
Frozen fish and cephalopod molluscs for fish bait - Ministry for ...
176
Supplementary Import Risk Analysis: Frozen fish and cephalopod molluscs for fish bait Approved for IHS development Prepared for Ministry for Primary Industries By the Animal Risk Assessment Team, Risk Assessment Group, Animal and Plant Health Directorate, Biosecurity New Zealand, Ministry for Primary Industries ISBN No:978-1-004381-9 (online) November 2020
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Frozen fish and cephalopod molluscs for fish bait - Ministry for ...

Supplementary Import Risk Analysis: Frozen fish and cephalopod molluscs for fish bait
Approved for IHS development
Prepared for Ministry for Primary Industries By the Animal Risk Assessment Team, Risk Assessment Group, Animal and Plant Health Directorate, Biosecurity New Zealand, Ministry for Primary Industries ISBN No:978-1-004381-9 (online) November 2020


Disclaimer
While every effort has been made to ensure the information in this publication is accurate, the Ministry for Primary Industries does not accept any responsibility or liability for error of fact, omission, interpretation or opinion that may be present, nor for the consequences of any decisions based on this information. Cover photo: Atlantic herring (Clupea harengus) (Photo credit: NOAA, Wikimedia Commons) Recommended citation: MPI (2020) Supplementary Import Risk Analysis: Frozen fish and cephalopod molluscs for fish bait. Ministry for Primary Industries, New Zealand. This publication is available on the Ministry for Primary Industries website at http://www.mpi.govt.nz/news-and-resources/publications/ © Crown Copyright - Ministry for Primary Industries


Supplementary Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Version 4.0
19 November 2020
Approved for IHS development
Enrico Perotti
Associate Director Animal and Plant Health, Risk Assessment Group,
Biosecurity New Zealand

ii Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Version information
Version number Comments Date of release
1.0 First draft version October 2019
1.1 Internally peer reviewed version January 2020
2.0 Peer-reviewed version March 2020
3.0 Revised version for second peer review May 2020
3.1 Revised version – Extended frozen storage September 2020
4.0 Approved for IHS development November 2020
New Zealand is a member of the World Trade Organisation and a signatory to the Agreement on the Application of Sanitary and Phytosanitary Measures (The Agreement). Under the Agreement, countries must base their measures on an International Standard or an assessment of the biological risks to plant, animal or human health.
This document extends and supplements a previous risk analysis (eviscerated or trunked fish for human consumption) (Blackwell 2019), to examine the risks associated with frozen fish and cephalopod molluscs imported for use as fish bait in commercial and recreational fishing. It provides a scientific analysis of the likelihood of entry, exposure, establishment and spread of various pathogens and diseases in the commodity and assesses the consequences of their establishment in New Zealand. The document has been internally and externally peer reviewed.

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait iii
Contributors to this risk analysis
The following people provided significant input into the development of this risk analysis:
Primary contributor
Ron Blackwell Senior Adviser, Animal Risk Assessment Team, Risk Assessment Group
Biosecurity New Zealand, Ministry for Primary Industries, Wellington, New Zealand
Internal Peer Review
Sudharma Leelawardana (Sue)
Manager, Animal Risk Assessment Team. Risk Assessment Group
Biosecurity New Zealand, Ministry for Primary Industries, Wellington, New Zealand
Don Leelawardana Senior Adviser, Animal Risk Assessment Team, Risk Assessment Group
Biosecurity New Zealand, Ministry for Primary Industries, Wellington, New Zealand
Shahid Haneef Senior Adviser, Animal Risk Assessment Team, Risk Assessment Group
Biosecurity New Zealand, Ministry for Primary Industries, Wellington, New Zealand
Dan Kluza Principal Adviser, Risk Assessment Group
Biosecurity New Zealand, Ministry for Primary Industries, Wellington, New Zealand
Vicki Melville
Manager, Animal Imports Team 1, Risk Assessment Group
Biosecurity New Zealand, Ministry for Primary Industries, Wellington, New Zealand
Nasser Ahmed Senior Adviser, Animal Imports Team 1, Risk Assessment Group
Biosecurity New Zealand, Ministry for Primary Industries, Wellington, New Zealand
External Scientific Review
Ramesh Perera Biosecurity consultant Griffith, ACT, Australia

iv Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Table of Contents
Index of Tables and Figures vi
Acronyms and abbreviations vii
Executive summary 1
Introduction 3 2.1 The importance of fish bait in New Zealand 5 2.2 The importance of aquaculture 7
Scope, commodity definition and assumptions 9 3.1 Scope 9 3.2 Commodity definition 10 3.3 Assumptions 12
Risk analysis methodology 13 4.1 General procedures 13
Hazard identification 16 5.1 Hazard list for risk assessment 35
General considerations 35 6.1 Risk assessment considerations 35 6.2 Risk management considerations 39
Birnaviridae: Marine aquabirnavirus 43 7.1 Technical review 43 7.2 Risk assessment 47 7.3 Risk management 48
Iridoviridae: Megalocytivirus (red sea bream iridovirus - RSIV) and associated viruses 50 8.1 Technical review 50 8.2 Risk assessment 52 8.3 Risk management 54
Iridoviridae: Erythrocytic necrosis virus (ENV) 55 9.1 Technical review 55 9.2 Risk assessment 57 9.3 Risk management 59
Nodaviridae: Nervous necrosis virus (NNV) 60 10.1 Technical review 60 10.2 Risk assessment 64 10.3 Risk management 66
Orthomyxoviridae: Infectious salmon anaemia virus (ISAV) 67 11.1 Technical review 67 11.2 Risk assessment 70 11.3 Risk management 71
Reoviridae: Piscine aquareovirus (PRV) and associated aquareoviruses 72 12.1 Technical review 72 12.2 Risk assessment 74 12.3 Risk management 75
Rhabdoviridae: Infectious haematopoietic necrosis virus (IHNV) 75 13.1 Technical review 75 13.2 Risk assessment 77 13.3 Risk management 78
Rhabdoviridae: Viral haemorrhagic septicaemia virus (VHSV) 80 14.1 Technical review 80 14.2 Risk assessment 83 14.3 Risk management 85

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait v
Edwardsiella spp. 86 15.1 Technical review 86 15.2 Risk assessment 89 15.3 Risk management 91
Francisella spp. 92 16.1 Technical review 92 16.2 Risk assessment 94 16.3 Risk management 94
Pseudomonas anguilliseptica 94 17.1 Technical review 94 17.2 Risk assessment 97 17.3 Risk management 97
Streptococcus spp. (S. agalactiae serotype III: 283, S. iniae) 97 18.1 Technical review 97 18.2 Risk assessment 101 18.3 Risk management 103
Myxozoan pathogens 104 19.1 Technical review 104 19.2 Risk assessment 110 19.3 Risk management 112
References 113
Appendices 156

vi Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Index of Tables and Figures
Table 1. Estimated total number of hooks per year set in commercial long lining, by vessel size group, for fishing years (1 Oct to 30 Sept) 2002-03, 2012-13 and 2018-19 5
Table 2. Estimated number of recreational fishing trips using fish bait during 2017-18 6
Table 3. Estimated volume (t) and source location of finfish imported as fish bait, 2018 8
Table 4. Estimated volume (t) of coleoid cephalopod molluscs imported as fish bait, 2018 9
Table 5. Finfish and coleoid cephalopod mollusc families and species considered as fish bait 12
Table 6. Hazard Identification table for specified1 wild marine finfish and coleoid molluscs used as fish bait17
Table 7. Fish bait species susceptible to marine aquabirnavirus (MABV) 45
Table 8. Fish bait species susceptible to red sea bream iridovirus (RSIV), infectious spleen and kidney necrosis virus (ISKNV) and associated iridoviruses 51
Table 9. Fish bait species susceptible to erythrocytic necrosis virus (ENV) 56
Table 10. Fish and coleoid cephalopod mollusc species susceptible to nervous necrosis virus (NNV) 63
Table 11. Fish bait species susceptible to strains of infectious salmon anaemia virus (ISAV) 69
Table 12. Fish bait species susceptible to piscine aquareovirus and related strains 73
Table 13. Fish bait species susceptible to infectious haematopoietic necrosis virus (IHNV) 76
Table 14. Fish bait species susceptible to viral haemorrhagic septicaemia virus (VHSV) 81
Table 15. Fish bait species susceptible to Edwardsiella spp. 88
Table 16. Fish bait species susceptible to Francisella spp. 93
Table 17. Fish bait species susceptible to Pseudomonas anguilliseptica 95
Table 18. Fish bait species susceptible to Streptococcus agalactiae III: 283 99
Table 19. Fish bait species susceptible to Streptococcus iniae 99
Table 20. Fish species susceptible to exotic myxozoan pathogens Enteromyxum leei, Kudoa clupeidae, K. iwatai. K. nova. K. thyrsites 107
Table 21. Taxonomy and geographical distribution of finfish imported as fish bait, 2008-2017 156
Table 22. Classification and distribution of coleoid cephalopod molluscs imported, 2008-2017 158
Table 23. Imports of coleoid cephalopod molluscs, 2013-2015 160
Table 24. Risk organisms and associated fish bait species where specific risk management measures are proposed 161
Table 25. Assumed reduction in pathogen load associated with risk management options 164
Figure 1. The general risk analysis process 14
Figure 2. Potential distribution pathways for frozen fish or coleoid cephalopod mollusc fish bait 38

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait vii
Acronyms and abbreviations
Term/Acronym Definition
Bait fish Whole, frozen, small sized, generally schooling pelagic finfish (e.g. herring, mackerel) imported for use as fish bait.
Berley Minced fish and fish offal that is scattered on the water or on the seabed to attract fish. Berley may be combined with bread, flour and additional attractants such as fish oil. Berley is also known as chum, or ground-bait.
Biosecurity New Zealand
A business unit of the Ministry for Primary Industries, New Zealand.
CFU Colony forming units of bacteria.
CIHEAM International Centre for Advanced Mediterranean Agronomic Studies.
Commercially prepared and packaged
A product that has been manufactured in a commercial manner by a commercial enterprise and is packaged in tamper-proof packaging (MPI 2011).
Competent Authority Government veterinary authority or other Government authority having the responsibility and competence for ensuring or supervising the implementation of animal health and welfare measures, international veterinary certification and other standards and recommendations in the OIE Code and Aquatic Code.
Derived from marine waters
Organisms derived from marine waters only (i.e. excluding species that spend some, or all of their life cycle in fresh water, such as salmonids).
EFSA European Food Safety Authority.
Epizootic An outbreak of a rapidly spreading disease that is temporarily prevalent and widespread in a population.
ESR Environmental Science and Research, Crown Research Institute Ltd., New Zealand.
Eviscerated finfish Bony fish of Family Actinopterygii which have been processed to remove the viscera (internal organs) but where the head and/or gills may be attached.
FAO Food and Agriculture Organisation of the United Nations, Rome, Italy.
Farm sourced All organisms held in confinement at any point in their life cycle, including stock collected from the wild and then on-grown or held in confinement.
Finfish Poikilothermic vertebrates that breathe by gills, including bony fish of Family Actinopterygii and cartilaginous fish of Class Chondrichthyes.
Fish A general term which includes finfish, as well as other vertebrates and invertebrate organisms.
Fish bait Whole, frozen marine bait fish (see above), small tuna, coleoid cephalopod molluscs (squid, cuttlefish and octopus) imported for use as bait. Fish bait includes eviscerated fish or coleoid cephalopod molluscs originally imported for use for human consumption but downgraded for use as bait.
FishBase The FishBase Project: World online database for fish species (2019).
Free of disease Fish and coleoid molluscs with no external signs of infection, which have not been harvested following an epizootic incident, or as disease control.

viii Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Term/Acronym Definition
IHS Import Health Standard.
Marine fish Any species of fish that does not spend any part of its life cycle in fresh water. Salmon, which spawn in fresh water, are NOT a marine fish. Brackish water is considered to be part of the marine environment (MPI 2011).
MPI Ministry for Primary Industries, New Zealand.
Meltwater Water released from the melting of ice, including frozen fish bait.
NIWA National Institute for Water and Atmospheric Research, Crown Research Institute, New Zealand.
Non-viable Organism is not capable of living, growing or reproducing
Not cause significant disease
Pathogen is benign, causing no clinical signs of disease in the host species and is not a reservoir host for another pathogenic organism.
Opportunistic pathogen
Only causes clinical disease where the host is weakened or stressed, or infection only occurs as a result of physical injury.
OIE World Organisation for Animal Health, Paris, France.
RMP Risk management proposal.
SeaLifeBase The SeaLife Project. World database for non-fish marine species.
Shelf-stable Not requiring refrigeration or freezing before opening (MPI 2011).
SPS Agreement Agreement on the Application of Sanitary and Phytosanitary Measures.
Trunked cartilaginous fish
Processed fish of Class Chondrichthyes, where the viscera, gills and head have been removed.
Wild-sourced Organisms collected from the wild. This excludes finfish and cephalopods that have been farmed or that have otherwise spent any part of their life cycle in confinement.
WoRMS World online Register of Marine Species (2019).
WTO World Trade Organisation, Geneva, Switzerland.

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 1
Executive summary
This supplementary import risk analysis (SRA) reviews the biosecurity risks associated with
imported frozen fish bait. This commodity includes non-viable frozen, whole (uneviscerated)
wild-caught marine finfish (Class Actinopterygii) and molluscs (Subclass Coloidea, Class
Cephalopoda - cuttlefish, octopus and squid). It is restricted to specified species that were
imported for use as fish bait in 2018, as listed in Table 5.
Fish bait is a non-human food item, caught, stored and traded as a bulk commodity. The current
import requirements are defined in the Import Health Standard (IHS) for “Fish bait and fish
food” (FISFOOIC.ALL) (MPI 2011). This commodity must be wild caught, non-viable marine
finfish, either frozen at -18 °C for 168 hours (7 days) or treated with ionising radiation (at 25
kGyA). As cuttlefish, octopus and squid are not included in the definition of finfish in
FISFOOIC.ALL, they are imported as a food grade item, which is then on-sold as fish bait.
This SRA supplements an import risk analysis (IRA) completed in 2019, that reviewed trunked
elasmobranch and eviscerated teleost finfish imported for human consumption. As this IRA did
not include whole (uneviscerated) finfish or coleoid molluscs, the SRA assesses the biosecurity
risks associated with these commodities where imported for use as bait for commercial and
recreational fishing. Further, this definition is restricted to the finfish and coleoid cephalopod
mollusc species imported during 2018 (see Table 5) from estimated volumes and location of
capture data provided by the fish bait industry. All other fish bait species are ruled out of scope
of this risk analysis. Any other fish bait species not included in Table 5 should be subject to
specific risk analysis when requested by the Biosecurity New Zealand (BNZ) Animal Imports
team.
The preliminary hazard list for this SRA was largely derived from the earlier finfish IRA. It was
supplemented by a literature review to identify the preliminary hazards associated with coleoid
cephalopod molluscs, as no previous risk analysis has been completed on these species.
The combined preliminary hazard list identified 164 organisms or organism groups. Of these, 19
organisms (9 viruses, 5 bacteria and 5 myxozoan pathogens) were retained for risk assessment
after the hazard identification step. After risk assessment, 14 organisms (6 viruses, 3 bacteria and
5 myxozoan pathogens) were identified as representing non-negligible risk, as described in the
following table. Of these, only nervous necrosis virus (NNV) was associated with both finfish
and squid species. No other risk organisms were associated with coleoid cephalopods.
Group Pathogen
Virus Birnaviridae: marine aquabirnavirus (MABV)
Iridoviridae: Megalocytivirus (red sea bream iridovirus RSIV/Infectious skin and kidney necrosis virus (ISKNV))
Iridoviridae: erythrocytic necrosis virus (ENV)
Nodaviridae: nervous necrosis virus (NNV)
Rhabdoviridae: infectious haematopoietic necrosis virus (IHNV)
Rhabdoviridae: viral haemorrhagic septicaemia virus (VHSV)
Bacteria Edwardsiella spp.
Exotic Streptococcus species complex (S. agalactiae (serotype 3:283) and S. iniae)
Myxozoa Enteromyxum leei, Kudoa clupeidae, K, nova, K, iwatai, K, thyrsites
A Irradiation with 25 kGy is equivalent to a dosage of 2.5 Mrads (MPI 2011).

2 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
This SRA proposes general and specific risk management options. The general risk management
measures include:
• Frozen storage (pre-export or post-arrival storage in a transitional facility, at a temperature
range from -18 °C to -20 °C, for at least 4 months.
• Health certification from the relevant Competent Authority declaring:
o the species included in the commodity;
o the region/country of origin;
o that the commodity is sourced from wild stock;
o that the commodity has not been harvested from populations experiencing an epizootic
disease;
o has no visible signs of disease;
o labelling commodity as “For bait use only, unfit for human consumption”.
Extension of frozen storage (from the 7 days defined in FISFOOIC.ALL to 4 months)
substantially or completely denatures some bacteria (Edwardsiella spp.) and myxozoans
(Enteromyxum leei, Kudoa clupeidae, K. iwatai. K. nova and K. thyrsites). Therefore, no specific
risk management measures are considered necessary for these species.
For the remaining risk organisms listed in the table above, additional risk management measures
are considered necessary. The species-specific risk management options proposed include:
• Sourcing fish bait from species not associated with pathogens of concern.
• Sourcing fish bait from regions/countries recognised by BNZ as being free of corresponding
pathogens of concern, through the BNZ Country Approval Procedures.
• Pre-export or post-arrival batch testing of fish bait shipments to ensure absence of the
corresponding pathogens of concern, through the BNZ Country Approval Procedures.
• Pre-export or post-arrival irradiation (ionising radiation up to 50 kGyB).
The species-specific risk management options are described in the relevant chapter of this SRA
and summarised in Appendix 2. An assessment of the likely effects on pathogen occurrence in
the commodity associated with the general and specific risk management options is provided in
Appendix 3.
B Irradiation at 25 kGy denatures most bacterial, fungal and metazoan pathogens, but not all viruses (DAFF, 2012).
A dose of 50 kGy will denature all risk organisms associated with the commodity and is consistent with Australian
biosecurity guidelines (DAFF, 2013). It is acknowledged that no suitable irradiation facilities exist in New Zealand.
Further, a lower irradiation dose may be appropriate to denature the risk organisms associated with a particular fish
bait species. This may be determined on a case-by-case basis, as necessary.

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 3
Introduction
This supplementary import risk analysis (SRA) examines the biosecurity risks associated with
whole (uneviscerated) frozen marine species of finfish (class Actinopterygii) and cephalopod
molluscs (squid, cuttlefish and octopus - Subclass Coleoidea of Class Cephalopoda). These
finfish and cephalopod species were sourced from wild stocks and were imported for use as fish
bait in 2018 (Table 5). The SRA supplements a previous import risk analysis (IRA) that
examined risks associated with imported non-viable fresh, chilled and frozen eviscerated bony
fish (Osteichththyes) and trunked cartilaginous fish (Chondrichthyes). These may be sourced
from marine, fresh or brackish waters and are intended for human consumption (Blackwell,
2019). The 2019 IRA did not assess the risk to New Zealand from the importation of whole
(uneviscerated) finfish, or of cephalopod molluscs, so the relevant species have been assessed in
this SRA.
Finfish and cephalopod fish bait species, other than Octopus spp., generally occur in large
schools. Some, including herring (Clupea spp.) and sprat (Sprattus spp.) commonly occur in
mixed schools based on fish size (Maes & Ollevier, 2020). These fish bait species are caught for
human consumption and also support competing large-scale industrial fisheries for fish meal,
fish food and fish oil (Merino et al., 2014).
The knowledge on the population dynamics and epidemiology (including reservoir species,
modes of transmission, prevalence, morbidity and mortality) of the major risk organisms in
regard to lower value marine species such as fish bait, is complex and poorly understood (Munro
et al., 1983; Suttle, 2007; Dunn et al., 2012; Engelhard et al., 2014). Pathogen-associated
mortality in wild fish is generally un-noticed and rarely reported (Lafferty et al., 2015).
Translocated fish bait species represent a direct pathway for the spread of aquatic animal
diseases into the aquatic environment (Hine & MacDiarmid, 1997; Athanassopoulou & Roberts,
2004; Suttle, 2007; Herve-Claude et al., 2008; Dunn et al., 2012; Phelps et al., 2013; Pearce et
al., 2014; Oidtmann et al., 2013, 2017).
Pathogen introduction and establishment through the commodity has resulted in epizootics in
naïve wild fish host populations (including conspecifics separated by geography), causing
population level changes (Dunn et al., 2012). Examples include:
• Pilchard herpesvirus (PHV), which was introduced into the pilchard (Sardinops sagax)
wild fisheries in Australia in 1995 and New Zealand during 1996. This was reported to
have occurred through the use of infected frozen bait fish imported from California as
aquaculture feed and fish bait in Australia, and as fish bait in New Zealand (Smith et al.,
1996; Ward et al., 2001; Gaughan, 2002; Diggles, 2011). Recreational and commercial
fishers in New Zealand were observed collecting moribund and dead pilchards for later use
as fish bait in 1996 (Smith et al., 1996; Diggles, 2011). This introduction resulted in a 70-
75% decrease in spawning biomass in South Australian and New Zealand pilchard stocks
(Smith et al., 1996; Jones et al., 1997; Ward et al., 2001).
• A previous epizootic incident had occurred in Australian pilchards during 1988-89
(Whittington et al., 2008; Diggles, 2011). The lack of mortalities in New Zealand pilchards
during this period was attributed to a temporary ban on frozen pilchards imported from
Australia during the entire course of this first epizootic (Diggles, 2011).

4 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
• White spot syndrome virus (WSSV), which affected wild and farmed wild prawns
(Penaeus spp.) in Australia. This was reported to have occurred through the use of
imported infected frozen prawns as fish bait by recreational fishers (McColl et al., 2004a;
Biosecurity Queensland, 2017).
• Epizootic haematopoietic necrosis virus (EHNV), which has been spread across the inland
waterways of Northern Australia through infected frozen redfin perch (Perca fluviatilis)
used as bait (Whittington et al., 2010; Diggles, 2011).
• Spring viraemia of carp virus (SVCV), which may be spread from infected farmed or wild
cyprinid finfish to wild carp by recreational fishers (Goodwin et al., 2004; Diggles, 2011).
The inter-connectedness of the aquatic environment means that pathogen transfer between wild
and farmed stocks can also occur (Harvell et al., 1999; Gaughan, 2002; Dunn et al., 2012;
Oidtmann et al. 2013, 2017; Georgiades et al., 2016; Groner et al., 2016). Exposure may be
direct, where infected fish bait is used as fish feed. It may also be indirect, where susceptible
wild stocks are initially infected and then these stocks act as reservoirs for infection of farmed
species (Diamant et al., 2007). Examples include:
• Pilchard orthomyxovirus (POMV), a disease of wild pilchards (Sardinops sagax) that
has transferred to farmed Atlantic salmon (Salmo salar) in Tasmanian aquaculture (Mohr et
al., 2020).
• Viral haemorrhagic septicaemia virus (VHSV) (Rhabdoviridae), which first became
established in the Mediterranean Sea during 2004 following the use of infected sprats
(Sprattus sp.) and herring (Clupea harengus) as feed for marine farmed rainbow trout
(Oncorhynchus mykiss). Epizootics then occurred in other wild and farmed species, including
farmed European seabass (Dicentrarchus labrax), a previously unknown host for VHSV
(Dixon, 1999; Skall et al., 2005a, 2005b).
• VHSV was introduced into the North America Great Lakes through use of live fish as
fish bait (Diggles, 2011).
• Crayfish plague (Aphanomyces astaci), which was introduced into European waters
through frozen fish tissue containing A. astaci. This pathogen remains infective unless the fish
is frozen for at least 72 hours (Oidtmann et al. 2002; Diggles, 2011; OIE, 2019b).
If an introduced marine pathogen with a wide host range (such as VHSV) becomes endemic, its
eradication may be difficult or impossible (Myers et al., 1992; Amos et al., 1998, 2010;
Hershberger et al., 1999; Gaughan, 2002; Hedrick et al., 2003; Arkush et al., 2006).
The process of disease establishment in a new host population is not however automatic. The
presence of a pathogen in a commodity in itself does not result in disease establishment.
Successful establishment depends on many factors involving the pathogen, the host and the
environment (Stevens, 1960; Salama & Rabe, 2013).
This SRA examines the risks associated with aquatic animal diseases exotic to New Zealand.
These include both the aquatic animal diseases listed in the OIE Aquatic Code (OIE, 2019a) and
other significant aquatic animal diseases (Blackwell, 2019). These diseases could have a major
effect on the economy, people (including society and culture) and environment of New Zealand.

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 5
2.1 The importance of fish bait in New Zealand
New Zealand is geographically isolated. It has the world’s fourth largest Exclusive Economic
Zone, comprising 1.3 million square kilometres, with a 15,100 km long coastline. New Zealand’s
marine fisheries (consisting of commercial, customary and recreational sectors) play an
important economic and societal role. Of the 16,000 marine species identified in New Zealand
waters, 130 species are commercially fished (FAO, 2020a).
2.1.1 Fish bait use in New Zealand commercial fisheries
The New Zealand commercial fisheries were valued at NZ$ 4.18 billion (annually on average
during 2000 to 2015) (BERL, 2017). In this period, seafood was the fifth largest export
commodity, by value, representing (on average) 3.2% of total annual exports. In this period, the
seafood sector directly employed 4,305 full time workers, with a total employment of 13,468
people, representing 0.7% of New Zealand’s total employment (BERL, 2017). New Zealand
fishery exports in 2018 were collectively valued at NZ$ 1,817 million (Seafood New Zealand,
2019).
Fish bait is an important component of commercial and recreational fishing operations.
Commercial fishing operations using fish bait caught an estimated NZ$345 million worth of fish
in New Zealand during 2010–2015 (BERL, 2017; MPI, 2018a, 2018b, 2018c). Fish bait is used
to attract marine fish in line fishing, pot fishing and purse seining methods. Baited hooks may be
set in offshore waters to attract pelagic fish (those that inhabit the water column) or discharged in
quantity as an attractant to hold an entire school of pelagic fish in position prior to capture by
purse seine. Baited hooks may also be used in offshore and inshore waters to attract demersal
(bottom dwelling) fish to baited hooks or discharged in quantity as ground bait (FAO, 2018,
2019).
The total number of baited hooks used in New Zealand waters has declined, from 65 million in
2002-03, to 43 million in 2018-19 (Table 1).
Fish bait is also used to attract and retain fish to baited pots used by commercial and recreational
fishers in coastal inshore waters (MPI 2018a, 2018b, 2018c). During the 2017-18 fishing year,
2,083,276 commercial pot fishing sets were conducted (J. Moriarty, BNZ, pers. comm., 2020).
Table 1. Estimated total number of hooks per year set in commercial long lining, by vessel size group, for fishing years (1 Oct to 30 Sept) 2002-03, 2012-13 and 2018-19
Fishing year Number of hooks per year (thousands) Vessels < 20 m Vessels 20-30 m Vessels > 34 m Total
2002-03 27,115,490 1,887,569 36,278,908 65,281,967
2012-13 32,525,000 9,992 5.635,005 38,169,997
2018-19 19,834,717 6,011,654 18,007,719 43,854,090
Source: 2000-01 to 2012-13 from Pierre et al., (2014). 2018-19 from Fisheries New Zealand, (2020; Moriarty, J, BNZ, pers. comm., 2020).
2.1.2 Fish bait use in New Zealand recreational fishing
Recreational marine fishing activity primarily uses fish bait. The contribution of recreational
marine fishing activity to the New Zealand GDP (Gross Domestic Product) is estimated at
NZ$ 638 million annually (Southwick et al., 2018). Data from the BNZ Recreational fishing
survey (Table 2) indicates that over 1.8 million fishing trips using fish bait were carried out

6 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
during 2017-18. Recreational fishing mainly occurs in coastal inshore and shallow shelf areas.
Recreational marine fishing often occurs adjacent to marine aquaculture structures, as sea cages
act as fish aggregation devices (P. Lamb, pers. comm., 2019).
Table 2. Estimated number of recreational fishing trips using fish bait during 2017-18
Recreational fishing method
Rod/line Longline/Kontiki Pot Total
Trailer motorboat 880,018 26,501 21,165 927,684
Large Motor launch 207,711 4,103 6,389 218,203
Trailer yacht 3,565 150 164 3,879
Large yacht 13,960 525 0 14,485
Kayak/rowboat 75,634 3,220 2,681 81,535
Off land 464,505 84,402 3,468 552,375
Other 8,457 207 475 9,139
Total 1,653,850 119,108 34,342 1,807,300
Source: National Recreational Fishing Panel Survey, 2017-18 (Fisheries New Zealand 2020b).
2.1.3 Bait loss from commercial and recreational fishing
The proportion of fish baits in recreational and commercial line fishing lost to the environment
may reach 50% (Skud et al., 1978). Bait loss may result from initial mis-hooking of the bait, loss
through mechanical abrasion during setting and retrieval, loss when baits hit the water surface, or
loss when passing through the water column. Bait loss may also occur where baits are partially
consumed by fish or invertebrates including shrimps and crabs.
The automatic baiting systems used on larger commercial vessels use thousands of hooks (Table
1) and the overall number of lost baits may be considerable (Skud et al., 1978; High, 1980;
Smith, 2001; Ward & Myers, 2007). Unwanted fish bait is commonly discarded at sea by
commercial and recreational fishers (High 1980; Lokkeborg et al., 2010, 2014; Kumar et al.,
2016; Wynn-Jones et al., 2019). It is possible that fish bait may be discarded by recreational
fishers adjacent to marine sea cages.
2.1.4 Finfish bait imports, 2018
Prior to 2008, New Zealand was essentially self-sufficient in fish bait. During 2008 – 2017, a
variety of fish and coleoid mollusc species have been imported (Appendix 1).
Available import data for frozen fish bait do not provide the location of capture. Industry-
sourced data on species, volume of trade and location of capture of fish bait imported in 2018
(Table 3) indicate this mainly comprised of species of families Clupeidae (58%), Scombridae
(20%) and Mugilidae (13%). In contrast to previous years (see Appendix 1), mackerel species
(Carangidae) were not imported in 2018. A minor amount (< 1 t) of flying fish (family
Exocoetidae) were imported as a trial in 2018 (B. Burney, pers. comm., 2019). This is considered
insignificant and flying fish are not considered further.
2.1.5 Coleoid cephalopod bait imports, 2018
Molluscs are not currently included in the definition of marine fish imported as frozen fish bait
(MPI, 2011). Import commodity data do not separate frozen coleoid molluscs used as fish bait
from imports for human consumption. Industry-sourced data on species, volume of trade and

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 7
location of capture of coleoid molluscs in 2018 (Table 4) indicate most (99%) cephalopod fish
bait consisted of squid (family Ommastrephidae).
2.2 The importance of aquaculture
The aquaculture sector employed 3000 people in 2015, with a contribution of $NZ 500 million to
the New Zealand economy. Of this total, $NZ 338.1 million was generated in exports
(Aquaculture, 2020). The New Zealand Government Aquaculture Strategy projects an increase
of this sector to reach $NZ 3 billion in annual sales by 2035 (Aquaculture, 2020). New Zealand
aquaculture is dependent upon access to clean marine and fresh waters (Lafferty et al., 2015;
Georgiades et al., 2016; Haenen, 2017).
Salmonid aquaculture in New Zealand is mainly focussed on Chinook (king) salmon
(Oncorhynchus tshawytscha), where hatchery-raised smolt are on-grown in fresh-water
aquaculture, or in sea-cages. Chinook salmon aquaculture was valued at $NZ 77 million in 2018
(Aquaculture New Zealand, 2019). Other developing marine finfish aquaculture species farmed
for food include yellowtail kingfish (Seriola lalandi), snapper (Sparus aurata), turbot (Colistium
nudipinnis) and grouper/bass (Polyprion oxygeneios) (NIWA, 2017a; Plant & Food, 2016).
While salmonid aquaculture uses purpose-designed pellet fish food, the aquaculture of non-
salmonid species may utilise fish bait species as aquaculture feed, as practiced in the Australian
aquaculture for Southern bluefin tuna (Thunnus maccoyii) (Ellis, 2016).
In fresh waters grass carp (Ctenopharyngodon idella) and silver carp (Hypophthalmichthys
molitrix) are also farmed for weed control purposes (Clayton & Wells, 1999; Blackwell, 2019).

8 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Table 3. Estimated volume (t) and source location of finfish imported as fish bait, 2018
Finfish family Common name Scientific name Volume 1 (t)
Volume 1
(t) by family
Reported location2 of capture
Clupeidae Herring, pilchard, sardine, sprat 600 Spotted sardinella Amblygaster (=Sardinella)
sirm 390 Malaysia
Round sardinella Sardinella aurita 100 China South American
pilchard Sardinops sagax 100 China, Japan,
South Africa Indian oil sardine Sardinella longiceps 10 Indonesia
Scombridae Tuna, mackerel 208 Blue mackerel Scomber australasicus 90 Fiji Skipjack tuna Katsuwonas pelamis 55 China, Indonesia Little tunny Euthynnus alletteratus 28 China, Indonesia Atlantic mackerel Scomber scombrus 25 Spain Chub mackerel Scomber japonicus 10 China
Mugilidae Mullet Flathead grey mullet Mugil cephalus 140 140 Australia,
Indonesia
Scomberesocidae Saury Pacific saury (Samna) Cololabis saira 57 57 China, Chinese
Taipei
Engraulidae Anchovy Californian anchovy Engraulis mordax 33 33 United States
Hemiramphidae Halfbeak Ballyhoo halfbeak Hemiramphus (Esox)
brasiliensis 4 4 Indonesia
Exocoetidae Flying fish a Japanese flying fish Cheilopogon agoo <1 Japan Tropical two-wing
flying fish Exocoetus volitans <1 Japan
Total 1042 Source New Zealand fish bait industry: B. Burney, pers. comm., 2019; R. Clark, pers. comm., 2019; D. Rutherford, pers. comm., 2019; Spencer, pers. comm., 2019; C. Williams, pers. comm., 2019.
Notes 1. Estimated total weight provided by fish bait industry. 2. Reported location of capture provided by fish bait industry.

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 9
Table 4. Estimated volume (t) of coleoid cephalopod molluscs imported as fish bait, 2018
Family Species (current scientific name)
Common name Previous scientific name
Volume 1
(t) Volume1
(t) by family
Location of capture2
Ommastrephidae
Subfamily Illicinae
Illex argentinus Argentine shortfin squid
Ommastrephes argentinus
276 Argentina, China
Illex coindetii Broadtail shortfin squid
Illex illicibrosus 80 United States
Subfamily Todarodinae
Todarodes pacificus Japanese flying squid
Ommastrephes pacificus
350 706 China
Loliginidae Doryteuthis opalescens
Opalescent inshore squid
Loligo opalescens
80 United States
Loligo spp. Common squid Loligo spp. 1 81 Chinese Taipei, United States
Uroteuthis duvaucelli Indian squid Loligo duvaucelii 03 Malaysia3
Sepiidae Sepia recurvirostra Curvespine cuttlefish
Sepia recurvirostra
2 Malaysia
Sepia spp. Cuttlefish Sepia spp. 1 3 Malaysia
Octopodidae Octopus spp. Octopus Octopus spp. 1 1 China, Japan
Source New Zealand fish bait industry: B. Burney, pers. comm. 2019; R. Clark, pers. comm., 2019.
Notes 1. Estimated weight (t) provided by fish bait industry
2. Location of capture as provided by fish bait industry
3. Uroteuthis duvaucelii pre-ordered in 2018 to be caught during 2018-2019.
Scope, commodity definition and assumptions
3.1 Scope
This SRA assesses the biosecurity risks associated with imported frozen, whole, wild-caught
marine fish (Class Actinopterygii) and coleoid cephalopod molluscs (Class Cephalopoda,
subclass Coleoidea), ‘imported for use as fish bait’ under the Biosecurity Act 1993C.
This risk analysis includes whole coleoid cephalopod molluscs, which are imported for human
consumption and redirected into the fish bait pathway. However, other imported aquatic animal
products may become unfit for human consumption due to post-border product spoilage
(including freezer burn, incorrect frozen storage, or contamination). Most rejected product is
discarded, or processed into fish meal, but some may be re-directed into the fish bait pathway
(Blackwell, 2019). As this occurs in an ad-hoc manner, the volume of transferred product is
unknown, but likely to be small. While this latter pathway is not considered further, the findings
in this SRA should be considered in any future review of aquatic animal products for human
consumption.
Processed “consumer-ready aquatic animal products” imported for human consumption (MPI,
2001, 2008), fish bait products derived from imported ornamental fish, live or preserved fish or
molluscs for display, zoos or for biological research purpose (Hine & Diggles, 2005), or fish
The Biosecurity Act 1993 and Fisheries Act 1996 are available online from the Parliamentary Counsel Office of the New Zealand Government at: http://www.legislation.govt.nz/act.

10 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
food derived from any other plant or animal sources (Cobb, 2008) are outside the scope of this
analysis.
Fish bait caught in fresh, brackish or estuarine waters is also outside the scope of this risk
analysis. Euryhaline species such as flathead grey mullet (Mugil cephalus) are considered in-
scope if caught in the marine environment, but not if caught in fresh or brackish waters.
Fish bait may also be imported as a “user-ready shelf-stable” product that has been chemically or
heat-processed (MPI, 2011). This commodity is outside the scope of this risk analysis.
3.2 Commodity definition
For the purposes of this risk analysis, the commodity definition is restricted to non-viable finfish
of Class Actinopterygii and coleoid cephalopod molluscs of Class Cephalopoda: Subclass
Coleoidea (squid, cuttlefish and octopus) currently imported into New Zealand for use as fish
bait.
The commodity definition for finfish (Class Actinopterygii) used in this SRA is consistent with
the definition used in the IRA for eviscerated or trunked fish for human consumption (Blackwell,
2019).
Coleoid cephalopod molluscs were not included in the IRA for eviscerated or trunked fish for
human consumption (Blackwell, 2019) and no previous risk analysis has been completed for this
commodity. For the purposes of this risk analysis, the definition of coleoid cephalopod molluscs
is consistent with the Fisheries Act, 1996 as:
“All species of the phylum Echinodermata and phylum Mollusca and all species of the
class Crustacea at any stage of their life history, whether living or dead.”
Not all molluscs are used as fish bait (B. Burney, pers. comm., 2019). For the purposes of this
risk analysis, molluscan fish bait is restricted to members of sub-class Coleoidea (squid,
cuttlefish and octopus). This excludes the shelled cephalopods of genera Allonautilus and
Nautilus (Subclass Nautiloidea) (WoRMS, 2019) as these are not used as bait.
Cephalopod molluscs are defined (Jereb et al., 2010) as:
“The class within the Phylum Mollusca, characterized by bilateral symmetry, internal
‘shell’ or absence of shell (except nautiluses), anterior head, appendages and funnel,
posterior mantle, mantle cavity with organs, and shell and fins when present.”
Sub-class Coleoidea is defined (MolluscaBase, 2019; WoRMS, 2019) as:
“molluscs within the Class Cephalopoda including squid, (families Loliginidae and
Ommastrephidae), cuttlefish (family Sepiidae) and octopus (family Octopodidae) but
excluding nautiloid cephalopods of Subclass Nautiloidea.”
Fish bait is a non-human food item currently imported as a whole (uneviscerated) bulk
commodity, under Section 7.2.4 of the Import Health Standard (IHS) for fish bait and fish food

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 11
(FISFOOIC.ALL) (MPI, 2011). Under this IHS, fish bait is restricted to marine finfish (Class
Actinopterygii) which is:
• Labelled with species and country of capture
• Non-viable and either:
o Frozen (to below -18 °C for at least 18 hours); or,
o Irradiated with 2.5 MradsD.
The IHS (FISFOOIC.ALL) is restricted to marine finfish. For the purposes of this risk analysis,
the definition of fish bait is extended to include coleoid molluscs (Class Cephalopoda, subclass
Coleoidea). Further, this definition is restricted to the finfish and coleoid cephalopod mollusc
species listed in Table 5. This has been determined from import data from 2018 and information
provided by the fish bait industry (B. Burney, pers. comm., 2019; R. Clark, pers. comm., 2019; P.
Lamb, pers. comm., 2019; M. Lyford, pers. comm., 2020; C. Williams, pers. comm., 2019).
All other fish bait species are ruled out of scope of this risk analysis. Any other fish bait species
not included in Table 5 would be subject to specific risk analysis when requested by the
Biosecurity New Zealand Animal Imports team.
This commodity includes fish bait that is sorted to species and frozen (at sea or on shore) into
blocks of varying size (typically around 20 kg). The blocks are each placed into cardboard
cartons and these are stored in frozen containers, ready for export (B. Burney, pers. comm.,
2020). Imported fish may be used in the whole form as fish bait or be further processed into
berley.
Frozen fish bait is transported in refrigerated containers. The standard refrigerated containers used
in international trade can be maintained within a temperature range from -20 °C to + 10 °C (Sea
Containers, 2020). In their frozen storage mode these containers are maintained within a
temperature range from -18 °C, to -20 °C (B. Burney, pers. comm., 2019; T. Leighton, BidFood
Ltd., Wellington, pers. comm., 2020).
D Irradiation with 2.5 Mrads is equivalent to a dosage of 25 kGy. It is suggested this be increased to 50 kGy, consistent with Australian Government biosecurity guidelines (DAFF, 2013).

12 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Table 5. Finfish and coleoid cephalopod mollusc families and species considered as fish bait
Category Family Species
Finfish Carangidae Trachurus japonicus, T. murphi, T. symmetricus, T. trachurus
Clupeidae Ambylygaster sirm, Clupea harengus, Clupea pallasii pallasii, Sardinella aurita, Sardinella lemuru, Sardinella longiceps, Sardinops sagax
Engraulidae Engraulis mordax, E. ringens
Hemiramphidae Hemiramphus balao, H. brasiliensis, H. dussumieri, H. lutkei
Mugilidae Chelon auratus, C. labrosus1, C. ramada, Mugil cephalus
Scomberosocidae Cololabias saira
Scombridae Auxis rochei, A. thazard, Euthynnus alletteratus, Katsuwonas pelamis, Sarda chilensis, S. lineolata, S. sarda, Scomber australasicus, S. japonicus, S. scombrus, Thunnus obesus
Cephalopods Loliginidae Doryteuthis opalescens, Loligo spp., Uroteuthis duvaucelli
Ommastrephidae Illex argentinus, I. coindetti, Todarodes pacificus
Sepiidae Sepia recurvirostrata, Sepia spp.
Octopodidae Octopus spp.
Notes
1 The thicklip grey mullet Chelon (Mugil) labrosus is included here because of potential confusion with the flathead grey mullet (Mugil cephalus). Source
New Zealand fish bait importers, Animal Imports team, Biosecurity New Zealand
3.3 Assumptions
For the purposes of this risk analysis, the following general assumptions are made:
• Susceptibility determined experimentally (where the infection process closely follows a
likely route of natural infection e.g. per os or cohabitation) represents a potential exposure
pathway.
• Pathogen epidemiology observed in aquaria or experiment follows similar processes to
disease in wild stocks of fish and coleoid molluscs.
• Organisms that have never been reported in the scientific literature or technical databases
(such as the New Zealand Organism Register (NZOR, 2017)) are assumed to be absent from
New Zealand (exotic), following the principles of Chapter 1.4 of the OIE Aquatic Code
(OIE, 2019a).
• Frozen storage (to at least -18 °C for at least 168 hours (7 days)) is assumed to denature
fungi, protozoa, nematodes, monogenean, digenean and cestode helminths, as well as most
metazoan parasites present in the commodity (Blackwell, 2019; USDA, 2019). These
pathogens are not considered further.
• Viral, bacterial and myxozoan pathogens may remain viable in fish bait frozen (to -18 °C
or -20 °C for 168 hours (7 days)) (Hine & MacDiarmid, 1997; Johnston, 2008).
• Host range may be wider than reported in the literature, particularly for low value species
such as fish bait (Blackwell, 2019; OIE, 2019a). As the scope of the commodity is confined
to listed fish bait species (Table 5), the rule that applied in Blackwell (2019) has been
modified, following the principles of Chapter 1.5 of the OIE Aquatic Code (OIE 2019a), as
follows:
o If only one species in a genus is susceptible to an identified risk organism, then only
that species is excluded in the species declaration option
o If two or more species in a genus are susceptible to an identified risk organism, then all
listed fish bait species in that genus are excluded in the species declaration option

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 13
o If two or more genera in a family are susceptible to an identified risk organism, then all
listed fish bait species in the family (or sub-family) are excluded in the species
declaration option.
Risk analysis methodology
4.1 General procedures
The methodology used in this risk analysis is guided by the Biosecurity New Zealand Risk
Analysis Procedures – Version 1 (Biosecurity New Zealand, 2006), the Handbook on Import
Risk Analysis for animals and animal products (OIE, 2010) and Chapter 2 of the Aquatic Animal
Health Code (OIE, 2019a). The risk analysis process comprises several steps: hazard
identification, risk assessment and risk management (Figure 1). It also includes risk
communication.
4.1.1 Hazard identification
A list of organisms of concern associated with the commodity (the preliminary hazard list) is
compiled from the OIE list of aquatic animal diseases (OIE, 2019a), published BNZ risk
analyses and from the relevant published scientific literature. For each organism on this list,
several steps are completed: formal identification (taxonomic classification), OIE status, New
Zealand status, together with a determination of whether or not the organism meets key
biological and epidemiological characteristics to be considered as a hazard.
Hazard identification concludes with an assessment of whether or not the organism is identified
as a hazard in the commodity. The results of the hazard identification are commonly summarised
as a table. All organisms deemed to be hazards are subjected to further risk assessment.
4.1.2 Risk assessment
Risk assessment (Figure 1) consists of four steps applied to each hazard: entry assessment,
exposure (and establishment) assessment, consequence assessment and risk estimation. At each
of the first two steps a qualitative assessment is made of the likelihood (of entry, exposure and
establishment, respectively), based on the available epidemiological information. The
consequence assessment is undertaken to determine the likely direct and indirect impacts of
entry, exposure and establishment of a risk organism in New Zealand. This includes effects on
people (including societal and cultural), the New Zealand environment and the New Zealand
economy (Biosecurity Act 1993, Section 23).

14 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Figure 1. The general risk analysis process
Organisms of concern
HAZARD IDENTIFICATION
Can the organism remain viable inthe commodity ?
Does transmission require a vector?
Is the vector present
in New Zealand?
Is the pathogen present in New Zealand
Are there different
strains overseas?
Not identified as a hazard in this
risk analysis
Identified as ahazard in thisrisk analysis
RISK ASSESSMENT
Entry AssessmentLikelihood of hazard entering New Zealand
on the pathway
Exposure AssessmentLikelihood of exposure and establishment in
New Zealand
Consequence AssessmentLikely impacts on economy,
environment andhuman health in New Zealand
Risk Estimation
Organism is assessed
to be a risk
Risk Estimation
Not assessed to
be a risk
RISK MANAGEMENT
What options are available to managethe risk ?
What is the effect of each measure on the
level of risk?
yes
yes
yes
yes
no
no
no
no
no
no
yes
negligible
negligible
negligible
-
non negligible
-non negligible
non negligible
Is the organism present in theCommodity ?
yes
RISK COMMUNICATION
In addition to using the terms negligible and non-negligible to describe risks, the entry, exposure
and consequence assessments also use qualitative descriptors (ranging from very low to very
high) to describe the comparative levels of likelihood (in the case of entry and exposure
assessments) or impact (in the case of consequence assessment) (Biosecurity New Zealand,
2006).
The Biosecurity New Zealand Risk Analysis Procedures (Biosecurity New Zealand, 2006)
provides a more detailed explanation of the terminology as follows:
Risk attributes Description
Negligible Not worth considering, insignificant
Non-negligible Worth considering, significant
Risk descriptors
Very low Close to insignificant
Low Less than average, coming below the normal level
Medium Around the normal or average level
High Extending above the normal or average level
Very high Well above the normal or average level
These qualitative levels of likelihood of entry, exposure and establishment and the levels of
impact (economic, social, and environmental) were combined to assess whether the associated
risk is negligible or non-negligible.

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 15
If, either the likelihood of entry or the likelihood of exposure of a pathogen is assessed as
negligible, then the overall risk is deemed negligible and the remaining steps, being redundant, are
not undertaken.
If the expected likelihood of entry or exposure are assessed to be non-negligible, but the impacts of
establishment are assessed as negligible, then the overall risk is also negligible, and no further
analysis is necessary.
Those pathogens considered to pose a non-negligible risk are deemed to require risk
management. The relative magnitude of the risk (very low to very high) assists in determining
the stringency of risk management measures that would be needed to reduce the risk to an
acceptable level.
4.1.3 Risk management
Risk management identifies the options available for managing non-negligible risks, based on
the epidemiology of the risk organism. Where the OIE Code (OIE, 2019a) lists recommendations
for the management of a risk, these are described alongside additional risk management options,
where available, from the scientific literature.
In addition to the options presented, prohibition may also be considered. Recommendations for
the appropriate sanitary measures to achieve the effective management of risks are not made in
this document. These will be determined when an Import Health Standard (IHS) is drafted.
As obliged under Article 3.1 of the World Trade Organisation (WTO) SPS Agreement (WTO,
2017), the measures adopted in IHSs will be based on international standards, guidelines and
recommendations where they exist, except as otherwise provided for under Article 3.3 of the
SPS Agreement. That is, measures providing a higher level of protection than international
standards can be applied if there is scientific justification, or if there is a level of protection that
the member country considers is more appropriate. These additional measures must be based on
a scientific risk analysis.
4.1.4 Risk communication
After a draft import risk analysis has been written, BNZ analyses the options available and
proposes draft measures for the effective management of the identified risks. These are then
presented in a draft IHS that is released for public comment, together with a risk management
proposal (RMP) that summarises the options analysis, the rationale for the proposed measures
and provides a link to the draft risk analysis.
Not every risk organism identified in the risk analysis may be associated with a particular
imported fish bait species and require risk management in an IHS. The RMP will take into
account specific information that would affect the need for risk management measures. For
instance, factors considered in an RMP would include (but not be limited to) the country of
origin of the commodity, the presence or absence of risk organisms in that country, the species
from which the commodity is derived, and any manufacturing processes that inactivate risk
organisms.
The document package (draft IHS, RMP and risk analysis) is then released for stakeholder
consultation. Stakeholder submissions in relation to these documents are reviewed and
published, including any supplementary risk analyses that may be required, before a final IHS is
issued.

16 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Hazard identification
Previous risk analyses have been completed for fish bait and fish food (Cobb, 2009) and for
eviscerated/trunked fish for human consumption (Blackwell, 2019). No previous risk assessment
has been completed for coleoid cephalopod molluscs (squid, cuttlefish, and octopus) imported
for any end use. The preliminary hazard list was compiled using Cobb (2009) and Blackwell
(2019), the OIE list of aquatic animal diseases (OIE, 2019a), the peer-reviewed scientific
literature, health databases, previous BNZ risk assessments and information provided by experts
and interested parties. Each organism was assessed in terms of eight key criteria:
• OIE status
• New Zealand status
• Viability in fish bait (frozen to -20 °C for 168 hours (7 days))
• Cause significant disease
• Presence of more virulent exotic strains
• Necessity for a vector or intermediate host in the life cycle
• Zoonotic potential
• Potential hosts in New Zealand.
Not all the organisms identified as hazards in the previous risk analyses are relevant to the
commodities considered here. Some organisms were not considered as hazards, Blackwell
(2019) because they were not present in eviscerated or trunked fish. These, however, may be
present in whole frozen fish or coleoid cephalopods.
Only the pathogens reported from the fish and coleoid cephalopod host species listed in Table 5
are considered for this analysis. To establish in New Zealand, a pathogen must be present and
viable in the commodity.
The findings of the Hazard Identification for pathogens derived from finfish and coleoid
cephalopod molluscs are presented in Table 6. All organisms identified as hazards were
subjected to risk assessment.

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 17
Table 6. Hazard Identification table for specified1 wild marine finfish and coleoid molluscs used as fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Viruses
Astroviridae
Eastern sea garfish astrovirus
Hemiramphidae Eastern sea garfish (Hyporhamphus australis)
Southwest Pacific, Australia
N N Y N N/A N N Y N Geoghegan et al., 2018
Birnaviridae
Marine aquabirnavirus (MABV)
Infectious pancreatic necrosis virus-like (IPNV-like strains) including viral deformity virus (VDV) and yellowtail ascites virus (YAV)
Carangidae Japanese amberjack (Seriola quinqueradiata), Japanese jack mackerel (Trachurus japonicus), Mediterranean horse mackerel (Trachurus mediterraneus). yellowtail amberjack (Seriola lalandi)
Widespread Western Atlantic (Gulf of Cadiz), Mediterranean Sea, Japan, North-central Pacific, Australia
N Y Y Y Y N N Y Y Sorimachi & Hara, 1985; Castric, 1997,;Hanlon & Forsythe, 1990; Nakajima et al.,1993; Diggles, 2004a, 2004b; Isshiki et al., 2004; Tisdall & Phipps, 1987; Davies et al., 2010;; Munro & Midtlyng, 2011;; Ogut & Altuntas, 2014,; Diggles, 2016
MABV (cont.) Clupeidae Atlantic herring (Clupea harengus), sardine (Sardinops spp.), Cllupeidae (wide host range)
West Atlantic (Gulf of Cadiz)
N N Y Y N/A N N Y Y Nakajima et al., 1998; Crane et al., 2000; Crane & Williams, 2008; Wallace et al., 2008; McColl et al., 2009; Crane & Hyatt, 2011; Diggles, 2011 2016; Fish Base, 2019
MABV (cont.) Engraulidae European anchovy (Engraulis encrasiciolus)
West Atlantic (Gulf of Cadiz), Mediterranean Sea
N N Y Y N/A N N Y Y Wallace et al., 2008; Ogut & Altuntas, 2014
MABV (cont.) Scombridae Atlantic mackerel (Scomber scombrus)
West Atlantic (Gulf of Cadiz)
N N Y Y N/A N N Y Y Wallace et al., 2008; Moreno et al., 2014
Bunyaviridae
Eastern sea garfish bunya-like virus
Hemiramphidae Eastern sea garfish (Hyporhamphus australis)
Southwest Pacific, Australia
N N Y N N/A N N Y N Geoghegan et al., 2018
Hepnaviridae
Eastern sea garfish hepatitis B virus
Hemiramphidae Eastern sea garfish (Hyporhamphus australis)
Southwest Pacific, Australia
N N Y N N/A N N Y N Geoghegan et al., 2018
Herpesviridae
Malacoherpesvirus (unclassified)
Octopodidae Octopus vulgaris United States N N Y N N/A N N Y N Prado-Alvarez & Garcia-Fernandez, 2019
Pilchard herpesvirus Clupeidae South American pilchard (Sardinops sagax) (= S. neopilchardus)
Southwest Pacific (Australia, New Zealand)
N Y Y Y N N N Y N Crockford et al. 2005; Hanson et al., 2011

18 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Iridoviridae
Subfamily Megalocytivirus
Red sea bream iridovirus (RSIV), infectious spleen and kidney necrosis virus ISKNV, heart and skeletal muscle inflammation (HSMI) and associated viruses
Carangidae Japanese jack mackerel (Trachurus japonicus), bigeye trevally (Caranx sexfasciens), Japanese scad (Decapterus maruadsi), shrimp scad (Alepes djedaba (= Carynx calla)), yellowstripe scad (Selaroides leptolepis), barred queenfish (Scomberoides tala (= Chorinemus hainanensis)). double spotted queenfish (Scomberoides lysan (= Chorinemus moadetta)
Northwest Pacific (China, Korea, Japan, southeast Asia)
Y N Y Y N/A N N Y Y Wang et al., 2007; Anon., 2017a; OIE, 2019a; Rimmer et al., 2015, 2017; CIFA, 2019
RSIV (cont.) Clupeidae Dotted gizzard shad (Konosirus (Clupanodon) punctatus), Pacific herring (Clupea pallasii pallasii)
Northwest Pacific, (China, Korea, Japan, southeast Asia)
Y N Y Y N/A N N Y Y Wang et al., 2007
RSIV (cont.) Mugilidae Flathead mullet (Mugil cephalus), longarm mullet ((Osteomugil cunnesius (= Mugil (Osteomugil) ophuyseni))
Widespread Indo-Pacific
Y N Y Y N/A N N Y Y Gibson-Kueh et al., 2004, Wang et al., 2007; OIE, 2019a; Rimmer et al., 2015, 2017
RSIV (cont.) Scombridae Chub mackerel (Scomber japonicus), greater amberjack (Seriola dummerili), yellowtail amberjack (Seriola lalandi), Japanese amberjack (S. quinqueradiata), Japanese Spanish mackerel (Scomberomorus niphonius), northern bluefin tuna (Thunnus thynnus)
Widespread Indo-Pacific, anti-tropical
Y N Y Y N/A N N Y Y Wang et al., 2007; Rimmer et al., 2015, 2017; Anon., 2017a; OIE, 2019a; CIFA., 2019
Viral erythrocytic necrosis virus (VEN or ENV) and associated viruses (Iridoviridae)
Clupeidae Atlantic herring (Clupea harengus), Pacific herring (C. pallasii pallasii)
Atlantic and Pacific Salish Sea, Alaska
N N Y Y N/A N N Y Y Reno et al., 1985; Emmenegger et al., 2014
VEN (Cont.). Engraulidae Californian anchovy (Engraulis mordax)
Northeast Pacific N N Y Y N/A N N Y Y Pagowski et al., 2019

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 19
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Subfamily Lymphocystivirus
Lymphocystis disease iridovirus (LCDV)
Clupeidae Indian oil sardine (Sardinella longiceps)
Widespread N Y Y N N/A N N Y N Matsusato, 1975; Leong & Colorni, 2002; Borrego et al., 2017; Vijapoopathy et al., 2016
Lymphocystis iridovirus (cont,)
Mugilidae Flathead grey mullet (Mugil cephalus)
Widespread N Y Y N N/A N N Y N Alexandrawicz,1951; Ovcharenko, 2016
Iridoviridae (unclassified)
Octopodidae, Sepiidae
Octopus vulgaris Western Atlantic N N Y N (not in wild stocks)
N/A N N Y N Rungger et al., 1971; Farley, 1978
Nodaviridae: Betanodavivus
Korean shellfish nervous necrosis virus (KSNNV) (Nodaviridae)
Sepiidae Sepia spp. Korea, China N N Y N N/A N N Y N Kim et al., 2018; Bandin & Souto, 2020
Viral encephalopathy and retinopathy (VER) and nervous necrosis virus (VNN) (NNV) Striped Jack nervous necrosis virus (SJNNV)
Carangidae Atlantic horse mackerel (Trachurus trachurus), greater amberjack (Seriola dummerili), Japanese jack mackerel (T. japonicus), permit (yellow-wax pompano) (Trachinotus falcatus), snub-nose pompano (Trachinotus blochii), white trevally (striped jack) (Pseudocaranx dentex), yellowtail amberjack (S. quinqueradiata)
Widespread N N Y Y N/A N N Y Y Mori et al., 1991, 1992; Danayadol et al., 1995; Nguyen et al., 1996; Tubbs et al., 2007; Gomez, 2010; Crane & Hyatt, 2011; Forrest et al., 2011; IDAAD, 2019; OIE 2019a
NNV (cont.) Clupeidae European pilchard (Sardina pilchardus)
Widespread N N Y Y N/A N N Y Y Panzarin et al., 2012
NNV (cont.) Engraulidae Japanese anchovy (Engraulis japonicus)
Northwest Pacific (Japan)
N N Y Y N/A Y N Y Y Gomez et al., 2006
NNV (cont.) Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon (Mugil) auratus) (= Liza aurata), leaping mullet (Chelon (=Liza, Mugil) saliens), thicklip grey mullet (Chelon (=Mugil) labrosus)
Mediterranean, Caspian Sea, Indo-Pacific (Israel, Iran)
N N Y Y N/A N N Y Y Ucko et al., 2004; Panzarin et al., 2012; Zorriehzahra et al., 2005, 2016
NNV (cont.) Octopodidae Octopus vulgaris Italy N N Y Y N/A N N Y Y Fichi et al., 2015; Bandin & Souto, 2020
NNV (cont.) Scombridae Chub mackerel (Scomber japonicus)
Korea N N Y Y N/A N N Y Y Gomez et al., 2008
NNV (cont.) Todaronidae Todarodes pacificus Widespread N N Y Y N/A N N Y Y Ford et al., 1986; Gomez, 2010; Fiorito et al., 2015; OIE, 2019b

20 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Betanodavirus (unclassified strains 1 and 2)
Octopodidae Octopus vulgaris Europe N N Y N N/A N N N N Fichi et al., 2015; Fiorito et al., 2015; Prado-Alvarez & Garcia-Fernandez, 2019
Orthomyxoviridae
Isavirus
Infectious salmon anaemia virus (ISAV) (HPR0 ISAV, HPR-deleted ISAV)
Clupeidae Atlantic herring (Clupea harengus)
Widespread Y N Y Y N/A N N Y Y Nylund et al., 2002; Hine & Diggles, 2005; Tubbs et al., 2007; Diggles, 2011; OIE, 2019a
Orthomyxovirus
Pilchard orthomyxovirus (POMV)
Clupeidae South American pilchard (Sardinops sagax) (=S. neopilchardus)
Southwest Pacific (Australia, New Zealand)
N Y Y N N N N Y N SCAAH, 2015; Diggles, 2016; PROMED, 2017; NIWA, 2018; Blackwell, 2019
Picornaviridae
Eastern sea garfish picornavirus
Hemiramphidae Eastern sea garfish (Hyporhamphus australis)
Southwest Pacific (Australia)
N N Y N N/A N N Y N Geoghegan et al., 2018
Reoviridae: Aquareovirus
Piscine aquareovirus (PRV) and associated viruses
Carangidae Atlantic horse mackerel (Trachurus trachurus)
Widespread N N Y Y N/A N N Y Y Fauquet et al., 2005; Cobb, 2008; King et al., 2011; Garseth et al., 2012; Roberts, 2012; Wiik-Nielsen et al., 2012; Carlile et al., 2014
PRV (cont.) Clupeidae Atlantic herring (Clupea harengus)
Northeast Atlantic N N Y Y N/A N N Y Y Wiik-Nielsen et al., 2012
Reoviridae (unclassified)
Sepiidae Sepia officianalis Europe N N Y N N N N Y N Devauchelle & Vago, 1971
Rhabdoviridae
Eastern sea garfish rhabdovirus
Hemiramphidae Eastern sea garfish (Hyporhamphus australis)
Southwest Pacific (Australia)
N N Y N N/A N N Y N Geoghegan et al., 2018
Infectious haematopoietic necrosis virus (IHNV)
Clupeidae Pacific herring (Clupea pallasii pallasii)
East, West Pacific, Northeast Atlantic
Y N Y Y N/A N N Y Y Hart et al., 2011; Dixon et al., 2016; OIE, 2019a
Viral haemorrhagic septicaemia virus (VHSV) (Rhabdoviridae)
Carangidae Mediterranean horse mackerel (Trachurus mediterraneus), Japanese amberjack (Seriola quinqueradiata)
Black Sea, Europe, Northwest Pacific (Japan, S. Korea)
Y N Y Y N/A N N Y Y Ito et al., 2004; Kim et al., 2013a; OIE, 2019a

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 21
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
VHSV (cont.). Clupeidae Atlantic herring (Clupea harengus)
Black Sea, Europe, Widespread, East and West Atlantic
Y N Y Y N/A N N Y Y Meyers et al., 1994; Traxler & Kieser, 1994; Kocan et al., 1997; Dixon et al., 1997; Marty et al., 1998; Mortensen et al., 1999; Traxler et al., 1999; Kocan et al., 2001; Biosecurity Australia, 2002; Hedrick et al., 2003; Goodwin et al., 2004; OIE, 2019a
VHSV (cont.). Clupeidae Pacific herring (Clupea pallasii pallasii)
Pacific Ocean Y N Y Y N/A N N Y Y Meyers et al., 1994; Hershberger et al., 1999; Meyers & Winton, 1995; Marty et al.,1998; Biosecurity Australia, 2002; Purcell et al., 2012; OIE, 2019a
VHSV (cont.). Clupeidae European sprat (Sprattus sprattus)
Northeast Pacific (Alaska, Canada)
Y N Y Y N/A N N Y Y Skall et al., 2005a, 2005b; Crane & Hyatt, 2011; Diggles, 2011; Ogut & Altunas, 2014; Sandlund et al., 2014; CIFA, 2019
VHSV (cont.). Clupeidae European pilchard (Sardinia pilchardus)
East Atlantic Y N Y Y N/A N N Y Y Herve-Claude et al., 2008
VHSV (cont.) Clupeidae South American pilchard (Sardinops sagax)
Pacific Y N Y Y N/A N N Y Y Biosecurity Australia, 2002; OIE, 2019a
VHSV (cont.) Engraulidae European anchovy (Engraulis encrasicolus)
Black Sea, Northeast Atlantic (Europe) Widespread
Y N Y Y N/A N N Y Y OIE, 2019a
VHSV (cont.) Mugilidae Flathead grey mullet (Mugil cephalus)
Northwest Pacific (S. Korea), Northeast Pacific (California)
Y N Y Y N/A N N Y Y Biosecurity Australia, 2002; Lee et al., 2007; Kim & Faisal, 2010; Kim et al., 2013a; OIE, 2019a
VHSV (cont.). Scombridae Chub mackerel (Scomber japonicus)
Northeast Pacific (California)
Y N Y Y N/A N N Y Y Hedrick et al., 2003; Kim et al., 2013a; Anon., 2017a; OIE, 2019a
Unassigned virus Octopodidae Macroctopus maorum, Octopus vulgaris
Widespread, New Zealand
N Y Y Y N N N Y N Prado-Alvarez & Garcia-Fernandez, 2019
Unassigned virus (possibly Ostreid herpesvirus (OsHV-1)
Loliginae Loligo pealei Northwest Atlantic, Northeast Pacific
N Y Y N N N N N N Hanlon & Forsyth, 1990; Prado-Alvarez & Garcia-Fernandez, 2019
Bacterial pathogens
Alphaproteobacteriaceae
Rickettsia-like organisms (RLO)
Octopodidae Octopus vulgaris Northwest Atlantic, Mediterranean
N Y Y Y N Y N Y N Gestal et al., 1988; Fiorito et al., 2015; Diggles, 2011

22 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Bacillaceae
Bacillus spp. Octopodidae Octopus vulgaris Mediterranean Sea, West Atlantic
N Y Y N N N Y Y N Ford et al., 1986; Farto et al., 2014; NZ Fungi, 2019
Betaproteobacteriaceae
Acinetobacter anitratus (= A. baumanii)
Octopodidae Enteroctopus dofleini Europe N Y N Y N N Y Y N Stoskopf et al., 1987; Fiorito et al., 2015; Anon., 2017b
Achromobacter sp. Mugilidae Flathead grey mullet (Mugil cephalus), Longarm mullet (Osteomugil (Mugil) cunnesius)
Northeast Pacific (California)
N Y Y N N N Y Y N Almedia et al., 1968; Paperna & Overstreet, 1981; Ovcharenko, 2016
Enterobacteriaceae
Aeromonas caviae (= A. punctata) (Aeromonas hydrophila species complex)
Loliginidae Loligo forbesi, Loligo sp. Indo-Pacific (Philippines)
N Y Y (20 days at -20⁰C)
Y N N Y Y N Baldrias & Alvero, 1999; Joseph et al., 2013; Fiorito et al., 2015
Aeromonas hydrophila (cont.)
Carangidae Mediterranean horse mackerel (Trachurus mediterraneus)
Mediterranean Sea (Turkey)
N Y Y Y N N Y Y N Boran et al., 2013; Ozturk & Altinok, 2014; DermNet NZ, 2014
Aeromonas hydrophila (cont.)
Mugilidae Flathead grey mullet (Mugil cephalus)
Mediterranean Sea (Turkey)
N Y Y Y N N Y Y N Paperna & Overstreet, 1981; Ovcharenko, 2016
Aeromonas hydrophila (cont.)
Octopodidae Octopus joubini Widespread N Y Y Y N N Y Y N Fiorito et al., 2015; NZ Fungi, 2019
Aeromonas hydrophila (cont.)
Octopodidae Octopus spp. Widespread N Y Y Y (Zoonotic) N N Y Y N DermNet NZ, 2014; Gestal et al., 2019
Aeromonas salmonicida var. salmonicida (atypical strains)
Clupeidae Atlantic herring (Clupea harengus), Pacific herring (C. pallasii pallasii)
Widespread N Y Y Y N N N Y N Anderson et al., 1994; Evelyn, 2001; Keeling et al., 2013; Brosnahan et al., 2018b
Aeromonas spp. Loliginidae Lollicuncula brevis Northeast Pacific N Y Y Y N N Y Y N Ford et al., 1986; Sherrill et al., 2000; Fiorito et al., 2015
Aeromonas spp. (cont.)
Sepiidae Sepia offinianalis Northeast Pacific N Y Y Y N N Y Y N Hanlon et al., 1984; Ford et al., 1986; Sherrill et al., 2000; Fiorito et al., 2015
Aeromonas spp. (cont.)
Octopodidae Octopus sp. Northeast Pacific N Y Y Y N N Y Y N Hanlon et al., 1984; Ford et al.,1986; Sherrill et al., 2000; Fiorito et al., 2015
Citrobacter freundii Sepiidae Sepia officinalis Europe N Y Y Y N N Y Y N Sherrill et al., 2000; DermNet NZ, 2014
Edwardsiella species complex
Clupeidae Indian oil sardine (Sardinella longiceps)
India N N Y Y N/A N Y Y Y Kumar et al., 2016

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 23
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Edwardsiella species complex (cont.)
Mugilidae Flathead grey mullet (Mugil cephalus)
Widespread N N Y Y N/A N Y Y Y Kusuda et al., 1976; Miyazaki & Egusa, 1976; Evans et al., 2011; Plumb & Hansen, 2011; Park et al., 2012; Kluzik & Woodford, 2016; Haenen, 2017; Reichley et al., 2017
Enterobacter cloacae Mugilidae Flathead grey mullet (Mugil cephalus)
Mediterranean Sea (Turkey)
N Y Y Y N N Y Y N Sekar et al., 2008; Ozturk & Altinok, 2014; Clark et al., 2018; Yow et al., 2018
Escherichia sp. Mugilidae Longarm mullet (Osteomugil (Mugil) cunnesius)
Indo-Pacific (Kuwait)
N Y Y Y N N Y Y N Almeida et al., 1968; Ovcharenko, 2016; NZFungi, 2019
Eubacterium tarantellae
Mugilidae Flathead grey mullet (Mugil cephalus)
Northeast Pacific (California)
N Y (sea lions)
Y Y N N Y N N Austin & Austin, 2007; Buller, 2014; Austin, 2005
Enterococcaceae
Lactococcus garviae (= Enterococcus seriolicida)
Loliginidae, Octopodidae
Squid (Loligo sp.) Octopus (Octopus vulgaris)
Northeast Pacific N Y Y Y N N Y Y N Wang et al., 2007; Farto et al., 2014; DermNet NZ, 2014
Lactococcus garviae (cont.)
Mugilidae Flathead grey mullet (Mugil cephalus)
Northwest Pacific (Chinese Taipei)
N Y Y Y (zoonotic) N N Y Y N Chen et al., 2002; Wang et al., 2007; NZOR, 2019
Chlamydiaceae
Epitheliocystis (Chlamydia-like intracellular disease)
Carangidae Greater amberjack (Seriola dummerili), greenback horse mackerel (Trachurus declivus), yellowtail amberjack (Seriola lalandi), Japanese amberjack (S. quinqueradiata)
Northwest Pacific (Australia, Japan), Southeast Pacific (South America)
N Y Y N N N N Y N Nowak & LaPatra, 2006; Stride & Nowak, 2013; Georgiades et al., 2016
Epitheliocystis spp. (cont.)
Clupeidae Pacific herring (Clupea pallasii pallasii), South American pilchard (Sardinops sagax)
Central Pacific (Australia)
N Y Y N N N N Y N Nowak & LaPatra, 2006; Stride & Nowak, 2013; Georgiades et al., 2016
Bacteroidetes: Flavobacteriaceae
Flavobacterium sp. Loliginidae Lolliguncula (Loligo) brevis Europe N Y Y N N N Y N N Ford et al., 1986; Farto et al., 2014; DermNet NZ, 2014
Tenacibaculum maritimum (Flexibacter maritimus, Cytophaga marina)
Clupeidae Pacific sardine (Sardinops sagax)
Mediterranean (Turkey), Northwest Pacific (Japan), North east Pacific (California)
N Y Y Y N N Y Y N Chen et al., 1995; Anderson, 1996; McGladdery, 1999; Santos et al., 1999; Brosnahan et al., 2017
Tenacibaculum maritimum (cont.)
Engraulidae Anchovy (Engraulis sp.), northern anchovy (E. mordax)
Northeast Atlantic (Europe), South Pacific
N Y Y Y N N Y Y N Diggles, 2011; Ozturk & Altinok, 2014; Diggles, 2016; Brosnahan et al., 2018a

24 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Gammaproteobacteria: Francisellaceae
Francisella spp. Scombridae Atlantic mackerel (Scomber scombrus)
Widespread N N Y Y N N/A Y Y Y Johnston, 2008; Colquhoun & Duodu, 2011
Gammaproteobacteria: Halomonadaceae
Halomonas aquamarina (= Achromobacter aquamarines), H. superficialis
Muglidae Flathead grey mullet (Mugil cephalus)
West Atlantic (USA, Gulf of Mexico)
N N Y N N N/A N Y N Almeida et al., 1968; Paperna & Overstreet, 1981; NZFungi, 2019
Shewanella spp. Octopodidae Octopus vulgaris Northwest Atlantic, Mediterranean
N Y Y N N N Y Y N Farto et al., 2014
Micrococcinaceae: Micrococcaceae
Micrococcus spp. Loliginidae Loligo forbesii, Sepioteuthis lessoniana
Northwest Atlantic, Mediterraneam
N Y Y N N N Y Y N Fiorito et al., 2015; DermNet NZ, 2014
Staphylococcus sp. Loliginidae Lolliguncula brevis Northwest Atlantic, Mediterranean
N Y Y N N N Y Y N Ford et al., 1986; Farto et al., 2014
Renibacterium salmoninarum
Clupeidae Atlantic herring (Clupea harengus) (by experimental infection, not present in wild stocks)
Widespread N N Y Y N N/A N Y N Evelyn, 1993; Wiens, 2011; Hershberger et al., 2013
Cornyebacteriaceae: Mycobacteriaceae
Mycobacterium (=Myxobacterium sp., Mycobacterium marinum, M. fortuitum))
Loliginidae Doryteuthis (Amerigo) pealeii (=Loligo pealei)
Widespread, Europe, Mediterranean
N Y Y N N N Y Y N Fiorito et al., 2015; DermNet NZ, 2014
Mycobacterium spp. (cont.)
Mugilidae Flathead grey mullet (Mugil cephalus)
Northeast Atlantic (Europe), Mediterranean
N Y Y Y (zoonotic)
N N Y Y N Paperna & Overstreet, 1981; Ngan et al., 2005; Varello et al., 2014; Ovcharenko, 2016; NZ Fungi, 2019
Spirochaetales: Leptospira
Leptospira icterohaemorhagiae
Muglidae Flathead grey mullet (Mugil cephalus)
West Atlantic (Gulf of Mexico)
N Y Y N N N Y Y N Paperna & Overstreet, 1981; Ovcharenko, 2016; Anon., 2012; MOH, 2019
Moritellaceae
Moritella (Vibrio) viscosa (winter ulcer disease)
Mugilidae Flathead grey mullet (Mugil cephalus)
Widespread N N Y N N N/A N Y N Austin & Austin, 2007; Tubbs et al., 2007; Björnsdóttir, 2011; Buller, 2014

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 25
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Pseudomonadaceae
Pseudomonas anguilliseptica (Serotype 2 isolates)
Clupeidae Atlantic (Baltic) herring (Clupea harengus membras)
Widespread N N Y Y N N/A Y Y Y Lonnstrom et al., 1994
Pseudomonas stutzeri Octopodidae Octopus (Octopus briareus, O. joubini)
Northwest Atlantic, Mediterranean
N Y Y Y N N Y Y N Ford et al., 1986; Sherrill et al., 2000
P. stutzeri (cont.) Sepiidae Sepia officinalis Northwest Atlantic, Mediterranean
N Y Y Y N N Y Y N Ford et al., 1986; Sherrill et al., 2000
Pseudomonas sp. Mugilidae Flathead grey mullet (Mugil cephalus)
Northeast Atlantic (Europe)
N Y Y Y N N Y Y N Lewis et al., 1970; Paperna & Overstreet, 1981; Ovcharenko, 2016; NZFungi, 2019
Pseudomonas sp. (cont.)
Octopodidae Octopus bimaculoides, Enteroctopus dofleini
Northwest Atlantic, Mediterranean
N Y Y N N N Y Y N Stoskopf et al., 1987; Fiorito et al., 2015; NZFungi, 2019
Pseudomonas sp. (cont.)
Sepiidae Loligo forbesii, Lolliguncula brevis, Sepioteuthis sp., Lessoniana sp.
Northwest Atlantic, Mediterranean
N Y Y N N N Y Y N Hanlon et al., 1984; Ford et al., 1986; Sherrill et al., 2000; Fiorito et al., 2015
Streptococcaceae
Streptococcus agalactiae (Group B type 283)
Clupeidae Pacific sardine (Sardinops sagax)
Indo-Pacific (Singapore, Japan, Kuwait)
N N Y Y Y N/A Y Y NZOR, 2017; Odhiambo et al., 2018
Streptococcus agalactiae (Group B type 283) (cont.)
Mugilidae Klunzinger's mullet (Liza klunzingeri), flathead grey mullet (Mugil cephalus)
Indo-Pacific (Singapore, Japan, Kuwait)
N N Y Y Y N/A Y Y Y Bunch & Bejerano, 1997; Evans et al., 2002; NZOR, 2017
Streptococcus iniae Clupeidae Sardine (Sardinops sagax (= S. melanostictus))
Widespread N N Y Y N N/A Y Y Y Minami et al.,1979; Salati, 2011
S. iniae (cont.) Engraulidae Anchovy (Engraulis japonicus) (= E. japonica)
Northwest Pacific (Japan)
N N Y Y N N/A Y Y Y Minami et al.,1979
S. iniae (cont.) Mugilidae Flathead grey mullet (Mugil cephalus)
Widespread N N Y Y N N/A Y Y Y Johnston, 2008; Kluzik & Woodford, 2016
Streptococcus sp. Mugilidae Flathead grey mullet (Mugil cephalus), thinlip grey mullet (Chelon (Liza) ramada)
Northwest Atlantic (United States), Mediterranean, Northwest Pacific (Chinese Taipei)
N Y Y Y N N Y Y N Plumb et al., 1974; Wang et al., 2001; Chen et al., 2002; Ovcharenko, 2016
Vibrionaceae: Photobacteriaceae
Photobacterium damsela (= P. damsela piscida)
Carangidae Mediterranean horse mackerel (Trachurus mediterraneus)
Mediterranean (Turkey) Widespread
N Y Y Y N N Y Y N Tanrikul & Cagirgan, 2001; Ozturk & Altinok, 2014; NZ Fungi, 2017

26 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Photobacterium damsela (= P. damsela piscida) (cont.)
Mugilidae Flathead grey mullet (Mugil cephalus)
Mediterranean (Turkey) Widespread
N Y Y Y N N Y Y N Lewis et al., 1970; Paperna & Overstreet. 1981; Black, 1997; Ozturk & Altinok, 2014; NZ Fungi, 2017
Vibrionaceae: Vibrio
Vibrio alginolyticus, V. carchariae, V. harveyi, V. parahaemolyticus, V. vulnificus
Carangidae Mediterranean horse mackerel (Trachurus mediterraneus), Atlantic horse mackerel (T. trachurus)
Widespread N Y Y Y N N Y Y N Ozturk & Altinok, 2014; NZOR, 2017
Vibrio spp. (cont.) Mugilidae Flathead grey mullet (Mugil cephalus)
Widespread N Y Y Y N N Y Y N Paperna & Overstreet, 1981
Vibrio spp., V. anguillarum, V. lentus, V. pelagicus, V. splendidus, V. neptunius
Mugilidae Flathead grey mullet (Mugil cephalus), Lebranche mullet (Mugil liza)
Widespread N Y Y Y N N Y Y N Burke & Rodgers, 1981; Paperna & Overstreet, 1981; De Sousa et al., 1999
Vibrio spp. (cont.) Sepiidae Sepia officinalis, Sepia spp., Sepioteuthis lessoniana, Loligo forbesii, Lolliguncula brevis
Northwest Atlantic, Mediterranean
N Y Y N N N Y Y N Reimschuessel et al., 1990; Sherrill et al., 2000; Farto et al., 2003, 2014; Fiorito et al., 2015; Fichi et al., 2015
Vibrio spp. (cont.) Octopodidae Octopus bimaculoides, O. O. briareus, O. maya, O. joubini, O. vulgaris
Northwest Atlantic, Mediterranean
N Y Y N N N Y Y N Reimschuessel et al., 1990; Sherrill et al., 2000; Farto et al., 2003, 2014; Fiorito et al., 2015; Fichi et al., 2015
Cnidaria: Myxozoa
Alataspora serenum Carangidae European horse mackerel (Trachurus trachurus)
Northeast Atlantic, Irish Sea
N N Y N (coelozoic) N/A Y N Y N Campbell, 2005
Alataspora solomoni Carangidae European horse mackerel (Trachurus trachurus), Mediterranean horse mackerel (T. mediterraneus)
Black Sea, Mediterranean
N N Y N (coelozoic) N/A Y N Y N Iurakhno, 1988; Campbell, 2005
Alataspora sp. Mugilidae Thinlip grey mullet (Chelon (Liza) ramada)
Mediterranean N N Y N (coelozoic) N/A Y N Y N Campbell, 2005; Yurakhno & Ovcharenko, 2014
Ceratomyxa auerbachi Clupeidae Atlantic herring (Clupea harengus), Pacific herring (C. pallasi pallasi)
Widespread N N Y N (coelozoic) N/A Y N Y N Rahimian, 2007; Koie et al., 2008
Ceratomyxa australis Carangidae Cape horse mackerel (Trachurus capensis)
Southeast Atlantic (Namibia)
N N Y N (coelozoic) N/A Y N Y N Gaevskaya & Kovaleva, 1979; Campbell, 2005; Sarkar, 2010
Ceratomyxa sardinellae
Clupeidae Indian oil sardine (Sardinella longiceps)
Indo-Pacific (Bay of Bengal)
N N Y N (coelozoic) N/A Y N Y N Sarkar, 2010
Chloromyxum kotorensis
Mugilidae Golden grey mullet (Chelon (Mugil) auratus (= Liza aurata)
Mediterranean, Black Sea
N N Y N (coelozoic) N/A Y N Y N Lubat et al., 1989

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 27
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Chloromyxum marinum
Hemiramphidae Japanese halfbeak Hyporhamphus (Hemirhamphus) sajori
Northwest Pacific (Japan)
N N Y N (coelozoic) N/A Y N Y N Shulman, 1966
Davisia donecae Carangidae Atlantic horse mackerel (Trachurus trachurus), Cape horse mackerel (T. capensis)
Southeast Atlantic, (Namibia)
N N Y N (coelozoic) N/A Y N Y N Gaevskaya & Kovaleva, 1979; Campbell, 2005
Enteromyxum leei (=Myxidium leei)
Mugilidae Flathead grey mullet (Mugil cephalus) and 46 other species from 16 genera
East Atlantic, Mediterranean, West Pacific
N N Y Y N/A N N Y Y Montero et al., 2007
Henneguya ouakamensis
Mugilidae Flathead grey mullet (Mugil cephalus)
East Atlantic (Senegal),
N N Y N N/A Y N Y N Kpatcha et al., 1997; Whipps et al., 2003
Henneguya species 3 Mugilidae Flathead grey mullet (Mugil cephalus)
Southeast Atlantic (Senegal)
N N Y N N/A Y N Y N Faye et al., 1997
Kudoa azevedoi Carangidae Horse mackerel (Trachurus trachurus)
Northeast Atlantic, Mediterranean (Tunisia)
N N Y N N/A Y N Y N Moran et al., 1999a; Eiras et al., 2005; Mansour et al., 2014
Kudoa bora (= Chloromyxum bora)
Mugilidae Flathead grey mullet (Mugil cephalus) (= M. japonicus), keeled mullet (Liza carinata)
West Pacific (Chinese Taipei)
N N Y N N/A Y N Y N Fujita, 1930; Eiras et al., 2005; Ovcharenko, 2016
Kudoa caudata Scombridae Chub mackerel (Scomber japonicus)
Pacific Ocean N N Y N N/A Y N Y N Kovaleva & Gayevskaya, 1983; Li et al., 2013
Kudoa (= Chloromyxum) clupeidae
Clupeidae Alewife (Alosa pseudoharengus), Atlantic menhaden (Brevoortia tyrannus), European herring (Clupea harengus) and 9 other species
Northwest, Southwest Atlantic
N N Y Y N/A Y N Y Y Hahn, 1917; Meglitsch, 1947; Reimschuessel et al., 2003; Langdon, 2007
Kudoa haridasae Carangidae European horse mackerel (Trachurus trachurus)
Indo-Pacific (Bay of Bengal)
N N Y N (coelozoic) N/A Y N Y N Sarkar & Ghosh, 1991; Campbell, 2005
Kudoa haridasae (cont.)
Mugilidae Goldspot mullet (Liza parisa)
Indo-Pacific (Bay of Bengal)
N N Y N (coelozoic) N/A Y N Y N Sarkar & Ghosh, 1991; Campbell, 2005
Kudoa hexapunctata Scombridae Pacific bluefin tuna (Thunnus orientalis)
Northwest Pacific (Japan)
N N Y N N/A Y N Y N Quiazon, 2015; Kasai et al., 2017
Kudoa intestinalis Mugilidae Flathead grey mullet (Mugil cephalus)
Northwest Pacific (Japan)
N N Y N N/A Y N Y N Maeno et al., 1993; Moran et al., 1999a
Kudoa iwatai Mugilidae Flathead grey mullet (Mugil cephalus) and 19 other species
Mediterranean N N Y N N/A Y N Y Y Egusa & Shiomitsu, 1983; Diamant et al., 2005; Ovcharenko, 2016
Kudoa nova (= Kudoa quadratum)
Carangidae European horse mackerel (Trachurus trachurus) and 20 other species
North Atlantic (Norway, Bay of Biscay), Mediterranean
N N Y Y N/A Y N Y Y Lom & Dykova, 1992; Longshaw et al., 2004; Campbell, 2005

28 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Kudoa prunusi (Occurs in farmed fish, but rare in wild stocks)
Scombridae Pacific bluefin tuna (Thunnus orientalis)
Northwest Pacific (Japan)
N N Y N N/A Y N Y N Meng et al., 2011
Kudoa shiomitsui Scombridae Pacific bluefin tuna (Thunnus orientalis)
Northwest Pacific (Japan)
N N Y N NA Y N Y N Zhang et al., 2010
Kudoa tetraspora Mugilidae Flathead grey mullet (Mugil cephalus)
Indo-Pacific (India)
N N Y N N/A Y N Y N Narasimhamurti & Kalavati, 1979
Kudoa thyrsites Clupeidae South African pilchard (Sardinops sagax ocellatus), S. sagax neopilchardus (Western Australia), Sardinella lemuruand 37 other species in 18 families
Southwest, Northwest Pacific (Australia, Japan), South Atlantic (Africa), Northeast Atlantic (England, Europe), Southeast Pacific (Chile), Northeast Pacific (Canada, Alaska)
N N Y Y N/A Y N Y Y Munday et al., 1998; Whipps & Kent, 2006; Henning et al., 2013; Levsen, 2015
Kudoa thyrsites (cont.) Engraulidae Japanese anchovy (Engraulis japonicus) and 18 other hosts
Pacific (Australia, Japan)
N Y Y Y N Y N Y Y Stehr & Whittaker, 1986; Moran & Kent, 1992; Hine et al., 2000; Diggles, 2011
Kudoa trachuri Carangidae Japanese jack mackerel (Trachurus japonicus)
Northwest Pacific N N Y N N/A Y N Y N Matsukane et al., 2011
Kudoa trifolia Mugilidae Flathead grey mullet (Mugil cephalus), thinlip grey mullet (Chelon (Mugil) ramada), Golden grey mullet (C. (Mugil) auratus (= Liza aurata)
Indo-Pacific, Mediterranean
N N Y N N/A Y N Y N Holzer et al., 2006; Ovcharenko, 2016
Kudoa unicapsula Mugilidae Golden grey mullet (Chelon (Mugil) auratus =Liza aurata)
Mediterranean N N Y N N/A Y N Y N Yurakhno et al., 2007
Kudoa yasunagi Scombridae Pacific bluefin tuna (Thunnus orientalis)
Northwest Pacific (Japan)
N N Y N N/A Y N Y N Zhang et al., 2010
Kudoa sp. Carangidae Horse mackerel (Trachurus trachurus)
East Atlantic, Mediterranean
N N Y N N/A Y N Y N Campbell, 2005
Kudoa sp. Mugilidae Flathead grey mullet (Mugil cephalus), fringelip mullet (Crenimugil (Mugil) crenilabis)
Widespread, West Atlantic, Red Sea
N N Y N (coelozoic) N/A Y N Y N Paperna & Overstreet, 1981
Kudoa sp. (cont.) Octopodidae Paroctopus dofleini Japan N N Y N N/A Y N Y N Yokoyama & Masuda, 2001; Lom & Dykova, 2006
Microspora sp. (Microsporaceae)
Sepiidae Sepia sp. Widespread N Y N N N N N Y N Hochberg, 1983; Novis, 2004

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 29
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Myxidium incurvatum Mugilidae Flathead grey mullet (Mugil cephalus)
West Atlantic (USA), Northwest Pacific
N Y Y N N/A Y N Y N Jaysari & Hoffman, 1982; Paperna & Overstreet, 1981; Cairns et al., 2009
Myxobolus adeli Mugilidae Golden grey mullet (Chelon (Mugil) auratus (= Liza) aurata)
Mediterranean, Black Sea, Azov Sea
N N Y N (coelozoic) N Y N Y N Yurakhno & Ovcharenko, 2014
Myxobolus bizerti (= M. hannensis), M. goensis, M. nile
Mugilidae Flathead grey mullet (Mugil cephalus)
Indo-Pacific, Mediterranean
N N Y N N Y N Y N Bahri & Marques, 1996; Eiras & D’Sousa, 2004; Sharon et al., 2019
Myxobolus branchialis (= M. branchiale, Myxosoma branchialis)
Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon (Mugil) auratus (= Liza aurata), leaping mullet (C. (Liza, Mugil) saliens)
Black, Caspian Seas, Northwest Pacific
N N Y N (coelozoic)
N Y N Y N Shulman, 1966; Paperna & Overstreet, 1981; Ibragimov, 1987; Iskov, 1989; Yurakhno & Ovcharenko, 2014
Myxobolus cephalus (= M cephalis)
Mugilidae Flathead grey mullet (Mugil cephalus)
Atlantic Ocean, Gulf of Mexico
N N Y N N Y N Y N Lom & Dykova, 1992; Yurakhno & Ovcharenko, 2014
Myxobolus cheni Mugilidae Flathead grey mullet (Mugil cephalus), So-iuy mullet (Planiliza haematocheila (= M.l haematocheilus, M. soiuy))
Black Sea, Northwest Pacific (China, Japan, Russia)
N N Y N (coelozoic) N Y N Y N Shulman, 1966; Paperna & Overstreet, 1981; Yurakhno & Ovcharenko, 2014
Myxobolus chiungchowensis
Mugilidae Flathead grey mullet (Mugil cephalus)
Northwest Pacific, China
N N Y N (coelozoic) N Y N Y N Eiras et al., 2005
Myxobolus episquamalis
Mugilidae Flathead grey mullet (Mugil cephalus)
East Atlantic, Mediterranean, Pacific (Australia, Korea, Chinese Taipei)
N N Y N N Y N Y N Bahri et al., 2003; Rothwell et al., 1997; Kim et al., 2013b; Lane et al., 2014; Yurakhno & Ovcharenko, 2014
Myxobolus exiguus (=Myxosporidium mugilis)
Mugilidae Golden grey mullet (Chelon (Mugil) auratus) (= Liza aurata), flathead grey mullet (M.l cephalus), thicklip grey mullet (C. labrosus (= M. l chelo)), thinlip grey mullet (C. (Mugil) ramada)
Black Sea, Russia, Mediterranean
N N Y N N Y N Y N Paperna & Overstreet, 1981; Bahri et al., 2003
Myxobolus goensis Mugilidae Flathead grey mullet (Mugil cephalus)
Indo-Pacific (Indian Ocean)
N N Y N N Y N Y N Eiras & D’Sousa, 2004; Eiras et al., 2005
Myxobolus goreensis Mugilidae Flathead grey mullet (Mugil cephalus)
Indo-Pacific (Indian Ocean)
N N Y N N/A Y N Y N Fall et al., 1997; Yurakhno & Ovcharenko, 2014
Myxobolus improvisus Mugilidae Golden grey mullet (Chelon (Liza) auratus)
Mediterranean, Black Sea, Azov Sea
N N Y N (coelozoic) N/A Y N Y N Yurakhno & Ovcharenko, 2014

30 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Myxobolus incurvatum, M. rotundus
Mugilidae Flathead grey mullet (Mugil cephalus)
North, South Pacific (California, New Zealand)
N Y Y N N Y N Y N Jaysari & Hoffman, 1982; Hine & Diggles, 2000; Yurakhno & Ovcharenko, 2014
Myxobolus mugauratus (= M. nile)
Mugilidae Golden grey mullet (Chelon (Mugil) auratus (= Liza aurata))
Indo-Pacific (India)
N N Y N N/A Y N Y N Eiras et al., 2005; Yurakhno & Ovcharenko, 2014
Myxobolus mugcephalus
Mugilidae Flathead grey mullet (Mugil cephalus)
Indo-Pacific (India)
N N Y N N/A Y N Y N Eiras et al., 2005; Yurakhno & Ovcharenko, 2014
Myxobolus mugilis (= M. mugili)
Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon (Mugil) auratus (= Liza aurata)), thicklip grey mullet (C. labrosus (= Mugil chelo))
Mediterranean (Greece, Egypt), Indian Ocean
N N Y N N/A Y N Y N Parenzan, 1966; Paperna & Overstreet, 1981; Eiras et al., 2005
Myxobolus muelleri Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon (Mugil) auratus (= Liza aurata)), thinlip grey mullet (C. (Mugil) ramada)
Black Sea, Russia, Mediterranean (Italy, Tunisia), Southeast Atlantic (Senegal)
N N Y N N/A Y N Y N Paperna & Overstreet, 1981; Bahri et al., 2003; Yurakhno & Ovcharenko, 2014; Sharon et al., 2019
Myxobolus mugchelo Mugilidae Thicklip grey mullet (Chelon labrosus (= Mugil chelo)
Mediterranean N N Y N N/A Y N Y N Eiras et al., 2005; Yurakhno & Ovcharenko, 2014
Myxobolus parvus Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon (Mugil) auratus = Liza aurata)), so-iuy mullet (Planiliza (Liza) haematocheila (= M. l haematocheilus, M. soiuy))
Widespread N N Y N N/A Y N Y N Shulman, 1966; Paperna & Overstreet, 1981; Ovcharenko, 2015
Myxobolus raibauti Mugilidae Flathead grey mullet (Mugil cephalus)
Southeast Atlantic (Senegal)
N N Y N (coelozoic) N/A Y N Y N Eiras et al., 2005
Myxobolus rhodei Mugilidae Flathead grey mullet (Mugil cephalus)
Southwest Pacific (Australia)
N N Y N (coelozoic) N/A Y N Y N Eiras et al., 2005
Myxobolus rotundus Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon (Mugil) auratus =Liza aurata))
Black Sea, Pacific Ocean (California, New Zealand)
N Y Y N (coelozoic) N Y N Y N Chernova, 1967; Jaysari & Hoffman, 1982; Hine & Diggles, 2000; Yurakhno & Ovcharenko, 2014
Myxobolus saranai (= M. brachialis)
Mugilidae Golden grey mullet (Chelon (Mugil) auratus = Liza aurata)
Black Sea N N Y N N/A Y N Y N Paperna & Overstreet, 1981; Ovcharenko, 2015
Myxobolus spinacurvatura
Carangidae Horse mackerel (Trachurus trachurus)
East Atlantic, Mediterranean
N N Y N (coelozoic) N/A Y N Y N Bahri et al., 2003; Campbell, 2005
Myxobolus (= Myxosoma) sp.
Mugilidae Flathead grey mullet (Mugil cephalus), squaretail mullet (Ellochelon vaigiensis (= M.l waigensis))
Indo-Pacific (India), East Atlantic, Mediterranean
N N Y N (coelozoic) N/A Y N Y N Paperna & Overstreet, 1981

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 31
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Myxodavisia (Davisia) donecae
Carangidae Atlantic horse mackerel (Trachurus trachurus), Cape horse mackerel (T. capensis)
Southeast Atlantic (Namibia)
N N Y N (coelozoic) N/A Y N Y N Gaevskaya & Kovaleva,1979; Campbell, 2005
Myxodavisia (Davisia) haldare
Clupeidae Sardinella longiceps Indo-Pacific N N Y N (coelozoic) N/A Y N Y N Sarkar, 2010
Myxosoma branchialis Mugilidae Flathead grey mullet (Mugil cephalus)
Northwest Pacific N N Y N (coelozoic) N/A Y N Y N Paperna & Overstreet, 1981
Ortholinea (Sphaerospora) divergens
Mugilidae Golden grey mullet (Chelon (Mugil) auratus (= Liza aurata))
Black Sea N N Y N (coelozoic) N/A Y N Y N Yurakhno & Ovcharenko, 2014; Ozer et al., 2015a
Ortholinea orientalis Clupeidae Atlantic herring (Clupea harengus), Pacific herring (C. pallasii pallasii), sprat (Sprattus sprattus)
East Atlantic (Denmark) Northeast Pacific (Alaska)
N N Y N (coelozoic) N/A Y N Y N Marty et al., 1998; Karlsbakk & Koie, 2011
Parvicapsula minibicornis
Clupea European sprat (Sprattus sprattus), European herring (Clupea harengus)
Northeast Pacific N N Y N (coelozoic) N/A Y N Y N Jones et al., 2003; Atkinson et al., 2011; Jorgensen et al., 2011; Koie et al., 2013
Polysporoplasma mugilis
Mugilidae Golden grey mullet (Chelon (Mugil) auratus (= Liza aurata)), thinlip grey mullet (C (Mugil) ramada), thicklip grey mullet (C. (Mugil) labrosus)
Mediterranean, Black Sea
N N Y N (coelozoic) N/A Y N Y N Sitja-Bobadilla & Alvarez-Pellitero, 1995; Yurakhno, 2011; Yurakhno & Ovcharenko, 2014; Ovcharenko, 2016
Pseudalataspora pontica
Mugilidae Golden grey mullet (Chelon (Mugil) auratus (= Liza aurata))
Black Sea N N Y N (coelozoic) N/A Y N Y N Dmitrieva & Gaevskaya, 2001
Sphaeromyxa argentinensis
Engraulidae Argentine anchovy (Engraulis anchoita)
Southwest Atlantic (Argentina, Uruguay)
N N Y N (coelozoic) N/A Y N Y N Timi & Sardella, 1998
Sphaeromyxa balbianii Clupeidae Allis shad (Alosa sardina), European pilchard (Sardina (Clupea) pilchardus)
Widespread N N Y N (coelozoic) N/A Y N Y N Lom, 2004
Sphaeromyxa bonaerensis
Engraulidae Argentine anchovy (Engraulis anchoita)
Southwest Pacific (Argentina, Uruguay)
N N Y N (coelozoic) N/A Y N Y N Timi & Sardella, 1998
Sphaeromyxa parva Scomberosocidae Pacific saury (Cololabias saira)
Northwest Pacific (Japan)
N N Y N (coelozoic) N/A Y N Y N Lom, 2004
Sphaeromyxa (= Sphaeromyxum) sabrazesi
Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon (Mugil) auratus (= Liza aurata))
Black Sea, Western Atlantic, Mediterranean (Spain)
N N Y N (coelozoic) N/A Y N Y N Dmitrieva & Gaevskaya, 2001; Yurakhno & Ovcharenko, 2014

32 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Sphaerospora dicentrachi (=S. mugili)
Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon (Mugil) auratus (= Liza aurata)), leaping mullet (C. (Liza, Mugil) saliens), so-iuy mullet (Planiliza (Liza) haematocheila (= Liza haematocheilus, M. soiuyi)), thicklip grey mullet (C. (Mugil) labrosus), thinlip grey mullet (C. (Liza ramada)
East Atlantic, Mediterranean Black Sea
N N Y N (coelozoic) N/A Y N Y N Quaglio et al., 2002; Yurakhno & Ovcharenko, 2014
Sphaerospora testicularis
Clupea European sprat (Sprattus sprattus), European herring (Clupea harengus)
Northeast Atlantic, Mediterranean
N N Y N (coelozoic) N/A Y N Y N Atkinson et al., 2011; Koie et al., 2013
Zschokkella admiranda
Mugilidae Flathead grey mullet (Mugil cephalus)
Black, Mediterranean Seas
N N Y N N/A Y N Y N Yurakhno & Ovcharenko, 2014
Zschokkella dogieli Mugilidae Flathead grey mullet (Mugil cephalus), fringelip mullet (Crenimugil crenilabis)
Indo-Pacific (Japan to Australia)
N N Y N N/A Y N Y N Paperna & Overstreet, 1981
Zschokkella mugilis Mugilidae Flathead grey mullet (Mugil. cephalus), leaping mullet (Chelon (Liza, Mugil) saliens), (thinlip grey mullet (C. (Liza, Mugil) ramada), thicklip grey mullet (C. (Mugil) labrosus)
Mediterranean (Italy, Spain)
N N Y N (coelozoic) N/A Y N Y N Paperna & Overstreet, 1981; Lubat et al., 1989; Qualigo et al., 2002; Yurakhno & Ovcharenko, 2014
Ascomycetes
Cladosporium sp. Octopodidae Eledone cirrhosa Widespread N Y N N N N N Y N Polglase et al., 1984; Simeca & Oestmann, 1995; Harms et al., 2006; Fiorito et al., 2015; NZ Fungi, 2019; Polglase, 2019
Cladosporium sp. (cont.)
Sepiidae European cuttlefish (Sepia officinalis)
Widespread N Y N N N N N Y N Polglase et al., 1984; Simeca & Oestmann, 1995; Harms et al., 2006; Fiorito et al., 2015; NZ Fungi, 2019; Polglase, 2019
Labyrinthula sp. Marine slime mould
Octopodidae Octopus vulgaris Northwest Atlantic, Mediterranean
N Y N N (in aquaria)
N N N Y N Fiorito et al., 2015; Armiger, 1964
Ulkenia amoeboidea Octopodidae Eledone cirrhosa, Octopus vulgaris
Widespread N N N N N/A N N Y N Polglase, 2019; Fiorito et al., 2015; NZFungi, 2019

Import risk analysis: Frozen fish and cephalopod molluscs for fish bait 33
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Schizochytrium sp. Illicinidae Illex illicibrosus Canada N Y N N N N N Y N Jones, 1981; Jones & O’Dor, 1983; NZ Fungi, 2019; Polglase, 2019
Ichthyobodo necator Octopodidae Octopus sp. Northeast Pacific N N N N N/A N Y Y N Forsythe et al., 1991; Poynton et al., 2009
Chromidina coronata Octopodidae Octopus vulgaris, Eledone cirrhosa, Scaeurgus unicirrhus
Mediterranean, Northeastern Atlantic
N N N Y N/A Y N Y N Hochberg, 1983; Souidenne & Furuya, 2019
C. coronate (cont.) Sepiidae Sepiola rondeletii Mediterranean, Northeast Atlantic
N N N Y N/A Y N Y N Hochberg, 1983; Souidenne & Furuya, 2019
C. coronate (cont.) Ommastrephidae Illex coindetiii Mediterranean, Northeast Atlantic
N N N Y N/A Y N Y N Hochberg, 1983; Souidenne & Furuya, 2019
Chromidina elegans Octopodidae Octopus salutii, Mediterranean, Northeast Atlantic
N N N Y N/A Y N Y N Hochberg, 1983; Souidenne & Furuya, 2019
C. elegans (cont.) Sepiidae Sepia elegans, S. orbignyana, Ilex coindetti,
North Pacific N N N Y N/A Y N Y N Hochberg, 1983; Souidenne & Furuya, 2019
C. elegans (cont.) Ilicinidae Dosidicus spp., Toderodes saggitatus
North Pacific N N N Y N/A Y N Y N Hochberg, 1983; Souidenne & Furuya, 2019
Opalinopsis sepiolae (= Opalinopsis octopi)
Octopodidae Octopus macropus, O. tetracirrhus
Mediterranean, Northeast Atlantic
N N N N N/A Y N Y N Hochberg, 1983; Souidenne & Furuya, 2019
O. sepiolae (cont.) Sepiidae Rossia macrosoma, Sepietta oweniana, Sepia rondeletii, S. elegans, Sepiola atlantica
Mediterranean, Notheast Atlantic
N N N N N/A Y N Y N Hochberg, 1983; Souidenne & Furuya, 2019
Conocyema polymorpha
Octopodidae Octopus vulgaris (= O. sinensis)
Mediterranean, Northeast Atlantic
N N N N N/A Y N Y N Furuya & Souidenne, 2019
Aggregata andresi Octopodidae Martialia hyadesi Southwest Atlantic
N N N Y N/A Y N Y N Gestal et al., 2005
Aggregata octopiana, A. spinosa, Aggregata spp.
Octopodidae Octopus vulgaris (= O. sinensis)
Mediterranean, Northwest Atlantic
N N N N N/A Y N Y N Gestal et al., 2002a, 2002b, 2007; Castellanos-Martinez et al., 2013, 2019
Aggregata bathytherma
Octopodidae Muusoctopus hydrothermalis (=Vulcanoctopus hydrothermalis)
Northeast Pacific N N N N N/A Y N Y N Gestal et al., 2010
Aggregata dobelli (Not in WoRMS 2019)
Octopodidae Enteroctopus dofleini Northeast Pacific N N N N N/A Y N Y N Poynton et al., 2002
Aggregata eberthi Sepiidae Sepia officinalis Mediterranean, Northeast Atlantic
N N N N N/A Y N Y N Castellanos-Martinez et al., 2013
Aggregata kudoi Sepiidae Sepia elliptica Northwest Indian Ocean
N N N N N/A Y N Y N Narasimhamurti, 1979
Aggregata millerorum (Not in WoRMS 2019)
Octopodidae Octopus bimaculoides Northeast Pacific N N N N N/A Y N Y N Poynton et al., 1992

34 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogen Fish host family Species known to be susceptible
Host distribution OIE listed disease
Reported from New Zealand
Viable in frozen fish bait
Causes significant disease
Virulent exotic strains
Vector/ intermediate host required
Zoonotic potential
Potential hosts in New Zealand
Retained for risk assessment
Reference
Aggregata saggitatus, Aggregata sp.
Ommastrephidae Todarodes sagittatus Northeast and Western Atlantic, Caribbean
N Y N N N Y N Y N Hochberg & Couch, 1971; Simeca & Oestmann, 1995; Gestal et al.,2002a, 2002b; Diggles et al., 2002; Castellanos-Martinez et al., 2013, 2019
Aggregata spp. Octopodidae Patagonian octopus (Octopus tehuelchus), Southern red octopus (Enteroctopus megalocyathus)
Southeast Atlantic N N N N N/A N N Y N Sardella et al., 2000
Dicyema paradoxum, D. typus, Dicyemennea lameerei
Octopodidae Octopus vulgaris (= O. sinensis)
Mediterranean, Northwest Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019; Gleadill 2016
Pleodicyema delamaeri
Octopodidae Bathypolypus sponsalis Mediterranean, Northwest Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019
Dicyemennea eledones (=Dicyemodeca eledones), D. lameerei
Octopodidae Eledone cirrhosa Mediterranean, Northeast Atlantic
N N N N N/A N N Y N Souidenne et al., 2016; Furuya & Souidenne, 2019
D. moschatum, D. eledones (cont.)
Octopodidae Eledone (Octopus) moschata
Mediterranean, Northeast Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019
D. macrocephalum Octopodidae Macrotritopus defilippi (=Octopus defilippi)
Mediterranean, Northeast Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019
D. macrocephalum (cont.)
Sepiidae Rondeletiola minor, Sepietta obscura, S. oweniana, Sepiola steenstrupiana, Sepia elegans
Mediterranean, Northeast Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019
D. paradoxum Octopodidae Octopus macropus Mediterranean, Northeast Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019
D. rondaletiolae Sepiidae Sepietta neglecta, S. oweniana
Mediterranean, Northeast Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019
D. banyulensis, D. benedeni, D. eledones
Octopodidae Octopus salutii Mediterranean, Northeast Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019
Dicyemennea gracile, Pseudicyema truncatum, D. whitmani, Microcyema vespa
Sepiidae Rossia macrosoma, Sepia officinalis, S. orbignyana
Mediterranean, Northeast Atlantic
N N N N N/A N N Y N Furuya & Souidenne, 2019
Symbols N/A Not applicable Shaded pathogens are retained for risk analysis Viable in frozen fish bait as defined in the current IHS for fish bait (MPI, 2011)

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 35
5.1 Hazard list for risk assessment
As per the findings of the hazard identification in Table 6 the following organisms were
identified as hazards associated with fish bait. Only nervous necrosis virus (NNV) was
associated with coleoid cephalopod molluscs. All organisms identified as hazards were subject to
a risk assessment.
Viral pathogens
Birnaviridae: Marine aquabirnavirus (MABV) including ‘IPNV-like’ strains
Iridoviridae: Red sea bream iridovirus (RSIV), Infectious spleen and kidney necrosis virus
(ISKNV), heart and skeletal muscle inflammation (HSMI) and associated iridoviruses;
Erythrocytic necrosis virus (ENV)/Viral erythrocytic necrosis (VEN).
Nodaviridae-Betanodavirus: nervous necrosis virus (NNV)/ viral nervous necrosis (VNN)/
viral encephalopathy and retinopathy (VER)
Orthomyxoviridae-Isavirus: Infectious salmon anaemia virus (ISAV)
Reoviridae-Aquareovirus: Piscine aquareovirus (PRV)
Rhabdoviridae: Infectious haematopoietic necrosis virus (IHNV), Viral haemorrhagic
septicaemia virus (VHSV)
Bacterial pathogens
Enterobacteriaceae: Edwardsiella spp.
Gammaproteobacteriaceae: Francisella spp.
Pseudomonadaceae: Pseudomonas anguilliseptica
Streptococcaceae: Streptococcus agalacteae, Streptococcus iniae
Myxozoan pathogens
Enteromyxum leei, Kudoa clupeidae, K. iwatai, K. nova, K. thyrsites
General considerations
6.1 Risk assessment considerations
The following presents general, non-disease specific information that was taken into
consideration in the risk assessment.
For the purposes of this risk analysis, fish bait is non-viable. It has either been frozen (to at least
-18 °C or -20 °C, for at least 168 hours (7 days)) consistent with (Johnston, 2008; USDA, 2019);
or, irradiated with 25 kGy (=2.5 Mrads) consistent with MP (2011). It is wild-sourced, caught in
marine waters (MPI, 2011) and limited to the species defined in the Commodity Definition
(Table 5).

36 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Fish bait has been derived from wild populations of marine finfish and cephalopods, which may
harbour pathogens. This commodity is recognised as an open or poorly regulated pathway for
pathogens through international trade (Biosecurity Australia, 2002; Dalton, 2004; Goodwin et
al., 2004; Arkush et al., 2006; Oidtmann et al., 2011; Phelps et al., 2013, 2014). As a traded
commodity, fish bait is caught, transported and stored in bulk, with no procedural steps (no
international standards) in the supply chain targeting pathogen reduction (Phelps et al., 2013,
2014; OIE, 2019b). Similarly, fish bait is imported in bulk and is not subject to any pathogen
reduction steps in the supply chain before entry into the aquatic environment.
Little information is available on the prevalence of most aquatic animal diseases in wild marine
fish stocks (Peeler et al., 2007; Peeler & Taylor, 2011), particularly for low value species such as
most fish bait (Dunn et al., 2012).
6.1.1 Entry assessment
A qualitative assessment of the likelihood of pathogen entry through the imported commodity is
provided for each identified hazard. Indicators may include, but are not limited to, the volume of
imports of the host species, the geographical distribution of the pathogen and the host species, as
well as the prevalence and persistence of infection in the host. The influence of climate change in
determining and extending the range of both pathogen and host is poorly understood (Harvell et
al., 2009; Burge et al., 2014).
6.1.2 Exposure assessment
Bait fish and coleoid cephalopods are schooling species (Maes & Ollevier, 2003; Merino et al.,
2014). They may be more susceptible to disease transmitted through close contact than solitary
fish species (Burge et al., 2014), but factors affecting the establishment of exotic pathogenic
organisms in susceptible aquatic hosts are poorly known (Munro et al., 1983; Peeler et al., 2007;
Jereb et al., 2010; Peeler & Thomas, 2011; Dunn et al., 2012 Engelhard et al., 2014).
Indicators affecting the assessment of likely exposure, establishment and spread include, but are
not limited to, persistence of the pathogen in the aquatic environment, infective dose, routes of
infection and methods of transmission (including vectors), host range (including conspecifics
and reservoir organisms), host density and geographical distribution, susceptible host life stages
and presence of obligate intermediate hosts (Phelps et al., 2013).
Exposure may also be influenced by natural risk mitigation features. These include abiotic
factors such as water temperature, dilution (of pathogen concentration below the infectious dose)
and distribution of the infective agent by ocean current hydrodynamics (Phelps et al., 2013;
Burge et al., 2014), while climate driven temperature changes may influence the host immune
response and affect the frequency of disease (Bowden, 2008; Burge et al., 2014).
While marine waters do provide a significant dilution factor, the distribution of both fish hosts
and pathogenic agents is not random (Suttle, 2007; Oidtmann et al. 2017). The fish hosts have
evolved behavioural responses to a complex set of coastal currents (Hume et al., 1992). Fishing
activity is also not random but is strongly directed to maximise contact with fish species (FAO,
2018). It is reasonable to assume that with the high number of fishhook sets, with an estimated
bait-loss of up to 50% (Skud et al., 1978) and the use of berley as a fish attractant (Wynn-Jones
et al., 2019), at least some infected fish bait may contact a suitable host species and allow a
pathogen to complete its life cycle.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 37
Biotic factors that may provide natural mitigation are complex. Available data for many marine
pathogens are usually confined to reported presence in a host. Comparatively little is known
about the epidemiology of aquatic infectious diseases compared to terrestrial vertebrate diseases
(Kermack & McKendrick, 1991; Burge et al., 2014). The processes leading to epizootics are
poorly understood (Bidegain et al., 2016).
Available data on host susceptibility, morbidity, mortality, or infectious dose have generally
been determined from experimental studies. Where studies have used unnatural infection
pathways (such as intra-peritoneal injection) the results must be treated with caution because
such data may not reflect the likely natural exposure potential in marine waters. Where study
methods reasonably simulate natural exposure pathways (such as infection per os),
epidemiological data (such as susceptibility of host species) the results are more reliable (OIE,
2019a).
Laboratory-based estimates of pathogen virulence and survival should be interpreted as
maximum estimates. They do not adequately take into consideration the influence of the aquatic
environment, including dilution factors, or predation. For example, viral and bacterial pathogens
may be consumed by protozoans in marine waters while infected host fish may be consumed by
predators before the pathogen life cycle is complete (Burge et al., 2014; Hallett et al., 2015;
Jones et al., 2015). Even where disease establishment occurs more quickly, infections may be
unapparent or unnoticed in wild fish stocks (Sindermann, 1987; Jones et al., 2015). This is most
likely to occur for diseases of lower economic importance, or for lower value host species (Vike
et al., 2014; Burge et al., 2014).
Given these uncertainties, the likelihood of exposure and establishment for pathogens associated
with fish bait has been capped as “medium” to better reflect the associated risk.
The potential distribution pathways whereby viable pathogens associated with fish bait can directly
enter natural waters are shown in Figure 2. In addition to the intended end use as fish bait, other
potential pathways include diversion of imported fish bait to other end uses (e.g. fish food) or
waste from further processing.
A smaller volume is destined for retail sale. This may be part-thawed and re-packaged through the
wholesale and retail distribution process, prior to retail sale. Pathogens may remain viable in liquid
wastes from thawing during this process and enter the aquatic environment. Where disposed of to
landfill, leachates and effluent harbouring pathogens may also enter aquatic environments either
directly or be dispersed through the action of piscivorous and scavenging birds (Blackwell, 2019).
The relative importance assigned to each pathway is shown in Figure 2, categorised as probable,
less significant or unlikely. These pathways are considered where relevant for each identified
pathogen. The likelihood of introduction of significant pathogens through redirection of fish from
the human consumption pathway (Blackwell, 2019) is likely to be very low and is not considered
further in this risk analysis.

38 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Figure 2. Potential distribution pathways for frozen fish or coleoid cephalopod mollusc fish bait
New Zealand marine fish aquaculture is largely focused on Chinook (King) salmon
(Oncorhynchus tshawytscha) (Family Salmonidae) on-grown in sea-cages. Chinook salmon
exports alone were valued at NZ$77 million in 2018 (Seafood New Zealand, 2019). Salmonid
aquaculture in New Zealand exclusively uses formulated fish food (Seafood New Zealand,
2019).
Wild fish species may become infected through imported fish bait. These may enter the sea
cages and be consumed by salmon. Recreational fishing activity commonly occurs immediately
adjacent to these sea cages, as these act as fish aggregation devices (Fisheries New Zealand,
2020). Discarded fish bait may also drift into a marine farm and recreational fishers may discard
their unused fish bait into the fish farm.
Other developing marine aquaculture fish species in New Zealand include snapper (Pagrus
auratus) (Family Sparidae), yellowtail kingfish (Seriola lalandi) (Family Carangidae), hapuku
(Polyprion oxygeneios) (Polyprionidae) (NIWA, 2020; Plant & Food, 2020). These developing
fisheries may use imported fish bait as fish food, as no artificial fish foods have yet been
developed. These activities represent viable risk pathways between wild fisheries and
commercial aquaculture.
6.1.3 Consequence assessment
A qualitative assessment of the consequences of pathogen exposure through the imported
commodity is provided for each identified hazard. This considers the impact on people
(including zoonotic potential, societal values and cultural values), the New Zealand environment
and the New Zealand economy (Biosecurity Act 1993).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 39
The economic consequences are comparatively easy to determine. Direct effects include, but are
not limited to, loss of volume or value of export trade, reduced productivity, increased mortality,
or reduced marketability of infected stocks (Dobson & May 1987). Indirect effects include
restrictions on New Zealand exports and trade access, as well as the costs of mitigation measures
and disease control processes (Biosecurity New Zealand 2006).
Little published information is available to determine the social and environmental consequences
of aquatic pathogen establishment (Munro et al. 1983; Peeler & Taylor 2011; Groner et al.
2016). Social effects include, but are not limited to, the direct consequences for human health of
zoonotic pathogens, the effects on traditional Maori customary fishing, cultural, spiritual
environmental and economic values, other cultural values and perceptions, as well as the effect
on recreational fishing activity. Indirect social effects include changed perceptions or job losses
in associated industries (such as tourism) resulting from the introduction of a risk organism
(Biosecurity New Zealand 2006). Indicators may be from non-market analysis (Southwick et al.
2018) or changes in social perception of amenity values (Biosecurity New Zealand 2006).
Environmental consequences may be direct or indirect. Direct consequences on indigenous fauna
include, but are not limited to, increased mortality and lower productivity, muscle degeneration,
liver dysfunction, interference with nutrition, cardiac disruption, nervous system involvement,
castration or mechanical interference with spawning, weight loss, and gross distortion of the
body (Sindermann 1987). These effects may only be apparent in wild fish stocks during
epizootics, due to high rates of predation on sick fish and scavenging of dead fish.
Indirect environmental effects may result from pathogen-induced changes in food webs. Bait fish
species such as clupeids feed on phytoplankton, zooplankton and detritus and represent a major
link between trophic levels in marine food webs. Bait fish also provide a significant food source
for other fish, seabirds and marine mammals (Dunn et al. 2012).
6.2 Risk management considerations
Risk management measures may be applied to reduce the risk associated with a specific risk
organism. These can be applied either pre-border (offshore) or post-arrival (where the shipment
is held in a transitional facility). Risk management measures are aimed at a particular point of the
risk pathway, typically the likelihood of entry.
Risk management measures may be applied singly or in combination with other measures.
The measures applied need to be commensurate with the perceived risk. Not all risk management
measures proposed may need to be applied at the present level of risk. Other identified measures
may be applied if and when the perceived level of risk changes.
6.2.1 General risk management measures
Identification of the species included in the commodity
This is intended to identify fish bait species that are not associated with any identified risk
organisms.
Declaration of the country of capture
Not all fish bait is caught within the territorial seas of a fishing nation. Some species such as
squid are taken in international waters. The declaration of country of capture is intended to

40 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
establish that the consignment has been sourced from areas where the risk organism has not
previously been reported.
Declaration that the commodity is sourced from wild stock
Fish bait species are of relatively low value. Species including flathead grey mullet (Mugil
cephalus) are farmed both for human consumption and to supply the international demand for
fish food and fish bait in the Mediterranean, United States and Southeast Asia (FAO, 2020b). It
is likely that the prevalence of infection in farmed fish bait may be higher than the assumed
prevalence of 2% to 4% generally associated with wild fish stocks (Corsin et al., 2009).
Declaration that the commodity has not been harvested from populations experiencing an
epizootic disease.
Fish bait species commonly occur in schools and wild stocks are subject to epizootics where risk
organisms may be endemic. The prevalence of infection during an epizootic may exceed 50%
(Reno et al., 1985; de la Pena et al., 2011). Where the commodity is used for human
consumption, diseased fish may be redirected to the fish bait pathway as a common method of
disposal. It is important to ensure that the commodity should only comprise healthy fish.
Declaration that the commodity has no visible signs of disease.
This implies the fish have been inspected under the oversight of the Competent Authority, where
appropriate standards have been established and followed. This does not mean actual inspection
is necessary.
Declaration of frozen storage
The current IHS for fish bait (MPI. 2011) requires a declaration that the commodity has been
frozen (to below -18 °C to -20 °C for at least 18 hours). The risk analysis for filleted
Oreochromis sp. (Johnston, 2008) proposed a minimum frozen storage period of at least 7 days
(168 hours, at -18°C to -20°C). Quick freezing and frozen storage will denature most metazoan
organisms (Hamed & Elias, 1970; Hauck, 1977; Barlow & Sleigh, 1979; Paperna & Overstreet,
1981; McVicar, 1982; Alderman & Polgase, 1986; Fayer & Nerad, 1996; Morris et al., 1998;
Olson et al., 1999; Shaw et al., 2000; Oidtmann et al., 2002; Racz & Zemankovics, 2002;
Johnston, 2008; Gregg et al., 2012; Leiro et al., 2012; Noga, 2012; Phelps et al., 2013;
Hershberger et al., 2015; Blackwell, 2019).
While the mechanisms of cell death are poorly understood (Pegg, 1987), factors include the
formation of ice crystals within and between cells, cell shrinkage, the destruction of cell
organelles and the concentrations of cellular metabolites. Ice crystals destroy external cell
membranes resulting in the loss of cellular integrity causing cell shrinkage. Ice crystals also
cause damage to internal cell membranes and disrupt organelle function. Ice formation also
dehydrates the cytoplasm, concentrating cell metabolites. These may reach toxic levels resulting
in cell death (Pegg, 1987; OHSU, 2020).
Viability may decrease over time, so extended frozen storage may be necessary to ensure
denaturation (Conlan et al., 2011). For many organisms, it is the freeze-thaw cycle, rather than
the length of freezing that leads to cell death (OHSU, 2020).
Most viruses and many gram-positive bacteria are unaffected by multiple freeze-thaw cycles and
extended frozen storage (Sourek, 1974; Meyers et al., 1999; Gaughan, 2001; Arkush et al., 2006;
Hervé-Claude et al. 2008; OIE, 2019b). Viruses are non-living and generally stable in
temperatures between + 20 °C and -80 °C, without alteration of their molecular structure

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 41
(Mortensen et al., 1998). Frozen storage (-20 C to -10°C) has been used for long-term
preservation of viruses in fish tissue (Plumb et al., 1973; Burke & Mulcahy 1983; Plumb &
Zilberg 1999) and for storage of pathogenic bacteria (Sourek, 1974).
The bacterium Edwardsiella spp. and myxozoan pathogens (Enteromyxum leei, Kudoa
clupeidae, K, iwatai, K. nova and K. thyrsites) are only slightly affected by short term (7 days)
frozen storage (Yurakhno. 2017; Castro et al., 2006; Johnston, 2008), but are substantially or
completely denatured following extended (at least 4 months) frozen storage (Brady &
Vinitnantharat, 1990; Arkush et al., 2013; Hoffman & Putz, 2011; Ohnishi et al., 2016; Anon.
2020b).
For the purposes of this risk analysis, it is proposed that the period of frozen storage be extended
to at least 4 months (17 weeks), for all fish bait, to denature Francisella spp., Enteromyxum leei,
Kudoa clupeidae, K. iwatai. K. nova and K. thyrsites. This may be achieved by either:
(a) a declaration that the commodity has been previously frozen for at least 4 months (17
weeks) at a temperature of -18 °C to -20 °C or below, from the date of capture; or,
(b) a requirement that the imported commodity must remain in frozen storage (at -18 °C to
-20 °C) in a BNZ-approved Transitional Facility, for this minimum time period, before
release.
Labelling the commodity as “Bait only, unfit for human consumption”.
While little actual control is possible post-border, the risk to human health can be reduced by
appropriate labelling of the commodity.
6.2.2 Pathogen specific risk management options
Sourcing fish bait from listed species not associated with pathogens of concern
Species prohibition includes identifying host species for identified pathogens of concern. From
the finfish and coleoid cephalopod molluscs identified as fish bait (Table 5) some species are
considered susceptible to identified risk organisms. Other fish bait species are unlikely to
represent a risk in the commodity.
Region/Country freedom
Fish bait may be sourced from regions or countries recognised by BNZ as being free of
corresponding pathogens of concern. Acceptance of region/ country freedom may be provided
through the BNZ Country Approval Procedures. For OIE-listed diseases, a certificate of country
freedom is provided by the Competent Authority of the exporting country. For non OIE-listed
diseases, acceptance of country freedom through the BNZ Country Approval Procedures would
be possible by a Health Certificate provided by the Competent Authority stating that the risk
organism had not been reported from the country of capture.
Pre-export or post-arrival irradiation (ionising radiation to 50 kGy)
Ionising radiation is commonly applied to fresh food commodities to extend shelf life by
denaturing food-spoiling bacteria (DAFF, 2013). Doses in the range of 1–10 kGy are unlikely to
change the nature of the commodity and 10 kGy is the maximum recommended for commodities
intended for human consumption (Anon., 1981, 1984; Crawford & Ruff, 1996). A minimum
dose of 3 kGy is recommended for frozen fish and shellfish product for human consumption
(IAEA, 2000). A dose of 1 kGy is recommended for denaturing microorganisms in fruit and
vegetables, and up to 30 kGy for bacterial decontamination of herbs and spices (FSANZ, 2020).

42 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
A dose of 10 kGy is equivalent to heat of energy of 10 J/kg and the heat capacity of water is 4.2
J/kg, therefore a dose of 10 kGy should raise the temperature by 2.4 °C (Loaharanu, 1997). For
food irradiation, a dose of 1 to 5 kGy is estimated to raise the temperature of the fish bait by 0.24
to 1.2 °C (Loaharanu, 1997).
Higher doses to 25 kGy are commonly used for the ‘bacterial sterilisation’ of medical equipment
and pharmaceuticals (DAFF, 2013) and as a biosecurity application to inactivate most bacterial,
fungal and metazoan pathogens likely to be present in imported fish and fish food (MPI, 2011).
Following Loraharanu (1997), a dose of 25 kGy would be expected to raise the product
temperature by 6 °C.
Viral pathogens present in frozen fish bait may require a higher dose to ensure denaturation.
Further, higher doses are generally required to achieve ‘radiation sterilisation’ and ensure viral
denaturation in frozen tissues, as compared to fresh or chilled commodities (DAFF, 2013). A
dose of 50 kGy, or the application of a 25 kGy dose twice is recommended for inactivation of
pathogens of biosecurity concern (DAFF, 2013). This seemingly high dose of radiation may be
suitable for frozen fish bait as the commodity is not intended for human consumption. Use of a
50 kGy dose could increase the product temperature by 12 °C (Loaharanu, 1997). However,
where the commodity is irradiated twice at 25 kGy (DAFF, 2013), the temperature increase
would remain at 6 °C. It is likely that the effect on frozen fish bait would be minimal.
It is acknowledged that no suitable irradiation facility is currently available in New Zealand.
However, an overseas facility may be approved by BNZ to undertake the irradiation of bait (N.
Ahmed, BNZ, pers. comm., 2020). Approved overseas irradiation facilities are used for
biosecurity irradiation by the Australian Government (DAWR, 2016).
Fish bait industry sector representatives (B. Burney, pers. comm., 2020; M. Lyford, pers. comm.,
2020) have expressed concerns that costs of overseas irradiation may make the product cost
prohibitive. Further, there is concern that irradiation may alter the nature of the product by
softening the tissues and reducing the volume of blood and fluids in the thawed product that
attract fish. Additional tests may be necessary to determine whether irradiated bait product
remains fit-for-purpose. Further, a lower irradiation dose may be appropriate to denature the risk
organisms associated with a particular fish bait species. This may be determined on a case-by-
case basis, as necessary. Irradiation, however, may be still be a viable risk management option
for the commodity.
Heat treatment
Heat treatment is a viable risk management option for fish for human consumption (OIE, 2019a;
Blackwell, 2019) or for processed fish food based on fish meal (Cobb, 2009). However, heating
alters the nature of the commodity, making it unfit for use as fish bait (B. Burney, pers. comm.,
2020; R. Clarke, pers. comm., 2020; M. Lyford, pers. comm., 2020). Heat treatment is not a
viable risk management option as the commodity definition is for frozen fish bait. Heat treatment
is not considered any further.
Chemical treatment
Surface disinfectant denature most risk organisms (Bovo et al., 2005; Yoshimizu et al., 2005;
Watanabe & Yoshimizu, 1998; OIE, 2019b). These chemicals have poor penetration and are
unsuitable for treatment of whole fish bait (Fraser et al., 2006; Phelps et al., 2013; Chua et al.,
2019).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 43
Chemical bath processing methods are effective against bacteria and viruses present in whole
fish bait in laboratory trials (Faisal et al., 2012; Phelps et al., 2013; Tang et al., 2016). These
methods are either impractical for commercial-scale use for whole fish bait (N. Phelps, pers.
comm., 2020), or too toxic for general use (A. Pearson, BNZ, pers. comm., 2019). Chemical
processing methods are not viable risk management options and are not considered further.
Pre-export or post-arrival batch testing
OIE-approved batch testing methods (Corsin et al., 2009; OIE, 2019a) and standard
microbiological or molecular testing protocols are available to certify the absence of identified
risk organisms in the commodity. Pre-border testing procedures may be carried out by a testing
laboratory certified by the Competent Authority of the exporting country. Acceptance of a
negative batch test result by BNZ through the BNZ Country Approval Procedures may be a
viable risk management option (N. Ahmed, BNZ, pers. comm., 2020).
These risk management options are evaluated for each risk organism in the appropriate chapter.
6.2.3 Estimated levels of pathogen reduction
Qualitative descriptors are used to categorise estimated levels of reduction in pathogen load
considered likely to result from the application of each risk management option and assist in the
selection of risk management options for each risk organism. The indicative levels are:
Eliminate > 95% reduction in pathogen load
Substantial 71–95% reduction in pathogen load
Moderate 51–70% reduction in pathogen load
Slight < 50% reduction in pathogen load
Birnaviridae: Marine aquabirnavirus
7.1 Technical review
7.1.1 Aetiological agent
Marine aquabirnavirus (MABV) is a double-stranded RNA virus in the Family Birnaviridae,
which includes infectious pancreatic necrosis virus (IPNV) and related strains. Genomic analysis
indicates the strains of IPNV identified from salmonids are clustered into 6 genogroups (MABV
genogroups 1-6). A seventh genogroup includes 28 MABV strains associated with non-salmonid
fish. As IPNV is only associated with salmonids, other related MABV strains are considered as
IPNV-like MABV strains (Blake et al., 2001).
IPNV-like MABV strains associated with fish bait include yellowtail ascites virus (YAV) and
viral deformity virus (VDV) isolated from yellowtail (Seriola quinqueradiata) in Japan
(Sorimachi & Hara, 1985; Nakajima et al., 1998). These strains have low host specificity.
Strains isolated from different non-salmonid hosts and geographical areas are highly homologous
(more than 98.5% identical) (Hosono et al., 1996; Zhang & Suzuki, 2004; (Crane & Hyatt, 2011;
Shin et al. 2011; B. Jones, pers. comm., 2015). Each strain exerts different mortality rates in
different hosts depending on the water quality (temperature, salinity) and stocking levels (Barja,
2004; Crane & Hyatt, 2011).

44 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
7.1.2 OIE status
Infection with MABV is not listed by the OIE (OIE, 2019a).
7.1.3 New Zealand status
Infection with IPNV is regarded as an exotic, notifiable disease in New Zealand (Johnston, 2008;
Anon., 2016). Related MABV strains of genogroup 5 have been reported from farmed New
Zealand turbot (Colistium nudipinnis), flounder (Rhombosolea spp.) and from wild Chinook
salmon (Oncorhynchus tshawytscha) (Tisdall & Phipps, 1987; Anderson, 1996; Johnston, 2008).
Other non-salmonid MABV strains reported from fish bait species overseas (including VDV and
YAV) have not been reported from New Zealand (Anderson, 1990; Diggles et al., 2002; Tubbs
et al., 2007).
7.1.4 Zoonotic disease
MABV strains are not zoonotic (CABI, 2019a).
7.1.5 Epidemiology
Distribution and host range
Over 100 species representing more than 30 families of marine finfish are susceptible to IPNV
and IPNV-like MABV strains (Blackwell, 2019). IPNV strains also occur naturally in mussels
(Mytilus galloprovincialis, M. edulis) and oysters (Pinctada fucata), with moderate clinical signs
(Hill & Alderman, 1979) while scallops (Pecten spp.) can be experimentally infected. Shellfish
isolates cause clinical signs of IPNV in rainbow trout (Oncorhynchus mykiss) in experimental
infection (Hill, 1982; Mortensen et al., 1992; Isshiki et al., 2001; Molloy et al., 2013).
MABV strains have low host specificity. The species associated with MABV strains in fish bait
and their geographic/pathogen range are given in Table 7 (Nakajima et al., 1998; Kahn et al.,
1999; Crane et al., 2000; Isshiki et al., 2001; Crane & Williams, 2008; McColl et al. 2009a;
Crane & Hyatt, 2011; Munro & Midtlyng, 2011; Diggles, 2011;Van Beurden et al., 2012).
Prevalence ranges from 0.1% to 12.5% in saithe (Pollachius virens) and flounder (Platichthys
flesus) (Wallace et al., 2008; Diggles, 2011). IPNV prevalence was higher in wild non-salmonid
fish caught less than 5 km from infected marine salmon farms in Scottish waters, suggesting fish
farms may act as a reservoir for infection (Wallace et al., 2008, 2017). MABV strains including
IPNV were not reported in European anchovy (Engraulis encrasiciolus) or Atlantic mackerel
(Scomber scombrus), from a survey of 37 species of marine fish undertaken in coastal Scottish
waters (Wallace et al., 2008).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 45
Table 7. Fish bait species susceptible to marine aquabirnavirus (MABV)
Family Host species Host geographical range1 Pathogen range2
Carangidae Japanese jack mackerel (Trachurus japonicus)
Northwest, Central Pacific West, Northwest Pacific (Asia, Japan, South Korea)
Clupeidae Atlantic herring (Clupea harengus), Bali sardinella (Sardinella lemuru), Indian oil sardine (Sardinella longiceps), Pacific herring (C. pallasii pallasii), round sardinella (Sardinella aurita), South American pilchard (Sardinops sagax), spotted sardinella (Ambylygaster sirm)
West Atlantic (USA to Argentina), Mediterranean, Western Pacific Japan, Indo-Pacific
East, Western Atlantic, Mediterranean, Western Pacific (Asia, Japan, South Korea)
Scombridae Atlantic mackerel (Scomber scombrus)
East Atlantic (Gulf of Cadiz) East Atlantic
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature)
Potential host species in New Zealand
Potential host species of MABV include farmed Chinook salmon (O. tshawytscha) and
yellowtail kingfish (Seriola lalandi). MABV potentially affects invertebrates including mussels
(Mytilus spp.) and scallops (Pecten spp.) (Tubbs et al., 2007).
Pathogenicity
MABV strains are associated with economically serious diseases affecting wild and farmed fish.
Mortalities of up to 100% occur in salmonids (Oncorhynchus spp., Salmo spp. and Salvelinus
spp.) (Barja, 2004; Ruane et al., 1997; Skjesol et al., 2011).
Mortalities in non-salmonid fisheries are variable. Mortality ranges from 5% to 100% in farmed
eels (Anguilla anguilla) and from 80% to 90% in farmed Japanese amberjack (Seriola
quinqueradiata) (Nakajima et al., 1998; Van Beurden et al., 2012).
Clinical infection is rarely associated with wild fish, but subclinical infection may be endemic
(Van Beurden et al., 2012). Surviving fish are carriers of disease (Kahn et al., 1999).
Pathogenicity varies with season, size and age of fish, where older fish are less susceptible to
clinical infection (Isshiki et al., 2001).
The minimum infectious dose is low (> 1 TCID50 mL-1 (= 0.7 × 101 pfu mL-1) as determined by
bath immersion (4 h at 10 °C) (Smith et al., 2000; Isshiki et al., 2001; Urquhart et al., 2008;
Murray et al., 2005; Bebak & McAllister, 2009; Munro & Midtlyng, 2011; Oidtmann et al.,
2017).
Host specificity is low and cross-infection occurs between host species (Kahn et al., 1999;
Isshiki et al., 2001; Munro & Midtlyng, 2011; Van Beurden et al., 2012). Strains isolated from
Atlantic cod (Gadus morhua) caused 20% mortality in Atlantic salmon (S. salar) (Urquhart et
al., 2009), while strains from Japanese amberjack (Seriola quinqueradiata) were pathogenic to
spotted knifejaw (= gold stripe parrotfish) (Oplegnathus punctatus), barred knifejaw (=Japanese
parrotfish) (O. fasciatus) and yellowtail amberjack (S. lalandi) (= gold stripe amberjack, S.
aureovittata) (Isshiki et al., 2001).

46 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Clinical signs
External clinical signs are variable. Subclinical carriers typically show no external signs.
Clinically infected fish may have a distended abdomen, moderate exophthalmia and
haemorrhages on the ventral skin surface and ventral fin base (Munro & Midtlyng, 2011).
Pathogen survival
MABV strains are unaffected by frozen storage (Kahn et al., 1999). They remain viable in
fomites (shipping containers, processing equipment and bait storage boxes) for at least 8 weeks
(Desautels & MacKelvie, 1975; Munro & Midtlyng, 2011). They remain viable for extended
periods in dead and decaying fish at high titre (> 106.0 pfu g-1) (plaque forming units) (Isshiki et
al., 2001; Munro & Midtlyng, 2011).
MABV strains remain viable in marine coastal waters for up to 35 days (at 10 °C to 15 °C)
(Toranzo et al., 1983; Kitamura & Suzuki, 2000; Kitamura et al., 2004; Oidtmann et al., 2017).
MABV strains also survive passage through piscivorous birds, where viral titre in faeces may
exceed the minimum infectious dose (McAllister & Owens, 1992). They can remain viable in
fresh waters for 17–43 days (at 10 °C to 15 °C) (Desautels & MacKelvie, 1975; Tu et al., 1975;
Ahne, 1982; Raynard et al., 2007; Oidtmann et al., 2017). However, data derived from
experiments may not reflect length of infectivity in natural waters (see section 6.1.2).
Transmission
Transmission is direct. This may be vertical (via infected eggs), or horizontal through
consumption of infected bait, cohabitation (through infected water, fish urine, faeces, and sexual
fluids) or through fomites (vehicles, nets, utensils associated with human activities such as
aquaculture and fish processing) (Munro & Midtlyng, 2011). Surviving fish are life-long latent
carriers and typically shed high levels of viral particles (Urquhart et al., 2008; Munro &
Midtlyng, 2011). Wild fish may function as a reservoir of infection for aquaculture species
(Castric, 1997).
MABV strains may be transported at least 30 km in coastal marine waters, even in slow currents
(2 cm sec-1), at titres exceeding the minimum infectious dose (Smith et al., 2000; Isshiki et al.,
2001; Urquhart et al., 2008; Murray et al., 2005; Bebak & McAllister, 2009; Munro & Midtlyng,
2011; Oidtmann et al., 2017).
MABV strains may be dispersed from fish bait discarded in landfill to waterways by piscivorous
birds (Munro & Midtlyng, 2011). They may also be dispersed between waterways (Desautels &
MacKelvie, 1975; Raynard et al., 2007). They are reported in rivers (at a titre of 1.3 × 104 pfu L-
1) up to 19 km from an infected freshwater site (Smith et al., 2000).
Infection and disease progression
Pathogen entry occurs by ingestion, usually through the intestinal epithelium or across the gill
membranes (Munro & Midtlyng, 2011). All life stages may be infected (DFO, 2017). Infection is
typically focused on tissues of the pancreas, kidney, spleen, pyloric caecae, intestine and liver,
but may also be present in the brain, gonads and gills (McColl et al., 2009; Munro & Midtlyng,
2011; Roberts, 2012).
Infected fish may shed high viral titres (from 101 to 107.5 pfu L-1) in body fluids, depending on
the virus strain, host species and age (Billi & Wolf, 1969; Johansen et al., 2011). However,
factors influencing morbidity appear complex and likely to also include environmental stressors

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 47
(e.g. temperature, salinity and oxygen concentration) (Munro & Midtlyng, 1999; Isshiki et al.,
2001).
Inactivation
MABV strains are unaffected by either short (30 days) or long-term (4 months) frozen storage (at
-20 °C) (Kahn et al., 1999). They are inactivated by UV light in seawater (Liltved et al., 2006)
and are likely to be denatured by ionising radiation at 50 kGy in frozen fish (DAFF, 2013).
7.2 Risk assessment
7.2.1 Entry assessment
MABV strains are associated with fish bait species (Table 7), including herrings (Clupea sp.,
Sardinella spp. and Sardinops spp.) (Clupeidae) imported in large quantities during 2018 (Table
3). The prevalence of clinical disease is low in wild fish but may be higher adjacent to infected
marine farms (Wallace et al., 2008). Subclinically infected fish remain lifelong carriers and shed
high volumes of virus (Munro & Midtlyng, 2011). Pathogenicity varies with season, size and age
of the host (Isshiki et al., 2001; Munro & Midtlyng, 2011).
MABV strains are likely to remain viable in fish bait. As prevalence of subclinical disease in
wild fish is likely to be low, the likelihood of entry is assessed as low.
7.2.2 Exposure and establishment assessment
MABV strains are environmentally stable and can be spread by fomites, including commercial
processing, storage and transport equipment (Munro & Midtlyng, 2011). They are likely to
remain viable in commercial fish processing and bait storage facilities, in bait discarded to
landfill or through meltwater discharges (Munro & Midtlyng, 2011) for up to 24 weeks
(Desautels & MacKelvie, 1975). They may be dispersed from landfill to waterways and between
waterways by piscivorous birds (McAllister & Owens, 1992).
MABV strains remain viable in dead fish tissues, in moist packaging materials at high titre (>
106.0 pfu g-1) and in water for up to 8 weeks (Desautels & MacKelvie, 2011; Munro & Midtlyng,
2011). The infectious dose is low (Urquhart et al., 2008; Johansen et al., 2011) and all life stages
may be infected (DFO, 2017). Host specificity is low which could allow exposure to occur
through cross-contamination with discarded packaging in the supply chain.
The likelihood of exposure and establishment is assessed as medium.
7.2.3 Consequence assessment
Economic consequences
MABV strains affect over 30 families of marine fish (Blackwell 2019), including major farmed
and fished species in New Zealand. Wild stocks may act as reservoirs of infection (Raynard et
al., 2007; Wallace et al., 2008; Lafferty et al., 2015). Host specificity is low, while morbidity
and mortality may be high, reaching 100% in farmed fish. All life stages can be infected (DFO,
2017).
Infection with MABV is not an OIE-listed disease, so establishment would not result in
restrictions on trade access for New Zealand. It is likely that establishment would result in
lowered productivity and increased mortality, with serious economic consequences through lost
production in salmonid aquaculture, which was valued at $77 million in 2018 (Aquaculture New

48 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Zealand, 2019). Establishment is also likely to adversely affect other aquaculture species,
including yellowtail kingfish (Seriola lalandi) (NIWA, 2017a) and snapper (Sparus aurata)
(Plant & Food, 2016). Establishment would also have indirect economic effects on businesses
reliant on aquaculture and commercial wild fisheries sectors (MPI, 2018a, 2018b, 2018c).
Social consequences
MABV strains are not zoonotic (CABI, 2019a). They are reported from a wide range of species
including several major inshore recreational target species such as snapper (Pagrus auratus) and
trevally (Pseudocarynx dentax) (Wynn-Jones et al., 2014, 2019). Introduction is likely to reduce
both the productivity and relative abundance of these already stressed inshore fish stocks (MPI,
2018a, 2018b, 2018c), with significant negative social impacts for customary and recreational
fishers.
Recreational fishing is the fifth most popular recreational activity for adults, involving 700,000
marine fishing trips annually, with a contribution of NZ$ 638 million to the GDP (Gross
domestic product) in 2015 (Southwick et al., 2018). The snapper and trevally recreational
fisheries contributed NZ$ 402 million and NZ$ 71 million respectively to the GDP in 2015
(Southwick et al., 2018).
Environmental consequences
MABV strains affect fish and invertebrates including mussels (Mytilus galloprovincialis) and
pearl oysters (Pinctada spp.). While scallops (Pecten spp.) can also be experimentally infected,
the environmental significance of infections in bivalves is unknown (Mortensen et al., 1992;
Isshiki et al., 2001; Molloy et al., 2013). Inshore fish and invertebrate species are considered
likely to encounter and consume discarded fish bait. Clupeids which feed on phytoplankton,
zooplankton and detritus, provide a significant food source for other fish, seabirds and marine
mammals and constitute a major link between trophic levels in marine food webs (Dunn et al.,
2012).
While host specificity and infectious dose are low, little is known of the effects of MABV strains
on wild fish stocks (Lafferty et al., 2015). MABV strains may be dispersed from marine to fresh
waters by piscivorous birds (Johansen et al., 2011) and may cause population level epizootics
that could result in significant environmental effects (Whittington et al., 2008; Paul et al., 2001;
Fish & Game, 2014).
The introduction of MABV strains would have moderately adverse economic, social and
environmental consequences. The consequences of establishment are assessed to be medium.
7.2.4 Risk estimation
The entry, exposure and consequence assessments are low to medium, so the risk estimate for
MABV strains is assessed to be medium. Under the procedures followed in this risk analysis,
risk management measures may be justified.
7.3 Risk management
MABV strains have been assessed to be a risk in fish bait. Infection is not an OIE-notifiable
disease, so the Aquatic Code (OIE 2019a) provides no specific guidance or processing
requirements that would ensure the destruction of the virus.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 49
MABV strains are associated with fish bait (Table 7). Therefore, restriction to non-susceptible
species should substantially reduce the risk associated with MABV strains and be a viable risk
management option.
MABV strains have a wide geographical range (Table 7). Their occurrence in wild stocks in
other regions is likely to be low or negligible (Munro & Midtlyng, 2011). Restriction through the
BNZ Country Approval Procedures, to regions where these strains have not been reported should
substantially reduce the MABV risk and be a viable risk management option.
The presence of MABV strains in a shipment of fish bait may be verified by batch testing using
standard sampling procedures (Corsin et al., 2009; OIE, 2019a). Standard diagnostic procedures
for detection of these strains are provided in McColl et al. (2004b, 2009). Verification of a
negative test from a shipment should substantially reduce the risks associated with MABV
strains and be viable risk management options.
MABV strains are unaffected by freezing, so frozen storage (at -18°C for either 168 hours, or for
4 months), is not a risk management option.
MABV strains are denatured by ionising radiation at 50 kGy (DAFF, 2013). This irradiation
treatment should substantially reduce the MABV risk and be a viable risk management option.
7.3.1 Risk management options
In addition to the general risk management measures that are proposed for all imported fish bait,
one or more of the following risk management options would reduce the MABV specific risk to
an acceptable level:
Option 1
• Competent Authority attestation that the commodity does not include: Japanese jack
mackerel (Trachurus japonicus) (Carangidae); Atlantic herring (Clupea harengus), Bali
sardinella (Sardinella lemuru), Indian oil sardine (Sardinella longiceps), Pacific herring
(Clupea pallasii pallasii), round sardinella (Sardinella aurita), South American pilchard
(Sardinops sagax), spotted sardinella (Ambylygaster sirm) (Clupeidae); and Atlantic
mackerel (Scomber scombrus) (Scombridae) should substantially reduce the risks
associated with MABV strains, and the commodity could be imported without any further
restrictions.
Option 2
• Acceptance of region/country freedom through the BNZ Country Approval Procedures
should substantially reduce the MABV risk so the commodity could be imported without
any further restrictions.
Option 3
• A negative test result from a shipment of the commodity, following approved sampling and
diagnostic procedures through the BNZ Country Approvals Procedures should substantially
reduce the MABV risk, so the commodity could be imported without further restrictions.
Option 4
• Treatment with ionising radiation (at 50 kGy) should substantially reduce the MABV risk, so
the commodity could be imported without further restrictions.

50 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Iridoviridae: Megalocytivirus (red sea bream iridovirus - RSIV) and associated viruses
8.1 Technical review
8.1.1 Aetiological agent
The Genus Megalocytivirus is one of five genera classified within the family Iridoviridae (OIE,
2019a). Four main species of Megalocytivirus are recognised:
• Infectious spleen and kidney necrosis virus (ISKNV) (two genotypes);
• Red sea bream iridovirus (RSIV) (two genotypes);
• Turbot reddish body iridovirus (TRBIV);
• Three-spine stickleback iridovirus (TSIV).
These represent a species complex consisting of at least 3 genotypes, with closely related strains
that infect a wide range of marine and freshwater fish species (Nolan et al., 2015; Blackwell,
2019). While other uncharacterised strains, including scale drop disease virus of Asian seabass
(Lates calcarifer) (SDDV) are reported (Halaly et al., 2019), natural viral recombinations within
this species complex also occur (Kurita & Nakajima, 2012; Sriwanayos et al., 2013; Nolan et al.,
2015; Blackwell, 2019). For example, the RSIV-Ku strain isolated from red sea bream (Pagrus
major) from aquaculture in Chinese Taipei is a natural recombination of ISKNV and RSIV
genotypes (Shui et al., 2018).
8.1.2 OIE status
Infection with red sea bream iridovirus (RSIV) and infectious spleen and kidney iridovirus
(ISKNV) are recognised as agents of the OIE-listed red sea bream iridovirus disease (RSIVD)
(OIE, 2019a). Other strains within the species complex are not agents of OIE-listed diseases.
8.1.3 New Zealand status
RSIV and ISKNV have not been reported from New Zealand from active monitoring
programmes for RSIVD. New Zealand is considered free from RSIVD (OIE 2019a) and this is a
notifiable disease in New Zealand (Anon, 2016). RSIV and ISKNV are considered exotic to New
Zealand.
8.1.4 Zoonotic disease
RSIV and ISKNV strains are not considered zoonotic (OIE, 2019a).
8.1.5 Epidemiology
Distribution and host range
RSIV and ISKNV strains have a very wide host range from 20 families of marine and freshwater
fish (Gibson-Kueh et al., 2004; Song et al., 2008; Xu et al., 2010; Blackwell, 2019; OIE 2019a,
2019b).
The species associated with RSIV and ISKNV in fish bait and their geographic/pathogen range
are given in Table 8. Prevalence in wild fish stocks is poorly known. (Gibson-Kueh et al., 2004;
Song et al., 2008; OIE, 2019b).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 51
Table 8. Fish bait species susceptible to red sea bream iridovirus (RSIV), infectious spleen and kidney necrosis virus (ISKNV) and associated iridoviruses
Family Host Species Host geographical range1 Pathogen range2
Carangidae Japanese jack mackerel (Trachurus japonicus)
Northwest Pacific, Japan, Korea, Southeast Asia
Northwest, Central Pacific
Clupeidae Pacific herring (Clupea pallasi pallasii)
Arctic, Northwest, Northeast Central Pacific
Northeast, Central Pacific
Mugilidae Flathead grey mullet (Mugil cephalus)
Cosmopolitan in tropical, subtropical and temperate waters
Northwest, Central Pacific
Scombridae Chub mackerel (Scomber japonicus)
Indo-Pacific, Eastern, Western, Central Pacific Southeast Atlantic (South Africa)
Northwest, Central Pacific
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature). The Northwest and Central Pacific includes Australia, China, Chinese Taipei,
Hong Kong, South Korea, Malaysia, the Philippines, Singapore and Thailand.
Potential hosts in New Zealand
A wide range of marine and freshwater fish are considered susceptible to RSIV and ISKNV
strains. These include snapper (Pagrus auratus) (Sparidae), silver trevally (Pseudocaranx
dentex), trevally (Caranx georgianus) and jack mackerel (Trachurus spp.) (Carangidae). ISKNV
also occurs in freshwaters, where susceptible species include galaxiids (Galaxias spp.) and
cyprinids such as grass carp (Ctenopharyngodon idella) (Diggles, 2003; Blackwell, 2019).
Pathogenicity
All host life stages are susceptible, but juveniles are more susceptible than adults (OIE, 2019a).
Pathogenicity varies among and between viral clades and strains. While pathogenicity ranging
from 50% to 100% occurs in farmed red sea bream (S. auratus) (Sparidae) (Sano et al., 2011),
strains affecting wild fish species are of lower pathogenicity (OIE, 2019a). Pathogenicity is
temperature related, with higher mortalities generally associated with temperatures over 25 °C
(Jun et al., 2009).
The infectious dose (determined experimentally by intra-peritoneal (i.p.) injection of S. auratus
as 103 TCID50 -1) resulted in mortalities from 20% to 60% (Shinmoto et al., 2009; Kurita &
Nakajima, 2012). The lethal dose (105 TCID50 -1) caused 100% mortality, 7–10 days post
infection. Infection was associated with liver, kidney, spleen, heart and muscle tissues (Shinmoto
et al., 2009). Experimental infection (by bath inoculation) indicated subclinical infection was
widespread in surviving carrier fish that remained carriers of disease (Ito et al., 2013). RSIV and
ISKNV strains are not zoonotic.
Clinical signs
Clinical signs of infection vary widely, and infection may progress with no external signs (Noga,
2010; Kurita & Nakajima, 2012; OIE, 2019a).
Pathogen survival
Experimental data indicate megalocytivirus strains survive in water for at least 7 days at 15 °C
(Ito et al., 2013) and for longer periods at 4 °C. They are unaffected by frozen storage in fish
tissues (to -80 °C) and remain viable after repeated freeze-thaw cycles (OIE, 2019a, 2019b).

52 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
While viability in natural waters is unknown (see section 6.1.2), RSIV strains remained viable in
frozen baitfish used as feed in Asian aquaculture (Laijimin et al., 2015).
Transmission
Disease transmission is horizontal, through the water column (OIE, 2019a).
Infection and disease progression
Infection occurs by direct contact with viral particles, or through consumption of infected fish
(Sano et al., 2011; OIE, 2019a). Disease progression is initially focused on the viscera (including
the heart). Infection then typically spreads to the gills, skeletal muscle, connective tissue and
spleen (Whittington et al., 2010; OIE, 2019a, Ito et al., 2013).
Infection with ISKNV strains may also be subclinical (Hine & Diggles, 2005), where overt
disease may occur when the host is stressed due to adverse environmental conditions such as
temperature or salinity (Mohr et al., 2015)
Inactivation
Megalocytiviruses are unaffected by short (16 hours) or medium-term frozen storage (to at least
4 months at -20 °C) (OIE, 2019a). They are likely to be inactivated by ionising radiation of
frozen fish bait (at 50 kGy) (DAFF, 2013).
8.2 Risk assessment
8.2.1 Entry assessment
RSIV and ISKNV strains are associated with fish bait species (Table 8), including Pacific
herring (Clupea pallasii pallasii) (Clupeidae) imported from the Asia-Pacific region in high
volumes during 2018 (Table 3). Prevalence in wild fish is unknown, but assumed to be low-
moderate, as these viruses remain viable in frozen fish bait (Laijimin et al., 2015). Surviving fish
may be carriers with no external signs of infection (Ito et al., 2013).
As the volume of susceptible fish bait derived from the Asia-Pacific region is high, the risk of
entry of RSIV and ISKNV strains is assessed as low-medium.
8.2.2 Exposure and establishment assessment
RSIV and ISKNV strains are unaffected by short or medium-term frozen storage and multiple
freeze-thaw cycles. They remain viable in seawater for up to 7 days at 15 °C (Ito et al. 2013) and
for longer periods at 4 °C (OIE 2019a). All life stages of the host are susceptible (OIE 2019a).
The infectious dose (determined experimentally) in red sea bream (S. auratus) is low (103
TCID50 -1), with mortalities varying from 20% to 60% (Kurita & Nakajima, 2012; Shinmoto et
al., 2009).
Pathogenicity in wild stocks is unknown, but these pathogens may be spread through fish bait,
where used as aquaculture feed (Sano et al., 2011; Laijimin et al., 2015; OIE, 2019a). A wide
variety of potential host species are present in New Zealand waters.
RSIV and ISKNV strains are environmentally persistent. The infectious dose is assumed to be
low and infection can be initiated through fish bait. The likelihood of exposure and establishment
is therefore assessed to be medium.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 53
8.2.3 Consequence assessment
Economic consequences
Infection with RSIV, ISKNV are OIE-listed diseases (OIE, 2019a), so their establishment would
result in loss of trade access for New Zealand. They have moderate-high morbidity and mortality
(Nakajima et al., 1998; Van Beurden et al., 2012). Their establishment could result in lower
productivity and increased mortality in over 20 families of marine and fresh water fish
(Blackwell, 2019), including snapper (Pagrus auratus) (exports valued at $33 million) and jack
mackerel (Trachurus spp.) (exports valued at $68 million) in 2018 (Aquaculture New Zealand,
2019).
Productivity losses and increased mortality would also negatively impact the developing
aquaculture of yellowtail kingfish (Seriola lalandi) and grouper (Polyprion oxygeneios)
(Diggles, 2003; Symonds et al., 2014).
Establishment of freshwater strains may also adversely affect grass carp (Ctenopharyngodon
idella) and silver carp (Hypophthalmichthys molitrix) aquaculture, used for weed control
(Clayton & Wells, 1999; Blackwell, 2019).
Social consequences
RSIV and ISKNV strains are not considered zoonotic. They have a wide host range (Hine &
Diggles, 2005; Blackwell, 2019), so establishment may negatively affect target species of
importance for customary and recreational fishing (Wynn-Jones et al., 2014, 2019). Recreational
fishing is the fifth most popular recreational activity for adults, involving 700,000 marine fishing
trips annually, with a contribution of NZ$ 638 million to the GDP (Gross domestic product) in
2015 (Southwick et al., 2018).
The snapper recreational target fishery contributed NZ$ 492 million to the GDP (Gross domestic
product) in 2015 (Southwick et al., 2018) and is a taonga for Maori culture (MPI, 2018c). In
fresh water, ISKNV infects galaxiids (Galaxias spp.), which forms a major part of customary
and recreational fisheries for whitebait (NIWA, 2019).
Many of these marine and freshwater fish stocks are considered stressed (MPI, 2018a, 2018b,
2018c). A reduction in productivity and increase in mortality due to the establishment of RSIV
and ISKNV strains would have significant social impacts for New Zealand.
Environmental consequences
RSIV and ISKNV strains are reported from over 20 fish families and 60 species of fish in marine
and fresh waters (Xu et al., 2010; OIE, 2019a, 2019b; Blackwell,2019). Inshore species
including clupeid, mugilid and scombroid fish are considered likely to encounter and consume
discarded fish bait. Clupeids feed on phytoplankton, zooplankton and detritus and represent a
major link between trophic levels in food webs (Varpe et al., 2005; DFO, 2009; Dunn et al.,
2012). Clupeid and mugilid species provide a significant food source for other fish, seabirds and
marine mammals (Dunn et al., 2012).
Reductions in Pacific herring (C. p. pallasii) abundance (from over-fishing) caused a reported
60% decrease in the relative abundance of wild salmon (Oncorhynchus tshawytscha) in coastal
British Columbia, with major biological, economic and social consequences for humans and wildlife (Pagowski et al., 2019). The wide host range of these pathogens may potentially cause
population level epizootics in wild fisheries with significant environmental effects on the New

54 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Zealand coastal environment (Whittington et al., 2008; Paul et al., 2001; Fish & Game, 2014;
Lafferty et al., 2015).
ISKNV strains may be dispersed into fresh waters by infected euryhaline fish species (Go &
Whittington, 2006; Go, 2015), or through the use and discarding of bait in fresh water
(Blackwell, 2013). If ISKNV strains established in wild cyprinids (such as goldfish, Carassius
auratus), these could act as reservoir hosts for infection of native galaxiids (Galaxias spp.)
(Galaxiidae) (Clayton & Wells, 1999; Blackwell, 2019), as well as threatened aquatic
amphibians (Hine & Diggles, 2005).
Infection may also become endemic, with overt infection occurring when the host is
environmentally stressed (Mohr et al., 2015). Hine & Diggles (2005) assessed the environmental
consequences of establishment of RSIV and ISKNV strains (through introduced ornamental fish)
as high to catastrophic.
The introduction of RSIV and ISKNV strains would have significant negative economic, social
and environmental consequences. The consequences of establishment are assessed to be high.
8.2.4 Risk estimation
The entry, exposure, establishment, and consequence assessments for RSIV and ISKNV strains
are medium to high, so the risk estimation is assessed to be medium-high. Under the procedures
followed in this risk analysis, risk management measures may be justified.
8.3 Risk management
RSIV and ISKNV strains have been assessed to be a risk in fish bait. Infection with these strains
is OIE-listed (OIE, 2019a), so the OIE Code provides some guidance on control measures
necessary for inactivation. However, the OIE-listed options require heat treatment which would
render the fish bait unfit for purpose. These are not considered further.
RSIV and ISKNV strains are associated with some fish bait species (Table 8), while other fish
bait species are not associated with RSIV and ISKNV. Therefore, restriction of the commodity to
species not associated with these strains should substantially reduce the risk and be a viable risk
management option.
RSIV and ISKNV strains have a wide geographical range (Table 8). Their occurrence in wild
stocks in other regions is likely to be low or negligible (OIE, 2019a). Restriction, through the
BNZ Country Approval Procedures, to regions where these strains have not been reported should
substantially reduce the risk and be a viable risk management option.
The presence of RSIV and ISKNV strains in a shipment of fish bait may be verified by batch
testing using standard sampling procedures (Corsin et al., 2009; OIE, 2019a). Standard
diagnostic procedures for detection of these strains are provided in McColl et al. (2004b, 2009).
Verification of a negative test for RSIV and ISKNV from a shipment should substantially reduce
the risk and be a viable risk management option.
RSIV and ISKNV are unaffected by frozen storage (OIE, 2019b). Therefore, frozen storage
(at -18°C) for either 168 hours or 4 months is not a risk management option.
RSIV and ISKNV strains are denatured by exposure to ionising radiation (to 50 kGy) (DAFF,
2013). This irradiation treatment should substantially reduce the risk and be a viable risk
management option.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 55
8.3.1 Risk management options
In addition to the general risk management measures that are proposed for all imported fish bait
(section 6.2.1), one, or more of the following risk management options would reduce the RSIV
and ISKNV specific risk to an acceptable level:
Option 1
• Competent Authority attestation that the commodity does not include: Japanese mackerel
(Trachurus japonicus) (Carangidae); Pacific herring (Clupea pallasii pallasii) (Clupeidae);
flathead grey mullet (Mugil cephalus) (Muglidae); and chub mackerel (Scomber japonicus)
(Scombridae) should substantially reduce the RSIV and ISKNV risk, so the commodity
could be imported without any further restrictions.
Option 2
• Acceptance of region/country freedom through the BNZ Country Approval Procedures,
should substantially reduce the RSIV and ISKNV risk, so the commodity could be
imported without any further restrictions.
Option 3
• A negative test result from a shipment of the commodity, following sampling and diagnostic
procedures approved through the BNZ Country Approvals Procedures should substantially
reduce the RSIV and ISKNV risk, so the commodity could be imported without further
restrictions.
Option 4
• Treatment with ionising radiation (to 50 kGy) should substantially reduce the RSIV and
ISKNV risk, so the commodity could be imported without any further restrictions.
Iridoviridae: Erythrocytic necrosis virus (ENV)
9.1 Technical review
9.1.1 Aetiology
Erythrocytic necrosis virus (ENV) is the agent of viral erythrocytic necrosis disease (VEN). It is
a double-stranded DNA virus, classified within family Iridoviridae, originally isolated from
salmonids and herring (Clupea spp.) (Walker & Sherburne, 1977; Winton & Hershberger, 2014).
It may represent a new species within the Iridoviridae (Emmenegger et al., 2014; Hick et al.,
2016; Purcell et al., 2016).
Several ENV strains exist among host species and within geographical ranges, but their genetic
relationships are unknown (Winton & Hershberger, 2014; Hick et al., 2016). Viral isolates from
Chinook salmon (O. tshawytscha) and Pacific herring (Clupea pallasii pallasii) were over 99%
similar, suggesting that cross-species infection occurs (Hershberger et al., 2006, 2009; Winton &
Hershberger, 2014; Pagowski et al., 2019).
9.1.2 OIE status
Infection with VEN is not an OIE-listed disease (OIE, 2019a).

56 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
9.1.3 New Zealand status
ENV has not been reported from New Zealand (Anderson, 1990; Diggles, 2011). Infection with
ENV is not a notifiable disease in New Zealand (Anon., 2016).
9.1.4 Zoonotic disease
ENV is not zoonotic (Blackwell, 2019).
9.1.5 Epidemiology
Distribution and host range
ENV is reported from over 10 families and 20 species of elasmobranch and teleost marine fish
(MacMillan et al., 1980; Dannevig & Thorud, 1999; Davies et al., 2009; Emmenegger et al.,
2014; Winton & Hershberger, 2014; Hick et al., 2016; Blackwell, 2019). It is reported from wild
and farmed fish (Hick et al., 2016). The species associated with ENV in fish bait and their
geographic/pathogen range are given in Table 9 (Nicholson & Reno, 1981; Hershberger et al.,
2011; Emmenegger et al., 2014).
ENV occurs in wild Pacific herring (C. p. pallasii) at a prevalence of 27%, reaching 67% in
epizootics (Reno et al., 1985). Prevalence in Californian anchovy (E. mordax) is 38%, reaching
50% in epizootics (Hershberger et al., 2009, 2011; Pagowski et al., 2019).
Table 9. Fish bait species susceptible to erythrocytic necrosis virus (ENV)
Family Host species Host geographical range1 Pathogen range2
Clupeidae Atlantic herring (Clupea harengus)
Baltic Sea, Northeast, Northwest Atlantic
Baltic Sea. Northeast Atlantic, (Europe, Spain, United Kingdom)
Clupeidae Pacific herring (C. pallasii pallasii)
Arctic, Northwest, Northeast Pacific (Alaska to Mexico)
Northwest, Northeast Pacific (Japan)
Engraulidae Californian anchovy (Engraulis mordax)
Northeast Pacific (Canada to Mexico)
Northeast Pacific (USA, Canada)
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature). ENV is reported from the North Atlantic and North Pacific Oceans, as well as
from Spain, Japan and the United Kingdom. It is not reported from the Southern Hemisphere (Hick et al., 2016).
Potential hosts in New Zealand
ENV may potentially affect a range of inshore fish in New Zealand, including dogfish
(Triakidae) (Tubbs et al., 2007), as well as farmed and wild salmonids.
Pathogenicity
ENV is unculturable, so relationships between virus presence and clinical disease are difficult to
determine experimentally (Emmenegger et al., 2014). It may remain dormant where water
temperatures are lower than 20 °C (Pagowski et al., 2019). Pathogenicity varies among ENV
strains, host species, route of infection and intensity of exposure (Reno et al., 1985; Winton &
Hershberger, 2014). All life stages of the host may be infected, and prevalence may be inversely
correlated to host age (Smail, 1982).
Transmission and infection details are uncertain (Winton & Hershberger, 2014; Pagowski et al.,
2019). ENV inclusion bodies are present in erythrocytes collected from kidney and spleen tissues

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 57
(Glenn et al., 2012) and infection can be experimentally induced by filtered preparations from
diseased fish (Evelyn & Traxler, 1978).
Clinical signs
Infection usually occurs without external clinical signs, but fish with acute infection (VEN) may
show signs of anaemia (Reno et al., 1985; Winton & Hershberger, 2014).
Pathogen survival
Iridoviruses are unaffected by frozen storage (OIE, 2019b). From experimental data, ENV
remains viable in erythrocytes in solution up to 2 weeks (Nicholson & Reno, 1981). However,
viability may be less in natural waters (see section 6.1.2).
Transmission
Disease transmission is mainly horizontal, but vertical transmission is also reported (Rohovec &
Amandi, 1981).
Infection and disease progression
ENV affects red blood cells, causing long-term systemic anaemia. This may eventually affect up
to 90% of erythrocytes in the host (Reno et al., 1985). Acute disease (VEN) is often associated
with co-infection by opportunistic pathogens (Hershberger et al., 2006; Glen et al., 2012).
Infection occurs through the water column, where the initial prevalence is likely to be low in
wild fish stocks (Hershberger et al., 2006). From a multi-year field study on wild Pacific herring
(C. pallasii pallasii), population-level VEN epizootics were an important component of natural
mortality (Hershberger et al., 2009; Pagowski et al., 2019).
Subclinical infection of iridoviruses rarely causes high morbidity or mortality in wild fish (Reno
et al. 1985). However, co-infection may occur with other viruses (Hershberger et al., 2006) and
overt disease may then develop where the host is stressed by environmental stressors, such as
low dissolved oxygen and low salinity (Meyers et al., 1986; Winton & Hershberger, 2014; Mohr
et al., 2015).
Inactivation
Iridoviruses including ENV are unaffected by either short (168 hours) or medium-term (4
months) frozen storage (at -20 °C) (OIE, 2019b). ENV is likely to be denatured by exposure to
ionising radiation (to 50 kGy) (DAFF, 2013).
9.2 Risk assessment
9.2.1 Entry assessment
ENV is reported from listed fish bait species from a wide geographical area (Table 9). Viral
prevalence ranges from 27% to 38% in wild bait fish, reaching 67% during epizootics
(Hershberger et al., 2009; Pagowski et al., 2019). As viral shedding rates increase when herrings
are stressed, viral particles may also be concentrated in the water used to freeze baitfish (Hick et
al., 2016). ENV is likely to remain viable in frozen fish bait (OIE 2019b).
ENV is associated with fish bait across a wide geographical area, at a moderate-high prevalence
and may also be present in meltwater. The likelihood of entry is therefore assessed to be high.

58 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
9.2.2 Exposure and establishment assessment
The transmission mechanisms of ENV are poorly known. Experimental data indicate iridoviruses
remain viable in seawater for up to 7 days at 15 °C (Ito et al., 2013) and for longer periods at
4 °C (Anon., 2018). However, the length of time that ENV may remain infective under natural
conditions is unknown (see section 6.1.2).
All life stages of the host may be infected (Smail, 1982). Infection of farmed salmonids,
including Chinook salmon (Oncorhynchus tshawytscha), through wild fish bait species is
reported, suggesting the infectious dose of ENV is likely to be low (Hershberger et al., 2006,
2009; Sano et al., 2011; Winton & Hershberger, 2014; OIE, 2019a; Pagowski et al., 2019).
A wide variety of potential hosts are present in New Zealand waters. ENV remains viable in
seawater and is environmentally resistant. Infection can be initiated in aquaculture species when
fish bait is used as feed. Therefore, the likelihood of exposure and establishment is assessed as
medium.
9.2.3 Consequence assessment
Economic consequences
Infection with VEN is not an OIE-listed disease, so establishment would not affect New
Zealand’s trade status. Subclinical infection commonly occurs in wild fish stocks, but rarely
causes high morbidity or mortality (Reno et al., 1985). However, ENV is the agent of viral
erythrocytic necrosis (VEN) in salmonids farmed in marine waters. ENV may be associated with
high mortality and morbidity (Hershberger et al., 2009; Winton & Hershberger, 2014; Pagowski
et al., 2019). It would likely have a significant effect on New Zealand marine aquaculture, which
is focussed on Chinook salmon (O. tshawytscha), valued at $77 million in export earnings in
2018 (Hick et al., 2016; Seafood New Zealand, 2019).
ENV has a wide host range, affecting over 10 families of elasmobranch and teleost marine fish
(Blackwell, 2019). Establishment would also have a major economic effect on the developing
aquaculture of yellowtail kingfish (Seriola lalandi) (Carangidae) (Diggles, 2003; Symonds et
al.,2014).
The effects of introduction of viral pathogens on wild fish stocks are unknown (Lafferty et al.,
2015), but are likely to be low (Reno et al., 1985; Kahn et al., 1999). ENV may affect
commercial inshore fisheries including jack mackerel (Trachurus spp.), valued at $68 million in
2018 (Seafood New Zealand, 2019).
Social consequences
ENV is not considered zoonotic. Establishment would have significant social implications for the
recreational hatchery-based fisheries for rainbow trout (O. mykiss) and brown trout (Salmo
trutta) (Marsh & Mkwra, 2013; Fish & Game, 2014), due to lost productivity and increased
mortality. The Lake Taupo trout fishery alone was valued in 2012 at NZ$ 70–80 million (Marsh
& Mkwra, 2013).
Recreational marine fishing is the fifth most popular recreational activity for adults, involving
700,000 marine fishing trips in 2015, with a contribution of NZ$ 638 million to the GDP (Gross
domestic product). Recreational yellowtail kingfish fishing contributed NZ$ 79.3 million to the
New Zealand GDP in 2015 (Southwick et al., 2018).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 59
Environmental consequences
ENV is reported from over 20 species of marine and anadromous fish (Hine & Diggles, 2005;
Blackwell, 2019). Little is known about the potential effects of virus infection of wild fish stocks
(Lafferty et al., 2015), but subclinical infection is rarely associated with high morbidity and
mortality (Reno et al., 1985; Kahn et al., 1999). However, inshore fish species including clupeid
and engraulid fish are considered likely to encounter and consume discarded fish bait. Clinical
infection in North American clupeid and engraulid fisheries has been associated with population
level epizootics (Hick et al., 2016). Reductions in herring abundance (from over-fishing) caused
a reported 60% decrease in the relative abundance of wild salmon (O. tshawytscha) in coastal
British Columbia. This resulted in major biological, economic and social consequences for
humans and wildlife (Pagowski et al., 2019). Pacific herring (C. p. pallasi) is a keystone species
in marine food webs (Varpe et al., 2005; DFO, 2009), while clupeid and engraulid species
represent significant feed for other marine organisms, including fish, seabirds and marine
mammals (Dunn et al., 2012). The introduction of agents capable of causing population level
epizootics would have significant environmental effects on New Zealand coastal fisheries
(Whittington et al., 2008; Paul et al., 2001).
The introduction of ENV would have significant negative economic, social and environmental
consequences. The consequences of establishment are assessed to be high.
9.2.4 Risk estimation
The entry, exposure, establishment, and consequence assessments are medium-high, so the risk
estimation for ENV is medium-high. Under the procedures followed in this risk analysis, risk
management measures may be justified.
9.3 Risk management
ENV has been assessed to be a risk in fish bait. Infection with VEN is a not an OIE-notifiable
disease, so the OIE Aquatic Code (OIE, 2019a) provides no specific guidance or processing
requirements that would ensure the destruction of the virus.
ENV is associated with fish bait (Table 9). Therefore, restriction to non-susceptible species in
the commodity should substantially reduce the ENV risk and be a viable risk management
option. ENV has a wide geographic range (Table 9). The occurrence is likely to be low in other
geographic regions. Restriction through the BNZ Country Approval Procedures to regions where
ENV has not been reported should substantially reduce the ENV risk and be a viable risk
management option.
The presence of ENV strains in a shipment of fish bait may be verified by batch testing using
standard sampling procedures (Corsin et al., 2009; OIE, 2019a). Standard diagnostic procedures
for detection of the virus are provided in Emmenegger et al. (2014) and Pagowski et al. (2019).
Verification of a negative test from a shipment should substantially reduce the ENV risk and be a
viable risk management option.
ENV is unaffected by freezing (OIE, 2019b) so frozen storage (at -18°C for either 168 hours, or
for 4 months) is not a viable risk management option. ENV is denatured by ionising radiation at
50 kGy (DAFF 2013). This irradiation treatment should substantially reduce the ENV risk and be
a viable risk management option.

60 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
9.3.1 Risk management options
In addition to the general risk management measures that are proposed for all imported fish bait
(section 6.2.1), one or more of the following risk management options would reduce the ENV
specific risk to an acceptable level:
Option 1
• Competent Authority attestation that the commodity does not include European herring
(Clupea harengus), Pacific herring (Clupea pallasii pallasii) (Clupeidae) and European
anchovy (Engraulis mordax) (Engraulidae), should substantially reduce the ENV risk, so
the commodity could be imported without any further restrictions.
Option 2
• Acceptance of country/zone freedom through the BNZ Country Approval Procedures
should substantially reduce the ENV risk so the commodity could be imported without any
further restrictions.
Option 3
• A negative test result from a shipment of the commodity, following sampling and diagnostic
procedures through the BNZ Country Approvals Procedures should substantially reduce the
ENV risk, so the commodity could be imported without further restrictions.
Option 4
• Treatment with ionising radiation (to 50 kGy) should substantially reduce the ENV risk, so
the commodity could be imported without any further restrictions.
Nodaviridae: Nervous necrosis virus (NNV)
10.1 Technical review
10.1.1 Aetiology
Nodaviruses are non-enveloped positive-stranded RNA viruses, classified as a Betanodavirus.
The type specimen is nervous necrosis virus (NNV). Four main genotypes are recognised by the
OIE (OIE, 2019b): SJNNV-type (striped jack nervous necrosis virus), TPNNV-type (tiger puffer
nervous necrosis virus), BFNNV-type (barfin flounder nervous necrosis virus) and RGNNV-type
(red-spotted grouper nervous necrosis virus) (OIE 2019b). These genotypes broadly correlate
with serotype A (SJNNV), serotype B (TPNNV) and serotype C (BFNNV and RGNNV) (Mori
et al., 2003; OIE, 2019b).
Several other unclassified serotypes have been proposed (Castric, 1997; Johansen et al., 2004;
Kim et al., 2011; Doan et al., 2017; Prado-Alvarez & Garcia-Fernandex, 2019; Bandin & Souto,
2020), but these are not currently recognised by the OIE (OIE, 2019a).
Serotypes are grouped by their optimum growth temperature in vitro (Iwamoto et al., 2000).
Serotype A is infective over a temperature range from 20 °C to 30 °C. Serotype B only affects
tiger puffer (Takifugu rubripes) and is optimal at 20 °C. Serotype C affects fish over a wide
temperature range, including cold water (from 15 °C to 20 °C) and warm water fish (from 25 °C
to 30 °C) (OIE, 2019b). Within each serotype, host specificity is low, as serotypes of one

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 61
genotype cause disease in host species that are predominantly infected by other genotypes
(Munday et al., 2002; Tanaka et al., 2003; Sano et al., 2011).
10.1.2 OIE status
Infection with NNV or other nodaviruses is described by the OIE as viral encephalopathy and
retinopathy (VER) (OIE, 2019b). It is not an OIE-listed disease (OIE, 2019a).
10.1.3 New Zealand status
No genotype of NNV have been reported from New Zealand (Hine & Diggles, 2005; Tubbs et
al., 2007; Moody & Horwood, 2008). Infection with NNV is not a notifiable disease in New
Zealand (Anon. 2016) but it is considered exotic (Tubbs et al., 2007).
10.1.4 Zoonotic disease
Betanodaviruses including NNV are not considered zoonotic (Adachi et al., 2008; OIE, 2019a).
10.1.5 Epidemiology
NNV causes viral nervous necrosis disease (VNN) or viral encephalopathy and retinopathy
(VER) (OIE, 2019b).
Distribution and host range
NNV strains are widely distributed (Table 10). They occur in 24 families of fish and coleoid
molluscs in marine and fresh waters. They are also reported from bivalve molluscs (blue mussel,
Mytilus galloprovincialis), crustaceans (blue swimming crab, Portunus pelagicus) and annelid
worms (Nereis spp.) adjacent to infected fish farms (Fichi et al., 2015; Fiorito et al., 2015; Costa
& Thompson, 2016; Doan et al., 2017; Volpe et al., 2018; OIE, 2019b; Blackwell, 2019). The
international trade in nereid worms for fish food may represent an infection pathway (OIE
2019b). NNV strains may also be dispersed by piscivorous birds (OIE 2019b).
NNV occurs in wild Mediterranean fish species at a prevalence of 21%, where the absence of
infected clupeids may reflect their higher susceptibility and mortality (Berzak et al., 2019). NNV
occurs at a prevalence up to 100% in wild mackerel (Scomber spp.) in the Philippines (de la Pena
et al., 2011). NNV strains are a major pathogen of larval and juvenile fish in Chinese, Asian,
Australian and European aquaculture (Castric, 1997; Chi et al., 2016).
The species associated with NNV strains in fish bait and their geographic/pathogen range are
given in Table 10 (Mori et al., 1991, 1992; Nguyen et al., 1996; Ucko et al., 2004; Gomez et al.,
2006; Sakamoto et al., 2008; Tubbs et al., 2007; Gomez et al., 2010; Crane & Hyatt, 2011;
Forrest et al., 2011; Panzarin et al., 2012; Fiorito et al., 2015; Zorriehzahra et al., 2016; OIE,
2019b; Bandin & Sousa, 2020).
Potential host species in New Zealand
Potential species at risk in New Zealand include eels (Anguilla spp.), flatfish (Pleuronectidae),
jack mackerel (Trachurus spp.), snapper (Pagrus auratus), striped trumpeter (Latris lineata),
trevally (Caranx georgianus) and yellowtail kingfish (Seriola lalandi) (Blackwell, 2019).
Pathogenicity
NNV causes economically significant neurological disease (VNN) in a wide range of wild and
farmed fish species (Munday et al., 2002; Sano et al., 2011). Mortalities may reach 100% in

62 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
aquaculture, particularly for larval and juvenile fish (OIE, 2019b). Infection in older, larger fish
is often subclinical, where surviving fish may represent reservoirs of disease (Sano et al., 2011;
Chi et al., 2016).
Mortality of farmed sea bream (S. auratus) infected with NNV may reach 50% in marine
aquaculture, with no clinical signs (Cherif et al., 2008).
Experimental infection (by injection) of sevenband grouper (Epinephelus septemfasciatus) at a
rate of 105.5 TCID50 fish-1 resulted in cumulative mortalities up to 44%. Viral titres between 109.5
and 1010.0 TCID50 g-1 were recovered from brain tissues. Viral titre in the tank seawater reached
105 TCID50 L-1. All surviving fish had viral titre in brain tissues of 104 to 107 TCID50 g
-1 (Nishi et
al. 2016). While these data indicate high mortality and high viral shedding rates in surviving
fish, little is known about infectivity in wild fish populations (Lafferty et al., 2015).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 63
Table 10. Fish and coleoid cephalopod mollusc species susceptible to nervous necrosis virus (NNV)
Family Host species Host geographical range1 Pathogen range2
Fish families
Carangidae Atlantic horse mackerel (Trachurus trachurus), Chilean jack mackerel (T. murphi)
Mediterranean, Eastern, Southwest Atlantic (Argentina), Pacific
Mediterranean, Indo-Pacific, Pacific, Northeast, Southwest Atlantic
Japanese jack mackerel (T. japonicus), Pacific jack mackerel (T. symmetricus)
Eastern, Central, South Pacific, Northwest Pacific, Japan, Southeast Asia
Indo-Pacific, North Pacific
Clupeidae Atlantic herring (Clupea harengus)
Northeast, Northwest Atlantic, North Sea, Baltic Sea, Black Sea
Northeast, Northwest Atlantic, Mediterranean
Mugilidae Flathead grey mullet (Mugil cephalus)
Cosmopolitan in coastal tropical, subtropical and temperate waters (Eastern and Western Atlantic and Pacific, Indo-Pacific, Black Sea, Mediterranean)
Indo-Pacific, North Pacific, Northeast, Northwest Atlantic Mediterranean
Thinlip grey mullet (C. ramada)
Indo-West Pacific, East, Southwest Pacific
Indo-Pacific
Golden grey mullet (Chelon (Mugil) auratus)
Eastern Atlantic, Black Sea, Mediterranean
Northeast Atlantic, Mediterranean
Thicklip grey mullet (Chelon (Mugil) labrosus)
Eastern Atlantic, Black Sea, Mediterranean
Northeast Atlantic, Mediterranean
Scombridae Atlantic mackerel (Scomber scombrus)
Eastern Atlantic, Mediterranean
Northeast Atlantic (Norway), Mediterranean (Greece)
Blue mackerel (S. australiasicus)
Indian Ocean, Indo-Pacific, Pacific
Northwest Pacific
Chub mackerel (Scomber japonicus)
Indo-Pacific, Eastern, Western Pacific
North Pacific (Japan)
Coleoid cephalopod mollusc families
Ommastrephidae Japanese flying squid (Todarodes pacificus (=Ommastrephes pacificus)
Indo-Pacific, North Pacific North Pacific
Octopodidae Common Octopus (Octopus vulgaris)
Europe, Mediterranean Mediterranean
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature). NNV strains are reported from the Indo-Pacific (Australia, China, India, and Southeast Asia), Northeast and Northwest Atlantic, Europe and North America. They have not been reported from South Africa, South America or New Zealand (Hine & Diggles, 2005; Moody & Horwood, 2008; Sano et al., 2011).
Clinical signs
Infection with NNV causes a neurological disease which usually progresses with no external
clinical signs (Sano et al., 2011; Blackwell, 2019).
Pathogen survival
Serotype A NNV can be transmitted in dead and decomposing fish tissues (Chi et al., 2016) and
through fomites in aquaculture (OIE, 2019b). From laboratory experiments using artificial and
sterilised seawater, NNV remains viable in seawater (below 25 °C) for at least 12 months

64 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
(Frerichs et al., 2000). Survival is assumed to be at least 6 months and possibly longer (OIE,
2019b), but may be influenced in natural waters by dilution and predation factors (see section
6.1.2).
Transmission
Disease transmission may be vertical and horizontal, but life cycle details are poorly known
(Sano et al., 2011; OIE, 2019b). Wild fish may be reservoirs for infection. Farmed gilthead sea
bream (Sparus aurata) in Israeli aquaculture were infected at a prevalence of 20%, where the
infection was derived from adjacent wild fish stocks in the Mediterranean Sea (Berzak et al.,
2019).
Infection and disease progression
Infection usually occurs through the water column. Disease progression may initially follow
several routes (Costa & Thompson, 2016). In clinical infection, NNV is neuro-invasive, affecting
the spinal cord, swim bladder, then progressing to the brain and retinal tissues. Viral particles are
released into the water column by carrier fish through gonad and digestive excretions (Sano et
al., 2011).
NNV infection is reported in aquaculture through use of infected fish bait of Family Clupeidae
and Japanese flying squid (Todarodes pacificus) (Family Ommastrephidae) as feed (Mori et al.,
2005; Gomez et al., 2010; OIE, 2019b).
Inactivation
No simple control methods are available (Doan et al., 2017). Nodaviruses are unaffected by
either short (168 hours), or medium-term (4 months) freezing (at -20 °C) (OIE, 2019b). NNV
strains are likely to be denatured by ionising radiation (at 50 kGy) (DAFF, 2013).
10.2 Risk assessment
10.2.1 Entry assessment
NNV has a wide geographical distribution and a wide host range (Table 10) (Moody &
Horwood, 2008; Sano et al., 2011). Host specificity is low (Munday et al. 2002) and each NNV
genotype commonly affects several species within a geographical region (Shetty et al., 2012).
Prevalence is low-moderate (up to 21%) in wild fish and mollusc stocks (Castric, 1997; Chi et
al., 2016; Berzak et al., 2019), including fish bait species imported in high volumes (Tables 1 &
2).
NNV strains remain viable in frozen fish. Experimental data suggest survival in marine or fresh
waters may be up to 6 months (Frerichs et al., 2000; OIE, 2019b). Survival in natural waters is
unknown (see section 6.1.2). Therefore, the likelihood of entry is assessed as medium.
10.2.2 Exposure assessment
NNV strains remain viable during frozen storage, transportation and processing (Sano et al.,
2011). They remain viable in water and in decaying tissues for extended periods (OIE, 2019b).
Infection in farmed fish has been traced to the use of frozen wild-sourced fish and coleoid
cephalopods as feed in aquaculture (Castric, 1997; Mori et al., 2005; Gomez-Casado et al., 2011;
Chi et al., 2016; OIE, 2019b).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 65
NNV strains may be dispersed by scavenging piscivorous birds, fomites, or through
contaminated fish processing equipment (Sano et al., 2011). The host range is wide (OIE, 2019b)
and the infectious dose is assumed to be low-moderate (105.5 TCID50 fish-1) (Nishi et al., 2016).
Viral titre is high in decaying fish (between 109.5 and 1010.0 TCID50 g-1) and in seawater (up to
105 TCID50 L-1) due to the high shedding rates of surviving fish (Nishi et al., 2016).
The likelihood of exposure and establishment is assessed as medium.
10.2.3 Consequence assessment
Economic consequences
Infection with NNV is not an OIE-listed disease (OIE, 2019a) so establishment would not result
in restrictions on trade access for New Zealand. NNV strains cause disease with moderate-high
morbidity and mortality in over 70 species of marine and freshwater fish (Castric, 1997; Munday
et al., 2002; Gomez-Casado et al., 2011; Sano et al., 2011; Doan et al., 2017; Blackwell, 2019).
Establishment would likely have economic consequences for the developing marine aquaculture
of yellowtail kingfish (Seriola lalandi), hapuku/grouper (Polyprion oxygeneios) and snapper
(Pagrus auratus). (NIWA, 2017a, 2017b; Plant & Food, 2017).
Little is known about the effect of introduced viruses on wild fish populations (Lafferty et al.,
2015). Clinical disease is uncommon in wild stocks, but the prevalence of subclinical infection
varies widely (from 20% to 100%) (de la Pena et al., 2011; Berzak et al., 2019). Establishment
may result in decreased productivity and increased mortality in inshore commercial fisheries
including jack mackerel (Trachurus spp.) (export value NZ $68 million in 2018) and snapper
(Pagrus auratus) (export value $33 million in 2018) (Plant & Food, 2017; MPI, 2018a, 2018b,
2018c; Seafood New Zealand, 2019). Infected wild species may also act as a reservoir for
infection of farmed fish (Berzak et al., 2019).
NNV strains may also be dispersed by piscivorous birds (OIE, 2019b) into the freshwater
environment. Establishment may result in lowered productivity and increased mortality in grass
carp (Ctenopharyngodon idella) and silver carp (Hypophthalmichthys molitrix) farmed for use in
weed control (Clayton & Wells, 1999; Blackwell, 2019), as well as developing aquaculture of
eels (Anguilla spp.) (NIWA, 2017c). Other introduced cyprinids including goldfish (Carassius
auratus) may act as reservoir hosts (Doan et al., 2017).
Social consequences
NNV strains are not considered to be zoonotic. Their establishment may cause reduced
productivity and increased mortality in marine and freshwater fish and shellfish species of
importance for customary and recreational fishing (Wynne-Jones et al., 2019, 2019; Blackwell,
2019).
Recreational fishing is the fifth most popular recreational activity for adults, involving 700,000
marine fishing trips annually, with a contribution of NZ$ 638 million to the GDP (Gross
domestic product) in 2015 (Southwick et al., 2018).
The snapper, hapuku and trevally recreational fisheries contributed NZ$ 402 million, NZ$ 12.5
million and NZ$ 71 million respectively to the GDP in 2015 (Southwick et al., 2018). Several
species including snapper and eels (Anguilla spp.) are taonga for Maori culture and important for
customary fishing (MPI, 2019c). These fish stocks are considered stressed (MPI, 2018a, 2018b,
2018c) and further reduction in their abundance would have significant social effects for New
Zealand.

66 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Environmental consequences
NNV strains affect a wide range of marine and freshwater fish (Blackwell, 2019). However, little
is known about the effects of viral infection in wild fish stocks (Lafferty et al., 2015). Species
such as clupeids perform an important role linking trophic levels in food webs and represent a
significant food source for other fish, seabirds and marine mammals (Varpe et al., 2005; Dunn et
al., 2012; Whitfield et al., 2012). Reduction in herring (C. p. pallasi) abundance (from over-
fishing) caused a reported 60% decrease in the relative abundance of wild salmon
(Oncorhynchus tshawytscha) in coastal British Columbia, with major biological, economic and
social consequences for humans and wildlife (Pagowski et al., 2019).
NNV strains affect invertebrates including bivalve and coleoid molluscs, crustaceans and annelid
worms (OIE, 2019b; Volpe et al., 2018; Blackwell, 2019), but the environmental significance is
unknown.
NNV strains may also be dispersed by piscivorous birds (OIE, 2019b) and affect freshwater
species including eels (NIWA, 2017c), where introduced cyprinids including goldfish (Carrasius
auratus) may act as reservoir hosts (Doan et al., 2017). The environmental significance of
establishment in freshwater species is unknown.
The establishment of NNV strains would have negative economic, social and environmental
consequences. The consequences of establishment are assessed to be medium.
10.2.4 Risk estimation
The entry, exposure, establishment and consequence assessments for NNV strains are medium,
so the risk estimate is medium. Under the procedures followed in this risk analysis, risk
management measures may be justified.
10.3 Risk management
NNV strains have been assessed to be a risk in the commodity. Infection with NNV is a not an
OIE-notifiable disease, so the Aquatic Code (OIE, 2019a) provides no specific guidance or
processing requirements that would ensure the destruction of the virus.
NNV strains are associated with several finfish and cephalopod species (Table 10). Therefore,
restriction to non-susceptible species in the commodity should substantially reduce the risk
associated with NNV strains and be a viable risk management option.
NNV strains have a wide geographic range (Table 10). Their occurrence in wild stocks in other
regions is likely to be low. Restriction through the BNZ Country Approval Procedures to regions
where NNV strains have not been reported should substantially reduce the NNV risk and be a
viable risk management option.
The presence of NNV strains in a shipment of fish bait may be determined by batch testing,
using standard sampling procedures (Corsin et al., 2009; OIE, 2019a). Standard diagnostic
procedures for detection of the virus are available (Arimoto et al., 1982; Panzarin et al., 2010;
OIE, 2019b). Verification of a negative test from a shipment should substantially reduce the
NNV risk and be a viable risk management option.
NNV strains are unaffected by freezing (OIE, 2019b). Therefore, frozen storage (at -18 °C to -20
°C for either 168 hours, or for 4 months) is not a viable risk management option.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 67
NNV strains are denatured by ionising radiation (at 50 kGy) (DAFF, 2013). This treatment
should substantially reduce the NNV risk and be a viable risk management option.
10.3.1 Risk management options
In addition to the general risk management measures that are proposed for all imported fish bait
(section 6.2.1), one or more of the following risk management options would reduce the NNV
specific risk to an acceptable level:
Option 1
• Competent Authority attestation that the commodity does not include: Atlantic horse
mackerel (Trachurus trachurus), Chilean jack mackerel (T. murphi), Japanese jack mackerel
(T. japonicus), Pacific jack mackerel (T. symmetricus) (Carangidae); Atlantic herring
(Clupea harengus) (Clupeidae); flathead grey mullet (Mugil cephalus), golden grey mullet
(Chelon auratus), thicklip grey mullet (Chelon labrosus), thinlip grey mullet (Chelon
ramada) (Mugilidae); blue mackerel (Scomber australasicus), chub mackerel (Scomber
japonicus), Atlantic mackerel (Scomber scombrus) (Scombridae), Japanese flying squid
(Todarodes pacificus) (Ommastrephidae); and common octopus (Octopus vulgaris)
(Octopodidae) should substantially reduce the risk associated with NNV strains, so the
commodity could be imported without any further restrictions.
Option 2
• Acceptance of region/country freedom through the BNZ Country Approval Procedures
should substantially reduce the risk associated with NNV strains, so the commodity could
be imported without any further restrictions.
Option 3
• A negative test result from a shipment of the commodity, following approved sampling and
diagnostic procedures, through the BNZ Country Approval Procedures should substantially
reduce the risk associated with NNV strains, so the commodity could be imported without
further restrictions.
Option 4.
• Treatment with ionising radiation (at 50 kGy) should substantially reduce the risk
associated with NNV strains, so the commodity could be imported without any further
restrictions.
Orthomyxoviridae: Infectious salmon anaemia virus (ISAV)
11.1 Technical review
11.1.1 Aetiological agent
Infectious salmon anaemia virus (ISAV) is classified in the Genus Isavirus, within the Family
Orthomyxoviridae (OIE, 2019a). Two forms of ISAV are recognised: highly pathogenic HPR
(highly polymorphic region)-deleted ISAV, and low-virulent HPR0 ISAV (the non-deleted HPR
form of ISAV). Both forms may be present in fish populations (Cárdenas et al., 2014). Outbreaks
of ISAV may have resulted from the emergence of HPR-deleted ISAV from low-virulent HPR0
ISAV (Cunningham et al., 2002; Mjaaland et al., 2002; Christiansen et al., 2017; Gagné &
LeBlanc, 2018; OIE, 2019a).

68 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Several distinct strains of each form have been isolated, based on geographical area (Miller &
Cipriano, 2003; OIE, 2019b).
11.1.2 OIE status
Infection with either HPR0 or HPR-deleted ISAV strain is an OIE-notifiable disease (OIE,
2019a).
11.1.3 New Zealand status
New Zealand is considered free from both HPR0 and HPR-deleted forms of ISAV (Stone et al.,
1997; Norman et al., 2013). Infection with either form of ISAV is a notifiable disease in New
Zealand (Anon., 2016).
11.1.4 Zoonotic disease
ISAV strains are not zoonotic (CABI, 2019b).
11.1.5 Epidemiology
Distribution and host range
The HPR0 and HPR-deleted strains of ISAV are widely distributed (Table 11). Prevalence varies
widely (from 5% to 90%), depending on pathogen strain and host species (Le Blanc et al. 2018).
The species associated with ISAV strains in fish bait and their geographic/pathogen range are
given in Table 11. Infection of Atlantic herring (C. harengus) with HPR0 strains mainly occurs in
close proximity to infected farmed salmonids (OIE, 2019b).
HPR-deleted strains are not reported from wild fish bait species (OIE, 2019a). However, Atlantic
herring (C. harengus) and Pacific herring (C. p. pallasii)) can be experimentally infected (by
bath immersion) with isolates from infected salmonids (Table 11) (Biacchesi et al., 2007; OIE,
2019a; CABI, 2019b). Other, as yet unknown marine host species may also function as
reservoirs of infection (OIE, 2019b)

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 69
Table 11. Fish bait species susceptible to strains of infectious salmon anaemia virus (ISAV)
HPR0 strains (low virulence)
Family Host Species Host geographical range1 Pathogen range2
Clupeidae Atlantic herring (Clupea harengus)
Atlantic, cosmopolitan Northwest, Northeast Atlantic (Canada, Europe, Norway, UK, USA)
HPR-deleted strains (high virulence)
Family Host species Host geographical range1 Pathogen range2
Clupeidae Atlantic herring (Clupea harengus)
Atlantic, cosmopolitan Not reported from wild fish3
Clupeidae Pacific herring (Clupea pallasii pallasii)
Indo-Pacific, Pacific, cosmopolitan
Not reported from wild fish3
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature). The HPR0 strains are widely reported from wild salmonids in marine and
freshwater salmon growing regions including Norway, Scotland, Ireland, New Brunswick, Nova Scotia, Maine and Chile (OIE, 2019a). The HPR-deleted strains are widely reported from Atlantic salmon (Salmo salar) in the Northeastern Atlantic Ocean (Europe, the Faroe Islands), United Kingdom (Scotland), Middle East (Iraq), Northwest Atlantic Ocean (Canada in the Bay of Fundy), the United States (Maine), and South America (Chile) (OIE 2019a; Blackwell 2019).
3 Reported experimental infection of HPR-deleted strains of ISAV
Potential host species in New Zealand
Species at risk in New Zealand include brown trout (Salmo trutta) and rainbow trout
(Oncorhynchus mykiss) in marine and fresh waters, while Pacific herring C. p. pallasii) may
represent a reservoir for infection. Other Pacific salmon species including Chinook salmon (O.
tshawytscha) are considered resistant to HPR0 and HPR-deleted strains of ISAV (Rimstad et al.,
2011; Norman et al., 2013; Kibenge & Godoy, 2016).
Pathogenicity
HPR0 strains cannot be maintained in cell culture, so infectivity and pathogenicity processes are
poorly known (OIE, 2019b). HPR0 strains are of low virulence. They cause transient subclinical
infections in apparently healthy salmonids (Salmo spp.) in marine and fresh waters. All ages of
fish may be infected (OIE, 2019b). While these fish are not considered to be carriers (EFSA,
2012; OIE, 2019a), the role of HPR0 strains in the re-emergence of pathogenic HPR-deleted
strains remains unclear (Cunningham et al., 2002; EFSA, 2012; OIE, 2019a).
HPR-deleted strains cause long-term chronic disease in farmed Atlantic salmon (S. salar) with
associated cumulative mortalities of up to 90% (OIE, 2019a; Blackwell, 2019). Infection in non-
salmonids usually progresses without clinical signs (Cunningham et al., 2002). HPR-deleted
strains isolated from experimentally infected Pacific herring (C. p. pallasii) are infective to
salmonids (OIE, 2019b).
Clinical signs
Infection with HPR0 strains occurs with no clinical signs in non-salmonids (OIE, 2019b).
Infection with HPR-deleted strains in salmonids is characterised by exophthalmia and darkening
of the skin usually within 2-4 weeks post-infection. Infection in non-salmonids typically occurs
with no clinical signs of infection (OIE, 2019a).

70 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
The presence of ISAV may be detected by PCR in a range of species, including Chinook salmon
(O. tshawytscha), in the absence of clinical signs (Kibenge & Godoy, 2016). The significance of
these benign infections is uncertain, but they may be the source of clinical disease in farmed
Atlantic salmon (Salmo salar) (Murray et al., 2002; Kibenge & Godoy. 2016).
Pathogen survival
From experimental data, ISAV strains remain viable in marine or fresh water, for up to 4 months
in seawater at 4 °C (Rimstad & Mjaaland, 2002) and up to 14 days at temperatures up to 15 °C
(Falk et al., 1997; MacLeod et al., 2003; Rimstad et al., 2011). These strains survive up to 6 days
in fish tissues held on-ice (Tubbs et al., 2007; OIE, 2019a). They are sensitive to UV radiation
(Oye & Rimstad, 2001), but their survival in natural waters may be influenced by dilution and
predation factors (see section 6.1.2).
ISAV strains may remain viable for up to 4 months in moist decaying fish tissues dumped in
landfill (Rimstad & Mjaaland, 2002; Rimstad et al., 2011).
Transmission
Transmission may be horizontal or vertical, but the epidemiology of ISAV is currently unclear
(OIE, 2019a, 2018b). Dispersal may occur by fomites including fishing boats and gear, transport
and factory processing equipment, as well as through fish wastes, blood, melt and wash water
(OIE, 2019b). ISAV strains survive passage through the avian digestive system (OIE, 2019a) and
may be dispersed between waterways through piscivorous and scavenging birds (OIE, 2019a).
Infection and disease progression
HPR0 strains targets the epithelial cells of the gills (Aamelfot et al. 2012), as well as kidney and
heart tissue (Christiansen et al., 2011; Lyngstad et al.,2011).
HPR-deleted strains cause chronic disease initially infecting the gills, skin surface and intestinal
tract. Disease then becomes systemic, affecting all major organs (OIE, 2019a).
Inactivation
ISAV strains are unaffected by short (168 hours), or medium-term (4 months) frozen storage (at
-18°C) and survive 5 freeze-thaw cycles with no loss in viability (Rimstad et al., 2011; OIE,
2019b). ISAV strains are assumed to be denatured by ionising radiation at 50 kGy (DAFF,
2013).
11.2 Risk assessment
11.2.1 Entry assessment
In fish bait, HPR0 strains are only associated with subclinically infected wild Atlantic herring
(C. harengus) (Table 11), where they are caught in close proximity to infected salmon farms
(OIE, 2019a). While no data are available on disease prevalence, it is reasonable to assume that
fishing for herrings is unlikely to take place close to marine salmon farms. The prevalence is
therefore likely to be low in wild stocks. The likelihood of entry of HPR0 strains is assessed to
be low.
HPR-deleted strains are not reported from fish bait (Table 11). The likelihood of entry of HPR-
deleted strains is assessed to be so low as to be negligible.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 71
11.2.2 Exposure and establishment assessment
HPR0 strains are unaffected by frozen storage. They may remain viable in thawed fish tissues for
up to 6 days and in the water column for a further 7 days, within a temperature range from 4 °C
to 15 °C (MacLeod et al., 2003; Tubbs et al., 2007; OIE, 2019a).
Infection may also occur through fomites including fishing boats and gear, transport and factory
processing equipment, as well as through discarded bait. HPR0 strains may also be dispersed
through offal and discards into waterways by piscivorous birds (OIE, 2019b).
The risk of exposure and establishment of HPR0 strains is considered to be medium.
11.2.3 Consequence assessment
Economic consequences
HPR0 ISAV is an OIE-listed disease, so establishment could result in restrictions on trade access
for New Zealand exports, as well as indirect economic consequences for the fishing, fish
processing and tourist industries. ISAV strains are associated with fish of families Clupeidae,
Gadidae and Salmonidae (Blackwell, 2019). New Zealand finfish aquaculture exports are
focused on Chinook salmon (O. tshawytscha) which is considered resistant to infection (OIE,
2019a), but ISAV presence may be detected by PCR analysis (Kibenge & Godoy, 2016).
While establishment of an OIE-listed disease may have serious economic consequences for New
Zealand trade, the HPR0 strains are of low pathogenicity (OIE, 2019a). The economic
consequences of their establishment on other commercial fisheries are assumed to be negligible.
Social consequences
ISAV strains are not zoonotic (CABI, 2019b). The HPR0 strains are unlikely to have a direct
effect on fish stocks considered significant for customary or recreational fishing (Wynne-Jones
et al., 2014, 2019). The social consequences of establishment are considered negligible.
Environmental consequences
The HPR0 strains of ISAV are associated with transient, subclinical infection which usually
occurs with no clinical signs (OIE, 2019a, OIE, 2019b). Prevalence in fish bait is likely to be
very low, so establishment would be unlikely to cause significant effects on wild fish (EFSA,
2012; OIE, 2019a, OIE, 3019b). The environmental consequences of establishment of the HPR0
strains are negligible.
The HPR0 strains of ISAV are of low pathogenicity and cause transient subclinical infection.
The economic, social and environmental consequences of establishment through fish bait are
assessed as negligible.
11.3 Risk management
As the entry and consequence assessments are assessed to be negligible, ISAV and other
orthomyxoviruses are not considered to be risks in the commodity. No further assessment is
necessary.

72 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Reoviridae: Piscine aquareovirus (PRV) and associated aquareoviruses
12.1 Technical review
12.1.1 Aetiological agent
Piscine aquareovirus (PRV) is a group of seven closely associated species (Aquareovirus A -
Aquareovirus G), each comprised of several strains (Lupiani et al., 1993; Bustos et al., 2011; Ke
et al., 2011; King et al., 2011; Blindheim et al., 2015). The relationships between these species
and other associated but uncharacterised strains is unclear (Fauquet et al., 2005; Palacios et al.,
2010; Biering & Garseth, 2012; Kristoffersen et al., 2013). PRV strains are agents of heart and
skeletal muscle inflammation (HSMI), cardiomyopathy syndrome (CMS) and erythrocytic body
inclusion syndrome (EBIS). Co-infection with multiple strains may also occur (Wiik-Nielsen et
al., 2011).
12.1.2 OIE status
Disease caused by PRV strains is not notifiable to the OIE (OIE, 2019a).
12.1.3 New Zealand status
PRV strains have not been reported from New Zealand (Cobb, 2008; B. Jones, pers. comm.,
2015). Infection with PRV is not a notifiable disease in New Zealand (Anon., 2016).
12.1.4 Zoonotic disease
PRV strains are not zoonotic (Anon., 2020a).
12.1.5 Epidemiology
Distribution and host range
PRV strains are widely distributed (Table 12). They are reported from 12 families of fish
(Blackwell, 2019). Infection involves multiple carrier species and virus reservoirs (Roberts,
2012; Wiik-Nielsen et al., 2012; Zhang & Gui, 2015).
The species associated with PRV strains in fish bait and their geographic/pathogen range are
given in Table 12. The prevalence of PRV strains in wild Atlantic horse mackerel (T. trachurus)
and Atlantic herring (C. harengus) is low (Wiik-Nielsen et al., 2012; Carlile et al., 2014).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 73
Table 12. Fish bait species susceptible to piscine aquareovirus and related strains
Family Host Species Host geographical range1 Pathogen range2
Carangidae Atlantic horse mackerel (Trachurus trachurus)
Mediterranean, East Atlantic Northeast, Northwest Atlantic (Canada, Denmark, Norway, United Kingdom, Spain)
Clupeidae Atlantic herring (Clupea harengus)
Atlantic Northeast Atlantic (Denmark, Norway, United Kingdom, Spain)
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature). PRV is reported from the Northeast Atlantic Ocean (Denmark, Norway,
Scotland, Spain), Northwest Atlantic (Canada); the Northeast and Central Pacific Ocean (Australia, China, Japan), and South America (Chile) (Chen & Jiang, 1984; Fauquet et al., 2005; King et al., 2011; Garseth et al., 2012; Roberts, 2012; Wiik-Nielsen et al., 2012; Carlile et al., 2014; Zainathan et al., 2014).
Potential host species in New Zealand
Susceptible species in New Zealand include salmonids, silverside (Argentina elongata),
mackerel (Trachurus spp.) and Pacific herring (C. p. pallasii) (Paul, 2000; Tubbs et al., 2007).
The New Zealand brill (Colistium guntheri) and turbot (C. nudipinnis) (Pleuronectidae) are
related to the Northern Hemisphere brill (Scophthalmus rhombus) and turbot (S. maximus)
(Pleuronectidae) (Chanet, 2003). Thus, it is reasonable to conclude that pathogens affecting
turbot and brill (Scophthalmus spp.) may infect endemic brill and turbot species (C. Johnston,
pers. comm., 2015).
Pathogenicity
PRV strains reported from salmonids are of high morbidity, low-medium pathogenicity, with a
mortality rate of 5–20% (Kongtorp et al., 2004a, 2004b; Wiik-Nielsen et al., 2012). Subclinical
infection also occurs in wild salmonids at low prevalence rates, with no clinical signs (Palacios
et al., 2010; Wiik-Nielsen et al., 2012; Anon., 2020a).
Pathogenicity is complex, varying among strains and between host species (Wiik-Nielsen et al.,
2012). It is also influenced by water temperature and the presence of co-infecting viral pathogens
(Crane & Carlisle, 2010; Wiik-Nielsen et al., 2012; Zainathan et al., 2014; Johansen et al., 2015;
Garver et al., 2016; Wessel et al., 2017).
PRV strains associated with Atlantic horse mackerel (T. trachurus) and Atlantic herring (C.
harengus) are of low pathogenicity (Carlile et al., 2004). Disease occurrence in these species is
not geographically related to disease occurrence in salmonids and their role as disease reservoirs
for adjacent farmed species is poorly understood (Wiik-Nielsen et al., 2012; Johansen et al.,
2015; Garver et al., 2016).
Clinical signs
Infection in Atlantic horse mackerel (T. trachurus) and Atlantic herring (C. harengus) occurs
with few or no external clinical signs (Garseth et al., 2012; Wiik-Nielsen et al., 2012; Carlile et
al., 2014).
Pathogen survival
From experimental data, PRV strains remain viable for almost 6 months in distilled water
(McDaniels et al., 1983), up to 13 days in seawater (Wiik-Nielsen et al., 2012) and up to 12
months in sewage sludge at 8 °C (Wellings et al., 1976). They remain viable in low moisture

74 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
processed foods for 2 months at 5 °C (Pirtle & Beran, 1991). Survival in natural waters may be
influenced by dilution and predation factors (see section 6.1.2), but PRV strains are likely to
remain viable in fish bait discarded into marine waters (Kongtorp et al., 2004a, 2004b; Wiik-
Nielsen et al., 2012).
Transmission
Transmission is horizontal and direct, through the water column (Watanabe et al., 2006; Garver
et al., 2016),
Infection and disease progression
Infection in salmonids initially focuses on skeletal and heart muscle, causing ventricular lesions,
necrosis and myositis. This spreads to the gills, causing haemorrhages and necrosis. Infection
then becomes systemic, affecting the liver, spleen, kidney and intestine. (Wessel et al., 2017),
with haemorrhages of the jaws, operculum and fin bases (Rivas et al., 1996a, 1996b; Aldrin et
al., 2010; Kibenge & Godoy, 2016).
Inactivation
PRV strains remain viable after long-term frozen storage (at -80 °C) and survive 3 freeze-thaw
cycles (Hauge et al., 2016).) They are therefore assumed to be unaffected by either short (168
hours), or medium-term (4 months) frozen storage (at -20 °C). PRV strains are assumed to be
denatured by ionising radiation (at 50 kGy) (DAFF, 2013).
12.2 Risk assessment
12.2.1 Entry assessment
PRV strains occur at low prevalence in fish bait but have a wide geographical distribution (Wiik-
Nielsen et al., 2012). They are associated with Atlantic herring (C. harengus) and Atlantic horse
mackerel (T. trachurus) (Table 12). Imports of T. trachurus as fish bait in 2018 (Table 3)
represented over 10% by volume, while C. harengus was not imported.
PRV strains are unaffected by frozen storage. Infected fish may be present in fish bait at a low
rate of prevalence (Wiik-Nielsen et al., 2012).
The likelihood of entry of PRV strains is assessed as medium.
12.2.2 Exposure and establishment assessment
PRV strains are assumed to be unaffected by frozen storage for at least 4 months (at -20 °C) and
survive multiple freeze-thaw cycles (Hauge et al., 2016). From experimental data, PRV strains
remain viable for up to 13 days in seawater and for up to 12 months in marine sediments
(Wellings et al., 1976; Wiik-Nielsen et al., 2012). Survival in natural waters is unknown (section
6.1.2). Potential alternative hosts exist in New Zealand (Paul, 2000; Tubbs et al., 2007).
Prevalence is low in wild stocks and the strains associated with fish bait are of low
pathogenicity. Disease establishment is complex and likely to require co-infection with other
viral pathogens (Johansen et al., 2011; Wiik-Nielsen et al., 2012; Carlile et al., 2014; Johansen et
al., 2015; Garver et al., 2016; Wessel et al., 2017).
The likelihood of exposure and establishment is therefore assessed to be so low as to be
negligible.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 75
12.3 Risk management
As the likelihood of exposure and establishment of PRV strains through the commodity is
assessed to be negligible, no further risk assessment is necessary.
Rhabdoviridae: Infectious haematopoietic necrosis virus (IHNV)
13.1 Technical review
13.1.1 Aetiological agent
Infectious haematopoietic necrosis virus (IHNV) is a single-stranded RNA virus classified in the
genus Novirhabdovirus, within the Family Rhabdoviridae (Bootland & Leong, 2011; OIE,
2019a). It was first identified from sockeye salmon (Oncorhynchus nerka) on the West Coast of
the United States (Rucker et al.,1953; Wingfield et al.,1969). One serotype is recognised
(Bootland & Leong, 2011), with several genogroups. These vary widely in pathogenicity and are
geographically based. They have a wide host range within each region (Bootland & Leong,
2011).
13.1.2 OIE status
Infection with IHNV is listed by the OIE as a notifiable disease (OIE, 2019a).
13.1.3 New Zealand status
IHNV has not been reported from New Zealand (Boustead et al., 1993; Stone et al., 1997). It is
considered an exotic notifiable disease (Anon., 2016).
13.1.4 Zoonotic disease
IHNV is not zoonotic (OIE, 2019a).
13.1.5 Epidemiology
Distribution and host range
IHNV occurs in 11 families of marine and freshwater fish (Blackwell, 2019), as well as from
marine and freshwater invertebrates (Bootland & Leong, 2011; OIE, 2019b).
IHNV is widely distributed (Table 13). Infection in non-salmonids commonly occurs without
clinical signs, so the host range is poorly known. Reports are based on targeted experimental
challenge and from incidental detection during surveillance programmes or surveys (Dixon et
al., 2016).
The species associated with IHNV in fish bait and their geographic/pathogen range are given in
Table 13 (Hart et al., 2011; OIE, 2019a).
Potential host species in New Zealand
Susceptible fish hosts in New Zealand include all salmonids and a wide range of other marine,
estuarine and freshwater fish (Tubbs et al., 2007).

76 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Table 13. Fish bait species susceptible to infectious haematopoietic necrosis virus (IHNV)
Family Host Species Host geographical range1 Pathogen range2
Clupeidae Pacific herring (Clupea pallasii pallasii)
Pacific Northeast, Northwest Pacific
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature). IHNV is widely distributed in marine and fresh waters of North America
(Pacific and Northwest USA), South America (Bolivia, Chile), Asia (China, Japan, South Korea and Chinese Taipei (Taiwan), Europe (Austria, Croatia, Czech Republic, France, Germany, Italy, Netherlands, Poland, Slovenia and Spain), Iran, Kuwait, Russia and Pakistan (Tubbs et al., 2007; Dixon et al., 2016; OIE, 2019b).
Pathogenicity
Mortality in farmed fish varies widely (from 50% to 90%), depending on viral strain, host
species and environmental factors (Munro & Midtlyng, 2011; Dixon et al., 2016). Epizootics are
associated with water temperatures between 8 °C and 14 °C (Dixon et al., 2012).
Pathogenicity is much lower in wild stocks, where chronically infected surviving fish may act as
carriers (Bootland & Leong, 2011).
Clinical signs
Infection may occur with no clinical signs, but external lesions, skin darkening and petechial
haemorrhages at the base of the fins, vent, gills and mouth may be present (Bootland & Leong,
2011).
Transmission
Transmission can be vertical or horizontal (OIE, 2019b). Piscivorous birds and invertebrates
including mayflies (Callibaetis spp.), salmon lice (Lepeophtheirus salmonis), salmon leeches
(Piscicola spp.) and parasitic copepods (Salmonicola spp.) may function as vectors in marine and
fresh waters (Bootland & Leong, 2011; Kent et al., 1998; OIE, 2019a, OIE, 2019b).
Infection and disease progression
The infective dose is relatively low (103 pfu ml-1) (Traxler et al., 1993; Bootland & Leong,
2011). Entry occurs through the epidermal cells of the gills, skin, fin bases, oral region, or
through the digestive system (Stone et al., 1997; Bootland & Leong, 2011).
Infection in salmonids is associated with the haematopoietic tissues (liver, spleen and kidney),
(Munro & Midtlyng, 2011). Infection may spread to brain tissue (Samuelsen et al., 2006).
The infectivity of IHNV in wild non-salmonid fish is poorly understood. Infection may be
transient and progress with no clinical signs (Garver & Wade, 2017; OIE, 2019b). The role of
non-salmonids as reservoir hosts for salmonids is uncertain (Garver & Wade, 2017).
From experimental data, strains isolated from eels (Anguilla spp.) are infective to salmonids
(OIE, 2019a). Experimental infection of salmonids (by bath immersion at a low dose rate (102 to
103 pfu ml-1 (plaque forming units) for 10 minutes) with strains from Pacific herring (C. p.
pallasii) did not cause clinical infection. However, an asymptomatic carrier state developed, with
low (<10%) mortality, at a prevalence of up to 70% (Bergmann et al., 2002; Bootland & Leong,
2011; Hart et al., 2011). These experimental data may not fully reflect disease progression in the
natural environment (Free et al., 2006; Dixon et al., 2016).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 77
Pathogen survival
IHNV remains infective over a wide temperature range (from 3°C to 18°C) (Bootland & Leong,
2011; OIE, 2019a). From experimental data, it remains viable for up to 30 days in fresh waters,
14 days in marine or estuarine waters and up to 9 weeks in sediment (Stone et al., 1997;
Yoshimizu et al., 2005; OIE, 2019b). However, survival in natural waters is unknown, and may
be influenced by dilution and predation factors (see section 6.1.2).
Inactivation
IHNV remains viable in frozen fish brain tissue (to -20 °C for 5 months) and is resistant to
multiple freeze-thaw cycles (Stone et al., 1997; Tubbs et al., 2007; Dixon et al., 2016). It is
therefore assumed to be unaffected by either short (168 hours), or medium-term (4 months)
frozen storage (at -20 °C).
IHNV is assumed to be denatured by ionising radiation (at 50 kGy) in frozen fish tissue (DAFF,
2013).
13.2 Risk assessment
13.2.1 Entry assessment
IHNV remains viable in the frozen commodity (Stone et al., 1997; Bootland & Leong, 2011;
Dixon et al., 2016) and infection may occur with no external signs. The prevalence in fish bait is
unknown, but likely to be low (Lafferty et al., 2015).
The likelihood of entry is therefore assessed as low.
13.2.2 Exposure and establishment assessment
IHNV remains viable for several months in marine or fresh waters and sediments (Kahn et al.,
1999; Yoshimizu et al., 2005). IHNV strains are geographically related, with low apparent host
specificity among fish and invertebrate hosts (Bootland & Leong, 2011; OIE, 2019b). The
infectious dose is low (103 pfu ml-1) (Traxler et al., 1993; Bootland & Leong, 2011), but strains
vary widely in pathogenicity, morbidity and mortality (OIE, 2019a).
The epidemiology of IHNV in Pacific herring (C. p. pallasii) is poorly known and disease may
be transient (Garver & Wade, 2017). Surviving fish may remain carriers of disease (Bootland &
Leong, 2011; Mussely & Goodwin, 2012; Sim-Smith et al., 2014; Symonds et al., 2014).
Experimental data indicate IHNV isolates from Pacific herring can infect salmonids (Foot et al.,
2006; Hart et al., 2011; Dixon et al., 2012), but their potential to act as a reservoir host is
uncertain (Garver & Wade, 2017).
The likelihood of exposure and establishment is therefore assessed as low.
13.2.3 Consequence assessment
Economic consequences
Infection with IHNV is an OIE-listed disease, so establishment would result in loss of market
access, with far-reaching consequences for the New Zealand economy. Establishment in
salmonid aquaculture would also result in lowered productivity and increased mortality, with
serious economic consequences through lost production. Salmonid aquaculture in New Zealand

78 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
was valued at $77 million in 2018 (Aquaculture New Zealand, 2019). Other significant indirect
costs would result from destocking, disinfection and ongoing monitoring to regain disease-free
status (OIE, 2019a).
IHNV has a wide host range affecting 11 families of fish (Blackwell, 2019; OIE, 2019a). Little is
known about the economic consequences on non-salmonid fish (Lafferty et al., 2015). These
may include losses in productivity and increased mortality in the developing aquaculture and
major inshore fishery for snapper (Sparus aurata) (Sparidae). Snapper exports from capture
fishing were valued at $33 million in 2018 (Seafood New Zealand, 2019).
Social consequences
IHNV is not zoonotic. Establishment in the major customary and recreational fisheries (Wynne-
Jones et al., 2014, 2019) including silver sea bream (snapper) (Pagrus auratus) in coastal waters,
and eels (Anguilla spp.) in fresh water (MPI, 2018b), would be likely to incur significant social
consequences. These species are considered taonga for Maori (MPI, 2013a, 2013b, 2013c).
Recreational fishing is the 5th most popular recreational activity for adults, involving 700,000
marine fishing trips annually, with a contribution of NZ$ 638 million to the GDP (Gross
domestic product) in 2015 (Southwick et al., 2018). The snapper recreational fishery contributed
NZ$ 402 million to the GDP (Southwick et al., 2018).
Environmental consequences
Inshore fish species including clupeids are considered likely to encounter and consume discarded
fish bait. IHNV has a wide host range, but pathogenicity varies widely among strains and
between hosts (Bootland & Leong, 2011; Garver & Wade, 2017). The environmental
consequences for wild fish stocks are essentially unknown (Lafferty et al., 2015; Garver &
Wade, 2017).
Clupeids provide a significant food source for other fish, seabirds and marine mammals and a
major link between trophic levels in the aquatic food web (Dunn et al., 2012), so reduction in
biomass due to disease could have significant environmental consequences.
IHNV is an OIE-listed disease affecting a wide host range, with major economic, social and
environmental consequences for New Zealand. The consequences of establishment are assessed
as medium-high.
13.2.4 Risk estimation
The entry, exposure, establishment and consequence assessments are low to medium-high. While
uncertainty remains about the pathogenicity, occurrence and epidemiology of the strains
associated with clupeid fish, the risk of entry and establishment of IHNV is estimated to be low-
medium. Under the procedures followed in this risk analysis, risk management measures may be
justified.
13.3 Risk management
IHNV has been assessed to be a risk in the commodity. As an OIE-listed disease, the OIE
Aquatic Code (OIE, 2019a) provides some guidance on processing requirements that would
ensure destruction of the virus. However, the proposed heat treatments would make the
commodity unfit for purpose. These options are not considered further.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 79
IHNV is unaffected by frozen storage (at -20 °C for at least 5 months) (Dixon et al., 2016).
Therefore, frozen storage at -20 °C, for either 168 hours, or for 4 months, is not a viable risk
management option.
Treatment of the commodity using ionising radiation (at 50 kGy) (DAFF, 2013) should
substantially reduce the IHNV risk and be a viable risk management option.
IHNV is associated with the commodity (Table 13). Therefore, restriction to non-susceptible
species in the commodity should substantially reduce the IHNV risk and be a viable risk
management option.
IHNV has a wide geographic range (Table 13). The occurrence is likely to be low or negligible
in other geographic regions. Restriction through the BNZ Country Approval Procedures to
regions where IHNV has not been reported should substantially reduce the IHNV risk and be a
viable risk management option.
The presence of IHNV in a shipment of the commodity may be determined by batch testing
using standard sampling procedures (Corsin et al., 2009; OIE, 2019a). Standard diagnostic
procedures for detection of the virus are provided in Emmenegger et al. (2000) and OIE (2019b).
Verification of a negative test from a shipment should substantially reduce the IHNV risk and be
a viable risk management option.
13.3.1 Risk management options
In addition to the general risk management measures that are proposed for all imported fish bait
(section 6.2.1), one, or more of the following risk management options would reduce the ENV
specific risk to an acceptable level:
Option 1
• Competent Authority attestation that the commodity does not include Pacific herring (Clupea
pallasii pallasii) (Clupeidae) should substantially reduce the IHNV risk, so the commodity
could be imported without any further restrictions.
Option 2
• Acceptance of region/country freedom through the BNZ Country Approval Procedures
should substantially reduce the IHNV risk so the commodity could be imported without
any further restrictions.
Option 3
• A negative test result from a shipment of the commodity, following approved sampling and
diagnostic procedures, through the BNZ Country Approvals Procedures should substantially
reduce the IHNV risk, so the commodity could be imported without further restrictions.
Option 4
• Treatment with ionising radiation (at 50 kGy) should substantially reduce the IHNV risk, so
the commodity could be imported without any further restrictions.

80 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Rhabdoviridae: Viral haemorrhagic septicaemia virus (VHSV)
14.1 Technical review
14.1.1 Aetiological agent
Viral haemorrhagic septicaemia virus (VHSV) is a single-stranded RNA virus, classified in the
Genus Vesiculovirus, within the Family Rhabdoviridae (OIE, 2019a). It is the agent of viral
haemorrhagic septicaemia (VHS), an acute and economically significant haemorrhagic viral
disease of marine and freshwater fish (Einer-Jensen et al., 2004; Skall et al., 2005a, 2005b; OIE,
2019b). Four genotypes of VHSV are recognised, with several sub-lineages of varying
pathogenicity (OIE, 2019a).
14.1.2 OIE status
Infection with VHSV is listed by the OIE as a notifiable disease (OIE, 2019a).
14.1.3 New Zealand status
Active surveillance is undertaken for VHSV in New Zealand, but VHS has not been reported
from the Southern Hemisphere (Stone et al., 1997; DAFF, 2005, 2014; OIE, 2019a). It has not
been identified in New Zealand (OIE, 2019a), and it is exotic. Infection with VHSV is a
notifiable disease in New Zealand (Anon., 2016).
14.1.4 Zoonotic disease
VHSV is not zoonotic (OIE, 2019a).
14.1.5 Epidemiology
VHS is one of the most economically important diseases affecting marine and freshwater fish
(OIE, 2019a). VHSV strains in European freshwater aquaculture appear to have been transferred
from northern Pacific and Atlantic oceanic waters through marine fish used as feedstock for
aquaculture (Hedrick et al., 2003; Phelps et al., 2014). Management of salmonid fisheries in the
USA now includes movement restrictions on bait fish species as a disease control measure (Skall
et al., 2005a, 2005b; Bain et al., 2010; Phelps et al., 2013, 2014).
Distribution and host range
VHSV is associated with over 44 families of marine and freshwater fish (Blackwell, 2019), but
may potentially affect all temperate marine species (EFSA, 2008; OIE, 2019b; Anon., 2017a). It
also remains viable in filter-feeding oysters (Crassostrea gigas) and mussels (Mytilus edulis) in
Korea (Choi et al., 2015) without clinical signs for up to 7 days. Extracts from the mollusc
digestive gland failed to initiate infection (by intraperitoneal injection) in olive flounder
(Paralichthys olivaceus), suggesting the likelihood of infection of fish through this pathway is
low (Kim et al., 2017).
VHSV is essentially a cold-water pathogen, occurring where temperatures do not exceed 15 °C,
but it has also been introduced into Iranian and Kuwaiti aquaculture (Hedrick et al., 2003; Tubbs
et al., 2007; Amos et al., 2010; Spikler, 2010; OIE, 2019b). New Zealand oceanic water

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 81
temperatures vary between 17 °C in the north, to 12 °C in southern waters (Pinkerton et al.,
2018).
The species associated with VHSV in fish bait and their geographic/pathogen range are given in
Table 14 (Meyers et al., 1994, 1995; Traxler & Kieser, 1994; Meyers & Winton, 1995; Kocan et
al., 1997; Dixon et al., 1997; Marty et al., 1998; Hershberger et al., 1999; Mortensen et al.,
1999; Meyers et al., 1999; Traxler et al., 1999; Kocan et al., 2001; Biosecurity Australia, 2002;
Hedrick et al., 2003; Goodwin et al., 2004; Skall et al., 2005a, 2005b; Lee et al., 2007; Herve-
Claude et al., 2008; Kim & Faisal, 2010; Crane & Hyatt, 2011; Diggles et al., 2011; Purcell et
al., 2012; Kim et al., 2013a; Phelps et al., 2013; Ogut & Altuntas, 2014; Sandlund et al., 2014;
CIFA, 2019; OIE, 2019a).
Table 14. Fish bait species susceptible to viral haemorrhagic septicaemia virus (VHSV)
Family Host species Host geographical range1 Pathogen range2
Clupeidae Atlantic herring (Clupea harengus)
East, West Atlantic, Black Sea, Europe
Northeast, Northwest Atlantic (Canada, Europe, USA)
Bali sardinella (Sardinella lemuru), Indian oil sardine (Sardinella. longiceps)
North, East, West Indian Ocean, Indo-Northeast Indian Ocean
Indo-Pacific, Northwest, Central Pacific, Northeast Indian Ocean
Pacific herring (Clupea pallasii pallasii), Spotted sardinella (Amblygaster sirm),
North, West, Central Pacific Northeast Pacific (USA)
South American pilchard (Sardinops sagax)
Pacific Northeast Pacific (Canada)
Round sardinella (Sardinella aurita)
East, West Atlantic Northeast, Northwest Atlantic
Mugilidae Flathead grey mullet (Mugil cephalus)
Northwest Pacific (Korea), Northeast Pacific (Southern California)
Northwest, Northeast Pacific (Canada, Japan, South Korea, USA)
Scombridae Chub mackerel (Scomber japonicus)
Indo-Pacific, Eastern, Western Pacific
Northeast Pacific (Korea), Northwest Pacific (Canada, USA)
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature). VHSV is reported from the North Atlantic Ocean (English Channel), Baltic
Sea, Northwest Pacific Ocean (Japan, South Korea), and Eastern Pacific Ocean (from Alaska to California). It has also been introduced into Iranian and Kuwaiti aquaculture (Hedrick et al., 2003; Tubbs et al., 2007; Amos et al., 2010; Spikler, 2010; OIE, 2019b).
The prevalence in wild Pacific herring (C. p. pallasii) varies from 0 to 4-8% (Hedrick et al.,
2003; Arkush et al., 2006), where viral titre in apparently healthy fish is 1,000 to 10,000 times
lower than from fish with active disease (Meyers et al., 1999). Epizootics in wild Pacific herring
are reported in Alaska (Meyers et al., 1999) and in Pacific sardine (S. sagax) in British
Columbia, and Canada (Hedrick et al., 2003).
Potential host species in New Zealand
A wide range of potential marine and freshwater hosts are present in New Zealand (Tubbs et al.,
2007).

82 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogenicity
VHSV causes mortalities ranging from 20% to 100% in marine and freshwater aquaculture.
Pathogenicity in wild fish is variable, but subclinically infected Pacific herring (C. p. pallasii)
are known to shed substantial quantities of virus. Wild fish species may act as reservoirs of
disease (Hershberger et al., 1999).
Mortality is temperature related (OIE 2019a). Experimental infection of salmonids, by
cohabitation with infected Pacific herring (C. p. pallasii) in conditions approximating natural
waters (at 10 °C) resulted in mortality of 81.5% (Gross et al., 2019). Similar experiments
undertaken at higher temperatures (20 °C and above) failed to initiate clinical disease (Arkush et
al., 2008).
Clinical signs
External clinical signs in salmonids include exophthalmia and widespread haemorrhages of the
skin, fin bases and eyes. However, clinically infected fish may show no external signs where
temperatures exceed 5 °C (Munday, 2002). Subclinical infection may progress with no external
signs (Arkush et al., 2006).
Pathogen survival
From experimental data, VHSV remains viable for up to 10 days in sediment at 4 °C (Afonso et
al., 2012; DAFF, 2005). Viability in marine waters varies from up to 13 days at 4 °C, to 1.5 days
at 20 °C). Viability in fresh water varies (from up to 40 days at 4 °C, to less than 1 day at 30 °C).
Infection in wild stocks is therefore considered unlikely where water temperatures exceed 18 °C
(Arkush et al., 2006; Hawley & Garver, 2008). VHSV is sensitive to UV radiation (Oye &
Rimstad, 2001). Viability in natural waters may also be influenced by dilution and predation
factors (see section 6.1.2).
Disease transmission
Transmission is horizontal and direct, through the water column (Arkush et al., 2005).
VHSV can be transferred through frozen fish bait (Phelps et al., 2013), although prevalence in
wild stocks is likely to be low (from 0 to 4-8%) (Arkush et al., 2006). VHSV can be dispersed by
fomites, through meltwater or discharges from commercial processing, ballast water discharges,
or by mechanical transfer through piscivorous birds (DAFF, 2005; Bain et al., 2010; Bowser et
al., 2010).
Frozen sardines (Clupea p. pallasii) from California have been imported as bait into South
Australia without the introduction of VHS. This has been attributed to: the low prevalence of
VHSV in wild fish stocks; the effect of medium-term frozen storage; and the temperature of the
receiving waters being higher than the threshold (18 °C) necessary for VHSV establishment
(DAFF, 2002; Arkush et al., 2006; Herve-Claude et al., 2008; Anon., 2010; Pinkerton et al.,
2018).
Infection and disease progression
Pathogen entry occurs through the epithelial cells of the gills, viscera or skin. Fish bait species
(such as Pacific herring) swim in dense shoals and the infectious dose is low (101.5 pfu mL-1)
(DAFF, 2005). From experimental data, infection may be initiated by cohabitation in healthy fish
after only 1 hour of water borne exposure (Kocan et al., 1997; Hershberger et al., 2007; Hawley
& Garver, 2008). The viral titre in infected South American pilchard (Sardinops sagax) ranged
from 1.26 × 105 to 1.56 × 108 pfu g–1 of tissue (Arkush et al., 2006).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 83
Disease may follow several alternative pathways, depending upon the VHSV strain and the host
species (DAFF, 2005; OIE, 2019a). Infection may be immediate and focussed at the point of
entry or be delayed until viral particles reach the endothelial cells of the vascular tissues, brain or
spleen (DAFF, 2005).
In acute infection, disease initially infects the haemopoietic tissues. It then spreads to the major
organs and musculature, causing extensive necrosis. Infection may also spread to the brain and
neural tissues (OIE, 2019a).
Chronic infection progresses slowly, affecting all major organs, usually with no external signs
(DAFF, 2005). Surviving fish are carriers and continue to shed viral particles through urine,
faeces, mucous and sexual fluids, for months or years (DAFF, 2005).
Homogenates derived from infected fish are infectious to a wide range of species (Oidtmann et
al., 2011). Strains isolated from European marine fish are of low virulence for Atlantic salmon
(Salmo salar) (Dale et al., 2009), but strains from Pacific herring are virulent to salmonids.
Caution is advised before assuming a species is resistant to infection (Skall et al., 2005a, 2005b).
Inactivation
Short-term frozen storage (at -20 °C for 16 hours) is only partially effective against VHSV
(Wolf, 1988; Arkush al., 2006; Phelps et al., 2014; OIE, 2019a). Viral titre is reduced (by up to
90%) following repeated freeze-thaw cycles (Meyers et al., 1994). Studies approximating
commercial frozen storage of sardine Sardinops sagax used as bait indicated VHSV was
completely inactivated after 50 days (7 weeks) frozen storage (at -20 °C) (Arkush et al. 2003).
However, other studies indicate sufficient VHSV may remain viable in thawed fish tissues to
initiate infection where viral titre is high (Meyers at al., 1994; Arkush et al., 2006; Herve-Claude
et al., 2008; Phelps et al., 2013). VHSV is also inactivated where the receiving water
temperature exceeds 18 °C (Herve-Claude et al., 2008; Anon., 2010) and both frozen storage and
the receiving water temperature are recognised as significant risk factors (Anon., 2010). VHSV
is therefore assumed to be substantially, but not completely inactivated by medium-term frozen
storage (for 4 months at -20 °C).
VHSV is assumed to be denatured by ionising radiation (at 50 kGy) of frozen fish tissue (DAFF,
2013).
14.2 Risk assessment
14.2.1 Entry assessment
VHSV may be present in the commodity (Table 14). While Pacific herring was not imported
during 2018, clupeids represented over 50% (by volume) of the finfish imported as bait in 2018
(Table 3).
Prevalence in wild stocks is low (from 4-8%) in wild Pacific herring (C. p. pallasii) except
during epizootics (Hedrick et al., 2003; Arkush et al., 2006), but VHSV occurs in both Pacific
herring (C. p. pallasii) and South American pilchard (S. sagax) (Meyers et al., 1999).
VHSV may be present in carrier fish (Hershberger et al., 1999; Hedrick et al., 2003; Arkush et
al., 2006; Oidtmann et al., 2011; Phelps et al., 2013). VHSV may also be introduced through
meltwater and water used in processing fish bait (OIE, 2019a). Viability is substantially reduced
following frozen storage (Arkush et al., 2006).

84 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
VHSV may remain viable in the commodity. However, prevalence is generally low in wild fish
bait stocks and viability is substantially reduced following frozen storage (Arkush et al. 2006;
Herve-Claude et al., 2008), The likelihood of entry is assessed to be low.
14.2.2 Exposure and establishment assessment
VHSV is recognised as a major risk organism in the international fish bait trade (Biosecurity
Australia, 2002; Dalton, 2004; Goodwin et al., 2004; Arkush et al., 2006; Anon., 2010;
Oidtmann et al., 2011; Phelps et al., 2013, 2014). VHSV remains viable in marine and fresh
water, or in sediments for extended periods, where water temperatures do not exceed 18 °C
(DAFF, 2005; Arkush et al., 2006; Herve-Claude et al., 2008; Hawley & Garver, 2008; Afonso
et al., 2012).
The infectious dose is low (101.5 pfu mL-1) (DAFF 2005) and VHSV has a wide host range (OIE,
2019a). Wild fish species may act as reservoirs of disease (Hershberger et al.,1999) and infection
may progress without clinical signs of disease (Arkush et al., 2006).
VHSV can be transferred through frozen fish bait (Herve-Claude et al., 2008; Anon., 2010;
Phelps et al., 2013). It is substantially inactivated by long-term storage (at -20 °C for at least 50
days) (Arkush et al., 2006; Herve-Claude et al., 2008; Anon., 2010), but sufficient VHSV may
remain viable where viral titre is high to establish infection where the temperature of the
receiving water is less than 18 °C (Arkush et al., 2006; Herve-Claude et al., 2008; Hawley &
Garver, 2008), such as Chile or New Zealand (Stone et al., 1997; Herve-Claude et al., 2007;
Pinkerton et al, 2018).
The use of frozen menhaden (Brevoortia tyrannus) as feed in Australian aquaculture is restricted
to marine waters where temperatures exceed 14 °C, due to concerns about VHSV infection
(Diggles, 2007). Movement controls (restricting bait to the water body in which it was caught)
have been placed on bait in the continental USA, while additional processing is also required for
imported fish bait in some US States (Phelps et al., 2013, 2014).
The likelihood of exposure and establishment through the commodity is assessed as low.
14.2.3 Consequence assessment
Economic consequences
Infection with VHSV is an OIE-listed disease primarily affecting salmonids (OIE, 2019a). Its
establishment would result in loss of market access for New Zealand, with significant direct
economic consequences. It is highly pathogenic, with mortalities up to 100% in salmonid
aquaculture (OIE, 2019a).
Other consequences of establishment include reduced productivity and increased mortality, as
Chinook salmon (O. tshawytscha) is particularly susceptible to VHSV (OIE, 2019a). New
Zealand salmonid aquaculture of Chinook salmon (O. tshawytscha) was worth $77 million in
export earnings in 2018 (Seafood New Zealand, 2019). Further indirect economic costs would
include fallowing and destocking, as well as disinfection of farms and processing equipment in
the supply chain.
VHSV has a wide host range in marine and fresh waters (OIE, 2019a, 2018b; Blackwell, 2019).
Wild stocks may represent reservoirs of infection for other farmed species (Hershberger et al.
1999). Establishment may affect the developing aquaculture of brill (Colistium guntheri) and

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 85
turbot (C. nudipinnis) (Pleuronectidae), hapuku/grouper (Polyprion oxygeneios) (Polyprionidae),
snapper (Sparus aurata) (Sparidae) and yellowtail kingfish (Seriola lalandi) (Carangidae).
Several wild fisheries may also be affected in marine and fresh waters (Diggles, 2003. 2004a,
2004b; Hickman & Tait, 2008; Symonds et al., 2014; NIWA, 2017b; Plant & Food, 2017; MPI,
2018a, 2018b, 2018c). For example, the snapper export fishery was valued at $33 million in
2018 (Seafood New Zealand, 2019).
Social consequences
VHSV is not zoonotic (OIE, 2019a). It has a wide host range, including many coastal marine fish
species (Tubbs et al., 2007). While effects on wild fish species are uncertain (Lafferty et al.,
2015), establishment is likely to result in lowered productivity and increased mortality for
susceptible species of significance for customary and recreational fishing (Wynn-Jones et al.,
2014, 2019; MPI, 2018a, 2018b, 2018c).
Recreational fishing is the fifth most popular recreational activity for adults, involving 700,000
marine fishing trips annually, with a contribution of NZ$ 638 million to the GDP (Gross
domestic product) in 2015 (Southwick et al., 2018). The snapper, hapuku and trevally
recreational fisheries contributed NZ$ 402 million, NZ$ 12.5 million and NZ$ 71 million
respectively to the GDP in 2015 (Southwick et al., 2018).
Environmental consequences
VHSV is highly pathogenic and potentially affects all temperate fish species (OIE, 2019b).
Establishment may result in lowered productivity and increased mortality in a wide range of
marine and freshwater fish species, many of which are considered stressed (MPI, 2018a, 2018b,
2018c). While effects of introduced viruses on wild fish stocks are uncertain (Lafferty et al.,
2015), VHSV has caused epizootics in Pacific herring (C. p. pallasii) and in South American
pilchard (S. sagax) (Meyers et al., 1999; Hedrick et al., 2003).
Inshore fish species including clupeids and mugilids provide a significant food source for other
fish, seabirds and marine mammals and are a major link between trophic levels in the aquatic
food web (Dunn et al., 2012). Wild fish may also act as reservoirs of disease (Hershberger et al.,
1999).
The establishment of VHSV is likely to have significant direct and indirect economic, social and
environmental effects for New Zealand. Therefore, the consequences of establishment are
assessed to be high.
14.2.4 Risk estimation
The entry and exposure assessments for VHSV are low, but the consequences are assessed to be
high. The risk estimate is assessed to be medium. Under the procedures followed in this risk
analysis, risk management measures may be justified.
14.3 Risk management
VHSV has been assessed as a risk in the commodity. VHS is an OIE-listed disease, so the OIE
Aquatic Code and Aquatic Manual provide some guidance to avoid the spread of VHSV (OIE,
2019a, 2019b). As the OIE-recommended heat treatment (OIE 2019a) would render the
commodity unfit for purpose as fish bait, this option is not considered further.

86 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
VHSV is associated with fish bait (Table 14). Short-term frozen storage (for 168 hours) is
unlikely to be effective in denaturing VHSV (Arkush et al., 2006; Phelps et al., 2013). Medium-
term frozen storage (-18 °C to -20 °C) for at least 4 months (16 weeks) should denature most
VHSV, but sufficient virus may remain present to initiate infection where viral titre is high
(Arkush et al., 2006; Herve-Claude et al., 2008; Anon., 2010).
The VHSV Working Group recommended batch testing of frozen fish bait (Anon., 2010). When
combined with medium-term frozen storage (see section 6.2.1), batch testing should substantially
reduce the risk associated with VHSV and be a viable risk management option.
Treatment of the commodity using ionising radiation (at 50 kGy) (DAFF, 2013) should
substantially reduce the VHSV risk and be a viable risk management option.
14.3.1 Risk management options.
In addition to the general risk management measures that are proposed for all imported fish bait
(section 6.2.1), one or more of the following risk management options would reduce the VHSV
specific risk to an acceptable level:
Option 1
• Competent Authority attestation that the commodity does not include Atlantic herring
(Clupea harengus), Bali sardinella (Sardinella lemuru), Indian oil sardine (Sardinella.
longiceps), Pacific herring (Clupea pallasii pallasii), spotted sardinella (Amblygaster sirm),
South American pilchard (Sardinops sagax), round sardinella (Sardinella aurita)
(Clupeidae); flathead grey mullet (Mugil cephalus) (Mugilidae); and Chub mackerel
(Scomber japonicus) (Scombridae) should substantially reduce the IHNV risk, so the
commodity could be imported without any further restrictions.
Option 2
• Acceptance of region/country freedom through the BNZ Country Approval Procedures
should substantially reduce the VHSV risk so the commodity could be imported without
any further restrictions.
Option 3
• A negative test result from a shipment of the commodity, following approved sampling and
diagnostic procedures through the BNZ Country Approvals Procedures, should substantially
reduce the VHSV risk, so the commodity could be imported without further restrictions.
Option 4
• Treatment with ionising radiation (at 50 kGy) should substantially reduce the VHSV risk, so
the commodity could be imported without any further restrictions.
Edwardsiella spp.
15.1 Technical review
15.1.1 Aetiological agent
Edwardsiella spp. are a species complex of Gram-negative rod-shaped filamentous bacteria
classified in the Family Enterobacteriaceae (Farmer et al., 1984, 1985). Five genetically distinct

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 87
but phenotypically ambiguous species are recognised (E. ictaluri, E. tarda, E. piscida, E.
hoshinae and E. anguillarum) with high intergenomic heterogeneity (Reichley et al., 2017;
Katharios et al., 2019). They vary widely in their responses to environmental parameters
(salinity, temperature, and pH), host specificity, geographical range, specific target organs, drug
resistance and pathogenicity (Park et al., 1983; Reichley et al., 2017).
High resolution phylogenetic methods, such as multilocus sequence analysis (MLSA),
multilocus sequence typing (MLST) and whole genome sequencing (Shao et al., 2015) are
necessary to identify species within the species complex (Katharios et al., 2019). For the
purposes of this risk analysis, these species are considered together as Edwardsiella spp.
15.1.2 OIE status
Infection with Edwardsiella spp. is not notifiable to the OIE (OIE, 2019a).
15.1.3 New Zealand status
Edwardsiella spp. are reported from migratory seabirds and cetaceans that inhabit Antarctic and
Sub-Antarctic waters and may occasionally enter New Zealand waters (Sakazaki & Tamura,
1975; Shao et al., 2015; EPA, 2015; Tindall, 2016).
Edwardsiella spp. have not been reported from fish in Antarctic and Sub-Antarctic waters (S.
Fenwick, Massey University, reported in Stone et al., 1997; Leotta et al., 2009; Cools et al.,
2013). They have not been reported from New Zealand marine or freshwater fish (Johnston,
2008; M. Bestbier, BNZ, pers. comm., 2020). Therefore, for the purposes of this risk analysis,
Edwardsiella spp. are assumed to be exotic. Infection with Edwardsiella spp. is not a notifiable
disease in New Zealand (Anon., 2016).
15.1.4 Zoonotic disease
Edwardsiella spp. are zoonotic. Bacterial gastroenteritis may result from eating undercooked
aquatic products. Wound infections associated with fishing and fish processing can cause
cellulitis and gas gangrene (Jordan & Hadley, 1969; Nagel et al., 1982). Infection is rarely
serious but may progress to systemic conditions including meningitis, septicaemia and
osteomyelitis (Janda et al., 1993; Hirai et al., 2015).
15.1.5 Epidemiology
Distribution and host range
Edwardsiella spp. are reported from marine and freshwater invertebrates, fish, amphibians,
reptiles and marine mammals. They also occur in oceanic birds, as well as terrestrial animals
including cattle and swine (Blackwell, 2019). Edwardsiella spp. have a wide geographical
distribution (Table 15).
The species associated with Edwardsiella spp. in fish bait and their geographic /pathogen range
are given in Table 15.

88 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Table 15. Fish bait species susceptible to Edwardsiella spp.
Family Host species Host geographical range1 Pathogen range2
Clupeidae Indian oil sardine (Sardinella longiceps)
Indo-Pacific Indo-Pacific (India)
Mugilidae Flathead grey mullet (Mugil cephalus)
Cosmopolitan in coastal waters of the tropical, subtropical and temperate waters of all seas.
Northwest Pacific (Japan)
Notes 1 Host geographical range (from FishBase, 2019) 2 Pathogen geographical range (from literature). Edwardsiella spp. are widely distributed, in over 30 countries, as well as from
the Antarctic, Atlantic, Indian and Pacific Oceans (Leotta et al., 2009).
Potential host fish species in New Zealand
A wide range of potential host fish species are present in New Zealand marine and freshwaters,
including members of Carangidae (yellowtail kingfish, Seriola lalandi), Pleuronectidae (turbot,
Colistium nudipinnis), Salmonidae (Oncorhynchus spp., Salmo spp., Salvelinus spp.), Sparidae
(snapper, Pagrus (Chrysophrys) auratus) and Polyprionidae (hapuku, Polyprion spp.) (Tubbs et
al., 2007; Blackwell, 2019).
Pathogenicity
Pathogenicity in farmed Chinook salmon (O. tshawytscha) varies from 5% to 15%. The lethal
dose (LD50) is low, estimated at 4.1 × 106 cells for chinook salmon (Park et al., 2012) and 1 ×
105 cells for gilthead seabream (Sparus aurata) (Baya et al., 1997).
Edwardsiella spp. are generally regarded as opportunistic pathogens of lower pathogenicity in
wild fish stocks (Amandi et al., 1982; Park et al., 2012). Epizootics occur in grey mullet (Mugil
cephalus) in Japanese coastal waters (Kusuda et al., 1976).
Clinical signs
External signs may include white skin patches and cutaneous lesions, but disease expression
varies widely between pathogen strains and among host species. Infection may also progress
with no external clinical signs (Evans et al., 2011).
Pathogen survival
Edwardsiella spp. are environmentally resistant. From experimental data, they remain viable in
marine or fresh waters up to 76 days, over a wide temperature range (from 45 °C to Antarctic
waters). They remain viable on dead and moribund fish tissues and survive in sediments in a
viable but not culturable (VBNC) state for extended periods (Evans et al., 2011). However, the
length of survival under natural conditions is unknown (see section 6.1.2).
Disease transmission
Disease transmission is horizontal and direct, through the water column (Amandi et al., 1982).
Edwardsiella spp. may be dispersed by fomites, through contaminated water, aerosol
transmission, or by piscivorous birds (Johnston, 2008; Evans et al., 2011).Edwardsiella spp. are
opportunistic pathogens with a wide host range and may become ubiquitous in endemic areas
(Buller, 2014; Wiedenmayer et al., 2006; Harai et al., 2015).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 89
Infection and disease progression
Edwardsiella spp. are significant bacterial pathogens affecting farmed marine and freshwater
fish of all ages (Plumb & Hansen, 2011). From experimental infection per os, the infectious dose
is very low (from 1 × 101 to 9 × 107 cells) for channel catfish (Ictalurus punctatus) (Park et al.,
2012). Edwardsiella spp. infection in aquaculture is usually associated with poor rearing
conditions including high water temperature, poor water quality and high organic content (Meyer
& Bullock, 1973; Rashid et al., 1994; Evans et al., 2011).
Disease progression may follow a renal or hepatic pathway. In renal infection, micro-abscesses
form in kidney tissues. In hepatic infection, large abscesses develop in the liver, with extensive
tissue liquefaction (Miyazaki & Egusa, 1976; Evans et al., 2011). In both pathways, infection
becomes systemic, infecting the spleen, liver, heart and digestive tissue mucosa. Necrotic lesions
may also form in the lateral musculature (Miyazaki & Kaige, 1985).
Inactivation
Edwardsiella spp. are moderately cold-resistant. They survive frozen storage (up to 50 days
at -20 °C) in frozen fish tissues (Castro et al., 2006; Johnston, 2008), but are rendered non-viable
by longer-term frozen storage (at least 50 days) (Brady & Vinitnantharat, 1990). It is assumed
therefore that Edwardsiella spp. would be inactivated by medium-term frozen storage (at -20 °C
for 4 months).
Ionising radiation (at 50 kGy) is also likely to denature Edwardsiella spp. in frozen fish tissues
(DAFF, 2013).
15.2 Risk assessment
15.2.1 Entry assessment
Edwardsiella spp. are associated with the commodity (Table 15). Prevalence may reach 14% in
wild Indian oil sardine (Sardinella longiceps) (Clupeidae) in the Bay of Bengal during epizootics
(Kumar et al. 2014).
The prevalence in flathead grey mullet (Mugil cephalus) (Mugilidae) in Japanese waters is
unknown but assumed to be low (Kusuda et al., 1976; Evans et al., 2011). M. cephalus
represented 13% and S. longiceps represented less than 1%, respectively, by volume of finfish
bait imports from all countries in 2018 (see Table 3). Edwardsiella spp. strains remain viable in
dead and moribund fish tissues (Evans et al., 2011) and in fish bait frozen for up to 50 days
(Castro et al., 2006; Johnston, 2008). Infected fish may show no external signs of disease (Brady
& Vinitnantharat, 1990).
Edwardsiella spp. have a limited distribution where associated with fish bait and the prevalence
in healthy wild stocks is assumed to be low.
The likelihood of entry is assessed as low.
15.2.2 Exposure and establishment assessment
Edwardsiella spp. are non-obligate opportunistic pathogens. From experimental data, they
remain viable in a resistant VBNC state in the aquatic environment for extended periods, over a
wide temperature range (from 0 °C to 45 °C) (Evans et al., 2011; Park et al., 2012). Survival in
natural waters is unknown (see section 6.1.2).

90 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Infection occurs through the water column, by cross-contamination of working surfaces (where
fish bait is used on fishing vessels), through the discharge of contaminated meltwater (Johnston,
2008; Evans et al., 2011). Edwardsiella spp. may be dispersed from landfill or between
waterways by piscivorous birds (Evans et al., 2011).
Edwardsiella spp. are zoonotic pathogens and human exposure may occur through cross-
contamination or handling of infected fish bait (Janda & Sharon, 1993; Hirai et al., 2015).
The host range in fish bait species is small and the prevalence is low in healthy wild fish stocks.
The likelihood of exposure and establishment is assessed as low-medium.
15.2.3 Consequence assessment
Economic consequences
Edwardsiella septicaemia is not listed by the OIE (OIE, 2019a), so establishment would not
result in trade restrictions for New Zealand. However, Edwardsiella spp. have a wide host range
affecting farmed marine and freshwater fish worldwide. Infection results in lowered productivity
and increased mortality. Chronic infection causes tissue spoilage that makes the product
commercially unmarketable (Evans et al., 2011; CABI, 2015).
Establishment may result in serious economic consequences for farmed and fished species in
New Zealand, including salmonids. These include Chinook salmon (Oncorhynchus tshawytscha)
(Salmonidae) which was valued at $77 million in exports in 2018 (Seafood New Zealand, 2019).
However, the prevalence of clinical disease in farmed fish overseas is commonly related to over-
stocking and poor management practices (Evans et al., 2011).
Other potentially affected farmed species include yellowtail kingfish (Seriola lalandi)
(Carangidae), brill (Colistium guntheri) and turbot (C. nudipinnis) (Pleuronectidae) and gilthead
sea bream (snapper) (Sparus aurata) (Sparidae) (Diggles, 2003; Hickman & Tait, 2008; Mussely
& Goodwin, 2012; Sim-Smith et al., 2014; Symonds et al., 2014; NIWA, 2017a, 2017b; Plant &
Food, 2016, 2017). Establishment could also affect grass carp (Ctenopharyngodon idella) and
silver carp (Hypophthalmichthys molitrix) aquaculture for weed control in fresh waters (Clayton
& Wells, 1999; NIWA, 2014).
While prevalence of Edwardsiella spp. is generally low in wild fish stocks, epizootics occur in
wild flathead grey mullet (Mugil cephalus) (Evans et al., 2011). The establishment of
Edwardsiella spp. may result in reduced productivity, increased mortality and increased product
rejection in New Zealand coastal inshore fisheries (MPI, 2018a, 2018b, 2018c). For example, the
snapper export fishery was valued at $33 million in 2018 (Seafood New Zealand, 2019).
Social consequences
Edwardsiella spp. are zoonotic, generally causing minor bacterial gastroenteritis and cellulitis
through handling infected fish (Jordan & Hadley, 1969; Nagel et al., 1982). Serious infection is
rare, but may include meningitis, septicaemia and osteomyelitis (Janda et al., 1993; Hirai et al.,
2015).
Establishment of Edwardsiella spp. may affect freshwater aquaculture. This includes brown trout
(Salmo trutta) and rainbow trout (Oncorhynchus mykiss) farmed for recreational fishing (Fish &
Game, 2014). The Lake Taupo trout fishery alone is valued at NZ$ 70–80 million annually
(Marsh & Mkwra, 2013).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 91
The establishment of Edwardsiella spp. may affect other species of significance for customary
and recreational fishing in marine and freshwater, including eels (Anguilla spp.). These are
considered stressed by the level of current fishing activity (Wynn-Jones et al., 2014, 2019;
NIWA, 2017c). Eels (Anguilla spp.) are considered a taonga in Maori culture (MPI, 2018a,
2018b, 2018c)
Recreational fishing is the 5th most popular recreational activity for adults, involving 700,000
marine fishing trips annually, with a contribution of NZ$ 638 million to the GDP (Gross
domestic product) in 2015. Of this, snapper contributed NZ$236 million and trevally NZ$46
million to the GDP in 2015. These fisheries provided an estimated total of NZ$ 402 million and
NZ$ 71 million respectively, to the New Zealand economy in 2015 (Holdsworth et al., 2016;
Southwick et al., 2018).
Environmental consequences
Edwardsiella spp. are opportunistic pathogens of generally low pathogenicity in wild fish stocks
(Park et al., 2012). The infectious dose is very low (Park et al., 2012) and the host range includes
20 families of marine and freshwater fish (Evans et al., 2011; Blackwell, 2019). Edwardsiella
spp. may become endemic, with reported prevalence of up to 60% in wild freshwater fish
(Vandepitte et al., 1980). Wild fish may also function as reservoir hosts for human and animal
infection, including aquaculture (Vandepitte et al., 1980; Evans et al., 2011; Cools et al., 2013).
Edwardsiella spp. are opportunistic, zoonotic pathogens with a wide host range. While of low
pathogenicity in wild fish stocks, they may become significant pathogens of farmed fish,
particularly when aquaculture occurs under sub-optimum conditions.
The consequences of establishment are assessed as low.
15.2.4 Risk estimation
As the entry, exposure and consequence assessments are low, or low-medium, the risk estimate
is for Edwardsiella spp. is assessed to be low. Under the procedures followed in this risk
analysis, risk management measures may be justified.
15.3 Risk management
Edwardsiella spp. have been identified as risk organisms in previous aquatic risk analysis
(Johnston, 2008). Infection with Edwardsiella spp. is not an OIE-notifiable disease, so the OIE
Aquatic Code (OIE, 2019a) provides no specific guidance or processing requirements that would
ensure the destruction of the pathogen.
Edwardsiella spp. are associated with fish bait (Table 15). Edwardsiella spp. are unlikely to be
denatured by short-term frozen storage (at -20 °C for 168 hours). They are likely to be
substantially or completely denatured by medium-term frozen storage (to -20 °C for at least 4
months) (Brady & Vinitnantharat, 1990). An extended period of frozen storage is consistent with
current fish bait industry practice (B. Burney, pers. comm., 2020). Adoption of this general risk
management option should substantially reduce the risk associated with Edwardsiella spp. and
be a viable risk management option.
15.3.1 Risk management options
If the general risk management measures proposed for all imported fish bait (section 6.2.1) are
adopted, including medium-term frozen storage (at -18 to -20 °C for four months), then no

92 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
pathogen-specific risk management measures would be necessary to reduce the risk associated
with Edwardsiella spp. to an acceptable level.
Francisella spp.
16.1 Technical review
16.1.1 Aetiological agent
Francisella spp. represent a species complex of rickettsia-like intracellular bacteria classified in
Family Francisellaceae, within the Proteobacteriaceae (Arkush & Bartholomew, 2011). The host
relationships and distribution within this species complex are unclear (Ottem et al., 2009; Arkush
& Bartholomew, 2011; Colquhoun & Duodu, 2011). Strains of Francisella piscida (= F.
noatunensis) associated with finfish include F. noatunensis orientalis in warm water fish species
and F. noatunensis noatunensis in cold water fish species (Colquhoun & Duodu, 2011).
16.1.2 OIE status
Infection with Francisella spp. is not listed by the OIE (OIE, 2019a).
16.1.3 New Zealand status
Francisella spp. are considered exotic (Johnston, 2008) and have not been reported from New
Zealand. Infection with Francisella spp. is not a notifiable disease in New Zealand (Anon.,
2016).
16.1.4 Zoonotic disease
Francisella spp. affecting fish are not considered zoonotic (Colquhoun & Duodu, 2011).
16.1.5 Epidemiology
Distribution and host range
Francisella spp. are reported from 9 families of marine and freshwater fish and invertebrates
(Johnston, 2008; Colquhoun & Duodu, 2011; Blackwell, 2019). They have a wide geographical
distribution (Table 16).
The species associated with Francisella spp. in fish bait and their geographic /pathogen range are
given in Table 16. Prevalence in wild Atlantic cod (Gadus morhua) reached 20% off the
Swedish coast in 2004 (Alfjorden et al., 2006), while lower levels of prevalence (from 7-11%)
were reported in Atlantic cod from coastal Norwegian waters (Ottem et al., 2008).
Francisella spp. is also reported at a prevalence of 11%, in Atlantic mackerel (S. scombrus) from
areas immediately adjacent to infected Atlantic cod farms. However, these prevalence data were
determined from a very small sample size (n=9 fish) The bacterial load in Atlantic mackerel was
much lower than for infected Atlantic cod (Colquhoun & Duodu, 2011).
Strains of Francisella spp. may be host specific. Given the small sample size available for
Atlantic mackerel (Ottem et al., 2008), the significance of this infection is difficult to estimate
(Colquhoun & Duodu, 2011).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 93
Table 16. Fish bait species susceptible to Francisella spp.
Family Host species Host geographical range1 Pathogen range2
Scombridae Atlantic mackerel (Scomber scombrus)
North Atlantic, Mediterranean Sea
Northeast Atlantic (Europe)
Notes 1 Host geographical distribution (from FishBase 2019) 2 Pathogen geographical range (from literature). Francisella spp. are reported from the East, South and West Atlantic (Central
and South America, Norway, United Kingdom, United States), and the North, Central and South Pacific (Asia, China, Hawaii, Central and South America) (Colquhoun & Duodu, 2011).
Potential fish host species in New Zealand
Potential hosts in New Zealand include Atlantic salmon (Salmo salar) and brown trout (S.
trutta), as well as mackerel (Scomber spp.) and flatfishes (Order Pleuronectiformes) (Colquhoun
& Duodu, 2011).
Pathogenicity
Francisella spp. infects farmed Atlantic cod (G. morhua) with mortalities up to 40%.
Subclinically infected wild fish, including Atlantic mackerel (S. scombrus), have been recovered
adjacent to Atlantic cod aquaculture in the Northeast Atlantic, since the 1980s (Ottem et al.,
2008). The role of these fish as reservoir hosts for farmed species is unclear (Colquhoun &
Duodu, 2011).
Clinical signs
Clinically infected fish may have darkened skin due to extensive haemorrhages of the dermal
musculature. Subclinical infection usually progresses with mild or no clinical signs (Nylund &
Ottem, 2006; Colquhoun & Duodu, 2011).
Pathogen survival
Francisella spp. may enter a VBNC state. Experimental data indicate they may survive for up to
30 days at 4 °C, and up to 16 days at 12 °C in marine and fresh waters, with longer survival in
sediment (Ottem et al., 2009; Colquhoun & Duodu, 2011). However, survival time in natural
waters is unknown and may be influenced by other factors (see section 6.1.2).
Disease transmission
Transmission is horizontal and direct, through the water column, but the exact mechanisms are
unclear (Colquhoun & Duodu, 2011). Francisella spp. may be dispersed by fomites including
fish bait processing equipment (Arkush & Bartholomew, 2011).
Infection and disease progression
Francisella spp. are considered emerging pathogens causing systemic, chronic, granulomatous
infections in wild and farmed fish (Colquhoun & Duodu, 2011).
Experimental data suggest the infectious dose in intracellular bacteria ranges from 10 cfu fish-1
(Nylund & Ottem, 2006; Colquhoun & Duodu, 2011), to 105 TCID50 ml-1 (Almendras et al.,
1997; Birbeck et al., 2004; Nylund et al., 2006). It is likely that a relatively high dose would be
necessary to induce infection with Francisella spp. (Johnston, 2008).
Pathogen entry occurs through the skin or viscera. Different strains target different organ
systems of the host. While clinical disease is associated with extensive necrotic granulomas of

94 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
the skin, mouth, gill and nasal tissue, the pathogenic mechanisms remain unclear (Arkush &
Bartholomew, 2011; Colquhoun & Duodu, 2011).
Inactivation
Francisella spp. remain viable after prolonged frozen storage (at -80 °C) (Colquhoun & Duodu,
2011). Related species survive for up to 75 days in frozen sheep muscle stored at -20 °C
(Johnston, 2008). Francisella spp. are unlikely therefore, to be affected by either short (168
hours) or medium-term (4 months) frozen storage (to -20 °C).
Francisella spp. are assumed to be denatured by ionising radiation (at 50 kGy) (DAFF, 2013).
16.2 Risk assessment
16.2.1 Entry assessment
In fish bait, Francisella spp. occur rarely in wild Atlantic mackerel (Scomber scombrus) with no
clinical signs. The prevalence estimates of 10% are based on a small sample size (n=9 fish) and
infection only occurs in fish taken immediately adjacent to Atlantic cod (G. morhua) fish farms
previously infected with Francisella spp. (Ottem et al., 2009). The significance of this infection
is unknown but is assumed to be minor (Colquhoun & Duodu, 2011).
Commercial fishing is a large-scale operation (FAO, 2014) and Atlantic mackerel represented
less than 3% of imported finfish bait in 2018 (Table 3). The likelihood of entry of Francisella
spp. through fish bait is therefore assessed to be so low as to be negligible.
16.3 Risk management
As the entry assessment is negligible, Francisella spp. is assessed as not representing a risk in
the commodity. No further risk assessment is necessary.
Pseudomonas anguilliseptica
17.1 Technical review
17.1.1 Aetiological agent
Pseudomonas anguilliseptica is a Gram-negative, rod-shaped motile bacterium, classified in
Family Pseudomonadaceae. Three main serotypes are recognised (I-III), with high phenotypical
homogeneity (Lopez-Romalde et al., 2003). Each serotype includes several isolates reported
from a wide range of marine and freshwater fish hosts and across geographical areas (Daly &
Akoi, 2011).
17.1.2 OIE status
Infection with P. anguilliseptica is not listed by the OIE (OIE, 2019a).
17.1.3 New Zealand status
P. anguilliseptica has not been reported from New Zealand and is considered exotic (Duignan et
al., 2003). Infection with P. anguilliseptica is not a notifiable disease in New Zealand (Anon.,
2016).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 95
17.1.4 Zoonotic disease
P. anguilliseptica is not zoonotic (Haenen, 2017).
17.1.5 Epidemiology
P. anguilliseptica is a significant emerging pathogen, causing “Winter disease” in farmed fish
(Daly, 1999; Daly & Akoi, 2011; Wheeler, 2012; Blackwell, 2019).
Distribution and host range
P. anguilliseptica infection is reported from over 17 families of wild and farmed marine and
freshwater fish (Blackwell, 2019). It has a wide geographical distribution (Table 17).
The species associated with P. anguilliseptica in fish bait and their geographic/pathogen range
are given in Table 17.
Table 17. Fish bait species susceptible to Pseudomonas anguilliseptica
Family Host Species Host geographical range1 Pathogen range2
Clupeidae Atlantic herring (Clupea harengus)
Atlantic Northeast Atlantic (Europe)
Notes 1 Host geographical range (from FishBase 2019) 2 Pathogen geographical range (from literature). P. anguilliseptica is reported from the North Atlantic and Mediterranean Sea
(Canada, Denmark, Egypt, Finland, France, Portugal, Spain and the United Kingdom), the Indo-Pacific and Northern Pacific Ocean (Australia, India, Japan, Malaysia and Chinese Taipei). It is also reported from deep sea microbial and hydroid communities in oceanic waters off Fiji (Berthe et al., 1995; Daly, 1999; Balboa et al., 2003; Romanenko et al., 2008; Wheeler, 2012; Andree et al., 2013; Mastan, 2013).
Prevalence in farmed fish varies from 12% (in salmonids), to 83% (in seabream, Sparus aurata)
(Wiklund & Lonnstrom, 1994; Fadel et al., 2018). Prevalence in wild Atlantic herring (C.
harengus) from coastal Finnish waters may reach 50% in epizootics (Lonnstrom et al., 1994)
(Table 17).
Infected wild fish may function as reservoir hosts (Daly & Akoi, 2011). P. anguilliseptica
infection has been transferred to farmed fish by herrings (Clupea spp.) (Clupeidae) when used as
aquaculture feed (Nash et al., 1987).
Potential fish host species in New Zealand
Potential hosts in New Zealand include jack mackerel (Trachurus sp.) (Carangidae), herrings
(Clupea spp.) (Clupeidae), salmonids (Oncorhynchus spp., Salmo spp., Salvelinus spp.), and
snapper (Pagrus auratus) (Sparidae) (Blackwell, 2019).
Pathogenicity
P. anguilliseptica is an opportunistic pathogen of wild fish (Lonnstrom et al., 1994). It causes
epizootic infections in a wide range of wild and farmed species (Daly & Akoi, 2011). Mortality
in marine aquaculture ranges from 2% (in Atlantic cod, Gadus morhua), to 50% (in Atlantic
salmon, Salmo salar) (Blackwell, 2019). However, pathogenicity is strongly influenced by water
quality and stocking levels (Nash et al.,1987) and varies between hosts (Muroga et al., 1975).
Experimental infection per os failed to initiate clinical disease in rainbow trout (Oncorhynchus
mykiss) (Lonnstrom et al., 1994), or in wild turbot (Scophthalmus (Psetta) maximus) (Magi et
al., 2009). In both studies, subsequent infection by intra-peritoneal (i.p.) injection resulted in

96 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
80% mortality in rainbow trout and 100% mortality in turbot, respectively. These data reflect the
opportunistic nature of P. anguilliseptica infection (Magi et al., 2009) and indicate P.
anguilliseptica is a secondary pathogen (requiring another agent to initiate infection) (Daly &
Akoi, 2011).
Strains of P. anguilliseptica associated with wild Atlantic herring (Clupea harengus) are of low
pathogenicity (Lonnstrom et al., 1994) where P. anguilliseptica may also be a secondary
pathogen, (Wiklund, 2016).
Clinical signs
External signs of infection may include exophthalmia, petechial haemorrhages, ulceration of the
ventral surface, mouth, anal region and fin bases, as well as eye lesions and punctured corneas
(Lonnstrom et al., 1994; Wiklund, 2016). The disease may also progress with no external clinical
signs (Mastan, 2013; Wiklund, 2016).
Pathogen survival
P. anguilliseptica survives in dead fish tissues (Sakr & El-Rhman, 2008). Experimental data
indicate P. anguilliseptica remains viable in marine and fresh water for extended periods (Ciric
et al., 2009), but its survival in natural waters is unknown (see section 6.1.2) Growth is limited
above 37 °C (Ferguson et al., 2004).
P. anguilliseptica survives in fish offal dumped in landfill and has been recovered from leachates
in groundwater (Tian et al., 2014). It is also present in deep-sea invertebrate communities
(Romanenko et al., 2008).
Disease transmission
Disease transmission is horizontal, through the water column. Surviving fish are carriers (Fadel
et al., 2018). P. anguilliseptica may be dispersed through infected discharge water, biofilm and
fomites (Lonnstrom et al., 1994).
Infection and disease progression
Infection occurs through cohabitation, or by consumption of infected fish, but the details of
infection are poorly understood (Austin, 2005; Austin & Austin, 2007). The infective dose varies
among isolates (Magi et al., 2009).
Disease may follow several pathways. Infection may be systemic, with necrotic vacuolation and
degeneration of liver, kidney and spleen tissues. This spreads to connective tissues, skeletal
musculature, the brain and cartilage, causing myolysis and pyogranulomatous inflammation
(Sakr & El-Rhman, 2008). Infection may also target the brain and nervous system, causing
corneal opacity, cephalic osteochondritis and meningitis. Surviving fish are carriers of disease
(Magi et al., 2009). Infection associated with the nervous system occurred at a prevalence of
33% in wild Atlantic herring (C. harengus) (Lonnstrom et al., 1994) and 40% in sea bream
(Sparus aurata) (Fadel et al., 2018).
Inactivation
P. anguilliseptica is resistant to freezing. It is unaffected by frozen storage (to -20 °C)
(Algammal et al., 2020) and remains viable after long-term storage (to -70 °C) (Hine &
MacDiarmid 1997; Magi et al. 2009). It is therefore assumed to be unaffected by either short
(168 hours), or medium-term (4 months) frozen storage (at -20 °C).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 97
P. anguilliseptica is likely to be denatured by ionising radiation (at 50 kGy) (DAFF 2013).
17.2 Risk assessment
17.2.1 Entry assessment
P. anguilliseptica is associated with Atlantic herring (C. harengus) from Finnish waters at a
prevalence of up to 50%. Infected fish show no external signs of infection (Lonnstrom et al.,
1994). P. anguilliseptica is likely to remain viable in the commodity (Sakr & El Rhman, 2008;
Magi et al., 2009). As the host range and geographical range associated with fish bait are limited,
the likelihood of entry is assessed as low.
17.2.2 Exposure and establishment assessment
P. anguilliseptica survives in marine and fresh waters for extended periods (Ferguson et al.,
2004; Ciric et al., 2009). It remains viable in landfill and can be dispersed between waterways by
mechanical vectors such as piscivorous birds (Tian et al., 2014).
Exposure may occur through consumption of infected fish bait (Nash et al., 1987; Lonnstrom et
al., 1994), or by cohabitation through infected meltwater (Magi et al., 2009). It has a wide range
of potential hosts in New Zealand waters (Tubbs et al., 1997).
The strains associated with Atlantic herring are of low pathogenicity (Lonnstrom et al. 1994).
and P. anguilliseptica may be a secondary pathogen (Wiklund, 2016). It may be introduced into
farmed fish stocks through the use of fish bait as aquaculture feed, but establishment is strongly
influenced by water quality and stocking levels (Lopez-Romalde et al., 2003; Sakr & El Rhman,
2008; Magi et al., 2009; Jansson & Vennerstrom, 2014).
The likelihood of exposure and establishment of P. anguilliseptica through fish bait is assessed
to be so low as to be negligible.
17.3 Risk management
As the likelihood of exposure and establishment is assessed to be negligible, P. anguilliseptica is
not assessed to be a risk in the commodity. No further risk assessment is necessary.
Streptococcus spp. (S. agalactiae serotype III: 283, S. iniae)
18.1 Technical review
18.1.1 Aetiological agent
Streptococcus agalactiae (serotype III: 283) and S. iniae are gram-positive non-spore forming
bacteria of the Streptococcus species complex, classified within the family Streptococcaceae
(Gao et al., 2014; Mishra et al., 2018). S. agalactiae is classified in Lancefield’s serotype group
B Streptococcus (GBS).
Pathogenicity and host specificity vary widely among the members of this species complex
which include free-living, commensal and zoonotic organisms (Agnew & Barnes, 2007;
Rajendram et al., 2016; Zhou et al., 2011, 2018; Mishra et al., 2018; Barkham et al., 2019).
Many Streptococcus spp. are considered opportunistic pathogens (Bunch & Bejerano, 1997). S.

98 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
agalactiae III: 283 and S. iniae are regarded as primary fish pathogens, causing streptococcosis
disease in fish (Salati, 2011; Mishra et al., 2018).
18.1.2 OIE Status
Streptococcosis is not an OIE-listed disease (OIE, 2019a).
18.1.3 New Zealand status
Serotypes of S. agalactiae are endemic in New Zealand, causing mastitis in dairy animals and
mild streptococcosis in wild and farmed fish (Hine & Diggles, 2005; Johnston, 2008; DermNet,
2014).
The pathogenic serotypes of S. agalactiae including serotype III: 283 reported from fish
(Barkham et al., 2018) have not been reported from New Zealand and are considered exotic
(Blackwell, 2019).
S. iniae has never been reported from New Zealand (Hine & Diggles, 2005; Johnston, 2008). It is
also considered to be exotic. Infection with Streptococcus spp. is not a notifiable disease in New
Zealand (Anon., 2016).
18.1.4 Zoonotic disease
S. iniae and S. agalactiae III: 283 are zoonotic pathogens. S. iniae infection is mainly associated
with puncture wounds arising from the handling of fish. This generally causes mild bacteremic
cellulitis. In rare cases, however, infection progresses to endocarditis, meningitis, arthritis,
sepsis, pneumonia, osteomyelitis and toxic shock (Shoemaker et al., 2001; Baiano & Barnes,
2009).
S. agalactiae infection occurs through consumption of under-cooked fish. Infection may result in
meningitis and pneumonia (Agnew & Barnes, 2007; Baiano & Barnes, 2009).
Of the 2,824 hospitalised cases in Australia attributed to atypical Streptococcus spp. during
1999–2000 and in 2006–2007, 278 cases in 1999–2000, and 430 cases in 2006–2007, were due
to S. iniae, respectively. As S. iniae is not included in routine biochemical testing or clinical
databases, these values are likely to be under-estimates (Baiano & Barnes, 2009).
18.1.5 Epidemiology
Distribution and host range
S. agalactiae (serotype III: 283) (= S. difficule) and S. iniae occur in over 20 families of fish
(Blackwell, 2019). They are widely distributed (Table 18 and 19).
The species associated with S. agalactiae III: 283 in fish bait and their geographic/pathogen
range are given in Table 18 (Eldar et al., 1995; Evans et al., 2002). Streptococcus infection also
occurs in South American pilchard (Sardinops sagax) in Kenyan coastal waters (Odhiambo et
al., 2018). This is not considered further as the species of Streptococcus is not defined.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 99
Table 18. Fish bait species susceptible to Streptococcus agalactiae III: 283
Family Host species Host geographical range1 Pathogen range2
Mugilidae Flathead grey mullet (Mugil cephalus)
East, West Atlantic, Mediterranean East, West Pacific
Northeast Atlantic (Florida) Mediterranean (Europe)
Notes 1 Host geographical range (from FishBase 2019) 2 Pathogen geographical range (from literature). Streptococcus agalactiae III; 283 is reported from Europe (Belgium, Israel,
Iran, Italy, Kuwait, Spain), Asia (Hong Kong, Singapore, South Korea, Japan, Thailand, Viet Nam), North America (USA), South America (Brazil, Columbia, Costa Rica) and South Africa, (Buchanan et al. 2008; Zhou et al. 2011; Delannoy et al., 2013; Chen, 2019).
The species associated with S. iniae in fish bait and their geographic/host range are given in
Table 19 (Minami et al., 1979; Yasunaga et al., 1982; Sako, 1984; Johnston, 2008; Salati, 2011;
Kluzik & Woodford, 2016). The prevalence in flathead grey mullet (M. cephalus) may reach
50% during epizootics (Plumb et al., 1974).
Table 19. Fish bait species susceptible to Streptococcus iniae
Family Host species Host geographical range1 Pathogen range2
Carangidae Japanese jack mackerel (Trachurus japonicus)
Northwest Pacific, Japan, Korea
Northwest Pacific
Clupeidae South American pilchard (Sardinops sagax (= S. melanostictus)
West Pacific West Pacific Ocean
Mugilidae Flathead grey mullet (Mugil cephalus)
East, West Atlantic, Pacific. Mediterranean Sea
Mediterranean Sea (Israel)
Scombridae Chub mackerel (Scomber japonicus)
Indo-Pacific, Eastern, Western Pacific
Northwest Pacific
Notes 1 Host geographical range (from FishBase 2019) 2 Pathogen geographical range (from literature) S. iniae is globally distributed (Susanna et al., 2003).
Potential fish host species in New Zealand
A wide range of marine and freshwater fish host species are present in New Zealand (Tubbs et
al., 2007; Blackwell, 2019).
Pathogenicity
S. agalactiae III: 283 affects wild species of marine and freshwater fish where water
temperatures exceed 17 °C (Salati, 2011) and rarely affects farmed species where temperatures
are lower than 25 °C (Tavares et al., 2018). New Zealand coastal water temperatures vary from
17 °C in the north, to 12 °C in the south (Pinkerton et al., 2018). While S. agalactiae may
survive in New Zealand waters, it is likely to only affect farmed fish species in Northern New
Zealand waters.
S. iniae infection occurs in a wide range of tropical and temperate waters from 10 °C to 45 °C.
Optimal pathogenicity is associated with water temperature range from 35°C to 45 °C, but
infection occurred in farmed Australian barramundi (Lates calcarifer) at 10 °C (Bromage et al.
1999; Bromage & Owens, 2002).
Mortalities range from 10% to 60% in the coastal marine aquaculture of rainbow trout
(Oncorhynchus mykiss) (Salmonidae), turbot (Scophthalmus maximus) (Scophthalmidae),

100 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
gilthead sea bream (Sparus aurata) (Sparidae) in Europe and Australia (Carson et al., 1993;
Prieta et al., 1993, cited in Musquiz et al., 1999; Toranzo et al., 2009). Mortality is strongly
influenced by water quality and stocking levels.
Clinical signs
External signs of infection may include exophthalmia, distended abdomen, haemorrhages in the
eyes, fin bases and opercula, darkening of the skin and spine displacement (Amal & Saad, 2011).
Infection may also progress with no external signs (Yanong & Francis-Floyd, 2002; Salati,
2011).
Pathogen survival
S. iniae survives in marine waters for up to 7 days (Bromage et al., 1999). Survival in natural
waters is unknown (see section 6.1.2).
Disease transmission
Transmission is horizontal and direct, either through consumption of infected fish tissues, or as
opportunistic infection, following abrasion and skin damage (Yasunaga, 1982; Shoemaker et al.,
2000). Subclinically infected or surviving fish may act as carriers of infection (Baiano & Barnes,
2009).
Infection and disease progression
Infectivity is maintained for long periods in the aquatic environment by sequential subclinical
infection (Kitao et al., 1979; Nguyen et al., 2002). S. iniae remained present in water and
sediment samples taken from a bastard halibut (Paralichthys olivaceus) farm for two years
following initial infection (Bromage et al., 1999).
Infection is internalised by invasion and survival within macrophage cells (Yoshida et al., 1996;
Bromage et al., 1999; Baiano & Barnes, 2009). Experimental infection per os indicate the
infectious dose is low, ranging from 1 × 103 cfu ml-1 (in for barramundi, Lates calcarifer), to 2 ×
107 cfu ml-1 (in tilapia, Oreochromis sp.), after 1-minute exposure (Bromage & Owens, 2002;
Shoemaker et al., 2000).
Acute infection mainly occurs in younger fish, but all ages may be infected. Disease progresses
from the skin to systemic septicaemia of the spleen, liver, heart and kidney tissues, eyes, brain
and nervous system (Amal & Saad, 2011; Salati, 2011). Chronic infection may develop into a
carrier state in older fish (Baiano & Barnes, 2009; Salati, 2011).
Wild fish may be reservoirs of infection for farmed species. Streptococcus sp. infection was
transferred from wild Klunzinger’s mullet (Liza klunzingeri) to gilthead seabream (Sparus
aurata) farmed in Kuwait (Zlotkin et al.,1998; Evans et al., 2002; Salati, 2011).
Levels of S. iniae in infected host tissues may reach 109 cfu g-1 in spleen, kidney and brain tissue
and 109 cfu mL-1 in blood (Nguyen et al., 2002; Shutou et al., 2007). Infection results in lowered
productivity, while the presence of blood-filled lesions in fish tissues renders the product
commercially unmarketable (Muzquiz et al., 1990; Salati, 2011).
Inactivation
Streptococcus spp. remain viable in fish tissues after short term frozen storage (at -70 °C for 180
days) and long-term frozen storage (at -20 °C for up to 9 months) (Evans et al., 2004; Hine &
Digges, 2005; Diggles, 2011). Streptococcus spp. remain viable in frozen sardine (S. sagax)

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 101
where used as feed for yellowtail (Seriola quinqueradiata) aquaculture in Japan (Yasunaga,
1982) and are likely to remain infective in frozen fish bait. It is assumed therefore, that
Streptococcus spp. would be unaffected by either short term (168 hours), or medium-term (4
months) frozen storage (at -20 °C).
Streptococcus spp. are likely to be denatured by ionising radiation (at 50 kGy) (DAFF, 2013).
18.2 Risk assessment
18.2.1 Entry assessment
S. agalactiae III: 283 and S. iniae are associated with fish bait (Table 18 and Table 19). Infected
fish may show no external signs of infection (Evans et al., 2011). Prevalence in wild fish bait is
unknown, but this may reach 50% following epizootics (Plumb et al., 1974; Lafferty et al.,
2015). Streptococcus spp. are unaffected by frozen storage (Evans et al., 2004; Hine & Diggles,
2005, Diggles, 2011) and may be present in the meltwater associated with fish bait (Bromage et
al., 1999).
The likelihood of entry is assessed as medium.
18.2.2 Exposure assessment
Experimental data indicate Streptococcus spp. survive for up to 7 days in the aquatic
environment (Bromage et al., 1999) within the temperature range of New Zealand coastal marine
waters (Pinkerton et al., 2018).
The pathogen load in tissues or in meltwater is high (Nguyen et al., 2001; Shutou et al., 2007),
while the infectious dose is low (Bromage & Owens 2002). Infectivity can be maintained in the
environment by sequential subclinical infection of a range of carrier hosts (Bromage et al., 1999;
Nguyen et al., 2002).
Infection has occurred through use of frozen fish bait (S. sagax) as feed in aquaculture
(Yasunaga, 1982). Infection can also be transferred from wild to farmed fish (Plumb et al., 1974;
Zlotkin et al., 1998; Salati, 2011). S. iniae was introduced into the USA in 1994, demonstrating
its ability to translocate (Perera et al., 1994; Johnston, 2008).
The likelihood of exposure and establishment of S. agalactiae III: 283 and S. iniae is assessed as
medium.
18.2.3 Consequence assessment
Economic consequences
Streptococcosis (including infection with S. iniae or S. agalactiae) is not an OIE-listed disease
(OIE, 2019a), so establishment would not result in loss of trade access for New Zealand.
Establishment may result in lowered productivity and increased mortality, as well as product loss
resulting from tissue contamination (Muzquiz et al., 1990; Salati, 2011). Streptococcosis causes
economic losses estimated at US$100 million annually in the combined aquaculture of Australia,
Israel, Japan and the USA (Shoemaker et al., 2001).
S. agalactiae III; 283 and S. iniae have a wide host range (Salati, 2011). Their establishment may
affect the aquaculture of Chinook salmon (Oncorhynchus tshawytscha) (Salmonidae), with

102 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
exports valued at $77 million in 2018 (Aquaculture New Zealand, 2019). It may also affect the
developing aquaculture of yellowtail kingfish (Seriola lalandi) (Carangidae) and New Zealand
snapper (silver seabream) (Pagrus auratus) (Sparidae) (Plant & Food, 2016, 2017; NIWA,
2017a).
Establishment may reduce productivity of important coastal inshore fisheries (MPI, 2018a,
2018b, 2018c). Wild fish are reservoirs of infection for farmed species, including gilthead sea
bream (Sparus auratus) (Sparidae) (Zlotkin et al., 1998). The New Zealand snapper (P. auratus)
export fishery was valued at $33 million in 2018 (Seafood New Zealand, 2019).
Establishment may affect farmed freshwater species, including grass carp (Ctenopharyngodon
idella) and silver carp (Hypophthalmichthys molitrix) used for weed control (Clayton & Wells,
1999). Introduced cyprinids such as the European (red fin) perch (Perca fluviatilis) may function
as reservoir hosts.
Social consequences
Exotic S. iniae and S. agalactiae strains are emerging zoonotic pathogens (Salati 2011). Most
cases are minor, but infection may be serious in older patients (> 50 years) or those with
compromised immune systems (Shoemaker, 2001; Agnew & Barnes, 2007; Baiano & Barnes,
2009; Salati, 2011).
Establishment may affect recreational trout fishing (brown trout, Salmo trutta) and rainbow trout
(Oncorhynchus mykiss) (Fish & Game, 2014). The Lake Taupo trout fishery alone was valued
annually at NZ$ 70–80 million (Marsh & Mkwra, 2013).
The effects of pathogens on wild fisheries are poorly known (Lafferty et al., 2015), but
establishment may affect other species of importance for customary and recreational fishing in
marine and fresh waters (Wynn-Jones et al., 2014, 2019). Recreational fishing is the 5th most
popular recreational activity for adults, involving 700,000 marine fishing trips annually, with a
contribution of NZ$ 638 million to the GDP in 2015 (Gross domestic product) (Southwick et al.
2018). The snapper and trevally recreational fisheries contributed NZ$ 402 million and NZ$ 71
million respectively to the GDP in 2015 (Southwick et al., 2018).
Environmental consequences
Streptococcus spp. may be introduced through frozen fish bait (Yasunaga, 1982). S. agalactiae
III: 283 and S. iniae have a wide host range. The pathogen load is high, and the infective dose is
low (Bromage & Owens, 2002). Infection has established through sequential subclinical
infection of susceptible wild fish, leading to Streptococcus spp. becoming endemic (Kitao et al.,
1979; Nguyen et al., 2002).
Epizootic infection is reported in flathead grey mullet (Mugil cephalus) (Plumb et al., 1974).
Establishment may cause lowered productivity in a wide range of marine and freshwater fish
species considered stressed at current levels of fishing activity (MPI, 2018a, 2018b, 2018c).
Inshore fish of families Clupeidae, Engraulidae and Mugilidae also represent a significant food
source for other fish, seabirds and marine mammals and a major link between trophic levels
(Dunn et al., 2012).
The consequences of the introduction of S. agalactiae III: 283 and S. iniae are assessed to be
medium.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 103
18.2.4 Risk estimation
As the entry, establishment and consequence assessments are medium, the risk estimates for S.
agalactiae III: 283 and S. iniae are assessed to be medium. Under the procedures followed in this
risk analysis, risk management measures may be justified.
18.3 Risk management
S. agalactiae III: 283 and S. iniae have been assessed to be risks in fish bait. Infection with
Streptococcus spp. is not an OIE-listed disease, so the OIE Aquatic Code (OIE, 2019a) provides
no specific guidance or processing requirements that would ensure the destruction of these
pathogens.
S. agalactiae III: 283 and S. iniae are associated with the commodity (Table 18 and Table 19).
Therefore, restriction to non-susceptible species in the commodity should substantially reduce
the risk associated with these pathogens and be a viable risk management option.
S. agalactiae III: 283 and S. iniae have a wide geographic range (Table 18 and Table 19). Their
occurrence is likely to be low in other geographic regions. Restriction through the BNZ Country
Approval Procedures to regions where these pathogens have not been reported should
substantially reduce the risk associated with these pathogens and be a viable risk management
option.
The presence of S. agalactiae III: 283 and S. iniae in a shipment of the commodity may be
verified by batch testing using standard sampling procedures (Corsin et al., 2009; OIE, 2019a).
Standard genomic tests for S. iniae are not available. Recognised diagnostic procedures are
provided in Weinstein et al. (1997), Pereira et al. (2013) and Buller (2014). Verification of a
negative test from a shipment should substantially reduce the risk associated with these
pathogens and be a viable risk management option.
Streptococcus spp. are unaffected by frozen storage (at -20 °C) for up to 9 months (Evans et al.,
2004; Hine & Digges, 2005; Diggles, 2011). Therefore, frozen storage, for either 168 hours or 4
months, is not a viable risk management option.
Streptococcus spp. are assumed to be denatured by ionising radiation (at 50 kGy) (DAFF 2013).
This irradiation treatment should substantially reduce the risk associated with these pathogens
and be a viable risk management option.
18.3.1 Risk management options
In addition to the general risk management measures that are proposed for all imported fish bait
(see section 6.2.1), one or more of the following risk management options would reduce the risk
specific to S. agalactiae III: 283 and S. iniae an acceptable level.
Option 1
• Competent Authority attestation that the commodity does not include Japanese jack mackerel
(Trachurus japonicus) (Carangidae); South American pilchard (Sardinops sagax)
(Clupeidae); flathead grey mullet (Mugil cephalus) (Mugilidae); and chub mackerel
(Scomber japonicus) (Scombridae) should substantially reduce the S. agalactiae III: 283 and
S. iniae risk, so the commodity could be imported without further restrictions.

104 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Option 2
• Acceptance of region/country freedom through the BNZ Country Approvals Procedures
should substantially reduce the S. agalactiae III: 283 and S. iniae risk, so the commodity
could be imported without further restrictions.
Option 3
• A negative test from a shipment of the commodity, following approved sampling and
diagnostic procedures through the BNZ Country Approvals Procedures should substantially
reduce the S. agalactiae III: 283 and S. iniae risk, so the commodity could be imported
without further restrictions.
Option 4
• Treatment with ionising radiation (at 50 kGy) of the frozen commodity should substantially
reduce the S. agalactiae III: 283 and S. iniae risk, so the commodity could be imported
without further restrictions.
Myxozoan pathogens
19.1 Technical review
19.1.1 Aetiological agents
Myxozoan (= Myxosporean) fish pathogens are highly specialised multicellular obligate
parasites. They are classified in Class Myxozoa of the Phylum Cnidaria (WoRMS, 2019). The
current classification within the Myxozoa is based on a complex and detailed description of
spore anatomy (Lom & Dykova, 1992). This classification has become controversial. Genomic
studies have shown the Class Myxozoa to be polyphyletic, with several major clades and species
complexes (e.g., Chloromyxum, Kudoa, Myxobolus spp.) (Okamura et al., 2011; Fiala et al.,
2015; Okamura & Gruhl, 2015; Chang et al., 2015; Okamura et al., 2018). Some, including the
Kudoa thyrsites species complex, have a wide host range (Feist & Longshaw, 2006; Fiala et al.,
2015; Vidal et al., 2017).
Myxozoans generally cause little apparent harm to their hosts (Feist & Longshaw, 2006).
However, tissue-inhabiting myxozoans cause extensive post-mortem myoliquefaction,
commonly resulting in post-harvest rejection of the commercial product. Infected wild fish may
become reservoir hosts for farmed species (Langdon et al., 1992; Hine & Jones, 1994; Moran et
al., 1999). Myxozoans have been previously identified as risk organisms in New Zealand fish
(Johnston, 2008; Blackwell, 2019).
This risk analysis is restricted to exotic myxozoans associated with post-mortem
myoliquefaction, where the host range includes fish species of economic, social or
environmental importance to New Zealand. These are reviewed together in this risk analysis, as
the host range is common for several identified myxozoan hazards.
19.1.2 OIE status
Diseases associated with myxozoan pathogens are not listed by the OIE (OIE, 2019a).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 105
19.1.3 New Zealand status
Myxozoans are reported from New Zealand fish. These are generally host-specific and are
primarily associated with coelomic spaces (bile duct, gall bladder, urinary bladder) (Hine. 1977;
Hewitt & Little, 1972; Boustead, 1982; McArthur & Sengupta, 1982; Landsber & Lom, 1991;
Hine et al., 2000; Diggles et al., 2002; Bingham, 2009; Lane et al., 2014). They are likely to
have a minimal effect on their host (Lom & Dykova, 1992).
Myxozoan species including Kudoa hexapunctata, K. prunusi, K. shiomitsui and K. yasunagi
(Table 6) are reported from highly migratory pelagic and oceanic tuna species. These tuna
species are widely reported from the Pacific Ocean, including New Zealand (Zhang et al., 2010;
Meng et al., 2011; Kasai et al., 2017). It is considered likely that the myxozoans associated with
these highly migratory species are present, but not reported in New Zealand waters. They are not
considered further.
Kudoa thyrsites has a wide geographical distribution and a wide host range. It has not yet been
reported from New Zealand waters (Hine & Jones, 1994; Stone et al., 1997; Hine et al., 2000;
Blackwell, 2019). It has not been reported from passive testing programmes (M. Bestbier, BNZ,
pers. comm., 2020). It is assumed that K. thyrsites is absent from New Zealand waters.
19.1.4 Zoonotic disease
Myxozoans are minor zoonotic pathogens (Boreham et al., 1998; Feist & Longshaw, 2006;
Iwashita et al., 2013; Kasai et al., 2015). Myxobolus spp. have caused zoonotic infection
following consumption of previously frozen fish fillets (Boreham et al., 1998; Ohnishi et al.,
2016).
19.1.5 Epidemiology
Distribution and host range
Blackwell (2019) identified several myxozoan species of concern likely to be associated with
eviscerated fish. The present risk analysis considers a wider range of myxozoans likely to be
present in whole (uneviscerated) fish bait species (Table 5). Most myxozoans are host-specific,
at least to the family level, with high tissue or organ-specificity (Lom & Dykova, 1995, 2006;
Molnar & Eszterbauer, 2015). They are of low or low-moderate pathogenicity and are not
regarded as significant fish pathogens in wild fish stocks (Langdon et al., 1992; Lom & Dykova,
1992, 2006; Diamant et al., 1994; Feist & Longshaw, 2006; Lom & Dykova, 2006; Gomez et al.,
2014). These have not been considered further.
Myxozoan species from genera Enteromyxum and Kudoa are associated with post-mortem
myoliquefaction and have a wide host range. E. leei occurs in 46 host species from 6 families. K.
clupeidae occurs in 13 host species from 2 families. K. iwatai occurs in 19 host species from 6
families. K. nova (= K. quadratum) occurs in 20 host species from 2 families, while the K.
thyrsites species complex occurs in 40 host species from 5 families (Lom & Dykova, 2006;
Blackwell, 2019).
The species associated with E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites in fish bait
and their geographic/host range are given in Table 20 (Hahn, 1917; Meglitsch, 1947; Heckman
& Jensen, 1978; Kpatcha et al., 1999; Egusa & Shiomitsu, 1983; Lom & Dykova, 1992; Munday
et al., 1998; Reimschuessel et al., 2003; Longshaw et al., 2004; Campbell, 2005; Diamant et al.,
2005; Whipps & Kent, 2006; Langdon, 2007; Montero et al., 2007; Matsukane et al., 2011;

106 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Henning et al., 2013; Mackenzie & Kalavati, 2014; Fall et al., 2015; Kasai et al., 2015; Levsen,
2015; Ovcharenko, 2016; Yurakhno, 2017; Dezfuli et al., 2020).
The reported geographical range of these myxozoans may underestimate their true distribution,
as some regions, such as the North Atlantic, have been more thoroughly investigated than others
(Mackenzie & Kalavati, 2014; Fiala et al., 2015).
Myxozoan prevalence varies widely but is likely to be low in wild fish stocks. Few data are
available, and the sample sizes are often small (Henning et al., 2013; Alama-Bermejo et al.,
2013). Reported prevalence of E. leei in the thinlip grey mullet (C. ramada) was estimated at
83% (from n= 43 fish) (Dezlui et al., 2020). K. thyrsites prevalence varies with season and with
host size in fish bait species. In the South American pilchard (S. sagax), prevalence varies from
17% during spring, to 91% in autumn (Reed et al., 2012); Henning et al., 2019). In Atlantic
mackerel (S. scombrus), overall prevalence was 0.8% (from 1,475 fish), but the prevalence was
higher (8.9%) in older, heavier (> 600 g) fish (Levsen et al., 2008).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 107
Table 20. Fish species susceptible to exotic myxozoan pathogens Enteromyxum leei, Kudoa clupeidae, K. iwatai. K. nova. K. thyrsites
Myxozoa Host family
Host species Host geographical range1
Pathogen range2
Enteromyxum leei
Mugilidae Flathead grey mullet (Mugil cephalus), golden grey mullet (Chelon auratus), thinlip grey mullet (C. ramada)
East Atlantic, West Atlantic, Mediterranean, West Pacific Ocean
East Atlantic, Mediterranean
Kudoa clupeidae
Clupeidae Atlantic herring (Clupea harengus)
Northwest Atlantic, Southwest Atlantic
Northeast Pacific (California), Southwest Atlantic
Kudoa iwatai Carangidae Japanese jack mackerel (Trachurus japonicus)
North Pacific Japan, Korea, Red Sea
Mugilidae Flathead grey mullet (Mugil cephalus)
Widespread, Mediterranean
Mediterranean, Japan
Kudoa nova Carangidae European horse mackerel (Trachurus trachurus)
North Atlantic, Northeast Atlantic, Black Sea, Mediterranean, Pacific
Pacific (Western Australia), Black Sea, North, Northeast Atlantic, West Atlantic, Mediterranean
Scombridae Bigeye tuna (Thunnus obesus), little tunny (Euthynnus alleteratus)
North Sea, Northeast, Northwest Atlantic, Mediterranean
North Sea, Northwest Atlantic, Mediterranean
Kudoa thyrsites
Clupeidae Bali sardinella (Sardinella lemuru), South American pilchard (Sardinops sagax)
Central, Northwest Pacific Northeast, Southeast Atlantic Southeast, Northeast Pacific, Mediterranean
Northwest, Eastern Pacific (Alaska, Australia, Japan, East, Northeast Northwest Atlantic, Mediterranean (Chile, England, South Africa)
Engraulidae Californian anchovy (E. mordax)
Northeast Pacific (California)
Northeast Pacific (California)
Scombridae European horse mackerel (Scomber scombrus), Chub mackerel (S. japonicus)
Northeast, Southeast Atlantic, Pacific (Australia, Canada, Chile, Japan)
Northeast, Southeast Atlantic (South Africa), Pacific (Australia, Japan, Southeast Pacific (Chile), Northeast Pacific (Canada, Alaska)
Notes 1 Host geographical range (from FishBase 2019) 2 Pathogen geographical range (from literature)
Potential fish host species in New Zealand
A wide range of potential hosts exist in New Zealand (Blackwell, 2019), but the following
commercially farmed fish species are relevant to this chapter: brown trout (Salmo trutta),
Chinook salmon (Oncorhynchus tshawytscha), rainbow trout (O. mykiss) (Salmonidae);
yellowtail kingfish (Seriola lalandi) (Carangidae); snapper (Pagrus auratus) (Sparidae); hapuku
(Polyprion oxygeneios) (Polyprionidae); turbot (Colistium nudipinnis) (Pleuronectidae). The host
range of the myxozoans E. leei, K. clupeidae, K. nova, K. iwatai and K. thyrsites includes finfish
species commercially farmed in New Zealand.

108 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pathogenicity
Myxozoans have little apparent effect on their fish hosts, even at high levels of infection (Lom &
Dykova, 1992, 2006; Alvarez-Pellitero & Sitja-Bobadilla, 1993; Feist & Longshaw, 2006;
Gomez et al., 2014; Diggles, 2011; Blackwell, 2019). Myxozoan species clades preferentially
infect specific tissue types, but the severity of infection varies among host species. (Egusa &
Nakajima, 1980; Burger et al., 2008).
Clinical signs
Infection in wild fish stocks usually progresses with few or no external signs of infection (Lom
& Dykova, 2006). Chronic infection is usually associated with aquaculture, where stressors may
include high temperatures, overcrowding and poor water quality. Heavily infected fish may
become emaciated, with visible cysts, lesions and necrosis of the skin or fins. These fish may
show poor weight gain and increased mortality (Lom & Dykova, 2006; Gomez et al., 2014;
Stilwell & Yanong, 2018).
Pathogen survival
Little is known about myxozoan survival in marine waters. Experimental data from fresh water
myxozoans such as M. cerebralis suggest infective actinospores may remain viable for up to 7
days in water (Hoffman, 1990; Eszterbauer et al., 2001), or up to 12 months in sediments
(Nehring et al., 2015). However, experimental data may not reflect survival in natural waters
(see section 6.1.2).
Disease transmission
The life cycle of marine myxozoans is complex (Chang et al., 2015; Fiala et al., 2015; Okamura
et al., 2015; Naldoni et al., 2019). Life cycle details are only known for 50 freshwater
myxozoans out of the 2,200 species associated with fish. These have an indirect life cycle,
usually involving a bryozoan or annelid intermediate host (Lom & Dykova, 2006).
Little is known about the life cycle details, viability and host range of marine myxozoans (Feist
& Longshaw, 2006; Lom & Dykova, 2006). These are assumed to also have an indirect life
cycle. The intermediate host may be a bryozoan or annelid (Lom & Dykova, 2006), or an
undescribed zooplankton species (Sindermann, 1990). Myxozoans may persist and replicate
through vertical transmission in their intermediate hosts (Feist & Longshaw, 2006; Hallett et al.,
2015; Nehring et al., 2015).
Some marine myxozoans, such as E. leei, have a direct fish-fish life cycle. Infection occurs by
cohabitation, or through consumption of infected tissues, but the details of infection are poorly
understood (Diamant, 1997; Redonto et al., 2002; Yasuda et al., 2002; Diamant et al., 2006;
Lom & Dykova, 2006).
Disease transmission in Kudoa spp. may also be direct, through an extra-sporogonic stage, or
stages in the life cycle. Experimental studies have shown infection with K. thyrsites can be
initiated in naïve Atlantic salmon (Salmo salar) by injection of blood from infected fish
containing these extra-sporogonic stages (Moran et al., 1999; Young & Jones, 2005). While the
significance of these life cycle stages in disease transmission is uncertain (Young & Jones,
2005), K. thyrsites could also be transmitted directly from fish to fish.
Myxozoans survive passage through avian digestive systems. The infective stages may be
dispersed in avian faeces between waterways by piscivorous birds (El-Matbouli & Hoffmann,

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 109
1991). Host specificity to the intermediate host is usually low (Gomez et al., 2014) and
alternative invertebrate hosts may be present in New Zealand waters.
Transmission of spores from the fish host may be delayed until the death or predation of the host.
Therefore, the time required for an exotic myxozoan to establish in a new region may be in the
order of decades. The process of establishment is complex and dependent on multiple factors.
These include temperature, salinity, pH, the rate of predation of the host fish relative to the
developmental stage of the pathogen, as well as on the distribution biology of both the
intermediate and final host (Hallett et al., 2015; Jones et al., 2015).
Infection and disease progression
Infective actinospores contact the fish host by invading cells of the skin, fins, buccal cavity,
digestive tract or gills (Lom & Dykova, 2006). Infection may remain localised in the epithelial
tissues, or become systemic, affecting multiple organs (Gomez et al., 2014).
Infection may be coelozoic (inhabiting body spaces) or histozoic infection (inhabiting organs or
muscle tissues). Chronic infection is usually lifelong. Infection generally progresses with little
apparent effect upon the host, where the infection rate is low (Lom & Dykova, 2009).
At higher rates of infection, tissue necrosis, cell detachment and vacuolisation may occur. This
results in chronic enteritis, anorexia, and cachexia (Gomez et al., 2014). Infection of the gill
tissues causes epithelial damage and cyst formation. The resulting hypertrophy and fusion of gill
lamellae may significantly reduce gill function (Lom & Dykova, 2006).
Where infection occurs in the musculature, open skin lesions develop that allow myxospores to
be released to the environment. When these lesions heal, an enclosed pseudocyst is formed. The
processes of pseudocyst formation are poorly understood (Sindermann, 1990; Rothwell et al.,
1997; Gomez et al., 2014). These pseudocysts decompose post-mortem, causing tissue
myoliquefaction, tissue spoilage and economic loss of the commercial fish product (Feist &
Longshaw, 2006). The loss rates from product rejection due to tissue myoliquefaction may be
significant. It is estimated that 4% to 7% of Canadian farmed Atlantic salmon (Salmo salar)
product is rejected due to myoliquefaction (Marshall et al., 2015).
Myxozoans present in deeper tissues of the brain, cartilage, or somatic muscle tissues also cause
little apparent harm to their host. These are released by decomposition, following the death of the
host (Diamant, 1997; Feist & Longshaw, 2006).
Inactivation
Myxospores are relatively resistant to frozen storage in fish tissues. Most inactivation data are
available for freshwater myxozoans. Spore viability of M. cerebralis is reduced following
medium-term (3 months) frozen storage (to -20 °C or below) (Boreham et al., 1998; El Matbouli
& Hoffmann 1991; Yurakhno, 2017). Infectivity is significantly reduced after 5 months storage
(at -20 °C), where only 2 of the 15 samples (13%) remained viable (Hoffman & Putz, 2011). M.
cerebralis spores in salmon tissue stored at -20 °C or -80°C were rendered non-viable after 2
months storage, while spores held at 4 °C and higher remained infective for the Tubifex tubifex
intermediate host (Hedrick et al., 2008).
For marine myxozoans, a 20% reduction in viability of Kudoa nova occurred after freezing
(to -20 °C) (Yurakhno, 2017). Kudoa septempunctata spores in olive flounder (Paralichthys
olivaceus) were rendered non-viable by frozen storage (to -20 °C for 4 hours) (Anon., 2020b).

110 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Marine myxozoans are likely to be unaffected by short-term (168 hours) frozen storage
(at -20 °C), but substantially, or completely denatured by medium-term frozen storage (for at
least 4 months) (El Matbouli & Hoffmann, 1991; Boreham et al., 1998; Lom & Dykova, 2006;
Hedrick et al., 2008; Reed et al., 2012; Ohishi et al., 2016; Henning et al., 2019; Anon., 2020b).
Myxozoan prevalence is generally low in wild fish stocks. Where pathogen load is high,
sufficient spores may remain viable to initiate infection after short-medium term frozen storage
(Hedrick et al., 2008; Levsen et al., 2008; Bartholomew & Kerans, 2015; Henning et al., 2019).
Myxozoans are likely to be denatured by ionising radiation (at 50 kGy) (DAFF, 2013).
19.2 Risk assessment
19.2.1 Entry assessment
The myxozoans E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites are associated with fish
bait, including species that are imported in large quantities (Table 3). Infected fish typically
show no external signs of infection (Lom & Dykova, 2006).
The prevalence in fish bait is highly variable depending on host species, fish size and season of
capture (Lom & Dykova, 2006; Levsen et al., 2008; Reed et al., 2012; Henning et al., 2019).
Myxozoans are likely to remain viable in the commodity. As myxozoan prevalence is highly
variable, the likelihood of entry is assessed as low.
19.2.2 Exposure assessment
The life cycle of marine myxozoans is poorly understood (Lom & Dykova, 2006). Some, such as
E. leei have a direct life cycle, with fish-fish transmission and a wide host range (Diamant et al.,
1997, 2006; Feist & Longshaw, 2006). Others, including Kudoa spp. may also have a direct life
cycle (Moran et al., 1999; Young & Jones, 2005). These may also use an intermediate host such
as an annelid, bryozoan, polychaete or other unknown zooplankton species (Sindermann, 1990;
Bartholomew & Kerans, 2015).
Little data are available on viability of the infective stages, infective dose, or the specificity of
the final, or intermediate hosts of Enteromyxum leei or Kudoa spp. (Feist & Longshaw,2006). It
is possible that alternative host species may be present in New Zealand waters (Blackwell,
2019). Myxozoans can also persist through vertical transmission in their intermediate hosts (Feist
& Longshaw, 2006; Hallett et al., 2015; Nehring et al., 2015). Myxozoans may be dispersed by
piscivorous birds between waterways through faeces or in discarded fish tissues (El-Matbouli &
Hoffmann, 1991).
It is reasonable to assume that infected fish bait used in inshore waters may infect coastal inshore
finfish, through a suitable benthic invertebrate host (if required), in quantities and duration
sufficient to establish an infection. However, the factors influencing establishment success of
myxozoans are complex. As infection is rarely fatal to the finfish host, successful establishment
in wild stocks may be unapparent, even over a time scale of decades (Hallett et al., 2015; Jones
et al., 2015).
Myxozoan establishment may only be apparent in susceptible marine farmed species, where
infection has become established in a susceptible wild fish host, that could act as a reservoir of
infection (Feist & Longshaw, 2006; Montero et al., 2007; Bartholomew & Kerans, 2015).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 111
The likelihood of exposure and establishment of E. leei, K. clupeidae, K. iwatai, K. nova and K.
thyrsites through fish bait is extremely uncertain but is assumed to be low, or very low.
19.2.3 Consequence assessment
Economic consequences.
Disease associated with E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites is not listed by
the OIE (OIE, 2019a), so establishment would not result in loss of general trade access for New
Zealand. However, the presence of K. thyrsites in Chinook salmon exports may result in loss of
access to some North American markets (M. Bestbier, BNZ, pers. comm., 2020).
E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites are likely to cause little apparent effect
in wild fish stocks at low levels of prevalence (Lom &Dykova, 2009). However, they may cause
economic losses through reduced productivity and increased mortality in aquaculture (Jones et
al., 2015). This would have significant consequence for New Zealand salmonid aquaculture,
including brown trout (Salmo trutta), Chinook salmon (Oncorhynchus tshawytscha) and rainbow
trout (Oncorhynchus mykiss). Losses in Canadian salmonid aquaculture resulting from Kudoa
thyrsites were estimated at CAD$ 6 million in 2015 (Braden et al., 2017). Chinook salmon
(Oncorhynchus tshawytscha) exports were valued at $77 million in 2018 (Aquaculture New
Zealand, 2019).
E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites may also affect aquaculture of non-
salmonid species including yellowtail kingfish (Seriola lalandi) (Carangidae), snapper (Pagrus
auratus) (Sparidae), hapuku (Polyprion oxygeneios) (Polyprionidae) and turbot (Colistium
nudipinnis) (Pleuronectidae) (Moran et al., 1999a; Rigos et al., 1999; Montero et al., 2007;
Rigos & Katharios, 2010; Berger & Adlard, 2011; ; Ovcharenko, 2016; NIWA, 2017a, 2017b,
2017c; Aquaculture New Zealand, 2019). For example, the snapper export fishery was valued at
$33 million in 2018 (Seafood New Zealand, 2019).
Other Kudoa spp. associated with mugilid and scombroid fish species have caused significant
economic losses in Asian, Australian and European aquaculture (Campbell, 2005; Lom &
Dykova, 2006; Montero et al., 2007; Yurakhno & Ovcharenko, 2014; Bartholomew & Kerans,
2015; Jones et al., 2015). These host species are not farmed in New Zealand, so the
consequences of establishment associated with these fish families are assumed to be negligible.
Social consequences
Myxozoans are minor zoonotic pathogens, causing gastroenteritis following consumption of
previously frozen fish fillets (Boreham et al., 1998).
The social consequences for customary and recreational fishing in New Zealand due to the
establishment of E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites are unknown, but likely
to be very low, at low rates of prevalence in wild fish stocks.
Environmental consequences
Evaluation of the consequences of myxozoan establishment in wild fish stocks is complex
(Lafferty et al., 2015). Myxozoans are of low pathogenicity (Egusa & Nakajima, 1980; Diamant
et al., 1994; Burger et al., 2008; Lom & Dykova, 2013; Gomez et al., 2014; Hallett et al., 2015;
Jones et al., 2015). The environmental consequences are unknown, but likely to be very low.
The consequences of establishment of E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites
are uncertain, but assessed to be low.

112 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
19.2.4 Risk estimation
The entry, exposure and consequence assessments for E. leei, K. clupeidae, K. iwatai, K. nova
and K. thyrsites are assessed to be low. The risk estimate is therefore assessed to be low, but with
high levels of uncertainty. Under the procedures followed in this risk analysis, risk management
measures may be justified.
19.3 Risk management
E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites have been assessed to be a risk in the
commodity. Infection with myxozoan pathogens is not an OIE-notifiable disease, so the OIE
Aquatic Code (OIE, 2019a) provides no specific guidance or processing requirements that would
ensure the destruction of these pathogens.
Myxozoans have been identified as risk organisms in previous IRAs in New Zealand
(MacDiarmid, 1993; Stone et al., 1997; Diggles, 2011; Blackwell, 2019) and in Australia (Kahn
et al., 1999). They have been identified as risk organisms in risk analyses for emerging
aquaculture species (Nowak et al., 2003; Hutson et al., 2007; Sanchez-Garcia et al., 2014;
Okamura et al., 2015).
Myxozoans E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites are associated with fish bait
(Table 21). These species are likely to be substantially or completely denatured by medium-term
frozen storage (to -20 °C for at least 4 months (17 weeks)) (El Matbouli & Hoffmann, 1991;
Boreham et al., 1998; Lom & Dykova, 2006; Hedrick et al., 2008; Reed et al., 2012; Ohishi et
al., 2016; Henning et al., 2019; Anon., 2020b). An extended period of frozen storage is
consistent with current fish bait industry practice. Adoption of this general risk management
option should substantially reduce the risk associated with these myxozoans and be a viable risk
management option.
19.3.1 Risk management options
If the general risk management measures proposed for all imported fish bait (section 6.2.1) are
adopted, then no pathogen-specific risk management measures would be necessary to reduce the
risk associated with E. leei, K. clupeidae, K. iwatai, K. nova and K. thyrsites to an acceptable
level.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 113
References
Aamelfot, M; Dale, O B; Weli, S; Koppang, E O; Falk, K (2012). Expression of 4-O-acetylated sialic acids on
Atlantic salmon endothelial cells correlates with cell tropism of infectious salmon anaemia virus. Journal of
Virology 86: 10571–10578.
Abollo, E; Novoa, B; Figueras, A (2005). SSU rDNA analysis of Kudoa rosenbuschi (Myxosporea) from the
Argentinean hake Merluccius hubbsi. Diseases of Aquatic Organisms 64: 135–139 [Online] Available from:
http://www.doi.10.3354/dao064135 [Accessed 6 December 2019].
Adachi, K; Ichinose, T; Watanabe, K; Kitazato, K; Kobayashi, N (2008). Potential for the replication of the
betanodavirus redspotted grouper nervous necrosis virus in human cell lines. Archives in Virology 153(1): 15–24
[Online] Available from: http://www.doi.10.1007/s00705-007-1051-9 [Accessed 17 Decamber 2019].
Afonso, L O B; Richmond, Z; Eaves, A A; Richard, J; Hawley, L M; Garver, K A (2012). Use of ultraviolet C
radiation (UVC) to inactivate infectious hematopoietic necrosis virus (IHNV) and viral haemorrhagic septicaemia
virus (VHSV) in fish processing plant effluent. Journal of Aquaculture Research and Development 3: 120 [Online]
Available from: http://omicsonline.org/use-of-ultraviolet-c-uvc-radiation-to-inactivate-infectious-hematopoietic-
necrosis-virus-ihnv-and-viral-hemorrhagic-septicemia-virus-vhsv-in%20fish-processing-plant-effluent-2155-
9546.1000120.php?aid=3943 [Accessed 24 April 2015].
Agnew, W; Barnes A C (2007). Streptococcus iniae: An aquatic pathogen of global veterinary significance and a
challenging candidate for reliable vaccination. Veterinary Microbiology 122: 1–15 [Online] Available from:
http://www.doi.10.1016/j.vetmic.2007.03.002 [Accessed 14 January 2020].
Ahne, W (1982). Vergleichende Untersuchung über die Stabilität von vier fischpatogenen Viren (VHSV, PFR,
SVCV, IPNV) (Comparative studies on the stability of four fish-pathogenic viruses [VHSV, PFR, SVCV, IPNV]).
Zentralbl. Veterinarmed. [B] (Journal of Veterinary Medicine Series B) 29: 457–476.
Alama-Bermejo, G; Sima, R; Raga, J A; Holzer, A S (2013). Understanding myxozoan infection dynamics in the
sea: Seasonality and transmission of Ceratomyxa puntazzi. International Journal for Parasitology 43(9): 771–780
[Online] Available from: https://doi.org/10.1016/j.ijpara.2013.05.003 [Accessed 6 December 2019].
Alderman, D J; Polgase, J L (1986). Are fungal diseases significant in the marine environment? pp 189–198 In:
Moss, S T (Ed.). The biology of marine fungi. Cambridge University Press, Cambridge, United Kingdom, 382 p.
Aldrin, M; Storvik, B; Frigessi, A; Vijiugrein, H; Jansen, P A (2010). A stochastic model for the assessment of
the transmission pathways of heart and skeleton muscle inflammation, pancreas disease and infectious salmon
anaemia in marine fish farms in Norway. Preventative Veterinary Medicine 93(1): 51–61.
Alexandrawicz, J S (1951). Lymphocystis tumours in the red mullet (Mullus surmuletus L.). Journal of the Marine
Biological Association of the United Kingdom 30: 315–332.
Alfjorden, A; Jansson, E; Johansson, K E (2006). A systemic granulomatous inflammatory disease in wild
Atlantic cod, Gadus morhua associated with a bacterium of the genus Francisella. In: Raynard, R; Wahili, T;
Vatsos, I; Mortensen, S (Eds.). Disease Interactions and Pathogen exchange between farmed and wild aquatic
animal populations - a European network (DIPnet). Final Report SSPSSP8-2004-006598).
Algammal, A M; Mabrok, M; Sivaramasamy, E; Youssef F M; Stwa, M H; Kholy, A W; Hetta, H F; Hozzein
W N (2020). Emerging MDR-Pseudomonas aeruginosa in fish commonly harbor oprL and toxA virulence genes
and blaTEM, blaCTX-M, and tetA antibiotic-resistance genes. Science Report 10: 15961 [Online] Available from:
https://doi.org/10.1038/s41598-020-72264-4 {Accessed 22 September 2020].
Almeida, L J; Da Silva, E J; Freitas, Y M (1968). Microorganisms from some tropical fish diseases. Journal of
the Fisheries Research Board of Canada. 25: 197-201.
Almendras, F E; Fuentealba, I C; Jones, S R M; Markham, F; Spangler, E (1997). Experimental infection and
horizontal transmission of Piscirickettsia salmonis in freshwater raised Atlantic salmon, Salmo salar L. Journal of
Fish Diseases 20(6): 409–418.
Alvarez-Pellitero, P; Sitja-Bobadilla, A (1993). Pathology of Myxosporea in marine fish culture. Diseases of
Aquatic Organisms 17: 229–238.
Amal, M N A; Saad, Z-S (2011). Streptococcosis in Tilapia (Oreochromis niloticus): A Review. Pertanika Jounal
of Tropical Agricultural Science 34(2): 195-206.
Amandi, A; Hiu, S F; Rohovec, J S; Fryer, J L (1982). Isolation and Characterization of Edwardsiella tarda from
Fall Chinook Salmon (Oncorhynchus tshawytscha). Applied and Environmental Microbiology 43(6): 1380–1384.

114 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Amos, K; Thomas, J; Hopper, K (1998). A case history of adaptive management strategies for viral hemorrhagic
septicaemia virus (VHSV) in Washington State. Journal of Aquatic Animal Health 10(2): 152–159 [Online]
Available from: https://doi.org/10.1577/1548-8667(1998)010%3C0152:ACHOAM%3E2.0.CO;2 [Accessed 7
October 2019].
Amos, K H; Bakal, R S; Barker, A A; Blair, M J; Bouchard, D A; Bruneau, N; Bowser, P R; Egrie, P G;
Ellis, S Kaisal, M; Garver, K A; Giray, C; Goodwin, A E; Gustafson, L L; House, N L; Karreman, G A;
Kebus, M J; Klotins, K C; LaPatra, S E; Marty, G D; Merrill, P D; Noyes, A D; Olesen, N J; Saksida, S M;
Snow, M; St-Hilaire, S; Tomlinson, S M; Uhland, S C; Vennerstrom, P; Wagner, B A; Warg, J V; Whelan, G
E; Winton, J R (2010). Viral haemorrhagic septicaemia virus (VHSV IVb) risk factors and association measures
derived by expert panel. Preventative Veterinary Medicine 94: 128–139.
Anderson, C (1990). Important diseases of salmonid fish and the risk they pose to New Zealand. Surveillance
17(2): 17–18.
Anderson, C (1996). Distribution of salmon diseases in New Zealand. Surveillance 23(4): 23–24.
Anderson, C; Knowles, G; De Lisle, G (1994). A survey for Edwardsiella ruckeri and Aeromonas salmonicida in
farmed and wild fish. Surveillance 21(3): 38–40.
Andree, K B; Rodgers, C J; Furones, D; Gisbert, E (2013). Co-infection with Pseudomonas anguilliseptica and
Delftia acidovorans in the European eel, Anguilla anguilla (L.): a case history of an illegally trafficked protected
species. Journal of Fish Diseases 36: 647–656.
Anonymous, (1981). Wholesomeness of irradiated food. Joint FAO/IAEA/WHO Expert Committee on Food
Irradiation. FAO Technical Report Series 659. World Health Organisation, Geneva, Switzerland.
Anonymous, (1984). General standard for irradiated food and recommended international code of practice for the
operation of radiation facilities used in the treatment of food. Codex Alimentarius, CAC/VOL XV, Food and
Agriculture Organisation of the United Nations, Rome, Italy.
Anonymous (2010). Viral hemorrhagic septicemia virus (VHSV IVb) risk factors and association measures derived
by expert panel. VHSV Expert Panel and Working Group. Preventative Veterinary Medicine 94: 128139 [Online]
Available from:
https://reader.elsevier.com/reader/sd/pii/S0167587709003638?token=49DAF55F616D30A27FD637A2BA5C189D4
6583907AC6E4095C8DA2DD2C9DC1B3333D4E5AC8047223845C85BC9A1E3CB4A [Accessed 21 September
2020].
Anonymous, (2012). Leptospirosis in New Zealand – Best practice recommendations for the use of vaccines to
prevent human exposure. Report for the New Zealand Veterinary Association, Massey University, Palmerston
North, New Zealand [Online] Available from: http://www.massey.ac.nz/massey/fms/Colleges/College%
20of%20Sciences/epicentre/Docs/Leptospirosis%20in%20NZ%20-%20Best%20Practice%20recommendations
%20vaccine%20use%20to%20prevent%20human%20exposure.pdf [Accessed 17 May 2019].
Anonymous, (2016). Biosecurity (Notifiable Organisms) Order 2016 SR 2016/73. New Zealand Government,
Wellington, 13 p.
Anonymous, (2017a). Aquatic Animal Diseases Significant to Australia: Identification Field Guide 4th Edition
Department of Agriculture, Fisheries and Forestry. Government of Australia. Canberra, Australia [Online] Available
from http://www.agriculture.gov.au/SiteCollectionDocuments/animal-plant/aquatic/field-guide/4th-
edition/finfish/viral-haemorrhagic-septicaemia.pdf [Accessed 31 July 2017].
Anonymous, (2017b). Acinetobacter baumanii species complex. General antimicrobial susceptibility data collected
from hospital and community laboratories. Public Health Surveillance, Environmental Science and Research (ESR),
Wellington, New Zealand [Online] Available from:
https://surv.esr.cri.nz/antimicrobial/general_antimicrobial_susceptibility.phpb [Accessed 28 May 2019].
Anonymous, (2020a). Aquareovirus. Diseases of wild and cultured fish in Alaska. Alaska Department of Fish and
Game, Juneau, Alaska, United States of America, 2 p. [Online] Available from:
https://www.adfg.alaska.gov/static/species/disease/pdfs/fishdiseases/aquareovirus.pdf [Accessed 28 June 2020].
Anonymous, (2020b). Kudoa septempunctata – Myxosporidia. Tokyo Food Safety Information Centre, Bureau of
Social Welfare and Public Health, Tokyo Metropolitan Government, 2-8-1 Nishishinjuku, Shinjuku-ku, Tokyo,
Japan [Online] Available from: https://www.fukushihoken.metro.tokyo.lg.jp/shokuhin/eng/musi/29.html [Accessed
29 July 2020].
Aquaculture New Zealand, (2019). New Zealand’s aquaculture industry key facts 2019 [Online] Available from:
http://www.seafoodnewzealand.org.nz/our-industry/key-facts/ [Accessed 17 December 2019].

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 115
AQUAVETPLAN, (2008). Operational Procedures Manual- decontamination, version 1.0 (2008). Department of
Agriculture, Fisheries and Forestry, Government of Australia, Canberra, Australia [Online] Available from:
http://www.daff.gov.au/aquavetplan [Accessed 24 September 2019].
Arimoto, M; Mushiake, K; Mizuta, Y; Nakai, T; Mw, K; Furusawa I (1992). Detection of striped jack nervous
necrosis virus (SJNNV) by enzyme-linked immunosorbent assay (ELISA). Fish Pathology 27: 191–195
Arkush, K D; Mendonca, H L; McBride, A M; Yun, S; McDowell, T S; Hedrick R P (2006). Effects of
temperature on infectivity and commercial freezing on survival of the North American strain of viral hemorrhagic
septicaemia virus (VHSV). Diseases of Aquatic Organisms 69: 145−151.
Arkush, K D; Bartholomew, J L (2011). Piscirickettsia, Francisella and epitheliocystis, pp 302–337 In: Woo, P T
K; Bruno, D W (Eds.). Fish Diseases and Disorders: Volume 3 - Viral, Bacterial and Fungal Infections, 2nd Edition.
CABI International, Wallingford, United Kingdom, 930 p.
Armiger, L C (1964). An occurrence of Labyrinthula in New Zealand Zostera. New Zealand Journal of Botany 2:
3–9.
Athanassopoulou, F; Roberts, R J (2004). Streptococcal infections of farmed fish. Journal of The Hellenic
Veterinary Medical Society 55(2): 136–144.
Atkinson, S D; Jones, S R; Adlard, R D; Bartholomew, J L (2011). Geographical and host distribution patterns
of Parvicapsula minibicornis (Myxozoa) small subunit ribosomal RNA genetic types. Parasitology 138(3): 969–977
[Online] Available from: http://www.doi.10.1017/S0031182011000734.[Accessed 13 September 2019].
Austin, B (2005). Bacterial Pathogens of Marine Fish. In: Belkin, S; Colwell, R R (Eds.). Oceans and health:
pathogens in the marine environment. Springer, Boston, Massachusetts, United States of America.
Austin, B; Austin, D A (2007). Bacterial fish pathogens: Diseases of wild and farmed fish. Springer Science,
580 p.
Bahri, S; Marques, A (1996). Myxosporean parasites of the genus Myxobolus from Mugil cephalus in Ichkeul
lagoon, Tunisia: description of two new species. Diseases of Aquatic Organisms 27: 115–122 [Online]
Available from: https://www.doi.org/10.3354/dao027115 [Accessed 20 September 2019].
Bahri, S; Andree, K B; Hedrick, R P (2003). Morphological and phylogenetic studies of marine Myxobolus spp.
from mullet in Ichkeul Lake, Tunisia. Journal of Eukaryotic Microbiology 50(6): 463–470 [Online] Available from:
http://www.doi.10.1111/j.1550-7408.2003.tb00272.x. [Accessed 22 October 2019].
Baiano, J C F; Barnes, A C (2009). Towards the control of Streptococcus iniae. Emerging Infectious
Diseases 15(12): 1891–1896 [Online] Available from: http://www.doi.10.3201/eid1512.090232 [Accessed 1
November 2019].
Bain, M B; Cornwell, E R; Hope, K M; Eckerlin, G E; Casey, R N; Groocock, G H; Getchell, R G; Bowser, P
R; Winton, J R; Batts, W N; Cangelosi, A; Casey, J W (2010). Distribution of an invasive aquatic pathogen (viral
haemorrhagic septicaemia virus) in the Great Lakes and its relationship to shipping. PLoS ONE 5 (4): e10156
[Online] Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2854150/ [Accessed 23 April 2015].
Balboa, S; Ferguson, H W; Romalde, J L (2003). Phenotypic, serological and genetic characterisation of
Pseudomonas anguilliseptica strains isolated from cod, Gadus morhua L., in northern Europe. Journal of Fish
Diseases 30: 657–664.
Baldrias, L R; Alvero, M A C (1999). Aeromonas species from raw squid balls: a preliminary study of a potential
food-borne pathogen and health hazard. Journal of Environmental Science and Management 2(1): 48–52.
Bandin, I; Souto, S (2020). Betanodavirus and VER Disease: A 30-year research review. Pathogens 9(106): 1–46
[Online] Available from: http://www.doi:10.3390/pathogens9020106 [Accessed 30 April 2020].
Barja, J L (2004). Report about fish viral diseases. pp 91-102. In: Alvarez-Pellitero P, Barja JL, Basurco B, Berthe
F, Toranzo AE (Eds.). Mediterranean aquaculture diagnostic laboratories. Zaragoza: CIHEAM, 2004. Options
Méditerranéennes: Série B. Etudes et Recherches, 49 p.
Barkham, T; Zadoks, R N; Azmai, M N A; Baker, S; Bich, V T N; Chalker, V; Chau, M L; Dance. D;
Deepak, R N; van Doorn, H R; Gutierrez, R A; Holmes, M A; Huong, L N P; Koh, T H; Martins, E;
Mehershahi, K; Newton, P; Ng, L C; Phuoc, N N; Sangwichian, O; Sawatwong, P; Surin, U; Tan, T Y; Tang,
W Y; Thuy, N V; Turner, P; Vongsouvath, M; Zhang, D; Whistler, T; Chen, S L (2019). One hypervirulent
clone, sequence type 283, accounts for a large proportion of invasive Streptococcus agalactiae isolated from
humans and diseased tilapia in Southeast Asia. PLoS Neglected Tropical Diseases 13(6): e0007421 [Online]
Available from: https://www.doi.org/10.1371/journal.pntd.0007421 [Accessed 14 January 2020].

116 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Barlow, D I; Sleigh M A (1979). Freeze substitution for preservation of ciliated surfaces for scanning electron
microscopy. Journal of Microscopy 115(1): 81–95.
Bartholomew, J L; Kerans, B (2015). Risk assessments and approaches for evaluating myxozoan disease
impacts, pp 379–395 In: Okamura, B; Gruhl, A; Bartholomew, J L (Eds.). Myxozoan Evolution, Ecology and
Development. Springer, Geneva, Switzerland, 441 p.
Baya, A M; Romalde, J M; Green, D E; Navarro, R B; Evans, J; May, E B; Toranzo, A E (1997).
Edwardsiellosis in wild striped bass from the Chesapeake Bay. Journal of Wildlife Disease 33(3): 517–525.
Bebak, J; McAllister, P E (2009). Continuous exposure to infectious pancreatic necrosis virus during early life
stages of rainbow trout, Oncorhynchus mykiss (Walbaum). Journal of Infectious Diseases 32: 173–181.
Berger, M A A; Adlard, R D (2011). Low host specificity in the Kudoidae (Myxosporea: Multivalvulida)
including seventeen new host records for Kudoa thalassomi. Folia Parasitologica 58(1): 1–16 [Online] Available
from: http://www.paru.cas.cz/folia [Accessed 1 November 2019].
Bergmann, S M; Fichtner, D; Skall, H F; Schlotfeldt, H J; Olesen, N J (2002). Age- and weight-dependent
susceptibility of rainbow trout Oncorhynchus mykiss to isolates of infectious haematopoietic necrosis virus (IHNV)
of varying virulence. Diseases of Aquatic Organisms 55: 205–210.
BERL, (2017). The economic contribution of commercial fishing to the New Zealand economy. Business and
Economic Research Limited, Wellington, New Zealand, 53 p. [Online] Available from:
https://www.inshore.co.nz/fileadmin/Documents/Other/BERL_Report_August_2017.pdf [Accessed 17 October
2019].
Berthe, F C J; Michael, C; Bernardet J F (1995). Identification of Pseudomonas anguilliseptica isolated from
several species in France. Diseases of Aquatic Organisms 21: 151–155.
Berzak, R; Scheinin, A; Davidovich, N; Regev, Y; Diga, R; Tchernov, D; Morick, D (2019). Prevalence of
nervous necrosis virus (NNV) and Streptococcus species in wild marine fish and crustaceans from the Levantine
Basin, Mediterranean Sea. Diseases of Aquatic Organisms 133(1): 7–17 [Online] Available from:
http://www.doi.10.3354/dao03339 [Accessed 17 December 2019].
Biacchesi, S; Le Berre, M; Le Guillou, S; Benmansour, A; Brémont, M; Quillet, E; Boudinot, P (2007). Fish
genotype significantly influences susceptibility of juvenile rainbow trout, Oncorhynchus mykiss (Walbaum), to
waterborne infection with infectious salmon anaemia virus. Journal of Fish Diseases 30: 631–636.
Bidegain, G; Powell, E N; Klinck, J M; Ben-Horin, T; Hoffmann, E E (2016). Marine infectious disease
dynamics and outbreak thresholds: contact transmission, pandemic infection, and the potential role of filter feeders.
Ecosphere 7(4): e01286 [Online] Available from http://www.doi.10.1002/ecs2.1286 [Accessed 3 April 2020].
Biering, E; Garseth, A H (2012). Heart and skeletal muscle inflammation (HSMI) of farmed Atlantic salmon
(Salmo salar L.) and the associated piscine reovirus (PRV). ICES Identification Leaflet for Diseases and Parasites
of Fish and Shellfish. No. 58, 6 p.
Billi, J H; Wolf, K (1969). Quantitative comparison of peritoneal washes and faeces for detecting infectious
pancreatic necrosis (IPN) virus in carrier brook trout. Journal of the Fisheries Research Board of Canada 26: 1459–
1465.
Bingham, P (2009). Myxozoan parasite confirmed. Surveillance 36: 38.
Biosecurity Australia, (2002). Policy review of fish importation resulting from the discovery of viral haemorrhagic
septicaemia virus (VHSV) in pilchards Sardinops sagax and mackerel Scomber japonicus from Californian waters:
Summary of technical information. Department of Agriculture, Fisheries and Forestry, Canberra, Australia.
Biosecurity New Zealand, (2006). Risk Analysis Procedures –Version 1, Biosecurity New Zealand, Ministry for
Primary Industries, Wellington, New Zealand [Online] Available from:
https://www.mpi.govt.nz/dmsdocument/2031/direct [Accessed 7 October 2019].
Birkbeck, T H; Rennie, S; Hunter, D; Laidler, L A; Wadsworth S (2004). Infectivity of a Scottish isolate of
Piscirickettsia salmonis for Atlantic salmon Salmo salar and immune response of salmon to this agent. Diseases of
Aquatic Organisms 60(2): 97–103.
Björnsdóttir, B; Gudmundsdottir, T; Gudmundsdottir, B K (2011). Virulence properties of Moritella viscosa
extracellular products. Journal of Fish Diseases 34(5): 333–343 [Online] Available from:
https://www.doi.org/10.1111/j.1365-2761.2011.01246.x [Accessed 22 January 2020].
Black, H (1997). Pasteurella isolates from sheep pneumonia cases in New Zealand. Surveillance 24(3): 5–8.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 117
Blackwell, R G (2013). Bait fish use in New Zealand: Recent trends. Memorandum prepared for the Animal
Imports team, Ministry for Primary Industries, Wellington, New Zealand (Unpublished report) 22 p.
Blackwell, R G (2019). Import risk analysis: Eviscerated or trunked fish for human consumption. Ministry for
Primary Industries, Wellington, New Zealand, 527 p.
Blake, S; Ma, JY; Caporale, D A; Jairath S; Nicholson B L (2001). Phylogenetic relationships of aquatic
birnaviruses based on deduced amino acid sequences of genome segment A cDNA. Diseases of Aquatic Organisms
45(2): 89–102 [Online] Available from: http://www.doi.10.3354/dao045089 [Accessed 21 October 2019].
Blindheim, S; Nylund, A; Watanabe. K; Plarre. H; Erstad, B; Nylund S (2015). A new aquareovirus causing
high mortality in farmed Atlantic halibut fry in Norway. Archives of Virology 10: 91–102.
Bootland, L M; Leong, J C (2011). Infectious haematopoietic necrosis. pp 66–109. In: Woo, P T K; Bruno, D W
(Eds.), Fish Diseases and Disorders: Volume 3 - Viral, Bacterial and Fungal Infections, 2nd edition. CABI
International, Wallingford, United Kingdom, 930 p.
Boran, H; Terzi, E; Altinok, I; Capkin, E; Bascinar, N (2013). Bacterial diseases of cultured Mediterranean
Horse mackerel (Trachurus mediterraneus) in sea cages. Aquaculture 396: 8-13.
Boreham, R E; Hendrick, S; O’Donoghue, P J; Stenzel, D J (1998). Incidental finding of Myxobolus spores
(Protozoa: Myxozoa) in stool samples from patients with gastrointestinal symptoms. Journal of Clinical
Microbiology 36: 3728–3730.
Borrego, J J; Valverde, EJ; Labella, A M; Castro, D (2017). Lymphocystis disease virus: its importance in
aquaculture. Aquaculture 9(2): 179–193 [Online] Available from: https://doi.org/10.1111/raq.12131 [Accessed 16
December 2019].
Boustead, N C (1982). Fish diseases recorded in New Zealand, with a discussion on potential sources and
certification procedures. Fisheries Research Division Occasional Publication 34, Ministry of Agriculture and
Fisheries, Wellington, New Zealand. 19 p.
Boustead, N C; Meyers, T R; Short, S (1993). Absence of infectious haematopoietic necrosis virus (IHNV) in
New Zealand sockeye salmon, Oncorhynchus nerka. New Zealand Journal of Marine and Freshwater Research 27:
55–60.
Bovo, G; Hill, B; Husby, A; Håstein, T; Michel, C; Olesen, L; Storset, A; Midtlyng, P J (2005). Pathogen
survival outside the host, and susceptibility to disinfection: Work package 3 report. Final report for project VESO-
1601, prepared by VESO, Norway for the European Commission, Brussels, 56 p [Online] Available from:
https://pdfs.semanticscholar.org/c396/ed2286949bfedd94842ee23f018a81ee9ade.pdf [Accessed 30 October 2019].
Bowden, T J (2008). Modulation of the immune system of fish by their environment. Fish and Shellfish
Immunology 25: 373–83.
Bowser, P R; Winton, J R; Batts, W N; Cangelosi, A; Casey, J W (2010). Distribution of an invasive aquatic
pathogen (viral haemorrhagic septicaemia virus) in the Great Lakes and its relationship to shipping. PLoS ONE 5
(4): e10156 [Online] Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2854150/ [Accessed 23 April
2015].
Braden. L M; Rasmussen, K J; Purcell, S L; Ellis, L; Mahoney, A; Cho, S; Whyte, S; Jones, S R M; Fast, M
D (2017). Acquired protective immunity in Atlantic salmon Salmo salar against the myxozoan, Kudoa thyrsites,
involves induction of MHIIβ+/CD83+ antigen presenting cells. Infection and Immunity 86(1): 1–18 [Online]
Available from: http://www.doi:10.1128/IAI.00556-17 [Accessed 6 December 2019].
Brady, Y J; Vinitnantharat, S (1990). Viability of bacterial pathogens in frozen fish. Journal of Aquatic Animal
Health 2(2): 149-150.
Bromage, E S; Thomas, A; Owens, L (1999). Streptococcus iniae, a bacterial infection in barramundi Lates
calcarifer. Diseases of Aquatic Organisms 36: 177–181.
Bromage, E S; Owens, L (2002). Infection of barramundi Lates calcarifer with Streptococcus iniae: effects of
different routes of exposure. Diseases of Aquatic Organisms 52: 199–205.
Brosnahan, C L; Ha, H J; Booth, K; McFadden, A M J; Jones, J B (2017). First report of a rickettsia‐like
organism from farmed Chinook salmon, Oncorhynchus tshawytscha (Walbaum), in New Zealand. New Zealand
Journal of Marine and Freshwater Research 51: 356–369 [Online] Available
from: https://doi.org/10.1080/00288330.2016.12420 [Accessed 22 October 2019].

118 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Brosnahan, C L; Munday, J S; Jeong-Ha, H; Preece, M; Jones, J B (2018a). New Zealand rickettsia‐like
organism (NZ‐RLO) and Tenacibaculum maritimum: Distribution and phylogeny in farmed Chinook salmon
(Oncorhynchus tshawytscha). Journal of Fish Diseases 42(1): 85-95 [Online] Available from:
https://doi.org/10.1111/jfd.12909 [Accessed 15 August 2019].
Brosnahan, C L; Pande, A; Keeling, S E; van Andel, M; Jones, J B (2018b). Lamprey (Geotria australis;
Agnatha) reddening syndrome in Southland rivers, New Zealand 2011–2013: laboratory findings and epidemiology,
including the incidental detection of an atypical Aeromonas salmonicida. New Zealand Journal of Marine and
Freshwater Research 53(3): 416–436 [Online] Available from: http://www.doi.1080/00288330.2018.1556167
[Accessed 1 November 2019].
Buchanan, J T; Colvin K M; Vicknair, M R; Patel, S K; Timmer, A M; Nizet, V (2008). Strain-associated
virulence factors of Streptococcus iniae in hybrid-striped bass. Veterinary Microbiology 131(1–2): 145–153
[Online] Available from: https://www.doi.org/10.1016/j.vetmic.2008.02.027 [Accessed 14 January 2020].
Buller, N B (2014). Bacteria and fungi from fish and other aquatic animals, 2nd edition: A practical identification
manual. CABI International, Wallingford, United Kingdom, 919 p.
Bunch, E C; Bejerano I (1997). The effect of environmental factors on the susceptibility of hybrid tilapia
Oreochromis niloticus × Oreochromis aureus to streptococcosis. The Israeil Journal of Aquaculture 49(2): 67–76.
Burge, C A; Eaken, C M; Friedman, C S; Froelich, B; Hershberger, P K; Hofmann, E E; Petes, L E; Prager,
K C; Weil, E; Wiis, B L; Ford, S E; Harvell, C D (2014). Climate change influences on marine infectious
diseases: implications for management and society. Annual Reviews in Marine Science 6: 249–277 [Online]
Available from: http://www.doi.10.1146/annurev-marine-010213-135029 [Accessed 3 April 2020].
Burger, M A; Barnes, A C; Adlard, R D (2008). Wildlife as reservoirs for parasites infecting commercial species:
host specificity and a redescription of Kudoa amamiensis from teleost fish in Australia. Journal of fish Diseases
31(11): 835–844 [Online] Available from: http://www.doi.10.1111/j.1365-2761.2008.00958.x [Accessed 1
November 2019].
Burke, J; Rodgers, L (1981). Identification of pathogenic bacteria associated with the occurrence of ‘red spot’ in
sea mullet, Mugil cephalus L., in south‐eastern Queensland. Journal of Fish Diseases 4(2): 153–159.
Burney, B (2019). Pendarves Ltd, Auckland, New Zealand. Personal communication.
Bustos, P; Rozas, M; Bohle, H; Ildenfonso, R; Sandoval, A; Gaete, A; Araya, C; Grothusen, H; Tapia, E;
Gallardo, A; Rojas, M; Lara, M; Labra, A; Gálvez, C (2011). Primer reporte de Piscine Reovirus (PRV) en
salmón del atlántico, Salmo salar, cultivado en Chile. Versión Diferente 15: 65–69.
CABI, (2015). Edwardsiella septicaemia (Edwardsiella tarda infection). Pathogen data sheet. CAB International,
Wallingford, United Kingdom [Online] Available from: http://www.cabi.org/isc/datasheet/84398 [Accessed 12
August 2015].
CABI, (2019a). Invasive Species Compendium: infectious pancreatic necrosis virus. CAB International,
Wallingford, United Kingdom [Online] Available from: https://www.cabi.org/isc/datasheet/79273 [Accessed 30
October 2019].
CABI, (2019b). Invasive Species Compendium: infectious salmon anaemia. CAB International, Wallingford,
United Kingdom [Online] Available from: http://www.cabi.org/isc/datasheet/79288#549B8683-3597-40D5-9D34-
49478C92FD9C [Accessed 30 October 2019].
Cairns, S D; Gershwin, L A; Brook, F; Dawson, E W; Oscar, O V; Vervoot, W; Williams, G; Watson, J;
Opresco, D M; Shuhert, P; Hine, M P; Gordon, D P; Campbell, H J; Wright, A J; Sanchez, J; Fautin, D G
(2009). New Zealand Inventory of Biodiversity. Volume 1. Kingdom Animalia: Radiata, Lophotrochozoa,
Deuterostomia, Canterbury University Press, Christchurch, New Zealand [online] Available from:
https://repository.si.edu/handle/10088/8431 [Accessed 20 September 2019].
Callinan, R B; Keep, J A (2006). Bacteriology and parasitology of red spot disease in sea mullet, Mugil cephalus
L., from eastern Australia. Journal of Fish Diseases 12: 349–356.
Callinan, R B; Fraser, G C; Virgona, J L (2006). Pathology of red spot disease in sea mullet, Mugil cephalus L.,
from eastern Australia. Journal of Fish Diseases 12(5):467–479.
Campbell, N (2005). The myxosporean parasitofauna of the Atlantic horse mackerel, Trachurus trachurus (L.) in
the North-East Atlantic Ocean and Mediterranean Sea. Acta Parasitologica 50(2): 97–101.
Cardenas, C; Carmona, M; Gallardo, A; Labra, A; Marshall, S H (2014). Coexistence in field samples of two
variants of the infectious salmon anaemia virus: a putative shift to pathogenicity. PLoS One, 9, e87832 [Online]
Available from: http://www.doi.10.1371/journal.pone.0087832 [Accessed 30 October 2019].

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 119
Carlile, G; East, I J; McColl, K A; Ellard, K; Browning, G F; Crane, M St J (2014). The spatial and temporal
variation of the distribution and prevalence of Atlantic salmon reovirus (TSRV) infection in Tasmania, Australia.
Preventive Veterinary Medicine 116: 214–219.
Carson, J; Gudkovs, N; Austin, B (1993). Characterisatics of an Enterococcus-like bacterium from Australia and
South Africa, pathogenic for rainbow trout, Oncorhynchus mykiss (Wallbaum). Journal of Fish Diseases 16: 381–
388.
Castellanos-Martinez, S; Perez-Losanda, M; Gestel, C (2013). Molecular phylogenetic analysis of the coccidian
cephalopod parasites Aggregata octopiana and Aggregata eberthi (Apicomplexa: Aggregatidae) from the NE
Atlantic coast using 18S rRNA sequences. European Journal of Protozoology 49 (3): 373– 380 [Online] Available
from: http://www.doi.10.1016/j.ejop.2012.11.005 [Accessed 9 May 2019].
Castellanos-Martinez, S; Gestal, C; Pascual, S; Mladineo, I; Azevedo, C (2019). Protist (Coccidia) and related
diseases, pp 153– 158 In: Gestal, C; Pascual, S; Guerra, A; Fiorito, G; Vietes J M (Eds.). Handbook of diseases in
Cephalopods. Springer Science, New York, United States of America, 230 p.
Castric, J (1997). Viral diseases in fish mariculture. Bulletin of the European Association of Fish Pathologists
17(6): 220–228.
Castro, N; Toranzo, A E; Barja, J L; Nunez, S; Magarinos, G B (2006). Characterization of Edwardsiella tarda
strains isolated from turbot, Psetta maxima (L.). Journal of Fish Diseases 29(9): 541– 547.
Chanet, B (2003). Interrelationships of scophthalmid fishes (Pleuronectiformes: Scophthalmidae). Cymbium 27(4):
275–286.
Chang, E S; Neuhof, M; Rubenstein, N D; Diamant, A; Philppe, H; Huchon, D; Cartwright, P (2015).
Genomic insights into the evolutionary origin of Myxozoa within Cnidaria. Proceedings of the National Academy of
Sciences of the United States 112(48): 14912–14917 [Online] Available from:
https://doi.org/10.1073/pnas.1511468112 [Accessed 1 November 2019].
Chen, M F; Henry-Ford, D; Groff, J M (1995). Isolation of Flexibacter maritimus from California. Fisheries
Health Society (FHS)/American Fisheries Society (AFS) Newsletter 22: 7–11.
Chen, S C; Liaw, L L; Su, H Y; Wu, C Y; Chaung, H C; Tsai, Y H; Yang, K L; Chen, Y C; Chen, T H; Lin,
G R; Chen, S Y; Lin, Y D; Lee, J L; Lai, C C; Weng, Y J; Chu, S Y (2002). Lactococcus garvieae, a cause of
disease in grey mullet, Mugil cephalus L., in Chinese Taipei. Journal of Fish Diseases 25(12): 727–732.
Chen, S L (2019). Genomic insights into the distribution and evolution of Group B Streptococcus. Frontiers of
Microbiology 10: 3389 [Online] Available from: https://www.frontiersin.org/article/10.3389/fmicb.2019.01447
[Accessed 16 July 2020].
Chen, Y; Jiang, Y (1984). Morphological and physio-chemical characterisation of the hemorrhagic virus of grass
carp. Chinese Science Bulletin 29: 832–835.
Cherif, N; Thiery, R; Castric, J; Biarchesi, S; Bremont, M; Thabti, T; Liemen, L; Hammani, S (2008). Viral
encephalopathy and retinopathy of Dicentrarchus labrax and Sparus aurata farmed in Tunisia. Veterinary Research
Communications 33: 345–353.
Chernova, T N (1967). To the studies of myxosporidia fauna in fish of some water reservoirs of the western
Georgia. Trudy nauch.-issled. ribokhozyaystvennoy stantsii Gruzii. 12: 12–20 (In Russian) (cited in Ozer A, Kirca
DY (2013). Parasite fauna of Golden Grey Mullet Liza aurata (Risso, 1810) collected from Lower Kızılırmak Delta
in Samsun, Turkey. Helminthologia 50(4): 269–280 [Online] Available from: http://www. DOI 10.2478/s11687-
013-0140-4 [Accessed 12 September 2019].
Chi, S C; Wu, YC; Hog, J R (2016). Nodaviruses of fish. pp, 371–393 In: Kibenge, F S B; Godoy, M C (Eds.).
Aquaculture Virology. Academic Press, New York, United States of America, 549 p.
Choi JC; Kim YC; Choi H J (2015). Detection and Analysis of the Potential Risk of VHSV in Bivalves in Korea.
Journal of Fish Pathology 28(1): 27–35 [Online] Available from: http://www.doi.10.7847/jfp.2015.28.1.027
[Accessed 22 October 2020].
Christiansen, D B; Mcbeath, A J A; Aamelfot, M; Matejusova, I; Fourrier M; White P; Petersen P E; Falk K
(2017). First field evidence of the evolution from a non-virulent HPR0 to a virulent HPR-deleted infectious salmon
anaemia virus. Journal of General Virology 98: 595–606.
Chua J, Bozue J A; Klimko, C P; Shoe, J L; Ruiz, S L; Jensen, C L; Tobery, S A; Crumpler, J M; Chabot, D
J; Quirk, A V; Hunter, M; Harbourt, D E; Friedlander, A M; Cote, C K (2019). Formaldehyde and
glutaraldehyde inactivation of bacterial tier 1 select agents in tissues. Emerging Infectious Diseases 25(5): 919–926
[Online] Available from: https://doi.org/10.3201/eid2505.180928 [Accessed 29 August 2019].

120 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
CIFA, (2019). Susceptible species of aquatic animals. Canadian Food Inspection Agency, Government of Canada,
Ottawa, Canada [Online] Available from: https://micor.agriculture.gov.au/Fish/Documents/2014_01_10_-
_Canada_-_Susceptible_Species_of_Aquatic_Animals_list.pdf [Accessed 14 June 2019].
Ciric, L; Philp, J C; Whitely, S (2009). Hydrocarbon utilisation within a diesel-degrading bacterial consortium.
FEMS Microbiology Letters 303(2): 116–122.
Clark, M; Haisman-Welsh, R; White, E; Houliston, P (2018). Infection prevention and control and management
of carbapenemase-producing Enterobacteriaceae: Guidelines for health care providers in New Zealand acute and
residential care facilities, Ministry of Health, Wellington, New Zealand, 59 p. [Online] Available from:
https://www.health.govt.nz/system/files/documents/publications/infection-prevention-control-management-
carbapenemase-producing-enterobacteriaceae-dec18.pdf [Accessed 21 October 2019].
Clark, R (2019). Marine Foods and Bulk Baits, Tauranga, New Zealand. Personal communication.
Clayton, J S; Wells, R D S (1999). Some issues in risk assessment reports on grass carp and silver carp.
Conservation Advisory Science Notes No. 257. Department of Conservation, Wellington, 16 p.
Cobb, S (2008). Import risk analysis: Fish food. Ministry of Agriculture & Forestry, Wellington, New Zealand, 57p.
Colquhoun, D J; Duodu, S (2011). Francisella infections in farmed and wild aquatic organisms. Veterinary
Research 42(1): 47 [Online] Available from: http://www.veterinaryresearch.org/content/42/1/47 [Accessed 24
September 2014].
Conlan, J W; Chen, W; Bosio, C N; Cowley, S C; Elkins, K L (2011). Infection of mice with Francisella as an
immunological model. Current Protocols in Immunology 93(1): 19:4.1 to 19: 4.6.
Cools, P; Haelters, J; dos Santos-Santiago, G L; Claeys G; Bolens, J; Leroux-Roels, I; Vaneechoutte M;
Deschaght P (2013). Edwardsiella tarda sepsis in a live-stranded sperm whale. (Physeter macrocephalus).
Veterinary Microbiology 166: 311–315 [Online]. Available from:
http://www.dx.doi.org/10.1016/j.vetmic.2013.05.020 [Accessed 29 January 2020].
Corsin, F; Georgiadis, M; Hammell, K L; Hill B (2009). Guide for Aquatic Animal Health Surveillance. The
World Organisation for Animal Health (OIE), Paris, France, 126 p [Online] Available from:
https://www.oie.int/doc/ged/d6714.pdf [Accessed 30 June 2020].
Costa, J Z; Thompson, K D (2016). Understanding the interaction between betanodavirus and its host for the
development of prophylactic measures for viral encephalopathy and retinopathy. Fish and Shellfish Immunity 53:
35–49.
Crane, M St J; Williams, L M (2008). Viruses of salmonids: Virus isolation in fish cell lines. Australia and New
Zealand Standard Diagnostic Procedure, AAHL Fish Diseases Laboratory, CSIRO, Canberra, Australia, 36 p.
Crane M St J, Carlile F (2010). Aquareoviruses. pp 429–437, In: Mahy, B W H; van Regenmortel, M H V (Eds.).
Desk encyclopaedia of animal and bacterial virology. Academic Press, New York, United States of America, 646 p.
Crane, M St J; Hyatt, A (2011). Viruses of fish: an overview of significant pathogens. Viruses 3(11): 2025–2046.
Crane, M St J; Smith, P H; Williams, L M; Hyatt, A D; Eaton, L M; Gould, A; Handlinger J; Kattenbelt J;
Gudkovs, N (2000). First isolation of an aquatic birnavirus from farmed and wild fish species in Australia. Diseases
of Aquatic Organisms 43: 1–14.
Crawford, L M; Ruff, E H (1996). A review of the safety of cold pasteurization through irradiation. Food Control
7(2): 87–97 [Online] Available from: https://doi.org/10.1016/0956-7135(96)00004-7 [Accessed 21 October 2020].
Crockford, M; Jones, J B; Crane, M S J; Wilcox, G E (2005). Molecular detection of a virus, Pilchard
herpesvirus, associated with epizootics in Australasian pilchards Sardinops sagax neopilchardus. Diseases of
Aquatic Organisms 68: 1–5.
Cunningham, C O; Gregory, A; Black, J; Simpson, I; Raynard, R S (2002). A novel variant of the infectious
salmon anaemia virus (ISAV) hemagglutinin gene suggests mechanisms for virus diversity. Bulletin of the European
Association of Fish Pathologists 22: 366–374.
DAFF, (2005). Disease strategy: Viral haemorrhagic septicaemia (Version 1.0), In: Australian aquatic veterinary
emergency plan (AQUAVETPLAN), Edition 2. DAFF (Department of Agriculture, Fisheries and Forestry),
Canberra, ACT, Australia, 63 p.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 121
DAFF, (2013). Gamma irradiation as a treatment to address pathogens of animal biosecurity concern: draft review,
Biosecurity advice 2013/04. Department of Agriculture, Fisheries and Forestry Biosecurity, Canberra, Australia
[Online] Available from:
https://www.agriculture.gov.au/sites/default/files/sitecollectiondocuments/ba/animal/2013/gamma-
irradiation/BA2013-04-Gamma-irradiation.pdf [Accessed 10 January 2020].
DAFF, (2014). Viral haemorrhagic septicaemia. Aquatic animal diseases significant to Australia: Identification field
guide 4th edition. DAFF (Department of Agriculture, Fisheries and Forestry), Canberra, ACT, Australia. [Online]
Available from: http://www.agriculture.gov.au/SiteCollectionDocuments/animal-plant/aquatic/field-guide/4th-
edition/aquatic-disease-user-guide-edition-4.pdf. [Accessed 10 January 2020].
Dale, O B; Orpetveit, I; Lyngstad, T M; Kahn, S; Skall, H F; Olesen, N J; Dannevig, B H (2009). Outbreak of
viral haemorrhagic septicaemia (VHS) in seawater-farmed rainbow trout in Norway caused by VHS virus Genotype
III. Diseases of Aquatic Organisms 85: 93–103.
Dalton, R (2004). Aquaculture: Fishing for trouble. Nature 431 (7008): 502–504.
Daly, J G (1999). Other bacterial pathogens, pp 577 –597. In: Woo, P T K; Bruno, D W (Eds.) Fish diseases and
disorders. Vol. 3, Viral, Bacterial and fungal infections. CAB International, London, New York, United States of
America, 930 p.
Daly, J G; Aoki, T (2011). Pasteurellosis and other bacterial diseases, pp 648–668. In: Woo, P T K; Bruno, D W
(Eds.). Fish diseases and disorders: Volume 3 - Viral, bacterial and fungal infections, 2nd edition. CAB
International, Wallingford, United Kingdom, 930 p.
Dannevig, B H; Thorud, K E (1999). Other viral diseases and agents of cold-water fish: infectious salmon
anaemia, pancreas disease and viral erythrocytic necrosis, pp 164–175. In: Woo, P T K; Bruno, D W (Eds.). Fish
diseases and disorders, volume 3. CAB International, New York. United States of America. 930 p.
Davies, A J; Curtis, L; Grutter, A S; Smit, N J (2009). Suspected viral erythrocytic necrosis (VEN) in a juvenile
blackbar triggerfish Rhinecanthus aculeatus, from Lizard Island, Great Barrier Reef, Australia. Marine Biodiversity
Records 2(1): e149 [Online] Available from: http://www.doi.10.1017/S1755267209990674 [Accessed 21 January
2020].
Davies, K R; McColl, K A; Wang, L F; Yu, M; Williams, L M; Crane, M St J (2010). Molecular
characterisation of Australasian isolates of aquatic birnaviruses. Diseases of Aquatic Organisms 93: 1–15.
Davies, Y M; Xavier de Oliveira, M G; Cunha, M P V; Franco, L S; Santos, S L P; Moreno, L Z; de Moura,
V; Gomes, V T; Sato, M I Z; Nardi, M S; Moreno, A M; Saidenberg, A B; Marques de Sa, L R; Knobl, T
(2018). Edwardsiella tarda outbreak affecting fishes and aquatic birds in Brazil. Veterinary Quarterly 38(1): 99–105
[Online] Available from: http://www.doi.10.1080/01652176.2018.1540070 [Accessed 4 May 2020].
DAWR, (2016). Offshore irradiation treatment providers scheme. Department of Agriculture and Water Resources,
Canberra, Australia [Online] Available from: http://www.agriculture.gov.au/import/before/prepare/treatment-
outside-australia [Accessed 25 May 2020].
Delannoy, C M; Crumlish, M; Fontaine, M C (2013). Human Streptococcus agalactiae strains in aquatic
mammals and fish. BMC Microbiology 13(1): 41 [Online] Available from:
https://bmcmicrobiol.biomedcentral.com/articles/10.1186/1471-2180-13-41 [Accessed 16 July 2020].
DermNet NZ, (2014). Database on diseases of the skin. New Zealand Dermatological Society [Online] Available
from: http://www.dermnetnz.org/about-us/ [Accessed 21 October 2019].
Desautels, D; MacKelvie, R M (1975). Practical aspects of survival and destruction of infectious pancreatic
necrosis virus. Journal of the Fisheries Research Board of Canada 32: 523–531.
Devauchelle, G; Vago, C (1971). (Particlues d’allure virale dans les cellules de l’estomac de la seiche) Particles of
viral aspect in stomach cells of the cuttlefish Sepia officinalis L. (Mollusca, Cephalopoda). CR Hebd Seances,
Academy of Science, Paris, France 272: 894–896 (In French, English abstract).
DFO, (2009). Stock assessment report on Pacific herring in British Columbia. Science Advisory Report 2009/059.
Canadian Science Advisory Secretariat, Department of Fisheries and Oceans, Ottawa, Canada, 14 p [Online]
Available from: http://www.dfo-mpo.gc.ca/Library/338390.pdf [Accessed 17 December 2019].
DFO, (2017). Residual infectious pancreatic necrosis (IPN) transmission risk from arctic char transfers into British
Columbia. Pacific Region Science Response 2017/041. Canadian Science Advisory Secretariat, Department of
Fisheries and Oceans, Ottawa, Canada, 34 p. [Online] Available from: https://waves-vagues.dfo-
mpo.gc.ca/Library/40655441.pdf [Accessed 10 January 2020].

122 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Diamanka, A; Fall, M; Diebakate, C; Faye, N; Toguebaye, B S (2008). Identification of Myxobolus episquamalis
(Myxozoa, Myxobolidae) in flathead grey mullet Mugil cephalus (Pisces, Teleostei, Mugilidae) from the coast of
Senegal (eastern tropical Atlantic Ocean). Acta Adriatica 49: 19–23.
Diamant, A (1997). Fish-to-fish transmission of a marine myxosporean. Diseases of Aquatic Organisms 30(2):
99–105 [Online] Available from: http://www.doi.10.3354/dao030099 [Accessed 13 September 2019].
Diamant, A; Colorni, A; Ucko, M (1997). Parasite and disease transfer between cultured and wild coastal marine
fish. Impact of mariculture on coastal ecosystems, Lisbon, 21-24 February 2007. CIESM Workshop Monograph 32:
49–53 [Online] Available from: http://ciesm.org/online/monographs/32/WM_32_49_53.pdf [Accessed 28 July
2020].
Diamant, A; Lom, J; Dyková, I (1994). Myxidium leei n. sp., a pathogenic myxosporean of cultured sea bream
Sparus aurata. Diseases of Aquatic Organisms 20: 137–141.
Diamant, A; Ram, S; Paperna, I (2006). Experimental transmission of Enteromyxum leei to freshwater fish.
Diseases of Aquatic Organisms 72: 171–178.
Diamant, A; Ucko, M; Paperna, I; Colorni, A; Lipshitz, A (2005). Kudoa iwatai (Myxosporea: Multivalvulida)
in wild and cultured fish in the Red Sea: Redescription and molecular phylogeny. Journal of Parasitology 91(5):
1175–1189 [Online] Available from: https://doi.org/10.1645/GE-491R.1 [Accessed 18 September 2019].
Diggles, B K; Hine, P M; Handley, S; Boustead, N C (2002). A handbook of diseases of importance to
aquaculture in New Zealand. NIWA Science and Technology Series 49. National Institute of Water and Atmospheric
Research, Auckland, New Zealand, 200 p.
Diggles, B K (2003). Import risk assessment: Juvenile yellowtail kingfish (Seriola Lalandi) from Spencer Gulf
aquaculture, South Australia. NIWA Client Report: WLG 2002/03. National Institute of Water and Atmospheric
Research, Wellington, New Zealand, 54 p.
Diggles, B K (2004a). Health assessment of cultured kingfish (Seriola lalandi) from Crail Bay, January 2004.
DigsFish Pathology Services Client Report DF04/ 03, 17 p.
Diggles, B K (2004b). Parasites as potential stock discriminators of kingfish (Seriola lalandi) (Preliminary Report).
DigsFish Pathology Services Client Report DF04/13, 12 p.
Diggles, B K (2007). Scientific submission to AQIS supporting an application for an import permit allowing entry
of whole, round menhaden (Brevoortia tyrannus) for use as lobster bait. DigsFish Services Client Report DF07-01,
39 p.
Diggles, B K (2011). Risk analysis: Aquatic animal diseases associated with domestic bait translocation. Report
prepared for FRDC Project 2009/072, DigsFish Services, Banksia Beach, Queensland, Australia, 296 p [Online]
Available from: https://apfa.com.au/wp-content/uploads/2017/03/Bait-and-Burley-pathogen-RA.pdf {Accessed 9
September, 2020]’
Diggles, B K (2016). Updated disease risk assessment report – relocation of salmon farms in Marlborough Sounds,
New Zealand. Report DF 16-01 prepared for the Ministry for Primary Industries, New Zealand, by DigsFish
Services, Queensland, Australia, 80 p.
Diggles, B K; Hine, P M; Handley, S; Boustead, N (2002). A handbook of diseases important to New Zealand.
NIWA Science and Technology Series 49, National Institute of Marine and Freshwater Research, Auckland, New
Zealand, 200 p.
Dixon, P F (1999). VHSV came from the marine environment: clues from the literature, or just red herrings.
Bulletin of the European Association of Fish Pathologists 19(2): 60–65.
Dixon, P F; Feist, S; Kehoe, E; Parry, L; Stone, D M; Way, K (1997). Isolation of viral haemorrhagic
septicaemia virus from Atlantic herring Clupea harengus from the English Channel. Diseases of Aquatic Organisms
30: 81–89.
Dixon, P; Paley, R; Alegri-Moran, R; Oidtmann, B (2016). Epidemiological characteristics of infectious
hematopoietic necrosis virus (IHNV): a review. Veterinary Research 47(63): 1–26 [Online] Available from:
http://www.doi.10.1186/s13567-016-0341-1 [Accessed 8 June 2018].
Dmitrieva, E V; Gaevskaya, A V (2001). Parasitological aspects of mugilid mariculture and of their introduction
into the Sea of Azov and the Black Sea. Ecologiya Morya 55: 73–78.
Doan, Q K; Vandeputte, M; Chatain, B; Morin, T; Allal, F (2017). Viral encephalopathy and retinopathy in
aquaculture: a review. Journal of Fish Diseases 40: 717–742 [Online] Available from:
http://www.doi:10.1111/jfd.1254 [Accessed 20 August 2019].

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 123
Dobson, A P; May, R M (1987). The effects of parasites on fish populations—theoretical aspects. International
Journal for Parasitology 17(2): 363–370 [Online] Available from: https://doi.org/10.1016/0020-7519(87)90111-1
[Accessed 3 April 2020].
Duignan, P J; Hine, P M; Joy, M; Gibbs, N; Jones, G W; Okeoma, C (2003). Disease surveillance in freshwater
fish from the lower North Island. Surveillance 30(3): 6–8.
Dunn, A M; Torchin, M E; Hatcher, M J; Kotanen, PM; Blumenthal, D M; Byers, J E; Coon, C A C;
Frankel, V M; Holt, R D; Hufbauer, R A; Kanarek, A R; Schierenbeck, K A; Wolfe, L M; Perkins, S E
(2012). Indirect effects of parasites in invasions. Functional Ecology 26(6): 1262–1274 [Online] Available from:
https://www.doi.org/10.1111/j.1365-2435.2012.02041.x [Accessed 7 January 2020].
EFSA, (2012). Scientific opinion on infectious salmon anaemia (ISA). EFSA Journal 10(11): 2971. EFSA Panel on
Animal Health and Welfare, European Food Safety Authority, 22 p.
Einer-Jensen, K; Ahrens, P; Forsberg, R; Lorenzen, N (2004). Evolution of the fish rhabdovirus viral
haemorrhagic septicaemia virus. Journal of General Virology 85(5): 1167–1179.
Eiras, J C; D’Souza, J (2004). Myxobolus goensis n.sp. (Myxozoa, Myxosporea, Myxobolidae), a parasite of the
gills of Mugil cephalus (Osteichthyes, Mugilidae) from Goa, India. Parasite 11: 243–248.
Eiras, J C; Molnar, K; Lu, Y S (2005). Synopsis of the species of Myxobolus Butschli, 1882 (Myxozoa:
Myxosporea: Myxobolidae). Systematic Parasitology 61: 1–46 [Online] Available from: http://www.doi. DOI
10.1007/s11230-004-6343-9 [Accessed 17 September 2019].
Egusa, S; Nakajima, K (1980). Kudoa amamiensis n. sp. (Myxosporea: Multivalvulida) found in cultured
yellowtails and wild damselfishes from Amami-Ohshima and Okinawa, Japan. Nippon Suisan Gakkaishi 46(10):
1193–1198 [Online] Available from: https://doi.org/10.2331/suisan.46.1193 [Accessed 1 November 2019].
Egusa, S; Shiomitsu, T (1983). Studies on myxosporidian parasites of the genus Kudoa found in marine fish
cultured in Japan. Fish Pathology 18(3): 163–171.
Eldar, A; Bejerano, Y; Livoff, A; Horovitcz, A; Bercovier, H (1995). Experimental streptococcal meningo-
encephalitis in cultured fish. Veterinary Microbiology 43(1): 33–40.
Ellis, D; Kiessling, I (2016). Ranching of Southern Bluefin Tuna in Australia, Chapter 9 pp 217– 232 In: Advances
in Tuna Aquaculture, Academic Press, New York, United States of America [Online] Available from:
http://www.doi.10.1016/B978-0-12-411459-3.00010-2 [Accessed 21 October 2020].
El-Matbouli, M; Hoffmann, R W (1991). Effects of freezing, aging and passage through the alimentary canal of
predatory animals on the viability of Myxobolus cerebralis spores. Journal of Aquatic Animal Health 3: 260–262.
Emmenegger, E J; Meyers, T R; Burton, T O; Kurath, G (2000). Genetic diversity and epidemiology of
infectious hematopoietic necrosis virus in Alaska. Diseases of Aquatic Organisms 40: 163 – 176.
Emmenegger, E J; Glen, J A; Winton, J R; Batts, W N (2014). Molecular identification of erythrocytic necrosis
virus (ENV) from the blood of Pacific herring (Clupea pallasii). Veterinary Microbiology 174: 16–26.
Engelhard, G C; Peck, M A; Rindorf, A; Smout, S C; van Deurs, M; Raab, K; Anderson, K H; Garthe, S;
Lauerberg, R A M; Scott, F; Brunel, T; Aarts, G; van Kooten, T; Dickey-Collas, M (2014). Forage fish, their
fisheries and their predators: Who drives whom? ICES Journal of Marine Science 71(1): 90–104.
EPA, (2015). Database of organisms present in New Zealand, Environmental Protection Authority, Wellington,
New Zealand [Online] Available from:
https://www.epa.govt.nz/assets/Uploads/Documents/NewOrganisms/Guidance/microbes.pdf [Accessed 1 November
2019].
Eszterbauer, E; Székely, C S; Molnár, K; Baska, F (2001). Development of Myxobolus bramae (Myxosporea:
Myxobolidae) in an oligochaete alternate host, Tubifex tubifex. Journal of Fish Diseases 23(1): 19–25.
Evans, J J; Klesius, P H; Gilbert, P M; Shoemaker, C A; Sarawi, M A; Landsberg, J; Duremdez, R;
Marjouk, A; Zenki, S A (2002). Characterization of b-haemolytic Group Β Streptococcus agalactiae in cultured
sea bream Sparus aurata L., and wild mullet Liza klunzingeri (Day) in Kuwait. Journal of Fish Diseases 25: 505–
509.
Evans, J J; Klesius, P H; Plumb, J A; Shoemaker, C A (2011). Edwardsiella septicaemias pp 512-569 In: Woo,
P T K; Bruno, D W (Eds.). Fish diseases and disorders; Volume 3 – Viral, bacterial and fungal infections, 2nd
edition, CAB International, Wallingford, United Kingdom, 930 p.
Evans, J J; Wiedenmayer, A A; Klesius, P H; Shoemaker, C A (2004). Survival of Streptococcus
agalactiae from frozen fish following natural and experimental infections. Aquaculture 233: 15–21.

124 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Evelyn, T P T (1993). Bacterial kidney disease, pp 177–195 In: Inglis V, Roberts RJ, Bromage NR (Eds.) Bacterial
diseases of fish. Halstead Press, New York, United States of America.
Evelyn, T P T (2001). The effects of chilling, freezing and cold-smoking on the infectious titre of certain microbial
fish pathogens that may occasionally be present in marketed salmonid flesh, pp 215–229 In: Rodgers, C J (Ed.) Risk
analysis in aquatic animal health - Proceedings of an international conference. World Organisation for Animal
Health (OIE); Paris, France.
Evelyn, T P T; Traxler, G S (1978). Viral erythrocytic necrosis: Natural occurrence in Pacific salmon and
experimental transmission. Journal of the Fisheries Research Board of Canada 35(6): 903–907 [Online] Available
from: https://doi.org/10.1139/f78-145 [Accessed 22 August 2019].
Fadel, A; Mabrok, H; Aly, S (2018). Epizootics of Pseudomonas anguilliseptica among cultured seabream (Sparus
aurata) populations: Control and treatment strategies. Microbial Pathogenesis 121: 1–8. [Online] Available from:
http://www.doi.10.1016/j.micpath.2018.04.021 [Accessed 9 September 2019].
Faisal, M; Shavalier, M; Kim, R K; Millard, E V; Gunn, M R; Winters, A D; Schulz, C A; Eissa, A; Thomas,
M V; Wolgamood, M; Whelan, G E; Winton, J (2012). Spread of the emerging viral hemorrhagic septicaemia
virus strain, genotype IVb, in Michigan, USA. Viruses 4(5): 734–760 [Online] Available from:
http://www.10.3390/v4050734 [Accessed 30 October 2019].
Falk, K; Namork, E; Rimstad, E; Mjaaland, S; Dannevig, B H (1997). Characterization of infectious salmon
anaemia virus, an orthomyxo-like virus isolated from Atlantic salmon (Salmo salar L). Journal of Virology 71:
9016–9023.
Fall, M; Kpatcha, K; Diebakate, C; Faye, N; Toguebaye, B (1997). Observations sur des myxosporidies
(Myxozoa) du genre Myxobolus parasites de Mugil cephalus (Poisson, Téléostéen) du Sénégal. Parasite. 4(2): 173–-
180 (In French, English abstract) [Online] Available from: https://doi.org/10.1051/parasite/1997042173 [Accessed
20 September 2019].
FAO, (2018). The state of the world’s fisheries and aquaculture, 2018. Food and Agriculture Organization of the
United Nations (FAO), Rome, Italy. [Online] Available from: http://www.fao.org/documents/card/en/c/I9540EN
[Accessed 17 October 2019].
FAO, (2019). Fishing gear types: hooks and lines, Food and Agriculture Organization of the United Nations (FAO),
Rome, Italy [Online] Available from: http://www.fao.org/fishery/geartype/109/en [Accessed 17 October 2019].
FAO, (2020a). New Zealand. Fishery and aquaculture country profiles. Fisheries and Aquaculture Department,
Food and Agriculture Organisation of the United Nations. Rome, Italy [Online] Available from:
http://www.fao.org/fishery/facp/NZL/en [Accessed 31 March 2020].
FAO, (2020b). Mullet, Mugil cephalus. Cultured Aquatic Species Fisheries and Aquaculture Department, Food and
Agriculture Organisation of the United Nations. Rome, Italy [Online] Available from:
http://www.fao.org/fishery/culturedspecies/Mugil_cephalus/en [Accessed 28 July 2020].
Farley, C A (1978). Viruses and virus like lesions in marine molluscs. Marine Fisheries Review 40: 18–20.
Farmer, J J; Davis, B R; Hickman-Brenner, F W; McWhorter, A; Huntley-Carter, G P; Asbury, M A;
Riddle, C; Wathen-Grady, H G; Elias, C; Fanning, G R; Steigerwalt, A G; O'Hara, C M; Morris, G K;
Smith, P B; Brenner, D J (1985). Biochemical identification of new species and biogroups of Enterobacteriaceae
isolated from clinical specimens. Journal of Clinical Microbiology 21(1): 46–76.
Farmer, J J; McWhorter, A C; Brenner, D J; Morris, D K (1984). The Salmonella-Arizona group of
Enterobacteriaceae: nomenclature, classification and reporting. Clinical Microbiology Newsletter 6: 63–66.
Farto, R; Armada, S P; Montes, M; Guisande, J A; Perez, M J; Nieto, T P (2003). Vibrio lentus associated with
diseased wild octopus (Octopus vulgaris). Journal of Invertebrate Pathology 83: 149–156 [Online} Available from:
https://doi.org/10.1016/S0022-2011(03)00067-3 [Accessed 9 May 2019].
Farto, R; Fichi, G; Gestal, C; Pascual, S; Nieto, T P (2014). Bacteria affecting cephalopods, pp 127–142 In:
Gestal, C; Pasual, S; Guerra, A; Fiorito, G; Veites, J M (Eds.) Handbook of pathogens and diseases in cephalopods.
Springer Science, New York, United States of America, 230 p.
Fauquet, C M; Mayo, M A; Maniloff, J; Desselberger, U; Bal, L A (2005). Virus taxonomy: Eighth Report of
the International Committee on the Taxonomy of Viruses. Academic Press, London, United Kingdom, 1162 p.
Faye, N; Kpatcha, T K; Fall, M; Toguebaye, B S (1997). Heart infections due to myxosporean (Myxozoa)
parasites in marine and estuarine fishes from Senegal. Bulletin of the European Association of Fish Pathologists
17(3-4): 115–117.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 125
Fayer, R; Nerad, T (1996). Effects of low temperatures on viability of Cryptosporidium parvum oocysts. Applied
and Environmental Microbiology 62(4): 1431–1433.
Feist, S W; Longshaw, M (2006). Phylum Myxozoa, pp 230–296 In: Woo PTK (Ed.).Fish diseases and disorders,
Volume 1. CAB International, Wallingford, United Kingdom, 791 p.
Ferguson, H W; Collins, R O; Moore, M; Coles, M; MacPhee, D D (2004). Pseudomonas anguilliseptica
infection in farmed cod, Gadus morhua L. Journal of Fish Diseases 27: 249–253.
Fiala, I; Bartosova-Sojkova, P; Whipps, C M (2015). Classification and phylogenetics of Myxozoa, pp 85–110
In: Okamura, B; Gruhi, A; Bartholomew J (Eds.). Myxozoan evolution and development. Springer Science, New
York, United States of America [Online] Available from: http://www.doi.10.1007/978-3-319-14753-6_5 [Accessed
21 January 2020].
Fichi, G; Cardeti, G; Perrucci, S; Vanni, A; Cersini, A; Lenzi, C; De Wolf, T; Fronte, B; Garducci, M;
Susini, F (2015). Skin lesion-associated pathogens from Octopus vulgaris: first detection of Photobacterium
swingsii, Lactococcus garvieae and betanodavirus. Diseases of Aquatic Organisms 115(2): 147–156 [Online]
Available from: http://www.doi.10.3354/dao02877 [Accessed 1 November 2019].
Fiorito, G; Affuso, A; Basil, A C; Girolamo, P; D’Angelo, L; Dickel, L; Gestal, C; Grasso, F; Kuba, F M;
Melillo, D; Osorio, D; Perkins, K; Ponte, G; Shashar, N; Smith, D; Smith, J; Andrews, P L R (2015).
Guidelines for the care and welfare of cephalopods in research –A consensus based on an initiative by CephRes,
FELASA and the Boyd Group. Laboratory Animals 49(52): 1–90 [Online] Available from:
http://www.doi.10.1177/0023677215580006 la.sagepub.com [Accessed 14 May 2019].
Fish and Game, (2014). New Zealand Fish and Game Council [Online] Available from:
http://www.fishandgame.org.nz/ [Accessed 24 September 2014].
FishBase, (2019). A global information system on fisheries, Quantitative Aquatics, Inc, [Online] Available from
http://www.fishbase.de/ [Accessed 7 October 2019].
Fisheries New Zealand, (2019a). Aquaculture Decision Report — Western Firth Marine Farming Consortium,
Coastal Permit CST60082314, Firth of Thames. Fisheries New Zealand, Ministry for Primary Industries,
Wellington, New Zealand [Online] Available from: https://www.fisheries.govt.nz/dmsdocument/33993/direct
[Accessed 28 July 2020].
Fisheries New Zealand, (2019b). National Panel Survey of Marine Recreational Fishers 2017–18. New Zealand
Fishery Assessment Report 2019/24, 108 p. Ministry for Primary Industries, Wellington, New Zealand [Online]
Available from: https://www.fisheries.govt.nz/dmsdocument/36792-far-201924-national-panel-survey-of-marine-
recreational-fishers-201718 [Accessed 29 July 2020].
Fisheries New Zealand, (2020). National Plan of Action – Seabirds 2020. Supporting document. Ministry for
Primary Industries, Wellington, New Zealand [Online] Available from: s
https://www.mpi.govt.nz/dmsdocument/40658-national-plan-of-action-seabirds-2020-supporting-document
[Accessed 29 July 2020].
Foot, J; Free, D; McDowell, T; Arkusk, K; Hedrick, R P (2006). Infectious hematopoietic necrosis virus
transmission and disease among juvenile Chinook salmon exposed in culture compared to environmentally relevant
conditions. San Francisco Estuary and Watershed Science 4(1): 1–16 [Online] Available from:
https://doi.org/10.15447/sfews.2006v4iss1art2 [Accessed 9 January 2020].
Ford, L A; Alexander, S K; Cooper, K M; Hanlon, R T (1986). Bacterial populations of normal and ulcerated
mantle tissue of the squid, Lolliguncula brevis. Journal of Invertebrate Pathology 48(1): 13–26.
Forrest, B; Hopkins, G; Webb, S; Tremblay, L (2011). Overview of marine biosecurity risks from finfish
aquaculture development in the Waikato Region. Waikato Regional Council Technical Report 2011/22, prepared by
the Cawthron Institute, Nelson, New Zealand 94 p. [Online] Available from:
http://www.waikatoregion.govt.nz/assets/WRC/WRC-2019/TR201122.pdf [Accessed 27 August 2019].
Forsythe, J W; Hanlon, R T; Bullis, R A; Noga, E J (1991). Octopus bimaculoides (Pickford & McConnaughey,
1949) - a marine invertebrate host for ectoparasitic protozoans. Journal of Fish Diseases 14(4): 431–442.
Fraser, D I; Munro, P D; Smail, D A (2006). Disinfection guide version IV: practical steps to prevent the
introduction and minimise the transmission of diseases of fish. Fisheries Research Services Internal Report 13/06.
Marine Laboratory, Aberdeen, Scotland, United Kingdom, 14 p.
Frerichs, G N; Tweedie, A; Starley, W G; Richards, R H (2000). Temperature, pH, and electrolyte sensitivity,
and heat, UV and disinfectant inactivation of sea bass (Dicentrarchus labrax) neuropathy
nodavirus. Aquaculture 185: 13–24.

126 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
FSANZ (2020). Food irradiation. Food standards Ausatralia New Zealand [Online] Available from:
https://www.foodstandards.gov.au/consumer/foodtech/irradiation/Pages/default.aspx [Accessed 21 October 2020].
Fujita, T (1930). On a new myxosporidia in the muscle of the grey-mullet Chloromyxum bora nov. sp. Rybnoe
Khozyaystvo 42: 45–48 (In Russian).
Furuya, H; Souidenne, D (2019). Dicyemids, pp 159–68 In: Gestal C, Pascual S, Guerra A, Fiorito G, Vieites J
(Eds.). Handbook of pathogens and diseases in cephalopods, Springer, New York, United States of America, 230 p.
Gaevskaya, A V; Kovaleva, A A (1979). Two new species of Myxosporidia Davisia donecae n. sp. and
Ceratomyxa australis n. sp. from the horse mackerel in the south-eastern Atlantic. Russian Journal of Marine
Biology 3: 80–83.
Gagné, N; Leblanc, F (2018). Overview of infectious salmon anaemia virus (ISAV) in Atlantic Canada and first
report of an ISAV North American-HPR0 subtype. Journal of Fish Diseases 41: 421–430 [Online] Available from:
http://www.doi.10.1111/jfd.12670 [Accessed 30 October 2019].
Gao, X Y; Zhi, X Y; Li, H W; Klenk, H P; Li, W J (2014). Comparative genomics of the bacterial genus
Streptococcus illuminates evolutionary implications of species groups. PLoS ONE 9(6): e101229 [Online] Available
from: http://www.doi.10.1371/journal.pone.0101229 [Accessed 14 January 2020].
Garseth, Å H; Fritsvold, C; Opheim, M; Skjerve, E; Biering, E (2012). Piscine reovirus (PRV) in wild Atlantic
salmon, Salmo salar L and seatrout, Salmo trutta L. in Norway. Journal of Fish Diseases 36: 483–493.
Garver, K A; Hawley, L M; McClure, C A; Schroeder, T; Aldous, S; Doig, F; Snow, M; Edes, S; Baynes, C;
Richard, J (2011). Development and validation of a reverse transcription quantitative PCR for universal detection
of viral hemorrhagic septicaemia virus. Diseases of Aquatic Organisms 95: 97–112.
Garver, K A; Johnson, S C; Polinski, M P; Bradshaw, J C; Marty, G D; Snyman, H N; Morrison, D B;
Richard, J (2016). Piscine orthoreovirus from Western North America Is transmissible to Atlantic Salmon and
Sockeye Salmon but fails to cause heart and skeletal muscle inflammation. PLoS One 5:11(1):e0146229 [Online\
Available from: http://www.doi. 10.1371/journal.pone.0146229 [Accessed 21 January 2020].
Garver, K; Wade, J (2017). Characterization of infectious hematopoietic necrosis virus (IHNV). Research
Document 2017/073, Canadian Science Advisory Secretariat (CSAS), Department of Fisheries and Oceans (DFO),
Ottawa, Canada, 32 p.
Gaughan, D J (2002). Disease-translocation across geographic boundaries must be recognized as a risk even in the
absence of disease identification: the case with Australian Sardinops. Reviews in Fish Biology and Fisheries 11(2):
113-123.
Geoghegan, J L; Di Giallonardo, F; Cousins, K; Shi, M; Williamson, J E; Holmes, E C (2018). Hidden
diversity and evolution of viruses in market fish. Virus Evolution 4(2): vey031 [Online] Available from:
http://www.doi.10.1093/ve/vey031 [Accessed 31 May 2019].
Georgiades, E; Fraser, R; Jones, J B (2016). Options to Strengthen On-farm Biosecurity Management for
Commercial and Non-commercial Aquaculture. Ministry for Primary Industries Technical Paper No: 2016/47,
prepared for the Aquaculture Unit, Ministry for Primary Industries, Nelson, New Zealand, 366 p [Online] Available
from: https://www. www.mpi.govt.nz/dmsdocument/13287/send [Accessed 11 June 2019].
Gestal, C; Abolla, E; Pascual, S (1988). Rickettsiales-like organisms in the gills of reared Octopus vulgaris
(Mollusca, cephalopoda). Bulletin of the European Association of Fish Pathologists 18: 13–14.
Gestal, C; Abollo, E; Pascual, S (2002a). Observations on associated histopathology with Aggregata octopiana
infection (Protista: Apicomplexa) in Octopus vulgaris. Diseases of Aquatic Organisms 50: 45–49.
Gestal, C; Guerra, A; Pascual, S; Azevedo, C (2002b). On the life cycle of Aggregata eberthi and observations on
Aggregata octopiana (Apicomplexa, Aggregatidae) from Galicia (NE Atlantic). European Journal of Protistology
37: 427–435.
Gestal, C; Guerra, A; Pascual, S (2007). Aggregata octopiana (Protista: Apicomplexa): a dangerous pathogen
during commercial Octopus vulgaris on growing. ICES Journal of Marine Science 64: 1743–1748.
Gestal, C; Nigmatullin, Ch M; Hochberg, F G; Guerra, A; Pascual, S (2005). Aggregata andresi n. sp.
(Apicomplexa: Aggregatidae) from the ommastrephid squid Martialia hyadesi in the SW Atlantic Ocean and some
general remarks on Aggregata spp. in cephalopod hosts. Systematic Parasitology 60: 65–73.
Gestal, C; Pascual, S; Hochberg, F H (2010). Aggregata bathytherma sp. nov. (Apicomplexa: Aggregatidae) a
new coccidian parasite associated with a deep-sea hydrothermal ventoctopus. Diseases of Aquatic Organisms 91:
237–242.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 127
Gestal, C; Pasual, S; Guerra, A; Fiorito, G; Veites, J M (2019). Handbook of pathogens and diseases in
cephalopods. Springer Science, New York, United States of America, 230 p.
Gibson-Kueh, S; Ngoh-Lim, G H; Netto, P; Kurita, J; Nakajima, K; Ng, M L (2004). A systemic iridoviral
disease in mullet, Mugil cephalus L. and tiger grouper, Epinephelus fuscoguttatus Forsskal: a first report and study.
Journal of Fish Diseases 27: 693–699.
Giulietti, L; Mattiucci, S; Paoletti, M; Grevskott, D H; Bao, M; Cipriani, P; Levsen, A (2019). Morphological
and molecular identification of a new Kudoa thyrsites isolate in Mediterranean silver scabbardfish Lepidopus
caudatus. Diseases of Aquatic Organisms 132: 125–134 [Online] Available from:
https://www.doi.org/10.3354/dao033161 [Accessed 1 November 2019].
Gleadill, I G (2016). Octopus sinensis d’Orbigny, 1841 (Cephalopoda: Octopodidae): valid species name for the
commercially valuable East Asian common octopus. Species Diver 21: 31–42.
Glen, J A; Emmenegger, E J; Grady, C A; Roon, S R; Gregg, J L; Conway, C M; Winton, J R; Hershberger,
P K (2012). Kinetics of viral load and erythrocytic inclusion body formation in Pacific herring artificially infected
with erythrocytic necrosis virus. Journal of Aquatic Animal Health 24(3): 195 –200 [Online] Available from:
https://doi.org/10.1080/08997659.2012.676592 [Accessed 21 August 2019].
Go, J (2015). Megalocytiviruses and Megalocytivirus-like infections of ornamental fish, pp 15–18 In:
Megalocytiviruses of freshwater ornamental fish and pathogenicity in marine and euryhaline species. PhD. thesis,
Farm Animal and Veterinary Public Health, Faculty of Veterinary Science, University of Sydney, Sydney, Australia
[Online] Available from: https://www.core.ac.uk/download/pdf/41241461.pdf [Accessed 21 January 2020].
Go, J; Whittington, R (2006). Experimental transmission and virulence of a megalocytivirus (Family Iridoviridae)
of dwarf gourami (Colisa lalia) from Asia in Murray cod (Maccullochella peelii peelii) in Australia. Aquaculture
258, 140-149.
Gomez, D K; Baeck, D W; Kim, J H; Choresca, C H; Park, S C (2008). Molecular detection of betanodavirus in
wild marine fish populations in Korea. Journal of Veterinary Diagnostic Investigation 20: 38–44.
Gomez, D K; Bartholomew, J; Sunyer, J O (2014). Biology and mucosal immunity to Myxozoans.
Developmental and Comparative Immunology 43(2): 243–256 [Online] Available from:
http://www.doi.10.1016/j.dci.2013.08.014 [Accessed 18 September 2019].
Gomez, K W; Lim, D J; Baek, G W; Youn, H J; Shin, N S; Youn, H Y; Hwang, C Y; Park, J H; Park, S C
(2006). Detection of betanodaviruses in apparently healthy aquarium fishes and invertebrates. Journal of Veterinary
Science 74: 369–374.
Gomez, D K; Baek, G W; Kim, J H; Choresca, C H; Park, S C (2008). Molecular detection of betanodavirus in
wild marine fish populations in Korea. Journal of Veterinary Diagnostic Investigation 20(1): 38–44 [Online]
Available from: http://www.doi.10.1177/104063870802000107 [Accessed 8 April 2020].
Gomez, D K; Mori, K; Okinaka, Y; Nakai, T; Park, S C (2010). Trash fish can be a source of betanodaviruses
for cultured marine fish. Aquaculture 302: 158–163 [Online] Available from: http://
www.doi.10.1016/j.aquaculture.2010.02.033 [Accessed 7 October 2019].
Gomez-Casado, E; Estepa, A; Coll, J M (2011). A comparative review on European-farmed finfish RNA viruses
and their vaccines. Vaccine 29(15): 2657–2671.
Goodwin, A E; Peterson, J E; Myers, T R; Money, D J (2004). Transmission of exotic fish viruses. Fisheries 29:
19−23.
Granados-Amores, J; García-Rodríguez, F J; Hochberg, F G; Salinas-Zavala, C A (2014). The taxonomy and
morphometry of squids in the family Loliginidae (Cephalopoda: Myopsida) from the Pacific Coast of Mexico.
American Malacological Bulletin 32(2): 198–208 [Online] Available from: https://doi.org/10.4003/006.032.0209
[Accessed 17 December 2019].
Gregg, J L; Grady, C A; Friedman, C S; Hershberger, P K (2012). Inability to demonstrate fish-to-fish
transmission of Ichthyophonus from laboratory infected Pacific herring Clupea pallasii to naïve conspecifics.
Diseases of Aquatic Organisms 99: 139–142 [Online] Available from: http://www.doi.10.3354/dao02458 [Accessed
11 September 2019].
Groner, M L; Mynard, J; Bretya, R; Carnegie, R B; Dobson, A; Friedman, C S; Froelich, B; Garren, M;
Gulland, F M D; Heron, S F; Noble, R T; Revie, C W; Shields, J D; Vanderstichel, R; Weil, E; Wyllie-
Echeverria, S W; Harvell, C D (2016). Managing marine disease emergencies in an era of rapid change
Philosophical Transactions of the Royal Society, Series B: 20150364 [Online] Available from:
http://dx.doi.org/10.1098/rstb.2015.0364 [Accessed 27 January 2020].

128 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Gross, L; Richard, J; Hershberger, P; Garver, K (2019). Low susceptibility of sockeye salmon Oncorhynchus
nerka to viral hemorrhagic septicaemia virus genotype Iva. Diseases of Aquatic Organisms 135: 201–209 [Online]
Available from: https://doi.org/10.3354/dao03398 [Accessed 10 December 2019].
Guk, S M; Shin, E H; Kim, J L; Sohn, W M; Hong, K S; Yoon, C H; Lee, S H; Chai, J L (2007). A survey of
Heterophyes nocens and Pygidiopsis summa metacercariae in mullets and gobies along the coastal areas of the
Republic of Korea. The Korean Journal of Parasitology 45(3): 205–211.
Haenen, O (2017). Major bacterial diseases affecting aquaculture. Presentation to the Aquatic AMR Workshop 1:
10-11 April 2017, Mangalore, India, Responsible management of bacterial diseases in Aquaculture
(FMM/RAS/298: Strengthening capacities, policies and national action plans on prudent and responsible use of
antimicrobials in fisheries. Food and Agriculture Organisation of the United Nations [Online] Available from:
http://www.fao.org/fi/static-media/MeetingDocuments/WorkshopAMR/presentations/07Haenen.pdf [Accessed 17
December 2019].
Hahn, C W (1917). On the sporozoan parasites of the fishes of Woods Hole and vicinity. III. On the Chloromyxum
clupeidae of Clupea harengus (Young), Pomolobus pseudoharengus (Young) and P. aestivalis (Young). Journal of
Parasitology 4: 13–20.
Halaly, M A; Subramaniam, K; Koda, S A; Popov, V L; Stone, D; Way, K; Waltzek, T B (2019).
Characterization of a novel Megalocytivirus isolated from European chub (Squalius cephalus). Viruses 11(5): pii: E
440 [Online] Available from: http://www.doi.10.3390/v11050440 [Accessed 24 October 2019].
Hallett, S L; Hartigan, A; Atkinson, S D (2015). Myxozoans on the move: Dispersal modes, exotic species and
emerging diseases, pp 343–362 In: Okamura B, Gruhl A, Bartholomew J (Eds.) Myxozoan Evolution, Ecology and
Development. Springer International, Geneva, Switzerland, 441 p.
Hamed, M G E; Elias, A N (1970). Effect of food processing methods upon survival of the trematode Heterophyes
sp. in flesh of mullet caught in brackish Egyptian waters. Journal of Food Science 35: 386.
Hanlon, R T; Forsythe, J W; Cooper, K M; Dinuzzo, A R; Folse, D S; Kelly, M T (1984). Fatal penetrating skin
ulcers in laboratory-reared octopuses. Journal of Invertebrate Pathology 44(1): 67–83.
Hanlon, R T; Forsythe, W (1990). Diseases of mollusca: Cephalopoda: Diseases caused by microorganisms, pp
23−46 In: Kinne O (Ed.) Diseases of marine animals, Vol III. Biologische Anstalt Helgoland, Hamburg, Germany.
Hanson, L; Dishon, A; Kotler, M (2011). Herpesviruses that Infect Fish. Viruses 3: 2160–2191.
Harai, Y; Ashata-Tago, S; Ainoda, Y; Fujita, T; Kikuchi, K (2015). Edwardsiella tarda bacteraemia. A rare but
fatal water- and foodborne infection: Review of the literature and clinical cases from a single centre. Canadian
Journal of Infectious Diseases and Medical Microbiology 26(6): 313–318 [Online] Available from
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4692300 [Accessed 26 June 2018].
Harms, C A; Lewbart, G A; McAlarney, R; Christian, L S; Geissler, K A; Lemons, C (2006). Surgical excision
of mycotic (Cladosporium sp.) granulomas from the mantle of a cuttlefish (Sepia officinalis). Journal of Zoo and
Wildlife Medicine 37(4): 524–530.
Hart, L M; Traxler, G S; Garver, K A; Richard, J; Gregg, J L; Grady, C A; Kurath, G; Hershberger, P K
(2011). Larval and juvenile Pacific herring Clupea pallasii are not susceptible to infectious hematopoietic necrosis
under laboratory conditions. Diseases of Aquatic Organisms 93:105–110 [Online] Available from:
https://doi.org/10.3354/dao02294 [Accessed 27 May 2019].
Hauck, A K (1977). Occurrence and survival of the larval nematode Anisakis sp. in the flesh of fresh, frozen,
brined, and smoked Pacific herring, Clupea harengus pallasi. Journal of Parasitology 63: 515–519.
Hauge, H; Dahle, M; Modal, T; Thoen, E; Gjevre, A G; Weli, S; Alarcon, M; Grove, S (2016). Piscine
orthoreovirus can infect and shed through the intestine in experimentally challenged Atlantic salmon (Salmo salar
L.) Veterinary Research 47: 57, 12 p. [Online] Available from: http://www.doi.1186/s13567-016-0343-z [Accessed
24 August 2020].
Harvell, C D; Kim, K; Burkholder, J M; Colwell, R R; Epstein, P R; Grimes, D J; Hofmann, E E; Lipp, E K;
Osterhaus, A D M E; Overstreet, R M; Porter, J W; Smith, G W; Vasta, G R (1999). Emerging marine diseases
– Climate links and anthropomogenic factors. Science 285: 1505–1510 [Online] Available from:
http://www.doi.10.1126/science.285.5433.1505 [Accessed 17 December 2019].
Hawley, L M; Garver, K A (2008). Stability of viral hemorrhagic septicaemia virus (VHSV) in freshwater and
seawater at various temperatures. Diseases of Aquatic Organisms 82: 171–178 [Online] Available from:
http://www.doi.10.3354/dao01998 [Accessed 1 November 2019].

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 129
Hedrick, R P; Batts, W N; Yun, S; Traxler, G S; Kaufman, J; Winton, J R (2003). Host and geographic range
extensions of the North American strain of viral haemorrhagic septicaemia virus. Diseases of Aquatic Organisms 55:
211–220.
Hedrick, R P; McDowell, T S; Mukkatira, K; MacConnell, E; Petrie, B (2008). Effects of freezing, drying,
ultraviolet radiation, chlorine and quaternary ammonium compounds on the infectivity of myxospores of Myxobolus
cerebralis for Tubifex tubifex. Journal of Aquatic Animal Health 20: 116– 125.
Henning, S S; Hoffman, L C; Manley, M (2013). A review of Kudoa-induced myoliquefaction of marine fish
species in South Africa and other countries. South African Journal of Science 109(11-12): 1–5 [Online] Available
from: http://www.dx.doi.org/10.1590/sajs.2013/20100320 [Accessed 16 September 2019].
Henning, S S; Krugel, K; Manley, M (2019). Prevalence of Kudoa thyrsites (Myxozoa) in South African sardine
Sardinops sagax, kingklip Genypterus capensis, and Cape hakes Merluccius capensis and M. paradoxus. South
African Journal of Marine Sciences 41(3): 261–268 [Online] Available from: [Accessed 6 December 2019].
Hershberger, P K; Elder, N E; Grady, C A; Gregg, J L; Pacheco, C A; Greene, C; Rice, C; Meyers, T R
(2009). Prevalence of viral erythrocytic necrosis in Pacific herring and epizootics in Skagit Bay, Puget Sound,
Washington. Journal of Aquatic Animal Health 21: 1–7.
Hershberger, P K; Gregg, J L; Grady, C A; Collins, RM; Winton, J R (2011). Susceptibility of three stocks of
Pacific Herring to viral hemorrhagic septicaemia. Journal of Aquatic Animal Health 22(1): 1–7.
Hershberger, P K; Gregg, J; Pacheco, C; Winton. J; Richard, J; Traxler, G (2007). Larval Pacific herring,
Clupea pallasii (Valenciennes), are highly susceptible to viral haemorrhagic septicaemia and survivors are partially
protected after their metamorphosis to juveniles. Journal of Fish Diseases 30: 445–458.
Hershberger, P K; Hart, A; Gregg, J; Elder, N; Winton, J (2006). Dynamics of viral hemorrhagic septicaemia,
viral erythrocytic necrosis and ichthyophoniasis in confined juvenile Pacific herring Clupea pallasii. Diseases of
Aquatic Organisms 70: 201–208.
Hershberger, P K; Hart, L M; MacKenzie, A H; Yanney, M L; Conway, C M; Elliott, D G (2015). Infecting
Pacific Herring with Ichthyophonus sp. in the laboratory. Journal of Aquatic Animal Health 27(4): 217–221 [Online]
Available from: https://doi.org/10.1080/08997659.2015.1095809 [Accessed 11 September 2019].
Hershberger, P K; Kocan, R M; Elder, N E; Meyers, T R; Winton, J R (1999). Epizootiology of viral
hemorrhagic septicaemia virus in Pacific herring from the spawn-on-kelp fishery in Prince William Sound, Alaska,
USA. Diseases of Aquatic Organisms 37: 23–31.
Hershberger, P; Rhodes, L; Kurath, G; Winton, J (2013). Infectious diseases of fishes in the Salish Sea.
Fisheries 38(9): 402.
Herve-Claude, L P; Carpenter, T E; Hedrick, R E (2008). Risk of introducing viral hemorrhagic septicaemia
virus (VHSV) to the Chilean South Pacific via sardine imports from Europe. Diseases of Aquatic Organisms 78:
199–207 [Online] Available from: https://www.doi.org/10.3354/dao01862 [Accessed 5 August 2019].
Hewitt. G C; Little, R W (1972). Whirling disease in New Zealand trout caused by Myxosoma cerebralis (Hofer,
1903) (Protozoa: Myxosporidia). New Zealand Journal of Marine and Freshwater Research 6: 1–10.
Hick, P; Becker, J; Whittington, R (2016). Iridoviruses of fish, pp 127–152 In: Kibenge FSB, Godoy MG (Eds.).
Aquaculture Virology, Academic Press, London, United Kingdom, 549 p.
Hickman, W R; Tait, J M (2008). Experimental rearing of juvenile New Zealand turbot Colistium nudipinnis
(Waite 1910): A potential new aquaculture species. Aquaculture Research 32(9): 727–737 [Online] Available from:
http://www.doi.10.1046/j.1365-2109.2001.00610.x [Accessed 27 August 2019].
High, W L (1980). Bait loss from halibut longline gear observed from a submersible. Marine Fisheries Review
42(2): 26–29.
Hill, B J (1982). Infectious pancreatic necrosis virus and its virulence. pp 91-114 In: Roberts, R J (Ed.) Microbial
diseases of fish. Academic Press, London.
Hine, P M (1977). Myxobolus iucundus n. sp. (Sporozoa: Myxosporidia) parasitic in Galaxias maculates (Jenyns,
1842) in New Zealand. Journal of the Royal Society of New Zealand 7: 157–161.
Hine, P M; MacDiarmid, S C (1997). Contamination of fish products: risks and prevention. OIE Reviews in
Science and Technology 16(1): 135–145 [Online] Available from:
https://pdfs.semanticscholar.org/4556/d1275f4260251a15459c0f50ef33215f28c5.pdf [Accessed 17 June 2020].
Hine, M P; Diggles, B K (2005). Import Risk assessment: Ornamental Fish, Ministry for Primary Industries,
Wellington, New Zealand, 262 p.

130 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Hine, M P; Jones, J B (1994). Bonamia and other aquatic parasites of importance to New Zealand, New Zealand
Journal of Zoology, 21(1): 49–56 [Online] Available from: http://www.doi.10.1080/03014223.1994.951797
[Accessed 16 September 2019].
Hine, M P; Jones, J B; Diggles, B K (2000). A checklist of the parasites of New Zealand fishes, including
previously unpublished records. NIWA Technical Report 75. National Institute for Water and Atmospheric Research
Ltd. Auckland, New Zealand 95 p.
Hine, M P; MacDiarmid, S C (1997). Contamination of fish products: risks and prevention. Scientific and
Technical Review (OIE) 16(1): 135–145.
Hirai, Y; Asahata-Tago, S; Ainoda, Y; Fujita, T; Kikuchi, K (2015). Edwardsiella tarda bacteraemia. A rare but
fatal water and foodborne infection: review of the literature and clinical cases from a single centre. Canadian
Journal of Infectious Diseases and Medical Microbiology 26(6): 313–318 [Online] Available from:
http://www.dx.doi.org/10.1155/2015/702615 [Accessed 14 January 2010].
Hochberg, F G (1983). The parasites of cephalopods: A review. Memoirs of the National Museum of Victoria 44:
109–145.
Hochberg, F G; Couch, J A (1971). Biology of cephalopods, pp. 221-228 In: Miller JW, Vanerwalker J, Walier
RA (Eds.). Scientists-in-the-sea, United States Department of the Interior, Washington D.C, United States of
America, 674 p.
Hoffman, G L (1990). Myxobolus cerebralis, a worldwide cause of salmonid whirling disease. Journal of Aquatic
Animal Health 2: 30–37.
Hoffman, G l; Putz, R E (2011). Effect of Freezing and Aging on the Spores of Myxosoma cerebralis, the
Causative Agent of Salmonid Whirling Disease. The Progressive Fish Culturalist 33(2): 95–98 [Online] Available
from: http://www.doi.10.1577/1548-8640(1971)33[95:EOFAAO]2.0.CO;2 [Accessed 28 July 2020].
Holdsworth, J; Rea, T; Southwick, R (2016). Recreational fishing in New Zealand – a billion-dollar industry.
Unpublished report produced for the New Zealand Marine Research Foundation, March 2016 [Online] Available
from: http://www. Nzmrf.org.nz/files/New-Zealand-Fishing-economic-Report.pdf [Accessed 31 August 2020].
Holzer, A S; Blasco-Costa, I; Sarabeev, V L; Ovcharenko, M O; Balbuelena, J A (2006). Kudoa trifolia sp. n. –
molecular phylogeny suggests a new spore morphology and unusual tissue location for a well-known genus. Journal
of Fish Diseases 29: 743–755 [Online] Available from: http://www/doi.10.1111/j.1365-2761.2006.00770.x
[Accessed 12 September 2019].
Hosono, N; Suzuki, S; Kusuda, R (1996). Genogrouping of birnaviruses isolated from marine fish: a comparison
of VP2/NS junction regions on genome segment A. Journal of Fish Diseases 19: 295–302.
Hume, T M; Bell, R G; de Lange, W P; Healy, T R; Hicks, D M; Kirk, R M (1992). Murray, D (1991). Coastal
oceanography and sedimentology in New Zealand, 1967–91. New Zealand Journal of Marine and Freshwater
Research 26(1): 1–36 [Online] Available from: https://doi.org/10.1080/00288330.1992.9516497 [Accessed 29 July
2020].
Hutson, K S; Ernst, I; Whittington, I D (2007). Risk assessment for metazoan parasites of yellowtail kingfish
Seriola lalandi (Perciformes: Carangidae) in South Australian sea-cage aquaculture. Aquaculture 271(1-4): 85–99
[Online] Available from: https://doi.org/10.1016/j.aquaculture.2007.03.020 [Accessed 7 May 2020].
IAEA, (2000). Irradiation of fish, shellfish and frogs legs. A compilation of technical data for authorisation and
control. International Consultative Group on Food Irradiation. IAEA Technical Document 1158, International
Atomic Energy Agency, Vienna, Austria 96 p.
Ibragimov, S R (1987). Forming of the parasite fauna of mullets in the Caspian Sea. Baku, Records of the Institute
of Zoology of AS Azerbaijan SSR (Deposited by VINITI 03.04.87, № 2407-B87): 1–14 (In Russian).
IDAAD, (2019). International database on aquatic animal diseases, Centre for Environment, Fisheries and
Aquacultural Science, Department for Environment, Food and Rural Affairs (DEFRA), Weymouth, United
Kingdom [Online] Available from: https://www.cefas.co.uk/international-database-on-aquatic-animal-diseases/
[Accessed 21 October 2019].
Inouye, K; Ikeya, F; Yamazaki, T; Hara, T (1990). Virucidal activities of various germicides to infectious
pancreatic necrosis virus (IPNV). Fish Pathology 25(2): 81–86 [Online] Available from
http://www.doi.10.3147/jsfp.25.81 [Accessed 6 August 2019].
Iskov, M P (1989). Myxosporidia (Myxosporea). In: Markevitch, A P; Schulman, S S (Eds.) Fauna Ukrainy 40(37)
(Invertebrate fauna of the Ukraine, in 40 volumes), Naukowa Dumka, Kiev, Ukraine, 212 p. (In Russian, with
English translation).

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 131
Isshiki, T; Nagano, T; Kanehira, K; Suzuki, S (2004). Distribution of marine birnavirus in cultured marine fish
species from Kagawa Prefecture, Japan. Journal of Fish Diseases 27(2): 89–98.
Isshiki, T; Nagano, T; Suzuki, S (2001). Infectivity of aquabirnavirus strains to various marine fish species.
Diseases of Aquatic Organisms 46(2): 109–114.
Ito, T; Mori, K; Arimoto, M; Nakajima, K (2004). Virulence of viral hemorrhagic septicaemia virus (VHSV)
isolates from Japanese flounder Paralichthys olivaceus in rainbow trout and several species of marine fish. Fish
Pathology 39(2): 103–104.
Ito, T; Yoshiura, Y; Kamaishi, T; Nakajima, K (2013). Dynamics of virus titre and quantity of red sea bream
iridovirus DNA using natural and autoclaved artificial sea water under different water temperatures. Bulletin of the
European Association of Fish Pathologists 33(4): 118–125.
Ito, T; Yoshiura, Y; Kamaishi, T; Yoshida, K; Nakajima, K (2013). Prevalence of red sea bream iridovirus
among organs of Japanese amberjack (Seriola quinqueradiata) exposed to cultured red sea bream iridovirus.
Journal of General Virology 94: 2094–2101 [Online] Available from: http://www.doi. 10.1099/vir.0.052902-0
[Accessed 7 April 2020].
Iurakhno, V M (1988). New data on Myxosporidia of Black Sea fishes. Parazitologiia 22(6): 521–524 (In
Russian).
Iwamoto, T; Nakai, T; Mori, K; Arimoto, M; Furusawa, I (2000). Cloning of the fish cell line SSN-1 for piscine
nodaviruses. Diseases of Aquatic Organisms 43: 81–89.
Iwashita, Y; Kamijo, Y; Nakahashi, S; Shindo, A; Yokoyama, K; Yamamoto, A; Yukinari, O; Ishikura, K;
Fujioka, M; Hatada, T; Takeda, T; Maruyama, K; Hiroshi, I (2013). Food poisoning associated with Kudoa
septempunctata. Journal of Emergency Medicine 44(5): 943–954.
Janda, J M; Sharon, L A (1993). Infections associated with the genus Edwardsiella: the role of Edwardsiella tarda
in human disease. Clinical and Infectious Diseases 17(4): 742–748 [Online] Available from:
http://www.doi.10.1093/clinids/17.4.742 [Accessed 1 November 2019].
Jansson, E; Vennerstrom, P (2014). Infectious diseases of coldwater fish in marine and brackish waters. Chapter 2
In: Woo P, Bruno D (Eds.), Diseases and disorders of finfish in cage culture, 2nd edition, CAB International,
Wallingford, United Kingdom, 342 p.
Jaysari, M; Hoffman, G L (1982). Review of Myxidium (Protozoa: Myxozoa: Myxosporea). Protozoological
Abstracts (England) 6(4): 61–91.
Jereb, P; Roper, C F E; Norman, M D; Finn, J K (2010). Cephalopods of the world. An annotated and illustrated
catalogue of species known to date. FAO Species Catalogue for Fishery Purposes 4(2). Food and Agriculture
Organisation of the United Nations, Rome, Italy [Online] Available from:
http://www.fao.org/docrep/014/i1920e/i1920e.pdf [Accessed 20 May 2019].
Jereb, P; Vecchione, M; Roper, C F E (2010). Family Loliginidae, pp 38–117 In: Jereb, P; Roper, C F E; Norman,
M D; Finn, J K (Eds.) Cephalopods of the world. An annotated and illustrated catalogue of species known to date.
Volume 2. Myopsid and Oegopsid Squids. FAO Species Catalogue for Fishery Purposes 4(2). Food and Agriculture
Organisation of the United Nations, Rome, Italy [Online] Available from:
http://www.fao.org/docrep/014/i1920e/i1920e.pdf [Accessed 20 May 2019].
Jeong, J B; Cho. H J; Jun, L J; Hong, S H; Chung, J K; Jeong, H D (2008). Transmission of iridovirus from
freshwater ornamental fish (pearl gourami) to marine fish (rock bream). Diseases of Aquatic Organisms 82: 27–36.
Johansen, L H; Jensen, I; Mikkelsen, H; Bjorn, P A; Jansen, P A; Bergh, O (2011). Disease interaction and
pathogens exchange between wild and farmed fish populations with special reference to Norway. Aquaculture 315:
167–186.
Johansen, R; Rove, S; Svendsen, N A K; Mohdal, I; Dannevoig, B (2004). A sequential study of pathological
findings in Atlantic halibut, Hippoglossus hippoglossus (L), throughout one year after an acute outbreak of viral
encephalopathy and retinopathy. Journal of Fish Diseases 27: 327–341.
Johnston, C (2008). Import risk analysis: Frozen skinless and boneless fillet meat of Pangasius spp. fish from
Vietnam for human consumption. Biosecurity New Zealand, Ministry of Agriculture and Forestry, Wellington, New
Zealand, 60 p.
Jones, G (1981). Thraustochytrid pathogens. Bulletin of the British Mycological Society 16 (Supplement 1): 5–6.
Jones, G; D’Or, R K (1983). Ultrastructural observations on a thraustochytrid fungus parasitic on the gills of squid
(Illex illecebrosus Lesuer). Journal of Parasitology 69: 903–911.

132 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Jones, J B; Hyatt, A D; Hine, P M; Whittington, R J; Griffin, D A; Bax, N Z (1997). Special topic review:
Australasian pilchard mortalities. World Journal of Microbiology & Biotechnology 13(4): 383–392.
Jones, S R (2003). Distribution, prevalence and severity of Parvicapsula minibicornis infections among
anadromous salmonids in the Fraser River, British Columbia, Canada. Diseases of Aquatic Organisms 54(1): 49–54
[Online] Available from: http://www.doi.10.3354/dao054049 [Accessed 17 December 2019].
Jones, S R M; Bartholomew, J L; Zhang, J Y (2015). Mitigating myxozoan disease impacts on wild fish
populations, pp 397–413 In: Okamura, B; Gruhl, A; Bartholomew, J (Eds.) Myxozoan Evolution, Ecology and
Development. Springer, Geneva, Switzerland, 441 p [Online] Available from:
https://link.springer.com/chapter/10.1007/978-3-319-14753-6_21 [Accessed 21 January 2020].
Jones, S R M; Long, A (2019). Host size influences prevalence and severity of Kudoa thyrsites (Cnidaria:
Myxosporea) infection in Atlantic salmon Salmo salar. Diseases of Aquatic Organisms 133: 99–105 [Online]
Available from: https://doi.org/10.3354/dao03335 [Accessed 8 May 2020].
Jordan, G W; Handley, W K (1969). Human infection with Edwardsiella tarda. Annals of Internal Medicine 70:
283–288.
Jorgensen, A; Nylund, A; Nikolaisen, V; Alexandersen, S; Karlsbakk, E (2011). Real-time PCR detection of
Parvicapsula pseudobranchicola (Myxozoa: Myxosporea) in wild salmonids in Norway. Journal of Fish Diseases
34: 365–371 [Online] Available from: http://www.doi.10.1111/j.1365-2761.2011.01248.x [Accessed 23 October
2013].
Joseph, A V; Sasidharan, R S; Nair, H P; Bhat, S G (2013). Occurrence of potential pathogenic Aeromonas
species in tropical seafood, aquafarms and mangroves off Cochin coast in South India. Veterinary World 6(6): 300–
306 [Online] Available from: http://www.doi.10.5455/vetworld.2013.300-306 [Accessed 28 May 2019].
Jun, L J; Jeong, J B; Kim, J H; Nam, J H; Shin, K W; Kim, J K; Kang, J C; Jeong, H D (2009). Influence of
temperature shifts on the onset and development of red sea bream iridoviral disease in rock bream Oplegnathus
fasciatus. Diseases of Aquatic Organisms 84: 201–208.
Kahn, S A; Beers, P T; Findlay, V L; Peebles, I R; Durham, P J; Wilson, D W; Gerrity, S E (1999). Import
risk analysis on non-viable salmonids and non-salmonid marine finfish. Australian Quarantine and Inspection
Service, Department of Agriculture, Fisheries and Forestry, Canberra, Australia, 409 p.
Kalavati, C; Nandi, N C (2007). Handbook on myxozoan parasites of Indian fishes, Department of Fisheries,
Kolkata, India 294 p. [Online] Available from: https://archive.org/stream/dli.zoological.hpg.035/index_djvu.txt, 294
p.
Karlsbakk, E (2001). Myxozoa, pp. 80-84 In: Costello, MJ. (Ed.) (2001). European register of marine species: a
check-list of the marine species in Europe and a bibliography of guides to their identification. Collection
Patrimoines Naturels 50: 80–84.
Karlsbakk, E; Koie, M (2011). Morphology and SSU rDNA sequences of Ortholinea orientalis (Shulman and
Shulman-Albova, 1953) (Myxozoa, Ortholineidae) from Clupea harengus and Sprattus sprattus (Clupeidae) from
Denmark. Parasitology Research 109(1): 139–145 [Online] Available from: http://www.doi.10.1007/s00436-010-
2237-8 [Accessed 23 October 2019].
Katharios, P; Kalatzis, P G; Kokkari, C; Pavlidis, M; Wang, Q (2019). Characterization of a highly virulent
Edwardsiella anguillarum strain isolated from Greek aquaculture, and a spontaneously induced prophage therein.
Frontiers in Microbiology 10(141), 6 p. [Online] Available from: https://doi.org/10.3389/fmicb.2019.00141
[Accessed 1 November 2019].
Kasai, A; Tsuduki, H; Jimenez, L A; Li, Y C; Tanaka, S; Sata, H (2017). Incidence of three Kudoa spp., K.
neothunni, K. hexapunctata, and K. thunni (Myxosporea: Multivalvulida), in Thunnus tunas distributed in the
Western Pacific Ocean. Parasitological Research 116: 1137–1150 [Online] Available from:
http://www.doi.10.1007/s00436-016-5369-7 [Accessed 14 May 2019].
Ke, F; He, L B; Pei, C; Zhang, Q Y (2011). Turbot reovirus (SMReV) genome encoding a FAST protein with a
non-AUG start site. BMC Genomics 12: 323, 13 p [Online] Available from: https://www.doi.org/10.1186/1471-
2164-12-323 [Accessed 12 August 2015].
Keeling, S E; Brosnahan, C L; Johnston, C; Wallis, R; Gudkovs, N; McDonald, W L (2013). Development and
validation of a real-time PCR assay for the detection of Aeromonas salmonicida. Journal of Fish Diseases 36: 495–
503.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 133
Kent, M L; Andree, K B; Bartholomew, J B; El-Matbouli, M; Desser, S S; Devlin, R H; Feist, S W; Hedrick,
R P; Hoffmann, R W; Khattra, J; Hallett, S L; Lester, J G; Longshaw, M; Palenzuela, O; Siddall, M E; Xiao,
C (2001). Recent advances in our knowledge of the Myxozoa. Journal of Eukaryotic Microbiology 48: 395–413.
Kent, M L; Traxler, G S; Kieser, D; Richard, J; Dawe, S C; Shaw, R W; Prosperi-Porta, G; Ketcheson, J;
Evelyn, T P T (1998). Survey of salmonid pathogens in ocean-caught fishes in British Columbia, Canada. Journal
of Aquatic Animal Health 10: 211–219.
Kibenge, F S B; Godoy, M (2016). Aquaculture virology. Academic Press, New York, United States of America,
568 p.
Kim, K; Kim, Y C; Kwon, W J; Jeong, H D (2017). Evaluation of blue mussel Mytilus edulis as vector for viral
hemorrhagic septicemia virus (VHSV). Diseases of Aquatic Organisms 126(3): 239 246 [Online] Available from:
http://www.doi.10.3354/dao03180 [Accessed 22 October 2020].
Kim, R; Faisal, M (2010). Comparative susceptibility of representative Great Lakes fish species to the North
American viral hemorrhagic septicaemia virus Sublineage IVb. Diseases of Aquatic Organisms 91: 23–24 [Online]
Available from: http://www.doi.10.3354/dao02217 [Accessed 21 October 2019].
Kim, Y C; Kwon, W J; Min, J G; Jeong, H D (2011). Isolation and initial characterization of new betanodaviruses
in shellfish. Transboundary and Emerging Diseases 65(6): 1557–1567 [Online] Available from:
https://doi.org/10.1111/tbed.12900 [Accessed 30 April 2020].
Kim, W S; Choi, S Y; Kim, D H; Oh, M J (2013a). A survey of fish viruses isolated from wild marine fishes from
the coastal waters of southern Korea. Journal of Veterinary Diagnostic Investigation 25(6): 750–755.
Kim, W S; Kim, J H; Oh, M J (2013b). Morphologic and genetic evidence for mixed infection with two
Myxobolus species (Myxozoa: Myxobolidae) in grey mullets, Mugil cephalus, from Korean Waters. The Korean
Journal of Parasitology 51(3): 369–373 [Online] Available from http://www.doi.10.3347/kjp.2013.51.3.369
[Accessed 7 January 2019].
King, A M Q; Lefkowitz, E; Adams, M; Carstens, E B (2011). Virus taxonomy: Ninth report of the international
Committee on the Taxonomy of Viruses. Elsevier Science, New York, United States, 1338 p.
Kitamura, S I; Kamata, S I; Nakano, S; Suzuki, S (2004). Solar UV radiation does not inactivate marine
birnavirus in coastal seawater. Diseases of Aquatic Organisms 58: 251–254.
Kitamura, S I; Suzuki, S (2000). Occurrence of marine birnavirus through the year in coastal seawater in the Uwa
Sea. Marine Biotechnology 2(2): 188–195.
Kitao, T; Aoki, T; Iwata, Κ (1979). Epidemiological study of streptococcosis of cultured yellowtail Seriola sp. 1.
streptococcosis of cultured yellowtail Seriola sp. 1. Distribution in seawater and muds around yellowtail farms.
Bulletin of the Japanese Society for Scientific Fisheries 45: 567–572.
Kluzik. N; Woodford, N (2016). Zoonoses associated with fish (including aquarium fish). Washington State
University, Washington, United States of America 2 p.
Kocan, R; Bradley, M; Elder, N; Meyers, T; Batts, W; Winton, J R (1997). North American strain of viral
hemorrhagic septicaemia virus is highly pathogenic for laboratory-reared Pacific herring. Journal of Aquatic Animal
Health 9: 279–290.
Kocan, R M; Hershberger, P K; Elder, N E (2001). Survival of the North American strain of viral hemorrhagic
septicaemia virus (VHSV) in filtered seawater and seawater containing ovarian fluid, crude oil and serum-enriched
culture medium. Diseases of Aquatic Organisms 44(1): 75–80 [Online] Available from: http://www.doi.
10.3354/dao044075 [Accessed 21 October 2019].
Koie, M; Karlsbakk, E; Einen, A C; Nylund, A (2013). A parvicapsulid (Myxozoa) infecting Sprattus sprattus
and Clupea harengus (Clupeidae) in the Northeast Atlantic uses Hydroides norvegicus (Serpulidae) as invertebrate
host. Folia Parasitologica 60(2) 149–154.
Køie, M; Karlsbakk, E; Nylund, A (2008). The marine herring myxozoan Ceratomyxa auerbachi (Myxozoa:
Ceratomyxidae) uses Chone infundibuliformis (Annelida: Polychaeta: Sabellidae) as invertebrate host. Folia
Parasitologica 55: 100–104.
Kongtorp, R T; Kjerstad, A; Taksdal, T; Guttvik, A; Falk, K (2004a). Heart and skeletal muscle inflammation
in Atlantic salmon, Salmo salar L.: a new infectious disease. Journal of Fish Diseases 27: 351–358.
Kongtorp, R T, Taksdal T, Lyngoy A (2004b). Pathology of heart and skeletal muscle infection (HSMI) in farmed
Atlantic salmon Salmo salar. Diseases of Aquatic Organisms 59(3): 217–224.

134 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Kovaleva A A, Gayevskaya, A V (1983). The first report of fish Myxosporidia from the open waters of the
southeastern Pacific Ocean. Vestnik Zoologii, 1: 6–11 (In Russian).
Kpatcha, T K; Faye, N; Diebakate, C; Fall, M; Toguebaye, B S (1997). Nouvelles espèces d'Henneguya
Thélohan, 1895 (Myxozoa, Myxosporea) parasites des poissons marins du Sénégal: Étude en microscopie
photonique et électronique. Annalesdes Sciences Naturelles, Zoologie, 13e (18): 81–91. (In French).
Kristoffersen, A B; Jensen, B B; Jansen, P A (2013). Risk mapping of heart and skeletal muscle inflammation in
salmon farming. Preventative Veterinary Medicine 109(1-2): 136–143.
Kuhnert, P; Bisgaard, M; Korczak, B M; Schwendener, S; Christensen, H; Frey, J (2012). Identification of
animal Pasteurellacea by MALDI-TOF mass spectrometry. Journal of Microbiological Methods 89(1):1–7.
Kumar, P; Abraham, T J; Adikesavalu, H (2016). Prevalence of Edwardsiella tarda in commercially important
finfish and shellfish of Bihar and West Bengal, India. Journal of Coast Life Medicine 4(1): 30–35 [Online]
Available from: http://www.doi.10.12980/jclm.4.2016apjtd-2014-0184 [Accessed 19 April 2020].
Kurita, J; Nakajima, K (2012). Megalocytiviruses. Viruses 4(4): 521–538 [Online] Available from:
http://www.doi.10.3390/v4040521 [Accessed 20 August 2019].
Kusuda, R; Toyoshima, T; Iwamura, Y; Sako, H (1976). Edwardsiella tarda from an epizootic of mullets Mugil
cephalus in Okisu Bay. Bulletin of the Japanese Society of Scientific Fisheries 42: 271–275.
Lafferty, K D; Harvell, C D; Conrad, J M; Friedman, C S; Kent, M L; Kuris, A M; Powell, E N; Rondeau, D;
Saksida, S M (2015). Infectious diseases affect marine fisheries and aquaculture economics. Annual review of
Marine Science 7: 471–496 [Online] Available from: https://www.doi.org/10.1146/annurev-marine-010814-015646
[Accessed 13 January 2020].
Laijimin, S; Razak, A A; Denil, D J; Ransagar, J; Wahid, M E A; Sade, A (2015). First detection of
megalocytivirus (Iridoviridae) in trash fish used for aquaculture feed in Sabah, Malaysia. International Journal of
Aquatic Science 6(1): 54–66.
Lamb, P (2019). Pete Lamb Fishing. Wellington, New Zealand. Personal communication.
Landsber, J H; Lom, J (1991). Taxonomy of the genera of the Myxobolus/Myxosoma group (Myxobolidae:
Myxosporea), current listing of species and revision of synonyms. Systematic Parasitology 18: 165–186.
Lane, H S; Booth, K; Pande, A; Jones, J B (2014). First report of the myxozoan parasite Myxobolus episquamalis
infecting grey mullet (Mugil cephalus) from New Zealand. New Zealand Journal of Marine and Freshwater
Research 49(2): 173–177 [Online] Available from: https://doi.org/10.1080/00288330.2014.984729 [Accessed 14
May 2019].
Langdon, J S (2007). Observations on new Myxobolus species and Kudoa species infecting the nervous system of
Australian fishes. Journal of Applied Ichthyology 6(2): 107–116 [Online] Available from:
http://www.doi.10.1111/j.1439-0426.1990.tb00508.x [Accessed 16 June 2019].
Langdon, J S; Thorne, T; Fletcher, W J (1992). Reservoir hosts and new clupeoid host records for the
myoliquefactive myxosporean parasite Kudoa thyrsites (Gilchrist). Journal of Fish Diseases 15(6): 459–471.
Landsberg, J H; Lom, J (1991). Taxonomy of the genera of the Myxosoma/Myxobolus group (Myxobolidae:
Myxosporea), current listing of species and revision of synonyms. Systematic Parasitology 18:165–186 [Online]
Available from: http://www.doi.10.1007/BF00009358 [Accessed 22 October 2019].
LeBlanc, F; Leadbeater, S; Laflamme, M; Gagne, N (2018). In vivo virulence and genomic comparison of
infectious Salmon Anaemia Virus isolates from Atlantic Canada. Journal of Fish Diseases 41(9): 1373–1384
[Online] Available from: http://www.doi.10.1111/ifd.12832 [Accessed 13 January 2020].
Lee, W L; Yun, H M; Kim, S R; Jung, S J; Oh, M J (2007). Detection of viral hemorrhagic septicemia virus
(VHSV) from marine fish in the South Western coastal area and East China Sea. Journal of Fish Pathology 20(3):
201–209.
Leighton, T (2020). BidFood Ltd, Wellington. Personal communication.
Leiro, J M; Piazzon, C; Dominguez, B; Mallo, N; Lamas, J (2012). Evaluation of some physical and chemical
treatments for inactivating microsporidian spores isolated from fish. International Journal of Food Microbiology
156(2): 152–160 [Online] Available from: http://www.doi.10.1016/j.ijfoodmicro.2012.03.017 [Accessed 19 June
2019].
Leong, T S; Colorni, A (2002). Infectious diseases of warmwater fish in salt water, pp 191–239 In: Woo, P T K;
Bruno, D W; Lim, L H S (Eds.) Diseases and disorders of fish in cage culture. CAB International, Wallingford,
United Kingdom, 354 p.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 135
Leotta, G A; Pineyro, P; Serena, S; Vigo, G B (2009). Prevalence of Edwardsiella tarda in Antarctic wildlife.
Polar Biology 32(5): 809–812.
Levsen, A (2015). A review of Kudoa species (Myxozoa, Multivalvulida) affecting seafood quality, with emphasis
on K. thyrsites in Atlantic mackerel (Scomber scombrus). Journal of Fisheries Science and Technology, Special
issue, Nha Trang University, Vietnam, 7 p.
Levsen, A; Jorgensen, A; Mo, T A (2008). Occurrence of postmortem myoliquefactive kudoosis in Atlantic
mackerel, Scomber scombrus L., from the North Sea. Journal of Fish Diseases 31(8): 601–611 [Online] Available
from: https://doi.org/10.1111/j.1365-2761.2008.00937.x [Accessed 24 September 2019].
Lewis, D H; Grumbles, L C; Mc Connell, S; Flowers, A I (1970). Pasteurella like bacteria from an epizootic in
menhaden and mullet in Galveston Bay. Journal of Wildlife Diseases 6: 160–162.
Li, Y C; Sato, H; Tanaka, S; Ohnishi, T; Kamata, Y; Sugita-Konishi, Y (2013). Characterization of the
ribosomal RNA gene of Kudoa neothunni (Myxosporea: Multivalvulida) in tunas (Thunnus spp.) and Kudoa
scomberi n. sp. in a chub mackerel (Scomber japonicus). Parasitological Research 112(5): 1991–2003.
Liltved, H; Vogelsang, C; Modahl, I; Dannevig, B H ((2006). High resistance of fish pathogenic viruses to UV
irradiation and ozonated seawater. Aquaculture Engineering 34: 72–82.
Loaharanu, P (1997). Irradiation as a cold pasteurization process of fish and seafood. Southeast Asian Journal of
Tropical Medicine and Public Health 28 (Supplement 1): 162–167.
Løkkeborg, S; Fernö, A; Humborstad, O B (2010). Fish behaviour in relation to longlines, pp 105–143 In: He, P.
(Ed.), Behaviour of marine fishes: Capture processes and conservation challenges. Wiley, Hoboken, New Jersey,
United States of America, 392 p.
Løkkeborg, S; Siikavuopio, S I; Humborstad, O B; Utne-Palm, A C; Ferter, K (2014). Towards more efficient
longline fisheries: Fish feeding behaviour, bait characteristics and development of alternative baits. Reviews in Fish
Biology and Fisheries 24: 985–1003.
Lom, J (2004). Morphology and ultrastructure of Sphaomyxa noblei sp. n. (Myxozoa), parasite of Heteroclinus
whiteleggii (Pisces) from Australian New South Wales coast. Folia Parasitologica 51: 19–26
Lom, J; Dykova, I (1992) Protozoan parasites of fishes. Elsevier Publishing, Amsterdam, The Netherlands, 315 p.
Lom, J; Dyková, I (1995). Myxosporea (Phylum Myxozoa), pp 97–148 In: Woo PTK (Ed.) Fish diseases and
disorders, Vol 1, Protozoan and Metazoan infections. CAB International, Wallingford, United Kingdom, 780 p.
Lom, J; Dykova, I (2006). Myxozoan genera: definition and notes on taxonomy, lifecycle terminology and
pathogenic species. Folia Parasitolica 53: 1–36.
Lom, J; Dykova, I (2013). Myxozoan genera: definition and notes on taxonomy, life-cycle terminology and
pathogenic species. Folia Parasitologica 53(1): 1–36 [Online] Available from:
http://www.doi.10.14411/fp.2006.001 [Accessed 1 November 2019].
Longshaw, M; Green, M J; Feist, S W (2004). Histopathology of parasitic infections in greater pipefish,
Syngnathus acus L., from an estuary in the UK. Journal of Fish Diseases 27: 245–248.
Lonnstrom, L; Wiklund, T; Bylund, G (1994). Pseudomonas anguilliseptica isolated from Baltic herring Clupea
harengus membras with eye lesions. Diseases of Aquatic Organisms 18: 143–147.
López-Romalde, S L; Magarinos, B; Nunez, S; Toranzo, A E; Romalde, J L (2003). Phenotypic and genetic
characterisation of Pseudomonas anguilliseptica strains isolated from fish. Journal of Aquatic Animal Health 15:
39–47.
Lowers, J M; Bartholomew, J R (2003). Detection of myxozoan parasites in oligochaetes imported as food for
ornamental fish. Journal of Parasitology 89(1): 84–91 [Online] Available from: https://doi.org/10.1645/0022-
3395(2003)089[0084:DOMPIO]2.0.CO;2 [Accessed 1 November 2019].
Lubat, V; Radujkovic, B; Marques, A; Bouix, G (1989). Parasites des poissons marins du Montenegro:
Myxosporidies. Acta Adriatica 30(1–2): 31–50.
Lupiani, B; Hetrick, F; Samal, S K (1993). Genetic analysis of aquareoviruses using RNA-RNA blot
hybridisation. Virology 197(1): 475–479.
Lyngstad, T M; Hjortaas, M J; Kristoffersen, A B; Markussen, T; Karlsen, E T; Jonassen, C M; Jansen, P A
(2011). Use of molecular epidemiology to trace transmission pathways for infectious salmon anaemia virus (ISAV)
in Norwegian salmon farming. Epidemics 3(1): 1–11 [Online] Available from:
http://www.doi.10.1016/j.epidem.2010.11.001 [Accessed 17 July 2020].

136 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
MacDiarmid, S C (1993). Risk analysis and the importation of animals and animal products. Revue scientifique et
technique 12(4): 1093–1097.
MacDiarmid, S C (1994). The risk of introducing exotic diseases of fish into New Zealand through importation of
ocean-caught Pacific salmon from Canada, MAF Regulatory Authority, Ministry of Agriculture and Forestry,
Wellington, New Zealand [Online] Available from: https://www.teururakau.govt.nz/dmsdocument/2799/direct
[Accessed 7 October 2019].
MacKelvie, R M; Desautels, D (1975). Fish Viruses -Survival and inactivation of infectious pancreatic necrosis
virus. Journal of the Fisheries Research Board of Canada 32(8): 1267–1273 [Online] Available from:
https://www.doi.org/10.1139/f75-147 [Accessed 6 August 2019].
MacKenzie, K; Kalavati, C (2014). Myxosporean parasites of marine fishes: their distribution in the world’s
oceans. Parasitology 141: 1709–1717.
MacLeod, L; Murray, Ag; Gregory, A (2003). Survival of infectious salmon anaemia virus (ISAV) in aquatic
environments. Abstract from 11th EAFP (European Association of Fish Pathologists) Conference, Malta, 2003.
MacMillan, J R; Mulcahy, D; Landolt, M (1980). Viral erythrocytic necrosis: some physiological consequences
of infection in chum salmon (Oncorhynchus keta). Canadian Journal of Fisheries and Aquatic Sciences 37: 799–
804.
Maeno, Y; Nagasawa, K; Sorimachi, M (1993). Kudoa intestinalis n. sp. (Myxosporea: Multivalvulida) from the
intestinal musculature of the stripped mullet, Mugil cephalus, from Japan. Journal of Parasitology 79: 190–192.
Maes, J M; Ollevier, R (2002). Size structure and feeding dynamics in estuarine clupeoid fish schools: field
evidence for the school trap hypothesis. Aquatic Living Resources 15(4): 211–216 [Online] Available from:
https://doi.org/10.1016/S0990-7440(02)01181-6 [Accessed 10 December 2019].
Magi, G E; Lopez-Romalde, S; Magarinos, G E; Lamas, J L; Toranzo, A E; Romalde, L (2009). Experimental
Pseudomonas anguilliseptica infection in turbot Psetta maxima (L.): a histopathological and immunohistochemical
study. European Journal of Histochemistry 53(2): 73–80 [Online] Available from:
http://www.doi.10.4081/ejh.2009.e9 [Accessed 30 August 2019].
Mansour, L; Harrath, A H; Abd-Elkader, O H; Alwasel, S; Abdel-Baki, A A; Al Omar, S Y (2014). Structural
and molecular characterization of Kudoa quraishii n. sp. from the trunk muscle of the Indian mackerel Rastrelliger
kanagurta (Perciforme, Scombridae) in Saudi Arabia coasts. Parasitology Research 113(4):1361–1370.
Marsh, D; Mkwra, L (2013). Review of freshwater non-market value studies. Department of Economics,
University of Waikato, Hamilton, New Zealand, 144 p.
Marshall, W L; Sitja-Bobadilla, A; Brown, H M; MacWilliam, T; Richmond, Z; Lamson, H; Morrison, DB,
Afonso, LOB (2015). Long‐term epidemiological survey of Kudoa thyrsites (Myxozoa) in Atlantic salmon (Salmo
salar L.) from commercial aquaculture farms. Journal of Fish Diseases 39(8): 929–946 [Online] Available from:
https://doi.org/10.1111/jfd.12429 [Accessed 8 May 2020].
Marty, G D; Freiberg, E F; Meyers, T R; Wilcock, J; Farver, T B; Hinton, D E (1998). Viral hemorrhagic
septicemia virus, Ichthyophonus hoferi, and other causes of morbidity in Pacific herring Clupea pallasi spawning in
Prince William Sound, Alaska, USA. Diseases of Aquatic Organisms 32(1): 15–40.
Mastan, S A (2013). Pseudomonas septicaemia in Labeo rohita (Ham.) and Cyprinus carpio (Linn.) in Andhra
Pradesh-natural occurrence and artificial challenge. International Journal of Pharmacy and Pharmaceutical
Sciences 5(2): 564–568.
Matsukane, Y; Sato, H; Tanaka, S; Kamata, Y; Sugita-Konishi, Y (2011). Kudoa iwatai and two novel Kudoa
spp., K. trachuri n. sp. and K. thunni n. sp. (Myxosporea: Multivalvulida), from daily consumed marine fish in
western Japan. Parasitology Research 108(4): 913–926 [Online] Available from:
https://www.doi.org/10.1007/s00436-010-2133-2 [Accessed 20 September 2019].
Matsusato, T (1975). On the lymphocystis disease in cultured yellowtail. Fish Pathology 10: 90–93.
McAllister, P E; Owens, W J (1992). Recovery of infectious pancreatic necrosis virus from the faeces of wild
piscivorous birds. Aquaculture 106(3-4): 227–232 [Online] Available from: https://www.doi.org/10.1016/0044-
8486(92)90254-I [Accessed 24 October 2019].
McArthur, C P; Sengupta, S (1982). Antigenic mimicry of eel tissues by a myxosporidian parasite? Zeitschrift für
Parasitenkunde 66: 249–255.
McColl K., Slater J., Jeyasekaran G., Hyatt A.D., Crane M.S (2004a). Detection of white spot syndrome virus and
yellow head virus in prawns imported into Australia, Australian Veterinary Journal 82: 69–74.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 137
McColl, K; Steeper, K; Crane, M (2004b). Aquatic Animal Health Subprogram: development of improved
procedures for the identification of aquatic birnaviruses. FRDC Project No. 2001/620 CSIRO Livestock Industries,
Australian Animal Health Laboratory, AAHL Fish Diseases Laboratory, Geelong, Victoria, Australia.
McColl, KA; Davies, K R; Young, J G; Crane, M St J (2009). Aquatic birnavirus infections of finfish. Australia
and New Zealand Standard Diagnostic Procedure, Department of Agriculture and Water Resources, Canberra,
Australia, 20 p. [Online] Available from:
http://www.agriculture.gov.au/SiteCollectionDocuments/animal/ahl/ANZSDP-Aquatic-birnaviruses.pdf [Accessed
15 August 2018].
McDaniels, A E; Cochran, K W; Gannon, J J; Williams, G W (1983). Rotavirus and reovirus stability in
microorganism free distilled and wastewaters. Water Research 17(10): 1349–1353.
McGladdery, S E (1999). Flexibacter maritimus, causal agent of flexibacteriosis in marine fish. ICES Identification
leaflet 44. ICES Working Group on Pathology and Diseases of Marine Organisms. International Council for the
Exploration of the Sea, Copenhagen, Denmark. 6 p.
McVicar, A H (1982). Ichthyophonus infection of fish, pp. 243-269 In: Roberts R J (Ed.), Microbial Disease of
Fish, Academic Press, London, United Kingdom.
Meglitsch, P A (1947). Studies on Myxosporidia from the Beaufort Region. II. Observations on Kudoa clupeidae
(Hahn), gen. nov. The Journal of Parasitology 33(3): 271–277.
Melville, V; Johnston, C (2010). Supplementary Risk Analysis: Extension of countries eligible to export frozen,
skinless, boneless fillet meat of Pangasius spp. under the import health standard fisfilic.spe. Ministry of Agriculture
and Forestry, Wellington, New Zealand, 15 p.
Meng, F; Yokoyama, H; Shirakashi, S; Grabner, D; Ogawa, K; Ishimaru, K; Sawada, Y; Murata, O (2011).
Kudoa prunusi n. sp. (Myxozoa: Multivalvulida) from the brain of Pacific bluefin tuna Thunnus orientalis
(Temminck & Schlegel, 1844) cultured in Japan. Parasitology International 60(1): 90–96 [Online] Available from:
https://www.doi.org/10.1016/j.parint.2010.11.006 [Accessed 17 September 2019].
Merino, G; Barange, M; Mullon, C (2014). Role of anchovies and sardines as reduction fisheries in the world fish
meal production: Overview of the interaction between the resource and environmental and socioeconomic drivers,
pp 285–307 In: Gainos, K (Ed.). Biology and ecology of sardines and anchovies, CRC Press, Taylor & France, New
York, United States of America, 394 p.
Meyer, F P; Bullock, G L (1973). Edwardsiella tarda, a new pathogen of channel catfish (Ictalurus punctatus).
Applied Microbiology 25(1): 155–156.
Meyers, T R; Hauck, A K; Blankenbeckler, W D; Minicucci, T (1986). First report pf viral erythrocytic necrosis
in Alaska, USA, associated with epizootic mortality in Pacific herring Clupea harengus pallasii. Journal of Fish
Diseases 9: 479–491.
Meyers, T R; Short, S; Lipson, K (1999). Isolation of the North American strain of viral hemorrhagic septicemia
virus (VHSV) associated with epizootic mortality in two new host species of Alaskan marine fish. Diseases of
Aquatic Organisms 38:81−86.
Meyers, T R; Short, S; Lipson, K; Batts, W N; Winton, J R; Wilcock, J; Brown, E (1994). Association of viral
hemorrhagic septicemia virus with epizootic haemorrhages of the skin in Pacific herring Clupea harengus pallasi
from Prince William Sound and Kodiak Island, Alaska, USA. Diseases of Aquatic Organisms 19: 27–37.
Meyers, T, Sullivan, J; Emmenegger, E; Follett, J (1992). Identification of viral hemorrhagic septicaemia virus
isolated from Pacific cod Gadus macrocephalus in Prince William Sound, Alaska, USA. Diseases of Aquatic
Organisms 12: 167–175 [Online] Available from: http://www.doi.10.3354/dao012167 [Accessed 7 October 2019].
Meyers, T R; Winton, J R (1995). Viral haemorrhagic septicaemia virus in North America. Annual Review of Fish
Diseases 5: 3–24.
Miller, O; Cipriano, R C (2003). International response to infectious salmon anaemia: prevention, control, and
eradication: Proceedings of a symposium, 3–4 September 2002, New Orleans, Louisiana. NMFS Technical Bulletin
1902, National Marine Fisheries Service, United States Department of Agriculture, Washington DC, United States
of America, 194 p.
Minami, T; Nakamura, M; Ikeda, Y; Ozaki, H (1979). A BetaHemolytic Streptococcus isolated from cultured
yellowtail. Fish Pathology 14: 33–38.
Mishra, A; Nam, G H; Gim, J A; Lee, H E; Jo, A; Kim, H S (2018). Current challenges of Streptococcus
infection and effective molecular, cellular, and environmental control methods in aquaculture. Molecules and Cells
41(6): 495–505.

138 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Miyazaki, T; Egusa, S (1976). Histopathological studies of Edwardsiellosis of the Japanese eel (Anguilla
japonica)-I. Suppurative interstitial nephritis form. Fish Pathology 11: 33–43.
Miyazaki, T; Kaige, N (1985). Comparative histopathology of Edwardsiellosis in fishes. Fish Pathology 20: 219–
227.
Mjaaland, S; Hungnes, O; Teig, A; Dannevig, B H; Thorud, K; Rimstad, E (2002). Polymorphism in the
infectious salmon anaemia virus hemagglutinin gene; importance and possible implications for evolution and
ecology of infectious salmon anaemia disease. Virology 302: 379–391.
MOH, (2019). Leptospirosis, Communicable disease control manual, Ministry of Health, Wellington, New Zealand
[Online] Available from: https://www.health.govt.nz/our-work/diseases-and-conditions/communicable-disease-
control-manual/leptospirosis [Accessed 5 June 2019].
Mohr, P G; Moody, N J G; Williams, L M; Hoad, J; Cummins, D M; Crane, M St J (2015). Molecular
confirmation of an ISKNV-like virus detected in farmed ornamental Platys (Xiphophorus maculates), Diseases of
Aquatic Organisms 116: 103–110 [Online] Available from: http://www.doi.10.3354/dao02896 [Accessed 13
January 2020].
Mohr, P G; Crane, M St J; Hoad J; Williams, L M; Cummins, D; Neave, M J; Shiel, B; Beddome, G;
Michaelski, W P; Peck, G R; Samsung, F; Wynne, J W; Cramen, S G; Hyatt, A G; Moody, N J G (2020).
Pilchard orthomyxovirus (POMV). I. Characterisation of an emerging virus isolated from pilchards Sardinops sagax
and Atlantic salmon Salmo salar. Diseases of Aquatic Organisms 139: 35–50 [Online] Available from:
http://www.doi.10.3354/dao03470 [Accessed 17 August 2020].
Molloy, S D; Pietrak, M R; Bricknell, I; Bouchard, D (2013). Experimental transmission of infectious pancreatic
necrosis virus from the blue mussel, Mytilus edulis, to cohabitating Atlantic salmon (Salmo salar) smolts. Applied
and Environmental Microbiology 79(19): 5882–5890 [Online] Available from:
http://www.doi.10.1128/AEM.01142- [Accessed 23 October 2019].
MolluscaBase, (2019). World taxonomic register of mollusc species, Flanders Marine Institute, Belgium [Online]
Available from: http://www.molluscabase.org [Accessed 17 December 2019].
Molnar, K; Eszterbauer, E (2015). Specificity of infection sites in vertebrate hosts, pp 295–313 In: Okamura, B;
Gruhl, A; Bartholomew, J (Eds.) Myxozoan Evolution, Ecology and Development. Springer, New York, United
States of America, 441 p.
Montero, F; Cuadrado, M; Padros, F; Crespo, S; Raga, J A (2007). Cryptocaryon irritans and Enteromyxum
leei, two threats for the culture of Diplodus puntazzo in the Mediterranean. Bulletin of the European Association of
Fish Pathologists 27: 242–247.
Moody, N J; Horwood, P F (2008). Betanodaviruses in finfish. Australian and New Zealand Standard Diagnostic
Procedures: Nodaviruses. Australian Government, Canberra, Australia, 18 p.
Moran, J D W; Kent, M L (1992). Diagnosing Kudoa thyrsites (Myxozoa) infection in fish, pp 329–345 In:
Cunningham CO (Ed.) Molecular diagnosis of salmonid diseases. Springer Science, New York 345 p.
Moran, J D W; Margolis, L; Webster, J M; Kent, M L (1999a). Development of Kudoa thyrsites (Myxozoa:
Myxosporea) in netpen-reared Atlantic salmon determined by light microscopy and a polymerase chain reaction test.
Diseases of Aquatic Organisms 37:185–193.
Moran JDW, Whitaker DJ. Kent ML (1999b). A review of the myxosporean genus Kudoa Meglitsch, 1947, and
its impact on the international aquaculture industry and commercial fisheries. Aquaculture 172: 163–196.
Moran, J D W; Whitaker, D J; Kent, M L (1999c). Natural and laboratory transmission of the marine myxozoan
parasite Kudoa thyrsites to Atlantic salmon. Journal of Aquatic Animal Health 11:110–115
Moreno, P; Oleveira, J G; Labella, A; Cutrin, J M; Baro, J C; Borrego, J J; Dopazo, C P (2014). Surveillance
of viruses in wild fish populations in areas around the Gulf of Cadiz (South Atlantic Iberian Peninsula). Applied and
Environmental Microbiology 80(20): 6560 – 6571 [Online] Available from:
http://www.aem.asm.org/content/aem/80/20/6560.full.pdf [Accessed 17 December 2019].
Mori, K; Nakai, T; Muroga, K; Arimoto, M; Mushiake, K; Furusawa, I (1992). Properties of a new virus
belonging to Nodaviridae found in larval striped jack (Pseudocaranx dentex) with nervous necrosis. Virology 187:
368– 371.
Mori, K; Nakai, T; Nagahara, M; Muroga, K; Mekuchi, T; Kanno, TA (1991). A viral disease in hatchery-
reared larvae and juveniles of redspotted grouper. Fish Pathology 26: 209–210.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 139
Mori, K; Sugaya, T; Nishioca, T; Gomez, D K; Fujunamy, Y; Oka, M; Arimoto, M; Okinaka, Y; Nakai,
T (2005). Detection of betanodaviruses from feed fish used in marine aquaculture. Abstract O-142 In: Proceedings
of the 12th International Conference Diseases of Fish and Shellfish, European Association of Fish Pathologists.
Copenhagen (Denmark), 11–16 September 2005.
Morris, G J; Smith, D; Coulson, G E (1988). A comparative study of the changes in the morphology of hyphae
during freezing and viability upon thawing for twenty species of fungi. Journal of General Microbiology 134: 2897–
2906.
Mortensen, H F; Heuer, O E; Lorenzen, N; Otte, L; Olesen, N J (1999). Isolation of viral haemorrhagic
septicaemia virus (VHSV) from wild marine fish species in the Baltic Sea, Kattegat, Skagerrak and the North Sea.
Virus Research 63 (1–2): 95–106.
Mortensen, S H; Bachere, E; Le Gall, G; Mialhe. E (1992). Persistence of infectious pancreatic necrosis virus
(IPNV) in scallops Pecten maximus. Diseases of Aquatic Organisms 12: 221–227.
Mortensen, S H; Hjeltnes, B; Rodseth, O; Krogsrud, J; Christie, J E (1990). Infectious pancreatic necrosis
virus, serotype N1, isolated from Norwegian halibut (Hippoglossus hippoglossus), turbot (Scophthalmus maximus)
and scallops (Pecten maximus). Bulletin of the European Association of Fish Pathologists 10(2): 42–43.
MPI, (2001). Import health standard for the importation into New Zealand of precooked heat-and-eat meals
containing animal products for human consumption from Australia. Ministry for Primary Industries, Wellington,
New Zealand, 11 p. [Online] Available from: https://www.biosecurity.govt.nz/dmsdocument/1817-heat-and-eat-
meals-containing-animal-products-from-australia-import-health-standard [Accessed 21 January 2020].
MPI, (2008). Import health standard for marine fisheries products for human consumption from all countries,
FISMARIC.ALL, 5 p [Online] Available from: https://www.mpi.govt.nz/dmsdocument/1779-marine-fisheries-
products-for-human-consumption-from-all-countries-import-health-standard [Accessed 27 August 2019].
MPI, (2010). Import risk analysis: Tropical, subtropical and temperate freshwater and marine ornamental fish and
marine molluscs and crustaceans. Review of submissions on supplementary risk analysis. Ministry for Primary
Industries, Wellington, New Zealand, 108 p.
MPI, (2011). Import health Standard for Fish bait and fish food (FISFOOIC.ALL), Ministry for Primary Industries,
Wellington, New Zealand, 11 p. [Online] Available from: https://www.biosecurity.govt.nz/dmsdocument/1778-fish-
food-and-fish-bait-from-all-countries-import-health-standard [Accessed 7 October 2019].
MPI, (2018a). Fisheries Assessment Plenary: Stock assessment and stock status– Volume 1: Introductory section to
groper [Online] Available from: https://www.fs.fish.govt.nz/Doc/24612/May%20Plenary%202018%20-
%20Volume%201.pdf.ashx [Accessed 7 October 2019].
MPI, (2018b). Fisheries Assessment Plenary: Stock assessment and stock status– Volume 2: Hake to pilchard
[Online] Available from: https://www.fs.fish.govt.nz/Doc/24613/May%20Plenary%202018%20-
%20Volume%202.pdf.ashx [Accessed 7 October 2019].
MPI, (2018c). Fisheries Assessment Plenary: Stock assessment and stock status –Volume 3: Pipi to yellow-eyed
mullet [Online] Available from: https://www.fs.fish.govt.nz/Doc/24614/May%20Plenary%202018%20-
%20Volume%203.pdf.ashx [Accessed 7 October 2019].
Munday, B L; Kwang, J; Moody, N (2002). Betanodavirus infections of teleost fish: a review. Journal of Fish
Diseases 25: 127–142.
Munday, L; Su, X Q; Hershberger, H C (1998). A survey of product defects in Tasmanian Atlantic salmon
(Salmo salar). Aquaculture 169: 297–302.
Munro, A L S; McVicar, A H; Jones, R (1983). The epidemiology of infectious disease in commercially
important wild marine fish Rapports et process-verbaux des reunions (Marine Science Symposia: International
Council for the Exploration of the Sea) 182: 21–32.
Munro, T S; Midtlyng, P J (2011). Infectious pancreatic necrosis and associated aquatic birnaviruses, pp 1–65 In:
Woo, P T K; Bruno, D W (Eds.). Fish diseases and disorders Volume 3: Bacterial and Fungal Infections (2nd
Edition), CAB International, Wallingford, United Kingdom, 930 p.
Muroga, K; Jo, Y; Sadawa, T (1975). Studies on red spot disease of pond cultured eels. II: Pathogenicity of the
causative bacterium Pseudomonas anguilliseptica. Fish Pathology (Japan) 9(2): 107–114.
Murray, A G (2002). Shipping and the spread of infectious salmon anaemia in Scottish aquaculture. Emerging
Infectious Diseases 8: 1–5.

140 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Murray, A G; Amundred, T L; Gillibrand, P A (2005). Models of hydrodynamic pathogen dispersal affecting
Scottish salmon production: modelling shows how Scotland eradicated ISA, but not IPN. Bulletin of the Aquaculture
Association of Canada 105: 79–86.
Mussely, H; Goodwin, E (2012). Feasibility of land-based aquaculture in New Zealand. Cawthron Report No.
2094, 20p.
Muzquiz, J L; Royo, F M; Ortega, C; De Blas, I; Ruiz, I; Alonso, JL (1999). Pathogenicity of streptococcosis in
rainbow trout (Oncorhynchus mykiss): dependence on age of diseased fish. Bulletin of the European Association of
Fish Pathologists 19(3): 114–120.
Nagel, D; Serritella, A; Layden, T J (1982). Edwardsiella tarda gastroenteritis associated with a pet turtle.
Gastroenterology 82: 1436-1437
Nakajima, K; Inouye, K; Sorimachi, M (1998). Viral diseases in cultured marine fish in Japan. Fish Pathology
33:181–188.
Nakajima, K; Maeno, Y; Arimoto, M; Inouye, K; Sorimachi, M (1993). Viral deformity of yellowtail
fingerlings. Fish Pathology 28: 125–129.
Naldoni, J; Adriano, E A; Hartigan, A; Sayer, C; Okamura, B (2019). Malacosporean myxozoans exploit a
diversity of fish hosts. Parasitology 146: 968–978 [Online] Available from:
https://www.doi.org.10.1017/S0031182019000246 [Accessed 17 December 2019].
Narasimhamurti, C C (1979). The eimeriid Aggregata kudoi n.sp. from Sepia elliptica Agnew. Parasitology 20:
154–158.
Narasimhamurti, C C; Kalavati. C (1979). Kudoa tetraspora n. sp. (Myxosporidea: Protozoa) parasitic in the
brain tissue of Mugil cephalus. Proceedings of the Indian Academy of Sciences. 88(1): 85–89.
Nash G, Anderson IA, Shariff M, Shamsudin M (1987). Bacteriosis associated with epizootic in the giant sea
perch, Lates calcarifer, and the estuarine grouper Epinephelus tauvina, cage culture in Malaysia. Aquaculture 67(1–
2): 105–111.
Nehring, B B; Schisler, G; Chiaramonte, L; Horton, A; Poole, B (2015). Assessment of the long-term viability
of the myxospores of Myxobolus cerebralis as determined by production of the actinospores by Tubifex tubifex.
Journal of Aquatic Animal Health 27(1): 50–56.
Ngan, N; Morris, A; de Chalain, T (2005). Mycobacterium fortuitum infection caused by a cat bite. The New
Zealand Medical Journal 118: 1211, 4 p [Online] Available from: http://www.nzma.org.nz/journal/118-1211/1354
[Accessed 15 May 2019].
Nguyen, H T; Kanai K; Yoshikoshi K (2001). Immunohistochemical examination of experimental Streptococcus
iniae infection in Japanese flounder Paralichthys olivaceus. Fish Pathology 36:169–178.
Nguyen, H; Kanai, K; Yoshikoshi, K (2002). Ecological investigation of Streptococcus iniae in cultured Japanese
flounder (Paralichthys olivaceus) using selective isolation procedures. Aquaculture 205: 7–17.
Nguyen, H D; Nakai, T; Muroga, K (1996). Progression of striped jack nervous necrosis virus (SJNNV) infection
in naturally and experimentally infected striped jack Pseudocaranx dentex larvae. Diseases of Aquatic Organisms
24: 99–105.
Nicholson, B L; Reno, P W (1981). Viral erythrocytic necrosis in marine fish. Fish Pathology 15: 129–134.
Nishi, S; Yamashita, H; Kawato, Y; Nakai, T (2016). Cell Culture Isolation of Piscine Nodavirus (Betanodavirus)
in Fish-Rearing Seawater. Applied and Environmental Microbiology 82(8): 2537–2544 [Online] Available from:
http://www.doi.10.1128/AEM.03834-15 [Accessed 17 December 2019].
NIWA, (2014). Grass carp Ctenopharyngodon idella (Valenciennes, 1844) [Online] Available from:
https://www.niwa.co.nz/freshwater-and-estuaries/nzffd/NIWA-fish-atlas/fish-species/grass_carp [Accessed 12
February 2015].
NIWA, (2017a). Aquaculture species: Yellowtail kingfish, National Institute for Water and Atmospheric Science,
Auckland, New Zealand. [Online] Available from: https://www.niwa.co.nz/aquaculture/aquaculture-
species/yellowtail-kingfish [Accessed 20 October 2019].
NIWA, (2017b). Aquaculture species: Hapuku, National Institute for Water and Atmospheric Science, Auckland,
New Zealand. [Online] Available from: https://www.niwa.co.nz/aquaculture/aquaculture-species/hapuku [Accessed
20 October 2019].

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 141
NIWA, (2017c). Aquaculture species: Eels, National Institute for Water and Atmospheric Science, Auckland, New
Zealand. [Online] Available from: https://www.niwa.co.nz/aquaculture/aquaculture-species/eels [Accessed 20
October 2019].
NIWA, (2018). Identification guides and fact sheets [Online] Available from: https://www.niwa.co.nz/freshwater-
and-estuaries/nzffd/NIWA-fish-atlas/fish-species/species/ [Accessed 10 July 2018].
NIWA, (2019). Rainbow trout, Oncorhynchus mykiss (Walbaum, 1792). Identification guides and fact sheets
[Online] Available from: https://www.niwa.co.nz/freshwater-and-estuaries/nzffd/NIWA-fish-atlas/fish-
species/rainbow trout [Accessed 10 July 2018].
NIWA, (2020). New Zealand freshwater fish database: Galaxiidae, NIWA Atlas of NZ freshwater fishes, National
Institute for Water and Environmental Research, Auckland, New Zealand [Online] Available from:
https://www.niwa.co.nz/freshwater-and-estuaries/nzffd/NIWA-fish-atlas/fish-species/galaxiidae [Accessed 8
January 2020].
Noga, E J (2010). Fish disease: diagnosis and treatment. John Wiley, New York, United States of America, 378 p.
Noga, E J (2012). A synopsis of mycotic diseases of marine fishes and invertebrates, pp 143–168 In: Perkins. F O;
Cheng, T C (Eds.). Pathology in Marine Science, Elsevier, New York, United States of America, 538 p.
Nolan, D; Stephens, F; Crockford, M; Jones, J B; Snow, M (2015). Detection and characterization of viruses of
the genus Megalocytivirus in ornamental fish imported into an Australian border quarantine premises: an emerging
risk to national biosecurity. Journal of Fish Diseases 38(2): 187–195.
Norman, M D; Finn, J K; Hochberg, F G (2014). Family Octopodidae, pp. 36-215 In: Jereb, P; Roper, C F E;
Norman, M D; Finn, J K (Eds.). Cephalopods of the world. An annotated and illustrated catalogue of cephalopod
species known to date. Volume 3. Octopods and Vampire Squids. FAO Species Catalogue for Fishery Purposes
4(3): 353 pp. Food and Agriculture Organisation of the United Nations (FAO), Rome, Italy 11 p.
Norman. R E; Brosnahan, C; Fischer, J; Frazer, J; Johnston, C; Jones, B; Keeling, S; Pande, A; Pulford, D;
Rawdon, T; Williams, R (2013). Salmon mortality investigation REW-1017, Pelorus Sound. MPI Technical Paper
2013/19. Ministry for Primary Industries, Wellington, New Zealand, 14 p.
Novis, P M (2004). A taxonomic survey of Microspora (Chlorophyceae, Chlorophyta) in New Zealand, New
Zealand Journal of Botany 42(1): 153–165 [Online] Available from:
http://www/doi.10.1080/0028825X.2004.9512896 [Accessed 28 May 2019].
Novoslavskij, A; Terentjeva, M; Eizenberga, I; Valcina, O; Bartkevics, V; Berzins, A (2015). Major foodborne
pathogens in fish and fish products: A review. Annals of Microbiology 66(1): 1–15.
Nowak, B; Rough, K; Ellis, D; Crane, M; Cameron, A; Clarke, S (2003). Aquafin CRC- Southern bluefin tuna
aquaculture subprogram: A risk assessment of factors influencing the health of southern bluefin tuna. FRDC &
Aquafin CRC, ISBN 1 86295 099 7, 139 p.
Nowak, B F; LaPatra, S E (2006). Epitheliocystis in fish. Journal of Fish Diseases 29(10): 573–636.
Nylund, A; Devold, M; Mullins, J; Plarre, H (2002). Herring (Clupea harengus): A host for infectious salmon
anaemia virus (ISAV). Bulletin of the European Association of Fish Pathologists 22: 311–318.
Nylund, A; Ottem, K F; Watanabe, K; Karlsbakk, E; Krossoy, B (2006). Francisella sp. (Family
Francisellaceae) causing mortality in Norwegian cod (Gadus morhua) farming. Archives of Microbiology 185(5):
38–-392.
Nylund, A; Ottem, K F (2006). Francisellosis a new cod (Gadus morhua) disease caused by Francisella n. sp.
DIPnet (Disease interactions and pathogen exchange between farmed and wild aquatic animal populations – a
European network), Newsletter 45. VESO (Norwegian Veterinary Centre for Contract Research), 2 p.
NZ Fungi, (2019). New Zealand Fungi (and Bacteria). New Zealand fungi and bacteria reported from New
Zealand database, Landcare Ltd, Lincoln, New Zealand [Online] Available from:
https://www.landcareresearch.co.nz/resources/data/nzfungi [Accessed 18 October 2019].
NZOR, (2017). New Zealand Organisms Register. Landcare Research Limited, Lincoln, New Zealand [Online]
Available from: http://www.nzor.org.nz/ [Accessed 7 October 2019].
Odhiambo, J A; Birgen, J K; Okembo, P O; Alaro, L O (2018). microbial quality of preserved sardines sold in
Mombasa. American Scientific Research Journal for Engineering, Technology and Sciences (ASRJETS) 41(1):
133–145.

142 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Ogut, H; Altuntas, C (2014). Survey of viral haemorrhagic septicaemia virus in wild fishes in the southeastern
Black Sea. Diseases of Aquatic Organisms 109(2): 99–106 [Online] Available from:
http://www.doi.10.3354/dao02728 [Accessed 6 May 2019].
OHSU (2020). Cryobiology. ONPC Saturday Academy, Part 1. Oregon National Primate Centre, Oregon Health
and Science University, Beaverton, Oregon, United States of America [Online] Available from:
https://www.ohsu.edu/sites/default/files/2019-02/8b.%20Cryobiology%20Lecture%20Part%201%20-
%20PowerPoint.pdf [Accessed 22 September 2020].
Oidtmann, B; Dixon, P; Way, K; Joiner, C; Bayley, A E (2017). Risk of waterborne virus spread – review of
survival of relevant fish and crustacean viruses in the aquatic environment and implications for control measures.
Reviews in Aquaculture 10(3): 641–669. [Online] Available from: https://www.doi.org/10.111/raq.12192 [Accessed
7 October 2019].
Oidtmann, B; Heitz, B E; Rogers, D; Hoffmann, R W (2002). Transmission of crayfish plague. Diseases of
Aquatic Organisms 52: 159–167.
Oidtmann, B; Johnston, C; Klotins, K; Mylrea, G; Van, P T; Cabot, S; Rosado-Martin, P; Ababouch, L;
Berthe, F (2013). Assessment of the safety of aquatic animal commodities for international trade: The OIE Aquatic
Animal Health Code, Transboundary and Emerging Diseases 60(1): 27–38.
Oidtmann, B; Joiner, C; Reese, R A; Stone, D; Dodge, M; Dixon, P (2011). Risks associated with commodity
trade: transmission of viral haemorrhagic septicaemia virus (VHSV) to rainbow trout fry from VHSV-carrying
tissue homogenates. Transboundary and Emerging Diseases 58: 224–231.
OIE, (2010). Handbook on import risk analysis for animals and animal products, Volume 1, 2nd Edition,
Introduction and qualitative analysis. World Organisation for Animal Health, Paris, France [Online] Available from:
https://www.oie.int/doc/ged/D6586.pdf [Accessed 7 October 2019].
OIE, (2019a). Aquatic animal health code. World Organisation for Animal Health, Paris, France [Online] Available
from: https://www.oie.int/standard-setting/aquatic-code/ [Accessed 6 May 2019].
OIE, (2019b). Manual of Diagnostic Tests for Aquatic Animals, World Organisation for Animal Health, Paris,
France [Online] Available from: https://www.oie.int/standard-setting/aquatic-manual/ [Accessed 6 May 2019].
OIE, (2019). Official disease status: New Zealand, WAHIS (World Animal Health Information System), World
Organisation for Animal Health, Paris, France [Online] Available from: https://www.oie.int/animal-health-in-the-
world/official-disease-status/ [Accessed 6 May 2019].
Okamura, B; Gruhl, A; Bartholomew, J L (2014). Myxozoan Evolution, Ecology and Development. Springer
Science, New York, United States of America. 441 p.
Okamura, B; Gruhl, A (2015). Myxozoan Affinities and Route to Endoparasitism. pp 23–44 In: Myxozoan
Evolution, Ecology and Development, Springer Science, Berlin, Germany.
Okamura, B; Gruhl, A; Bartholomew, J (2015). An Introduction to Myxozoan Evolution, Ecology and
Development, pp 1–20 In: Okamura, B; Gruhl, A; Bartholomew, J (Eds.). Myxozoan evolution, ecology and
development, Springer Science, New York, United States of America.
Okamura, B; Hartigan, A; Naldoni, J (2018). Extensive uncharted biodiversity: the parasite
dimension. Integrative and Comparative Biology 58:1132–1145 [Online] Available from:
https://academic.oup.com/icb/article/58/6/1132/5026008 [Accessed 18 August 2020].
Olson, M E; Goh, J; Phillips, M; Guselle, N; McAllister, T A (1999). Giardia cyst and Cryptosporidium oocyst
survival in water, soil, and cattle faeces. Journal of Environmental Quality 28(6): 1991–1996.
Ohnishi, T; Fujiwara, M; Tomaru, A; Yoshinari, T; Sugita-Konishi, Y (2017). Cryopreservation of Kudoa
septempunctata sporoplasm using commercial freezing media. Parasitology Research 116: 425–427 [Online]
Available from: https://doi.org/10.1007/s00436-016-5262-4 [Accessed 29 July 2020].
Ottem, K F; Nylund, A; Isaksen, T E; Karlsbakk, E; Bergh, O (2008). Occurrence of Francisella piscicida in
farmed and wild Atlantic cod, Gadus morhua L., in Norway. Journal of Fish Diseases 31: 525–534 [Online]
Available from: http://www.doi. 10.1111/j.1365-2761.2008.00930.x. [[Accessed 15 April 2020].
Ottem, K F; Nylund, A; Karlsbakk, E; Fris-Moller, A; Kamaishi, T (2009). Elevation of Francisella
philomiragia subsp. noatunensis Mikalsen et al. (2007) to Francisella noatunensis comb. nov. [syn. Francisella
piscicida Ottem et al. (2008) syn. nov.] and the characterisation of Francisella noatunensis subsp. orientalis subsp.
nov., two important fish pathogens. Journal of Applied Microbiology 106: 1231–1243.
Ovcharenko, M (2016). Microparasites of worldwide mullets. Annals of Parasitology 61(4): 229–239.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 143
Oye, A K; Rimstad, E (2001). Inactivation of infectious salmon anaemia virus, viral haemorrhagic septicaemia
virus and infectious pancreatic necrosis virus in water using UVC irradiation. Diseases of Aquatic Organisms 48: 1–
5.
Ozer, A; Ozkan, H; Guneydag, S; Yurakhno, V (2015a). First report of several myxosporean (Myxozoa) and
monogenean parasites from fish species off Sinop coasts of the Black Sea Turkish Journal of Fisheries and Aquatic
Sciences 15: 737– 744 [Online] Available from: http://www. doi.10.4194/1303-2712-v15_3_18 [Accessed 20
September 2019].
Ozer, A; Ozkan, H; Yurakhno, V (2015b). New Host and Geographical Records of Ortholinea orientalis
(Schulman and Schulman-Albova, 1953) (Myxozoa, Myxosporea), a Parasite of Marine Fishes. Acta Zoologica
Bulgarica 67(4): 595–597.
Ozturk, R C; Altinok I (2014). Bacterial and viral fish diseases in Turkey. Turkish Journal of Fisheries and
Aquatic Sciences 14: 275–297 [Online] Available from: http://www.doi. 10.4194/1303-2712-v14_1_30 [Accessed 8
May 2019].
Padros, F; Palenzuela, O; Hispano, C; Tosas, O; Zarza, C; Crespo, S; Alvarez-Pellitero, P (2001). Myxidium
leei (Myxozoa) infections in aquarium reared Mediterranean fish species. Diseases of Aquatic Organisms 47(1): 57–
62.
Pagowski, V A; Merdecai, G J; Miller, K M; Schulze, A D; Kaukinen, K H; Ming, T J; Li, S; Teffer, A K;
Tabata, A A; Suttle, C A (2019). Distribution and phylogeny of erythrocytic necrosis virus (ENV) in salmon
suggests marine origin. Viruses 11(4): 358 [Online] Available from: http://www/ doi.org/10.3390/v11040358
[Accessed 21 August 2019].
Panzarin, V; Patarnello, P; Mori, A; Rampazzo, E; Cappellozza, E; Bovo, G; Cattoli, G (2010). Development
and validation of a real-time TaqMan PCR assay for the detection of betanodavirus in clinical specimens. Archives
of Virology 155:1193 – 1203.
Panzarin, V; Fusaro, A; Monne, I; Cappellozza, E; Paternello, P; Bovo, G; Capua, I; Holmes, E C; Cattoli, G
(2012). Molecular epidemiology and evolutionary dynamics of betanodavirus in southern Europe. Infection,
Genetics and Evolution 12: 63–70 [Online] Available from http://www.doi.10.1016/j.meegid.2011.10.007
[Accessed 28 May 2019].
Pagowski, V A; Mordecai, G J; Miller, K M; Schulze, A D; Kaukinen, K H; Ming, T J; Li, S; Teffer, A K;
Tabata, A; Suttle, C A (2019). Distribution and phylogeny of erythrocytic necrosis virus (ENV) in salmon suggests
marine origin. Viruses 11: 358 [Online] Available from: http://www.doi.10.3390/v11040358 [Accessed 29 October
2019].
Palacios, G; Lovoll, M; Tengs, T; Hornig, M; Hutchison, S; Hui, J; Kongtorp, RT; Savji N; Bussetti AV;
Solovyov, A; Kristoffersen, A B; Celone, C; Street, C; Trifonov, V; Hirschberg, D L; Rabadan, R; Egholm,
M; Rimstad, E; Lipkin, W I (2010). Heart and skeletal muscle inflammation of farmed salmon is associated with
infection with a novel reovirus. PLoS ONE 5(7): e11487.doi.10.1371/journal.pone.0011487 [Online] Available
from: http://www.journals.plos.org/plosone/article?id=10.1371/journal.pone.0011487 [Accessed 22 April 2015].
Paperna, I (1998) Mariculture versus environment: risks and impacts. Biologia Marina Mediterranea 5(1):381–
389.
Paperna, I; Overstreet, R (1981). Parasites and Diseases of Mullets (Mugilidae). Faculty Publications from the
Harold W. Manter Laboratory of Parasitology 579: 411–493.
Parenzan, P (1966). Myxobolus mugilis e Myxobolus branchialis nuovi missosporidi parassiti di Mugil chelo dello
Jonio. Bulletin of the Society Natural Napoli 73: 3–8.
Park, S; Wakabayashi, H; Watanabe, Y (1983). Serotype and virulence of Edwardsiella tarda isolated from eel
and their environment. Fish Pathology 18: 85–89.
Park, S B; Takashi, A; Jung, T S (2012). Pathogenesis of and strategies for preventing Edwardsiella tarda
infection in fish. Veterinary Research 43(67), 11 p [Online] Available from:
http://www.veterinaryresearch.org/content/43/1/67 [Accessed 8 August 2015].
Paul, L J (2000). New Zealand Fishes. Reed Books, Second edition. Auckland, New Zealand, 253 p.
Paul, L J; Taylor, P R; Parkinson, D M (2001). Pilchard (Sardinops neopilchardus) biology and fisheries in New
Zealand, and a review of pilchard (Sardinops, Sardina) biology, fisheries, and research in the main world fisheries.
New Zealand Fisheries Assessment Report 2001/37, 45 p.

144 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Pearce, F M; Oidtmann, B C; Thrush, M A; Dixon, P F; Peeler, E J (2014). Do imports of rainbow trout
carcasses risk introducing viral haemorrhagic septicaemia virus into England and Wales? Transboundary and
Emerging Diseases 61: 247–257.
Pearson, A (2019). New Zealand Food Safety, Ministry for Primary Industries, Wellington, New Zealand. Personal
communication.
Peeler, E J; Murray, A G; Thebault, A; Brun, E; Giovaninni, A; Thrush, M A (2007). The application of risk
analysis in aquatic animal health management. Preventative Veterinary Medicine 81: 3–20 [Online] Available from:
http://www.doi.10.1016/j.prevetmed.2007.04.012 [Accessed 29 November 2019].
Peeler, E J; Taylor, N G H (2011). The application of epidemiology in aquatic animal health -opportunities and
challenges. Veterinary Research 42: 94 [Online] Available from:
https://veterinaryresearch.biomedcentral.com/articles/10.1186/1297-9716-42-94 [Accessed 29 November 2019].
Pegg, D E (1987). Mechanisms of freezing damage. Symposia of the Society for Experimental Biology 41: 363-78.
Pena, L D de la; Suamaba, V S; Capulos, G C; Santos, M N M (2011). Prevalence of viral nervous necrosis
(VNN) virus in wild-caught and trash fish in the Philippines. Bulletin of the European Association of Fish
Pathologists 31(4): 129–138.
Perera, R P; Johnson, S K; Collins, M D; Lewis, D H (1994). Streptococcus iniae associated with mortality of
Tilapia nilotica x T. aurea hybrids. Journal of Aquatic Animal Health 6: 335–340.
Pereira, U P; Mian, G F; Oliveira, I C M; Benchetrit, L C; Costa, G M; Figueiredoa, H C P (2010).
Genotyping of Streptococcus agalactiae strains isolated from fish, human and cattle and their virulence potential in
Nile tilapia. Veterinary Microbiology 140(1–2): 186–192 [Online] Available from:
https://doi.org/10.1016/j.vetmic.2009.07.025 [Accessed 6 May 2020].
Phelps, N B; Goodwin, A E; Marecaux, E; Goyal, S M (2013). Comparison of treatments to inactivate viral
hemorrhagic septicemia virus (VHSV-IVb) in frozen bait fish. Diseases Of Aquatic Organisms 102: 211–216
[Online] Available from: http://www.doi.10.3354/dao02549 [Accessed 12 June 2019].
Phelps, N B; Craft, M E; Travis, D; Pelican, K; Goyal, S M (2014). Risk-based management of viral
hemorrhagic septicemia virus in Minnesota. North American Journal of Fisheries Management 34(2): 373–379
[Online] Available from: http://www.doi.10.1080/02755947.2014.880766 [Accessed 3 September 2019].
Pierre, J P; Thompson, F N; Cleal, J (2014). Seabird interactions with the deepwater bottom-longline fleet:
Characterisation of smaller vessel deepwater bottom-longline operations in relation to risk factors for seabird
capture. Final Report for Project MIT2013-03. 33 p. [Online] Available from:
https://files.dragonfly.co.nz/publications/pdf/Pierreetal_2014_mit2013-03.pdf [Accessed 29 July 2020].
Pinkerton, M; Sutton, P; Wood, S (2019). Satellite indicators of phytoplankton and ocean surface temperature for
New Zealand. NIWA Client Report 2018180WN, prepared for the Ministry for the Environment by the National
Institute for Water and Atmosphere, Auckland, New Zealand [Online] Available from:
https://www.mfe.govt.nz/sites/default/files/media/Marine/satellite-indicators-of-phytoplankton-and-ocean-surface-
temperature.pdf [Accessed 21 October 2020].
Pirtle, E C; Beran, G W (1991). Virus survival in the environment. Scientific and Technical Review (OIE) 10(3):
733–748.
Pittman, B; Shaw, E B; Cherry, W B (1977). Isolation of Francisella tularensis from infected human blood.
Journal of Clinical Microbiology 5(6): 621–624.
Plant & Food, (2016). Sustainable snapper piped out to sea by Plant and Food [Online] Available from:
http://www.stuff.co.nz/business/farming/aquaculture/87522652/sustainable-snapper-piped-out-to-sea [Accessed 14
August 2017].
Plant & Food, (2017). Snapper aquaculture. Rural Delivery, June 2017. Plant and Food Research, Nelson, New
Zealand [Online] Available from: https://www.ruraldelivery.net.nz/stories/Snapper-Aquaculture [Accessed 15
August 2017].
Plumb, J A; Hansen, L A (2011). Health maintenance and principal microbial diseases of cultured fishes. John
Wiley, New York, United States of America, 400 p.
Plumb, J A; Schachte, J H; Gaines, J; Peltier, W; Carroll, B (1974). Streptococcus sp. from marine fishes along
the Alabama and northwest Florida coast of the Gulf of Mexico. Transactions of the American Fisheries Society
103: 358–361 [Online] Available from: https://www.doi.org/10.1577/1548-
8659(1974)103<358:SSFMFA>2.0.CO;2 [Accessed 5 September 2019].

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 145
Pogorel'tseva, T P (1964). Material on the study of protozoan parasites of fish of the Black Sea. Transactions of
Ukrainian Research Publications of Parasitological Observations (Tr. Ukrainsk. Respub, Nauch. Obsh. Parazit.) 3:
16–29 (in Russian).
Polglase, J L (2019). Cephalopod diseases caused by Fungi and Labyrinthulomycetes, pp 111–122, In: Gestal, C;
Pasual, S; Guerra, A; Fiorito, G; Veites, J M (Eds.) Handbook of pathogens and diseases in cephalopods. Springer
Science, New York, United States of America, 230 p.
Polglase, J L; Dix, N J; Bullock, A M (1984). Infection of skin wounds in the lesser octopus Eledone cirrhosa by
Cladosporium sphaerospermum. Transactions of the British Mycological Society 82: 577–580.
Poynton, S; Reimschuessel, R; Stoskopf, M K (2002). Aggregata dobelli n. sp. And Aggregata millerorum n. sp.
(Apicomplexa: Aggregatidae) from two species of Octopus (Mollusca: Octopodidae) from the Eastern North Pacific
Ocean. The Journal of Protozoology 39(1): 248–256.
Poynton, S L; Schultz, D; Trembly. S; Kelly. K; Montali, RJ; Hadfield, C A; Delannoy, M; Litaker, W (2009).
Host-parasite interaction, ultrastructure, and molecular taxonomy of a pathogenic Ichthyobodo Sp. (Kinetoplastida:
Ichthyobodonidae): Infection in captive giant pacific octopus, Octopus (Enteroctopus) dofleini. Proceedings of the
40th Conference of the International Association of Aquatic Animal Medicine (IAAAM), SeaWorld, San Antonio,
Texas, United States of America 2009) [Online] Available from:
https://www.vin.com/apputil/content/defaultadv1.aspx?meta=Generic&pId=11285 [Accessed 1 November 2019].
Prado-Alvarez, M; Garcia-Fernandez, P (2019). Virus and virus-like particles affecting cephalopods, pp 123–127
In: Gestal C, Pasual S, Guerra A, Fiorito G, Veites JM (Eds.) Handbook of pathogens and diseases in cephalopods.
Springer Science, New York, United States of America, 230 p.
Prieta, J; Domenech, A M; Fernandez-Garayzabal, J F; Collins, M D; Rodrigues, U M; Jones, D; Rodriguez,
A; Dominguez, L (1993). Lactococcosis de la trucha arco iris (Oncorhynchus mykiss). Medical Veterinology 10:
367–373 (cited in Muzquiz et al. 1999).
PROMED, (2017). Pilchard orthomyxovirus – Australia (Tasmania) Farmed salmon [Online] Available from:
http://www.promedmail.org/direct.php?id=20171221.5517876 [Accessed 9 July 2018].
Purcell, M K; Bromage, E S; Silva, J; Hansen, J D; Badil, S M; Woodson, J C; Hershberger, P K (2012).
Production and characterization of monoclonal antibodies to IgM of Pacific herring (Clupea pallasii). Fish and
Shellfish Immunology 33(3): 552–558 [Online] Available from: https://doi.org/10.1016/j.fsi.2012.06.006 [Accessed
27 May 2019].
Purcell, M K; Pearman-Gillman, S; Thompson, R L; Gregg, J L; Hart, L M; Winton, J R; Emmenegger, J;
Hershberger, P K (2016). Identification of the major capsid protein of erythrocytic necrosis virus (ENV) and
development of quantitative real-time PCR assays for quantification of ENV DNA. Journal of Veterinary
Diagnostic Investigation 28(4): 382–391.
Quaglio, F; Delgado, M L; Caffara, M; Florio, D; Marcer, F; Fioravanti, M L; Restani, R (2002).
Histopathological observations in marine farmed fish infected by Myxosporidia. (II Osservazioni istopatologiche in
pesci marini d’allevamento affetti da mixosporidiosi. II). Boll Soc Ital Patol Ittica (Italian Journal of Parasitology)
14(34): 44–67.
Quiazon, K M A (2015). Updates on aquatic parasites in fisheries: implications to food safety, food security and
environmental protection. Journal of Coastal Zone Management 18(1): 396, 7 p. [Online] Available from:
http://www.doi.10.4172/2473-3350.1000396 [Accessed 14 June 2019].
Racz, Z O; Zemankovics, E (2002). Survival of metacercariae of Metagonimus yokogawai (Digenea:
Heterophyidae) on fish from River Danube. Magyar Allatorvosok Lapja 124(7): 437–444 (Cited in Johnston 2008).
Rahimian, H (2007). Parasites of fingerling herring Clupea harengus L.: ecology and fine morphology. Journal of
Helminthology 81(2): 199–207 [Online] Available from: http://www.doi.10.1017/S0022149X07735381 [Accessed
22 October 2019].
Rajendram, P; Kyaw, W M; Leo, Y S; Ho, H; Chen, W K; Lin, R; Pratim, P; Badaruddin, H; Ang, B;
Barkham, T; Chow, A (2016). Group B Streptococcus sequence type 283 disease linked to consumption of raw
fish in Singapore. Emerging Infectious Diseases 22(11): 1974–1977 [Online] Available from:
http://www.dx.doi.org/10.3201/eid2211.160252 [Accessed 24 July 2018].
Rashid, M M; Honda, K; Nakai, T; Muroga, K (1994). An Ecological Study on Edwardsiella tarda in Flounder
Farms. Fish Pathology 29(4): 221–227.

146 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Raynard, R; Wahli, T; Vatsos, I; Mortensen, S (2007). Review of disease interactions and pathogen exchange
between farmed and wild finfish and shellfish in Europe, p. 459 In: Final Report for the DIPNET project (Disease
interactions and pathogen exchange between farmed and wild aquatic animal populations - a European network).
European Commission [Online] Available from: http://www.dipnet.info/docs/doc.asp?id=48 [Accessed 13 January
2020].
Reed, C; MacKenzie, K; van der Lingen, C D (2012). Parasites of South African sardines, Sardinops sagax, and
an assessment of their potential as biological tags. Bulletin of the European Association of Fish Pathologists 32(2):
41–48.
Reichley, S R; Ware, C; Steadman, J; Gaunt P S; García, J C; LaFrentz, B R; Thachil, A; Waldbieser, G C;
Stine, C B; Buján, N; Arias, C R; Loch, T; Welch, T J; Cipriano, R C; Greenway, T E; Khoo, L H; Wise, D J;
Lawrence, M L; Griffin, M J (2017). Comparative phenotypic and genotypic analysis of Edwardsiella isolates
from different hosts and geographic origins, with emphasis on isolates formerly classified as E. tarda, and
evaluation of diagnostic methods. Journal of Clinical Microbiology 55: 3466–3491 [Online] Available from:
https://www.doi.org/10 .1128/JCM.00970-17 [Accessed 3 September 2019].
Reimschuessel, R; Gieseker, C M; Driscoll, C; Baya, A; Kane, A S; Blaser, V S; Evans, J J; Kent, M L;
Moran, J D; Poynton, S L (2003). Myxosporean plasmodial infection associated with ulcerative lesions in young-
of-the-year Atlantic menhaden in a tributary of the Chesapeake Bay, and possible links to Kudoa clupeidae.
Diseases of Aquatic Organisms 53(2): 143–166.
Reimschuessel, R; Stoskopf, M K; Bennett, R O (1990). Myocarditis in the common cuttlefish (Sepia officinalis).
Journal of Comparative Pathology 102(3): 291–297.
Reno, P W; Serreze, D V; Hellyer, S K; Nicholson, B L (1985). Hematological and physiological effects of viral
erythrocytic necrosis (VEN) in Atlantic cod and herring. Fish Pathology 20(2-3): 353–360.
Rigos, G; Christophilogiannis, M; Yiagnisi, M; Andriopoulou, A; Koutsodimou, M; Negas, I; Alexis, M
(1999). Myxosporean infections in Greek mariculture. Aquaculture International 7: 361–364.
Rigos, G; Katharios, P (2010). Pathological obstacles of newly-introduced fish species in Mediterranean
mariculture: A review. Reviews in Fish Biology and Fisheries 20(1): 47–70 [Online] Available from:
http://ww.doi.10.1007/s11160-009-9120-7 [Accessed 1 November 2019].
Rimmer, A E; Becker, J A; Tweedie, A; Lintermans, M; Landos, M; Stephens, F; Whittington, R J (2015).
Detection of dwarf gourami iridovirus (Infectious spleen and kidney necrosis virus) in populations of ornamental
fish prior to and after importation into Australia, with the first evidence of infection in domestically farmed Platy
(Xiphophorus maculatus). Preventative Veterinary Medicine 122(1-2): 181–194 [Online] Available from:
http://www.doi: 10.1016/j.prevetmed.2015.09.008 [Accessed 20 February 2018].
Rimmer, A E; Whittington, R J; Tweedie, A; Becker, J A (2017). Susceptibility of a number of Australian
freshwater fishes to dwarf gourami iridovirus (Infectious spleen and kidney necrosis virus). Journal of Fish Diseases
40(3): 293–310 [Online] Available from: http://www.doi.10.1111/jfd.12510 [Accessed 17 December 2019].
Rimstad, E; Dale, O B; Dannevig, B H; Falk, K (2011. Infectious salmon anaemia, pp 143–165 In: Woo PTK,
Bruno DW (Eds.) Fish Diseases and Disorders, Volume 3, CAB International, Wallingford, United Kingdom, 800 p.
Rimstad, E; Mjaaland, S (2002). Infectious salmon anaemia virus: An orthomyxovirus causing an emerging
infection in Atlantic salmon. Journal of Pathology, Microbiology and Immunology (APMIS) 110(4): 273–282.
Rivas, C; Bandin. I; Noya, M; Cepeda, C; Barja, J L; Dopazo, C P (1996a). In vitro and in vivo replication of
turbot aquareovirus (TRV) in turbot tissues. Diseases of Aquatic Organisms 25: 217–223.
Rivas, C; Bandin, I; Noya, M; Dopazo, C P; Cepeda, C; Barja, J L (1996b). Effect of the turbot aquareovirus on
fish macrophages using an in vitro model. Diseases of Aquatic Organisms 25: 209–216.
Roberts, R G (2012). Fish Pathology. John Wiley, London, United Kingdom, 581 p.
Rocha, S; Azevedo, C; Oliveira, E; Alves, A; Antunes, A; Rodrigues, P; Casal, G (2019). Phylogeny and
comprehensive revision of mugiliform-infecting myxobolids (Myxozoa, Myxobolidae), with the morphological and
molecular redescription of the cryptic species Myxobolus exiguus. Parasitology 146(4): 479–496 [Online] Available
from: https://www.doi.org/10.1017/S0031182018001671 [Accessed 24 September 2019].
Rodgers, L J; Burke, J B (1981). Seasonal variation in the prevalence of ‘‘red spot’’ disease in estuarine fish with
particular reference to the sea mullet, Mugil cephalus L. Journal of Fish Diseases 4: 297–307.
Rohovec. J S; Amandi, A (1981). Incidence of viral erythrocytic necrosis among hatchery reared salmonids in
Oregon. Fish Pathology 15: 135–141.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 147
Romanenko, L; Uchino, M; Tebo, R M; Tanaka, N; Frolova, G M; Mikhailov, V V (2008). Pseudomonas
marincola sp. nov., isolated from marine sediments. International Journal of Systematic and Evolutionary
Microbiology 58: 706–710.
Rothwell, J T; Virgona, J L; Canninan, R B (1997). Occurrence of cutaneous infections of Myxobolus
episquamalis (Myxozoa: Myxobolidae) in sea mullet, Mugil cephalus L, in Australia. Australian Veterinary Journal
75(5): 349–352 [Online] Available from: https://www.doi.org/10.1111/j.1751-0813.1997.tb15709.x [Accessed 15
May 2019].
Rothwell, J T; Virgona, J L; Callinan, R B; Nicholls, P J; Langdon, J S (2008). Occurrence of cutaneous
infections of Myxobolus episquamalis (Myxozoa: Myxobolidae) in sea mullet, Mugil cephalus L, in Australia.
Australian Veterinary Journal 75(5): 349–352.
Ruane, N; Geoghegan, F; Cinneide, M O (1997). Infectious pancreatic necrosis virus and its impact on Irish
salmon aquaculture and wild fish sectors. Marine Environment and Health, Series 30, Marine Institute, Galway,
Ireland, 56 p.
Rucker, R R; Whipple, W J; Parvin, J R; Evans, C A (1953). A contagious disease of salmon possibly of virus
origin. Fishery Bulletin 76: 35–46.
Rungger, D; Rastelli, M; Braendle, E; Malsberg, R G (1971). A viruslike particle associated with lesions in
muscles of Octopus vulgaris. Journal of Invertebrate Pathology 17: 72–80. [Online] Available from:
https://www.doi.org/10.1016/0022-2011(71)90128-5 [Accessed 20 June 2019].
Rutherford, D (2019). Lyford & Burkhart Taspac Ltd, Auckland, New Zealand. Personal communication.
Sakamoto, T; Okinaka, Y; Mori, K I; Sugaya, T; Nishioka, T; Oka, M; Yamashita, H; Nakai, T (2008).
Phylogenetic analysis of betanodavirus RNA2 identified from wild marine fish in oceanic regions. Fish Pathology
43(1): 19–27 [Online] Available from: https://home.hiroshima-u.ac.jp/fishpath/okinaka/PDF/Sak%202008.pdf
[Accessed 30 June 2020].
Sakazaki, R; Tamura, K (1975). Priority of the specific epithet anguillimortiferum over the specific epithet tarda
in the name of the organism presently known as Edwardsiella tarda. International Journal of Systematic and
Evolutionary Microbiology 25: 219–220.
Sakr, S F M; El-Rhman, A M M (2008). Contribution on Pseudomonas septicaemia caused by Pseudomonas
anguilliseptica in cultured Oreochromis niloticus, pp 1177–1197. In: From the Pharos to the Future: Proceedings of
the Eighth International Symposium on Tilapia in Aquaculture, October 12–14 2008, Cairo, Egypt [Online].
Available from: http://www.ag.arizona.edu/azaqua/ista/ ISTA8/ProceedingsISTA8.htm [Accessed 3 October 2014].
Salama, N K G; Rabe, B (2013). Developing models for investigating the environmental transmission of disease-
causing agents within open-cage salmon aquaculture. Aquaculture Environment Interactions 4: 91–115 [Online]
Available from: http:/www.doi.10.3354/aei99977 [Accessed 2 April 2020].
Salati, F (2011). Enterococcus seriolicida and Streptococcus spp. (S. iniae, S. agalactiae and S. dysgalactiae), pp
375–396 In: Woo, P T K; Bruno, D W (Eds.) Fish diseases and disorders: Volume 3, Viral, bacterial and fungal
infections. 2nd Edition. CAB International, Wallingford, United Kingdom, 930 p.
Samuelsen, O B; Nerland, A H; Jorgensen, T; Schroder, M B; Svasand, T; Bergh, O (2006). Viral and bacterial
diseases of Atlantic cod Gadus morhua, their prophylaxis and treatment: a review. Diseases of Aquatic Organisms
71: 239–254.
Sánchez-García, N; Raga, J A; Montero, F E (2014). Risk assessment for parasites in cultures of Diplodus
puntazzo (Sparidae) in the Western Mediterranean: prospects of cross infection with Sparus aurata. Veterinary
Parasitology 204(3-4):120–133.
Sandlund, N; Gjerset, B; Bergh, O; Modahi, I; Olesen, N J; Johansen, R (2014). Screening for viral
hemorrhagic septicaemia virus in marine fish along the Norwegian coastal line. PLoS One 9(9): e108529 [Online]
Available from http://www. doi.10.1371/journal.pone.0108529 [Accessed 8 May 2019].
Sano, M; Nakai, T; Fijan, N (2011) Viral diseases and agents of warm water fish, pp 168–244 In: Woo, P T K;
Bruno, D W (Eds.). Fish Diseases and Disorders: Volume 3 - Viral, Bacterial and Fungal Infections, 2nd edition.
CAB International, Wallingford, United Kingdom, 930 p.
Santos, Y; Pazos, F; Barja, J L (1999). Flexibacter maritimus, causal agent of flexibacteriosis in marine fish, pp.
1–6 In: Oliver, G. (Ed.). ICES Identification leaflet for diseases and parasites of fish and shellfish. International
Council for the Exploration of the Sea, Copenhagen, Denmark.

148 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Sardella, N H (1988). Descripciôn de la espora y de los estadios previos del mixosporidio Kudoa rosenbuschi,
parâsito muscular de Merluccius hubbsi. Ciclo de vida. (Description and lifecycle of the spore and previous stages
of myxosporidium Kudoa rosenbuschi, muscle parasite of Merluccius hubbsi). Parasitologia al Dia 12: 8–13 (In
Spanish).
Sardella, N H; Re, M E; Timi, J T (2000). Two new Aggregata species (Apicomplexa: Aggregatidae) infecting
Octopus tehuelchus and Enteroctopus megalocyathus (Mollusca: Octopodidae) in Patagonia, Argentina. Journal of
Parasitology 86: 1107–1113.
Sardella, N H; Timi, J T (1996). Parasite communities of Merluccius hubbsi from the Argentinean-Uruguayan
common fishing zone. Fisheries Research 27: 81–88.
Sarkar, N M (2010). Myxodavisia haldarae sp. n. and Ceratomyxa sardinellae sp. n. (Myxosporea, Bivalvulida)
from the gallbladder of Indian Sardine Sardinella longiceps collected in the Bay of Bengal. Protistology 6(4): 290–
296.
Sarkar, N; Ghosh, S (1991). Two new coelozoic Myxosporidia (Myxozoa: Myxosporea) from estuarine teleost
fishes (Mugilidae) of West Bengal, India. Proceedings of the Zoological Society of Calcutta 44: 131–135.
Sato, H (1984). Streptococcan infections in cultured yellowtail. NOAA Technical Report 111: 125–139. U.S.
Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service,
Silver Springs, Maryland, United States of America.
SCAAH, (2015). Sub-committee on Aquatic Animal Health SCAAH Newsletter No. 5, June 2015 [Online]
Available from: http://www.apfa.com.au/wp=content/uploads/2015/01/SCAAH-Newsletter-2015-7-July-2015.pdf
[Accessed 9 July 2018].
Sea Containers, (2020). Refrigerated shipping containers [Online] Available from:
https://www.seacontainers.co.nz/container-sales/refrigerated-shipping-containers/?gclid=EAIaIQobChMIkJmhr-
v96wIV16mWCh1LwwZQEAAYAyAAEgJO3vD_BwE [Accessed 23 Septemer 2020].
Seafood New Zealand, (2019). Economic review to December 2018. Seafood New Zealand, Nelson, New Zealand
[Online] Available from:
https://www.seafoodnewzealand.org.nz/fileadmin/documents/Economic_reviews/Economic_review_to_
December_2018.pdf [Accessed 29 November 2019].
Sekar, V T; Santiago, T C; Vijayan, K K; Alavandi, S V; Raj, V S; Rajan, J J S; Sanjuktha, M; Kalaimani, N
(2008). Involvement of Enterobacter cloacae in the mortality of the fish, Mugil cephalus. Letters in Applied
Microbiology 46: 667-672
Shao, S; Lai, Q; Liu, Q; Wu, H; Xiao, J; Shao, Z; Wang, Q; Zhang, Y (2015). Phylogenomics characterization
of a highly virulent Edwardsiella strain ET080813T encoding two distinct T3SS and three T6SS gene clusters:
propose a novel species as Edwardsiella anguillarum sp. nov. Systematic and Applied Microbiology 38: 36–47
[Online] Available from: http://www.doi.10.1016/j.syapm.2014.10.008 [Accessed 1 November 2019].
Sharon, G; Ucko, M; Tamir, B; Diamant, A (2019). Co-existence of Myxobolus spp. (Myxozoa) in grey mullet
(Mugil cephalus) juveniles from the Mediterranean Sea. Parasitology Research 118: 157–167 [Online] Available
from: https://www.doi.org/10.1007/s0043 [Accessed 13 September 2019].
Shaw, R W; Kent, M L; Adamson, M L (2000). Viability of Loma salmonae (Microsporidia) under laboratory
conditions. Parasitology Research 86(12): 978–981.
Sherrill, J; Spelman, L H; Reidel, C L; Montali, R L (2000). Common cuttlefish (Sepia officinalis) mortality at
the national zoological park: implications for clinical management. Journal of Zoo and Wildlife Medicine 31: 523–
531.
Shetty, M; Maiti, B; Santhosh, K S; Venugopal, M N; Karunasagar, I (2012). Betanodavirus of marine and
freshwater fish: Distribution, genomic organisation, diagnosis and control measures. Indian Journal of Virology
23(2): 114–123.
Shin, S P; Gomez, D K; Kim, J H; Choresca, C H; Han, J E; Woo, J; Park, S E (2011). Detection and genetic
analysis of aquabirnaviruses in subclinically infected aquarium fish. Journal of Veterinary Diagnostics and
Investigation 23: 325–329.
Shin, S P; Sohn, H C; Jin, C N; Kang, B J; Lee, J (2018). Quantitative investigation of Enteromyxum
leei (Myxozoa: Myxosporea) infection and relative condition factor in cultured olive flounder Paralichthys
olivaceus (Temminck and Schlegel). Journal of Fish Diseases 42(2): 159–165 [Online] Available from:
https://doi.org/10.1111/jfd.12912 [Accessed 6 May 2020].

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 149
Shinmoto, H; Taniguchi, K; Ikawa, T; Kawai, K; Oshima, S (2009). Phenotypic diversity of infectious Red Sea
bream iridovirus isolates from cultured fish in Japan. Applied and Environmental Microbiology 75(11): 3535–3541
[Online] Available from: http://www.doi.10.1128/AEM.02255-08 [Accessed 16 December 2019].
Shoemaker, C A; Evans, J J; Klesius, P H (2000). Density and dose: factors affecting mortality of Streptococcus
iniae infected tilapia (Oreochromis niloticus). Aquaculture 188: 229–235.
Shoemaker, C A; Klesuis, P H; Evans, J J (2001). Prevalence of Streptococcus iniae in tilapia, hybrid striped
bass, and channel catfish on commercial fish farms in the United States. American Journal of Veterinary Research
62(2): 174–177 [Online] Available from: http://www. doi.10.2460/ajvr.2001.62.174 [Accessed 17 December 2019].
Shui, J Y; Hong, J R; Ku, C C; Wen, C M (2018). Complete genome sequence and phylogenetic analysis of
megalocytivirus RSIV-Ku: A natural recombination infectious spleen and kidney necrosis virus. Archives of
Virology 163(4): 1037–1042 [Online] Available from: https://doi.org/10.1007/s00705-017-3689-2 [Accessed 24
October 2019].
Shukgalter, O (2009). Unknown disease Of European pilchard (Sardina pilchardus Walbaum, 1792) from the
Atlantic coast of North Africa, In: Bruckner, A W; Cipriano, R C; Shchelkunov, I S (Comps.), Bridging America
and Russia with shared perspectives on aquatic animal health. Proceedings of the third bilateral conference between
Russia and the United States, July 2009. Shepherdstown, West Virginia, United States of America, 360 p.
Shulman, S S (1966). Myxosporidia of the USSR. Izd. Nauka, Akad. Sci. USSR, Moskva-Leningrad, 504 pp. (In
Russian.), English translation Balkema AA (Ed.) (1990), Rotterdam: 631 pp. (Cited in Lom 2004).
Shutou, K; Kanai, K; Yoshikoshi, K (2007). Virulence attenuation of capsular polysaccharide-deleted mutants of
Streptococcus iniae in Japanese flounder Paralichthys olivaceus. Fish Pathology 42: 41–48.
Simeca, J M; Oestmann, D (1995). Selected diseases of captive and laboratory reared cephalopods, p 79 In:
Proceedings of the International Association for Aquatic Animal Medicine 26: 79.
Sim-Smith, C; Faire, S; Lees, A (2014). Managing biosecurity risk for business benefit. Report prepared for the
Ministry for Primary Industries by Coast and Catchment Ltd, Auckland, New Zealand, 214 p.
Sindermann, C J (1987). Effects of parasites on fish populations: Practical considerations. International Journal
for Parasitology 17(2): 371–382 [Online] Available from: https://doi.org/10.1016/0020-7519(87)90112-3 [Accessed
4 April 2020].
Sitja-Bobadilla, A; Alvarez-Pellitero, P (1995). Light and electron microscopic description of Polysporoplasma n.
g. (Myxosporea: Bivalvulida), Polysporoplasma sparis n. sp. from Sparus aurata (L.), and Polysporoplasma mugilis
n. sp. from Liza aurata L. European Journal of Protistology 31: 77–89 [Online] Available from:
http://www.doi.10.1016/S0932-4739(11)80360-3 [Accessed 12 September 2019].
Skall, H F; Olesen, N J; Mellergaard, S (2005a), Prevalence of viral haemorrhagic septicaemia virus in Danish
marine fishes and its occurrence in new host species. Diseases of Aquatic Organisms 66: 145–151.
Skall, H F; Olesen, N J; Mellergaard, S (2005b). Viral haemorrhagic septicaemia virus in marine fish and its
implications for fish farming – a review. Journal of Fish Diseases 28: 509–529.
Skjesol, A; Skjæveland, I; Elnæs, M; Timmerhaus, G; Fredriksen, B N; Jørgensen, S M; Krasnov, A;
Jørgensen, J B (2011). IPNV with high and low virulence: host immune responses and viral mutations during
infection. Journal of Virology 8: 396. [Online] Available from: http://www.doi.10.1186/1743-422X-8-396
[Accessed 26 July 2017].
Skud, B E (1978). Bait loss and competition. In: Factors affecting longline catch and effort. Scientific Report 64, 66
p. International Pacific Halibut Commission, Seattle, Washington, United States of America [Online] Available
from: http://docs.streamnetlibrary.org/IPHC/SciReport0064.pdf [Accessed 29 July 2020].
Smail, D A (1982). Viral erythrocytic necrosis in fish: a review. Proceedings of the Royal Society of Edinburgh,
Section B: Biological Sciences 81(3): 169–176.
Smith, N W McL (2001). Longline sink rates of an autoline vessel, and notes on seabird interactions. Science for
Conservation 183, Department of Conservation, Wellington, New Zealand, 32 p [Online] Available from:
https://www.doc.govt.nz/globalassets/documents/science-and-technical/sfc183.pdf [Accessed 29 July 2020].
Smith, P J; Holdsworth, J; Anderson, C; Hine, P M; Allen, D; Gibbs, W; McKenzie, L; Taylor, P R;
Blackwell, R G; Williamson, S H (1996). Pilchard deaths in New Zealand, 1995. New Zealand Fisheries Data
Report No. 70. 52 p.
Smith, G; Bebak, J; McAllister, P E (2000). Experimental infectious pancreatic necrosis infections: propagative or
point-source epidemics. Journal of Preventative Veterinary Medicine 47: 221–241.

150 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Snow, M; Bain, N; Black, J; Taupin, V; Cunningham, C O; King, J A; Skall, H F; Raynard, R S (2004).
Genetic population structure of marine viral haemorrhagic septicaemia virus (VHSV). Diseases of Aquatic
Organisms 61: 11–21.
Song, J Y; Kitamura, S I; Jung, S J; Miyadai, T; Tanaka, S; Fukuda, Y; Kim, S R; Oh, M J (2008). Genetic
variation and geographic distribution of megalocytiviruses. The Journal of Microbiology 46: 29–33.
Sorimachi, M; Hara. T (1985). Characteristics and pathogenicity of a virus isolated from yellowtail fingerlings
showing ascites. Fish Pathology 19: 231–238.
Souidenne, D; Florent, I; Dellinger, M; Romdhane, M S; Grellier, P; Furuya, H (2016). Redescription of
Dicyemennea eledones (Wagener, 1857) (Phylum Dicyemida) from Eledone cirrhosa (Lamarck, 1798) (Mollusca:
Cephalopoda: Octopoda). Systematic Parasitology 93: 905–915 [Online] Available from:
http://www.doi.10.1007/s11230-016-9659-3 [Accessed 31 May 2019].
Souidenne, D; Furuya, H (2019). Protist (Ciliates) and related diseases, pp 153–158 In: Gestal, C; Pasual, S;
Guerra, A; Fioriti, G; Vietes, J (Eds.). Handbook of pathogens and diseases in cephalopods, Springer Science, New
York, United States of America, 230 p.
Sourek, J (1974). Long-term preservation by freeze-drying of pathogenic bacteria of the Czechslovak National
Collection of Type Cultures. International Journal of Systematic Bacteriology 24(3): 358–365.
Sousa. J De; Eiras, J C; Ranzani-Paiva, M J T; Alexandrino, A C (1999). Bacteriology of wild grey mullets,
Mugil platanus Günther, from Cananéia, São Paulo State, Brazil. Revista Brasileira de Zoologia 16(4): 1065–1069
[Online] Available from: http://www.dx.doi.org/10.1590/S0101-81751999000400015 [Accessed 21 October 2019].
Southwick, R; Holdsworth, J C; Rea, T; Bragg, L; Allen, T (2018). Estimating marine recreational fishing’s
economic contributions in New Zealand. Fisheries Research 208: 116–123 [Online] Available from:
https://doi.org/10.1016/j.fishres.2018.06.016 [Accessed 30 January 2020].
Spencer, A (2019). Southern Baits Ltd, Auckland, New Zealand. Personal communication.
Spikler, A R (2010). Emerging and infectious diseases of animals. Iowa State University Press, Ames, Iowa, United
States of America, 383 p.
Sriwanayos, P; Francis-Floyd, R; Stidworthy, M F; Petty, B D; Kelly, K; Waltzek, T B (2013).
Megalocytivirus infection in orbiculate batfish Platax orbicularis. Diseases of Aquatic Organisms 105: 1–8.
Stehr, C; Whittaker. D J (1986). Host-parasite interaction of the myxosporeans Kudoa paniformis Kabata &
Whittaker, 1981 and Kudoa thyrsites (Gilchrist, 1924) in the muscle of Pacific whiting, Merluccius productus
(Ayres): an ultrastructural study. Journal of Fish Diseases 9: 505–517.
Stevens, R B (1960) Cultural practices in disease control, pp 357–429 In: Horsfall JG, Dimond AE (Eds.) Plant
pathology, an advanced treatise, Vol 3. Academic Press, New York, NY, United States of America.
Stevenson, A B; Roberston, D A (1976). The New Zealand Species of Trachurus (Pisces: Carangidae). Journal of
the Royal Society of New Zealand 7(2): 243–253 [Online] Available from:
http://www.doi.10.1080/03036758.1977.10427162 [Accessed 9 January 2020].
Stilwell, J; Yanong, R P E (2018). Myxosporidiosis (Myxozoan Infections) in Warmwater Fish. Fish parasites.
Institute of Food and Agricultural Sciences (IFAS) Extension, University of Florida, Gainesville, Florida, United
States of America [Online] Available from: https://edis.ifas.ufl.edu/fa201 [Accessed 8 May 2020].
Stone, M A B; MacDiarmid, S C; Pharo, H J (1997). Import health risk analysis: salmonids for human consumption.
Ministry of Agriculture Regulatory Authority, Wellington, New Zealand, 269 p.
Stoskopf, M K; Nevy, S; Flynn, S (1987). Treatment of ulcerative mantle disease due to Pseudomonas spp. in
Octopus dofleini and Octopus bimaculoides with oxytetracycline. Proceedings of the Annual meeting of the
International Association of Aquatic Animal Medicine (IAAM), Monterey, California, United States of America.
Stride, M C; Nowak, B F (2013). Epitheliocystis in three wild fish species in Tasmanian waters. Journal of Fish
Diseases 37(2): 157–162 [Online] Available from: https://doi.org/10.1111/jfd.12124 [Accessed 27 May 2019].
Susanna, K; Lau, P; Woo, P C Y; Tse, H; Leung, K W; Wong, S S Y; Yuen K Y (2003). Invasive Streptococcus
iniae infections outside North America. Journal of Clinical Microbiology 41(3): 1004–1009 [Online] Available
from: http://www.doi.10.1128/JCM.41.3.1004-1009.2003 [Accessed 16 July 2020].
Suttle, C A (2007). Marine viruses - major players in the global ecosystem. Reviews in Nature: Microbiology 5:
807–812.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 151
Suzuki, J; Murata, R; Yokoyama, H; Sadamasu, K; Kai, A (2015). Detection rate of diarrhoea-causing Kudoa
hexapunctata in Pacific bluefin tuna Thunnus orientalis from Japanese waters. International Journal of Food
Microbiology 194: 1–6.
Symonds, J E; Walker, S P; Pether, S; Gublin, Y; McQueen, D; King, A; Irvine, G W; Setiawan, A N;
Forsythe, J A; Bruce, M (2014). Developing yellowtail kingfish (Seriola lalandi) and hapuku (Polyprion
oxygeneios) for New Zealand aquaculture. New Zealand Journal of Marine and Freshwater Research 48(3): 371–
384.
Tanaka, S; Kuruyama, I; Nakai, T; Miyazaki, T (2003). Susceptibility of cultured juveniles of several marine
fish to the sevenband grouper nervous necrosis virus. Journal of Fish Diseases 26(2): 109–115.
Tang, L; Kang, H; Duan, K; Guo, M; Lian, G; Wu, Y; Li, Y; Gao, S; Jiang, Y; Yin, J; Liu, M (2016). Effects
of three types of inactivation agents on the antibody response and immune protection of inactivated IHNV vaccine
in rainbow trout. Viral Immunology 29(7): 430–435 [Online] Available from:
http://www.doi.10.1089/vim.2016.0035 [Accessed 6 August 2019].
Tanriul, T T; Çağirgan, H (2001). Doğadaki kefal balıklarında görülen pasteurellosis salgını (A Natural Outbreak
of Pasteurellosis in Mugil spp.). EU Su Ürünleri Dergisi (EU Journal of Fisheries & Aquatic Sciences) 18 (3–4):
509–512 [Online] Available from: http://www.egejfas.org/en/download/article-file/58137 [Accessed 20 January
2020] (referenced in Ozturk & Altinok 2014).
Tavares, G C; Carvalho, A F; Pereira, F L; Rezende, C P; Azevedo, V A C; Leal; Figuerido, H C P (2018).
Transcriptome and Proteome of Fish-Pathogenic Streptococcus agalactiae Are Modulated by Temperature.
Frontiers in Microbiology [Online] Available from: https://doi.org/10.3389/fmicb.2018.02639 [Accessed 22
September 2020].
Tian, J; Lu, J; Zhang, Y; Cheng-Li, J; Chen-Sun, L; Li-Hu, Z (2014). Microbial community structures and
dynamics in the 03/BAC drinking water treatment process. International Journal of Environmental Research and
Public Health 11(6): 6281–6290.
Timi, J T; Sardella, N H (1998). Myxosporeans and coccidians parasitic on engraulid fishes from the coasts of
Argentina and Uruguay. Parasite 5(4): 331–339 [Online] Available from:
http://dx.doi.org/10.1051/parasite/1998054331 [Accessed 17 September 2019].
Tindall, B Y (2016). On the synonymy of Edwardsiella tarda Ewing and McWhorter 1965 (Approved Lists 1980)
and Edwardsiella anguillimortifera (Hoshina 1962) Sakazaki and Tamura 1975 (Approved Lists 1980), an old
problem in need of a solution. International Journal of Systematic and Evolutionary Microbiology 66(11): 4910–
4912 [Online] Available from: http://www.doi.10.1099/ijsem.0.001434 [Accessed 29 January 2020].
Tisdall, D J; Phipps, J C (1987). Isolation and characterisation of s marine birnavirus from returning Chinook
salmon (Oncorhynchus tshawytscha) in the South Island of New Zealand. New Zealand Veterinary Journal 35: 217–
218.
Toranzo, A; Barja, J; Lemos, M; Hetrick, F M (1983). Stability of infectious pancreatic necrosis virus (IPNV) in
untreated, filtered and autoclaved estuarine water. Bulletin of the European Association of Fish Pathologists 3: 51–
53.
Toranzo, A E; Romalde, J L; Margarinos, B; Barja, J L (2009). Present and future of aquaculture vaccines
against fish bacterial diseases, pp 155–176 In: Rogers, C (Ed.). The use of veterinary drugs and vaccines in
Mediterranean Aquaculture, Zaragosa CIHEAM, Options Mediterraneennes; Serie A, Seminaires Mediterraneens,
86 p [Online] Available from: http://om.ciheam.org/article.php?IDPDF=801069 [Accessed 14 January 2020].
Traxler, G S; Kieser, D (1994). Isolation of the North American strain of viral hemorrhagic septicaemia virus
(VHSV) from herring (Clupea harengus pallasi) in British Columbia. Fish Health Section: American Fish Society
Newsletter 22: 8.
Traxler, G S; Kieser, D; Richard, J (1999). Mass mortality of pilchard and herring associated with viral
haemorrhagic septicaemia virus in British Columbia, Canada. American Fisheries Society Fish Health Section
Newsletter 27: 3–4.
Traxler, G S; Roome, J R; Kent, M L (1993). Transmission of infectious haematopoietic necrosis virus in
seawater. Diseases of Aquatic Organisms 16: 111–114.
Tu, K-C; Spendlove, R S; Goede, R W (1975). Effect of temperature on survival and growth of infectious
pancreatic necrosis virus. Infection and Immunity 11: 1409–1412.

152 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Tubbs, L; Lee, P; Diggles, B; Jones, J B; Sheppard, M; Sim-Smith, C (2007). A review of aquatic diseases of
significance to New Zealand. Final research report for project ZBS 2005-17. National Institute of Water and
Atmospheric Research, Wellington, New Zealand, 452 p.
Ucko, M; Colorni, A; Diamant, A (2004). Nodavirus infection in Israel mariculture. Journal of Fish Diseases 27:
459–469.
Urquhart, K; Bowden, T J; Buckett, B E; Fryer, R J; Ellis, A E (2009). Experimental study of the susceptibility
of Atlantic cod, Gadus morhua L, to infection with an IPNV strain pathogenic for Atlantic salmon Salmo salar L.
Journal of Fish Diseases 32: 447–456.
Urquhart, K; Murray, A C; Gregory, A; O’Dea, M; Munro, L A; Smail, D A; Shanks, A M; Raynard, R S
(2008). Estimation of infectious dose and viral shedding rates for infectious pancreatic necrosis virus in Atlantic
salmon. Salmo salar L, post-smolts. Journal of Fish Diseases 31: 879–887.
USDA, (2019). Freezing and food safety, Food Safety and Inspection Service, United States Department of
Agriculture, Washington D.C. United States of America [Online] Available from:
https://www.fsis.usda.gov/wps/portal/fsis/topics/food-safety-education/get-answers/food-safety-fact-sheets/safe-
food-handling/freezing-and-food-safety/CT_Index [Accessed 7 October 2019].
Van Beurden, S J; Engelsma, M Y; Roozenburg, I; Voorbergen-Laarman, M A; van Tulden, P W; Kerkhoff,
S; van Nieuwstadt, A P; Davidse, A; Haenen, O L M (2012). Viral diseases of wild and farmed European eel
Anguilla anguilla with particular reference to the Netherlands. Diseases of Aquatic Organisms 101: 69–86 [Online]
Available from: http://www.doi.10.3354/dao02501 [Accessed 21 January 2020].
Vandepitte, J; Van Damme, L; Desmyter, F Y (1980). Edwardsiella tarda and Plesiomonas shigelloides. Their
role as diarrhea agents and their epidemiology. Bulletin de la Société de pathologie Exotique Filiales 73(2): 139–
149 (In French, with English abstract).
Varello, K; Prearo, M; Serracca, L; Meloni, D; Rossini, I; Righetti, M; Pezzolato, M; Fioravanti, M L;
Ercolini, C; Bozzetta E (2014). Granulomatous lesions in a wild mullet population from the eastern Ligurian Sea
(Italy): mycobacteriosis vs. pseudotuberculosis. Journal of Fish Diseases 37: 553–558.
Varpe, Ø; Fiksen, Ø; Slotte, A (2005). Meta-ecosystems and biological energy transport from ocean to coast: The
ecological importance of herring migration. Oecologia 146: 443 [Online] Available from: http://www.doi.
10.1007/s00442-005-0219-9 [Accessed 17 December 2019].
Vennerström, P (2015). Pseudomoniasis (P. anguilliseptica) in farmed fish. ICES Identification Leaflet for
Diseases and Parasites of Fish and Shellfish, No. 63. International Council for the Exploration of the Sea (ICES),
Copenhagen, Denmark, 4 pp.
Vidal, L; Iannacone, J; Whipps, C M; Luque, J L (2017). Synopsis of the species of Myxozoa GRASSÉ, 1970
(Cnidaria: Myxosporea) in the Americas. Neotropical Helminthology 11(2): 413–511.
Vijapoopathy, S; Atyyaru, A; Kuzhanthaivel, R; Syed, A (2016). Lymphocystis in Indian oil sardine, Sardinella
longiceps (Valenciennes 1847). Asian Pacific Journal of Tropical Disease 6(8): 611–614 [Online] Available from:
https://doi.org/10.1016/S2222-1808(16)61095-2 [Accessed 16 December 2019].
Volpe, E; Grodzki, M; Panzarin, V; Guercio, A; Purpari, G; Serratore, P; Ciulli, S (2018). Detection and
molecular characterization of betanodaviruses retrieved from bivalve molluscs. Journal of Fish Diseases 41:
603−611.
Walker, R; Sherburne, S W (1977). Piscine erythrocytic necrosis virus in Atlantic cod Gadus morhua, and other
fish: ultrastructure and distribution. Journal of the Fisheries Research Board of Canada 34: 1188–1195.
Wallace, I S; Gregory, A; Murray, A G; Munro, E S; Raynard, RS (2008). Distribution of infectious pancreatic
necrosis virus (IPNV) in wild marine fish from Scottish waters with respect to clinically infected aquaculture sites
producing Atlantic salmon, Salmo salar L. Journal of Fish Diseases 31(3): 177–186 [Online] Available from: http://
https://www.doi.org/10.1111/j.1365-2761.2007.00886.x [Accessed 27 May 2019].
Wang, C S; Wen, C M (2016). Megalocytiviruses, pp 825–836 In: Liu, D (Ed.) Molecular Detection of Animal
Viral Pathogens, CRC Press, Taylor & France, New York, United States of America, 72 p.
Wang, C Y C; Shie, H S; Chen, S C; Huang, J P; Hseih, I C; Wen, M S; Lin, F C; Wu, D (2007). Lactococcus
garvieae infections in humans: possible association with aquaculture outbreaks. The International Journal of
Clinical Practice 61(1): 68–73 [online] Available from: https://www.doi.org/10.1111/j.1742-1241.2006.00855.x
[Accessed 9 May 2019].

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 153
Wang, H; Lee, L; Chang, H Y; Wang, C T; Wu, C H; Tasi, K H (2001). A survey of the diseases in cultured
brood grey mullet (Mugil cephalus Linnaeus) in the Yun-Chia-Nan area of Chinese Taipei. Journal of the Chinese
Society of Veterinary Science 27: 89–93.
Wang, Y Q; Lu, L; Weng, S P; Huang, J N; Chan, S M; He, J G (2007). Molecular epidemiology and
phylogenetic analysis of a marine fish infectious spleen and kidney necrosis virus-like (ISKNV-like) virus. Archives
of Virology 152: 763–773.
Ward, P; Myers, R A (2007). Bait loss and its potential effects on fishing power in pelagic longline fisheries.
Fisheries Research 86(1): 69–76 [Online] Available from: https://doi.org/10.1016/j.fishres.2007.05.002 [Accessed
29 July 2020].
Ward, T M; Hoedt, F; McLeay, L; Dimmlich, W F; Kinloch, M; McGarvey, R; Jackson, G; Rogers, P J;
Jones, K (2001). Effects of the 1995 and 1998 mass mortalities on the spawning biomass of Sardinops sagax in
South Australia. ICES Journal of Marine Science 58: 865-875.
Watanabe, K; Karlsen, M; Devold, M; Isdal, E; Litlabo, A; Nylund, A (2006). Virus-like particles associated
with heart and skeletal muscle inflammation (HSMI). Diseases of Aquatic Organisms 70: 183–192.
Watanabe, K; Yoshimizu, M (1998). Disinfection of equipment for aquaculture and fertilized eggs by ozonated sea
water. Fish Pathology 33: 145–146.
Webb, S C (1990). Pap pilchards and protozoa. South African Shipping News and Fishing Industry Review 45: 35.
Weinstein, M R; Litt, M; Kertesz, D A; Wyper, P; Rose, D; Coulter, M; McGeer, A; Facklam, R; Ostach, C;
Willey, B M; Borczyk, A; Low, D E (1997). Invasive infections due to a fish pathogen, Streptococcus iniae. New
England Journal of Medicine 337: 589–594.
Wellings, F M; Lewis, A L; Mountain, C W (1976). Demonstration of solids-associated virus in wastewater and
sludge. Applied and Environmental Biology 31: 354–358.
Wessel, O; Braanen, S; Alarcon, M; Hartveit, H; Roos, M; Markussen, M; Tengs, T; Dahle, M K; Rimstad, E
(2017). Infection with purified piscine orthomyxovirus demonstrates a causal relationship with heart and skeletal
muscle infection in Atlantic salmon. PLoS One 12(8): e0183781 16 p. [Online] Available from:
http://www.doi.10.5281/zenodo.343744 [Accessed 13 January 2020].
Wheeler, M (2012). Perth Metro lakes surveyed for introduced fish. Fisheries and Water, Science Network WA,
Australia, Government of Western Australia [Online] Available from: http://sciencewa.net.au/topics/fisheries-a-
water/item/2764-perth-metro-lakes-surveyed-for-introduced-fish [Accessed 3 October 2014].
Whipps, C M; Adlard, R D; Bryant, M S; Lester, R J G; Findlay, V; Kent, M L (2003). First report of three
Kudoa species from eastern Australia: Kudoa thyrsites from Mahi mahi (Coryphaena hippurus), Kudoa amamiensis
and Kudoa minithyrsites n. sp. from Sweeper (Pempheris ypsilychnus). The Journal of Eukaryotic Microbiology
50(3): 215–219 [Online] Available from: https://www.doi.org/10.1111/j.1550-7408.2003.tb00120.x [Accessed 14
June 2019].
Whipps, C M; Kent, M L (2006). Phylogeography of the cosmopolitan marine parasite Kudoa thyrsites (Myxozoa:
Myxosporea). Journal of Eukaryotic Microbiology 53(5): 364–373 [Online] Available from:
https://doi.org/10.1111/j.1550-7408.2006.00114.x [Accessed 16 September 2019].
Whitfield, A K; Panfili, J; Durand, J D (2012). A global review of the cosmopolitan flathead mullet Mugil
cephalus Linnaeus 1758 (Teleostei: Mugilidae), with emphasis on the biology, genetics, ecology and fisheries
aspects of this apparent species complex. Reviews in Fish Biology and Fisheries 22(3): 641–681 [Online] Available
from: http://www.doi.10.1007/s11160-012-9263-9 [Accessed 6 January 2019].
Whittington, R J; Crockford, M; Jordan, D; Jones, B (2008). Herpesvirus that caused epizootic mortality in
1995 and 1998 in pilchard, Sardinops sagax neopilchardus (Steindachner), in Australia is now endemic. Journal of
fish Diseases 31(2): 97–105.
Whittington, R J; Becker, J A; Dennis, M M (2010). Iridovirus infections in finfish – critical review with
emphasis on ranaviruses. Journal of Fish Diseases 33: 95–122.
Wiedenmayer, A A; Evans, J J; Klesius, P H (2006). Experimental Edwardsiella tarda infection in non-abraded
channel catfish Ameiurus punctatus by immersion. Fisheries Science 72(5): 1124–1126.
Wiens, G D (2011). Bacterial kidney disease, pp 338–374 In: Woo, P T K; Bruno, D W (Eds.) Fish diseases and
disorders: Volume 3, Viral, bacterial and fungal infections. 2nd. Edition. CAB International, Wallingford, United
Kingdom, 930 p.

154 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Wiik-Nielsen, C R; Lovoll, M; Sandlund, N; Faller, R; Wiik-Nielsen, J; Jensen, B B (2012). First detection of
piscine reovirus (PRV) in marine fish species. Diseases of Aquatic Organisms 97: 255–288 [Online] Available
from: http://www.doi.10.3354/dao02425 [Accessed 11 June 2019].
Wiklund, T (2016). Pseudomonas anguilliseptica infection as a threat to wild and farmed fish in the Baltic Sea:
Under the microscope. Microbiology Australia, September 2016, 2 p [Online] Available from: http://www.doi.
10.1071/MA16046 [Accessed 14 January 2020].
Wiklund, T; Bylund, G (1990). Pseudomonas anguilliseptica as a pathogen of salmonid fish in Finland. Diseases
of Aquatic Organisms 8: 13–19.
Williams, C (2019). Sanford Ltd, Auckland. Personal communication.
Wingfield, W H; Fryer, J L; Pilcher, K S (1969). Properties of the sockeye salmon virus (Oregon strain).
Proceedings of the Society for Experimental Biology and Medicine 130: 1055–1059.
Winton, J R; Hershberger, P K (2014). Viral Erythrocytic Necrosis. Chapter 2.2.7. The Blue Book 2014. Fish
Health Section, American Fisheries Society [Online] Available from: http://afs-
fhs.org/perch/resources/14069231372.2.7ven2014.pdf [Accessed 6 October 2017].
Wolf, K (1988). Fish viruses and fish viral diseases. Cornell University Press, Ithaca, New York, United States of
America, 476 p.
WoRMS, (2019). World Register of Marine Species- An authoritative classification and catalogue of marine names,
Editorial Board, Flanders Marine Institute, Belgium, [Online] Available from:
http://www.doi:10.14284/170ww.marinespecies.org at VLIZ. [Accessed 7 October 2019].
WTO, (2017). Review of the operation and implementation of the SPS Agreement, G/SPS/62, World Trade
Organisation, Geneva, Switzerland, 47 p [Online] Available from:
https://www.wto.org/english/tratop_e/sps_e/sps_e.htm [Accessed 17 December 2019].
Wynne-Jones, J; Gray, A; Heinemann, A; Hill, L; Walton, L (2019). National panel survey of marine
recreational fishers 2017–2018. New Zealand Fisheries Assessment Report 2019/24, 104 p. Fisheries New Zealand,
Ministry for Primary Industries, Wellington, New Zealand. [Online] Available from:
https://fs.fish.govt.nz/Doc/24728/FAR-2019-24-National-Panel-Survey-Marine-Recreational-Fishers.pdf.ashx
[Accessed 21 January 2020].
Wynne-Jones, J; Gray, A; Hill, L; Heinemann, A (2014). National Panel Survey of Marine Recreational Fishers
2011–12: Harvest Estimates. New Zealand Fisheries Assessment Report 2015/48, 94 p. Ministry for Primary
Industries, Wellington, New Zealand [Online] Available from:
https://fs.fish.govt.nz/Doc/23936/FAR_2015_48_2853_MAF2010-01.pdf.ashx [Accessed 21 January 2020].
Xu, X; Huang, L; Weng, S; Wang, J; Lin, T; Tang, J; Li, Z; Lu, Q; Xia, Q; Yu, X; He, J (2010). Tetraodon
nigroviridisas a nonlethal model of infectious spleen and kidney necrosis virus (ISKNV) infection. Virology 406:
167–175 [Online] Available from: https://doi.org/10.1016/j.virol.2010.07.003 [Accessed 28 January 2020].
Yamada, Y; Wakabayashi, H (1999). Identification of fish-pathogenic strains belonging to the genus Edwardsiella
by sequence analysis of sodB. Fish Pathology 34 (3): 145-150.
Yanong, R P E; Francis-Floyd, R (2019). Streptococcan infections of fish. IFAS Extension circular 57, School of
Forest Resources and Conservation, Program in Fisheries and Aquatic, UF/IFAS Extension, University of Texas,
Gainesville, Texas, United States of America, 5 p [Online] Available from:
https://www.edis.ifas.ufl.edu/pdffiles/FA/FA05700.pdf [Accessed 14 January 2020].
Yasunaga, Ν (1982). Occurrence- of Streptococcus sp. a pathogen of cultured yellowtail in muscle of sardine for
diets. Fish Pathology 17: 195–198.
Yokoyama, H; Inoue, D; Sugiyama, A; Wakabayashi, H (2000). Polymerase chain reaction and indirect
fluorescent antibody technique for the detection of Kudoa amamiensis (Multivalvulida: Myxozoa) in
yellowtail Seriola quinqueradiata. Fish Pathology 35: 157–162.
Yokoyama, H; Grabner, D; Shirakashi, S (2012). Transmission biology of the Myxozoa, pp. 3-42 In: Carvalho
ED, David GS, Silva RJ (Eds.), Health and Environment in Aquaculture, InTech, Rijeka, Croatia [Online] Available
from: http://www.intechopen.com/books/health-and-environment-in-aquaculture [Accessed 1 November 2019].
Yokoyama, H; Shirakashi, S (2007). Evaluation of hyposalinity treatment on infection with Enteromyxum leei
(Myxozoa) using anemonefish Amphiprion spp. as experimental host. Bulletin of the European Association of Fish
Pathologists 27: 74–78.

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 155
Yoshida, T; Yamada, Y; Sakai, M; Inglis, V; Xie, X J; Chen, S C; Kruger, R (1996). Association of the cell
capsule with anti-opsonophagocytosis in I-haemolytic Streptococcus spp. isolated from rainbow trout. Journal of
Aquatic Animal Health 8: 223–222.
Yoshimizu, M; Yoshinaka, T; Hatori, S; Kasai, H (2005). Survivability of fish pathogenic viruses in
environmental water and the inactivation of fish viruses. Bulletin of the Fisheries Research Agency of Japan,
Supplement 2: 47–54.
Young, C A; Jones, S R M (2005). Epitopes associated with mature spores not recognized on Kudoa thyrsites from
recently infected Atlantic salmon smolts. Diseases of Aquatic Organisms 63: 267−271.
Yow, B; McBride, S; Mowbray, J; Caughey, W; Wong, E; Luey, C; Siddiqui, A; Alexander, Z; Playle, V;
Askelund, T; Hopkins, C; Quek, N; Ross, K; Holland, D (2018). Enterobacteriaceae native joint septic arthritis.
Section 54: Open joint infections: Open Forum Infectious Diseases 5(1): s124 [Online] Available from:
https://www.doi.org/10.1093/ofid/ofy210.316 [Accessed 7 June 2019].
Yurakhno, V M (2011). Two new families and a new species of myxosporeans (Myxozoa, Myxosporea) of the
Mediterranean and Black Sea fishes. Vestnik zoologii, 45(6): 48–52.
Yurakhno, V M; Ovcharenko N A (2008). Myxosporeans of the world ocean mullets, pp. 231–234 In:
Parasitology in XXI century-problems, methods, solutions, Volume 3. Proceedings of the IV Congress of the
Russian Society of Parasitologists-Russian Academy of Sciences: 20-25 October 2008, St. Petersburg, Russia.
Yurakhno, V M; Ovcharenko, M O (2014). Study of Myxosporea (Myxozoa), infecting worldwide mullets with
description of a new species. Parasitological Research 113: 3661–3674 [Online] Available from:
http://www.doi.10.1007/s00436-014-4031-5 [Accessed 22 October 2019].
Yurakhno, V M; Ovcharenko, M O; Holzer, A S; Sarabeev, V L; Balbuena, J A (2007). Kudoa unicapsula n.
sp. (Myxosporea: Kudoidae) a parasite of the Mediterranean mullets Liza ramada and L. aurata (Teleostei:
Mugilidae). Parasitological Research 101: 1671–1680 Online] Available from: http://www.doi.10.1007/s00436-
007-0711-8 [Accessed 12 September 2019].
Zainathan, S C; Carlile, G; Carson, J; McColl, K A; Crane, M St J; Williams, L M; Hoad, J; Moody, N J G;
Aiken, H M; Browning, G F; Nowak, B F (2014). Development and application of molecular methods (PCR) for
detection of Tasmanian Atlantic salmon reovirus. Journal of Fish Diseases 38(8): 739–754 [Online] Available from:
https://doi.org/10.1111/jfd.12291 [Accessed 21 January 2020].
Zorriehzahra, M J; Adel, M; Dadar, M; Ullah, S; Ghasemi, M (2016). Viral nervous necrosis (VNN) an
emerging disease caused by Nodaviridae in aquatic hosts: Diagnosis, control and prevention: A review. Iranian
Journal of Fisheries Sciences [Online] Available from: http://www.doi.10.22092/ijfs.2018.117849 [Accessed 21
October 2019].
Zorriehzahra, M; Nakai, T; Sharifpour, I; Gomez. D G; Chi, C S; Soltani, M; Hassan, H M D; Rohani, M S;
Saidi, A A (2005). Mortality of wild golden grey mullet (Liza auratus) in Iranian waters of the Caspian Sea,
associated with viral necrosis-like agent. Iranian Journal of Fisheries Sciences 4: 43–58.
Zhang, C X; Suzuki, S (2004). Aquabirnaviruses isolated from marine organisms form a distinct genogroup from
other aquabirnaviruses. Journal of Fish Diseases 27: 633 643.
Zhang, J; Meng, F; Yokoyama, H; Miyahara, J; Takami, I; Ogawa, K (2010). Myxosporean and
microsporidian infections in cultured Pacific bluefin tuna Thunnus orientalis in Japan. Fisheries Science 76(6): 981–
990.
Zhang, Q Y; Gui, J F (2015). Virus genomes and virus-host interactions in aquaculture animals. Special Topic:
Fish biology and biotechnology: A review. Science China Life Sciences 58(2): 156–169.
Zhou, S M; Xie, M; Zhu, X Q; Ma, Y; Tan, Z L; Li, A X (2011). Rapid identification of Streptococcus iniae by
specific PCR assay utilising genetic markers in ITS rDNA. Journal of Fish Diseases 34(4): 265–271.
Zhou, Y; Millhouse, E; Shaw, T; Lappin, D F; Rajendran, R; Bagg, J; Lin, H; Ramage, G (2018). Evaluating
Streptococcus mutans Strain Dependent Characteristics in a Polymicrobial Biofilm Community. Frontiers in
Microbiology 9: 1498 [Online] Available from: https://www.doi.org/10.3389/fmicb.2018.01498 [Accessed 14
January 2020].
Zlotkin, A; Hershko, H; Eldar A (1998). Possible Transmission of Streptococcus iniae from Wild Fish to Cultured
Marine Fish. Aquatic and Environmental Microbiology 64(10): 4065–4067.

156 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Appendices
Appendix 1. Historical fish bait imports, 2008-2017
Prior to 2008, New Zealand’s commercial fishing industry was largely self-sufficient for fish bait
(Blackwell, 2013). Between 2008 and 2017, increases in demand from recreational and
commercial fishers resulted in the importation of a wide range of finfish (and cephalopod species
as fish bait.
1.1 Historical imports of finfish as bait, 2008-2017
The location of capture varied for each species between years depending on price and
availability (Table 21). Many fish bait species are widely distributed and the fishing fleets range
over a wide geographical area. (B. Burney, pers. comm., 2019; R. Clark, pers. comm., 2019; P.
Lamb, pers. comm., 2019; A. Spencer, pers. comm., 2019). Importantly, the reported country of
export or purchase of imported fish bait species may differ from the actual catch location (Jereb
et al., 2010; Granados-Amores et al., 2014; Norman et al., 2014; FAO, 2018; FishBase, 2019; B.
Burney, pers. comm., 2019; R. Clark, pers. comm., 2019).
Imports of redbait/Cape bonnetmouth (Emmelichthys nitidus nitidus) (Emmelichthyidae), silver
warehou (S. punctata) (Centrolophidae), and coalfish (Pollachius virens) (Gadidae) were minor
(< 1 tonne). These species are no longer imported (B. Burney, pers. comm., 2019). Other species
such as the European pilchard (Sardinia pilchardus) have not been recently imported. These
species were therefore not included in the list of species considered as fish bait (Table 5) and are
not considered further.
Table 21. Taxonomy and geographical distribution of finfish imported as fish bait, 2008-2017
Family Common name Species Geographical distribution1
Carangidae Pacific jack mackerel Trachurus symmetricus Eastern Pacific (Alaska to Mexico, Galapagos Is.)
Chilean jack mackerel Trachurus murphi South and Southwest Pacific, Southwest Atlantic
Japanese jack mackerel Trachurus japonicus Northwest Pacific, Japan, Korea
Centrolophidae Silver warehou Seriolella punctata Eastern Indian Ocean, Southwest and Southeast Pacific Ocean
Clupeidae Atlantic herring Clupea harengus Northwest (USA) and Northeast Atlantic Ocean, Europe
Pacific herring Clupea pallasii pallasii (Clupea pallasii)
Northeast and Northwest Pacific, Pacific Ocean
South American pilchard or sardine
Sardinops sagax Cosmopolitan: S. sagax ocellatus (S. Africa), S. s. neopilchardus (Australia), S. s. caerulus (California), S. s. melanostictus (China, Japan)
Round sardinella Sardinella aurita East and west Atlantic Ocean, Mediterranean Sea
Indian oil sardine Sardinella longiceps South and West Indian Ocean (East Indian coast)
Bali sardinella Sardinella lemuru Pacific Ocean (Asia, SE Asia, Australia)
Spotted sardinella Amblygaster sirm Indo-Pacific (Red Sea to Japan, New Guinea, Northern Australia, Fiji)

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 157
Family Common name Species Geographical distribution1
European pilchard Sardina pilchardus Northeast Atlantic, North Sea, Mediterranean
Emmelichthyidae Redbait/Cape bonnetmouth
Emmelichthys nitidus nitidus
Indo-Pacific, Eastern Atlantic Ocean
Engraulidae Californian anchovy Engraulis mordax Northeast Pacific Ocean (Vancouver to Mexico)
Peruvian anchovy Engraulis ringens Southeast Pacific Ocean (Peru to Chile)
Exocoetidae Tropical two-wing flying fish
Exocoetus volitans Widespread tropical, subtropical oceans
Sailfin flying fish Parexocoetus brachypterus
Indo-Pacific, West and East Atlantic Oceans
Gadidae Coalfish (saithe) Pollachius virens Northeast, Northwest Atlantic Ocean
Hemiramphidae Balao halfbeak Hemiramphus balao West Atlantic Ocean
Dussumier’s halfbeak Hyporamphus dussumieri Indo-Pacific (India to Okinawa)
Ballyhoo halfbeak Hemiramphus brasiliensis
West Atlantic (New York to Brazil), East Atlantic Oceans (Cape Verde to Angola)
Luke’s halfbeak Hemiramphus lutkei (= H. marginatus)
North Pacific (China, Japan), Southeast Asia, Pacific Islands
Jumping halfbeak Hemiramphus archipelagus (= H. marginatus)
Indo-Pacific, southeast Asia, Polynesia
Southern garfish Hyporhamphus melanochir
East Indian Ocean, Pacific Ocean (Australia)
Japanese halfbeak Hyporhamphus sajorii Northwest Pacific Ocean (Japan to China, Korea)
Mugilidae Flathead grey mullet Mugil cephalus Widespread in tropical, sub-tropical and temperate waters
Sand grey mullet, Myxus elongatus Southwest Pacific, Australia
South African mullet Chelon (Liza) richardsonii Southeast Atlantic, South Africa
Yellow eye mullet Aldrichetta forsteri Southwest Pacific, Australia, New Zealand
Scomberesocidae Sanmar, Pacific saury Cololabis saira Pacific Ocean (Japan, Alaska south to Mexico)
Scombridae Bonito (Bullet tuna) Auxis rochei Atlantic, Indian and Pacific Oceans
Bonito (Frigate tuna) Auxis thazard Atlantic, Indian and Pacific Oceans
Little tunny Euthynnus alletteratus Atlantic Ocean (widespread tropical, subtropical)
Skipjack tuna Katsuwonus pelamis Cosmopolitan (Atlantic, Indian and Pacific Oceans)
Atlantic bonito Sarda sarda Cosmopolitan (East and West Atlantic Ocean, Europe, Mediterranean Sea)
Eastern Pacific bonito Sarda chiliensis East Pacific Ocean (Chile to Peru)
Pacific bonito Sarda lineolata Northeast Pacific Ocean (Alaska to California)
Blue mackerel Scomber australasicus Indo-West Pacific
Chub (Pacific) mackerel Scomber japonicus Indo-Pacific, temperate

158 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Family Common name Species Geographical distribution1
Atlantic mackerel Scomber scombrus Atlantic Ocean, Mediterranean Sea
Bigeye tuna Thunnus obesus Cosmopolitan (tropical, subtropical waters)
Source New Zealand fish bait industry: B. Burney, pers. comm., 2019; R. Clark, pers. comm., 2019); D Rutherford, pers. comm., 2019. Notes 1 Reported distribution (FishBase, 2019)
1.2 Historical imports of coleoid cephalopods as bait, 2008-2017
Coleoid cephalopod molluscs (squid, cuttlefish and octopus) support extensive commercial
fisheries worldwide, both for use as bait and for human consumption. Historical imports of
coleoid molluscs into New Zealand during 2008-2017 were mainly squid of the families
Loliginidae and Ommastrephidae (Table 22). As previously noted, the reported country of export
or purchase of imported bait species may differ from the actual catch location.
Coleoid cephalopod molluscs were not considered in the previous IRA for fish food and fish bait
(Cobb 2008) and are not specifically covered in the IHS for fish bait (FISFOOIC.ALL) (MPI
2011). They have historically been imported as a food-grade product under the IHS
(FISMARIC.ALL) (MPI 2008), of which a variable proportion has then been on-sold as bait.
This is because the quality of industrial (non-food grade) product is considered too low for
commercial use as bait (B. Burney, pers. comm. 2018). No trade import data are available
separately for coleoid molluscs imported as fish bait. Of the total imports (by volume) of coleoid
cephalopod molluscs during 2013-2015 (Notes
1 Jereb & Roper (2010).
2 Within the Loglinidae, the genus Loligo is paraphyletic, with species based largely on geographical distribution 3 The classification of species within the Octopus species complex is currently under review, with 350 named species reported
(Norman et al. 2014) and over 150 potentially undescribed species known from tropical and subtropical waters (FishBase 2019).
Table 23), industry sources estimate that 700-800 t, representing approximately 35% of these
annual totals, were redirected annually for use as fish bait (B. Burney, pers. comm. 2019).
Table 22. Classification and distribution of coleoid cephalopod molluscs imported, 2008-2017
Family Common name Species Geographical distribution1
Loliginidae (Long-finned squids)2.
Patagonian squid Doryteuthis gahi (= Amerigo (Loligo) gahi)
Southeast Atlantic, Falkland Islands, Southwest Pacific
California market squid, opalescent inshore squid
Doryteuthis opalescens (= Loligo opalescens)
East Pacific (Alaska to Mexico)
Long-finned inshore squid
Doryteuthis pealeii (= Loligo (Amerigo) pealeii)
West Atlantic (Newfoundland to Venezuela)
Loligo squid undefined Loligo vulgaris Europe, Mediterranean, Northeast Atlantic
Spear squid Heterololigo (Loligo) bleekeri
Northwest Pacific (Japan to Korea)
Common squid Loligo forbesii Mediterranean Sea, Northeast Atlantic (Scotland to the Azores)
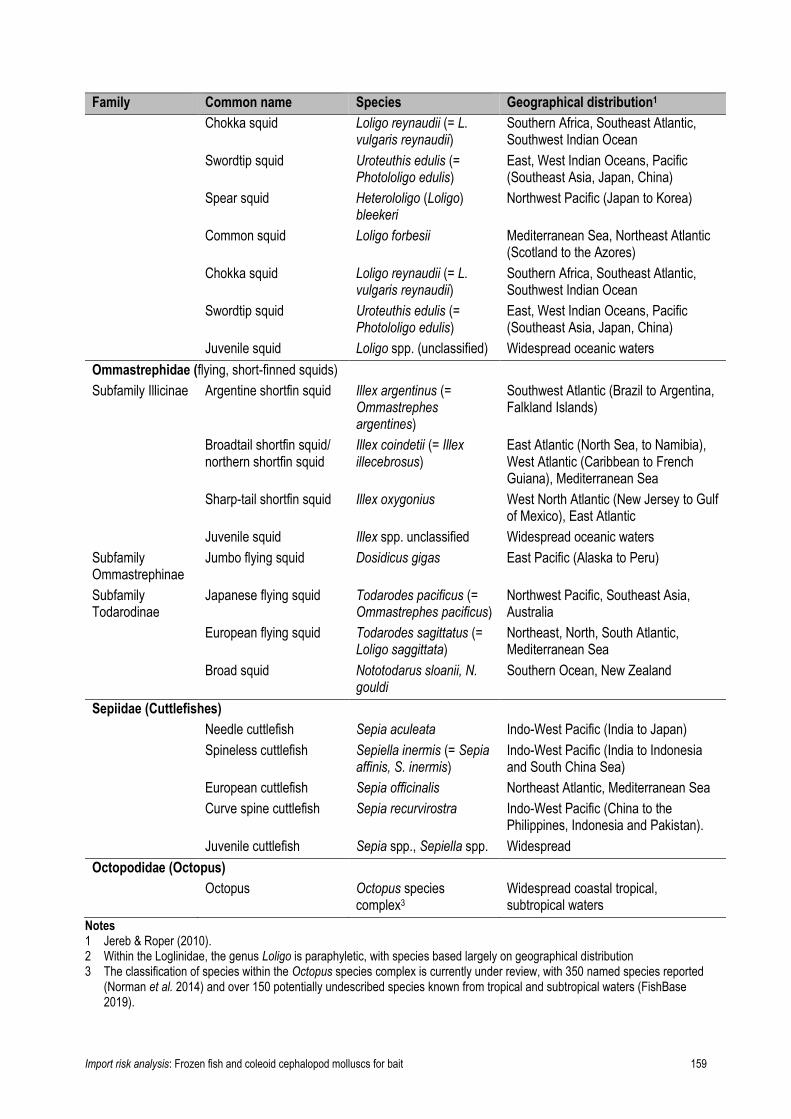
Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 159
Family Common name Species Geographical distribution1
Chokka squid Loligo reynaudii (= L. vulgaris reynaudii)
Southern Africa, Southeast Atlantic, Southwest Indian Ocean
Swordtip squid Uroteuthis edulis (= Photololigo edulis)
East, West Indian Oceans, Pacific (Southeast Asia, Japan, China)
Spear squid Heterololigo (Loligo) bleekeri
Northwest Pacific (Japan to Korea)
Common squid Loligo forbesii Mediterranean Sea, Northeast Atlantic (Scotland to the Azores)
Chokka squid Loligo reynaudii (= L. vulgaris reynaudii)
Southern Africa, Southeast Atlantic, Southwest Indian Ocean
Swordtip squid Uroteuthis edulis (= Photololigo edulis)
East, West Indian Oceans, Pacific (Southeast Asia, Japan, China)
Juvenile squid Loligo spp. (unclassified) Widespread oceanic waters
Ommastrephidae (flying, short-finned squids)
Subfamily Illicinae Argentine shortfin squid Illex argentinus (= Ommastrephes argentines)
Southwest Atlantic (Brazil to Argentina, Falkland Islands)
Broadtail shortfin squid/ northern shortfin squid
Illex coindetii (= Illex illecebrosus)
East Atlantic (North Sea, to Namibia), West Atlantic (Caribbean to French Guiana), Mediterranean Sea
Sharp-tail shortfin squid Illex oxygonius West North Atlantic (New Jersey to Gulf of Mexico), East Atlantic
Juvenile squid Illex spp. unclassified Widespread oceanic waters
Subfamily Ommastrephinae
Jumbo flying squid Dosidicus gigas East Pacific (Alaska to Peru)
Subfamily Todarodinae
Japanese flying squid Todarodes pacificus (= Ommastrephes pacificus)
Northwest Pacific, Southeast Asia, Australia
European flying squid Todarodes sagittatus (= Loligo saggittata)
Northeast, North, South Atlantic, Mediterranean Sea
Broad squid Nototodarus sloanii, N. gouldi
Southern Ocean, New Zealand
Sepiidae (Cuttlefishes)
Needle cuttlefish Sepia aculeata Indo-West Pacific (India to Japan)
Spineless cuttlefish Sepiella inermis (= Sepia affinis, S. inermis)
Indo-West Pacific (India to Indonesia and South China Sea)
European cuttlefish Sepia officinalis Northeast Atlantic, Mediterranean Sea
Curve spine cuttlefish Sepia recurvirostra Indo-West Pacific (China to the Philippines, Indonesia and Pakistan).
Juvenile cuttlefish Sepia spp., Sepiella spp. Widespread
Octopodidae (Octopus)
Octopus Octopus species complex3
Widespread coastal tropical, subtropical waters
Notes 1 Jereb & Roper (2010). 2 Within the Loglinidae, the genus Loligo is paraphyletic, with species based largely on geographical distribution 3 The classification of species within the Octopus species complex is currently under review, with 350 named species reported
(Norman et al. 2014) and over 150 potentially undescribed species known from tropical and subtropical waters (FishBase 2019).

160 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Table 23. Imports of coleoid cephalopod molluscs, 2013-2015
Total volume1 (t)
Year 2013 2014 2015
Family Common name Scientific name
Loliginidae, Ommastrephidae
Squid Illex spp., Ommastrephes spp., Loligo spp., Nototodarus spp., Sepioteuthis spp.
1,900 2,019 1,949
Sepiidae Cuttlefish Sepia spp. 44 30 41
Octopodidae Octopus Octopus spp. 52 71 47
Total 1,996 2,140 2,037
Source Trade data, Department of Statistics (2017). Notes 1 Estimated volume (t) of imports for human consumption and for use as bait

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 161
Appendix 2. Pathogen association by fish bait host species considered in this risk analysis (Table 5)
Table 24. Risk organisms and associated fish bait species where specific risk management measures are proposed
Host MABV RSIV/ISKNV ENV NNV IHNV VHSV Streptococcus agalactiae III: 283
Streptococcus iniae
OIE-listed N Y N N Y Y N N
Family Carangidae
Trachurus japonicus √ √ √ √
Trachurus murphi √
Trachurus symmetricus √
Trachurus trachurus √
Family Clupeidae
Ambylygaster sirm √ √
Clupea harengus √ √ √ √
Clupea pallasii pallasii √ √ √ √ √
Sardinella aurita √ √
Sardinella lemuru √ √
Sardinella longiceps √ √
Sardinops sagax √ √ √
Family Engraulidae
Engraulis mordax √
Engraulis ringens
Family Hemiramphidae
Hemiramphus balao
Hemiramphus brasiliensis
Hemiramphus dussumieri
Hemiramphus lutkei

162 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Host MABV RSIV/ISKNV ENV NNV IHNV VHSV Streptococcus agalactiae III: 283
Streptococcus iniae
Family Mugilidae
Chelon auratus √
Chelon labrosus √
Chelon ramada √
Mugil cephalus √ √ √ √ √
Family Scomberosocidae
Cololabias saira
Family Scombridae
Auxis rochei
Auxis thazard
Euthynnus alletteratus
Katsuwonas pelamis
Sarda chilensis
Sarda lineolata
Sarda sarda
Scomber australasicus √
Scomber japonicus √ √ √ √
Scomber scombrus √ √
Thunnus obesus
Family Loliginae
Doryteuthis opalescens
Loligo spp.
Uroteuthis duvaucelli
Family Ommastrephidae
Illex argentinus
Illex coindetti
Todarodes pacificus √

Import risk analysis: Frozen fish and coleoid cephalopod molluscs for bait 163
Host MABV RSIV/ISKNV ENV NNV IHNV VHSV Streptococcus agalactiae III: 283
Streptococcus iniae
Family Sepiidae
Sepia recurvirostrata
Sepia spp.
Family Octopodidae
Octopus vulgaris √
Octopus spp.
Note: General risk management options are likely to provide sufficient reduction in pathogen occurrence for Edwardsiella spp.,Enteromyxum leei, Kudoa clupeidae, K. iwatai, K. nova and K. thyrsites. No specific risk management options are therefore necessary.

164 Import risk analysis: Frozen fish and cephalopod molluscs for fish bait
Appendix 3. Summary and evaluation of risk management options
Table 25 summarises the qualitative evaluation of the expected reduction in pathogen load
associated with each risk management option in the relevant risk organism chapter.
The estimated levels of reduction in occurrence are:
Eliminate > 95% reduction in pathogen load
Substantial 71-95% reduction in pathogen load
Moderate 51-70% reduction in pathogen load
Slight < 50% reduction in pathogen load
Table 25. Assumed reduction in pathogen load associated with risk management options
Risk organism MABV RSIV/
ISKNV ENV NNV IHNV VHSV
General risk management options
Health certification Moderate Moderate Moderate Moderate Moderate Moderate
Frozen storage (-18 ° to
-20 ° for 4 months (17 weeks)
No effect No effect No effect No effect No effect Slight
Species specific risk management options
Source from species not associated with risk
organism Substantial Substantial Substantial Substantial Substantial Substantial
Source from region/country free of
risk organism Substantial Substantial Substantial Substantial Substantial Substantial
Pre-export or post-arrival batch testing
Substantial Substantial Substantial Substantial Substantial Substantial
Ionising radiation Substantial Substantial Substantial Substantial Substantial Substantial
Risk organism Edwardsiella
spp.
Streptococcus agalactiae III:
283
Streptococcus iniae
Enteromyxum leei
Kudoa clupeidae, K. iwatai, K. nova, K.
thyrsites
General risk management options
Health certification Moderate Moderate Moderate Moderate Moderate
Frozen storage ( -18 °
to -20 °C) for 4 months (17 weeks)
Substantial No effect No effect Substantial Substantial
Species specific risk management options
Source from species not associated with risk
organism N/A Substantial Substantial N/A N/A
Source from region/country free of
risk organism N/A Substantial Substantial N/A N/A
Pre-export or post-arrival batch testing
N/A Substantial Substantial N/A N/A
Ionising radiation N/Al Substantial Substantial N/A N/A
Notes “No effect” means a risk management option is unlikely to reduce pathogen occurrence N/A” means a risk management option is not applicable.




















