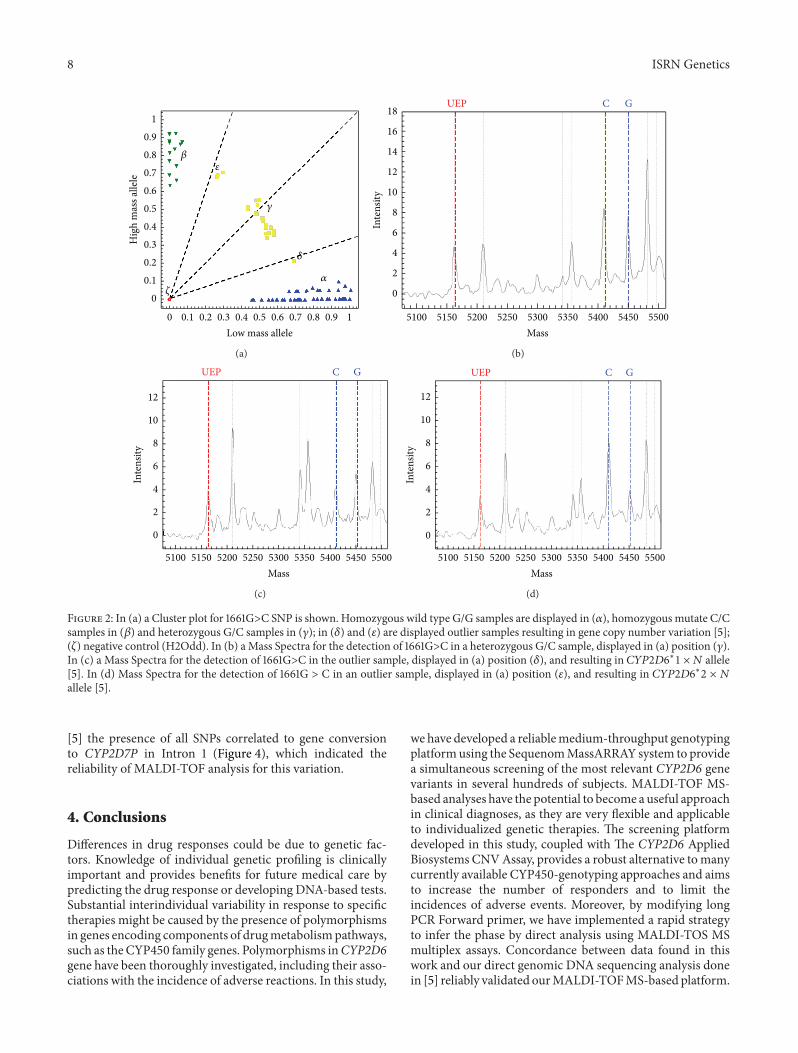
8 ISRN Genetics
120572
120573
120574
120577
120575
120576
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0010203040506070809
1
(a)
0
2
4
6
8
10
12
14
16
18
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(b)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(c)
0
2
4
6
8
10
12
5100 5150 5200 5250 5300 5350 5400 5450 5500
Inte
nsity
Mass
UEP C G
(d)
Figure 2 In (a) a Cluster plot for 1661GgtC SNP is shown Homozygous wild type GG samples are displayed in (120572) homozygous mutate CCsamples in (120573) and heterozygous GC samples in (120574) in (120575) and (120576) are displayed outlier samples resulting in gene copy number variation [5](120577) negative control (H2Odd) In (b) aMass Spectra for the detection of 1661GgtC in a heterozygous GC sample displayed in (a) position (120574)In (c) a Mass Spectra for the detection of 1661GgtC in the outlier sample displayed in (a) position (120575) and resulting in 11986211988411987521198636lowast1 times 119873 allele[5] In (d) Mass Spectra for the detection of 1661G gt C in an outlier sample displayed in (a) position (120576) and resulting in 11986211988411987521198636lowast2 times 119873allele [5]
[5] the presence of all SNPs correlated to gene conversionto CYP2D7P in Intron 1 (Figure 4) which indicated thereliability of MALDI-TOF analysis for this variation
4 Conclusions
Differences in drug responses could be due to genetic fac-tors Knowledge of individual genetic profiling is clinicallyimportant and provides benefits for future medical care bypredicting the drug response or developing DNA-based testsSubstantial interindividual variability in response to specifictherapies might be caused by the presence of polymorphismsin genes encoding components of drugmetabolismpathwayssuch as theCYP450 family genes Polymorphisms inCYP2D6gene have been thoroughly investigated including their asso-ciations with the incidence of adverse reactions In this study
we have developed a reliablemedium-throughput genotypingplatformusing the SequenomMassARRAY system to providea simultaneous screening of the most relevant CYP2D6 genevariants in several hundreds of subjects MALDI-TOF MS-based analyses have the potential to become a useful approachin clinical diagnoses as they are very flexible and applicableto individualized genetic therapies The screening platformdeveloped in this study coupled with The CYP2D6 AppliedBiosystems CNVAssay provides a robust alternative tomanycurrently available CYP450-genotyping approaches and aimsto increase the number of responders and to limit theincidences of adverse events Moreover by modifying longPCR Forward primer we have implemented a rapid strategyto infer the phase by direct analysis using MALDI-TOS MSmultiplex assays Concordance between data found in thiswork and our direct genomic DNA sequencing analysis donein [5] reliably validated ourMALDI-TOFMS-based platform
ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml

ISRN Genetics 9
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1
120572
120573
120574
120577
(a)
0 01 02 03 04 05 06 07 08 09 1Low mass allele
Hig
h m
ass a
llele
0
01
02
03
04
05
06
07
08
09
1120572
120573
120574120577
(b)
Figure 3 MALDI-TOF MS cluster plot distribution for 3176CgtT [17] and 3948GgtT [18] newly discovered SNPs [5] In each cluster thereare visualized 32 samples presenting heterozygous status for minus1584GgtC SNP and submitted to both types of PCR analysis LongRange PCR(Section 2 paragraph 1) and the two single allele PCR (Section 2 paragraph 2) for a total of 96 PCR analyses (a) Cluster plot for 3176CgtTSNP (120572) homozygous wild type CC samples (120573) one out of the two heterozygous CT samples presenting heterozygous status for minus1584GgtCSNP also (120574) the same heterozygous sample analysed in single allele PCR (120577) negative control (H
2Odd) (b) Cluster plot for 3948TgtG SNP
(120572) homozygous wild type TT samples (120573) the only heterozygous TG sample (120574) the only heterozygous sample analysed in single allelePCR (120577) negative control (H
2Odd)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T G C T G A G G C T C C C C T T T T TA C C C CA A A A A A AAG G G G GGG G G G
170 180 190 200 210
(a)
214 221 223 227 232 233 245G A G G G C G G C A G A G G T CC T G A AG G GT C C C C T T T TA C C C C C CA A A A AA G G G G GGG G G G
170 180 190 200 210 220
(b)
Figure 4 (a) Electropherograms of a homozygous wild type and (b) a homozygous mutate sample for CYP2D6CYP2D7P gene conversionin Intron 1 analyzed in [5] SNPs are circled in purple reference positions are indicated under each SNP For all samples presenting 214GgtCSNP in homo- or heterozygous status in MALDI-TOF MS analysis it was confirmed the presence of all SNPs correlated to gene conversionin Intron 1 by sequencing analysis
10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml

10 ISRN Genetics
Conflict of Interests
The authors declare not to have a financial relation with thecommercial identitiesmentioned in the paper thatmight leadto a conflict of interests
Acknowledgments
The authors gratefully acknowledge Professor FrancescoCucca INN-CNR Cagliari Director for gently furnishingSardinian DNAs Dr Luisella Saba and Dr Elena Congeddufor useful initial information in MALDI-TOF MS technol-ogy Dr Enrico Sorisio PharmaNess Sole Director for hishelpful suggestions Professor Annalisa Marchi Professor ofGenetics at the Faculty of Biology and Pharmacy Universityof Cagliari for her valuable feedback and support
References
[1] M Ingelman-Sundberg S C Sim A Gomez and C Rod-riguez-Antona ldquoInfluence of cytochrome P450 polymorphismson drug therapies pharmacogenetic pharmacoepigenetic andclinical aspectsrdquo Pharmacology amp Therapeutics vol 116 no 3pp 496ndash526 2007
[2] M Ingelman-Sundberg ldquoPharmacogenetics of cytochromeP450 and its applications in drug therapy the past present andfuturerdquo Trends in Pharmacological Sciences vol 25 no 4 pp193ndash200 2004
[3] J Ragoussis ldquoGenotyping technologies for allrdquo Drug DiscoveryToday Technologies vol 3 no 2 pp 115ndash122 2006
[4] M Falzoi A Mossa E Congeddu L Saba and L PanildquoMultiplex genotyping of CYP3A4 CYP3A5 CYP2C9 andCYP2C19 SNPs using MALDI-TOF mass spectrometryrdquo Phar-macogenomics vol 11 no 4 pp 559ndash571 2010
[5] M Falzoi L Pira P Lazzari and L Pani ldquoAnalysis of CYP2D6allele frequencies and identification of novel SNPs and sequencevariations in Sardiniansrdquo ISRN Genetics vol 2013 Article ID204560 10 pages 2013
[6] D van den Boom andM Ehrich ldquoDiscovery and identificationof sequence polymorphisms and mutations with MALDI-TOFMSrdquoMethods in Molecular Biology vol 366 pp 287ndash306 2007
[7] Human Cytochrome P450 Allele Nomenclature Committeehttpwwwcypalleleskise
[8] NCBI Single Nucleotide Polymorphism dbSNP httpwwwncbinlmnihgovprojectsSNP
[9] J Sistonen A Sajantila O Lao J Corander G Barbujaniand S Fuselli ldquoCYP2D6 worldwide genetic variation showshigh frequency of altered activity variants and no continentalstructurerdquo Pharmacogenetics and Genomics vol 17 no 2 pp93ndash101 2007
[10] S Fuselli I Dupanloup E Frigato et al ldquoMolecular diversityat the CYP2D6 locus in the Mediterranean regionrdquo EuropeanJournal of Human Genetics vol 12 no 11 pp 916ndash924 2004
[11] C Sachse J Brockmoller S Bauer and I Roots ldquoCytochromeP450 2D6 variants in a Caucasian population allele frequenciesand phenotypic consequencesrdquo American Journal of HumanGenetics vol 60 no 2 pp 284ndash295 1997
[12] S Raimundo C Toscano K Klein et al ldquoA novel intronicmuta-tion 2988GgtA with high predictivity for impaired function ofcytochrome P450 2D6 in white subjectsrdquoClinical PharmacologyandTherapeutics vol 76 no 2 pp 128ndash138 2004
[13] L D Bradford ldquoCYP2D6 allele frequency in European Cau-casians Asians Africans and their descendantsrdquo Pharmacoge-nomics vol 3 no 2 pp 229ndash243 2002
[14] D Marez M Legrand N Sabbagh et al ldquoPolymorphism ofthe cytochrome P450 CYP2D6 gene in a European populationcharacterization of 48 mutations and 53 alleles their frequen-cies and evolutionrdquo Pharmacogenetics vol 7 no 3 pp 193ndash2021997
[15] M Ingelman-Sundberg ldquoImplications of polymorphic cyto-chrome P450-dependent drug metabolism for drug develop-mentrdquo Drug Metabolism and Disposition vol 29 no 4 part 2pp 570ndash573 2001
[16] T Shimada F Tsumura H Yamazaki F P Guengerich andK Inoue ldquoCharacterization of (+minus)-bufuralol hydroxylationactivities in liver microsomes of Japanese and Caucasian sub-jects genotyped for CYP2D6rdquo Pharmacogenetics vol 11 no 2pp 143ndash156 2001
[17] ss469415642 CYP2D6 3176CgtT 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415642
[18] ss469415643 CYP2D6 3948TgtG 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=469415643
[19] I Johansson E Lundqvist M L Dahl and M Ingelman-Sundberg ldquoPCR-based genotyping for duplicated and deletedCYP2D6 genesrdquo Pharmacogenetics vol 6 no 4 pp 351ndash3551996
[20] QIAGEN LongRange PCR Handbook 2008 httpwwwqiagencomdefaultaspx
[21] ss470263991 CYP2D6 -948CgtA 2011 httpwwwncbinlmnihgovprojectsSNPsnp sscgiss=470263991
[22] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 1 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cds GenBank JN7163731 2011httpwwwncbinlmnihgovnuccoreJN7163731
[23] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 2 cytochrome P450 2D6 variant (CYP2D6) geneCYP2D6lowast2M allele complete cdsGenBank JN7163741 2011httpwwwncbinlmnihgovnuccoreJN7163741
[24] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 3 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163751 2011 httpwwwncbinlmnihgovnuccoreJN7163751
[25] M Falzoi L Pira P Lazzari and L Pani Homo sapienshaplotype 4 cytochrome P450 2D6 variant (CYP2D6)gene CYP2D6lowast2MCYP2D6lowast41 hybrid allele completecds GenBank JN7163761 2011 httpwwwncbinlmnihgovnuccoreJN7163761
[26] POethM Beaulieu C Park et al SequenomApplicationNoteApril 2005
[27] iPLEX Gold Application Guide 2009 httpwwwsequenomcomFilesGenetic-Analysis-FilesiP-LEX-Application-PDFsiPLEX-Gold-Application-Guide-v2r1
[28] Sequenom Assay Designer Suite httpsseqpws1sequenomcomAssayDesignerSuitehtml
[29] Ensembl Genome Browser httpwwwensemblorgindexhtml




















