
Nanosculpting reversed wavelength sensitivity into a photoswitchable iGluR
Upload
independentCategory
view
0download
0
Original article
Microbes and Infection 8 (2006) 2492e2500www.elsevier.com/locate/micinf
Functional and phenotypic changes in monocytes frompatients with tuberculosis are reversed with treatment
Marıa D. Sanchez1, Yoenis Garcıa1, Carlos Montes, Sara C. Parıs,Mauricio Rojas, Luis F. Barrera, Mauricio A. Arias2, Luis F. Garcıa*
Grupo de Inmunologıa Celular e Inmunogenetica, Facultad de Medicina, Sede de Investigacion Universitaria,Universidad de Antioquia, Cra 53 No. 61-30, Lab. 410, Medellın, Colombia
Received 14 March 2006; accepted 23 June 2006
Available online 13 July 2006
Abstract
Alterations of monocyte/macrophages have been reported in patients with tuberculosis (TB), but their significance is poorly understood.Blood mononuclear cells from patients with different clinical forms of TB, at various times of anti-TB treatment, and healthy tuberculin positiveindividuals, were double-stained for CD14 plus CD206, TLR-2, IFN-gR1, CD40, HLA-DR, CD36 and CD163, and analyzed by flow cytometry.Monocytes were infected with Mycobacterium tuberculosis H37Rv and 24 h later the phenotype, induction of necrosis and apoptosis and pro-duction of tumor necrosis factor TNFa, interleukin (IL)-10 and IL-12p40 were determined. TB patients presented higher percentage of CD14þ
cells but lower percentage of CD14þDRþ and CD14þCD36þ cells. Expression of CD14, HLA-DR and CD36 was decreased in TB patients.Normal percentages and expression were restored during anti-TB treatment. Monocytes from TB patients underwent necrosis and apoptosis afterM. tuberculosis infection, whereas monocytes from healthy controls exhibited only apoptosis. Anti-TB treatment reverted necrosis. There wereno differences between the various clinical forms of TB. In vitro M. tuberculosis infection decreased expression of the membrane moleculesstudied. HLA-DR and CD36 inhibition correlated with induction of apoptosis. Restoration of monocyte alterations during anti-TB treatmentsuggests that such alterations may be caused by the high M. tuberculosis load present during active disease.� 2006 Elsevier SAS. All rights reserved.
Keywords: Tuberculosis; Monocytes; Cytokines; Apoptosis; Necrosis
1. Introduction
The establishment of active tuberculosis (TB) and its clin-ical manifestations are closely linked to the host’s immuneresponse, but the mechanisms underlying the loss of the pro-tective response in patients with TB are still poorly understood[1]. The spectrum of immune response ranges from a protectiveresponse in latent TB, to the absence of a response and dissem-ination of mycobacteria in miliary TB. In pulmonary TB, there
* Corresponding author. Tel.: þ57 4 210 6446; fax: þ57 4 210 6455.
E-mail address: [email protected] (L.F. Garcıa).1 These authors contributed equally to this work.2 Present address: Department of Cellular and Molecular Medicine,
St. George’s, University of London, London SW17 0RE, UK.
1286-4579/$ - see front matter � 2006 Elsevier SAS. All rights reserved.
doi:10.1016/j.micinf.2006.06.005
is an ineffective anti-mycobacterial immune response witha clinically progressive disease, while in pleural TB there isa localized response that can, in some cases, lead to resolutionof the disease even without treatment [2].
Monocytes/macrophages play a determinant role in theanti-mycobacterial defense, the inflammatory granulomatousreaction, and the tissue damage that occurs in TB [3]. Thesecells are the primary target for Mycobacterium tuberculosisinfection, and their innate capacity to deal with mycobacteriadefines the early progression of the infection. Accumulation ofmacrophages and their precursors, circulating monocytes, atthe infection foci is an important feature of the host responseto mycobacteria and granuloma formation [4]. However, in-fected phagocytes within the granuloma eventually die eitherby apoptosis or necrosis [5], and the type of cell death may

2493M.D. Sanchez et al. / Microbes and Infection 8 (2006) 2492e2500
differentially condition the progression of the infection. Thereis evidence that apoptotic macrophages contain the mycobac-teria [6] and the derived apoptotic bodies can be phagocytosedby dendritic cells as a source for antigen presentation to T cells[7]. Contrariwise, necrotic macrophages may release myco-bacteria, able to infect incoming phagocytes, and inflamma-tory components that contribute to tissue damage [8].
Thus alterations in monocyte/macrophage function couldfavor the establishment of active TB. These changes mayalter: (1) the expression of membrane molecules able to bindM. tuberculosis or to transmit activating signals, such ascomplement receptors, mannose receptor (CD206), scavengerreceptors (ScR), Toll-like receptors (TLR), or Interferon-gamma receptor (IFNgR); (2) the pattern of cytokineproduction in response to mycobacterial stimuli, particularlyinterleukin (IL)-12, tumor necrosis factor (TNF) a andIL-10; (3) their antigen presenting capacity because dimin-ished expression of major histocompatibility complex antigensand/or co-stimulatory molecules; (4) their capacity to becomeproperly activated by T-cell derived cytokines; (5) the produc-tion of the anti-mycobacterial effector molecules; and (6) thepattern of cell death by infected macrophages leading tonecrosis, rather than apoptosis.
Therefore, a more detailed characterization of the monocytephenotype and function in patients with TB is needed to betterunderstand the level of immune dysfunction and possiblyprovide some prognostic information that would help to estab-lish a more effective treatment. It is also important to determinethe effect of anti-mycobacterial treatment on these alterations.In this report we show that monocytes from TB patients havereduced expression of CD14, HLA-DR and CD36. This re-duction correlated with M. tuberculosis-induced monocytedeath. Importantly, M. tuberculosis infection induces necrosisin monocytes from TB patients. The abnormalities in pheno-type and monocyte function were reversed with the anti-TBtreatment.
2. Materials and methods
2.1. Study population
Patients with a diagnosis of TB were recruited from differ-ent health units in the metropolitan area of Medellın, Colom-bia. These patients included 127 with pulmonary TB, 45 withpleural TB, 19 with miliary TB, and 14 with other forms of TBincluding meningitis, joint, renal, and ganglionary. TB wasdiagnosed by sputum smear staining for acid fast bacilli andmycobacteria culturing. Clinical and epidemiological criteria,X-ray, biopsy, and adenosine deaminase levels in pleural ef-flux, were considered when direct visualization of mycobacte-ria was not possible. One hundred and seventy patients werestudied at time of diagnosis (0 months of treatment), and 76of them (cohort group) were followed during the 6 monthsof anti-TB treatment. There were also 131 patients whowere studied independently at 0, 3 or 6 months of anti-TBtreatment. HIV-positive individuals, and patients with otherimmunodeficiencies, cancer, autoimmune diseases, as well as
pregnant women, were excluded. As controls, 113 tuberculinpositive healthy individuals were recruited from medical andlaboratory personnel from the Facultad de Medicina, Universi-dad de Antioquia, and other institutions where the patientswere recruited. The tuberculin skin test consists of the intra-dermal injection of 0.1 ml (2 U) of purified protein derivative(RT 23, SSI Copenhagen, Denmark). Indurations �10 mm,measured after 72 h, were considered positive. Written in-formed consent was obtained from all subjects studied. Thestudy was approved by the Ethics Committee of the Facultadde Medicina, Universidad de Antioquia.
2.2. Reagents and antibodies
RPMI-1640 and phosphate-buffered saline (PBS) werepurchased from Gibco-BRL (Grand Island, NY); lymphocyteseparation medium, pooled human serum (PHS) from Bio-Whittaker (Walkersview, MD); fetal calf serum (FCS) fromHyClone (South Logan, UT); trypan blue, NaN3, and bovineserum albumin (BSA) from SigmaeAldrich (St. Louis,MO); ethidium bromide (EtBr) from BE ICN (Cleveland,OH); terminal deoxynucleotidyl transferase (TdT) dUTP bio-tin Nick End Labeling (TUNEL) from Mebstain (Medicaland Biological Laboratories, Nagoya, Japan); DIOC6 (3,3 0-dihexyloxacarbocyanine iodide) from Molecular Probes(Eugene, OR); paraformaldehyde from Fisher Scientific (Pitts-burgh, PA); Middlebrook 7H9 and 7H10 media from BectonDickinson (Cockeysville, MD); and glycerol from Mallin-ckrodt Baker (Xalostoc, Mexico). Antibodies used in flow cy-tometry are shown in Table 1.
2.3. Culture and maintenance of M. tuberculosis H37Rv
Mycobacterium tuberculosis H37Rv was obtained from theInstituto Nacional de Salud (Bogota, Colombia), and grown in7H9 liquid medium. Cultures were harvested at exponentialgrowing phase. To disaggregate clumps, mycobacteria weresuspended in DMEM, and sonicated at 2.5 W output for3 min at 4 �C (Sonics Vibra Cell, model CV33, Newtown,CT), centrifuged for 5 min at 400 � g, the supernatant dilutedin PBS at 0.1 OD and frozen at �70 �C in DMEM containing30% glycerine and 10% FCS. The number of colony formingunits was determined as described [9].
2.4. Phenotyping of circulating monocytes
Mononuclear cells (PBMCs) were separated from 35 ml ofdefibrinated blood on Lymphocyte separation medium, washedand resuspended in RPMI-1640 supplemented with 0.5% PHSplus 2 mM L-glutamine and 10 mM HEPES. Cell viability wasalways �95% as tested by trypan blue exclusion. Five hundredthousand PBMC were washed and resuspended in 50 ml ofstaining buffer (PBS plus 1% PHS and 0.1% NaN3), contain-ing anti-CD14-PE plus antibodies against either HLA-DR orCD40, involved in antigen presentation [10], CD36 andCD163, two scavenger receptors associated with removal ofapoptotic cells [11], or mannose receptor (CD206), TLR-2

2494 M.D. Sanchez et al. / Microbes and Infection 8 (2006) 2492e2500
Table 1
Antibodies
Antibodies to: Clone Isotype Source
CD3-Cy UCHT1 IgG1* BD PharMingen, San Diego, CA
CD14-PE M5E2 IgG2a BD PharMingen
CD36-FITC CB38 (NL07) IgM BD PharMingen
CD40-FITC 5C3 IgG1 BD PharMingen
HLA-DR-FITC TU36 IgG2b Caltag Laboratories, Burlingame, CA
Mannose receptor-Cy-Chrome (CD206) 19.2 IgG1 BD PharMingen
TLR2-FITC TL2.1 IgG2a BD Biosciences
IFNg-R1-FITC 2E2 Hamster IgG Santa Cruz Biotechnology, Santa Cruz, CA
CD163-PE GHI/61 IgG1 BD PharMingen
Mouse IgG-FITC/PE/Cy-Chrome MOPC-21 IgG1 BD PharMingen
Mouse IgG- FITC/PE G155e178 IgG2a BD PharMingen
Mouse IgG-FITC 27e35 IgG2b BD PharMingen
Mouse IgM-FITC G155e228 IgM BD PharMingen
Mouse IgG-FITC BPC-4 IgG2b Ancell, Bayport, MN
Hamster IgG-FITC IgG Santa Cruz Biotechnology
and IFN-gRa, associated with phagocytosis and macrophageactivation [12], labeled as described in Table 1. Isotype controlantibodies were used as control for non-specific binding. Cellswere incubated on ice for 20 min, washed, resuspended in PBSand fixed with 2% paraformaldehyde. Ten thousand eventswere acquired in an EPICS XL flow cytometer (Coulter, Hia-leah, FL), and the percentage of positive cells and the meanfluorescence intensity (MFI) was analyzed using the softwareWindows Multiple Document Interface 2.8 (WinMDI, ScriptsResearch Institute, La Jolla, CA.).
2.5. Culture of monocytes and in vitroinfection with M. tuberculosis
PBMC (5 � 105 CD14þ cells) were cultured in 1 ml ofmedium in 24-well plates (BD Falcon, Bedford, MA) for 2 hat 37 � C. After incubation, the wells were extensively washedwith warm PBS containing 0.5% PHS. Adherent cells werecultured in RPMI-1640 supplemented with 10% PHS plus2 mM L-glutamine and 10 mM HEPES for 24 h. The percent-age of contaminating CD3þ cells was always �5%. Thenumber of adherent cells was determined in replicate wellby counting the nuclei [13]. The cells were infected withM. tuberculosis at a multiplicity of infection (MOI) of 5 for24 h. After incubation, culture supernatants were collectedand frozen at �70 �C, and the cells fixed with 2% paraformal-dehyde. Thereafter, the cells were harvested using rubberscrapers (Sarstedt, Newton, NC) and stained for determinationof surface markers and cell death by flow cytometry.
2.6. Cytokine measurement
TNFa, IL-10 and IL-12p40 levels in culture supernatantsfrom M. tuberculosis-infected and non-infected monocyteswere determined by ELISA using commercial kits (Duoset,R&D, Minneapolis, MN) following the manufacturer’sinstructions.
2.7. Determination of cell death ofM. tuberculosis-infected monocytes
Apoptosis and necrosis were determined by simultaneousstaining with DIOC6 and EtBr [14]. M. tuberculosis-infectedmonocytes (5 � 105) were resuspended in 2 ml PBS and in-cubated with 25 ml (1.3 mg/ml) of EtBr and 10 ml of 7 mMDIOC6 for 20 min at room temperature in darkness, followedby fixation with 2% paraformaldehyde. The cells werewashed and resuspended in PBS for flow cytometry analysis.Cells with high fluorescence intensity for DIOC6 and ne-gative for EtBr were considered alive. Cells with low fluores-cence intensity for DIOC6 and EtBr negative were consideredapoptotic, and cells positive for EtBr were considerednecrotic [14].
The TUNEL technique was used together with FITC-labeled anti-CD36 and anti-HLA-DR to determine the ex-pression of HLA-DR and CD36 on apoptotic cells. For thispurpose, 1 � 106 M. tuberculosis-infected monocytes werefixed with 2% paraformaldehyde, washed and permeabilizedwith PBS containing 0.5% Tween-20 and 0.2% BSA. TheTdT reagent (TdT buffer, dUTPs-PE, and TdT enzyme)was added in 18:1:1 ratio. After incubating for 1 h at37 �C, the cells were washed and resuspended in PBS con-taining 0.1% NaN3. Anti-CD36-FITC, anti-HLA-DR-FITC,or the respective isotype controls, were added at a finalconcentration of 5 mg/ml. The cells were washed and re-suspended in deionized water plus 0.2% BSA before flowcytometry analysis. Cells in absence of TdT enzyme wereused as negative control.
2.8. Statistical analysis
Comparisons between groups were done by one- and two-way ANOVA. Comparison between infected and non-infectedcultures was done by two-tailed paired t-test. Results wereconsidered significant when P < 0.05. All analyses weredone using Prism 4 software (Graphpad, San Diego, CA).
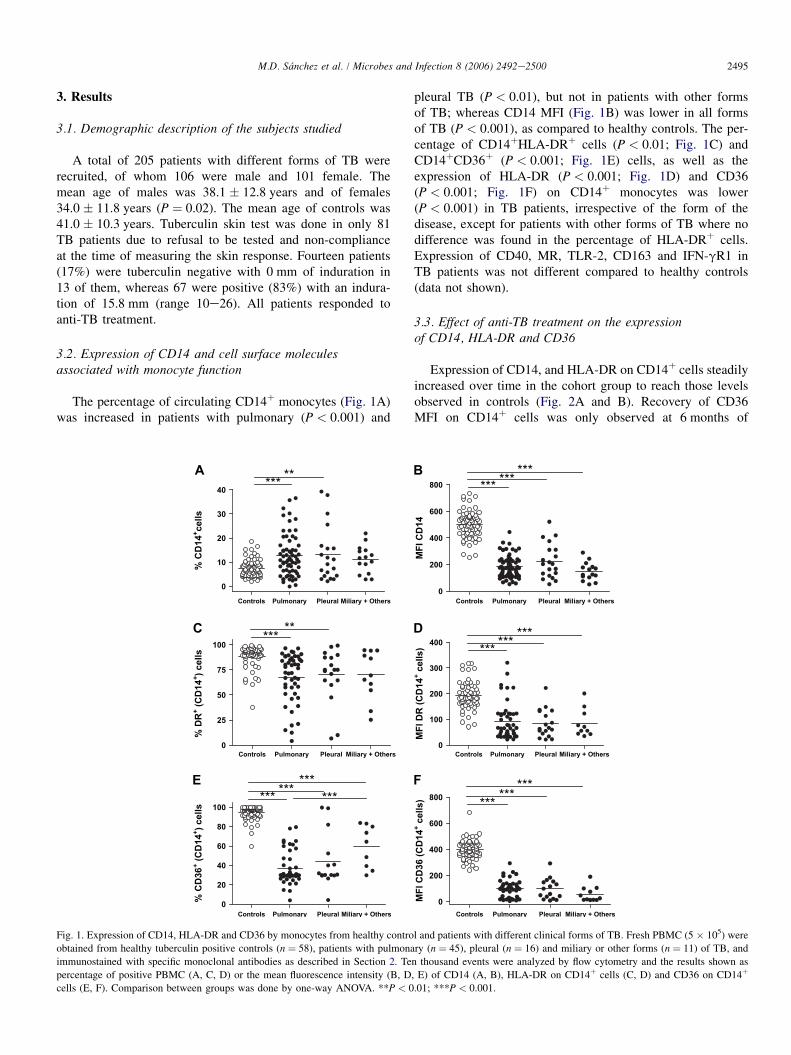
2495M.D. Sanchez et al. / Microbes and Infection 8 (2006) 2492e2500
3. Results
3.1. Demographic description of the subjects studied
A total of 205 patients with different forms of TB wererecruited, of whom 106 were male and 101 female. Themean age of males was 38.1 � 12.8 years and of females34.0 � 11.8 years (P ¼ 0.02). The mean age of controls was41.0 � 10.3 years. Tuberculin skin test was done in only 81TB patients due to refusal to be tested and non-complianceat the time of measuring the skin response. Fourteen patients(17%) were tuberculin negative with 0 mm of induration in13 of them, whereas 67 were positive (83%) with an indura-tion of 15.8 mm (range 10e26). All patients responded toanti-TB treatment.
3.2. Expression of CD14 and cell surface moleculesassociated with monocyte function
The percentage of circulating CD14þ monocytes (Fig. 1A)was increased in patients with pulmonary (P < 0.001) and
pleural TB (P < 0.01), but not in patients with other formsof TB; whereas CD14 MFI (Fig. 1B) was lower in all formsof TB (P < 0.001), as compared to healthy controls. The per-centage of CD14þHLA-DRþ cells (P < 0.01; Fig. 1C) andCD14þCD36þ (P < 0.001; Fig. 1E) cells, as well as theexpression of HLA-DR (P < 0.001; Fig. 1D) and CD36(P < 0.001; Fig. 1F) on CD14þ monocytes was lower(P < 0.001) in TB patients, irrespective of the form of thedisease, except for patients with other forms of TB where nodifference was found in the percentage of HLA-DRþ cells.Expression of CD40, MR, TLR-2, CD163 and IFN-gR1 inTB patients was not different compared to healthy controls(data not shown).
3.3. Effect of anti-TB treatment on the expressionof CD14, HLA-DR and CD36
Expression of CD14, and HLA-DR on CD14þ cells steadilyincreased over time in the cohort group to reach those levelsobserved in controls (Fig. 2A and B). Recovery of CD36MFI on CD14þ cells was only observed at 6 months of
A
Controls Pulmonary Pleural Miliary + Others Controls Pulmonary Pleural Miliary + Others
Controls Pulmonary Pleural Miliary + OthersControls Pulmonary Pleural Miliary + Others
Controls Pulmonary Pleural Miliary + Others Controls Pulmonary Pleural Miliary + Others
0
10
20
30
40*** **
% C
D14
+cells
B
0
200
400
600
800 *** *** ***
MF
I C
D14
C
0
25
50
75
100
*** **
% D
R+ (C
D14
+) cells
D
0
100
200
300
400
E
0
20
40
60
80
100
*** *********
% C
D36
+ (C
D14
+) cells
F
0
200
400
600
800 *** ******
MF
I C
D36 (C
D14
+ cells)
*** *** ***
MF
I D
R (C
D14
+ cells)
Fig. 1. Expression of CD14, HLA-DR and CD36 by monocytes from healthy control and patients with different clinical forms of TB. Fresh PBMC (5 � 105) were
obtained from healthy tuberculin positive controls (n ¼ 58), patients with pulmonary (n ¼ 45), pleural (n ¼ 16) and miliary or other forms (n ¼ 11) of TB, and
immunostained with specific monoclonal antibodies as described in Section 2. Ten thousand events were analyzed by flow cytometry and the results shown as
percentage of positive PBMC (A, C, D) or the mean fluorescence intensity (B, D, E) of CD14 (A, B), HLA-DR on CD14þ cells (C, D) and CD36 on CD14þ
cells (E, F). Comparison between groups was done by one-way ANOVA. **P < 0.01; ***P < 0.001.
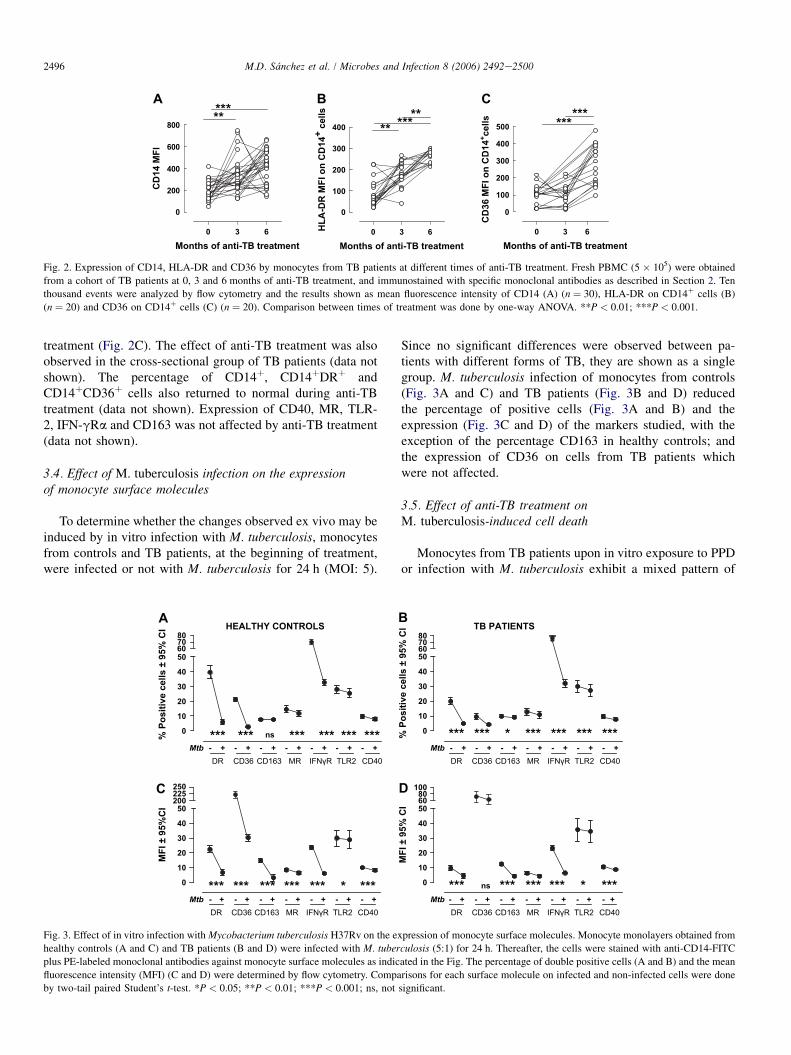
2496 M.D. Sanchez et al. / Microbes and Infection 8 (2006) 2492e2500
A
0 3 6
0
200
400
600
800
*****
Months of anti-TB treatment
CD
14 M
FI
C
0 3 6
0
100
200
300
400
500 ***
***
CD
36
M
FI o
n C
D1
4+c
ells
B
0 3 6
0
100
200
300
400***
**
**
HL
A-D
R M
FI o
n C
D1
4+
c
ells
Months of anti-TB treatment Months of anti-TB treatment
Fig. 2. Expression of CD14, HLA-DR and CD36 by monocytes from TB patients at different times of anti-TB treatment. Fresh PBMC (5 � 105) were obtained
from a cohort of TB patients at 0, 3 and 6 months of anti-TB treatment, and immunostained with specific monoclonal antibodies as described in Section 2. Ten
thousand events were analyzed by flow cytometry and the results shown as mean fluorescence intensity of CD14 (A) (n ¼ 30), HLA-DR on CD14þ cells (B)
(n ¼ 20) and CD36 on CD14þ cells (C) (n ¼ 20). Comparison between times of treatment was done by one-way ANOVA. **P < 0.01; ***P < 0.001.
treatment (Fig. 2C). The effect of anti-TB treatment was alsoobserved in the cross-sectional group of TB patients (data notshown). The percentage of CD14þ, CD14þDRþ andCD14þCD36þ cells also returned to normal during anti-TBtreatment (data not shown). Expression of CD40, MR, TLR-2, IFN-gRa and CD163 was not affected by anti-TB treatment(data not shown).
3.4. Effect of M. tuberculosis infection on the expressionof monocyte surface molecules
To determine whether the changes observed ex vivo may beinduced by in vitro infection with M. tuberculosis, monocytesfrom controls and TB patients, at the beginning of treatment,were infected or not with M. tuberculosis for 24 h (MOI: 5).
Since no significant differences were observed between pa-tients with different forms of TB, they are shown as a singlegroup. M. tuberculosis infection of monocytes from controls(Fig. 3A and C) and TB patients (Fig. 3B and D) reducedthe percentage of positive cells (Fig. 3A and B) and theexpression (Fig. 3C and D) of the markers studied, with theexception of the percentage CD163 in healthy controls; andthe expression of CD36 on cells from TB patients whichwere not affected.
3.5. Effect of anti-TB treatment onM. tuberculosis-induced cell death
Monocytes from TB patients upon in vitro exposure to PPDor infection with M. tuberculosis exhibit a mixed pattern of
A
0
10
20
30
40
50
- - - - - - -+ + + + + + +
****** *** *** *** ***
Mtb
ns
60
70
80
% P
ositive cells ± 95%
C
I
B
0
10
20
30
40
50
*** *** * *** *** *** ***
60
70
80
% P
os
itive cells ± 95%
C
I
C
0
10
20
30
40
50
*** *** *** *** *** * ***
200
225
250
MF
I ± 95%
CI
D
0
10
20
30
40
50
*** ns *** *** *** * ***
60
80
100
HEALTHY CONTROLS TB PATIENTS
DR CD36 CD163 MR IFNγR TLR2 CD40- - - - - - -+ + + + + + +Mtb
DR CD36 CD163 MR IFNγR TLR2 CD40
- - - - - - -+ + + + + + +Mtb
DR CD36 CD163 MR IFNγR TLR2 CD40- - - - - - -+ + + + + + +Mtb
DR CD36 CD163 MR IFNγR TLR2 CD40
MF
I ± 95%
C
I
Fig. 3. Effect of in vitro infection with Mycobacterium tuberculosis H37Rv on the expression of monocyte surface molecules. Monocyte monolayers obtained from
healthy controls (A and C) and TB patients (B and D) were infected with M. tuberculosis (5:1) for 24 h. Thereafter, the cells were stained with anti-CD14-FITC
plus PE-labeled monoclonal antibodies against monocyte surface molecules as indicated in the Fig. The percentage of double positive cells (A and B) and the mean
fluorescence intensity (MFI) (C and D) were determined by flow cytometry. Comparisons for each surface molecule on infected and non-infected cells were done
by two-tail paired Student’s t-test. *P < 0.05; **P < 0.01; ***P < 0.001; ns, not significant.

2497M.D. Sanchez et al. / Microbes and Infection 8 (2006) 2492e2500
necrosis and apoptosis, whereas cells from healthy controlsonly show apoptosis [14]. Here, we tested whether the typeof monocyte death varies in the different clinical forms ofTB, and if the type of death changes during anti-TB treatment.After 24 h of infection, nearly half of M. tuberculosis-infectedmonocytes from healthy controls died from apoptosis(Fig. 4A), whereas fewer than 10% died from necrosis. In TBpatients, irrespective of the form of the disease, about one thirdof the cells died from necrosis, and a similar percentage of cellsshowed apoptosis. This death pattern was reverted by anti-TBtreatment (Fig. 4B), with percentages of necrosis and apoptosissimilar to healthy controls at the third month of treatment.
3.6. Effect of apoptosis on HLA-DR and CD36expression by M. tuberculosis-infected monocytes
We next examined whether the down-regulation of expres-sion of the molecules studied induced by M. tuberculosis infec-tion could be associated with induction of apoptosis. Monocytesfrom healthy controls were infected or not with M. tuberculosis,and double stained with TUNEL and anti-HLA-DR or anti-CD36. A high percentage of monocytes expressed HLA-DR(Fig. 5A) and CD36 (Fig. 5D) prior to infection. After 24 h,non-infected cells showed a marked decrease in the expressionof both molecules, although the cells remain viable, as shown bythe negligible number of TUNELþ cells (Fig. 5B and E). M. tu-berculosis infection further inhibited HLA-DR and CD36 ex-pression (Fig. 5C and F), but this inhibition was accompaniedby great increase of TUNELþ-CD36� and -HLA-DR� cells,suggesting that inhibition of the expression of monocyte surfacemolecules after in vitro M. tuberculosis infection may be due tocell death induced by the infection.
3.7. Cytokine production by M. tuberculosis-infectedmonocytes
Induction of apoptosis and necrosis is modulated by theTNFa/IL-10 ratio and M. tuberculosis induces production ofboth cytokines [14,15]. IL-12 also plays an important role
during M. tuberculosis infection favoring a Th1 response [16].Thus, we measured TNFa, IL-10 and IL-12p40 in supernatantsof monocyte cultures infected with M. tuberculosis fromhealthy controls and TB patients with the clinical forms studied(Fig. 6). M. tuberculosis infection induced production of TNFaby healthy controls and TB patients, with no differences be-tween the various forms of disease (Fig. 6A). M. tuberculosis in-duced IL-10 production by the different groups (Fig. 6B), withthe exception of patients with other forms of TB in whom thedifference between infected and non-infected cultures was notsignificant. Increased IL-12p40 production was observed inhealthy controls and patients with pulmonary and other formsof TB, but not in patients with pleural TB (Fig. 6C).
4. Discussion
Our results demonstrate that active TB, irrespective of theclinical form of the disease, results in changes of the exp-ression, as well as in the percentage of monocytes expressingsurface molecules known to play important functions in theanti-TB response. The results also confirm previous observa-tions [14] that TB patients have an altered pattern of monocytedeath upon in vitro infection with M. tuberculosis.
Patients with pulmonary and pleural TB had increased per-centage of circulating CD14þ cells, but CD14 expression wassignificantly decreased in all clinical forms of TB. However,the expression levels and the percentage of CD14þ monocytesrecovered to normal values with anti-TB treatment. We [17]and others [18] have reported that TB patients have augmentedserum levels of soluble CD14 that return to normal values dur-ing anti-TB treatment. It is possible that during active TB,CD14 is released from the monocyte membrane into circula-tion [19], explaining its low expression on monocyte mem-brane, and the increased levels of serum sCD14. Successfulanti-TB treatment may result in retention of mCD14 on thecell membrane with subsequent decrease of sCD14. Impor-tantly, sCD14 has immunomodulatory effects, such as inhibi-tion of IL-2 and IFNg production [20], which have beenreported in patients with TB [21].
A
Control Pulmonar Pleural Miliary
0
20
40
60
80
100
Live
NecroticApoptotic
B
0 3 6
Time of treatment
% cells ± 95%
C
I
% cells ± 95%
C
I
0
20
40
60
80
100
Fig. 4. Effect of in vitro infection with M. tuberculosis and anti-tuberculosis treatment on monocyte death. (A) Monocytes obtained from healthy individuals
(n ¼ 18), and TB patients with pulmonary (n ¼ 13), pleural (n ¼ 4) and other forms of TB (n ¼ 2) were cultured at 5 � 105 cells/well and infected for 24
with M. tuberculosis at a MOI of 5. Then the cells were washed and incubated in the presence of EtBr and DIOC6, and 5000 events were acquired by flow cy-
tometry. The cells with high fluorescence intensity for DIOC6 and negative for EtBr were considered alive. The cells with low fluorescence intensity for DIOC6 and
EtBr negative were considered apoptotic, and the cells positive for EtBr were considered necrotic. (B) Monocyte monolayers from TB patients at 0 (n ¼ 19), 3
(n ¼ 19) and 6 (n ¼ 16) months of anti-TB treatment were infected and the percentage of live, apoptotic and necrotic cells detected as described above.

2498 M.D. Sanchez et al. / Microbes and Infection 8 (2006) 2492e2500
TUNEL
CD
36
DR
60%
2 h non-infected 24 h non-infected 24 h M.tb-infected
79% 22% 2% 5%
80%
15% 3% 1%
82%
A B C
D E F
Fig. 5. Effect of M. tuberculosis infection and cell adherence on CD36 and HLA-DR expression by blood-derived monocytes. Monocytes were allowed to adhere
onto plastic wells for 2 h (A and D) and then infected (C and F) or not (B and E) with M. tuberculosis at a MOI of 5 and cultured for 24 h. After culture, the cells
were tested for expression of HLA-DR (A, B, C) and CD36 (D, E, F) by staining with specific monoclonal antibodies, and analyzed by flow cytometry.
Figure shows one representative experiment of three.
Our group has previously reported that monocytes from TBpatients have decreased ability to phagocytose M. tuberculosis[22], and here we show that such patients have diminished per-centage of CD14þDRþ circulating monocytes and low expres-sion of HLA-DR on CD14þ cells. This evidence suggests thatduring active TB, monocytes, and presumably their descen-dent macrophages and myeloid DC, have impaired capacityto engulf, process and present mycobacterial antigens to Tcells, as shown by other authors [23]. These abnormalitiesmay be partially responsible for the depressed specific T-cellresponses observed in TB patients.
CD36 was also found to be severely impaired, both in thepercentage of positive cells and its expression on CD14þ cells.CD36 is a member of the scavenger receptors (SR) type B fam-ily and its expression is up-regulated during monocyte to mac-rophage maturation [11]. CD36 binds oxLDL and HDL andparticipates in removal of apoptotic cells. There is evidencethat transforming growth factor (TGF)-b down-regulatesCD36 expression [24], suggesting that the high serum levelsof TGF-b present in TB patients [25] may be responsible forthe low expression and decreased percentage of CD36þ mono-cytes. The reduction of CD36þ positive monocytes and its de-creased expression suggest that monocytes/macrophages fromTB patients may have an altered ability to recognize andremove apoptotic cells, allowing these cells to become latenecrotic and favoring bacterial dissemination.
The induction of apoptosis and necrosis of monocytes by invitro infection with M. tuberculosis confirms our previous re-ports [14]. Monocytes from individuals with latent TB under-went apoptosis upon infection with M. tuberculosis, whereasmonocytes from TB patients presented both apoptosis and ne-crosis, irrespective of the type of the disease. Anti-TB treat-ment reversed the induction of necrosis. These resultsfurther suggest that apoptosis may be a protective mechanismin M. tuberculosis infection whereas monocyte necrosis is animportant event in the deregulated inflammatory responsethat leads to the tissue damage and mycobacterial spreadthat occurs in active TB.
One important question that arises from our results relatesto the cause(s) of monocyte alterations in TB patients. The dif-ferences between healthy tuberculin-positive individuals andTB patients and the reversibility of the observed alterationswith anti-TB treatment provide strong evidence that changesin monocytes are caused by active infection. Results also sug-gest that systemic factors, from bacterial or host origin, maybe responsible for such effects. Circulating mycobacterialproducts, such as ManLAM [26], may participate in the mono-cyte abnormalities found in TB patients. On the host side, sys-temic mediators (cytokines, hormones, sCD14, among others)may be responsible for monocyte alterations in TB patients.They may affect the generation of new monocytes requiredat the granulomatous inflammation. They may also change

2499M.D. Sanchez et al. / Microbes and Infection 8 (2006) 2492e2500
the subsets of circulating monocyte by affecting their state ofmaturation [27], or inducing an alternative type of activation[28]. Chemokines [29] and adhesion molecules [30] inducedduring active TB may also define the migration of monocytesubsets from circulation into affected tissues.
Although down-regulation of the membrane molecules afterin vitro infection with M. tuberculosis cannot explain the exvivo findings, it may be relevant at the granuloma level. Thedown-regulation of HLA-DR and CD36 expression by non-infected monocytes, with no evidence of diminished cell viabil-ity, is in agreement with our own previous observation [14].However, in M. tuberculosis-infected cultures the decrease in
TNF-α
Controls Pulmonary Pleural Miliary + Others
1
10
100
1000
10000 -+
*** *** ** *
TB
Controls Pulmonary Pleural Miliary + Others
TB
Controls Pulmonary Pleural Miliary + Others
TB
pg
/m
l (G
eo
metric m
ean
± 95%
CI)
pg
/m
l (G
eo
metric m
ean
± 95%
CI)
pg
/m
l (G
eo
metric m
ean
± 95%
CI)
IL-10
1
10
100
1000
10000
-+
*** *** * ns
IL-12p40
1
10
10
1000
-+
* ** ns **
Fig. 6. Cytokine production by monocytes from patients with different clinical
forms of TB, infected in vitro with M. tuberculosis. Monocyte monolayers
from healthy controls (n ¼ 23), and patients with pulmonary (n ¼ 33), pleural
(n ¼ 8) and miliary plus other TB forms (n ¼ 5) were infected or not with
M. tuberculosis at a MOI of 5 during 24 h. Thereafter, the culture supernatants
were collected and assayed for TNFa (A), IL-10 (B) and IL-12p40 (C) by
ELISA. Differences between infected and non-infected cultures in the different
groups were analyzed by two-way ANOVA. *P < 0.05; **P < 0.01;
***P < 0.001; ns, not significant.
HLA-DR and CD36 expression on apoptotic cells was morestriking, suggesting that it may be a consequence of the M. tu-berculosis-induced monocyte death. However, more detailedexperiments are required to differentiate the effects mediatedby cell death and those mediated by M. tuberculosis infectionindependently of cell death.
Finally, we cannot exclude that the absence of significantdifferences between different clinical forms of TB may bedue to either the high variability within the groups of patients,or to the small number of patients in some groups (i.e. mili-ary). However, results suggest that monocyte phenotypes arenot predictive for TB susceptibility, and that monocytes arenot responsible for the spectrum of immune responses foundin the various forms of TB. Rather, these differences may bedue to variable alterations of the T-cell dependent acquiredimmune response.
Acknowledgements
This work was supported by Colciencias (Bogota, Colom-bia), grant 1115-04-11957. The authors acknowledge thepatients and control individuals that participated in this study,the collaboration of the personnel of the institutions wherethe patients were recruited, and the personnel of the FlowCytometry Unit.
References
[1] S.H.E. Kaufmann, Recent findings in immunology give tuberculosis
vaccines a new boost, Trends Immunol. 26 (2005) 660e667.
[2] K.V. Jalapathy, C. Prabha, S.D. Das, Correlates of protective immune
response in tuberculous pleuritis, FEMS Immunol Med. Microbiol. 40
(2004) 139e145.
[3] M.J. Fenton, M.W. Vermeulen, Immunopathology of tuberculosis: Roles
of macrophages and monocytes, Infect. Immun. 64 (1996) 683e690.
[4] M. Gonzalez-Juarrero, T.S. Shim, A. Kipnis, A.P. Junqueira-Kipnis,
I.M. Orme, Dynamics of macrophage cell populations during murine
pulmonary tuberculosis, J. Immunol. 171 (2003) 3128e3135.
[5] R. Placido, G. Mancino, A. Amendola, F. Mariani, S. Vendetti,
M. Piacentini, A. Sanduzzi, M.L. Bocchino, M. Zembala, V. Colizzi, Ap-
optosis of human monocyte/macrophages in Mycobacterium tuberculosis
infection, J. Pathol. 181 (1997) 31e38.
[6] L. Duan, H. Gan, D.E. Golan, H.G. Remold, Critical role of mitochon-
drial damage in determining outcome of macrophage infection with
Mycobacterium tuberculosis, J. Immunol. 169 (2002) 5181e5187.
[7] U.E. Schaible, F. Winau, P.A. Sieling, K. Fischer, H.L. Collins,
K. Hagens, R.L. Modlin, V. Brinkmann, S.H.E. Kaufmann, Apoptosis
facilitates antigen presentation to T lymphocytes through MHC-I and
CD1 in tuberculosis, Nat. Med. 9 (2003) 1039e1046.
[8] L. Danelishvili, J. McGarvey, Y.J. Li, L.E. Bermudez, Mycobacteriumtuberculosis infection causes different levels of apoptosis and necrosis
in human macrophages and alveolar epithelial cells, Cell Microbiol. 5
(2003) 649e660.
[9] M. Rojas, L.F. Barrera, G. Puzo, L.F. Garcia, Differential induction of ap-
optosis by virulent Mycobacterium tuberculosis in resistant and suscepti-
ble murine macrophages: role of nitric oxide and mycobacterial products,
J. Immunol. 159 (1997) 1352e1361.
[10] F.M. Brodsky, L.E. Guagliardi, The cell biology of antigen processing
and presentation, Annu. Rev. Immunol. 9 (1991) 707e744.
[11] L. Peiser, S. Gordon, The function of scavenger receptors expressed by
macrophages and their role in the regulation of inflammation, Microbes
Infect. 3 (2001) 149e159.

2500 M.D. Sanchez et al. / Microbes and Infection 8 (2006) 2492e2500
[12] R. van Crevel, T.H.M. Ottenhoff, J.W.M. van der Meer, Innate Immu-
nity to Mycobacterium tuberculosis, Clin. Microbiol. Rev. 15 (2002)
294.
[13] A. Nakagawara, C.F. Nathan, A simple method for counting adherent
cells: application to cultured human monocytes, macrophages and multi-
nucleated giant cells, J. Immunol. Methods 56 (1983) 261e268.
[14] D.P. Gil, L.G. Leon, L.I. Correa, J.R. Maya, S.C. Paris, L.F. Garcia,
M. Rojas, Differential induction of apoptosis and necrosis in monocytes
from patients with tuberculosis and healthy control subjects, J Infect. Dis.
189 (2004) 2120e2128.
[15] C. Barreto Pereira, M. Palaci, O.H.M. Leite, A.J.S. Duarte, G. Benard,
Monocyte cytokine secretion in patients with pulmonary tuberculosis dif-
fers from that of healthy infected subjects and correlates with clinical
manifestations, Microbes Infect. 6 (2004) 25e33.
[16] I.S. Leal, B. Smedegard, P. Andersen, R. Appelberg, Interleukin-6 and
interleukin-12 participate in induction of type I protective T-cell response
during vaccination with a tuberculosis subunit vaccine, Infect. Immun. 67
(1999) 5747e5754.
[17] E. Pacheco, C. Fonseca, C. Montes, J. Zabaleta, L.F. Garcia, M.A. Arias,
CD14 gene promoter polymorphism in different clinical forms of tuber-
culosis, FEMS Immunol. Med. Microbiol. 40 (2004) 207e213.
[18] S.D. Lawn, M.O. Labeta, M. Arias, J.W. Acheampong, G.E. Griffin, El-
evated serum concentrations of soluble CD14 in HIV� and HIVþ patients
with tuberculosis in Africa: prolonged elevation during anti-tuberculosis
treatment, Clin. Exp. Immunol. 120 (2000) 483e487.
[19] J.-J. Durieux, N. Vita, O. Popescu, F. Guette, J. Calzada-Wack,
R. Munker, R.E. Schmidt, J. Lupker, P. Ferrara, H.W.L. Ziegler-
Heitbrock, M.O. Labeta, The two soluble forms of the lipopolysaccharide
receptor, CD14: characterization and release by normal human mono-
cytes, Eur. J. Immunol. 24 (1994) 2006e2012.
[20] J.E. Rey Nores, A. Bensussan, N. Vita, F. Stelter, M.A. Arias, M. Jones,
S. Lefort, L.K. Borysiewicz, P. Ferrara, M.O. Labeta, Soluble CD14 acts
as a negative regulator of human T cell activation and function, Eur.
J. Immunol. 29 (1999) 265e276.
[21] F.O. Sanchez, J.I. Rodriguez, G. Agudelo, L.F. Garcia, Immune respon-
siveness and lymphokine production in patients with tuberculosis and
healthy controls, Infect. Immun. 62 (1994) 5673e5678.
[22] J. Zabaleta, M. Arias, J.R. Maya, L.F. Garcıa, Diminished adherence and/
or ingestion of virulent Mycobacterium tuberculosis by monocyte-
derived macrophages from patients with tuberculosis, Clin. Diagn. Lab.
Immunol. 5 (1998) 690e694.
[23] J. Gercken, J. Pryjma, M. Ernst, H.-D. Flad, Defective antigen presenta-
tion by Mycobacterium tuberculosis-infected monocytes, Infect. Immun.
62 (1994) 3472e3478.
[24] G. Draude, R.L. Lorenz, TGF-beta1 downregulates CD36 and scavenger
receptor A but upregulates LOX-1 in human macrophages, Am. J. Phys-
iol. Heart Circ. Physiol. 278 (2000) H1042eH1048.
[25] Z. Toossi, P. Gogate, H. Shiratsuchi, T. Young, J.J. Ellner, Enhanced pro-
duction of TGF-b by blood monocytes form patients with active tubercu-
losis and presence of TGF-b in tuberculous granulomatous lung lesions,
J. Immunol. 154 (1995) 465e473.
[26] E. Sada, D. Aguilar, M. Torres, T. Herrera, Detection of liporabinomann as
a diagnostic test for tuberculosis, J. Clin. Microbiol. 30 (1992) 2415e2418.
[27] E. Grage-Griebenow, H.-D. Flad, M. Ernst, Heterogeneity of human
peripheral blood monocyte subsets, J. Leukoc. Biol. 69 (2001) 11e20.
[28] S. Gordon, Alternative activation of macrophages, Nat. Rev. Immunol. 3
(2003) 23e35.
[29] Y. Lin, J. Gong, M. Zhang, W. Xue, P.F. Barnes, Production of monocyte
chemoattractant protein 1 in tuberculosis patients, Infect. Immun. 66
(1998) 2319e2322.
[30] G.M. Lopez Ramırez, W.N. Rom, C. Ciotoli, A. Talbot, F. Martiniuk,
B. Cronstein, J. Reibman, Mycobacterium tuberculosis alters expression
of adhesion molecules on monocytic cells, Infect. Immun. 62 (1994)
2515e2520.
Copyright © 2022 FDOKUMEN




















