
Function of the rat carotid body chemoreceptors in ageing
13
Function of the rat carotid body chemoreceptors in ageing Silvia V. Conde,* , Ana Obeso,* Ricardo Rigual,* Emilia C. Monteiro and Constancio Gonzalez* *Departamento de Bioquı ´mica y Biologı ´a Molecular y Fisiologı ´a, Facultad de Medicina. Universidad de Valladolid, Valladolid, Spain Departamento de Farmacologı ´a, Faculdade de Cie ˆncias Me ´dicas, Universidade Nova de Lisboa, Lisbon, Portugal Abstract Some age-related deficits in the ventilatory responses have been attributed to a decline in the functionality of the carotid body (CB) arterial chemoreceptors, but a systematic study of the CB function in ageing is lacking. In rats aged 3– 24 months, we have performed quantitative morphometry on specific chemoreceptor tissue, assessed the function of che- moreceptor cells by measuring the content, synthesis and release of catecholamines (a chemoreceptor cell neurotrans- mitter) in normoxia and hypoxia, and determined the func- tional activity of the intact organ by measuring chemosensory activity in the carotid sinus nerve (CSN) in normoxia, hypoxia and hypercapnic acidosis. We found that with age CBs en- large, but at the same time there is a concomitant decrease in the percentage of chemoreceptor tissue. CB content and turnover time for their catecholamines increase with age. Hypoxic stimulation of chemoreceptor cells elicits a smaller release of catecholamines in rats after 12 months of age, but a non-specific depolarizing stimulus elicits a comparable release at all ages. In parallel, there was a marked decrease in the responsiveness to hypoxia, but not to an acidic-hypercapnic stimulus, assessed as chemosensory activity in the CSN. We conclude that in aged mammals chemoreceptor cells become hypofunctional, leading to a decreased peripheral drive of ventilation. Keywords: ageing, carotid body, catecholamines, dopamine, hypoxia, Parkinson’s disease. J. Neurochem. (2006) 99, 711–723. Parenchymatous chemoreceptor cells of the carotid bodies (CBs) detect changes in arterial blood gases. They are activated when PO 2 decreases and when PCO 2 and/or [H + ] increase, and respond with an increase in the rate of release of their neurotransmitters. Released neurotransmit- ters set the action potential frequency in the carotid sinus nerve (CSN). Among chemoreceptor cells neurotransmit- ters, catecholamines (CA), particularly dopamine (DA), are released in proportion to stimulus intensity and in parallel to the CSN chemosensory activity (Fidone et al. 1982; Rigual et al. 1986, 1991; Gonzalez et al. 1992). CSN conveys the information to the brainstem where, in hypoxia and/or hypercapnia, it promotes hyperventilation aimed to restore blood gases to normality (Loeschcke 1974; Fidone and Gonzalez 1986; Gonzalez et al. 1994). CBs also contribute to set resting ventilation and exercise hyperpnea, as humans and experimental animals, when glomectomized, exhibit increased resting arterial PCO 2 , and a reduced tolerance to physical exercise as a result of reduced ventilation (Bisgard et al. 1976; Honda 1985; Fordyce 1987; Forster et al. 2000). These modifications associated with glomectomy reflect the situation in aged subjects (Chan and Welsh 1998). Ageing produces a gradual decline in the functional reserve of bodily organs and systems, with the result of a reduction in the ability to maintain homeostasis. Age- dependent reductions in breathing capacity, O 2 supply to tissues and arterial PO 2 have been observed (Sorbini et al. 1968; Cerveri et al. 1995; Gunnarsson et al. 1996; Chan and Welsh 1998; Knapowski et al. 2002). The reduced capacity of hypoxia to elicit an arousal reaction in aged sleeping dogs has been attributed to CB malfunction (Phillipson and Kozar 1993) and linked to the increased frequency of obstructive sleep apnoea in humans (Janssens et al. 1999; Behan et al. 2002). Semiquantitative histological studies of CBs in aged animals have revealed an increase in non-specific tissue with Received April 21, 2006; revised manuscript received June 6, 2006; accepted June 26, 2006. Address correspondence and reprint requests to Professor Dr Con- stancio Gonza ´lez, Departamento de Bioquı ´mica y Biologı ´a Molecular y Fisiologı ´a, Facultad de Medicina, Universidad de Valladolid, C/Ramo ´ny Cajal n°7, 47005 Valladolid. Spain. E-mail:[email protected] Abbreviations used: CA, catecholamines; CB, carotid body; CSN, carotid sinus nerve; DA, dopamine; NE, norepinephrine; PB, phosphate buffer; PBS, phosphate-buffered saline; TH, tyrosine hydroxylase. Journal of Neurochemistry , 2006, 99, 711–723 doi:10.1111/j.1471-4159.2006.04094.x Ó 2006 The Authors Journal Compilation Ó 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723 711
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Function of the rat carotid body chemoreceptors in ageing

Function of the rat carotid body chemoreceptors in ageing
Silvia V. Conde,*,� Ana Obeso,* Ricardo Rigual,* Emilia C. Monteiro� and Constancio Gonzalez*
*Departamento de Bioquımica y Biologıa Molecular y Fisiologıa, Facultad de Medicina. Universidad de Valladolid, Valladolid, Spain
�Departamento de Farmacologıa, Faculdade de Ciencias Medicas, Universidade Nova de Lisboa, Lisbon, Portugal
Abstract
Some age-related deficits in the ventilatory responses have
been attributed to a decline in the functionality of the carotid
body (CB) arterial chemoreceptors, but a systematic study of
the CB function in ageing is lacking. In rats aged 3–
24 months, we have performed quantitative morphometry on
specific chemoreceptor tissue, assessed the function of che-
moreceptor cells by measuring the content, synthesis and
release of catecholamines (a chemoreceptor cell neurotrans-
mitter) in normoxia and hypoxia, and determined the func-
tional activity of the intact organ by measuring chemosensory
activity in the carotid sinus nerve (CSN) in normoxia, hypoxia
and hypercapnic acidosis. We found that with age CBs en-
large, but at the same time there is a concomitant decrease in
the percentage of chemoreceptor tissue. CB content and
turnover time for their catecholamines increase with age.
Hypoxic stimulation of chemoreceptor cells elicits a smaller
release of catecholamines in rats after 12 months of age, but
a non-specific depolarizing stimulus elicits a comparable
release at all ages. In parallel, there was a marked
decrease in the responsiveness to hypoxia, but not to an
acidic-hypercapnic stimulus, assessed as chemosensory
activity in the CSN. We conclude that in aged mammals
chemoreceptor cells become hypofunctional, leading to a
decreased peripheral drive of ventilation.
Keywords: ageing, carotid body, catecholamines, dopamine,
hypoxia, Parkinson’s disease.
J. Neurochem. (2006) 99, 711–723.
Parenchymatous chemoreceptor cells of the carotid bodies(CBs) detect changes in arterial blood gases. They areactivated when PO2 decreases and when PCO2 and/or [H+]increase, and respond with an increase in the rate ofrelease of their neurotransmitters. Released neurotransmit-ters set the action potential frequency in the carotid sinusnerve (CSN). Among chemoreceptor cells neurotransmit-ters, catecholamines (CA), particularly dopamine (DA), arereleased in proportion to stimulus intensity and in parallelto the CSN chemosensory activity (Fidone et al. 1982;Rigual et al. 1986, 1991; Gonzalez et al. 1992). CSNconveys the information to the brainstem where, inhypoxia and/or hypercapnia, it promotes hyperventilationaimed to restore blood gases to normality (Loeschcke1974; Fidone and Gonzalez 1986; Gonzalez et al. 1994).CBs also contribute to set resting ventilation and exercisehyperpnea, as humans and experimental animals, whenglomectomized, exhibit increased resting arterial PCO2,and a reduced tolerance to physical exercise as a result ofreduced ventilation (Bisgard et al. 1976; Honda 1985;Fordyce 1987; Forster et al. 2000). These modificationsassociated with glomectomy reflect the situation in agedsubjects (Chan and Welsh 1998).
Ageing produces a gradual decline in the functionalreserve of bodily organs and systems, with the result of areduction in the ability to maintain homeostasis. Age-dependent reductions in breathing capacity, O2 supply totissues and arterial PO2 have been observed (Sorbini et al.1968; Cerveri et al. 1995; Gunnarsson et al. 1996; Chan andWelsh 1998; Knapowski et al. 2002). The reduced capacityof hypoxia to elicit an arousal reaction in aged sleeping dogshas been attributed to CB malfunction (Phillipson and Kozar1993) and linked to the increased frequency of obstructivesleep apnoea in humans (Janssens et al. 1999; Behan et al.2002). Semiquantitative histological studies of CBs in agedanimals have revealed an increase in non-specific tissue with
Received April 21, 2006; revised manuscript received June 6, 2006;accepted June 26, 2006.Address correspondence and reprint requests to Professor Dr Con-
stancio Gonzalez, Departamento de Bioquımica y Biologıa Molecular yFisiologıa, Facultad de Medicina, Universidad de Valladolid, C/Ramon yCajal n�7, 47005 Valladolid. Spain. E-mail:[email protected] used: CA, catecholamines; CB, carotid body; CSN,
carotid sinus nerve; DA, dopamine; NE, norepinephrine; PB, phosphatebuffer; PBS, phosphate-buffered saline; TH, tyrosine hydroxylase.
Journal of Neurochemistry, 2006, 99, 711–723 doi:10.1111/j.1471-4159.2006.04094.x
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723 711

maintenance or decrease in parenchymatous cells (Partanenet al. 1984; Hurst et al. 1985; Pokorsky et al. 2004).
These observations prompted us to study the effect of ageingon the basic morphometric parameters of the CB, on thefunction of chemoreceptor cells in the rat CB by monitoringtheir ability to store, synthesize and release CA in normoxiaand hypoxia, and on the capacity of the CB to drive respirationby recording the activity in the CSN in normoxic, hypoxic andhypercapnic conditions. We have found that, in spite of anenlargement of the organ, there is an age-dependent decrease inparenchymatous tissue after 12 months of age. The enlargedCBs exhibit a high CA content and turnover time and adiminished responsiveness to hypoxic stimuli to evoke therelease of this biogenic amine. Similarly, the chemosensoryactivity elicited by hypoxia, but not by hypercapnic acidosis, inthe CSN decreases significantly after 12 months.
Materials and methods
Animals and surgery
The experiments were performed in Sprague–Dawley rats of both
sexes aged 3 months (controls) and 12, 18 and 24 months
(experimental animals). All along their lifespan, the animals were
kept in the vivarium of our university with a seasonal day/light
rhythm, with temperature controlled between 23�C and 26�C and
with water and food ad libitum. The Institutional Committee of the
University of Valladolid for Animal Care and Use approved the
protocols.
For the experiments aiming to measure endogenous CA content,
and synthesis and release of [3H]CA, we used 8–12 CBs per
experiment. TheCBswere cleaned free of CSNand nearby connective
tissue following procedures previously described (Vicario et al.2000). For the recording of CSN activity, the preparation CB-CSN
was identified under a dissecting microscope and a block of tissue,
including the carotid bifurcation and the glossopharyngeal nerve, was
removed and placed in a Lucite chamber in ice-cold/100% O2-
equilibrated Tyrode (in mM: NaCl 140, KCl 5, CaCl2 2, MgCl2 1.1,
HEPES 10, glucose 5.5, pH 7.40) for further dissection of tissue
surrounding the CB and CSN. The CB-CSN preparation was digested
over 3–5 min in collagenase type I (1 mg/mL) solution to loosen the
perineurium (Rigual et al. 2002). The CB-CSN preparation was
maintained in ice-cold 100% O2-equilibrated Tyrode until it was
transferred to the recording chamber. In all instances, animals were
killed by an intracardiac overdose of sodium pentobarbital.
Labelling of CA stores to measure the rate of synthesis and
release of [3H]CA
General procedures for the synthesis and release experiments have
been described in prior publications (Fidone and Gonzalez 1982;
Vicario et al. 2000). In brief, the stores of CA in the CB were
labelled by incubating the organs over 2 h in a solution containing
tyrosine, the natural precursor of CA, labelled with tritium (3,5-
[3H]tyrosine, Amersham Pharmacia Biotech, Piscataway, NJ, USA).
After completion of the labelling period, in the experiments devoted
to measuring the rate of [3H]CA synthesis, the CBs were
homogenized and [3H]CA synthesized and free [3H]tyrosine present
in the tissues were measured in the supernatants, and [3H]tyrosine
incorporated into proteins was measured in the pellets.
In the release experiments, once the labelling period was
completed the CB was transferred individually to glass vials kept
in a shaker bath at 37�C for the entire experiment. The CBs were
gently moved by a continuous bubbling of the incubating solutions
with a gas mixture containing 20% O2/5% CO2/75% N2 saturated
with water vapour, except during the 10-min periods of hypoxic
stimulation. Solutions were collected at selected times (see Results)
for subsequent analysis of their [3H]CA content. Hypoxic and
depolarizing stimuli consisted in the incubation of the CBs for
10 min with low PO2-equilibrated (2% or 5% O2) and high K+-
containing solutions. All collected solutions were acidified with
glacial acetic acid to a pH of 3.0 and maintained at 4�C to prevent
degradation of the [3H]CA release until analysis. On completing the
release experiments, the CBs were processed for their analysis in
[3H]CA content as in the synthesis experiments.
Analytical procedures
[3H]Tyrosine incorporated into proteins was determined by liquid
scintillation spectrometry of the pellets resulting from centrifugation
of the acid homogenates of the CB. Free [3H]tyrosine accumulated
by the CBs during the period of synthesis was also measured by
liquid scintillation counting: an aliquot of the supernatants was
counted and the radioactivity present in it minus radioactivity in the
form of [3H]CA was taken as free intracellular [3H]tyrosine.
For the analysis of endogenous unlabelled and [3H]-labelled CA
in the tissue extracts, aliquots (10–50 lL) of supernatants were
directly injected into an HPLC system composed of a Milton Roy
CM 400 pump (Milton Roy Co., Rivera Beach, FL, USA), a Waters
C18 (particle size 4 lm) column (Waters Corporation, Barcelona,
Spain), a Waters U6K injector, a Bioanalytical Systems LC-4A
electrochemical detector (set at a holding potential of 0.75 mVand a
sensitivity of 1–5 nA; Bioanalytical Systems, West Lafayette, IN,
USA). The signal coming out of the detector was fed to an analogue-
to-digital converter controlled by Peak Sample Chromatography
System Software (Buck Scientific, East Northwalk, CT, USA).
Identification and quantification of endogenous CA in tissue
samples were carried out against external standards. In the case of
the synthesis experiments, identification of [3H]catechols was
similarly performed against external standards, but quantification
was made by collection of the HPLC column effluents correspond-
ending to the peaks of interest and scintillation counting of the
collected effluents.
The analysis of [3H]catechols present in the collected incubating
solutions in the release experiments included: adsorption to alumina
(100 mg) at a pH of 8.6, washing of alumina with distilled water,
bulk elution of all catechols (1 mL, 1 N HCl) and scintillation
counting. To raise the pH in the collected solutions from 3 to 8.6,
under continuous shaking we added to the vials 3.6 mL of 2 M
TRIS-buffer at a pH of 8.7.
Recording of CSN activity
The CB-CSN preparation was transferred to a recording chamber
mounted on a dissection microscope (Nikon Corporation, Tokyo,
Japan) and superfused (37�C) with bicarbonate/CO2 buffered saline
(in mM: NaCl 120, NaHCO3 24, KCl 3, CaCl2 2, MgCl2 1.1,
glucose 5, pH 7.40). Recordings of single or few fibres of CSN were
712 S. V. Conde et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723� 2006 The Authors
made using a suction electrode. The pipette potential was amplified
(Neurolog Digimiter, Hertfordshire, UK), displayed on an oscillo-
scope and stored on a personal computer (PC; 200 Hz acquisition
rate; Axonscope; Axon Instruments, Sunnyvale, CA, USA). Chemo-
receptor activity was identified (spontaneous generation of action
potentials at irregular intervals) and confirmed by its increase in
response to hypoxia (normoxia 20% O2 + 5% CO2 + 75% N2;
hypoxia 0% or 5% O2 + 5% CO2 + balanced N2). CSN activity was
digitalized, summed every second and converted in a voltage
proportional to the sum. Hypoxic and hypercapnic stimulation were
achieved, respectively, by perfusion of the preparations with solutions
equilibrated with gas mixtures containing 0% or 5% O2 + 5%
CO2 + balanced N2 and 20% O2 + 20% CO2 + balance of N2.
Morphometric studies of the CB: percentage of tissue sections
occupied by tyrosine hydroxylase positive chemoreceptor cells
Anaesthetized rats were heparinized (500 U/kg, i.v.) and then
perfused by gravity through the left ventricle at a pressure of 100 cm
of water. Perfusion consisted of 150 mL of phosphate-buffered
saline (PBS; 10 mM; pH 7.40) at 37�C, followed by 250 mL of 4%
paraformaldehyde in 0.1 M phosphate buffer (PB; pH 7.40) at 4�C.The carotid artery bifurcations were removed and the CBs cleaned
under a dissecting microscope of surrounding tissues. CBs were
post-fixed for 1 h in 4% paraformaldehyde in PB at 4�C and
transferred to a 30% sucrose in PB, also at 4�C, for cryoprotection.After embedding in tissue-tek� (Sakura Finetek Europe B.V.,
Zoeterwoude, The Netherlands), the CBs were frozen at )20�C.Serial sections 10-lm thick were obtained in a Leitz Cryostat
(model 1720) and collected in glass slides coated with 3-amino-
propyltriethoxy-silane (Sigma, St Louis, MO, USA). The sections
were washed in PBS at room temperature (22–23�C) for 5 min and
incubated in permeabilizing-blocking solution (PBS containing
0.1% Triton X-100 and 2% non-immunized goat serum) for
30 min. The incubation with the primary antibody (mouse anti-
tyrosine hydroxylase; Abcam, Barcelona, Spain) at a dilution
1 : 1000 in permeabilizing-blocking solution was made at 4�Cand carried overnight. After extensive (3 · 10 min) washing with
PBS, the sections were incubated with the secondary antibody (goat
anti-mouse-FITC; Sigma) at 1 : 1000 in permeabilizing-blocking
solution for 1 h at room temperature (22–23�C). Finally, the sectionswere washed with PBS (4 · 5 min) and with distilled water and
mounted with Vectashield (Vector Laboratories, Burlingame, CA,
USA). Negative controls were similarly incubated but in the absence
of primary antibody.
The sections were examined with a fluorescence microscope
(Axioscop 2 mot plus; Zeiss, Pacisa-Girarlt, Madrid, Spain) with the
excitation and emission filters for FITC. Images were captured using
the Photometric CoolSnap colour digital camera (Roper Scientific
GmbH, Ottobrunn, Germany) and stored in a computer. With
adequate software (Metamorph 6.3; Universal Imaging Corporation,
Dowingtown, PA, USA) the tyrosine hydroxylase-positive (TH+)
areas and the entire area of the CB tissue in each section were
measured and used to calculate the percentage area immunopositive
to the rate-limiting enzyme in CA biosynthesis.
Presentation of the data and statistical analysis
Most of the data are presented as means ± SEM, although some
prototype experiments are presented individually. When the data
are expressed per unit weight, it is intended to normalize and to
correct by the differences in CB weight, both intragroup and
intergroups and thereby to reduce the dispersion. However, in
some instances it is also necessary to express the data per CB:
this is important to understand what should be expected when a
CB is transplanted as it might occur in Parkinson’s disease.
Finally, in the release experiments it is very convenient to express
the evoked release as a percentage of the tissue content: it
corrects for the potential variations in CB size and/or rate of
[3H]CA synthesis.
One-way analysis of variance with Dunnett’s multiple comparison
test between groups was used to compare means of the different
groups. Differences were considered significant at the p £ 0.05 level.
Fig. 1 Age-dependent variation in body and carotid body weight in rat.
(a) From 3 to 12 months there is a large increase in body weight which
thereafter is maintained for up to 24 months. (b) In the CB, the in-
crease in organ size is only noticeable at 18 months of age. Data
represent means ± SEM of n individual values given in the drawing.
***p < 0.001.
Carotid body function in ageing 713
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723

Results
Age-related evolution of the body and of the CB weight
Figure 1(a) shows the age-related increase in body size inrats between 3 and 24 months. The group of 3 months ofpost-natal age comprised rats aged 80–100 days and theirmean body weight was 260.1 ± 8.4 g (n ¼ 75). At12 months of age, mean body weight reached 421.4 ±19.8 g (n ¼ 41), and at older ages there were only verymodest increases in body size to reach weights very closeto 460 g at 18 and 24 months of age.
The pattern of weight variation in the CB weight wasdifferent. In 111 CB control animals the mean CB weightwas 50.22 ± 1.97 lg, which was not different from that of12-month-old rats (45.91 ± 2.20 lg; n ¼ 56). However, at18 and 24 months the weights of the CBs were signifi-cantly larger, reaching weights of 70–75 lg (Fig. 1b).These unparalleled variations in body and carotid bodyweights determine that the ratios of CB weight to totalbody weight, that in control animals was 1.97 · 10)7,dropped to 1.09 · 10)7 at 12 months and returned to theintermediate values of 1.6 and 1.52 · 10)7 at 18 and24 months of age.
Content of CA in the CBs at different ages
Figure 2(a) shows the mean content of CA, DA andnorepinephrine (NE) in CBs of the rats of the different ages;data are expressed as pmol/CB. Mean DA content in controlCBs was 16.27 ± 1.47 pmol/CB (n ¼ 27). At 12 months ofage there was a moderate 10% increase in DA content thatdid not reach statistical significance. However, at 18 and24 months the content of DA in the CBs more than doubledto reach 41.83 ± 5.60 pmol/CB (n ¼ 12) and 55.71 ±7.16 pmol/CB (n ¼ 10), respectively. In all chromatograms,a small amount of dihydroxyphenyl acetic acid (the maincatabolite of DA), representing between 0.5 and 2% of theDA, was also detected (not shown). Contrary to DA, NElevels were similar at all ages with contents of 4.57 ±0.32 pmol/CB at 3 months and a tendency to increasemoderately with age to reach 6.16 ± 0.73 pmol/CB at24 months of age.
When the data are expressed per unit of CB weight(Fig. 2b), the same trend is evident regarding the DAcontent, which increases with age from 326.20 ±25.98 pmol/mg tissue at 3 months up to 650.80 ±60.29 pmol/mg tissue at 24 months. The small tendencyfor NE to increase, observed when the data are expressed perorgan, disappeared when expressed per mg of tissue: incontrol CBs the NE content amounted to 88.80 ± 7.35 pmol/CB and at 24 months it amounted 82.89 ± 7.56 pmol/CB.
Figure 3(c) shows mean DA : NE ratios. It is evident thatthe rat CB is a dopaminergic organ at all ages, and that thedopaminergic quality increases with age. Thus, in the CBs of
3-month-old control animals, the DA : NE ratio was3.94 ± 0.20 and it increased and reached values of8.24 ± 0.69 in the organs of 24-month-old rats.
Fig. 2 Age-dependent variations in the levels of catecholamines (CA)
and in the dopamine (DA) to norepinephrine (NE) ratios. (a, b) Absolute
levels of DA and NE in the CBs of 3–24-month-old rats expressed,
respectively, as pmol/CB and pmol/mg tissue. Note that the tendency
for DA to increase with age reaches statistical significance at 18 and
24 months of age. (c) DA/NE ratios increase in an age-dependent
manner. The dihydroxyphenyl acetic acid (DOPAC) content of the CB
was included to estimate the ratios as it is the only CA catabolite
present in significant amounts. Data represent means ± SEM of
n individual values shown. *p < 0.05, **p < 0.01, ***p < 0.001.
714 S. V. Conde et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723� 2006 The Authors

CA synthesis and CA turnover time at different ages
The rate of [3H]CA synthesis in CBs of different ages, aswell as the amount of free [3H]tyrosine present in the tissue,and the amount of [3H]tyrosine incorporated into proteins,are shown in Fig. 3. Figure 3(a) shows the data expressedper CB. The rate of [3H]DA synthesis in control CBs was1.56 ± 0.17 (n ¼ 12) pmol/CB/h and, while at 12 months
there was a small tendency to decrease, at 18 and 24 monthsthere was a clear tendency to increase, reaching rates of2.57 ± 0.0.82 (n ¼ 8) pmol/CB/h in the CBs of the oldestanimals. A similar pattern was noticeable with the rate ofsynthesis of [3H]NE, with the incorporation of free [3H]tyro-sine into the tissue and with the incorporation of [3H]tyrosineinto proteins. When the data are expressed per unit of tissueweight (Fig. 3b), there were not statistical differences orage-rated tendencies, except for an apparent decrease in theamount of [3H]tyrosine incorporated into proteins at18 months of age. In Fig. 3(c), we present the turnovertimes for DA and NE at all ages. They have been estimatedby dividing the endogenous content by the rate of CAsynthesis {i.e. pmol/CB/[(pmol/CB)/ h] or pmol/mg tissue/[(pmol/mg tissue)/h]}. While in the case of NE there werenot significant age-related changes in the turnover time, at allages being in the vicinity of 60–70 h, in the case of DA therewas a clear trend for the turnover time to increase with age:in control CBs obtained from 3-month-old animals, theturnover time was 9–10 h and at 24 months it reached 22 h(p < 0.01). Although there are not additional statisticaldifferences using the one-way ANOVA with Dunnett’s multi-comparison test, if we used a two-way unpaired t-test tocompare the data of 3 versus 12, 3 versus 18 and 3 versus 24-month-old rats the values of p reach 0.058, 0.003 and 0.001,respectively.
Absence of seasonal variations in CA content and
rate of synthesis in the CB
As the experiments reported here were spanned over a longperiod of time, it was necessary to exclude the possibility thatthe observed differences were produced by the seasonality ofthe experiments or other uncontrolled variables. To uncoversuch a possibility, we included a group of control animals ineach of the experiments aimed to measure the content ofendogenous CA and the rate of [3H]CA synthesis. The dataobtained in the control groups at the different times of thestudy are shown in Table 1. We did not observe significantseasonal differences except for an apparently more rapidturnover of NE (a smaller turnover time) in the group usedduring the summer time.
Release of [3H]CA in normoxia and in response to
hypoxia and high external K+ in the CBs of rats of
different ages
Figure 4(a–d) shows typical experiments carried out withindividual CBs obtained, respectively, from 3-, 12-, 18- and24-month-old animals. Note that in all cases the normoxicrelease of [3H]CA (20% O2-equilibrated solutions;PO2 � 140 mmHg; open bars) increases abruptly in thefraction correspondent to 40–50 min of the experimentswhen the organs were stimulated with hypoxia by incubationwith 2% O2-equilibrated solutions (PO2 � 22 mmHg; filledbars). In 2–3 normoxic fractions, following hypoxic stimu-
Fig. 3 Synthesis rate and turnover time of catecholamine (CA), and
accumulation of the natural precursor tyrosine in the CB at different
ages. (a) Data are expressed as pmol/organ/h and it can be seen that
there is a tendency for the synthesis of CA and the accumulation of
free precursor to increase at 18 and 24 months. (b) Such a tendency
disappears when data are expressed per unit weight. (c) While the NE
turnover time does not vary with age, the DA turnover time exhibits a
clear age dependency that reached statistical significance (**p < 0.01)
at 24 months using a one-way ANOVA test, but if data of 12, 18 and
24 months are individually compared with the values of control 3-
month-old rats p-values were, respectively, 0.058, 0.003 and 0.001.
Data represent means ± SEM of n individual values shown.
Carotid body function in ageing 715
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723

lation, the release declines slowly to pre-stimulation levels asa result of the washout of the [3H]CA and their labelledcatabolites (Gonzalez et al. 1987). The release evoked by thestimulus corresponds to the d.p.m. above the dotted line inthe graphs, and can be estimated mathematically bysubtracting the basal release from each of the stimulus andpost-stimulus fractions. From the release observed in theseindividual CBs, there appears to exist an age-dependentrelease of [3H]CA in hypoxic conditions (note differentscales in each individual experiment). Figure 5(a) shows thatthe mean basal or normoxic release obtained in all the releaseexperiments, and quantified in the sample immediately priorto the application of the stimuli, shows a non-significanttendency to increase at 24 months (22.08 ± 2.88 fmol/CB/10 min vs. 12.83 ± 1.32 fmol/CB/10 min; p ¼ 0.1067).When the basal release is expressed per mg of tissue, or aspercentage of tissue content (Figs 5b and c), any age-dependent tendency disappears.
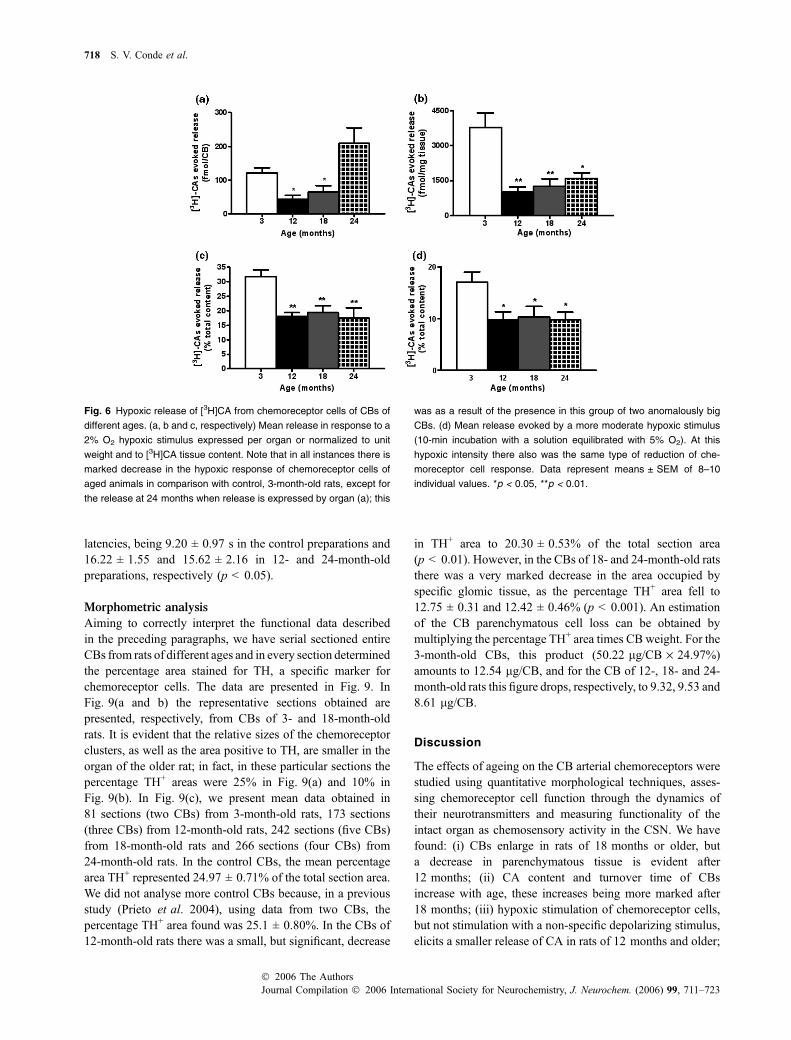
The mean release evoked by hypoxic stimulation (10-minincubation with 2% O2; PO2 � 22 mmHg) are shown inFig. 6(a–c). From Fig. 6(a), where the evoked release isexpressed per CB, it is shown that, at 12 and 18 months,each CB releases a smaller amount of [3H]CA than thecontrol 3-month-old organs, while in the organs of 24-month-old animals there is a clear tendency for the evokedrelease to be higher than in control animals. It should benoted, however, that this tendency is as a result of thepresence in this group of two unusually large CBs (180 and240 lg) which we have not excluded because their rate ofrelease expressed per unit weight or as percentage of tissuecontent is in the range of the release obtained in the rest ofthe CBs of the group (eight CBs in total). Figure 6(b and c)present the evoked release of [3H]CA induced by thehypoxic stimulus expressed as fmol/mg tissue and aspercentage of the [3H]CA content in the organs. Thispresentation of the data show that, when the differences in
Table 1 Seasonal variations in the content and rate of synthesis of CA in the rat CB
Season NE content [3H]NE synthesis NE turnover time DA content [3H]DA synthesis DA turnover time
Winter 2001 88.80 ± 7.35
(n ¼ 16)
1.23 ± 0.15
(n ¼ 12)
70.21 ± 4.94
(n ¼ 16)
326.20 ± 25.98
(n ¼ 16)
35.08 ± 4.63
(n ¼ 12)
9.20 ± 0.81
(n ¼ 16)
Summer 2002 84.52 ± 4.94
(n ¼ 6)
1.85 ± 0.16
(n ¼ 4)
45.76* ± 2.68
(n ¼ 6)
355.30 ± 54.17
(n ¼ 6)
44.95 ± 5.44
(n ¼ 4)
7.90 ± 1.20
(n ¼ 6)
Winter 2002 83.83 ± 8.31
(n ¼ 10)
1.46 ± 0.29
(n ¼ 11)
60.71 ± 5.02
(n ¼ 10)
317.54 ± 47.35
(n ¼ 10)
31.27 ± 5.54
(n ¼ 11)
10.18 ± 1.52
(n ¼ 10)
NE and DA content are expressed as pmol/mg tissue. [3H]NE and [3H]DA synthesis is expressed as (pmol/mg tissue)/h. Turnover times are
expressed in hours and were calculated by dividing each individual datum of NE and DA content by the mean rate of synthesis for each CA.
*p < 0.05.
Fig. 4 Sample release experiments in CBs
of 3-, 12-, 18- and 24-month-old rats (a, b, c
and d, respectively). In all cases, the time
course of [3H]CA release is indicated with
the black bars representing a hypoxic sti-
mulation of the organs (10-min incubation
with a solution equilibrated with 2% O2).
The dotted lines throughout the histograms
separate the basal normoxic (below the
line) from the hypoxia-induced release
(above). Note the different scales at differ-
ent ages.
716 S. V. Conde et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723� 2006 The Authors

the weight of the CB are eliminated, the control CBs releasegreater amounts of [3H]CA in response to hypoxia than theorgans of old animals, i.e. the ability of hypoxia to induce
the release of [3H]CA decreases at 12 months of age andremains low at older ages. Figure 6(d) shows the meanrelease of CA (% of tissue content) evoked by a less intensehypoxic stimulus (10-min incubation with 5% O2-equili-brated solutions PO2 � 36 mmHg), being evidently an age-dependent behaviour identical to that exhibited in responseto the stronger hypoxic stimulus.
Contrary to the situation with hypoxia, the release of[3H]CA evoked by high external K+ (35 mM) in a new groupof CBs from 3-, 12-, 18- and 24-month-old rats is notstatistically different, whether the data refer to organ, unitweight or [3H]CA tissue content (Figs 7a–c). We have notperformed experiments testing the effects of hypercapnia onthe release of CA at different ages, because the small potencyof this stimulus to elicit a significant CA release response(see Fig. 5 in Vicario et al. 2000) will require a great numberof experiments, and consequently a disproportionate greaternumber of animals, to obtain meaningful data.
Effects of hypoxia and hypercapnia on the chemosensory
activity in the CSN in rats of different ages
The basal activity recorded in a few fibre preparations of theCSN of rats of different ages, although comparable, variedfrom preparation to preparation, indicating that a differentnumber of fibres were included in each recording. Meanbasal activity was 9.08 ± 1.51 (n ¼ 21), 6.58 ± 1.50 (n ¼8),8.17 ± 1.25 (n ¼ 13) and 9.43 ± 1.62 (n ¼ 23) Hz in con-trol, 12-, 18- and 24-month-old rats; therefore, to correct forthe variable number of fibres in each preparation, the activityelicited by any given stimulus was calculated as times thebasal activity. Figure 8(a) shows sample neurograms andtypical responses obtained in a control (3 months) and an 18-month-old preparation for the two intensities of hypoxiatested (0% and 5% O2), evidencing that, in the preparationsof the old animals, hypoxic stimuli elicit a smaller response.Below are shown PO2 recorded with a microelectrode placedat the side of the preparations. Figure 8(b) summarizes meanresponses elicited by the moderate and the intense hypoxicstimuli and by a hypercapnic stimulus (20% CO2, pH 6.8) inpreparations of the four ages studied. In comparison with thecontrol, hypoxia produced a significantly smaller CSNresponse in preparations of the CSN of 12-, 18- and 24-month-old rats, but the response to the hypercapnic-acidstimulus was statistically identical at all ages. For each givenstimulus, we also measured the latency of the response andthe time to peak response (data not shown). The latenciesranged from 13 to 21 s for the intense hypoxia and from 18to 22 s for the less intense hypoxia. The times to peak rangedbetween 39 and 54 s for the intense hypoxia and 38 and 59 sfor the moderate hypoxia. There were not any age-relatedstatistical differences in these two parameters. The responseto hypercapnic stimulus had lower latencies and times topeak than the hypoxic stimuli, 9–16 s and 24–48 s, respect-ively; there was an age-related significant difference in the
Fig. 5 Basal normoxic release of [3H]CA from chemoreceptor cells of
CBs of different ages. (a–c) Basal release corresponds to the d.p.m.
present in the sample immediately prior to the application of the
stimuli. In (a), when expressed as fmol/CB, there is a tendency for the
release to increase at 18 and 24 months, but this tendency is abol-
ished when data are normalized to unit weight (b) and to [3H]CA tissue
content. Data represent means ± SEM of 42 individual values
(3 months) and 24–30 for other ages.
Carotid body function in ageing 717
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723
latencies, being 9.20 ± 0.97 s in the control preparations and16.22 ± 1.55 and 15.62 ± 2.16 in 12- and 24-month-oldpreparations, respectively (p < 0.05).
Morphometric analysis
Aiming to correctly interpret the functional data describedin the preceding paragraphs, we have serial sectioned entireCBs from rats of different ages and in every section determinedthe percentage area stained for TH, a specific marker forchemoreceptor cells. The data are presented in Fig. 9. InFig. 9(a and b) the representative sections obtained arepresented, respectively, from CBs of 3- and 18-month-oldrats. It is evident that the relative sizes of the chemoreceptorclusters, as well as the area positive to TH, are smaller in theorgan of the older rat; in fact, in these particular sections thepercentage TH+ areas were 25% in Fig. 9(a) and 10% inFig. 9(b). In Fig. 9(c), we present mean data obtained in81 sections (two CBs) from 3-month-old rats, 173 sections(three CBs) from 12-month-old rats, 242 sections (five CBs)from 18-month-old rats and 266 sections (four CBs) from24-month-old rats. In the control CBs, the mean percentagearea TH+ represented 24.97 ± 0.71% of the total section area.We did not analyse more control CBs because, in a previousstudy (Prieto et al. 2004), using data from two CBs, thepercentage TH+ area found was 25.1 ± 0.80%. In the CBs of12-month-old rats there was a small, but significant, decrease
in TH+ area to 20.30 ± 0.53% of the total section area(p < 0.01). However, in the CBs of 18- and 24-month-old ratsthere was a very marked decrease in the area occupied byspecific glomic tissue, as the percentage TH+ area fell to12.75 ± 0.31 and 12.42 ± 0.46% (p < 0.001). An estimationof the CB parenchymatous cell loss can be obtained bymultiplying the percentage TH+ area times CB weight. For the3-month-old CBs, this product (50.22 lg/CB · 24.97%)amounts to 12.54 lg/CB, and for the CB of 12-, 18- and 24-month-old rats this figure drops, respectively, to 9.32, 9.53 and8.61 lg/CB.
Discussion
The effects of ageing on the CB arterial chemoreceptors werestudied using quantitative morphological techniques, asses-sing chemoreceptor cell function through the dynamics oftheir neurotransmitters and measuring functionality of theintact organ as chemosensory activity in the CSN. We havefound: (i) CBs enlarge in rats of 18 months or older, buta decrease in parenchymatous tissue is evident after12 months; (ii) CA content and turnover time of CBsincrease with age, these increases being more marked after18 months; (iii) hypoxic stimulation of chemoreceptor cells,but not stimulation with a non-specific depolarizing stimulus,elicits a smaller release of CA in rats of 12 months and older;
Fig. 6 Hypoxic release of [3H]CA from chemoreceptor cells of CBs of
different ages. (a, b and c, respectively) Mean release in response to a
2% O2 hypoxic stimulus expressed per organ or normalized to unit
weight and to [3H]CA tissue content. Note that in all instances there is
marked decrease in the hypoxic response of chemoreceptor cells of
aged animals in comparison with control, 3-month-old rats, except for
the release at 24 months when release is expressed by organ (a); this
was as a result of the presence in this group of two anomalously big
CBs. (d) Mean release evoked by a more moderate hypoxic stimulus
(10-min incubation with a solution equilibrated with 5% O2). At this
hypoxic intensity there also was the same type of reduction of che-
moreceptor cell response. Data represent means ± SEM of 8–10
individual values. *p < 0.05, **p < 0.01.
718 S. V. Conde et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723� 2006 The Authors

(iv) CSN chemosensory activity elicited by hypoxia, butnot hypercapnic acidosis, decreases significantly after12 months. In the following paragraphs we will discuss: (i)our morphometric findings in relation to the general ageing
process and to data available on the CB; (ii) the meaning ofthe CA dynamics in the CB, emphasizing the loss of theability of chemoreceptor cells to respond to hypoxia; (iii) andwe will relate our findings on the chemosensory activity inthe CSN to ventilation in aged mammals; and, finally, (iv) wewill discuss the significance of our findings for the use of CBautographs to treat Parkinson’s disease.
Ageing is the sum of changes that occur in a livingorganism with the passage of time leading to functionalimpairment and death (Timiras 1972). Although not identi-cal, age-related changes share common traits in all organsand include a loss of parenchymatous cells, an increase inconnective tissue/collagen deposition and a decrease in bloodflow and functional performance (Muhlberg and Platt 1999;De Souza 2002; Sato et al. 2002; Serot et al. 2003). We havefound that there is an age-related decrease in chemoreceptorcell mass that, in the rat CB, occurs within the first year ofage. Previous studies on age-dependent changes in themorphometry of the CBs yield comparable conclusions,although in all of them the quantification was derived from areduced number of sections. For example, Hurst et al. (1985;see also Heath and Smith 1991) in haematoxylin/eosin-stained sections (‘five sections cut at levels through the blockand selecting the one with the largest area’) of human CBobtained post-mortem estimated that, in young humans (14–40 years), the parenchyma occupied 45% of the sectionsarea; in middle-aged (41–65 years) and old (66–100 years)the percentages decreased to 39 and 29%, respectively.Considering that blood vessels (mostly capillaries) in well-perfused CB occupy around 20–25% (McDonald 1981),most of the discrepancy in the absolute percentages ofparenchymatous tissue between rat and human appear to beas a result of the lack of perfusion in the study of Hurst et al.(1985). Hurst et al. (1985) also found that mean maximalarea of the CB sections increased by a factor of 1.6 in the oldgroup, being mainly as a result of an increase in connectivetissue; once again their findings match the increase in CBweight that we have determined well (Fig. 1). In a qualitativestudy, Di Giulio et al. (1998, 2003) also concluded that inageing there is a reduction of chemoreceptor cell volume andan increase in extracellular matrix. However, Partanen et al.(1984) reported that CB cell number does not change in ratsbetween 3 and 33 months; yet the number of cells/CB givenis around 250, a meaningless figure, as the estimated numberof chemoreceptor cells in the rat CB is �10 000 (McDonald1981).
The age-dependent changes in the metabolism of CAobserved in the CB have several aspects which deservecomment. The CB of rats 18 months and older have a highercontent and a higher concentration of DA than the organ ofthe control, the 3-month-old rats (Fig. 2). This high DAcontent is associated with a rate of DA synthesis (turnoverrate) which, when expressed by organ tends to be higher, butwhen expressed by unit weight is similar to that in control
Fig. 7 Release of [3H]CA induced by high external K+ in chemore-
ceptor cells of CBs of different ages. Release experiments using high
external K+ followed a protocol identical to that used for hypoxia
(Fig. 4), except for the stimulus period incubation which consisted of a
solution containing 35 mM KCl. (a, b and c, respectively) Mean release
in response to 35 mM KCl expressed per organ or normalized to
unit weight and to [3H]CA tissue content. There were not statistical
differences in any form of expressing the data. Data represent
means ± SEM of 8–10 individual values.
Carotid body function in ageing 719
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723

CBs (Fig. 3). The result is that the DA turnover timeincreases with age (Fig. 3c). Paralleling the rate of synthesis,the basal normoxic release of CA tends to be higher in theCBs from 18- and 24-month-old rats, but normalization ofdata by unit weight or tissue content abolishes the differ-ences. This observation on the basal release was expected
from the synthesis experiments. In steady-state conditions,synthesis replenishes from instant to instant what is beingused (released). Using radiometric methods, as we did, theabsolute rate of synthesis (31 pmol/mg/h for control CBs;Fig. 3b) exceeds the rate of release (350 fmol/mg/10 min ¼2.1 pmol/mg/h; Fig. 4b) because it is measured the actual
Fig. 8 Age-dependent decrease in the magnitude of the carotid sinus
nerve response to hypoxic stimulation and the absence of differences
in the responses to hypercapnic acidosis. (a) Sample neurograms
(top) at times and preparations selected as indicated in the records of
the integrated activity in the carotid sinus nerve responses (middle
row) at PO2 shown below (lower row). Note the marked differences in
the hypoxic responses recorded in preparations from control (3-month-
old) versus aged (18-month-old) rats. Note also that, in the control
preparations (middle row, left), the response elicited by the intense
hypoxic stimulus adapts. (b) Mean responses to intense and moderate
hypoxia and to hypercapnic acidosis obtained in preparations of rats of
different ages. Data represent means ± SEM of n individual values
given in the drawing. *p < 0.05, **p < 0.01.
720 S. V. Conde et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723� 2006 The Authors

rate of synthesis (the only precursor available is [3H]tyrosine)and only [3H]CA are quantified when the CB is releasingCA + [3H]CA.
CBs of 12-month and older rats exhibit a marked decrease inthe release of [3H]CA induced by hypoxia (Fig. 6). When a
comparison is made with the release elicited by high externalK+, this difference is more striking and gains particularsignificance (Fig. 7). The schema of the hypoxic transductionin chemoreceptor cells is as follows: (i) lowPO2, (ii) O2 sensor,(iii) coupling mechanisms, (iv) O2-sensitive K+ channelsinhibition; (v) chemoreceptor cell depolarization, (vi) activa-tion of voltage-operated Ca2+ channels, (vii) increase inintracellular Ca2+, (viii) exocytosis of neurotransmitters, (ix)increase in the CSN chemosensory activity and (x) increase inventilation (Gonzalez et al. 1992; He et al. 2005; Lahiri et al.2005). Comparison of hypoxic and high K+ responsesindicates that the age-related alteration resides in a step ofthe transduction cascade prior to chemoreceptor cell depolar-ization, i.e. ageing reduces the gain made by the early steps ofthe hypoxic transduction cascade. Uncertainties on themolecular nature of steps 2 and 3 and multiplicity of O2-sensitive K+ channels, renders endeavours in these directionsout of the scope of the present study.
Consistent with [3H]CA release findings, there areminimal age-related differences in the normoxic CSNactivity and a marked decrease in the hypoxic responses in12-month and older ages (Fig. 8b). Thus, a decrease in thecapacity of chemoreceptor cells to respond to hypoxia istranslated into a parallel decrease in the CB signal impinginginto the brainstem to drive ventilation. However, the decreasein the CSN response to hypoxia exceeds the decrease inhypoxic ventilation observed by Soulage et al. (2004). Threefactors may contribute to this difference: (i) the greatredundancy of the CB drive (unilateral CB denervation doesnot affect the ventilatory response to hypoxia; Busch et al.1983; Cragg and Khrisanapant 1994; Prieto-Lloret et al.2004); (ii) age-related changes at brainstem level integration(Mukai et al. 2003) can compensate the reduced drive fromthe CB chemoreceptors; (iii) additional age-dependent var-iables (efferent, neural or muscular, components of thechemoreflex arch or thoracic skeleton) might change the finalventilatory output. Chemosensory activity elicited by acidic-hypercapnic stimulus in the CSN does not show age-dependent differences, suggesting different hypoxic andacidic transductions mechanisms (Gonzalez et al. 1992,1994). Although out of the scope of the present study, wewant to point out that clear differences in the transductioncascades for hypoxic and hypercapnic stimuli have beennoted. For example, hypoxic stimuli are more powerful thanhypercapnic stimuli in eliciting activity in the CSN (Fitz-gerald and Parks 1971; see Fig. 8). This difference at thelevel of CSN activity is paralleled by a lower potency of thehypercapnic stimuli to elicit release of CA (Vicario et al.2000) and to increase the levels of cAMP in the CB (Perez-Garcia et al. 1990). Finally, other differences in the trans-duction cascade for both stimuli, such as Ca2+-dependencyon the release of CA (Obeso et al. 1992) and the subtypes ofvoltage-operated Ca2+ channels involved (Rocher et al.2005) have been noticed. Therefore, it is not surprising that
Fig. 9 Quantification of the tyrosine hydroxylase-positive areas
in sections of CBs of different ages. (a, b) Two sample sections
stained for tyrosine hydroxylase obtained from CBs of a control rat
(3 months old; a) and an aged rat (18 months old; b). Note the small
sizes of the clusters of parenchymatous cells and the decrease in
stained area (10 vs. 25%) in the section of the older CB. (c) Mean
percentages of tyrosine hydroxylase-positive areas present in serially
sectioned CBs (two, three, five and four CBs from 3-, 12-, 18- and
24-month-old rats, respectively). Data represent means ± SEM.
**p < 0.01, ***p < 0.001.
Carotid body function in ageing 721
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723

ageing causes differential modifications in the response tohypoxic and hypercapnic stimuli.
The present data evidence once again the parallelismexisting between the release of CA (mostly DA) bychemoreceptor cells and the level of activity in the CSN.Based on observations comparable with those presented inthis study, namely, that for every experimental situation andfor every stimuli tested there is an extraordinary parallelismbetween the release of DA and the activity in the CSN, ourlaboratory has championed the view that DA may act as theneurotransmitter between chemoreceptor cells and CSNsensory endings (Gonzalez et al. 1994). Yet, many laborat-ories have shown that, in spite of these parallelisms, the useof DA agonists and antagonists do not support the role of DAas the instant-to-instant neurotransmitter at this sensorysynapse (Zapata 1997; and see Gonzalez et al. 2003). Wecould speculate that DA released in direct proportion to thestimulus intensity might help to match the metabolic rate ofchemoreceptor cells to their level of activity, probably bycontributing the functional hyperaemia (Fidone and Gonz-alez 1986) or to fine-tuning oxidative metabolism (Maxwellet al. 1985; Gluck et al. 2002) of CB cells.
An additional interest to our study derives from the useof CB tissue autoimplants to treat Parkinson’s disease (seeFernandez-Espejo et al. 2005), which is typically an illnessof the elderly. Grafts of CB cells have been found to beeffective in ameliorating functional deficits in rodent andprimate models of Parkinson’s disease developed in younganimals (there are no studies in older animals), but theonly clinical study available revealed a limited improve-ment in patients grafted with autologous CB fragments(Arjona et al. 2003; Fernandez-Espejo et al. 2005). Thereduced amelioration of symptoms in the auto-implantedpatients might result from the limited capacity of chemo-receptor cells from aged animals to release DA in responseto hypoxia (normal brain PO2 represents a moderate/intense hypoxic stimulus for chemoreceptor cells; Obesoet al. 1997). In addition, the situation in patients may beworse because Parkinson’s disease affects basically allaminergic systems (Harnois and Di Paolo 1990; Bosboomet al. 2003), making it plausible that chemoreceptor cellsalso are altered; this latest suggestion is supported by thereduced ventilatory response to hypoxia, with normalventilation to hypercapnia observed in Parkinson’s diseasepatients (Onodera et al. 2000).
Acknowledgements
We would like to thank Ma de los Llanos Bravo for technical
assistance. The work was supported by DGICYT grant BFU2004-
06394, by FISS grant PI042462, by JCyL grant VA011C05 and by
an AI grant (MEC Spain/CRUP Portugal-HP03-26). SVC is funded
by a PhD grant from the Portuguese Foundation for Science and
Technology (FCT).
References
Arjona V., Minguez-Castellanos A., Montoro R. J. et al. (2003) Auto-transplantation of human carotid body cell aggregates for treatmentof Parkinson’s disease. Neurosurgery 53, 321–328.
Behan M., Zabka A. G. and Mitchell G. S. (2002) Age and gendereffects on serotonin-dependent plasticity in respiratory motorcontrol. Respir. Physiol. Neurobiol. 13, 65–77.
Bisgard G. E., Forster H. V., Orr J. A., Buss D. D., Rawlings C. A. andRasmussen B. (1976) Hypoventilation in ponies after carotid bodydenervation. J. Appl. Physiol. 40, 184–190.
Bosboom J. L., Stoffers D. and Wolters E. Ch (2003) The role of acet-ylcholine and dopamine in dementia and psychosis in Parkinson’sdisease. J. Neural Transm. Suppl. 65, 185–195.
Busch M. A., Bisgard G. E., Mesina J. E. and Forster H. V. (1983) Theeffects of unilateral carotid body excision on ventilatory control ingoats. Respir. Physiol. 54, 353–361.
Cerveri I., Zoia M. C., Fanfulla F., Spagnolatti L., Berrayah L., GrassiM. and Tinelli C. (1995) Reference values of arterial oxygen ten-sion in the middle-aged and elderly. Am. J. Respir. Crit. Care Med.152, 934–941.
Chan E. D. and Welsh C. H. (1998) Geriatric respiratory medicine. Chest114, 1704–1733.
Cragg P. A. and Khrisanapant W. (1994) Is the second carotid bodyredundant? Adv. Exp. Med. Biol. 360, 297–299.
De Souza R. R. (2002) Aging of myocardial collagen. Biogerontology 3,325–335.
Di Giulio C., Di Muzio M., Sabatino G., Spoletini L., Amicarelli F., DiIlio C. and Modesti A. (1998) Effect of chronic hyperoxia onyoung and old rat carotid body ultrastructure. Exp. Gerontol. 33,319–329.
Di Giulio C., Cacchio M., Bianchi G., Rapino C. and Di Ilio C. (2003)Selected contribution: Carotid body as a model for aging studies: isthere a link between oxygen and aging? J. Appl. Physiol. 95, 1755–1758.
Fernandez-Espejo E., Armengol J. A., Flores J. A., Galan-Rodriguez B.and Ramiro S. (2005) Cells of the sympathoadrenal lineage:biological properties as donor tissue for cell-replacementtherapies for Parkinson’s disease. Brain Res. Brain Res. Rev. 49,343–354.
Fidone S. and Gonzalez C. (1982) Catecholamine synthesis in rabbitcarotid body in vitro. J. Physiol. 333, 69–79.
Fidone S. and Gonzalez C. (1986) Peripheral chemoreceptors: initiationand control of discharges, in Handbook of Physiology. The Res-piratory System II (Fishman, A. P., ed.), pp. 247–312. AmericanPhysiology Society, Bethesda, MD.
Fidone S., Gonzalez C. and Yoshizaki K. (1982) Effects of low oxygenon the release of dopamine from the rabbit carotid body in vitro.J. Physiol. 333, 93–110.
Fitzgerald R. S. and Parks D. C. (1971) Effect of hypoxia on carotidchemoreceptor response to carbon dioxide in cats. Respir. Physiol.12, 218–229.
Fordyce W. E. (1987) Hypoxic ventilatory control in the awake cat5 years after carotid body resection. Respir. Physiol. 69,209–225.
Forster H. V., Pan L. G., Lowry T. F., Serra A., Wenninger J. andMartino P. (2000) Important role of carotid chemoreceptor afferentsin control of breathing of adult and neonatal mammals. Respir.Physiol. 119, 199–208.
Gluck M., Ehrhart J., Jayatilleke E. and Zeevalk G. D. (2002) Inhibitionof brain mitochondrial respiration by dopamine: involvement ofH2O2 and hydroxyl radicals but not glutathione-protein-mixeddisulfides. J. Neurochem. 82, 66–74.
722 S. V. Conde et al.
Journal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723� 2006 The Authors

Gonzalez E., Rigual R., Fidone S. J. and Gonzalez C. (1987) Mecha-nisms for termination of the action of dopamine in carotid bodychemoreceptors. J. Auton. Nerv. Syst. 18, 249–259.
Gonzalez C., Almaraz L., Obeso A. and Rigual R. (1992) Oxygen andacid chemoreception in the carotid body chemoreceptors. TrendsNeurosci. 15, 146–153.
Gonzalez C., Almaraz L., Obeso A. and Rigual R. (1994) Carotid bodychemoreceptors: from natural stimuli to sensory discharges.Physiol. Rev. 74, 829–898.
Gonzalez C., Rocher R. and Zapata P. (2003) Arterial chemoreceptors:cellular and molecular mechanisms in the adaptative and homeo-static function of the carotid body. Rev. Neurol. 36, 239–254.
Gunnarsson L., Tokics L., Brismar B. and Hedenstierna G. (1996)Influence of age on circulation and arterial blood gases in man.Acta Anaesthesiol. Scand. 40, 237–443.
Harnois C. and Di Paolo T. (1990) Decreased dopamine in the retinas ofpatients with Parkinson’s disease. Invest. Ophthalmol. Vis. Sci. 31,2473–2475.
He L., Dinger B., Sanders K., Hoidal J., Obeso A., Stensaas L., FidoneS. and Gonzalez C. (2005) Effect of p47phox gene deletion onROS production and oxygen sensing in mouse carotid bodychemoreceptor cells. Am. J. Physiol. Lung Cell Mol. Physiol. 289,L916–L924.
Heath D. and Smith P. (1991) Histological changes associated withageing of the adult carotid body, in Diseases of the Human CarotidBody. (Heath, D. and Smith, P., eds), pp. 25–22. Springer-Verlag,London.
Honda Y. (1985) Role of carotid chemoreceptors in control of breathingat rest and in exercise: studies on human subjects with bilateralcarotid body resection. Jpn J. Physiol. 35, 535–544.
Hurst G., Heath D. and Smith P. (1985) Histological changes associatedwith ageing of the human carotid body. J. Pathol. 147, 181–187.
Janssens J. P., Pache J. C. and Nicod L. P. (1999) Physiological changesin respiratory function associated with ageing. Eur. Respir J. 13,197–205.
Knapowski J., Wieczorowska-Tobis K. and Witowski J. (2002) Patho-physiology of ageing. J. Physiol. Pharmacol. 53, 135–146.
Lahiri S., Roy A., Baby S. M., Hoshi T., Semenza G. L. and PrabhakarN. R. (2005) Oxygen sensing in the body. Prog. Biophys. Mol.Biol. (published online ahead of print).
Loeschcke H. H. (1974) Central nervous chemoreceptors, in RespiratoryPhysiology (Widdicombe, J. G., ed.), pp. 167–196. Butterworths,London.
Maxwell G., Crompton S. and Smyth C. (1985) The effect of dopamineupon oxidative metabolism of brown fat adipocytes. Eur. J.Pharmacol. 116, 293–297.
McDonald D. M. (1981) Peripheral chemoreceptors. Structure–functionrelationships of the carotid body, in Regulation of Breathing(Hornbein, T. F., ed.), pp. 105–319. Marcel Dekker, New York.
Muhlberg W. and Platt D. (1999) Age-dependent changes of the kidneys:pharmacological implications. Gerontology 45, 243–253.
Mukai S., Gagnon M., Iloputaife I., Hamner J. W. and Lipsitz L. A.(2003) Effect of systolic blood pressure and carotid stiffness onbaroreflex gain in elderly subjects. J. Gerontol. A Biol. Sci. Med.Sci. 58, 626–630.
Obeso A., Rocher A., Fidone S. and Gonzalez C. (1992) The role ofdihydropyridine-sensitive Ca2+ channels in stimulus-evoked cate-cholamine release from chemoreceptor cells of the carotid body.Neuroscience 47, 463–472.
Obeso A., Rocher A., Herreros B. and Gonzalez C. (1997) Oxygenconsumption and energy metabolism in the carotid body, in TheCarotid Body Chemoreceptors (Gonzalez, C., ed.), pp. 28–43.Springer-Verlag, Berlin, Germany.
Onodera H., Okabe S., Kikuchi Y., Tsuda T. and Itoyama Y. (2000)Impaired chemosensitivity and perception of dyspnoea in Parkin-son’s disease. Lancet 356, 739–740.
Partanen M., Linnoila I., Hervonent A. and Rapoport S. I. (1984) Theeffect of aging on extra-adrenal catecholamine storing cells ofthe rat. Neurobiol. Aging 5, 105–110.
Perez-Garcia M. T., Almaraz L. and Gonzalez C. (1990) Effects ofdifferent types of stimulation on cyclic AMP content in the rabbitcarotid body: functional significance. J. Neurochem. 55, 1287–1293.
Phillipson E. A. and Kozar L. F. (1993) Effect of aging on metabolicrespiratory control in sleeping dogs. Am. Rev. Respir. Dis. 147,1521–1525.
Pokorsky M., Walski M., Dymecka A. and Marczak M. (2004) Theaging carotid body. J. Physiol. Pharmacol. 55, 107–113.
Prieto-Lloret J., Caceres A. I., Obeso A. et al. (2004) Ventilatory re-sponses and carotid body function in adult rats perinatally exposedto hyperoxia. J. Physiol. 554, 126–144.
Rigual R., Gonzalez E., Gonzalez C. and Fidone S. (1986) Synthesis andrelease of catecholamines by the cat carotid body in vitro: effects ofhypoxic stimulation. Brain Res. 374, 101–109.
Rigual R., Lopez-Lopez J. R. and Gonzalez C. (1991) Release ofdopamine and chemoreceptor discharge induced by low pH andhigh PCO2 stimulation of the cat carotid body. J. Physiol. 433,519–531.
Rigual R., Rico A. J., Prieto-Lloret J., de Felipe C., Gonzalez C. andDonnelly D. F. (2002) Chemoreceptor activity is normal inmice lacking the NK1 receptor. Eur. J. Neurosci. 16, 2078–2084.
Rocher A., Geijo-Barrientos E., Caceres A. I., Rigual R., Gonzalez C.and Almaraz L. (2005) Role of voltage-dependent calcium chan-nels in stimulus-secretion coupling in rabbit carotid body chemo-receptor cells. J. Physiol. 562, 407–420.
Sato A., Sato Y. and Uchida S. (2002) Regulation of cerebral corticalblood flow by the basal forebrain cholinergic fibers and aging.Auton. Neurosci. 96, 13–19.
Serot J. M., Bene M. C. and Faure G. C. (2003) Choroid plexus,aging of the brain, and Alzheimer’s disease. Front. Biosci. 8,515–521.
Sorbini C. A., Grassi V., Solinas E. and Muiesan G. (1968) Arterialoxygen tension in relation to age in healthy subjects. Respiration25, 3–13.
Soulage C., Pequignot J. M. and Perrin D. (2004) Breathing pattern andhypoxic sensitivity during ageing in a new model of obesity-resistant rat. Respir. Physiol. Neurobiol. 144, 45–57.
Timiras P. S. (1972) Development Physiology and Aging, pp. 408–461.Macmillan Co., New York.
Vicario I., Rigual R., Obeso A. and Gonzalez C. (2000) Characteri-zation of the synthesis and release of catecholamine in the ratcarotid body in vitro. Am. J. Physiol. Cell Physiol. 278, C490–C499.
Zapata P. (1997) Chemosensory activity in the carotid nerve: effects ofpharmacological agents, in The Carotid Body Chemoreceptors(C. Gonzalez, ed.), pp. 119–146. Springer-Verlag, Heidelberg,Germany.
Carotid body function in ageing 723
� 2006 The AuthorsJournal Compilation � 2006 International Society for Neurochemistry, J. Neurochem. (2006) 99, 711–723




















