
Genome-wide fungal stress responsive miRNA expression in wheat
14
1 23 Planta An International Journal of Plant Biology ISSN 0032-0935 Volume 240 Number 6 Planta (2014) 240:1287-1298 DOI 10.1007/s00425-014-2153-8 Genome-wide fungal stress responsive miRNA expression in wheat Behçet Inal, Mine Türktaş, Hakan Eren, Emre Ilhan, Sezer Okay, Mehmet Atak, Mustafa Erayman & Turgay Unver
Transcript of Genome-wide fungal stress responsive miRNA expression in wheat

1 23
PlantaAn International Journal of PlantBiology ISSN 0032-0935Volume 240Number 6 Planta (2014) 240:1287-1298DOI 10.1007/s00425-014-2153-8
Genome-wide fungal stress responsivemiRNA expression in wheat
Behçet Inal, Mine Türktaş, Hakan Eren,Emre Ilhan, Sezer Okay, Mehmet Atak,Mustafa Erayman & Turgay Unver

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL ARTICLE
Genome-wide fungal stress responsive miRNA expression in wheat
Behcet Inal • Mine Turktas • Hakan Eren •
Emre Ilhan • Sezer Okay • Mehmet Atak •
Mustafa Erayman • Turgay Unver
Received: 20 June 2014 / Accepted: 12 August 2014 / Published online: 26 August 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract MicroRNAs (miRNAs) are small non-coding
class of RNAs. They were identified in many plants with
their diverse regulatory roles in several cellular and met-
abolic processes. A number of miRNAs were involved in
biotic and abiotic stress responses. Here, fungal stress
responsive wheat miRNAs were analyzed by using miR-
NA-microarray strategy. Two different fungi (Fusarium
culmorum and Bipolaris sorokiniana) were inoculated on
resistant and sensitive wheat cultivars. A total of 87 dif-
ferentially regulated miRNAs were detected in the
8 9 15 K array including all of the available plant miR-
NAs. Using bioinformatics tools, the target transcripts of
responsive miRNAs were predicted, and related biological
processes and mechanisms were assessed. A number of the
miRNAs such as miR2592s, miR869.1, miR169b were
highly differentially regulated showing more than 200-fold
change upon fungal-inoculation. Some of the miRNAs
were identified as fungal-inoculation responsive for the first
time. The analyses showed that some of the differentially
regulated miRNAs targeted resistance-related genes such
as LRR, glucuronosyl transferase, peroxidase and Pto
kinase. The comparison of the two miRNA-microarray
analyses indicated that fungal-responsive wheat miRNAs
were differentially regulated in pathogen- and cultivar-
specific manners.
Keywords Fungal stress � Microarray � Micro RNA �Wheat
Abbreviations
B Resistant wheat cv. Bezostaja
BC Bezostaja control
BI Bezostaja inoculated
EST Expressed sequence tag
M Susceptible wheat cv. Mizrak
MC Mizrak control
MI Mizrak inoculated
Introduction
Wheat (Triticum aestivum L.) is one of the major crops
grown throughout the world (Tanaka et al. 2013) covering
17 % of all the cultivated land, and supplying ca. 55 % of
the carbohydrates consumed by humans (Han et al. 2013;
Meng et al. 2013). By the year of 2050, it is proposed that
the need for wheat will increase with 60 %; albeit the
production will decrease with 29 % due to the environ-
mental stress factors (Manickavelu et al. 2012). Fusarium
culmorum, a ubiquitous soil-borne fungus, is one of the
biotic stress agents in small-grain cereals, particularly in
wheat and barley. It causes foot- and root-rot, a disease
B. Inal and H. Eren authors have equally contributed.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00425-014-2153-8) contains supplementarymaterial, which is available to authorized users.
B. Inal � M. Turktas � S. Okay � T. Unver (&)
Department of Biology, Faculty of Science, Cankiri Karatekin
University, 18100 Cankiri, Turkey
e-mail: [email protected]; [email protected]
H. Eren � E. Ilhan � M. Erayman
Department of Biology, Faculty of Arts and Science, Mustafa
Kemal University, Hatay, Turkey
M. Atak
Department of Field Crops, Faculty of Agriculture, Mustafa
Kemal University, Hatay, Turkey
123
Planta (2014) 240:1287–1298
DOI 10.1007/s00425-014-2153-8
Author's personal copy

designated as Fusarium head blight (FHB), resulting in a
significant reduction in crop yield and quality as well as
contamination of the grain with mycotoxins (Scherm et al.
2013) such as trichothecenes leading to considerable
human and animal health risks (Kammoun et al. 2010).
Another biotic stress agent in wheat is Bipolaris sorokini-
ana Sacc. Shoemaker [teleomorph Cochliobolus sativus Ito
and Kuribayash (Drechs.)], a fungal phytopathogen with a
short biotrophic phase followed by successful tissue
infection in the necrotrophic growth phase, causing spot
blotch, root-rot, leaf-spot disease, black point, seedling and
head blight in cereal crops (Nizam et al. 2012; Zhang et al.
2012).
MicroRNAs (miRNAs) are an important class of
endogenous, single-stranded, non-coding, small RNAs
(sRNAs) regulating the gene expression at the post-tran-
scriptional levels in eukaryotes and viruses (Eldem et al.
2013; Han et al. 2013). Plant miRNAs are short (20–22 bp)
RNAs produced from imperfectly complementary (stem-
loop) RNA precursors (Meng et al. 2013). The genes
coding for miRNAs are transcribed by RNA polymerase II
or III as pri-miRNAs which are processed by a dicer-like
enzyme (DCL1) to form pre-miRNAs, and further cleaved
by DCL1 into a miRNA:miRNA* duplex. These molecules
are methylated by the RNA methylase HUA ENHANCER
1. Eventually, the mature miRNA is incorporated into a
ribonucleoprotein complex known as RNA-induced
silencing complex (RISC) (Eldem et al. 2013; Han et al.
2013).
Currently, 24,521 miRNA sequences from various
organisms are deposited in the publicly available miRNA
database (miRbase, Release 20.0, June 2013, http://www.
mirbase.org). Among 7,385 mature plant miRNAs pre-
sented in miRbase, 42 of them belong to bread wheat,
while 756 and 713 miRNAs are found for Medicago
truncatula and rice, respectively.
Considering the large genome size of the bread wheat
(Brenchley et al. 2012), a number of wheat miRNAs are
expected to be increased by the further studies. The prop-
erties of the miRNAs, such as their genome location can be
found via next-generation survey sequencing. Tanaka et al.
(2013) reported that some miRNAs are located in repeat
regions of wheat chromosome 6B and 1,805 out of 2,906
miRNA genes were identified in a DNA transposon-Mari-
ner. Deep-sequencing of sRNAs in developing grains
revealed 605 miRNAs representing 540 families in wheat
(Meng et al. 2013). Moreover, in silico analyses can also be
used for miRNA prediction (Pandey et al. 2013). Mani-
ckavelu et al. (2012) analyzed ca. 1 million ESTs and
found that wheat miRNA target sequences had maximum
homology with rice followed by maize and M. truncatula.
Similarly, Han et al. (2013) analyzed wheat ESTs, and
identified 48 new miRNAs belonging to 20 miRNA fami-
lies. Involvement of miRNAs in stress response was
reported by a number of studies. Wang et al. (2013)
reported that miR159, miR167a, and miR171 were signif-
icantly up-regulated in wheat upon UV-B radiation.
Additionally, miR164 was shown to be targeted a novel
NAC transcription factor from the NAM subfamily, nega-
tively regulating resistance of wheat to stripe rust (Feng
et al. 2013b).
The involvement of miRNAs in biotic stress response in
wheat has been exhibited in recent studies. Tae-miR159
targeted TaMYB3 (Feng et al. 2013a), tae-164 targeted a
NAC transcription factor, TaNAC21/22 (Feng et al. 2014),
and tae-miR408 targeted a chemocyanin-like protein gene
(TaCLP1) (Feng et al. 2013b). Additionally, miR167,
miR171, miR444, miR1129 and miR1138 were determined
as players in resistance of wheat against stripe rust (Gupta
et al. 2012). Moreover, 24 miRNAs were identified as
being responsive to powdery mildew infection in wheat via
Solexa high-throughput sequencing Xin et al. (2010).
The aim of this study was to identify the fungal-stress
responsive miRNAs in wheat. The miRNA expression
profiling of fungus inoculated resistant cultivars and sus-
ceptible cultivars were analyzed by miRNA-microarray
approach. Using bioinformatics tools, the target transcripts
of responsive miRNAs were predicted, and related bio-
logical processes and mechanisms were evaluated.
Materials and methods
Plant materials, growth conditions and pathogen
inoculation
In this study, two hexaploid bread wheat Triticum aestivum
L. cultivars with different levels of resistance response
against B. sorokiniana and F. culmorum were used. The
seeds were kindly provided by Research Institute of Field
Crops (TARM) in Ankara, Turkey. Two replicates of the
susceptible cv. Mizrak (M), and resistant cv. Bezostaja
(B) were used for fungal-inoculations. The seeds were
sterilized with 0.35 % (w/v) ethyl methane sulphonate
(EMS). They were planted in sterile peat, sand, and soil
mixed in a ratio of 1:1:1 by weight, and were kept for
8 days at climate cabinet under 24 �C 16/8 h light/dark
periods. Leaves and stems of four-week-old wheat were
individually sprayed with a suspension of 1 9 105 ml-1 B.
sorokiniana and F. culmorum spores. Plants sprayed with
sterile water were used as mock inoculation. 48 h after
inoculation (hai), leaves were harvested, and stored in
-80 �C until use. After 2 weeks, the plants were observed
for pathogen growth and resistance symptoms.
1288 Planta (2014) 240:1287–1298
123
Author's personal copy

RNA extraction, labeling, hybridization and washing
Total RNA was extracted from the leaves of two biological
replicates of pathogen and mock inoculated samples.
TRIzol reagent (Invitrogen) was used for the RNA isolation
according to the manufacturer’s instructions. The integrity
of RNA was evaluated on 1.5 % agarose gel. The amount
of the RNA was determined by NanoDrop 2000c spectro-
photometer (ThermoFisherScientific).
Two separate microRNA-Microarray analyses were
performed for the two different pathogen treatments. One
array slide was hybridized with two replicates of each M
and B cultivars inoculated with B. sorokiniana and their
controls, while the other slide was used for the corre-
sponding samples inoculated with F. culmorum and the
controls. The whole experimental steps for the two arrays
were performed simultaneously.
A total of 100 ng RNA from each sample were labeled
with cyanin-3 (Cy3). Labeling and hybridization steps were
performed according to the miRNA-microarray System
with miRNA Complete Labeling and Hyb Kit (Agilent
Technologies). The arrays were incubated at 55 �C for 20 h
in Agilent hybridization oven rotating at 20 rpm. After
hybridization, washing process was started immediately as
described in the manual. The images of hybridized
microarrays were acquired with MS 200 Microarray
Scanner System (NimbleGen). Scanning of images was
achieved with 5 lm resolution in a 61 9 21.6 mm scan
area, and 532 nm TIFF images were generated.
miRNA-microarray design strategy
The 8 9 15 K custom miRNA-microarrays (Agilent
Technologies) were designed according to manufacturer’s
protocol. To obtain a broad representation of miRNAs, a
total of 11,861 plant miRNAs obtained from the latest
miRBase v. 20 (http://www.mirbase.org, released June,
2013), and PMRD (Plant microRNA Database) http://
bioinformatics.cau.edu.cn/PMRD databases were used for
the probe design. Each miRNA was printed in 20 replicates
on the array. Including internal controls and replicated
probes, the custom 8 9 15 K array contained 237,580
probes, which were derived from all of the available plant
miRNAs in the current databases.
Data processing
To calculate the signal intensities of the spots, the TIFF
images were extracted using Feature Extraction v. 9. 5
(Agilent Technologies). For expression analysis, mean
value of the each probe intensity was used. The raw probe
signals were processed for background-correction with a
median polish algorithm (Irizarry et al. 2003), and were
normalized by quantile normalization method (Bolstad
et al. 2003). The signal intensities were transformed to
log2-ratio data. To obtain the miRNA expression mea-
surement, GeneSpringGX v. 11.5.1 (Agilent Technologies)
software was used. For one-color experiments, P values of
the quantile tests, and the expression ratios between the
two sets were computed. The probes with expression value
more than 1.45-fold and P B 0.05 between the two data
sets were designated as differentially expressed.
Target transcript prediction and onthology analysis
To identify the targets of the differentially expressed
miRNAs, psRNAtarget (http://plantgrn.noble.org/psRNA
Target/), a plant miRNA target finder software web tool,
was used. The parameters were set to T. aestivum unigene
library (v. 12, released on 2010_04_18) to BLAST the
target sites against miRNA sequences.
To annotate the target genes responsible to miRNAs
which were significantly differentially expressed, Blast to
Gene Ontology (Blast2GO) http://www.blast2go.com/
b2glaunch tool was used against the National Center for
Biotechnology Information (NCBI) database.
Validation of miRNA expression
To verify the miRNA-microarray data, 9 miRNAs were
selected based on differential expression patterns and
miRNA expression levels were measured using quantita-
tive RT-PCR (qRT-PCR). To make miRNA quantification
by qRT-PCR, 1,500 ng of RNA was used as template
(Yanik et al. 2013). Stem-loop reverse transcription reac-
tion was carried out in a total volume of 10 ll containing
0.4 mM of dNTP mix, 1 lM of the stem-loop reverse-
transcription (RT) primer. The mixture was incubated at
65 �C for 5 min, and then put on ice for 2 min. Then, 19
first-strand buffer, 0.01 M dithiothreitol (DTT), 0.4 U
RNAseOUT (Invitrogen) and 50 U SuperScriptIII (Invit-
rogen) were added to the mixture. The RT reaction was
achieved as follows: 30 min at 16 �C; and 60 cycles (30 �C
for 30 s, 42 �C for 30 s and 50 �C for 1 s). The RT reac-
tion was ended with 85 �C for 5 min. cDNA synthesis of
PCR control samples including no-RT primer, and no-RNA
template were also prepared (Varkonyi-Gasic et al. 2007;
Unver et al. 2010a).
The real-time qRT-PCR was performed using a SYBR
Green I Master mix from Roche on the LightCycler480 II
Real-Time PCR (Roche). The reaction mix was prepared in
a final volume of 20 ll containing 19 Master mix, 1 pmol
of the forward and reverse primers. Specifically designed
forward primers for each individual miRNA and a uni-
versal reverse-primer (50-GTGCAGGGTCCGAGGT-30)were used for qRT-PCR reactions (Yanik et al. 2013).
Planta (2014) 240:1287–1298 1289
123
Author's personal copy
Sequences of the primers used in the qRT-PCR were listed
in suppl. Table S1. The qRT-PCR conditions were setup as
follows: initial denaturation at 95 �C for 5 min, followed
by 50 cycles at 95 �C for 10 s, 55 �C for 20 s, and 72 �C
for 10 s. The melting curves were adjusted at 95 �C for 5 s
and 55 �C for 1 min, and then cooled to 40 �C for 30 s. All
reactions were repeated as triplicates.
Expression analysis of miRNA targeted transcripts
To quantify the expression level of target transcripts of the
selected stress responsive miRNAs, qRT-PCR analyses
were performed. The reverse transcription reactions of the
target genes were performed with the Fermentas First-
Strand cDNA Synthesis kit (ThermoFisher Scientific) as
mentioned in Turktas et al. (2013). PCR primers specific to
target genes were designed by using Primer3Plus software
v. 2.3.3 (http://primer3plus.com) (Untergasser et al. 2012).
The quantification was also performed using 18SrRNA as
internal reference (GenBank ID): AF147501; forward pri-
mer: 50-GTGACGGGTGACGGAGAATT-30; reverse pri-
mer: 50-GACACTAATGCGCCCGGTAT-30 (Unver et al.
2010b; Eldem et al. 2012; Turktas et al. 2013). Relative
expression levels of the target genes were calculated using
the 2-DDCt method (Schmittgen and Livak 2008).
Results
To identify F. culmorum and B. sorokiniana stress
responsive miRNAs in wheat, two separate 8 9 15 K
custom miRNA-microarray assays were conducted. Com-
parative analyses of the two array datasets were performed
to evaluate how the miRNA patterns are affected by dif-
ferent pathogens. The data obtained from the both miRNA-
microarray experiments were analyzed based on three
comparisons: (1) Bezostaja control (BC) vs Bezostaja
inoculated (BI), (2) Mizrak control (MC) vs Mizrak inoc-
ulated (MI) and (3) Bezostaja inoculated (BI) vs Mizrak
inoculated (MI).
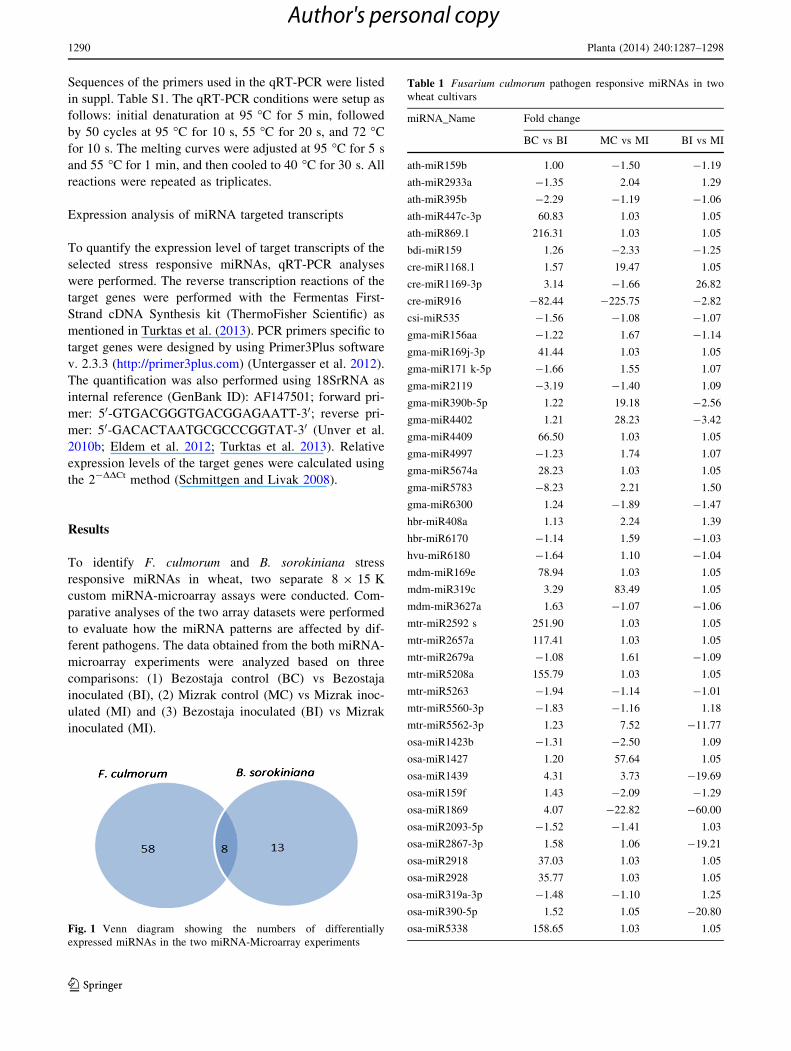
Fig. 1 Venn diagram showing the numbers of differentially
expressed miRNAs in the two miRNA-Microarray experiments
Table 1 Fusarium culmorum pathogen responsive miRNAs in two
wheat cultivars
miRNA_Name Fold change
BC vs BI MC vs MI BI vs MI
ath-miR159b 1.00 -1.50 -1.19
ath-miR2933a -1.35 2.04 1.29
ath-miR395b -2.29 -1.19 -1.06
ath-miR447c-3p 60.83 1.03 1.05
ath-miR869.1 216.31 1.03 1.05
bdi-miR159 1.26 -2.33 -1.25
cre-miR1168.1 1.57 19.47 1.05
cre-miR1169-3p 3.14 -1.66 26.82
cre-miR916 -82.44 -225.75 -2.82
csi-miR535 -1.56 -1.08 -1.07
gma-miR156aa -1.22 1.67 -1.14
gma-miR169j-3p 41.44 1.03 1.05
gma-miR171 k-5p -1.66 1.55 1.07
gma-miR2119 -3.19 -1.40 1.09
gma-miR390b-5p 1.22 19.18 -2.56
gma-miR4402 1.21 28.23 -3.42
gma-miR4409 66.50 1.03 1.05
gma-miR4997 -1.23 1.74 1.07
gma-miR5674a 28.23 1.03 1.05
gma-miR5783 -8.23 2.21 1.50
gma-miR6300 1.24 -1.89 -1.47
hbr-miR408a 1.13 2.24 1.39
hbr-miR6170 -1.14 1.59 -1.03
hvu-miR6180 -1.64 1.10 -1.04
mdm-miR169e 78.94 1.03 1.05
mdm-miR319c 3.29 83.49 1.05
mdm-miR3627a 1.63 -1.07 -1.06
mtr-miR2592 s 251.90 1.03 1.05
mtr-miR2657a 117.41 1.03 1.05
mtr-miR2679a -1.08 1.61 -1.09
mtr-miR5208a 155.79 1.03 1.05
mtr-miR5263 -1.94 -1.14 -1.01
mtr-miR5560-3p -1.83 -1.16 1.18
mtr-miR5562-3p 1.23 7.52 -11.77
osa-miR1423b -1.31 -2.50 1.09
osa-miR1427 1.20 57.64 1.05
osa-miR1439 4.31 3.73 -19.69
osa-miR159f 1.43 -2.09 -1.29
osa-miR1869 4.07 -22.82 -60.00
osa-miR2093-5p -1.52 -1.41 1.03
osa-miR2867-3p 1.58 1.06 -19.21
osa-miR2918 37.03 1.03 1.05
osa-miR2928 35.77 1.03 1.05
osa-miR319a-3p -1.48 -1.10 1.25
osa-miR390-5p 1.52 1.05 -20.80
osa-miR5338 158.65 1.03 1.05
1290 Planta (2014) 240:1287–1298
123
Author's personal copy
Fungal attack responsive miRNA expression patterns
Upon both fungal-inoculations, a total of 87 differentially
expressed wheat miRNAs were identified (Fig. 1). Out of 87
miRNAs, 66 were differentially expressed in wheat upon F.
culmorum pathogen inoculation. The remaining 21 miRNAs
were detected as B. sorokiniana pathogen-responsive.
Additionally, 8 of the 87 miRNAs (athmiR869.1, cre-
miR1169-3p, mtr-miR2592 s-3p, osa-miR1427, osa-
miR319a-3p.2-3p, ptc-miR169b-3p, vvi-miR3624-5p,
miR482e) were found to be common in both miRNA-
microarray analyses.
Identification F. culmorum responsive miRNAs
in wheat
A total of 59,430 probes produced detectable signals in the
array were hybridized with F. culmorum treated plants and
their controls. Although a large number of probes gener-
ated signals, 66 miRNAs were found to be differentially
expressed in at least one comparison between the samples
(Fig. 2; Table 1). These 66 miRNAs were also detected on
16 different plant species including 15 Oryza sativa, 11
Glycine max, 5 Arabidopsis thaliana, 5 Populus tricho-
carpa miRNAs.
The cluster analysis of the signals revealed that the
inoculated samples were grouped together which were
close to susceptible mock inoculation, while resistant mock
inoculation was clustered in a distant branch (Fig. 2).
Out of 66, 9 miRNAs (miR156, miR159, miR169,
miR171, miR319, miR390, miR396, miR408, miR535)
belong to conserved miRNA families. While 19 miRNAs
were found to be up-regulated, 29 miRNAs were detected
as down-regulated in BI compared to BC. On the other
hand, comparison between MI and MC revealed that
expression of 13 miRNAs were found to be induced, and
16 miRNAs were reduced.
The results indicated that some of the differentially
expressed miRNAs showed reverse expression patterns
between the two cultivars. Among the 19 up-regulated
miRNAs in BI vs BC comparison, eight of them (ath-
miR2933a, gma-miR171 k-5p, gma-miR5783, hvu-
miR6180, osa-miR5819, peu-miR2911, ptc-miR474b, vvi-
miR3626-3p) were found to be down-regulated in MI vs
MC comparison. Similarly, ath-miR159b, bdi-miR159, cre-
miR1169-3p, gma-miR6300, osa-miR159f, osa-miR1869,
ptc-miR482c-5p were identified as down-regulated in BI vs
BC library; however, they were observed as up-regulated in
MI vs MC (Table 1).
Additionally, irrelevant from resistance nature of the
cultivars, three miRNAs (cre-miR916, osa-miR5510, ptc-
miR916e-3p) were found to be up-regulated in inoculated
samples compared to control samples (Table 1).
To identify cultivar specific expression, we compared
miRNA patterns of BI vs MI. We found that vvi-miR3626-
3p and cre-miR1169-3p were 166 and 27-fold overex-
pressed, respectively, in BI. On the other hand, eight
miRNAs were measured as highly down-regulated in BI
(Table 1). Interestingly, 30 % of those 66 miRNAs were
highly down-regulated in BI compared to BC, while no
expression difference was observed for those miRNAs
comparing MI vs MC.
Identification B. sorokiniana responsive miRNAs
in wheat
The second microarray slide was subjected to the small
RNAs of B. sorokiniana and mock-inoculated samples.
Similar to the microarray experiment performed using F.
culmorum, 25 % of the all probes produced detectable
signals. The inoculated and control samples were clearly
clustered into different branches (Fig. 3).
The results showed that a total of 21 miRNAs were
differentially expressed between the samples (Table 2).
These 21 miRNAs belong to 11 different plant species
including A. thaliana (2), Chlamydomonas reinhardtii (1),
Table 1 continued
miRNA_Name Fold change
BC vs BI MC vs MI BI vs MI
osa-miR5510 -1.77 -1.67 -1.18
osa-miR5819 -1.56 1.19 -1.02
osa-miR5832 91.42 1.03 1.05
peu-miR2911 -19.92 1.01 1.66
ppt-miR1057 -1.52 -1.02 -1.31
ppt-miR901 20.78 1.76 -56.82
ptc-miR169b-3p 212.16 1.03 1.05
ptc-miR396e-3p -2.46 -1.47 1.07
ptc-miR474b -1.68 1.06 1.14
ptc-miR482c-5p 1.20 -19.50 -18.95
ptc-miR6436 -1.06 -112.18 -109.02
sbi-miR5565d 96.92 1.03 1.05
sbi-miR5565e 43.84 1.03 1.05
sbi-miR6228-5p 94.15 1.03 1.05
sbi-miR6229-5p -1.09 -1.70 1.16
vvi-miR3624-5p 98.31 1.03 1.05
vvi-miR3626-3p -166.78 1.03 166.15
zma-miR169a-3p 137.70 1.03 1.05
zma-miR169r-3p 33.50 1.03 1.05
zma-miR396a-3p -1.97 -1.12 1.04
Fold changes between the samples based on log2 were represented.
The signals were considered as differentially expressed with P B 0.05
and 1.45-fold change values
Planta (2014) 240:1287–1298 1291
123
Author's personal copy

G. max (2), M. truncatula (2), O. sativa (7), Physcomitrella
patens (1), P. trichocarpa (2), Salvia sclarea (1), Sorghum
bicolor (1), Vitis vinifera (1), and Zea mays (1) miRNAs.
The identified miRNAs were found to be the members of
four conserved miRNA families such as miR164, miR169,
miR319, and miR398.
Comparison between BI and BC showed that seven
miRNAs were identified as up-regulated, while 12 miR-
NAs were observed as down-regulated in the fungal-inoc-
ulated sample. On the other hand, fewer numbers of
miRNAs were found to be differentially regulated in sus-
ceptible cultivar M than those found in B. Five miRNAs
were found to be down-regulated, and seven miRNAs were
identified as up-regulated in MI vs MC comparison.
The analyses showed that majority of the miRNAs
represent similar expression patterns in both cultivars. For
instance, osa-miR1427, osa-miR319a-3p, osa-miR528-5p,
osa-miR5525, ssl-miR398, and zma-miR164 g-3p miRNAs
showed high induction in response to fungal attack in both
resistant and susceptible cultivars (Table 2).
On the other hand, ath-miR771, ath-miR869.1, mtr-
miR2592 s, mtr-miR2634, osa-miR6253, ptc-miR169b-3p,
ptc-miR477d-3p, and vvi-miR3624-5p expressions were
found to be down-regulated in BI compared to BC. How-
ever, downregulation of those miRNAs were observed in
MI compared to MC, while their expression differences
were not significant.
To identify cultivar-specific expression, we compared
miRNA patterns of BI vs MI. The gma-miR482e showed
35 fold higher expression in MI compared to BI. On the
other hand, compared to MI, BI had 4.6-fold upregulation
of ssl-miR398 (Table 2).
Target gene analysis of the miRNAs
Using bioinformatic tools, 546 potential target genes were
predicted as 60 F. culmorum responsive miRNAs, and any
potential targets were observed for 6 miRNAs (suppl.
Table S2). The discovered target genes were subjected to
gene ontology (GO) analyses to interpret their biological
functions and cellular organizations. The BLAST analysis
showed that the target genes were involved in a broad
spectrum of processes (Fig. 4). Most of the genes were
involved in metabolic process, protein metabolism, regu-
lation of biological quality, and response to stimulus. The
rest were responsible for cellular process, carbohydrate
metabolism, gene expression, developmental process,
transport, signal transduction, oxidation–reduction process,
RNA metabolic process, reproductive process and other
categories.
Fig. 2 The heat map of F. culmorum responsive miRNAs in wheat miRNA-Microarray assay. Normalized signal intensities of 66 differentially
expressed miRNAs were illustrated in color scheme ranged from red to blue. The signal intensities decreases from red to blue color
1292 Planta (2014) 240:1287–1298
123
Author's personal copy

We found a total of 155 transcripts targeted by 18 dif-
ferently expressed B. sorokiniana pathogen responsive
miRNAs (suppl. Table 3). The analyses showed that the
identified miRNAs target numerous transcripts with diverse
functions (Fig. 5). The targets were involved in response to
stimulus which was followed by transport, response to
stress, carbohydrate metabolism, cellular process, protein
metabolism, gene expression, oxidation–reduction process,
peroxidase reaction and other processes. The majority of
the miRNAs were involved in regulation of numerous
target genes. Among the responsive miRNA, ssl-miR398
targets the highest number of genes with 101 transcripts,
while ath-miR869 targets only one transcript (probable
glucuronoxylan glucuronosyltransferase f8h-like).
Validation of miRNA-microarray data
To validate the results of the two miRNA-microarrays,
expression of the selected nine fungal-responsive miRNAs
and their target transcripts were measured by qRT-PCR
(Fig. 6). Out of nine, five miRNAs (bdi-miR159, osa-
miR1869, ppt-miR901, ptc-miR169b-3p, and ptc-
miR482c-5p) were selected among F. culmorum respon-
sive miRNAs, and the remaining four miRNAs (cre-
miR1169-3p, osa-miR319a-3p, ssl-miR398, and zma-
miR164 g-3p) were selected from B. sorokiniana respon-
sive miRNAs. Expression profiles of most of the selected
miRNAs were comparable with the results of the micro-
arrays. Through miRNA-microarray measurement, ppt-
miR901 expression was found to be down-regulated upon
fungal-inoculation in the both cultivars, similarly, the
expression repression was also confirmed by qRT-PCR
analysis. Moreover, the expression of osa-miR319 was
detected as relatively high expressed in MI the BI in both
assays. Additionally, ssl-miR398 was expressed higher
upon inoculation in both cultivars which was validated by
qRT-PCR (Fig. 6).
The expected negative correlation between the expres-
sion pattern of the eight selected miRNAs and the target
transcripts were successfully confirmed by qRT-PCR
(Fig. 6). However, we detected an unexpected result only
for target transcript of miR398.
Discussion
In recent years, using different approaches, several studies
were conducted to understand functions of miRNAs in
stress tolerance in plants (Li et al. 2011; Zhou et al. 2012;
Chen et al. 2012). Being a high-throughput system,
Fig. 3 The heat map of B. sorokiniana responsive miRNAs in wheat miRNA-Microarray assay. Normalized signal intensities of 21 differentially
expressed miRNAs were illustrated in color scheme ranged from red to blue. The signal intensities decreases from red to blue color
Planta (2014) 240:1287–1298 1293
123
Author's personal copy

miRNA-arrays have an important and comprehensive role
in the identification of miRNAs responsive to different
stress factors. Although functions of plant miRNAs in
virus-plant interactions has been investigated, information
of miRNAs induced by fungal or bacterial pathogens is
limited (Zhao et al. 2012). In literature, there are only two
reports about fungal-responsive wheat miRNAs (Xin et al.
2010; Gupta et al. 2012). The current study aimed to
identify the wheat miRNAs involved in response to two
different fungi attacks. Comparing the results of two dis-
tinct wheat pathogens, F. culmorum and B. sorokiniana,
this study enabled us to identify how different wheat plants
responded to two different fungi.
Up to date, few numbers of miRNAs were arrayed in the
majority of the microarray based studies. Here, all the
known plant miRNAs were used to obtain more compre-
hensive data set. To our knowledge, this is the broadest
range miRNA-microarray study performed in wheat. A
previous study indicated the miRNAs can be transferred
from one species to another (Liang et al. 2013). Similarly, a
quarter of the probes originated from different species also
produced signals in our array study.
Here, two disease causing fungi were inoculated on two
wheat cultivars having different resistance levels. A num-
ber of miRNAs were found to be differentially regulated in
both assays. In which 66 and 21 miRNAs were observed as
responsive to F. culmorum and B. sorokiniana, respec-
tively. Two different miRNA expression profiles showed
that miRNA responses were significantly affected by
genetic background of the cultivars as well as fungal
agents.
Expression analysis revealed that fungal-inoculation
itself causes miRNA response in wheat. Heat-map illus-
trated a correlation between stressed and unstressed sam-
ples (Figs. 2, 3). Differentially expressed miRNAs detected
in B and M cultivars were clustered together, while the
signals obtained from control samples were grouped into a
different branch.
Comparing inoculated and control samples of resistant
cultivar, among 66 F. culmorum responsive miRNAs,
expression of 21 miRNAs was significantly reduced after
inoculation in the range between 20 and 251 folds. How-
ever, this pattern was not observed in susceptible cultivars
upon pathogen attack (Table 1). A similar expression
profile was also observed among the B. sorokiniana
responsive miRNAs. Among 21 differentially expressed
miRNAs, six miRNAs were down-regulated only in the
infected resistant B cultivar relative to the susceptible M
cultivar upon pathogen inoculation (Table 2). Ontology
analysis of the transcripts targeted by these miRNAs
revealed that some of the targets were included in disease
resistance processes (suppl. Tables S2, S3). Previous
studies were reported that a number of those target genes
were highly expressed in plants upon pathogen stress (DIaz
et al. 2002; Wagacha and Muthomi 2007; Xiao et al. 2013;
Gupta et al. 2014). The main reason behind the miRNA
regulation on fungal disease response in resistant cultivars
may be associated with the following hypothesis: Upon
fungal-inoculation, repression of these miRNAs causes
relatively higher accumulation of target transcripts. In the
end, resistant plant response against the pathogen attack.
Based on the ontology analysis, some of the miRNAs
have not been reported as stress responsive, while some
were known to be involved in fungal stress (Zhao et al.
2012). Roles of miR156, miR159, miR169, miR164,
miR319, and miR398 in response to fungal stress were
identified in pine, poplar, and wheat (Xin et al. 2010).
These results pointed that conserved miRNAs have vital
roles in pathogen defense mechanism in wheat. It was
reviewed that these miRNAs were also found to be
responsive to various abiotic stresses (Eldem et al. 2013).
However, the expression profiles of those miRNAs were
not fully consistent with each other indicating a species and
infection-specific miRNA regulation. Specifically, con-
flicting results were reported about the expression of
Table 2 B. sorokiniana pathogen-responsive miRNAs in two wheat
cultivars
miRNA_Name Fold change
BC vs BI MC vs MI BI vs MI
ath-miR771 1.70 1.17 -1.10
ath-miR869,1 129.71 1.04 -1.02
cre-miR1169-3p 274.31 10.44 -5.23
gma-miR4398 -1.45 -1.00 -1.79
gma-miR482e 2.02 -41.84 -34.90
mtr-miR2592 s 151.14 1.04 -1.02
mtr-miR2634 119.45 1.04 -1.02
osa-miR1427 -8.97 -32.38 -1.88
osa-miR1881 158.49 40.26 -2.56
osa-miR319a-3p,2-3p -9.51 -30.28 -1.66
osa-miR528-5p -2.18 -1.56 1.52
osa-miR5525 -2.03 -31.14 -1.06
osa-miR5800 -1.23 1.04 1.47
osa-miR6253 1.74 1.23 -1.78
ppt-miR894 1.37 1.58 1.16
ptc-miR169b-3p 102.68 1.04 -1.02
ptc-miR477d-3p 1.58 1.19 -1.17
sbi-miR169r-5p 2.12 2.80 1.51
ssl-miR398 -145.98 -29.78 4.64
vvi-miR3624-5p 58.70 1.04 -1.02
zma-miR164 g-3p -16.86 -49.34 1.31
Fold changes between the samples based on log2 were represented.
The signals were considered as differentially expressed with P B 0.05
and 1.45-fold change values
1294 Planta (2014) 240:1287–1298
123
Author's personal copy

miR169 family members against pathogen stress. miR169
was found to be down-regulated in soybean in response to
pathogen stress (Subramanian et al. 2008), while its
expression was induced in poplar against fungal attack
(Gupta et al. 2014). miR169 family members were signif-
icantly down-regulated in inoculated wheat cultivars.
Fig. 4 Gene ontology terms of biological function and molecular processes of F. culmorum responsive miRNA target transcripts in wheat
Fig. 5 Gene ontology terms of biological function and molecular processes of B. sorokiniana responsive miRNA target transcripts in wheat
Planta (2014) 240:1287–1298 1295
123
Author's personal copy
miR169 potentially targets a number of transcripts
including Pto kinase 1 which is a well-known resistance
gene (suppl. Table S2). Therefore, its expression level is
expected to be higher in inoculated samples compared to
controls. Since all these studies presented in silico miR169
target transcript prediction, it is not possible to identify
whether the same transcripts were targeted by miR169,
detailed target gene analysis is required to better under-
stand the exact reason of miR169 suppression upon fungal-
inoculation. Similarly, expression level of ath-miR869.1
was down-regulated in the infected wheat. The target
analysis showed that glucuronosyltransferase (GT)
(TC457607), which has a novel role in plant defense to
fungal pathogen, was regulated by ath-miR869.1 (suppl.
Table S2). GT was found to be participated in resistance
against deoxynivalenol (DON) which is a mycotoxin pro-
duced by fungal of the genus Fusarium (Bowles et al.
2005). Consequently, the higher level of GT in infected
plant is required to provide resistance.
To further evaluate miRNA regulation upon fungal
attack, we also analyzed miRNAs that showed opposite
expression in B and M cultivars. F. culmorum inoculation
caused downregulation of osa-159f, cre-1169-3p, and osa-
1869 miRNAs in BI, but induction of those in MI com-
pared to mock inoculations. Based on the target analysis,
those miRNAs were predicted to regulate SQUAMOSA
promoter binding proteins (SBP) family TFs, leucine-rich
repeat receptor, and copine family. It was reported that
expression of the target genes were induced upon pathogen
attack (Jambunathan et al. 2001; Kulcheski et al. 2011;
Eldem et al. 2013). Similarly, gma-miR482e was identified
as up-regulated in B. sorokiniana infected susceptible M
cultivar, but down-regulated in treated resistant B cultivar.
It was reported that a nucleotide-binding site leucine-rich
repeat (NBS-LRR) protein, targeted by miR482, was sup-
pressed in tomato when inoculated with Pseudomonas
syringe (DC3000) (Gupta et al. 2014). Furthermore, it was
identified that miR482bd-3p was down-regulated in a
susceptible soybean cultivar, which was treated with
Phakopsora pachyrhizi pathogen (Kulcheski et al. 2011).
In another study, expression of several ghr-miR482 family
members were reduced when infected with Verticillium
dahliae which caused upregulation of their NBS-LRR
targets (Zhu et al. 2013). Since a resistant cultivar needs to
synthesize more defense proteins than a susceptible culti-
var does, higher expression levels of such target genes in B
cultivar was expected.
On the other hand, three miRNAs (gma- miR5783, gma-
miR171 k-5p, ath- miR2933) were expressed more in F.
culmorum treated resistant B cultivar compared to its
Fig. 6 qRT-PCR validation of selected miRNAs and their target transcripts. The histograms show the relative values for the quantified
expressions of the miRNAs and the target genes. The analyses were performed as triplicates, and the error bars were indicated on each column
1296 Planta (2014) 240:1287–1298
123
Author's personal copy

control, while the opposite pattern was observed in sus-
ceptible M cultivar. A similar pattern was also observed for
gma-miR4398 upon B. sorokiniana infection. Previous
studies identified miR171 as adaptive responsive to stress
(Zhou et al. 2007; Liu et al. 2008). It was described that
conserved miR171 family was found to be down-regulated
in response to powdery mildew in wheat (Xin et al. 2010;
Khraiwesh et al. 2012). Although the two cultivars pre-
sented different expression profiles in our study, miR171
might take role in fungal pathogen stress. Peroxiredoxin
(Prx) (BE418197), predicted target of gma-miR5783,
functions as peroxidase which plays various functions
including response to pathogen infections (Rouhier et al.
2004). Recently, it was reported that WRKY transcription
factor was found to be the target of miR4398 (Zhang et al.
2014), and WRKYs in sunflower were identified as defense
response in biotic stress (Giacomelli et al. 2010).
Comparing the results of the two fungal-inoculation
miRNA expression assays, we found that eight miRNAs
were differentially regulated against both inoculations.
Besides, five of them showed the same expression profiles
in the two analyses. Although different pathogen caused
specific stress response, a common stress resistance
mechanism might also be considered based on this
outcome.
The overall analyses indicated that fungal-responsive
wheat miRNAs were differentially regulated in a pathogen
specific manner. In addition to inoculation-based miRNA
expression pattern, cultivar-related miRNA regulation
against pathogen response was also observed. This sug-
gested that such genetic variation among cultivars could be
amenable to selection for disease resistance in terms of
miRNAs expression upon fungal stresses. Some of the
miRNAs were identified as fungal-inoculation responsive
for the first time. Coupled with the genetic variation among
cultivars, understanding the genetic mechanisms of fungal
agents will enable us to better deal with the disease in
wheat.
Acknowledgments The study was funded by TUBITAK with Grant
Number: 113O546.
References
Bolstad BM, Irizarry RA, Astrand M, Speed TP (2003) A comparison
of normalization methods for high density oligonucleotide array
data based on variance and bias. Bioinformatics 19:185–193
Bowles D, Isayenkova J, Lim E-K, Poppenberger B (2005) Glyco-
syltransferases: managers of small molecules. Curr Opin Plant
Biol 8:254–263
Brenchley R, Spannagl M, Pfeifer M, Barker GL, D’Amore R, Allen
AM, McKenzie N, Kramer M et al (2012) Analysis of the bread
wheat genome using whole-genome shotgun sequencing. Nature
491:705–710
Chen L, Wang T, Zhao M, Tian Q, Zhang W-H (2012) Identification
of aluminum-responsive microRNAs in Medicago truncatula by
genome-wide high-throughput sequencing. Planta 235:375–386
Dıaz J, ten Have A, Van Kan JA (2002) The role of ethylene and
wound signaling in resistance of tomato to Botrytis cinerea. Plant
Physiol 129:1341–1351
Eldem V, Akcay UC, Ozhuner E, Bakır Y, Uranbey S, Unver T
(2012) Genome-wide identification of miRNAs responsive to
drought in peach (Prunus persica) by high-throughput deep
sequencing. PLoS One 7:e50298
Eldem V, Okay S, Unver T (2013) Plant microRNAs: new players in
functional genomics. Turk J Agric For 37:1–21
Feng H, Zhang Q, Li H, Wang X, Wang X, Duan X, Wang B, Kang Z
(2013a) vsiRNAs derived from the miRNA-generating sites of
pri-tae-miR159a based on the BSMV system play positive roles
in the wheat response to Puccinia striiformis f. sp. tritici through
the regulation of taMyb3 expression. Plant Physiol Biochem
68:90–95
Feng H, Zhang Q, Wang Q, Wang X, Liu J, Li M, Huang L, Kang Z
(2013b) Target of tae-miR408, a chemocyanin-like protein gene
(TaCLP1), plays positive roles in wheat response to high-
salinity, heavy cupric stress and stripe rust. Plant Mol Biol
83:433–443
Feng H, Duan X, Zhang Q, Li X, Wang B, Huang L, Wang X, Kang Z
(2014) The target gene of tae-miR164, a novel NAC transcrip-
tion factor from the NAM subfamily, negatively regulates
resistance of wheat to stripe rust. Mol Plant Pathol 15:284–296
Giacomelli JI, Ribichich KF, Dezar CA, Chan RL (2010) Expression
analyses indicate the involvement of sunflower WRKY tran-
scription factors in stress responses, and phylogenetic recon-
structions reveal the existence of a novel clade in the Asteraceae.
Plant Sci 178:398–410
Gupta OP, Permar V, Koundal V, Singh UD, Praveen S (2012)
MicroRNA regulated defense responses in Triticum aestivum L.
during Puccinia graminis f.sp. tritici infection. Mol Biol Rep
39:817–824
Gupta OP, Sharma P, Gupta RK, Sharma I (2014) Current status on
role of miRNAs during plant–fungus interaction. Physiol Mol
Plant Pathol 85:1–7
Han J, Kong ML, Xie H, Sun QP, Nan ZJ, Zhang QZ, Pan JB (2013)
Identification of miRNAs and their targets in wheat (Triticum
aestivum L.) by EST analysis. Genet Mol Res 12:3793–3805
Irizarry RA, Hobbs B, Collin F, Beazer-Barclay YD, Antonellis KJ,
Scherf U, Speed TP (2003) Exploration, normalization, and
summaries of high density oligonucleotide array probe level
data. Biostatistics 4:249–264
Jambunathan N, Siani JM, McNellis TW (2001) A humidity-sensitive
Arabidopsis copine mutant exhibits precocious cell death and
increased disease resistance. Plant Cell 13:2225–2240
Kammoun LG, Gargouri S, Barreau C, Richard-Forget F, Hajlaoui
MR (2010) Trichothecene chemotypes of Fusarium culmorum
infecting wheat in Tunisia. Int J Food Microbiol 140:84–89
Khraiwesh B, Zhu J-K, Zhu J (2012) Role of miRNAs and siRNAs in
biotic and abiotic stress responses of plants. Biochim Biophys
Acta 1819:137–148
Kulcheski F, de Oliveira L, Molina L, Almerao M, Rodrigues F,
Marcolino J, Barbosa J, Stolf-Moreira R, Nepomuceno A,
Marcelino-Guimaraes F, Abdelnoor R, Nascimento L, Cara-
zzolle M, Pereira G, Margis R (2011) Identification of novel
soybean microRNAs involved in abiotic and biotic stresses.
BMC Genomics 12:307
Li H, Dong Y, Yin H, Wang N, Yang J, Liu X, Wang Y, Wu J, Li X
(2011) Characterization of the stress associated microRNAs in
Glycine max by deep sequencing. BMC Plant Biol 11:170
Liang H, Zen K, Zhang J, Zhang C-Y, Chen X (2013) New roles for
microRNAs in cross-species communication. RNA Biol 10:1–4
Planta (2014) 240:1287–1298 1297
123
Author's personal copy

Liu H, Tian X, Li Y, Wu C, Zheng C (2008) Microarray-based
analysis of stress-regulated microRNAs in Arabidopsis thaliana.
RNA 14:836–843
Manickavelu A, Kawaura K, Oishi K, Shin IT, Kohara Y, Yahiaoui N,
Keller B, Abe R, Suzuki A, Nagayama T, Yano K, Ogihara Y
(2012) Comprehensive functional analyses of expressed
sequence tags in common wheat (Triticum aestivum). DNA
Res 19:165–177
Meng F, Liu H, Wang K, Liu L, Wang S, Zhao Y, Yin J, Li Y (2013)
Development-associated microRNAs in grains of wheat (Triti-
cum aestivum L.). BMC Plant Biol 13:140
Nizam S, Verma S, Singh K, Aggarwal R, Srivastava KD, Verma PK
(2012) High reliability transformation of the wheat pathogen
Bipolaris sorokiniana using Agrobacterium tumefaciens.
J Microbiol Meth 88:386–392
Pandey B, Gupta OP, Pandey DM, Sharma I, Sharma P (2013)
Identification of new stress-induced microRNA and their targets
in wheat using computational approach. Plant Signal Behav
8:e23932
Rouhier N, Gelhaye E, Gualberto JM, Jordy M-N, De Fay E,
Hirasawa M, Duplessis S, Lemaire SD, Frey P, Martin F (2004)
Poplar peroxiredoxin Q. A thioredoxin-linked chloroplast anti-
oxidant functional in pathogen defense. Plant Physiol
134:1027–1038
Scherm B, Balmas V, Spanu F, Pani G, Delogu G, Pasquali M,
Migheli Q (2013) Fusarium culmorum: causal agent of foot and
root rot and head blight on wheat. Mol Plant Pathol 14:323–341
Schmittgen TD, Livak KJ (2008) Analyzing real-time PCR data by
the comparative CT method. Nat Protoc 3:1101–1108
Subramanian S, Fu Y, Sunkar R, Barbazuk WB, Zhu J-K, Yu O
(2008) Novel and nodulation-regulated microRNAs in soybean
roots. BMC Genomics 9:160
Tanaka T, Kobayashi F, Joshi GP, Onuki R, Sakai H, Kanamori H,
Wu J, Simkova H, Nasuda S, Endo TR, Hayakawa K, Dolezel J,
Ogihara Y, Itoh T, Matsumoto T, Handa H (2013) Next-
generation survey sequencing and the molecular organization of
wheat chromosome 6B. DNA Res 21:103–114
Turktas M, Inal B, Okay S, Erkilic E, Dundar E, Hernandez P, Dorado
G, Unver T (2013) Nutrition metabolism plays an important role
in the alternate bearing of the olive tree (Olea europaea L.).
PLoS One 8:e59876
Untergasser A, Cutcutache I, Koressaar T, Ye J, Faircloth BC, Remm
M, Rozen SG (2012) Primer3—new capabilities and interfaces.
Nucleic Acids Res 40:e115
Unver T, Bakar M, Shearman RC, Budak H (2010a) Genome-wide
profiling and analysis of Festuca arundinacea miRNAs and
transcriptomes in response to foliar glyphosate application. Mol
Genet Genomics 283:397–413
Unver T, Parmaksız I, Dundar E (2010b) Identification of conserved
micro-RNAs and their target transcripts in opium poppy
(Papaver somniferum L.). Plant Cell Rep 29:757–769
Varkonyi-Gasic E, Wu R, Wood M, Walton EF, Hellens RP (2007)
Protocol: a highly sensitive RT-PCR method for detection and
quantification of microRNAs. Plant Methods 3:12
Wagacha JM, Muthomi JW (2007) Fusarium culmorum: infection
process, mechanisms of mycotoxin production and their role in
pathogenesis in wheat. Crop Prot 26:877–885
Wang B, Sun YF, Song N, Wang XJ, Feng H, Huang LL, Kang ZS
(2013) Identification of UV-B-induced microRNAs in wheat.
Genet Mol Res 12:4213–4221
Xiao J, Jin X, Jia X, Wang H, Cao A, Zhao W, Pei H, Xue Z, He L,
Chen Q, Wang X (2013) Transcriptome-based discovery of
pathways and genes related to resistance against Fusarium head
blight in wheat landrace Wangshuibai. BMC Genom 14:197
Xin M, Wang Y, Yao Y, Xie C, Peng H, Ni Z, Sun Q (2010) Diverse
set of microRNAs are responsive to powdery mildew infection
and heat stress in wheat (Triticum aestivum L.). BMC Plant Biol
10:123
Yanik H, Turktas M, Dundar E, Hernandez P, Dorado G, Unver T
(2013) Genome-wide identification of alternate bearing-associ-
ated microRNAs (miRNAs) in olive (Olea europaea L.). BMC
Plant Biol 13:10
Zhang Z, Liu X, Wang X, Zhou M, Zhou X, Ye X, Wei X (2012) An
R2R3 MYB transcription factor in wheat, TaPIMP1, mediates
host resistance to Bipolaris sorokiniana and drought stresses
through regulation of defense- and stress-related genes. New
Phytol 196:1155–1170
Zhang N, Yang J, Wang Z, Wen Y, Wang J, He W, Liu B, Si H, Wang
D (2014) Identification of novel and conserved microRNAs
related to drought stress in potato by deep sequencing. PloS One
9(4):e95489. doi:10.1371/journal.pone.0095489
Zhao J-P, Jiang X-L, Zhang B-Y, Su X-H (2012) Involvement of
microRNA-mediated gene expression regulation in the patho-
logical development of stem canker disease in Populus tricho-
carpa. PloS One 7(9):e44968. doi:10.1371/journal.pone.0044968
Zhou X, Wang G, Zhang W (2007) UV-B responsive microRNA
genes in Arabidopsis thaliana. Mol Syst Biol 3:103. doi:10.1038/
msb4100143
Zhou ZS, Zeng HQ, Liu ZP, Yang ZM (2012) Genome-wide
identification of Medicago truncatula microRNAs and their
targets reveals their differential regulation by heavy metal. Plant
Cell Environ 35:86–99
Zhu Q-H, Fan L, Liu Y, Xu H, Llewellyn D, Wilson I (2013) miR482
regulation of NBS-LRR defense genes during fungal pathogen
infection in cotton. PLoS One 8:e84390
1298 Planta (2014) 240:1287–1298
123
Author's personal copy




















