
Functional magnetic resonance mapping of intracerebroventricular infusion of a neuroactive peptide...
10
Journal of Neuroscience Methods 142 (2005) 115–124 Functional magnetic resonance mapping of intracerebroventricular infusion of a neuroactive peptide in the anaesthetised rat Alessandro Gozzi a,∗ , Adam J. Schwarz a , Torsten Reese a , Valerio Crestan b , Simone Bertani b , Giuliano Turrini b , Mauro Corsi c , Angelo Bifone a a Department of Neuroimaging, Psychiatry Centre of Excellence in Drug Discovery, GlaxoSmithKline Medicines Research Centre, Via Fleming 4, 37135 Verona, Italy b Laboratory Animal Sciences, Psychiatry Centre of Excellence in Drug Discovery, GlaxoSmithKline Medicines Research Centre, Via Fleming 4, 37135 Verona, Italy c Department of Neuropharmacology, Psychiatry Centre of Excellence in Drug Discovery, GlaxoSmithKline Medicines Research Centre, Via Fleming 4, 37135 Verona, Italy Received 14 April 2004; received in revised form 31 July 2004; accepted 11 August 2004 Abstract Pharmacological magnetic resonance imaging (phMRI) methods map the cerebral haemodynamic response to challenge with psychotropic agents as a surrogate for drug-induced changes in brain activity. However, many neuroactive compounds present low blood–brain barrier penetration and thus systemic administration may result in insufficient brain concentration. Intracerebroventricular (ICV) administration has been long used as an effective way of bypassing the blood–brain barrier in studies with non-brain-penetrant compounds, such as neuropeptides. In order to extend the range of pharmacological substances accessible to phMRI, we have developed methods to map relative cerebral blood volume (rCBV) changes induced by in situ ICV administration of neuroactive agents in the anaesthetised rat. We have applied this method to study for the first time the phMRI response to central administration of a neuropeptide, the metabolically stable and potent NK1 receptor agonist GR-73632. ICV administration of 4.2 pmol of GR-73632 produced a rapid onset and sustained rCBV increase in several brain structures, such as the amygdala, the caudate putamen and the cortex. These results demonstrate the feasibility of phMRI as a tool to study the functional correlates of brain activity induced by central administration of neuroactive agents. © 2004 Elsevier B.V. All rights reserved. Keywords: phMRI; GR-73632; NK1 receptor agonist; Rat; ICV; rCBV 1. Introduction Functional magnetic resonance imaging (fMRI) tech- niques map the spatial and temporal patterns of haemody- namic response to local changes in neuronal activity (Morris, 1999). A variant known as pharmacological MRI (phMRI) has recently been applied to map the functional response to systemic administration of various pharmacological agents in animal studies, including amphetamine (Chen et al., 1997; Silva et al., 1995), cocaine (Chen et al., 1999; Marota ∗ Corresponding author. Tel.: +39 045 921 9233; fax: +39 045 921 8073. E-mail address: [email protected] (A. Gozzi). et al., 2000), apomorphine (Chen et al., 1996; Zhang et al., 2000), bicuculline (Reese et al., 2000), nicotine (Stein et al., 1998), heroin (Xu et al., 2000), and serotonin re- ceptor ligands (Houston et al., 2001; Scanley et al., 2001). However, many neuroactive agents, including most peptidic neuromodulators, present low blood–brain barrier penetra- tion and their systemic administration may result in an in- sufficient brain penetration (Banks and Kastin, 1996; Von Bohlen and Dermietzel, 2002). Moreover, many physiolog- ical processes regulated by neuropeptides are also mediated by widely distributed peripheral receptors. This limits the use of systemic administration as a means of assessing the central activity of neuropeptides, given the difficulty of sep- 0165-0270/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.jneumeth.2004.08.013
Transcript of Functional magnetic resonance mapping of intracerebroventricular infusion of a neuroactive peptide...

Journal of Neuroscience Methods 142 (2005) 115–124
Functional magnetic resonance mapping of intracerebroventricularinfusion of a neuroactive peptide in the anaesthetised rat
Alessandro Gozzia,∗, Adam J. Schwarza, Torsten Reesea, Valerio Crestanb,Simone Bertanib, Giuliano Turrinib, Mauro Corsic, Angelo Bifonea
a Department of Neuroimaging, Psychiatry Centre of Excellence in Drug Discovery, GlaxoSmithKline Medicines Research Centre,Via Fleming 4, 37135 Verona, Italy
b Laboratory Animal Sciences, Psychiatry Centre of Excellence in Drug Discovery, GlaxoSmithKline Medicines Research Centre,Via Fleming 4, 37135 Verona, Italy
c Department of Neuropharmacology, Psychiatry Centre of Excellence in Drug Discovery, GlaxoSmithKline Medicines Research Centre,Via Fleming 4, 37135 Verona, Italy
Received 14 April 2004; received in revised form 31 July 2004; accepted 11 August 2004
Abstract
sychotropica rain barrierp ration hasb uropeptides.I rebral bloodv method tos tor agonistG tures, sucha e functionalc©
K
1
nn1hsiS
g
-1tidicetra-in-n-
diatedtheg the
ep-
0d
Pharmacological magnetic resonance imaging (phMRI) methods map the cerebral haemodynamic response to challenge with pgents as a surrogate for drug-induced changes in brain activity. However, many neuroactive compounds present low blood–benetration and thus systemic administration may result in insufficient brain concentration. Intracerebroventricular (ICV) administeen long used as an effective way of bypassing the blood–brain barrier in studies with non-brain-penetrant compounds, such as ne
n order to extend the range of pharmacological substances accessible to phMRI, we have developed methods to map relative ceolume (rCBV) changes induced by in situ ICV administration of neuroactive agents in the anaesthetised rat. We have applied thistudy for the first time the phMRI response to central administration of a neuropeptide, the metabolically stable and potent NK1 recepR-73632. ICV administration of 4.2 pmol of GR-73632 produced a rapid onset and sustained rCBV increase in several brain strucs the amygdala, the caudate putamen and the cortex. These results demonstrate the feasibility of phMRI as a tool to study thorrelates of brain activity induced by central administration of neuroactive agents.2004 Elsevier B.V. All rights reserved.
eywords:phMRI; GR-73632; NK1 receptor agonist; Rat; ICV; rCBV
. Introduction
Functional magnetic resonance imaging (fMRI) tech-iques map the spatial and temporal patterns of haemody-amic response to local changes in neuronal activity (Morris,999). A variant known as pharmacological MRI (phMRI)as recently been applied to map the functional response toystemic administration of various pharmacological agentsn animal studies, including amphetamine (Chen et al., 1997;ilva et al., 1995), cocaine (Chen et al., 1999; Marota
∗ Corresponding author. Tel.: +39 045 921 9233; fax: +39 045 921 8073.E-mail address:[email protected] (A. Gozzi).
et al., 2000), apomorphine (Chen et al., 1996; Zhanet al., 2000), bicuculline (Reese et al., 2000), nicotine (Steinet al., 1998), heroin (Xu et al., 2000), and serotonin receptor ligands (Houston et al., 2001; Scanley et al., 200).However, many neuroactive agents, including most pepneuromodulators, present low blood–brain barrier pention and their systemic administration may result in ansufficient brain penetration (Banks and Kastin, 1996; VoBohlen and Dermietzel, 2002). Moreover, many physiological processes regulated by neuropeptides are also meby widely distributed peripheral receptors. This limitsuse of systemic administration as a means of assessincentral activity of neuropeptides, given the difficulty of s
165-0270/$ – see front matter © 2004 Elsevier B.V. All rights reserved.oi:10.1016/j.jneumeth.2004.08.013

116 A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124
arating central neuromodulatory effects from those gener-ated by a direct action at peripheral receptor sites. As aconsequence, functional neuroimaging of the specific brainstructures on which neuropeptides exert their action remainselusive.
To obtain a rapid and efficient distribution of compoundswithin the central nervous system (CNS), direct injectioninto the cerebrospinal fluid (CSF) of the brain ventriclesmay be performed (Ghersi-Egea et al., 1996; Proescholdtet al., 2000). Intracerebroventricular (ICV) administrationhas been long used as an effective way of bypassing theblood–brain barrier, while minimising the problems of pres-sure damage and osmotic imbalance that may occur afterfocal intracerebral injection (Greenshaw, 1998). These fea-tures make ICV infusion a powerful technique that has pavedthe way to the investigation of the role of neuropeptidesin vivo.
In order to extend the range of pharmacological substancesaccessible to phMRI, we have developed methods to map rel-ative cerebral blood volume (rCBV) changes induced by insitu ICV administration of neuroactive agents in the anaes-thetised rat, permitting phMRI studies of compounds withpoor CNS penetration such as neuropeptides. A potent and se-lective NK1 receptor agonist, GR-736321 (Hagan et al., 1991;Rupniak et al., 2003) was used in combination with a rCBVp R-7 sub-s milyi esisi2 uesh ouralse ichs is im-p withi n anda
2
2
Ital-i Pro-t nimalc Prin-c 23,r
0 g)( nty-f uid
distribution and anatomical changes induced by the ICVsurgery, and 13 were used to investigate GR-73632 haemo-dynamic response (8 injected with GR-73632, 5 with PBScontrol, see below). Animals had free access to standard ratchow and tap water and were housed in groups of five insolid bottom cages with sawdust litter. Room temperature(20–22◦C), relative humidity (45–65%) and dark–light cy-cles (12 h each, lights on at 06:00 h) were automatically con-trolled. After arrival rats were allowed to acclimatise for atleast 5 days.
The procedure for the ICV cannulation is similar to thatreported byGreenshaw (1998). Plastic, MR compatible guidecannulae were used instead of the standard stainless steelones. Metallic inserts can in fact induce severe artefacts in MRimages and can absorb and shield radiofrequency radiationused to excite the nuclear spin system for NMR detection(Schenck, 1996).
On the day of surgery, rats were pre-anaesthetisedwith an intramuscular injection of 50�g/kg medetomidine(Dormitor®, Pfizer, Italy). This was followed after 10 min byan intramuscular injection of 10 mg/kg each of tiletamine andzolazepam (Zoletil 100®, Virbac, Italy) to induce anaesthesia.Subsequently, animals were given a subcutaneous injectionof benzilpennicillin (12500 IU/rat) and dihydrostreptomycin(5 mg/rat, Rubrocillina Veterinaria®, Nuova ICC, Italy) top werea /kg,R lec-t iumc o-d
d se-c longt ovedb entlys rileg wasd gen augen Inc.,U im-pa ma(d st dhe-s tha Co.,U auges SA),f OneI erei f de-b sur-g alsw ibed
hMRI protocol to prove the feasibility of the method. G3632 is a metabolically stable synthetic analogue oftance P, a neuropeptide belonging to the tachykinin famplicated in the regulation of affective behaviour and emn the brain, as well as nociception in the spinal cord (Mantyh,002). ICV administration of substance P and its analogas been experimentally used in a number of behavitudies to induce anxiety-related behaviour in rodents (Celliert al., 1999). In particular, the NK1 receptor system, on whubstance P preferentially acts as endogenous ligand,licated in numerous physiological functions associated
mportant psychiatric disease states such as depressionxiety.
. Materials and methods
.1. Implantation of the ICV cannula
All experiments were carried out in accordance withan regulations governing animal welfare and protection.ocols were also reviewed and consented to by a local aare committee, in accordance with the guidelines of theiples of Laboratory Animal Care (NIH publication 86–evised 1985).
A total of 37 male Sprague-Dawley rats (250–30Charles River, Como, Italy) were used in this study. Tweour animals were employed to study intraventricular fl
1 H2N-(CH2)4-CO-Phe-Pro-NMe-Leu-Met-NH2, Mw 766.0.
revent the development of bacterial infections; animalslso treated with the anti-inflammatory carprofen (5 mgimadyl®, Pfizer, Italy). The head was shaved with an e
ric clipper and the skin was disinfected with benzalconhloride (Citrosil®, Manettei & Roberts, Italy) and 10% iine (Betadine®, Viatris, Italy).
The anaesthetised animals were then mounted anured in a stereotaxic frame. An incision was made ahe midline to expose the skull, and periosteum was remy using cotton swabs. The skull surface was then gcraped in order to identify Bregma, and dried with steauze and cotton tips. A burr hole (diameter 0.9 mm)rilled with a handheld micro-drill. A blunted-tip 26 Gaueedle was used to perforate the Dura Mater. A 22 Gon-metallic guide cannula (C313GT/Spc, Plastic OneSA, outer diameter = 0.69 mm; length = 6 mm) waslanted using the following stereotaxic co-ordinates (Paxinosnd Watson, 1998): 5.6 mm posterior distance from bregAP), 1.5 mm lateral distance from the midline (L), −3.8 mmepth from the level of the skull (V). The guide cannula wa
hen secured to the skull with a few drops of tissue aive (Vetbond® 3M, Italy) which were then covered wiuniform layer of dental cement (Stratfrod–CooksonSA). One animal was cannulated with a standard 22 Gtainless steel guide cannula (C311G Plastic One Inc., Uor comparison. Stainless-steel stylets (C313DC, Plasticnc., USA) 0.5 mm longer than the indwelling cannulae, wnserted into the guide cannulae to keep them free oris and the skin was then sutured closed with 2/0 silkical thread. Twenty-four hours after surgery the animere given an additional injection of antibiotic as descr

A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124 117
above. After recovery rats were singly housed in flat wirelids without feed hopper to avoid damage to the surgicalimplant. On the following days rats were handled to pre-vent the development of hyperactivity and aggression. Oc-casionally, stylets were removed and immediately put backin place to avoid the development of tissue adhesions to thedevice.
During recovery, animals exhibited normal eating anddrinking behaviour. None of the ICV devices were dislodgedor damaged by rat behaviour. The skin healed within 4–5 dayswith minor inflammation. In cases of persistent inflammationor in presence of signs of infection, animals were subjectedto additional anti-inflammatory and antibiotic therapy. Noneof the animals showed any behavioural or physical signs ofdiscomfort.
2.2. Animal preparation for phMRI
A recovery from ICV skull surgery of at least 7 days wasallowed to exclude carry-over effects of the injectable anaes-thetic on cerebrovascular reactivity. On the day of MR exper-iment, rats were anaesthetised with 3% halothane (Wyeth,Italy) in a 30:70% O2:N2 gas mixture, tracheotomised andartificially ventilated with a mechanical respirator (Inspira,Oxford, UK). Upon tracheotomy and throughout surgeryt orala wasa( in-f thea ereo-t ingt tylusw oin-j bingw sm ml endo ithw eat weent in-j r toi
s forep siono oxy-g ).F tersw aluesw9 nceb eng were
monitored continually. Heart rate was calculated from arterialpressure pulses by the physiological data acquisition software(Biopac Systems Inc., Goweta, CA, USA). During the imageacquisition the maintenance level of anaesthesia was set to0.8%.
2.3. phMRI experiment
MRI data were acquired using a Bruker Avance 4.7T sys-tem. The animals were positioned prone in the manufacturer’sholding support and a “Rat Brain” curved quadrature receivecoil (Bruker, Ettlingen, Germany) was mounted on top ofthe animal skull and fixed to the animal holder. A 20 mm×12 mm opening in the center of the surface coil allowed ac-cess to the cannula protruding out of the animal skull. Theanimal holder was then fitted into a 72 mm birdcage resonator(Bruker, Ettlingen, Germany) which was used for radiofre-quency transmit only. Both coils are standard componentsprovided by the manufacturer.
The phMRI experiment was performed acquiringT2-weighted functional images using the RARE sequence, soas to minimise sensitivity to large blood vessels, and in-homogeneities of the static magnetic field (Ogawa et al.,1993; Weisskoff et al., 1994). High-resolutionT2-weightedanatomical images were acquired using the RARE sequencew et uiredi pos-t eri-m AREs41 trew or-d the8 e oft+ -t oodp d in-t , tos mind em,W e of4 anI 7.4,7 tionr xi-m pre-v ac-t ,1 isedw mento cting3 h,I
he anaesthetic level was maintained at 2%. Left femrtery and vein were cannulated and animal paralysischieved by a 0.25 mg/kg intravenous bolus ofd-tubocurarineSigma–Aldrich, Milano, Italy) followed by a continuoususion of 0.25 mg/kg/h (in 25 IU/ml of heparin) throughrtery. The animal was then moved into a customised st
actic holder (Bruker, Ettlingen, Germany). Before movhe animal into the magnet room, the stainless steel sas removed from the ICV guide cannula and the micr
ection system was assembled. A 70-cm piece of PE50 tuas connected with a 50�l syringe (Hamilton, USA). A glasicroinjection unit (C313/FS, Plastic One Inc., USA) 1 m
onger than the guide cannula was fitted into the otherf PE50 tubing. The microinjection system was filled water and than used to aspirate 7�l of the drug solution to bdministered. A small air bubble (corresponding to 3�l on
he syringe scale) was used to provide an interface bethe two liquids to prevent dilution of the drug. The microection unit was then fitted into the guide cannula prionstalling the animal into the magnet.
In order to monitor arterial blood gases, 200�l bloodamples were withdrawn from the arterial catheter beositioning the animal in the magnet and at the concluf the experiment and analysed for partial pressure ofen, carbon dioxide and pH (AVL, Roswell, GA, USAollowing the former measurement, ventilation parameere adjusted in order to keep the arterial blood gases vithin physiological range (30 <pCO2 < 53 mmHg;pO2 >0 mmHg). There was no systematic or significant differeetweenpCO2 or pO2 values pre- and post-MRI, or betweroups. Rectal temperature and arterial blood pressure
ith RARE factor 32, matrix 256× 256, FOV 40 mm, slichickness 1 mm, 32 contiguous slices. These were acqn the coronal plane, centred 10 mm caudal from theerior edge of the olfactory bulb. The time series expent comprised 256 sequential time frames using the R
equence with RARE factor 32, matrix 128× 128, FOV0 mm, slice thickness 2 mm, 8 contiguous slices, TEeff =10 ms, TR = 2700 ms,δt = 20 s. The geometrical cenas set identical to that of the anatomical images, iner that the central 16 slices of the former align withslices of the time series. The rostro-caudal coverag
he time series acquisition was approximately fromzbregma4 mm (anterior edge of slice 1) tozbregma− 12 mm (pos
erior edge of slice 8). A 2.67 ml/kg dose of Endorem blool contrast agent (Guerbet, France) was administere
ravenously (IV) following five reference image framesensitise the acquisition to changes in CBV. After a 30elay the ICV challenge was delivered. GR-73632 (Bacheil am Rhein, Germany) was administered at a dos
.2 pmol/rat (7�l/rat,n= 8). Five control animals receivedCV injection of vehicle (Phosphate Buffered Saline pH�l/rat). Compounds were infused manually at an injec
ate of 7�l/min. rCBV changes were tracked out to approately 32 min post-injection. The dose of GR-73632 was
iously reported to be effective at stimulating locomotorivity upon focal cerebral administration (Elliott et al., 1991992). At the end of the experiment rats were euthanith an overdose of anaesthetic agent. The correct placef the ICV cannula was assessed post-mortem by inje�l of a colouring agent (Methylen Blue, Sigma–Aldric
taly).
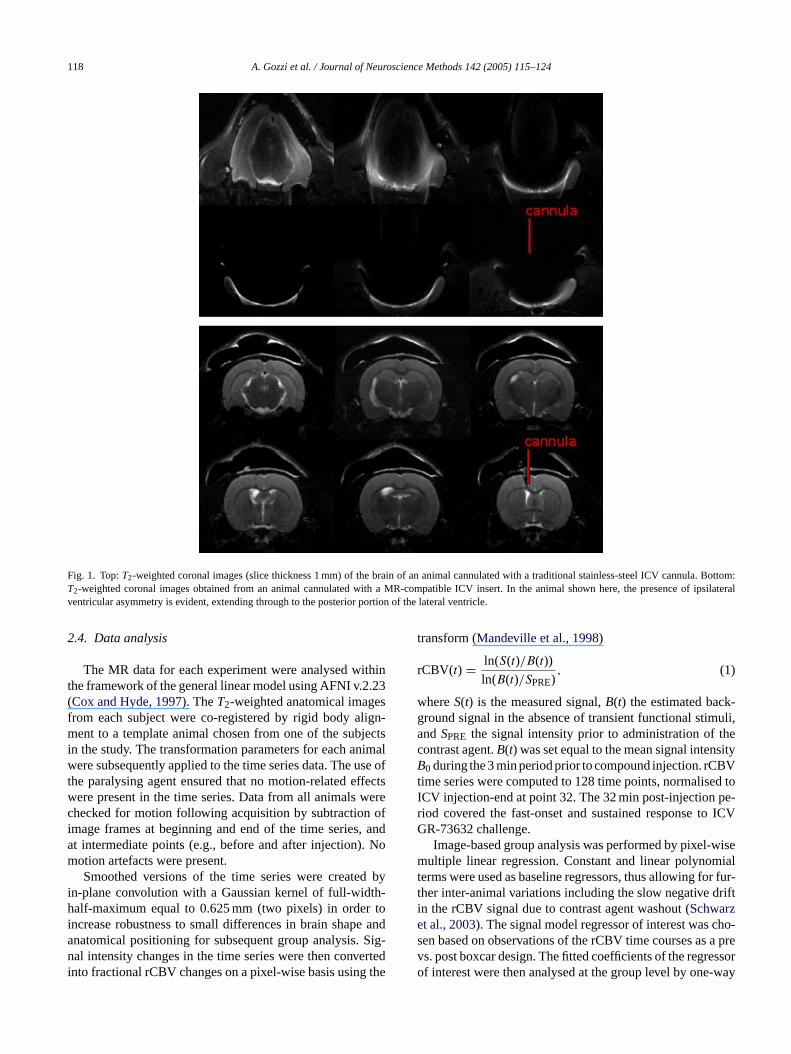
118 A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124
Fig. 1. Top:T2-weighted coronal images (slice thickness 1 mm) of the brain of an animal cannulated with a traditional stainless-steel ICV cannula. Bottom:T2-weighted coronal images obtained from an animal cannulated with a MR-compatible ICV insert. In the animal shown here, the presence of ipsilateralventricular asymmetry is evident, extending through to the posterior portion of the lateral ventricle.
2.4. Data analysis
The MR data for each experiment were analysed withinthe framework of the general linear model using AFNI v.2.23(Cox and Hyde, 1997). TheT2-weighted anatomical imagesfrom each subject were co-registered by rigid body align-ment to a template animal chosen from one of the subjectsin the study. The transformation parameters for each animalwere subsequently applied to the time series data. The use ofthe paralysing agent ensured that no motion-related effectswere present in the time series. Data from all animals werechecked for motion following acquisition by subtraction ofimage frames at beginning and end of the time series, andat intermediate points (e.g., before and after injection). Nomotion artefacts were present.
Smoothed versions of the time series were created byin-plane convolution with a Gaussian kernel of full-width-half-maximum equal to 0.625 mm (two pixels) in order toincrease robustness to small differences in brain shape andanatomical positioning for subsequent group analysis. Sig-nal intensity changes in the time series were then convertedinto fractional rCBV changes on a pixel-wise basis using the
transform (Mandeville et al., 1998)
rCBV(t) = ln(S(t)/B(t))
ln(B(t)/SPRE), (1)
whereS(t) is the measured signal,B(t) the estimated back-ground signal in the absence of transient functional stimuli,andSPRE the signal intensity prior to administration of thecontrast agent.B(t) was set equal to the mean signal intensityB0 during the 3 min period prior to compound injection. rCBVtime series were computed to 128 time points, normalised toICV injection-end at point 32. The 32 min post-injection pe-riod covered the fast-onset and sustained response to ICVGR-73632 challenge.
Image-based group analysis was performed by pixel-wisemultiple linear regression. Constant and linear polynomialterms were used as baseline regressors, thus allowing for fur-ther inter-animal variations including the slow negative driftin the rCBV signal due to contrast agent washout (Schwarzet al., 2003). The signal model regressor of interest was cho-sen based on observations of the rCBV time courses as a prevs. post boxcar design. The fitted coefficients of the regressorof interest were then analysed at the group level by one-way
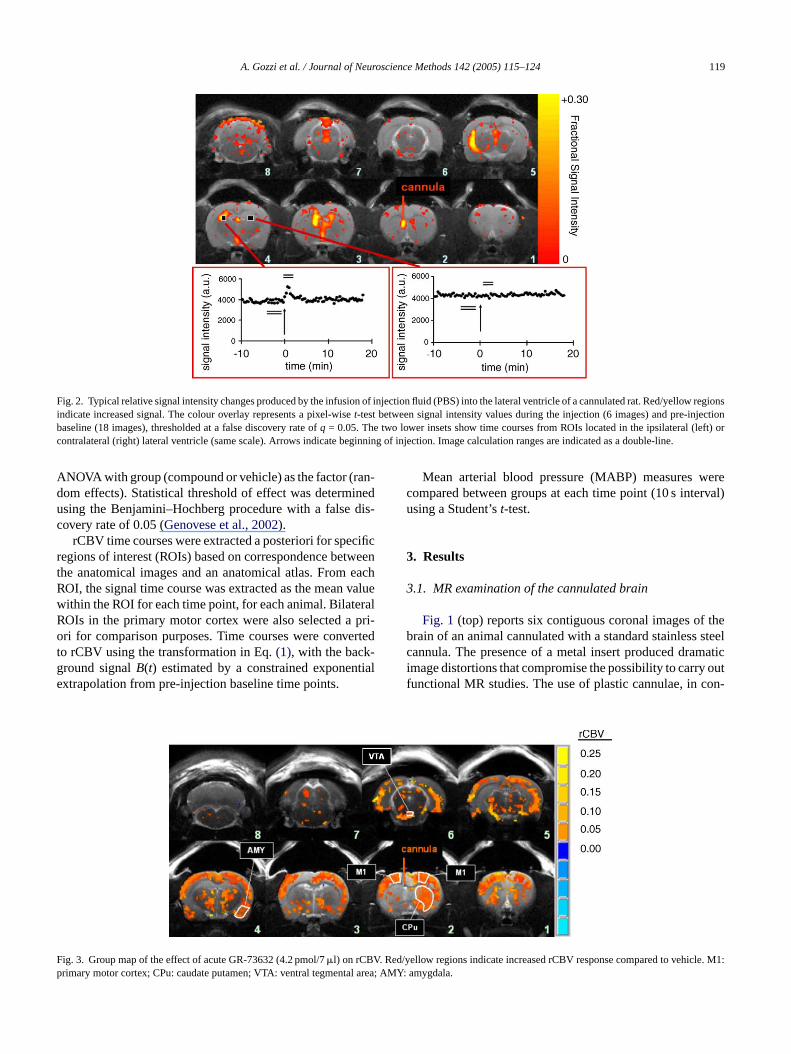
A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124 119
Fig. 2. Typical relative signal intensity changes produced by the infusion of injection fluid (PBS) into the lateral ventricle of a cannulated rat. Red/yellow regionsindicate increased signal. The colour overlay represents a pixel-wiset-test between signal intensity values during the injection (6 images) and pre-injectionbaseline (18 images), thresholded at a false discovery rate ofq = 0.05. The two lower insets show time courses from ROIs located in the ipsilateral (left) orcontralateral (right) lateral ventricle (same scale). Arrows indicate beginning of injection. Image calculation ranges are indicated as a double-line.
ANOVA with group (compound or vehicle) as the factor (ran-dom effects). Statistical threshold of effect was determinedusing the Benjamini–Hochberg procedure with a false dis-covery rate of 0.05 (Genovese et al., 2002).
rCBV time courses were extracted a posteriori for specificregions of interest (ROIs) based on correspondence betweenthe anatomical images and an anatomical atlas. From eachROI, the signal time course was extracted as the mean valuewithin the ROI for each time point, for each animal. BilateralROIs in the primary motor cortex were also selected a pri-ori for comparison purposes. Time courses were convertedto rCBV using the transformation in Eq.(1), with the back-ground signalB(t) estimated by a constrained exponentialextrapolation from pre-injection baseline time points.
F . Red/ e. M1:p l area;
Mean arterial blood pressure (MABP) measures werecompared between groups at each time point (10 s interval)using a Student’st-test.
3. Results
3.1. MR examination of the cannulated brain
Fig. 1 (top) reports six contiguous coronal images of thebrain of an animal cannulated with a standard stainless steelcannula. The presence of a metal insert produced dramaticimage distortions that compromise the possibility to carry outfunctional MR studies. The use of plastic cannulae, in con-
ig. 3. Group map of the effect of acute GR-73632 (4.2 pmol/7�l) on rCBVrimary motor cortex; CPu: caudate putamen; VTA: ventral tegmenta
yellow regions indicate increased rCBV response compared to vehiclAMY: amygdala.

120 A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124
trast, did not appear to affect the quality of images (Fig. 1,bottom). In 17 out of 37 animals, high-resolution MR anatom-ical images of cannulated brains highlighted visible asymme-try in the ipsilateral ventricles, which appeared enlarged andoversized. Asymmetric areas showed a bright, oedematousappearance, and extended throughout the ipsilateral cannu-lated ventricle (Fig. 1, bottom). In some instances, a dark areawas observed within the cannulated ventricle, proximate tothe tip of the cannula, probably the result of an air bubble.Cannulae appeared correctly placed into the left lateral ven-tricle in all the animals examined. This observation was con-firmed by the post-mortem analysis of the ventricles with acolouring agent.
3.2. Effect of ICV injection on the fluid dynamics in theventricles
Upon ICV infusion of vehicle or drug solutions a strong,focal increase in signal intensity was observed in the cannu-lated ventricle, probably due to the injection of fresh fluid(Fig. 2, P < 0.01). Local signal changes were also detectedin other ventricular areas of the brain, but the extent andthe profile of these signal fluctuations was highly variable.Typically, stronger alterations were seen ipsilaterally (Fig. 2,slices 2–5) suggesting an initial inhomogeneous distributiono l de-c theC
e re-g ven-tt thes cale hows e in-j fromt rrela-t BVr
3
in-c puta-m rea( –4),t slice4 rticals weda s. Al ctedi as-s entsi
n ins area,
the amygdala, the striatum, and the motor cortex (Fig. 4).Activation was sustained throughout the 32 min monitoredafter injection. The amplitude and profile of the rCBV timecourses in the motor cortex did not show substantial differ-ences between ipsilateral and contralateral areas (Fig. 4a). Ahigh degree of similarity between ipsilateral and contralat-eral time profile, was also present in the amygdala and othersub-cortical structures (data not shown). ICV infusion of PBS(control) produced no significant changes in rCBV. A slightlate increase in rCBV in M1 motor cortex of control ani-mals (Fig. 4a) was observed, probably the result of imperfectdetrending at late time points.
Neither GR-73632 nor vehicle (PBS) affected the meanarterial blood pressure. Occasionally, a short-lived hypoten-sion was observed upon the administration of the solutionin both groups of treatment. Statistical comparison of bloodpressure traces showed no significant compound-related ef-fects (Student’st-test,P > 0.1, all time points).
4. Discussion
The present study was carried out in order to demonstratethe feasibility of phMRI studies of centrally administeredneuroactive compounds. Central administration was achievedv iblec ingd NS,c ody-n r lowb smento giver tem-i
igh-l sity,c dom-i ex-a y ip-s ricle,m noto iculara 32 ini hesea lim-i rtainp gnif-i ymaw t thatt d toa ovas-c fort maco-l
inl is is
f the solution in the ventricles. Subsequently the signaayed, reflecting the equilibration of the excess fluid withSF in the interconnected ventricles.Strong spatial co-localisation was found between th
ions with the highest intraventricular changes, and thericular enlargements induced by the cannulation (Fig. 1, bot-om, andFig. 2). In three animals, after an initial increase,ignal did not go back to base-line level, indicating a lontrapment of the injected liquid. One animal did not signal alterations in any of the ventricles, a sign that thection was not successful, and was therefore excludedhe study. None of these patterns showed significant coion with the group of treatment or the extent of the rCesponse to the ICV challenge.
.3. phMRI of GR-73632
GR-73632 (4.2 pmol) produced significant rCBVreases in several brain structures including the caudateen (P < 0.01,Fig. 3, slices 1–2), the ventral tegmental a
slices 5–6), the lateral hypothalamic nucleus (slices 3he ventral subiculum and (slice 6) and the amygdala (). Significant changes were also detected in several cotructures (slices 1–6). The overall activation pattern shofairly bilateral distribution among the two hemisphere
oss of bilaterality in the rCBV response was only deten brain regions proximal to the cannulated ventricle orociated with the presence of the ventricular enlargemnduced by the cannulation (Figs. 2 and 3– slices 2 and 5).
rCBV increased rapidly upon GR-73632 administratioeveral brain structures, including the ventral tegmental
ia the insertion of a permanent indwelling MR compatannula into the lateral ventricle. This technique, allowirect administration of psychotropic agents into the Can be applied to map the spatial and temporal haemamic response produced by neuropeptides and otherain-penetrant compounds. It also permits the assesf centrally mediated effects of compounds that wouldise to confounding peripheral effects if administered syscally.
High-resolution coronal images of cannulated brains highted the presence of asymmetric foci of hyperintenonsistent with oedema or ventricular enlargements, prenantly in the ipsilateral ventricle of 46% of the animalsmined. These morphological alterations, being mostlilateral and extending throughout the cannulated ventay be ascribed to the implantation procedure. We didbserve any correlation between the presence of ventrnomalies and rCBV response to the injection of GR-736
ndividual subjects. However, at a group analysis level tbnormalities are a source of inter-animal variability that
ts the statistical significance of signal changes in ceeriventricular structures (see discussion below). No si
cant anatomo-pathological alterations in brain parenchere detected otherwise. Overall, these findings sugges
he cannulation protocol used in this study, when couplen appropriate recovery time, does not affect the cerebrular reactivity of the animals, and is therefore suitablehe assessment of the haemodynamic response to pharogical challenges.
Central infusion of vehicle or drug solutions resultedocal increase in the intraventricular signal intensity. Th
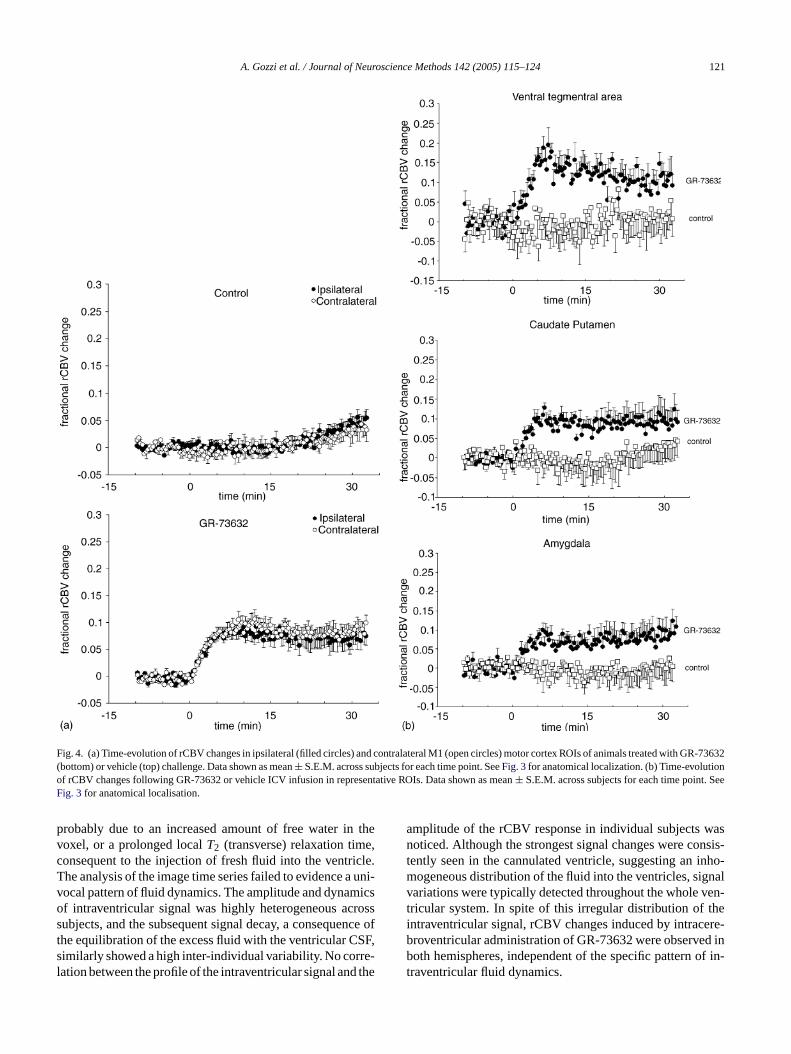
A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124 121
Fig. 4. (a) Time-evolution of rCBV changes in ipsilateral (filled circles) and contralateral M1 (open circles) motor cortex ROIs of animals treated with GR-73632(bottom) or vehicle (top) challenge. Data shown as mean± S.E.M. across subjects for each time point. SeeFig. 3for anatomical localization. (b) Time-evolutionof rCBV changes following GR-73632 or vehicle ICV infusion in representative ROIs. Data shown as mean± S.E.M. across subjects for each time point. SeeFig. 3for anatomical localisation.
probably due to an increased amount of free water in thevoxel, or a prolonged localT2 (transverse) relaxation time,consequent to the injection of fresh fluid into the ventricle.The analysis of the image time series failed to evidence a uni-vocal pattern of fluid dynamics. The amplitude and dynamicsof intraventricular signal was highly heterogeneous acrosssubjects, and the subsequent signal decay, a consequence ofthe equilibration of the excess fluid with the ventricular CSF,similarly showed a high inter-individual variability. No corre-lation between the profile of the intraventricular signal and the
amplitude of the rCBV response in individual subjects wasnoticed. Although the strongest signal changes were consis-tently seen in the cannulated ventricle, suggesting an inho-mogeneous distribution of the fluid into the ventricles, signalvariations were typically detected throughout the whole ven-tricular system. In spite of this irregular distribution of theintraventricular signal, rCBV changes induced by intracere-broventricular administration of GR-73632 were observed inboth hemispheres, independent of the specific pattern of in-traventricular fluid dynamics.

122 A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124
To maximise the sensitivity of the phMRI measurement, ablood pool contrast agent was administered intravenously tomap cerebral blood volume change as a surrogate for neuralactivation. Previous investigators (Mandeville et al., 2001)have demonstrated that the use of contrast agent in phMRIstudies in rodents provides increased functional sensitivity,compared to blood oxygen level dependent signal measure-ments. A typical CBV experiment comprises aT2
∗- or T2-weighted image time series, near the beginning of whichthe contrast agent is injected, upon which the signal dropssharply due to the contrast agent-enhanced relaxivity, and re-flects the local resting state blood volume. Subsequently, thefunctional or pharmacological challenge(s) are applied. Typ-ically, the systemic administration of the contrast agent is fol-lowed by an equilibration period (10–20 min) so to achieve amonotonic washout amenable to detrending. This delay posesrisks of peptidolytic and oxidative degradation if metaboli-cally vulnerable neuropeptides such as substance P are used(Dusser et al., 1988; Endo et al., 1988; Horsthemke et al.,1984; O’Neil et al., 2003). To overcome this problem, GR-73632, a synthetic pentapeptide with a marked resistance topeptidolysis, was chosen as selective NK1 agonist in placeof the endogenous ligand substance P.
Central administration of substance P has been demon-strated to induce a range of fear-related behaviours and de-f ;U larc stra-t l.,1 at-e V re-s rticals gment dala.m -gs boves tived entso de-p rietac
32s ures,p sucha f ther thep da ea thev rationA withe milartp istri-b trig-
gered by shallow ependymal and periventricular receptors.Haffmans et al. (1983)recorded the onset of enkephalin-induced seizures within 2.5 min from the intracerebroven-tricular administration of the compound in parenchymal brainstructures such as the cortex and the hippocampus. Consistentwith this, several other electrophysiology and microdialysisstudies provide evidence of a very rapid onset of neuronalactivity and neurotransmitters release even in deep parenchy-mal structures following intracerebroventricular administra-tion of various neuromodulatory compounds and peptides(Bier et al., 1987; Doi et al., 2001; Erhardt and Engberg, 2000;Kagamiishi et al., 2003; Lejeune and Millan, 2003; Mondaet al., 1998; Nyitrai et al., 1999; Shimizu et al., 1994). More-over, a rapid onset of activity is also in agreement with inhouse and published studies in which GR-73632 and otherNK1 receptor agonists have been used to induce anxiety-related behaviour in rodents (Rupniak et al., 2003).
A predominant area of asymmetry in the activation pat-tern of GR-73632 was observed, with a seemingly strongeractivation in the striatum contralateral to the ICV admin-istration. However, it must be emphasised that the activa-tion maps are the result of group statistical analysis over 13animals. As reported above, implantation of intraventricu-lar cannulae often results in permanent, unilateral ventricularenlargements, thus introducing an element of morphologi-c , thei ran-s sig-n atlyr e att tri-c tieso s ar-g enceb -ga ctiva-t inb . them aso
teb f thed fects.H surew lingo lyingt limi-n neu-r har-a ingfe poralp 32,t ever,a neu-
ensive cardiovascular changes in conscious rats (Saria, 1999nger et al., 1988). Similar behavioural and cardiovascuhanges were observed following focal cerebral adminiion of GR-73632 in rats (Elliott et al., 1991; Mongeau et a998). Administration of GR-73632 (4.2 pmol/rat) in the lral ventricle produced a rapid onset and sustained rCBponse showing signal increases in the cortex and sub-cotructures such as the caudate putamen, the ventral teal area, the lateral hypothalamic nucleus, and the amyg-RNA hybridisation (Maeno et al., 1993) and autoradioraphic localisation studies (Saffroy et al., 1988, 2003)howed the presence of NK1 receptors in all of the atructures. According with a role of substance P in affecisorders, the activation pattern includes critical componf the circuitry that appears to be dysfunctional in bothression and anxiety, such as the amygdala and the paortex (Davidson et al., 1999).
rCBV time courses of animals infused with GR-736howed rapid bilateral increases in several brain structeaking 5–10 min after the injection. The presence ofrapid onset of response may be surprising, in view o
elatively slow penetration of large neuropeptides intoarenchyma.Proescholdt et al. (2000)reported a fairly rapind rather uniform distribution of [14C]-inuline (a substratpproximately eight times larger than GR-73632) withinentricular space, but a subsequent slower tissue penetfaster brain parenchymal penetration was observed
nkephalin, a neuropeptide whose molecular size is sio that of GR-73632 (Haffmans et al., 1983). However, theattern of activation does not necessarily reflect the dution of the compound, as brain activation might be
-
l
.
al asymmetry in about 50% of the animals. Moreoverntracerebroventricular injection of fresh fluid induces a tient and variable local increase in intraventricular MRal intensity. Both sources of inter-animal variability greeduce the statistical significance of the rCBV responshe group level in the proximity of the cannulated venle, and make it difficult to directly compare the intensif ipsi- and contralateral activation in these areas. Thiument is corroborated by the strong spatial correspondetween the ventricular enlargements (Fig. 1, bottom), the reions showing large intraventricular signal changes inFig. 2,nd areas associated with an asymmetric pattern of a
ion in the rCBV map (Fig. 3, slices 2 and 5). Consistently,rain structures farther from the cannulated region (e.gotor cortex,Fig. 4a) a symmetrical activation pattern wbserved.
In phMRI experiments, it is often difficult to discriminaetween rCBV changes induced by the central activity orug, and those induced by peripheral cardiovascular efowever, no significant changes in arterial blood presere detected upon ICV infusion of GR-73632, thus ruut peripheral effects as the dominant mechanism under
he central haemodynamic response to this drug. In preary experiments, we have applied this method to otheroactive compounds, like bicuculline, a potent and well ccterised GABAA antagonist, and corticotropin-releas
actor (CRF), a large polypeptide (Gozzi et al., 2004). Inter-stingly, these compounds induced different spatio-tematterns of brain activity from that observed with GR-736
hus suggesting that the effect may be drug specific. Howdditional work is needed to fully elucidate the precise

A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124 123
rometabolic and neurovascular events underlying the rCBVresponse induced by the NK1 agonist GR-73632.
In conclusion, we have developed methods to study thephMRI response to neuroactive substances centrally admin-istered via a permanent indwelling MR compatible cannulae.We have applied this method to study for the first time therCBV changes induced by ICV infusion of a synthetic pep-tidic neurokinin receptor agonist. We believe this paradigmrepresents a valuable tool for studying non-invasively thefunctional correlates of the central activity induced by neu-ropeptides and other compounds characterised by poor brainpenetration.
References
Banks WA, Kastin AJ. Passage of peptides across the blood–brain barrier:pathophysiological perspectives. Life Sci 1996;59:1923–43.
Bier MJ, Goldfarb J, Orlowski M, Wilk S. Intracerebroventricular infusionof inhibitors of endopeptidase-24.11 (‘enkephalinase’) increases thespontaneous firing frequency of an identifiable set of cells in thesubstantia nigra. Eur J Pharmacol 1987;142:321–30.
Cellier E, Barbot L, Iyengar S, Couture R. Characterization of centraland peripheral effects of septide with the use of five tachykinin NK1receptor antagonists in the rat. Br J Pharmacol 1999;127:717–28.
Chen Q, Andersen AH, Zhang Z, Ovadia A, Gash DM, Avison MJ. Map-ping drug-induced changes in cerebral R2* by Multiple Gradient Re-
76.C eal
plan-ment.
C ac-sing
hav-
C MRI
D rainbiol
D en-oxy-acol
D Air-uced
E tionrain
E l andine
ptides
E sub-okyo)
E theglu-
G ps inage
Ghersi-Egea JF, Finnegan W, Chen JL, Fenstermacher JD. Rapid distribu-tion of intraventricularly administered sucrose into cerebrospinal fluidcisterns via subarachnoid velae in rat. Neuroscience 1996;75:1271–88.
Gozzi A, Schwarz AJ, Reese T, Crestan V, Bertani S, Turrini G, etal. Brain functional imaging of intracerebroventricular injection ofcorticotropin-releasing factor (CRF) in the anaesthetised rat. In: Pro-ceedings of the 12th ISMRM Scientific Meeting and Exhibition onBook of Abstracts, vol. 12; 2004. p. 226.
Greenshaw AJ. Electrical and Chemical Stimulation of Brain Tissue inVivo, vol. 32. Totowa: In Vivo Neuromethods Humana Press; 1998.
Haffmans J, Blankwater YJ, Ukponmwan OE, Zijlstra FJ, Vincent JE,Hespe W, et al. Correlation between the distribution of 3H-labelledenkephalin in rat brain and the anatomical regions involved inenkephalin-induced seizures. Neuropharmacology 1983;22:1021–8.
Hagan RM, Ireland SJ, Jordan CC, Beresford IJ, Deal MJ, Ward P.Receptor-selective, peptidase-resistant agonists at neurokinin NK-1and NK-2 receptors: new tools for investigating neurokinin function.Neuropeptides 1991;19:127–35.
Horsthemke B, Schulz M, Bauer K. Degradation of substance Pby neurones and glial cells. Biochem Biophys Res Commun1984;125:728–33.
Houston GC, Papadakis NG, Carpenter TA, Hall LD, Mukherjee B, JamesMF, et al. Mapping of brain activation in response to pharmacologicalagents using fMRI in the rat. Magn Reson Imaging 2001;19:905–19.
Kagamiishi Y, Yamamoto T, Watanabe S. Hippocampal serotonergic sys-tem is involved in anxiety-like behavior induced by corticotropin-releasing factor. Brain Res 2003;991:212–21.
Lejeune F, Millan MJ. The CRF1 receptor antagonist, DMP695, abol-ishes activation of locus coeruleus noradrenergic neurones by CRF inanesthetized rats. Eur J Pharmacol 2003;464:127–33.
M re-Res
M ennded
M derere-
ed
M Clin
M B,pro-age
M idsrmia.
M ectsuctalrats.
M spec-and
N le ofrosci
O annvel-of sig-803–
O Ann&
called Echo functional MRI. Magn Reson Imaging 1996;14:469–hen Y-CI, Brownell A-L, Galpern W, Isacson O, Bogdanov M, B
MF, et al. Detection of dopaminergic cell loss and neural transtation using pharmacological MRI, PET and behavioural assessNeuroReport 1999;10:2881–6.
hen YC, Galpern WR, Brownell AL, Matthews RT, Bogdanov M, Isson O, et al. Detection of dopaminergic neurotransmitter activity upharmacologic MRI: correlation with PET, microdialysis, and beioral data. Magn Reson Med 1997;38:389–98.
ox RW, Hyde JS. Software tools for analysis and visualization of fdata. NMR Biomed 1997;10:171–8.
avidson RJ, Abercrombie H, Nitschke JB, Putnam K. Regional bfunction, emotion and disorders of emotion. Curr Opin Neuro1999;9:228–34.
oi N, Brown CH, Cohen HD, Leng G, Russell JA. Effects of thedogenous opioid peptide, endomorphin 1, on supraoptic nucleustocin and vasopressin neurones in vivo and in vitro. Br J Pharm2001;132:1136–44.
usser DJ, Umeno E, Graf PD, Djokic T, Borson DB, Nadel JA.way neutral endopeptidase-like enzyme modulates tachykinin-indbronchoconstriction in vivo. J Appl Physiol 1988;65:2585–91.
lliott PJ, Mason GS, Graham EA, Turpin MP, Hagan RM. Modulaof the rat mesolimbic dopamine pathway by neurokinins. Behav BRes 1992;51:77–82.
lliott PJ, Mason GS, Stephens-Smith M, Hagan RM. Behaviourabiochemical responses following activation of midbrain dopampathways by receptor selective neurokinin agonists. Neurope1991;19:119–26.
ndo S, Yokosawa H, Ishii S. Purification and characterization of astance P-degrading endopeptidase from rat brain. J Biochem (T1988;104:999–1006.
rhardt S, Engberg G. Excitation of nigral dopamine neurons byGABA(A) receptor agonist muscimol is mediated via release oftamate. Life Sci 2000;67:1901–11.
enovese CR, Lazar NA, Nichols T. Thresholding of statistical mafunctional neuroimaging using the false discovery rate. NeuroIm2002;15:870–8.
aeno H, Kiyama H, Tohyama M. Distribution of the substance Pceptor (NK-1 receptor) in the central nervous system. Brain1993;18:43–58.
andeville JB, Jenkins BG, Kosofsky BE, Moskowitz MA, RosB, Marota JJA. Regional sensitivity and coupling of BOLD aCBV changes during stimulation of rat brain. Magn Res M2001;45:443–7.
andeville JB, Marota JJA, Kosofsky BE, Keltner JR, WeissleR, Rosen B, et al. Dynamic functional imaging of relative cbral blood volume during rat forepaw stimulation. Magn Res M1998;39:615–24.
antyh PW. Neurobiology of substance P and the NK1 receptor. JPsychiat 2002;63:6–10.
arota JJA, Mandeville JB, Weisskoff R, Moskowitz MA, RosenKosofsky BE. Cocaine activation discriminates dopaminergicjections by temporal response: an fMRI study in rat. NeuroIm2000;11:13–23.
onda M, Viggiano A, Sullo A, De VL. Aspartic and glutamic acincrease in the frontal cortex during prostaglandin E1 hypertheNeuroscience 1998;83:1239–43.
ongeau R, De Oca BM, Fanselow MS, Marsden CA. Differential effof neurokinin-1 receptor activation in subregions of the periaquedgray matter on conditional and unconditional fear behaviors inBehav Neurosci 1998;112:1125–35.
orris PG. Magnetic resonance imaging and magnetic resonancetroscopy assessment of brain function in experimental animalsman. J Psychopharmacol 1999;13:330–6.
yitrai G, Kovacs I, Szarics E, Skuban N, Juhasz G, Kardos J. Rointracellular Ca(2+) stores shaping normal activity in brain. J NeuRes 1999;57:906–15.
gawa S, Menon RS, Tank DW, Kim SG, Merkle H, EllermJM, et al. Functional brain mapping by blood oxygenation ledependent contrast magnetic resonance imaging. A comparisonnal characteristics with a biophysical model. Biophys J 1993;64:12.
’Neil MJ, Smith A, Heckelman PE, Obenchain GR, Gallipeau JAR,D’Arecca M. Merck Index: An Encyclopedia of Chemicals, Drugs

124 A. Gozzi et al. / Journal of Neuroscience Methods 142 (2005) 115–124
Biologicals. Whitehouse Station, NJ, USA: Merck & Co. Inc.; 2003p. 1270–1271.
Paxinos G, Watson C. The Rat Brain in Stereotactic Coordinates. SanDiego: Academic Press; 1998.
Proescholdt MG, Hutto B, Brady LS, Herkenham M. Studies of cere-brospinal fluid flow and penetration into brain following lateral ven-tricle and cisterna magna injections of the tracer [14C]inulin in rat.Neuroscience 2000;95:577–92.
Reese T, Bjelke B, Porszasz R, Baumann D, Bochelen D, Sauter A, et al.Regional brain activation by bicuculline visualized by functional mag-netic resonance imaging. Time-resolved assessment of bicuculline-induced changes in local cerebral blood volume using an intravascularcontrast agent. NMR Biomed 2000;13:43–9.
Rupniak NM, Carlson EJ, Shepheard S, Bentley G, Williams AR, HillA, et al. Comparison of the functional blockade of rat substance P(NK1) receptors by GR205171, RP67580, SR140333 and NKP-608.Neuropharmacology 2003;45:231–41.
Saffroy M, Beaujouan JC, Torrens Y, Besseyre J, Bergstrom L, GlowinskiJ. Localization of tachykinin binding sites (NK1, NK2, NK3 ligands)in the rat brain. Peptides 1988;9:227–41.
Saffroy M, Torrens Y, Glowinski J, Beaujouan J-C. Autoradiographicdistribution of tachykinin NK2 binding sites in the rat brain: compar-ison with NK1 and NK3 binding sites. Neuroscience 2003;116:761–73.
Saria A. The tachykinin NK1 receptor in the brain: pharmacology andputative functions. Eur J Pharmacol 1999;375:51–60.
Scanley BE, Kennan RP, Gore JC. Changes in rat cerebral blood volumedue to modulation of the 5-HT1A receptor measured with suscepti-bility enhanced contrast MRI. Brain Res 2001;913:149–55.
Schenck JF. The role of magnetic susceptibility in magnetic resonanceimaging: MRI magnetic compatibility of the first and second kinds.Med Phys 1996;23:815–50.
Schwarz AJ, Reese T, Gozzi A, Bifone A. Functional MRI using in-travascular contrast agents: detrending of the relative cerebrovascular(rCBV) time course. Magn Reson Imaging 2003;21:1191–200.
Shimizu N, Nakane H, Hori T, Hayashi Y. CRF receptor antagonist at-tenuates stress-induced noradrenaline release in the medial prefrontalcortex of rats. Brain Res 1994;654:145–8.
Silva AC, Zhang W, Williams DS, Koretsky AP. Multi-slice MRI of ratbrain perfusion during amphetamine stimulation using arterial spinlabeling. Magn Reson Med 1995;33:209–14.
Stein EA, Pankiewicz J, Harsch HH, Cho J-K, Fuller SA, Hoffmann RG,et al. Nicotine-induced limbic cortical activation in the human brain:a functional MRI study. Am J Psychiat 1998;155:1009–15.
Unger T, Carolus S, Demmert G, Ganten D, Lang RE, Maser-Gluth C,et al. P induces a cardiovascular defense reaction in the rat: pharma-cological characterization. Circ Res 1988;63:812–20.
Von Bohlen O, Dermietzel R. Neurotransmitters and Neuromodulators.Berlin: Wiley/VCH; 2002.
Weisskoff RM, Zuo CS, Boxerman JL, Rosen BR. Microscopic suscep-tibility variation and transverse relaxation: theory and experiment.Magn Reson Med 1994;31:601–10.
Xu H, Li S-J, Bodurka J, Zhao X, Xi Z-X, Stein EA. Heroin-induced neu-ronal activation in rat brain assessed by functional MRI. NeuroReport2000;11:1085–92.
Zhang Z, Andersen AH, Avison MJ, Gerhardt GA, Gash DM. FunctionalMRI of apomorphine activation of the basal ganglia in awake rhesusmonkeys. Brain Res 2000;852:290–6.




















