
ENHANCING POISON BAIT ACCEPTANCE AND TRAPPING OF Rattus rattus USING POULTRY EGG COMPONENTS AS BAIT...
101
CHAPTER – I INTRODUCTION Rodents have been identified as the most important mammalian pests at the global level (Cuong et al 2002). They cause significant damage to crops at pre- and post-harvest stages throughout the world (Prakash 1988, Buckle and Smith 1994, Parshad 1999a, Singleton et al 1999, Amusa et al 2005, Fayenuwo et al 2007, Meerburg and Kijlstra 2008, Singla and Parshad 2010, Singla and Babbar 2010 and 2012), with yield losses of 5-15% in most of the countries (Singleton and Petch 1994, Singleton 2003). Damages caused by rodents lead to huge amount of food shortages (Palis et al 2007). The rodent fauna of the Indian sub-continent is represented by 46 genera and 128 species (Ellerman 1961, Roonwal 1987). Of these 18 species are commensal and agricultural pests. Present checklist in India reports 103 species and 89 subspecies under 46 genera which belong to 7 families (Pradhan and Talmale 2011). Of these, 20 species have been reported to be of economic importance in India (Sridhara and Tripathi 2005). Their habitat, distribution, abundance and economic significance vary in different crops, seasons and geographical regions of the country (Rana et al 2006). Despite the development of a wide variety of rodent control strategies to limit their damage (Parshad 1999a, Singleton et al 1999, Parshad et al 2006, Weihong et al 1999), rodent control has not yet become an integral component of crop production and storage strategies in India. Analysis of the information available on the damage and economic losses caused by rodents in various crop fields, horticulture and forestry, poultry farms, rural and urban dwellings and storage facilities showed that chronic damage
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of ENHANCING POISON BAIT ACCEPTANCE AND TRAPPING OF Rattus rattus USING POULTRY EGG COMPONENTS AS BAIT...

CHAPTER – I
INTRODUCTION
Rodents have been identified as the most important
mammalian pests at the global level (Cuong et al 2002). They
cause significant damage to crops at pre- and post-harvest
stages throughout the world (Prakash 1988, Buckle and Smith
1994, Parshad 1999a, Singleton et al 1999, Amusa et al 2005,
Fayenuwo et al 2007, Meerburg and Kijlstra 2008, Singla and
Parshad 2010, Singla and Babbar 2010 and 2012), with yield
losses of 5-15% in most of the countries (Singleton and Petch
1994, Singleton 2003). Damages caused by rodents lead to huge
amount of food shortages (Palis et al 2007). The rodent fauna of
the Indian sub-continent is represented by 46 genera and 128
species (Ellerman 1961, Roonwal 1987). Of these 18 species are
commensal and agricultural pests. Present checklist in India
reports 103 species and 89 subspecies under 46 genera which
belong to 7 families (Pradhan and Talmale 2011). Of these, 20
species have been reported to be of economic importance in
India (Sridhara and Tripathi 2005). Their habitat,
distribution, abundance and economic significance vary in
different crops, seasons and geographical regions of the
country (Rana et al 2006). Despite the development of a wide
variety of rodent control strategies to limit their damage
(Parshad 1999a, Singleton et al 1999, Parshad et al 2006, Weihong
et al 1999), rodent control has not yet become an integral
component of crop production and storage strategies in India.
Analysis of the information available on the damage and
economic losses caused by rodents in various crop fields,
horticulture and forestry, poultry farms, rural and urban
dwellings and storage facilities showed that chronic damage

`
ranging from 2 to 15% persisted throughout the country
(Parshad 1999a). There are reports on rodents causing 6-8%
losses in rice crop and 10-12% in wheat crop in India (Santra
and Manna 2008, Chattopadhyay et al 2010). Annual food losses
caused by rodents in India are estimated to be 10-15% of the
total national production (Malhi and Sheikher 1989). Storage
losses to rodents in India alone are 25-30% costing at least
$5 billion annually (Cao et al 2002). In addition, rodents also
destroy food by contaminating it with their urine, faecal
droppings and hair. They are also important vectors or
reservoirs of numerous diseases that infect humans, domestic
animals and other wildlife species (Gratz 1994, Singla et al
2003, 2008a and 2012, Pai et al 2005, Meerburg et al 2009). They
are known to serve as hosts for at least 60 diseases (Hugh-
Jones et al 1995). The need to control commensal rodents is
rarely controversial, as the public has always associated rats
with lethal, disease-causing organisms such as bubonic plague.
While plague has long been absent from some countries,
including India, the potential health threat from commensal
rodents is now focused on other zoonotic diseases, such as
leptospirosis and salmonellosis (Cowan et al 2003).
The reasons for the subsistence of large rodent
populations in most of the storage premises and human
dwellings is the inadequate maintenance of buildings combined
with lack of hygiene, poor handling of food materials leading
to spillage and serious neglect of rodent proofing. The house
rat, Rattus rattus (Linnaeus) is one of the most common commensal
rodent pest worldwide. It often damages, contaminates and
spoils packed food and non-food materials in transit and
storage (Parshad 1999a). It is purely an indoor pest in
2

`
Pakistan (Roberts 1977, Brooks et al 1987, Ahmed et al 1995, Khokhar
et al 1999, Hussain and Iqbal 2002). It is the predominant pest
species infesting and depredating poultry farms in India with
highest annual productivity of 69.59 young/female/year
reported for any Indian rodent species (Sridhara and
Krishnamurthy 1992, Parshad 1999b).
Poultry is one of the important meat industries of India.
Poultry farms provide a most favourable and stable habitat
throughout the year for large populations of R. rattus (Parshad et
al 1987). It causes severe economic losses to poultry
production directly by feeding on poultry feed, contaminating
it with their excrements, damaging eggs, attacking and killing
chicks, causing structural damage to buildings, doors, windows
and feed containers and indirectly by transmitting or carrying
several diseases (Ahmad et al 1984, Chopra and Dhindsa 1987,
Parshad et al 1987, Soni and Rana 1988, Chopra 1992, Sridhara and
Krishnamurthy 1992, Gomez Villafane et al 2001, Hussain et al
2006, Chopra et al 2008a and b). The species acts as a wild
reservoir horizontally transmitting infectious organisms to
other rodent species and arthropod vectors living closer to
anthropized environments, thus leading to inevitable exchange
of pathogens between rodents, animals and humans (Chopra et al
2008b). Hence it is very important to control this species.
A wide range of control measures have been used from time
to time for the control of commensal rodents depending upon
the ecological conditions. Conventional methods of rodent
control all over the world include proofing, trapping and the
use of rodenticides (Parshad 1999b, Weihong et al 1999, Parshad
et al 2006). Trapping rats is an age old method which is
ecologically sound and environment friendly. Some scientists
3

`
have shown that trapping can, under some circumstances, be an
effective method of rodent pest management (Gebauer et al 1992,
Tobin et al 1993, Islam and Karim 1995). The use of appropriate
baits is, however, important when attempting to attract
certain species into traps (Sivaprakasam and Durairaj 1995,
Szocs et al 2004, Fuller et al 2005). Currently, the use of
rodenticides is the major method of rodent pest management.
Acute rodenticides and anticoagulants are the most effective
and widely used groups (Buckle 1994). Both have their own
merits and demerits and are recommended for use against
rodents under different conditions. Out of these two kinds of
rodenticides, acute poisons are more preferred and frequently
applied as people are anxious to see a rapid kill and get rid
of damage caused by them (Sterner et al 1996, Steven 2008).
Rodent control with rodenticide baits in crop lands and
other premises has often been found ineffective in reducing
rodent densities due to several factors (Pank 1976). Although
zinc phosphide has been reported to be an effective acute
rodenticide (Matschke et al 1982, Sterner et al 1996), numerous
researchers have reported bait acceptance problems related to
bitter taste, sub-lethal toxicosis and subsequent conditioned
aversion after rodents have ingested minimal levels of zinc
phosphide bait (Reidinger and Mason 1983, Prakash and Ghosh
1992, Reidinger 1997) resulting in reduced efficacy of
rodenticide baiting (Rzoska 1953, Prakash and Jain 1971,
Berdoy and MacDonald 1991, Sterner et al 1996). In no-choice
tests, zinc phosphide often produces 100% mortality, but in
bi-choice tests that more closely simulate its use in crop
fields, mortality levels can range from 50 to 75%.
4

`
Low bait uptake often occurs in connection with enhanced
neophobia (Berdoy 1994). There is, however, no evidence for a
genetic basis of different levels of neophobia in rat
populations (Macdonald et al 1999). Rats show initial avoidance
towards unfamiliar bait and new bait boxes in a familiar
environment (Barnett 1958). After initial intake of sublethal
dose, rats rapidly learn to avoid eating a poisonous mixture
or particular cereals used as bait base and become bait shy
(Howard and Marsh 1970, Bhardwaj and Khan 1979). Bait-shyness
induced through conditioned taste aversion, can last more than
a year, even when the rodenticides have been removed from the
baits (Howard and Marsh 1970, Owan 1978, Owan et al 1979,
Shepherd and Inglis 1993). Once shy, the rats prefer to remain
hungry than eating an apprehensive food (Sood and Gill 1980,
Prescott et al 1992). Several factors may influence selection of
bait by different species such as taste (Frank 1988),
olfactory sensitivity (Vander Wall et al 2003) and learned
feeding behavior (Galef and Whiskin 2001).
Several researchers have reported the need for an
additive that could be added to rodenticide baits and traps
(Smythe 1976, Marsh 1988, Shafi et al 1990, Parshad and Jindal
1991, Shafi et al 1991, 1992a, b, c and 1993, Koehler et al 1994,
Reidinger 1997, Khan et al 2000, Pervez et al 1999, 2003 and 2005)
for increasing their efficacy. Effective attractants would
allow managers to attract rodents to remote detection devices,
bait stations, or traps. The bait stations, in turn, might
have oral baits containing rodenticides or fertility control
materials (where non-target hazards preclude lethal control).
Most research along these lines, however, has been on food
flavor additives to enhance palatability and the amount of
5

`
time spent feeding on toxic baits (Meehan 1984, Marsh 1988).
Unfortunately, identification of effective rat attractants has
eluded researchers to date. Bullard (1985) reported that rats
were more interested in familiar foods than in a wide array of
odours tested. Additionally, much more effort has been put
into research on rodent repellents than on attractants (Meehan
1984). Bait additives have a long, if not ancient history.
But, till date, no rat attractant has ever been developed. The
best "attractant" so far is just a good bait.
Different workers have suggested the efficacy of
different locally available bait additives against different
rodent species (Kumar and Gangwar 2001, Kamal and Hossain
2003, Pervez et al 2003, Kaur and Parshad 2005, Alam et al 2007,
Witmer et al 2010, Naeem et al 2011, Kok et al 2013, Mushtaq et al
2013). Usually it is recommended that poultry feed may be usedas baiting medium to contain rodent menace in poultry farms
(Bhardwaj 1983). There are some reports on preference for
poison bait containing egg shells and egg contents by rodents
in rice and wheat crop fields, respectively (Khan et al 2000,
Pervez et al 2005). Usually it is recommended that poultry feed
may be used as baiting medium for preparation of rodenticide
baits to curtail rodent menace in poultry farms (Bhardwaj
1983). These additives if added in zinc phosphide bait may
benefit the farming community in increasing its acceptance as
it still remains one of the most widely used rodenticides in
the subcontinent.
Since R. rattus is the predominant rodent pest species found
in poultry farms and feeding on poultry eggs, the bait
containing poultry egg components may be more acceptable to
it. Present study was therefore proposed to formulate a cereal
6

`
based bait package for R. rattus using poultry egg components such
as egg shell and egg albumin for enhancing poison bait
acceptance and trapping of R. rattus with following objectives:
i) To study the acceptance of different concentrations of egg
shell and egg albumin by R. rattus in laboratory experiments.
ii) To study the acceptance of best concentration of two
bait additives by R. rattus under food scale consumption
monitor.
iii) To determine the effect of best concentration of best
bait additive in enhancing poison bait acceptance and
trapping by R. rattus in poultry farms.
7

CHAPTER – II
REVIEW OF
LITERATURE
Rodents inflict incalculable losses to standing crops,
harvested crops in threshing floors and to stored food grains
and other commodities. Their damaging propensities in
rangelands, afforested lands, fruit orchards and plantation
crops and poultry farms are well known (Parshad 1999b). No
country in the world is free from quantitative and qualitative
losses inflicted by rodents. But the losses are more
pronounced in the tropical countries where rodents play a
significant role in retarding agricultural production through
damage to field crops (Jackson 1977, Parshad 1999a, Singla and
Parshad 2010, Singla and Babbar 2010, Borah and Bora 2012,
Gogoi and Borah 2013). The damage caused by field rodents maysometimes be estimated roughly for local districts where a
measurable proportion of various crops is destroyed by the
rodents. Commensal rodent damage is, however, difficult to
assess primarily because so many different items can be
involved and rodents can invade almost any type of structure
(Lund 1994). In food stores and warehouses, rodents can give
great problems, not only by consuming or fouling a substantial
part of the food, but also because they destroy sacks, bags,
boxes and other packaging material. In the slum areas of large
towns and certain villages in developing countries, there is a
constant fight between humans and rodents when hungry rats try
to gnaw fingers and toes of elderly and helpless people during
the night or babies when not looked after (Lund 1994). Rodent
consumption of stored food grains and damage to storage

`
structures and containers, and indirect losses caused by
spillage, spoilage, or contamination by hair or faecal
droppings that results in condemnation or rejections of
shipments by the importing country are important economic and
public health problems worldwide (Jackson 1977, Brooks and
LaVoie 1990, Conover et al 1995).
Rodents are very intelligent animals with a strong
smelling and tasting sense, which prevents them from instant
ingestion of poisons applied directly. The rodenticides are
therefore applied with different bait materials. But again
their behavioral characteristic termed as neophobia defends
and restricts them to immediately switch on to poison baits.
Such behavioral phenomena render the rodents control very
tricky, because once shy, the rats prefer to remain hungry
than eating an apprehensive food (Canby 1977, Riley and Clarke
1977, Owan 1978, Owan et al 1979, Sood and Gill 1980, Prescott et
al 1992). Many workers have tried to find out the most
attractive bait materials (Shumake 1978). Because of the large
space or diastema behind their incisors, rodents can use
incisors to investigate or nibble unfamiliar materials without
actually taking them inside their mouths. Rodents are
omnivorous, exhibiting choices and preferences in their diet,
but often selecting the most abundant, palatable food
available. They readily learn to reject or avoid unpalatable
foods or those containing toxins, which presents a problem for
the development of bait materials for effective delivery of
rodenticides.
2.1 Commensal rodents
Rodent pest species as commensal, literally means ‘eating
at the same table’. Pest rodents eat man’s foods, damage his
9

`
possessions and use his transport to distribute all over the
globe. Rodent species of the genus, Rattus have probably been
responsible for more human sufferings than any other group of
vertebrates, not only through their destructive impact on food
crops, but also through their role in the transmission of
fatal or debilitating diseases. However, the genus also
contains species of immense scientific and clinical
significance, such as the familiar Norway or laboratory rat
(Rattus norvegicus), as well as a host of lesser known but
presumably beneficial species that inhabit a wide range of
natural habitats from mangrove forests to subalpine
grasslands. The genus Rattus, with a total of 61 currently
recognised species, is the only single largest mammalian genus
of all which is most complex and least well understood. R.
rattus and R. norvegicus, both are the best-known commensal species
having global distribution (Aplin et al 2003). The house mouse
(Mus musculus) is also widely distributed and together with the
house rat and Norway rat, are known as commensal rodents
because of their close association with human habitation.
Since most rodent species involved in stored product damage
are nocturnal, heavy infestations may persist unnoticed
without careful inspection of stores or premises (Jackson
1990).
2.1.1 The house rat, Rattus rattus
The house rat, R. rattus (Rodentia: Muridae), is one of the
most commonly encountered and economically important commensal
rodents. It is nocturnal and prefers to forage for food above
ground in elevated areas indoors and outdoors. It is an agile
climber and travels through trees and along vines, wires,
rafters, and rooftops. It often uses trees and utility lines
10

`
to reach food and to enter buildings, but can also be found
foraging in dense ground cover. It can swim and may use sewer
systems to disperse to new areas. Outdoors, it can travel
several hundred feet in a single night to find its survival
resources. It prefers to nest in secluded areas above ground
in such places as attics, soffits, overhead garage storage, in
the vine cover of fences or buildings, and in wood piles or
other stored materials where harborage can be found (Askham
and Leonard 1987). It favours dense non-deciduous trees or
trees with hollow cavities and the crowns of palm trees,
especially when old fronds are not removed. It sometimes
burrows in the ground especially in hot, dry environments. In
these areas, it may use trees, materials stored on the ground,
concrete slabs and sidewalks to support shallow burrows. Roof
rat signs include smudge marks on surfaces from oil and dirt
rubbing off their fur as they travel. Sounds in the attic are
often the first indication of the presence of house rats in a
residence. At night when the house is quiet, the rats may be
heard scurrying about (Sullivan 2002).
The house rats have a high reproductive potential and may
breed year-round in warmer areas (Chattopadhyay et al 2010).
They can reproduce when they are 3 to 5 months old. Females
produce 5 to 8 pups per litter with average 4 to 5 litters per
year. The number of litters depends on the area and varies
with nearness to the limit of their climatic range,
availability of nutritious food, density of the local rat
population and the age of the rat (Marsh 1994). The gestation
period is between 21 and 29 days. The young are born in a
nest. At birth they are hairless and their eyes are closed.
The young ones in the litter develop rapidly, growing hair
11

`
within a week (Sullivan 2002). Between 9 and 14 days, their
eyes open and they begin to explore for food and move about
near their nest. In the third week, they begin to take solid
food.
The house rat has been implicated in the transmission of
a number of diseases to humans and domestic animals (Gratz
1997). Examples include murine typhus (rickettsial disease),
various spirochetal diseases (lyme disease, leptospirosis) and
some protozoal diseases (leishmaniasis, toxoplasmosis). Of
these, leptospirosis is of special concern as an emergent
infectious disease, with a recent upsurge in the rate of
diagnosis of this previously ‘hidden’ disease (Singleton and
Petch 1994). Commensal house rat populations have been found to
carry heavy leptospira infections in widespread regions of
Africa, Asia, Europe and North America (Gratz 1997). These are
also suspected in the transfer of ectoparasites from one place
to another. The role of R. rattus as an agent of disease is most
spectacularly illustrated through the example of the great
plague pandemics. House rats or possibly just their fleas,
harbouring in cloth and other trade goods, first carried the
plague organism (Yersinia pestis) from Central Asia to the Middle
East and Europe in the 5th century AD. Shipping routes
provided even more effective means of transporting plague
bearing rats around the world, leading to the infamous Black
Death of the 14th century. Plague remains endemic in Central
and Southeast Asia, many parts of Africa and South America and
across much of the United States of America. In each area,
rodents serve as the primary or enzootic hosts (Biggins and
Kosoy 2001), with the pathogenic agent maintaining a sensitive
balance between infection rate and death rate in order to
12

`
persist indefinitely in the wild population. During the latest
resurgence of plague in India in 1994, about 4000 persons
suffered from infection and about 100 persons died and the
resulting panic led to tremendous loss of agricultural and
industrial production (Ramalingaswami 1994).
In Hawaii, North and South America and in Africa, house
rats are seen damaging ripening sugarcane crops (Taylor 1984,
Fiedler 1988, Prakash and Mathur 1988, Wood 1994). Coconuts
grown commercially in many tropical areas are subject to
damage particularly by R. rattus and R. tanezumi (Tobin et al 1990).
These species climb palms of all ages, primarily to feed on
developing nuts, which then fall prematurely on the ground
(Fiedler et al 1982, Wood 1994). The proportion of nuts that drop
prematurely due to rat damage can be quite high. However,
impacts on yield may not be proportional to the number of
developing coconuts that fall to the ground (Williams 1974,
Reidinger and Libay 1981, Fiedler et al 1982). Trees in some
areas may compensate for early damage by increasing the size
and weight of remaining nuts (Tobin et al 1997). The economic
impact of this damage is thus not clear (Tobin and Richmond
1993). In situations where rats feed on coconut flowers or
damage very small nuts, yield losses may be underestimated by
counts of fallen, maturing nuts. Macadamia orchards in Hawaii
and Australia sustain extensive damage from R. rattus (Tobin 1992,
White et al 1997). Five to ten percent of developing nuts are
damaged by rats in some Hawaiian orchards. The rats feed on
the fruit and vegetative portions of many landscape and garden
plants including the bark of trees. Their feeding and gnawing
may completely girdle young trees.
13

`
The house rat poses a serious threat to poultry
operations by feeding on poultry feed, contaminating it with
their excrements, damaging eggs, attacking and killing chicks,
causing structural damage to buildings, doors, windows, feed
containers and transmitting or carrying several diseases
(Ahmad et al 1984, Khatri and Veda 1984, Parshad et al 1987, Soni
and Rana 1988, Gora et al 1995, Gomez Villafane et al 2001,
Hussain et al 2006, Chopra et al 2008a and b). Even poultry
buildings with fibre glass or polystyrene insulated walls are
vulnerable to damage (Turner 1986). In addition, they also
cause general nuisance for birds in the poultry house due to
their noise and movements. The birds may be frightened, which
results in poor performance. However, due to financial
limitations many farmers build poor quality sheds inside the
crop fields and close to villages which naturally attracts
both field and commensal rodents leading to considerable
financial losses (Chopra and Sabharwal 1992, Gora et al 1995,
Munjal 2000, Chopra et al 2008a). A high carrying capacity of
poultry farms for rodents is evident from the reports of 292
rodents/3600 sq. ft floor area (Malhi et al 1991) and 72
rodents/100m2 floor area (Ahmad et al 1992). Damage to poultry
feed is a major cause of economic loss as feed accounts for
50–75% of the operational cost of a poultry farm. By causing
frequent structural damage to wooden doors, windows and
electric cables (by gnawing) and to floor and foundations (by
burrowing) the rats increase the maintenance costs
of the building.
2.2 Management methods
Different methods of commensal rodent management include
environmental sanitation, habitat manipulation, rodent-
14

`
proofing, use of chemosterilants, ultrasonic and
electromagnetic devices, traps, repellents and rodenticides
(Greaves and Rowe 1969, Kassa and Jackson 1984, Buckle 1994,
Smith 1994, Prakash et al 2003, Tobin and Fall 2006, Singla and
Parshad 2000 and 2009, Singla and Mittal 2012, Singla and Garg
2013, Singla et al 2008b and 2013). There has been heavy
reliance on rodenticides to control rat populations, although
other methods such as traps and exclusion are also used (Timm
1994). Traditional baiting or trapping on the ground or floor
may intercept very few house rats unless bait and/or traps are
placed at the very points that rats traverse from and to a
food resource. House rats have a strong tendency to avoid new
objects in their environment and this neophobia can influence
control efforts, for it may take several days before they will
approach a bait station or trap. Neophobia is more pronounced
in house rats than in Norway rats (Marsh 1994). The use of
rodenticides, in the form of poison bait is inexpensive and
effective. The success of this programme is, however, based on
the type of poisons and their formulations (Prakash and Mathur
1992). Baiting technique if appropriately applied is the most
reliable strategy to control rodent pests. Behavior modifying
components may play a significant role in developing the most
attractive baits. The criteria for food selection in rats are
complex and may depend upon many factors including
palatability (Young 1946). Taste of food plays a significant
role in food preference. According to Marsh (1986) taste often
supersedes any earlier influence of odour in food selection to
a degree that is not paralleled in humans.
2.2.1 Attractancts for rodenticide baits
15

`
Rodent control with poison baits in crop lands is often
ineffective in reducing rodent densities due to several
factors. Poor bait acceptance, sub-lethal dosing and dietary
preferences can reduce the efficacy of rodenticide baiting
(Berdoy and MacDonald 1991). Although zinc phosphide has been
reported to be an effective acute rodenticide (Matschke et al
1982, Sterner et al 1996), numerous researchers have reported
bait acceptance problems related to bitter taste, sub-lethal
toxicosis and subsequent conditioned aversion after rodents
have ingested minimal levels of zinc phosphide bait (Reidinger
and Mason 1983, Prakash and Ghosh 1992, Reidinger 1997). Bait-
shyness, induced through conditioned taste aversion, can last
more than a year, even when the zinc phosphide has been
removed from the baits (Shepherd and Inglis 1993). It has been
established that field as well as commensal rodents develop
poison aversion and bait shyness after a single exposure to
the widely used rodenticide, zinc phosphide (Prakash and Jain
1971, Prakash et al 1975, Bhardwaj and Khan 1979, Saxena and
Mathur 1995). The persistence of shyness in rodents which are
not pre-baited prior to poison baiting is of a higher
magnitude than in those which are exposed to the toxicant
after prebaiting (Bhardwaj and Prakash 1982). The development
of poison aversion and bait-shyness among rodents of economic
importance makes the control of any residual population
surviving a zinc phosphide baiting operation a difficult task,
since bait carrying the same poison on the second day is not
accepted by them (Bhardwaj and Prakash 1979). Sampling of food
is well known in R. rattus (Barnett 1966), R. norvegicus (Cowan 1977,
Barnett et al 1978) and B. bengalensis (Parshad and Jindal 1991).
Even in natural habitats, this sampling behaviour has survival
16

`
value as it enables the rats not only in finding new sources
of food but also in avoiding toxic baits (Barnett 1966,
Siddiqui and Khan 1982, Chopra et al 1984 and 2009).
Several researchers have noted the need for an attractant
odour that could be added to zinc phosphide baits to
successfully compete with the alternate field crop foods
(Koehler et al 1994, Reidinger 1997, Khan et al 2000). A more
rapid consumption rate for rodenticide baits may also lead to
a lessened need to leave baits exposed to non-target species
over extended time periods (Watkins et al 1999).
2.2.1.1 Pheromones as attractants
Pheromones are known to play an important role in the
social behaviour of mammals like attraction to opposite sex,
aggregation, parental care, individual identification,
territorial marking etc. The major source of chemical cues
involved for pheromonal communication are the secretions of
the specialized scent glands, urine and feces. In rats, the
preputial glands and cheek glands are reported to be important
as they stimulate a variety of social functions. Preputial
glands, in particular, appear to play an important role in
production of olfactory substances which attract the opposite
partner. When faced with a choice among several novel foods,
naïve (observer) rats choose novel foods that have previously
been ingested by conspecifics (demonstrators) with whom they
have interacted (Strupp and Levitsky 1984). This socially
mediated transfer of food preference has been observed even
when demonstrators are anesthetized and wire-mesh barriers are
placed between demonstrator and observer (Galef and Wigmor
1983). Such findings and results of other experiments (Galef et
17

`
al 1985) suggest that diet preference is mediated in part by the
volatile chemical cues.
Researchers have shown that rodents will respond to the
biologically-derived odors of conspecifics, but the responses
are variable depending on the age, sex, social dominance and
breeding condition of the animal (Salmon and Marsh 1989,
Drickamer 1997). Consequently, in terms of management tools,
we cannot expect rodents to respond reflexively and
consistently to pheromones as do some insects (Howard 1988).
One of the few odours that some researchers have found to be
attractive to rodents is carbon disulphide (Galef et al 1988,
Shumake et al 2002), however, other researchers did not find
this material to be particularly attractive (Koehler et al
1994). Carbon disulphide present in the rodent breath is
responsible for inducing diet preferences in rodents (Galef et
al 1988). The addition of carbon disulphide to a bait has also
been shown to increase bait consumption and time spent at that
site (Bean et al 1988). Carbon disulphide is therefore a
biologically meaningful odour to rodents that increases
attractiveness of foods. However, despite the publication of
these results, carbon disulphide was never used in commercial
baits. This may in part be due to its volatility and/or
toxicity. Although environmental marking by animals generally
relays social, sexual, or reproductive status, it has also
been shown that scent marking by Norway rats may play an
additional function in communicating food preferences.
Excretory deposits that surround food sites render those
sites, and the food, more attractive to Norway rats than
unmarked sites (Laland and Plotkin 1991). Further manipulation
18

`
of this behaviour at a bait station may not lure a rat to the
site but, should assist in bait uptake.
Conspecific urine odour is reported to improve poison
bait acceptance by reducing neophobia and masking poison
aversion and bait shyness behaviour in Bandicota bengalensis,
Meriones hurrianae and Tatera indica (Kumari and Prakash 1980 and 1988,
Kaur and Parshad 2006a). Scent gland secretions (Selvaraj and
Archunan 2002) and urine (Soni and Prakash 1987) have been
reported to overcome poison bait shyness to some extent in
laboratory rats and soft furred field rats, respectively.
The sex pheromone which is found in female rodent urine
or vaginal discharge could be an important substance for
attracting males to traps or to stations with toxic bait or
chemosterilants. An attempt to evaluate this potential
possibility has been made by Field (1971) who reported
increased acceptability of chemosterilant bait by rats on
treatment with estrous urine. However, more research is needed
to quantify the attractant capacity of these pheromones, and
to isolate and identify the chemical components that are
responsible for this presumed attraction. Odour plays a
considerable part in sexual activity and sex lures have been
used successfully in insect control. Proprietary rodent baits
are available “with added sex attractant" but the sex for
which the attractant is designed is not specified. The use of
sex attractants has considerable appeal to the public but
their use in rodent control has yet to be substantiated (Bull
1972).
2.2.1.2 Poultry egg components as bait additives
Pervez et al (2000) reported 3% egg yolk to be the
preferred bait additive over the yeast powder, minced meat and
19

`
fish meal, for controlling short tailed mole-rat, Nesokia indica
in laboratory studies. Shafi et al (1990) reported the
preference of egg yolk over egg shell powder, against the wild
caught R. rattus in Pakistan, while Pervez et al (1999) suggested
preference of egg shell powder over egg yolk by field rodents
(B. bengalensis, N. indica, Millardia meltada and Mus spp.). The egg, when
used as additive (2%), allowed a higher consumption of poison
bait for a successful control (Pervez et al 2005) and 3% egg
additive could mask the undesirable characters of the bait
material, rendering it palatable to commensal R. rattus and field
rodents (B. bengalensis and N. indica) (Pervez 2007). Abbas (2003),
however, reported preference of rats towards peanut butter
than egg shell powder as additives to bait in B. bengalensis in
wheat crop.
Shafi et al (1990) recommended that 2% egg yolk as additive
in poison bait can be useful in enhancing bait acceptance by R.
rattus in an effort to reduce its infestation in various
situations. It increased acceptability by 72.7 and 67.9% of
the bait containing brodifacoum and bromadiolone, respectively
against reference poison bait. Khan et al (2000) reported
preference for poison bait containing egg shells by rodents in
rice crop fields. Two second generation anticoagulants i.e.
brodifacoum and bromadiolone and one acute rodenticide i.e. zinc
phosphide each mixed with whole egg were tested in three
separate blocks of wheat crop (Pervez et al 2005). High intake
of egg mixed zinc phosphide bait by rodents was reported by
Prakash (1976). The improved formulation of zinc phosphide may
benefit the farming community as it still remains one of the
most widely used rodenticides in the subcontinent. Shafi et al
(1992b) reported highest intake of brodifacoum mixed with egg
20

`
shell by field rodents of wheat, over other additive baits.
Similar pattern of bait intake was recorded in bromadiolone
and zinc phosphide baits.
Khan et al (2000) conducted field trials in rice crop to
control rodent populations by improving poison bait acceptance
using the additives viz., minced meat, egg-yolk, egg-shell and
yeast. Poison baits, brodifacoum (0.005%), bromadiolone
(0.005%) and zinc phosphide (2%) were employed as
rodenticides. Egg-shell ranked first in the preference over
the other additives and non-additive poison baits. Mixing of
additive with brodifacoum reduced the population of M. musculus
to 92.4%, with bromadiolone to 89.7% and with zinc phosphide
to 94.7%. Similarly, for B. bengalensis, a reduction of 91.5% with
brodifacoum, 92.0% with bromadiolone and 94.7% with zinc
phosphide was recorded, while for both N. indica and M. meltada,
the respective decrease of 87.5, 90.5, 94.2, and 88.7, 85.7
and 94.7% was reported. Two baitings, one at flowering and
other at maturity stage of the crop reduced damage to 94.0%
with brodifacoum, 92.0% with bromadiolone and 91.5% with zinc
phosphide. It was concluded that the two baitings, one at the
flowering stage, and the other at maturity stage may be
employed to obtain a robust production.
It was suggested that the texture of the egg shell marks
an attractive taste for the rodents. For the zinc phosphide,
more preference for the egg shell bait than the non-additive
bait was significant as fundamentally, zinc phosphide causes an
immense "bait-shyness" and poison aversion (Prakash 1976). It
was hypothesized that an improved formulation using zinc
phosphide and egg shell may prove beneficial to the farmers as
it is one of the widely employed acute rodenticide against the
21

`
control of rodent species. However, with lack of expertise, its
grievous consequences cannot be done away with readily.
Addition of egg shell and fish meal did not show a significant
increase in relative consumption for the cereal base by B.
bengalensis (Naeem et al 2011). Rather they observed an inverse
trend indicating decreased preference for plain millet based
bait when mixed with egg shell and fish meal. Shafi et al (1993)
observed high preference toward minced meat by B. bengalensis than
egg shell and egg yolk mixed bait.
2.2.1.3 Other bait additives as attractants
Meehan (1984) cautioned that it is impossible to say
which particular foodstuff will be preferred by individual
rats or even whole populations. There is no such thing as
‘universally acceptable bait’. Rowe (1973) made a similar
comment about mice. These statements have been confirmed by
studies of consumption of foods by wild rats (Clark 1982).
Lund (2008) gave a general summary of the ideal
characteristics of rodent baits and additives. No magical
additives that made baits irresistible were identified. Marsh
(1988) summarised that sugars and vegetable oils and animal
fats are the most universally effective additives for cereal
baits to improve acceptance and palatability. Flavour
additives to bait have often decreased rather than increased
consumption.
A variety of methods were used to control rodent
populations and to reduce their damage (Witmer et al 1995).
Effective odour attractants would allow managers to attract
rodents to remote detection devices, bait stations, or traps.
The bait stations, in turn, might have oral baits containing
rodenticides or fertility control materials. Witmer et al (2008)
22

`
tested 3 odor attractants i.e. almond, lemon and ginger, that
appeared to have potential for effectively attracting Norway
rats but, none of the 3 potential attractants were found
effective. Wines (2004) examined the efficacy of liquid
concentrated banana extract as a potential attractant for
trained Gambian rats.
Bullard and Shumake (1977) used the responses of
Philippine rice field rats (R. rattus mindanensis) to show that
intensifying the flavour cues of familiar or favoured foods
could be a useful way of enhancing bait intake. Such
attractants help to keep rodents feeding longer at food
sources. They may be effective because they mask the taste of
a rodenticide and/or they are palatable in their own right
(Meehan 1984). Sugar is a well-known effective additive for
rats and mice, while bitter flavours tend to be rejected (Rowe
1961, Howard et al 1972, Shimizu et al 1980, Marsh 1988, Yamaguchi
1995). Norway rats in trials preferred salty and sweet tastes
over sour and bitter ones (Karasawa and Muto 1978, Kolody et al
1993). Singla et al (2010) studies the efficacy of sodium
chloride in improving acceptance of bait by rodents. The most
attractive concentrations of sugars have also been assessed in
various trials (Collier and Bolles 1968). When given the
choice between sugar on its own and flour, Norway rats
consumed only small amounts of the sugar (Barnett 1956).
Inverted sugars like maltose, dextrose, fructose and levulose
also are acceptable to rodents (Smythe 1976). Of the
artificial sweeteners, saccharin was favoured by laboratory
rats over cyclamate, while wild Norway and house rats
preferred glucose to saccharin (Wagner 1971a and b). Sugars
also help preserve the baits, potentially increasing shelf and
23

`
field life, but also increase bait palatability to
invertebrates and reptiles. Babbar et al (2009) found higher
acceptance of zinc phosphide bait containing 2% sugar by R.
rattus. Mice in a study exhibited a preference for monosodium
glutamate and sodium chloride (common salt), but more so for
sugars (Yamaguchi 1995). Salt and monosodium glutamate are
variable in their effect, and were disliked at concentrations
above 0.5% (Ohara and Naim 1977, Marsh 1986 and 1988, Kolody et
al 1993). Female rats consumed more salt than male rats (Flynn
et al 1993).
Oils can be effective bait additives. Meehan (1984) found
that the higher was the level of oil in bait, the more readily
it was taken. Mice have exhibited a preference for high-fat
foods (Imiazumi et al 2001) and rats have an appetite for
dietary fats and oils (Elizalde and Sclafani 1990, Ramirez
1993). Glycerine, corn oil, arachis oil and mineral oil were
more palatable to mice than olive, linseed or cod-liver oils
(Rowe et al 1974). Groundnut oil probably has a neutral flavour
to rodents (neither attractive nor repellent), but may act to
mask the flavour of cereals to which rats have developed bait
shyness (Bhardwaj and Khan 1979). In conjunction with gur,
groundnut oil increased bait consumption by mice more than
just the gur alone (Pathak and Saxena 1995). Olive oil
enhanced the acceptance of baits to male house rats in no-
choice tests, while soybean oil, sunflower oil, mustard oil,
groundnut oil and coconut oils were all preferred over plain
bait in choice tests, and groundnut oil was preferred over
coconut and mustard oils (Ahmad et al 1994). However, in the
same study, female rats preferred coconut, mustard and to some
extent, sunflower and soybean oil in mixed diets. In a study
24

`
in the USA, coconut, peanut and corn oils were most preferred
by Norway rats, whilst corn oil was preferred over peanut oil
by house rats (Meehan 1984).
In Australia, fishmeal was a successful additive to bait
for mice (Robards and Saunders 1998, Jacob et al 2003). These
authors listed a range of other additives such as chocolate,
cheese, aniseed, peanut oil and honey, that made no difference
to consumption. By contrast, chocolate is used elsewhere as an
effective trap attractant for mice to such an extent that UK
researchers have developed a chocolate mouse trap (Singh
2003). The addition of 2% onion pulp improved consumption of
bajra flour by mice in India (Saxena et al 1995). Toasted
coconut has been used successfully as bait in traps in
Rarotonga (Robertson et al 1998). Tabuchi et al (1991) listed
black pepper, milk and coffee as highly preferred food-related
odours, while, nut, peppermint, plum, orange and cheese also
elicited a bar-pressing response from rats. Raw fish and beef,
dried dog food, coconut oil, fresh or dried blood, chicken
offal, cinnamaldehyde, raspberry, aniseed and other commercial
products have all been attributed with attractive properties
for Norway rats (Bull 1972). McFadden (1984) listed aniseed,
banana, coconut, clove, eucalyptus and vanilla as acceptable
lures for Kiore. Schisla et al (1970) described the success of
‘Dexide’ - a carbohydrate with flavour material that increased
the consumption and efficacy of warfarin to mice, house rats
and Norway rats. Witmer et al (2008) reported almond, ginger and
lemon extract as potential attractants against Norway rats,
but they were not effective under field conditions. Kamal and
Hossain (2003) reported paddy grain as the most preferred bait
by rats in rice field, while dry fish and coconut was the
25

`
second and third preferred bait, respectively. The comparison
of average daily intake of rats between foods with and without
additives (2% sugar and 2% groundnut oil) revealed that the
rats showed significant preference for food items with
additives over their plain alternatives in most of the cases.
This preference for foods with additives was maintained over
different days of exposure. Taste of food has already been
reported to influence feeding response of R. rattus both in
laboratory conditions and poultry farms (Khan 1974, Prakash et
al 1980, Munjal 2000).
The preference of rats for a particular food depends more
upon its flavour than its nutritional value (Naganuma et al
1973). Additives like sugar and vegetable oils at 1 to 3%
concentration increase the food intake of rodents as they make
foods more acceptable and palatable to rats (Brooks and LaVoie
1990, Malhi and Kaur 1995). Moreover, selection of sweet food
has also an adaptive value as in natural food. Bitter taste is
associated with occurrence of toxic alkaloids or poisonous
foods while sweetness indicates presence of starch and
carbohydrates and the latter form the major component of diet
of rodents (Barnett and Prakash 1975, Rana et al 1992). Based on
overall mean daily food consumption, different foods were
eaten by R. rattus in the preference order of cracked maize >
cracked wheat > wheat flour > poultry feed > fish meal. Maize
is a major constituent of the poultry feed and (Bhardwaj 1983)
reported high preference for cracked maize than poultry feed
by R. rattus. This contradicts previous report that poultry feed
may be used as bait in poultry farms. Interestingly, cereals
and pulses have been recorded with more consumption than that
of poultry feed in Punjab and Rajasthan (Parshad et al 1987 and
26

`
1991, Mathur et al 1992). Thus, high preference of rats for
cracked maize than poultry feed appeared to relate to its
texture and taste qualities.
Naeem et al (2011) reported that dietary preferences of B.
bengalensis depend upon many factors including particle size,
palatability, taste, flavor and nutritious value of bait base
and the taste enhancers. Experiments performed to evaluate
inclination of the rodents towards cereal bases revealed that
millet was the most preferred over the other cereals. The
cereal bait bases preferred by the B. bengalensis were in the
order of Millet > wheat > maize > rice. The use of 2% butter
oil, however, greatly enhanced intake of cracked millet.
Jackson (1965) revealed that texture, odour and taste play a
significant role in selection of a particular bait base.
Preference for millet as compared to other cereals is also
reported by Parshad and Jindal (1991) who enunciated that B.
bengalensis prefers soft and small sized grains. Watson and Perry
(1954) found that millet was preferred by N. indica due to its
small size. Similar inferences were also drawn by Prakash et al
(1970) and Kumar and Khan (1978). Shafi (1991) argued that
particle size of grain plays an important role in enhancing
bait acceptability. Use of poison baits is still the most
reliable strategy for controlling field as well as commensal
rodents. However, baiting techniques should be modified
according to the psychological characteristics like neophobia,
bait shyness as well as feeding behaviour, exploratory
behaviour, transporting, hoarding and territoriality behaviour
of the target species (Lund 2008).
2.2.2 Bait attractants for traps
27

`
Trapping rodents in fields and premises is a common old
practice (Fitzwater and Prakash 1989). Two basic types of
traps are being used, the snap or kill trap and the live trap.
Among the snap traps, the Tanjore bow trap, a low cost bamboo
trap generally used by professional trappers in rice fields
(Neelanarayanan et al 1995); the wooden snap trap, a locally
fabricated trap using timber splinters (Srihari and
Chakravarthy 1992); the urang or arrow trap (Prakash and Mathur
1987) and the break-back spring loaded snap traps with wooden
or jawed iron base (Prakash and Mathur 1987) have been used
traditionally. Among these the break-back snap trap is most
popular. Glue boards are also effective for trapping indoor
rodents (Srivastava and Srivastava 1985). However, these have
not become popular so far because of the cost, hygiene
problems and inhumane method of killing.
Trapping efficiency is dependent on trap type chosen
(Wiener and Smith 1972), type of bait and pre-baiting method
(Chitty and Kempson 1949), social interaction between the
individuals (Ylenon et al 1990) and on effects of recent
trapping efforts. The later includes the effect of odour of
previously trapped conspecifics (Krebs and Boonstra 1984),
competitors or predators (Stodddart 1982) and the previous
experience of being caught in a trap (Tanaka 1963). Alam et al
(2007) evaluated six different baits for trapping rodents in
the poultry farms. Among them the preferred order was dry
fish, coconut meat, potato, bread, soap and wax. Hasanuzzaman
et al (2009) evaluated the effectiveness of bread, potato,
coconut meat, dry fish and paddy grains in trapping B. bengalensis
at different stages of wheat crop. Witmer et al (2010) tested 15
attractants for use in traps for capturing or detecting
28

`
Gambian rats. They found conspecific scents from faeces and
urine as the best attractants for single and paired Gambian
rats. They suggested that peanut butter, anise, ginger and
fatty acid scents can also be used as attractants in traps.
Kaur and Parshad (2006b) reported the efficacy of conspecific
urine in enhancing trapping of rodents. Kok et al (2013) used
different baits in traps for surveying small mammals at high
altitude in South Africa and concluded that the most suitable
bait is peanut butter and oats.
CHAPTER – III
MATERIAL AND METHODS
The present study was carried out at Animal House
Laboratory and Rodent Research Laboratory, Department of
Zoology, Punjab Agricultural University, Ludhiana and poultry
farms situated in the campus, Guru Angad Dev Veterinary and
Animal Science University (GADVASU), Ludhiana and village
Ghutani Kalan, District Ludhiana. Approval of the
Institutional Animal Ethics Committee was obtained for the use
of animals.
3.1 Laboratory experiments
3.1.1 Collection and maintenance of animals
For present studies, the house rat, R. rattus of both sexes
were trapped with the help of single catch and multi catch rat
traps from store houses and poultry farms in and around
Ludhiana. In laboratory, rats were acclimatized individually
in cages of size 36 x 23 x 23cm (Plate 1a) for 10-15 days
before the commencement of experiment with food and water
provided ad libitum. Food consisted of preferred cereal based
29

`
bait containing loose mixture of cracked wheat, powdered sugar
and groundnut oil (WSO bait) in ratio 96: 2: 2. Proper
hygienic conditions were maintained during the period of
experimentation.
3.1.2 Bait additives used
During present studies, poultry egg components such as
egg shell powder (ESP), egg albumin (EA) and crushed egg
shells (CES) were used as bait additives.
3.1.3 Experimental design of bi-choice feeding tests
During experimentation, mature and healthy rats of both
sexes were weighed and divided into four groups of ten rats
each (5 of each sex in each group). In first experiment, rats
of groups I, II and III were exposed to WSO bait containing 2,
5 and 10% ESP, respectively as additive in bi-choice with WSO
bait for 5 days. Rats of group IV kept as untreated control
were fed on WSO bait without additive. There was no
significant difference in average body weight of rats in
treated and untreated groups. Bait was kept in bowls and water
was provided ad libitum. The position of bowls was changed daily
to avoid any difference in consumption due to site preference.
In second experiment, rats of groups I, II and III were
exposed to WSO bait containing 2, 5 and 10% EA as bait
additive, respectively, whereas in experiment third, similar
groups of rats were exposed to WSO bait containing 2, 5 and
10% CES as bait additive, respectively. In all the three
experiments, rats of group IV were kept as untreated control
and were fed on WSO bait without additive. Before and after
the treatment period, all the rats were fed on WSO bait.
30

`
3.1.3.1 Bait acceptance
In all the three experiments, bait consumption was
recorded after every 24 h and every time, bait was replaced to
the original 30g. Before weighing, the bait of all the treated
and untreated rats was cleared of faecal pellets and dried if
needed. Mean daily consumption of food (g/100g bw) was
calculated separately for each group of rats during pre-
treatment, treatment and post-treatment periods. The percent
acceptance of treated bait over WSO bait consumed during
treatment period was determined as per the formula given
below:
Percentacceptance =
Consumption of WSO bait with additiveduring treatment period x
100Total bait consumed during treatment period
3.1.4 Experimental design of no-choice feeding tests
Four rats of each sex were exposed to WSO bait containing
bait additives in no-choice feeding test under Food Scale
Consumption Monitor (FSCM) (Columbus Instruments, USA) (Plate
1b). Each rat was kept in a cage provided with a feeding bowl
and a feeding sensor below. In first experiment, four female
rats were first exposed to WSO bait for four days. Every day
rats were exposed to bait for two hours. Before exposure to
baits, rats were kept without food overnight. Subsequently,
these rats were exposed to WSO bait containing 2%
concentration each of ESP, EA and CES. WSO bait containing
each type of bait additive was exposed to rats for four days.
Behaviour of rats was recorded in the form of food consumption
(g/kg bw), total number of feeding bouts made and total time
spent (s) in bouting in 2 hr a day by means of software of
31

`
FSCM loaded in the computer. Similar experiment was carried
out on male rats also.
3.2 Experiments in poultry farms
3.2.1 Bait additives in rodenticide baits
Three blocks (I, II and III) of poultry farm, with each
block further consisting of four replicated sheds were
selected at village Ghutani Kalan, District Ludhiana. One
block (IV) consisting of four sheds selected at the campus,
GADVASU, Ludhiana was kept as untreated control. Before
treatment, pre-census bait consumption was recorded from all
the sheds of four blocks by keeping WSO bait @ 500g/shed in
two rows of ten bait points each to record level of rodent
activity before treatment. After pre-census bait consumption,
sheds of blocks I, II and III were treated with 2% zinc
phosphide bait prepared in WSO without any bait additive, WSO
containing 2% egg albumin and WSO containing 2% egg shell
powder, respectively. Treatment bait was also kept @ 500g/shed
in two rows of ten bait points each. The treatment bait was
exposed to rats for two days after which the remaining bait
was collected and weighed to record the consumption (g/100g
bait) of different types of treatment baits. After one week of
treatment, post census bait consumption was recorded from all
the poultry sheds of each and every block by the same method
as used during pre-census. Reduction in level of rodent
activity (%) with respect to untreated control sheds was
determined as per the formula of Henderson and Tilton (1955)
which is as follows:
Percent reduction in rodent activity = 100 {1−t2 x r1t1 x r2 }
Where, t1 and t2 represent pre- and post-treatment bait
32

`
consumptions, respectively in treated sheds and r1 and r2
represent pre- and post-treatment bait consumptions,
respectively in untreated reference sheds.
Reduction in level of rodent activity (%) was also
determined with respect to same shed as per the formula given
below:
Pre-census bait consumption – Post-censusbait consumption x 100
Pre-census bait consumption
3.2.2 Bait additives in traps
Six poultry sheds were selected at the poultry farm
located in the campus of GADVASU, Ludhiana. All the sheds were
predominantly infested with R. rattus. Rodent trapping was
carried out in all the sheds by placing eight single rat traps
in each shed. Each trap was baited with a piece of chapatti.
The effect of 2% poultry egg albumin and 2% egg shell powder
in enhancing rodent trapping was studied by smearing these
components on the chapatti pieces which were then used for
baiting the traps. In first experiment, trapping was carried
out from all the sheds by alternatively placing traps
containing chapatti pieces without additive and those
containing chapatti pieces smeared with 2% egg shell powder.
In the second experiment, trapping was carried out from all
the sheds by alternatively placing traps containing chapatti
pieces without additive and those containing chapatti pieces
smeared with 2% egg albumin. In the third experiment, trapping
was carried out from all the sheds by alternatively placing
traps containing chapatti pieces smeared with 2% egg shell
powder and those containing chapatti pieces smeared with 2%
33

`
egg albumin. Every time, trapping was carried out for one week
and the number of rats trapped of each sex was recorded. From
the data on number of rats trapped of each sex, trap index in
the form of total number of rats trapped, number of male rats
trapped and number of female rats trapped per 100 trap nights
per day were calculated.
3.3 Statistical analyses
All values were expressed as mean ± SE. The data on food
consumption for two sexes, three bait additives, three
concentrations of each bait additive, five days of treatment
and five replicated rats was collected using factorial
experiments in Completely Randomized Design. Analysis was done
using General Linear Model in SAS 9.3. All pair wise treatment
comparisons were made using Tukeys’ HSD test at 5% level of
significance.
34

CHAPTER – IV
RESULTS AND DISCUSSION
Present study on evaluation of poultry components (egg
shell and egg albumin) as bait additives for increasing the
acceptance of cereal based bait was carried out in no-choice
and bi-choice feeding tests in laboratory. The effective
concentrations of these components were then mixed in 2% zinc
phosphide bait and tested in poultry farms to evaluate their
acceptance and efficacy in comparison to 2% zinc phosphide
bait without additive. The effect of these poultry components
was also studied on rodent trapping. The results obtained are
presented herewith.
4.1 Laboratory experiments
4.1.1 Bi-choice experiments
In bi-choice laboratory experiments, rats of both sexes
were exposed to WSO bait containing 2, 5 and 10%
concentrations each of ESP, EA and CES. Results revealed no
significant difference in mean daily consumption (g/100g bw)
of WSO bait during pre- and post-treatment periods among all
the treated and untreated groups of rats of both sexes in all
the three experiments using ESP, EA and CES.
4.1.1.1 Acceptance of egg shell powder (ESP) as bait
additive
During treatment period, significantly (P ≤ 0.05) higher
mean daily consumption (g/100g bw) of WSO bait containing 2%
(group I) and 5% (group II) ESP from that of WSO bait alone
was observed in female rats. There was no significant
difference in consumption between WSO bait without additive
and that containing 10% ESP by female rats of treated group

`
III (Table 1, Figure 1). Also there was no significant
difference in mean daily consumption of WSO bait containing
egg shell powder among the three treated groups I, II and III.
Percent acceptance of WSO bait with additive over WSO bait
alone was also not found to differ significantly among the
three treated groups in female rats (Figure 10). The percent
acceptance was found to be 67.21, 58.04 and 49.18%,
respectively in rats of groups I, II and III. Apparently,
there was a decrease in acceptance of bait with increasing
concentration of the bait additive in female rats (Table 1).
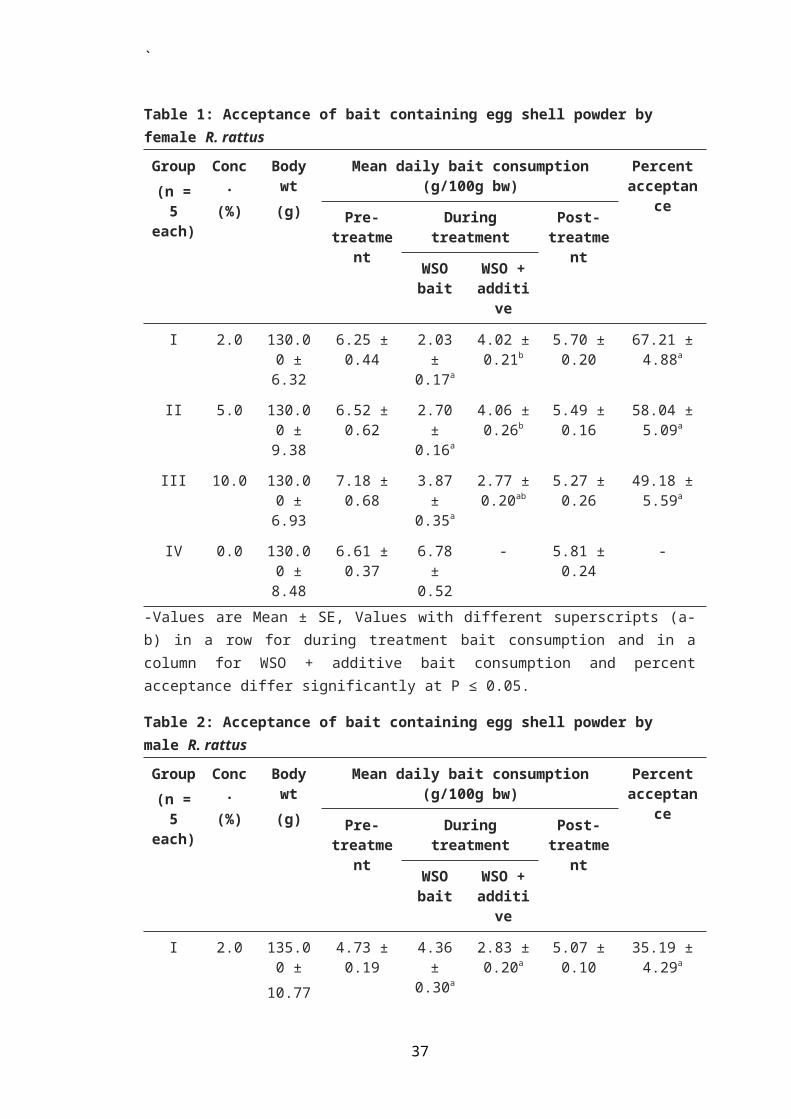
In male rat, there was no significant difference observed
in mean daily consumption of WSO bait containing bait additive
and that without any additive at all the concentrations of egg
shell powder tested (Table 2, Figure 2). Also there was no
significant difference in mean daily consumption of WSO bait
containing egg shell powder among the three treated groups I,
II and III. Percent acceptance of WSO bait with additive over
WSO bait alone was also not found to differ significantly
among the three treated groups (Figure 11). The percent
acceptance was found to be 35.19, 41.20 and 47.32%,
respectively in rats of groups I, II and III. Apparently,
there was an increase in acceptance of bait with increasing
concentration of the bait additive in male rats (Table 2).
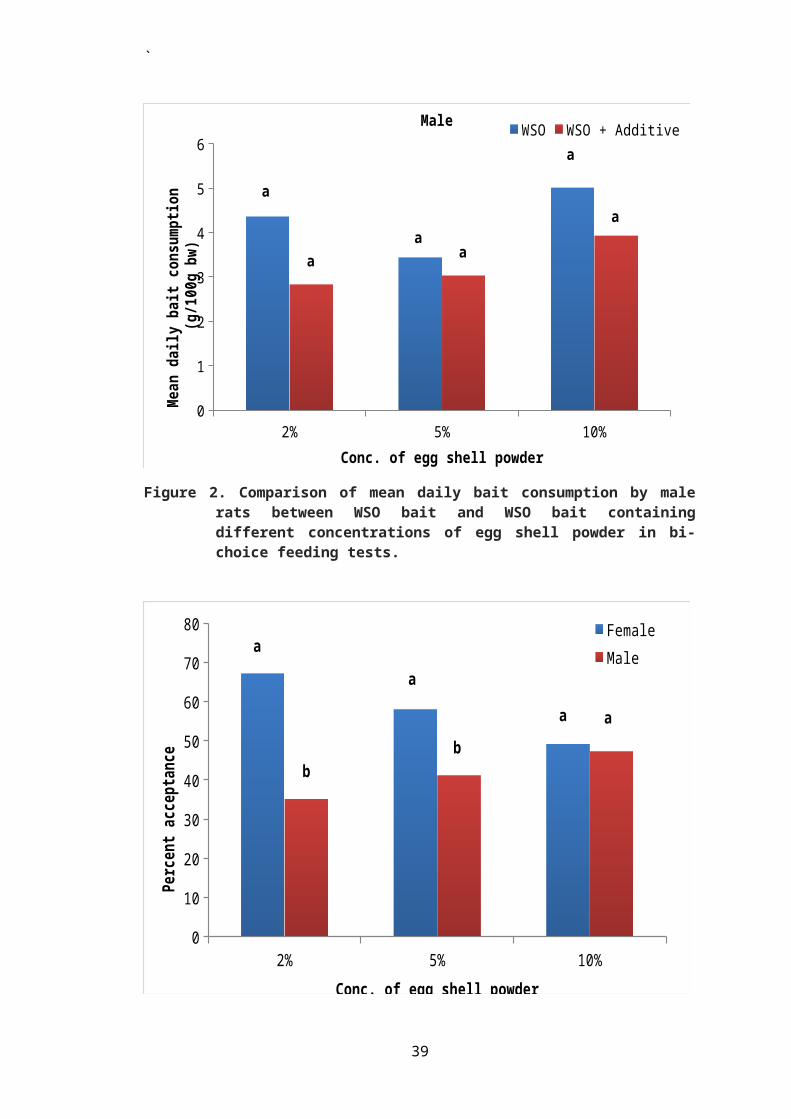
Percent acceptance of WSO bait containing ESP was
generally low in male rats as compared to female rats. This
difference in acceptance of bait between male and female rats
was found to be statistically significant (P ≤ 0.05) when rats
were exposed to WSO bait containing 2 and 5% ESP in choice
with WSO bait without additive (Figure 3).
36
`
Table 1: Acceptance of bait containing egg shell powder by female R. rattus
Group(n =5
each)
Conc.
(%)
Bodywt(g)
Mean daily bait consumption(g/100g bw)
Percentacceptan
cePre-treatme
nt
Duringtreatment
Post-treatme
ntWSObait
WSO +additive
I 2.0 130.00 ±6.32
6.25 ±0.44
2.03±
0.17a
4.02 ±0.21b
5.70 ±0.20
67.21 ±4.88a
II 5.0 130.00 ±9.38
6.52 ±0.62
2.70±
0.16a
4.06 ±0.26b
5.49 ±0.16
58.04 ±5.09a
III 10.0 130.00 ±6.93
7.18 ±0.68
3.87±
0.35a
2.77 ±0.20ab
5.27 ±0.26
49.18 ±5.59a
IV 0.0 130.00 ±8.48
6.61 ±0.37
6.78±
0.52
- 5.81 ±0.24
-
-Values are Mean ± SE, Values with different superscripts (a-b) in a row for during treatment bait consumption and in acolumn for WSO + additive bait consumption and percentacceptance differ significantly at P ≤ 0.05.
Table 2: Acceptance of bait containing egg shell powder by male R. rattus
Group(n =5
each)
Conc.
(%)
Bodywt(g)
Mean daily bait consumption(g/100g bw)
Percentacceptan
cePre-treatme
nt
Duringtreatment
Post-treatme
ntWSObait
WSO +additive
I 2.0 135.00 ±
10.77
4.73 ±0.19
4.36±
0.30a
2.83 ±0.20a
5.07 ±0.10
35.19 ±4.29a
37

`
II 5.0 133.00 ± 6.26
5.04 ±0.26
3.44±
0.17a
3.03 ±0.27a
4.94 ±0.03
41.20 ±4.39a
III 10.0 135.00 ±
10.20
5.37 ±0.43
5.01±
0.64a
3.93 ±0.27a
6.41 ±0.25
47.32 ±6.34a
IV 0.0 141.00 ± 6.39
5.76 ±0.40
5.85±
0.27
- 4.13 ±0.08
-
-Values are Mean ± SE, Values with similar superscript (a) ina row for during treatment bait consumption and in a columnfor WSO + additive bait consumption and percent acceptance donot differ significantly.
2% 5% 10%0
0.51
1.52
2.53
3.54
4.5 FemaleWSO WSO + Additive
Conc. of egg shell powder
Mean
dai
ly b
ait
cons
umpt
ion
(g/1
00g
bw)
a
b
b
a
a
a
Figure 1. Comparison of mean daily bait consumption by femalerats between WSO bait and WSO bait containingdifferent concentrations of egg shell powder in bi-choice feeding tests.
38
`
2% 5% 10%0
1
2
3
4
5
6Male WSO WSO + Additive
Conc. of egg shell powder
Mean
dai
ly b
ait
cons
umpt
ion
(g/1
00g
bw)
a
aa
a
a
a
Figure 2. Comparison of mean daily bait consumption by malerats between WSO bait and WSO bait containingdifferent concentrations of egg shell powder in bi-choice feeding tests.
2% 5% 10%0
10
20
30
40
50
60
70
80 FemaleMale
Conc. of egg shell powder
Perc
ent
acce
ptan
ce
a
b
a
b a a
39
`
Figure 3. Comparison of percent acceptance of baitcontaining different concentrations of egg shellpowder between male and female rats in bi-choicefeeding tests
Table 3: Acceptance of bait containing egg albumin by female R.
rattus
Group(n =5
each)
Conc.
(%)
Bodywt(g)
Mean daily food consumption(g/100g bw)
Percentacceptan
cePre-
treatment
Duringtreatment
Post-treatme
ntWSObait
WSO +additi
ve
I 2.0 131.00± 0.89
11.05 ±0.43
6.39 ±0.79a
10.04± 0.32b
8.92 ±0.46
67.45 ±3.83a
II 5.0 136.00±
14.31
12.77 ±1.03
7.25 ±0.45a
13.04± 0.57b
11.21 ±0.21
69.13 ±4.55a
III 10.0 134.00±
14.58
12.95 ±0.24
8.09 ±0.58a
12.71± 0.52b
8.72 ±0.38
65.00 ±3.86a
IV 0.0 127.00± 6.87
11.15 ±0.17
10.52± 0.52
- 9.39 ±0.37
-
-Values are Mean ± SE, Values with different superscripts (a-b) in a row for during treatment bait consumption and in acolumn for WSO + additive bait consumption and percentacceptance differ significantly at P ≤ 0.05.
40

`
Table 4: Acceptance of bait containing egg albumin by male R.
rattus
Group(n =5
each)
Conc.
(%)
Bodywt(g)
Mean daily food consumption(g/100g bw)
Percentacceptan
cePre-
treatment
Duringtreatment
Post-treatme
ntWSObait
WSO +additi
ve
I 2.0 130.00± 5.65
12.70 ±0.81
8.17 ±0.60a
9.98 ±0.40ab
10.61 ±0.75
57.12 ±4.36a
II 5.0 138.00±
17.06
9.58 ±0.40
5.79 ±0.32a
10.97± 0.05b
9.01 ±0.14
70.63 ±3.81b
III 10.0 132.00± 3.34
10.49 ±0.49
4.69 ±0.42a
11.57± 0.48b
7.88 ±0.11
76.22 ±4.11b
IV 0.0 123.00± 3.89
10.33 ±0.46
9.94 ±0.41
- 8.79 ±0.34
-
-Values are Mean ± SE, Values with different superscripts (a-b) in a row for during treatment bait consumption and in acolumn for WSO + additive bait consumption and percentacceptance differ significantly at P ≤ 0.05.
41

`
2% 5% 10%0
2
4
6
8
10
12
14 FemaleWSO WSO + Additive
Conc. of egg albumin
Mean
dai
ly b
ait
cons
umpt
ion
(g/1
00g
bw)
a aa
bb
b
Figure 4. Comparison of mean daily bait consumption by femalerats between WSO bait and WSO bait containingdifferent concentrations of egg albumin in bi-choicefeeding tests.
2% 5% 10%0
2
4
6
8
10
12
14 Male WSO WSO + Additive
Conc. of egg albumin
Mean
dai
ly b
ait
cons
umpt
ion
(g/1
00g
bw)
a
a
aa
bb
42

`
Figure 5. Comparison of mean daily bait consumption by malerats between WSO bait and WSO bait containingdifferent concentrations of egg albumin in bi-choicefeeding tests
2% 5% 10%0
102030405060708090
Female Male
Conc. of egg albumin
Perc
ent
acce
ptan
ce
a
a
a aa
a
Figure 6. Comparison of percent acceptance of bait containingdifferent concentrations of egg albumin between maleand female rats in bi-choice feeding tests
4.1.1.3. Acceptance of crushed egg shell (CES) as bait
additive
No significant difference in mean daily consumption
between WSO bait containing 2, 5 and 10% CES and WSO bait
without additive was observed in female rats. The consumption
of two kinds of baits was almost similar (Table 5, Figure 7).
Also there was no significant difference in mean daily
consumption of WSO bait containing CES among the three treated
groups I, II and III. Percent acceptance of WSO bait with
additive was also not found to differ significantly among the
three treated groups (Figure 10). The percent acceptance was
43

`
found to be 55.34, 54.25 and 53.62%, respectively in rats of
groups I, II and III (Table 5).
In male rats also no significant difference in mean daily
consumption between WSO bait containing 2, 5 and 10% CES and
WSO bait without additive was observed. The consumption of two
kinds of baits was almost similar (Table 6, Figure 8). Also
there was no significant difference in mean daily consumption
of WSO bait containing CES among the three treated groups I,
II and III. Percent acceptance of WSO bait with additive was
also not found to differ significantly among the three treated
groups (Figure 11). The percent acceptance was found to be
62.39, 55.35 and 58.94%, respectively in rats of groups I, II
and III (Table 6).
Percent acceptance of WSO bait containing CES by male
rats as compared to female rats was generally high at all the
three concentrations of CES tested. This difference in
acceptance of bait between male and female rats was, however,
not found to differ significantly at any of the concentrations
tested (Figure 9).
Table 5: Acceptance of bait containing crushed egg shells by female R. rattus
Group(n =5
each)
Conc.
(%)
Bodywt(g)
Mean daily food consumption(g/100g bw)
Percentacceptan
cePre-
treatment
Duringtreatment
Post-treatme
ntWSObait
WSO +additi
ve
I 2.0 143.00± 2.68
12.85 ±0.37
7.00±
0.42a
8.67 ±0.47a
11.54 ±0.29
55.34 ±6.10a
II 5.0 144.00 14.66 ± 8.28 9.67 ± 11.78 ± 54.25 ±
44

`
± 2.19 0.51 ±0.65a
0.62a 0.18 4.16a
III 10.0 142.00± 1.79
12.88 ±0.42
6.61±
0.49a
7.13 ±0.52a
10.99 ±0.36
53.62 ±3.62a
IV 0.0 146.00±
10.80
14.20 ±0.74
15.02±
0.40
- 11.09 ±0.18
-
-Values are Mean ± SE, Values with similar superscript (a) ina row for during treatment bait consumption and in a columnfor WSO + additive bait consumption and percent acceptance donot differ significantly.
Table 6: Acceptance of bait containing crushed egg shells by male R. rattus
Group(n =5
each)
Conc.
(%)
Bodywt(g)
Mean daily food consumption(g/100g bw)
Percentacceptan
cePre-
treatment
Duringtreatment
Post-treatme
ntWSObait
WSO +additi
ve
I 2.0 149.00± 5.72
11.17 ±0.63
7.62±
0.78a
9.92 ±0.30a
10.28 ±0.24
62.39 ±4.69a
II 5.0 149.00± 5.72
13.89 ±0.33
8.18±
0.64a
8.98 ±0.48a
13.84 ±0.42
55.35 ±5.07a
III 10.0 146.00± 7.79
13.12 ±0.37
7.08±
0.39a
10.46±
0.75a
11.82 ±0.39
58.94 ±3.31a
IV 0.0 144.00± 4.56
13.85 ±0.11
12.92±
0.26
- 12.36 ±0.42
-
-Values are Mean ± SE, Values with similar superscript (a) ina row for during treatment bait consumption and in a column
45

`
for WSO + additive bait consumption and percent acceptance donot differ significantly.
2% 5% 10%0
2
4
6
8
10
12 FemaleWSO WSO + Additive
Conc. of crushed egg shells
Mean
dai
ly b
ait
cons
umpt
ion
(g/1
00g
bw)
a
a aa
aa
Figure 7. Comparison of mean daily bait consumption by femalerats between WSO bait and WSO containing differentconcentrations of crushed egg shells in bi-choicefeeding tests.
46

`
2% 5% 10%0
2
4
6
8
10
12Male WSO WSO + Additive
Conc. of crushed egg shells
Mean
dai
ly b
ait
cons
umpt
ion
(g/1
00g
bw)
a
a
aa
a
a
Figure 8. Comparison of mean daily bait consumption by malerats between WSO bait and WSO bait containingdifferent concentrations of crushed egg shells in bi-choice feeding tests.
2% 5% 10%48
50
52
54
56
58
60
62
64FemaleMale
Conc. of crushed egg shells
Perc
ent
acce
ptan
ce
aa
a a aa
47

`
Figure 9. Comparison of percent acceptance of bait containingdifferent concentrations of crushed egg shells betweenmale and female rats in bi-choice feeding tests.
ESP EA CES0
10
20
30
40
50
60
70
80 Female2% 5% 10%
Perc
ent
acce
ptan
ce
aa
a
a aa
a a a
Figure 10. Comparison of percent acceptance of bait amongdifferent concentrations of egg shell powder, eggalbumin and crushed egg shells in female rats in bi-choice feeding tests.
48

`
ESP EA CES0
10
20
30
40
50
60
70
80
90 Male2% 5% 10%
Perc
ent
acce
ptan
ce
aa
aa
bb
aa a
Figure 11. Comparison of percent acceptance of bait amongdifferent concentrations of egg shell powder, eggalbumin and crushed egg shells in male rats in bi-choice feeding tests.
In female rats, the percent acceptance of WSO bait
containing 2% ESP, EA and CES was not found to differ
significantly. However, the percent acceptance of WSO bait
containing 5% EA was significantly (P ≤ 0.05) high from WSO
bait containing 5% CES and non-significantly high from WSO
bait containing 5% ESP (Figure 12). The percent acceptance of
WSO bait containing 10% EA was significantly (P ≤ 0.05) high
from WSO bait containing 10% ESP and non-significantly high
from WSO bait containing 10% CES (Figure 12). There was no
significant difference in percent acceptance of WSO bait
containing ESP and CES at all the concentrations tested.
In male rats, percent acceptance of WSO bait containing
EA and CES was found to be significantly (P ≤ 0.05) high from
that of WSO bait containing ESP at all the concentrations
49

`
tested (Figure 13). At 5 and 10% concentrations, percent
acceptance of WSO bait containing EA was comparatively more
than WSO bait containing similar concentrations of CES.
Statistically, there was no significant difference in percent
acceptance of WSO bait containing EA and CES.
In overall, the percent acceptance of WSO bait containing
2 and 5% ESP and 2, 5 and 10% EA was significantly more than
WSO bait alone in female rats. Whereas, in male rats, the
percent acceptance of WSO bait containing 2 and 5% EA was
significantly more than WSO bait alone. So from present
studies it can be concluded that addition of EA in WSO bait
increases the acceptance of bait by both male and female rats.
This may be due to high nutritional value of EA than ESP and
CES.
2% 5% 10%0
10
20
30
40
50
60
70
80Female WSO + ESP
WSO + EA
Conc.of bait additives
Perc
ent
acce
ptan
ce
a a
a ab
a
ba
b
ab
Figure 12. Comparison of percent acceptance of baitscontaining different bait additives by female rats inbi-choice feeding tests.
50
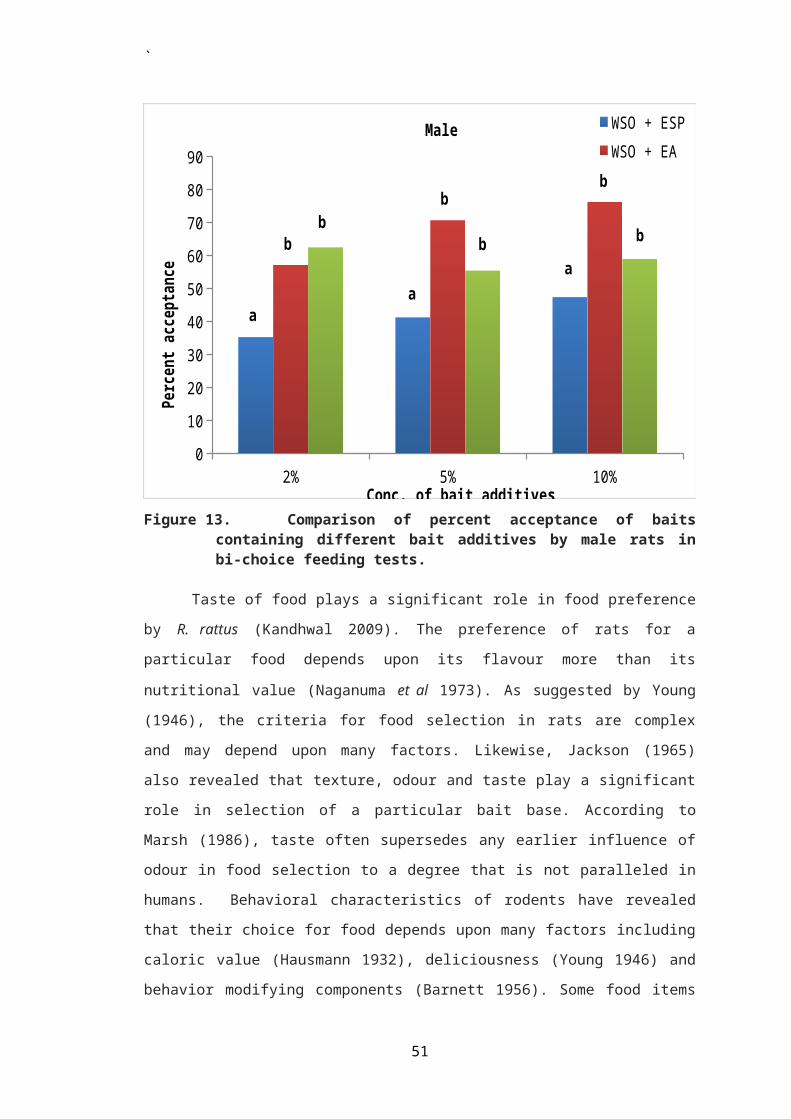
`
2% 5% 10%0
102030405060708090
Male WSO + ESPWSO + EA
Conc. of bait additives
Perc
ent
acce
ptan
ce
a
bb
a
b
b
b
ba
Figure 13. Comparison of percent acceptance of baitscontaining different bait additives by male rats inbi-choice feeding tests.
Taste of food plays a significant role in food preference
by R. rattus (Kandhwal 2009). The preference of rats for a
particular food depends upon its flavour more than its
nutritional value (Naganuma et al 1973). As suggested by Young(1946), the criteria for food selection in rats are complex
and may depend upon many factors. Likewise, Jackson (1965)
also revealed that texture, odour and taste play a significant
role in selection of a particular bait base. According to
Marsh (1986), taste often supersedes any earlier influence of
odour in food selection to a degree that is not paralleled in
humans. Behavioral characteristics of rodents have revealed
that their choice for food depends upon many factors including
caloric value (Hausmann 1932), deliciousness (Young 1946) and
behavior modifying components (Barnett 1956). Some food items
51

`
are intermittently preferred because of the energy they
provide (Stenseth 1977), chiefly due to their carbohydrate and
protein contents (Smythe 1976). These findings support present
results where we observed huge variations among the food
preferences for different taste enhancers. Like, there was no
significant difference observed between the consumption of WSO
bait alone and that containing 2, 5 and 10% crushed egg shell
by both male and female rats. There was no significant
difference observed between consumption of WSO bait alone and
that containing 2, 5 and 10% egg shell powder by male rats,
however, a significantly higher consumption of WSO bait
containing 2 and 5% egg shell powder was observed by female
rats. Also significantly higher consumption was observed of
WSO bait containing 5 and 10% egg albumin by male rats and
that of WSO containing all the three concentrations of egg
albumin by female rats. Present results thus indicate sex
specific differences in acceptance of cereal based bait
containing different poultry egg components as bait additives
in bi-choice. Similar sex specific variations in response of R.
rattus have also been reported earlier (Kaur et al 2008).
4.1.2 No-choice experiments under FSCM
In female rats (110-155g bw), no significant difference
was observed in average consumption (g/kg bw) of bait in a
period of 2 hr among WSO bait without additive and WSO baits
containing 2% concentration each of ESP, EA and CES in no-
choice feeding tests under FSCM (Table 7, Figure 14). However,
the average consumption of WSO bait with all the three types
of additives was non-significantly high than WSO bait without
additive. Further, among the three treated baits, the average
consumption of bait containing 2% EA was non-significantly
52

`
high than the other two bait additives. The total number of
bouts made by female rats in 2 hr towards WSO bait without
additive and those with three bait additives were also not
found to differ significantly (Table 7, Figure 14). The
number of bouts by female rats were highest towards bait
containing 2% EA and least towards bait containing 2% CES.
Total time spent (s) in bouting in 2 hr a day by female rats
was also maximum in bait containing 2% EA and least in bait
containing 2% CES (Table 7). The total time spent by rats in 2
hr among WSO bait without additive and baits containing three
additives was also not found to differ significantly (Figure
14). Also there was no significant difference in total time
spent towards bouting among the four replicated rats when
exposed to both treated and untreated baits.
Table 7. Acceptance of bait containing different additives byfemale R. rattus as recorded under FSCM
Parameters WSO bait WSO +2% ESP WSO + 2% EA WSO + 2% CES
Foodconsumptio
n(g/kg bw)
1.96±0.49a 4.01±1.00a 6.33±1.58a 6.01±1.50a
Number ofbouts made
4.85±1.48a 4.50±1.43a 6.20± 3.16a 2.45±0.51a
Time spenttowardsbouting(s)
175.10±78.38a
156.50±63.39a 181.50±92.21a
69.10±18.88a
-Values are Mean ± SE, Values with similar superscript (a) ina row do not differ significantly.
53
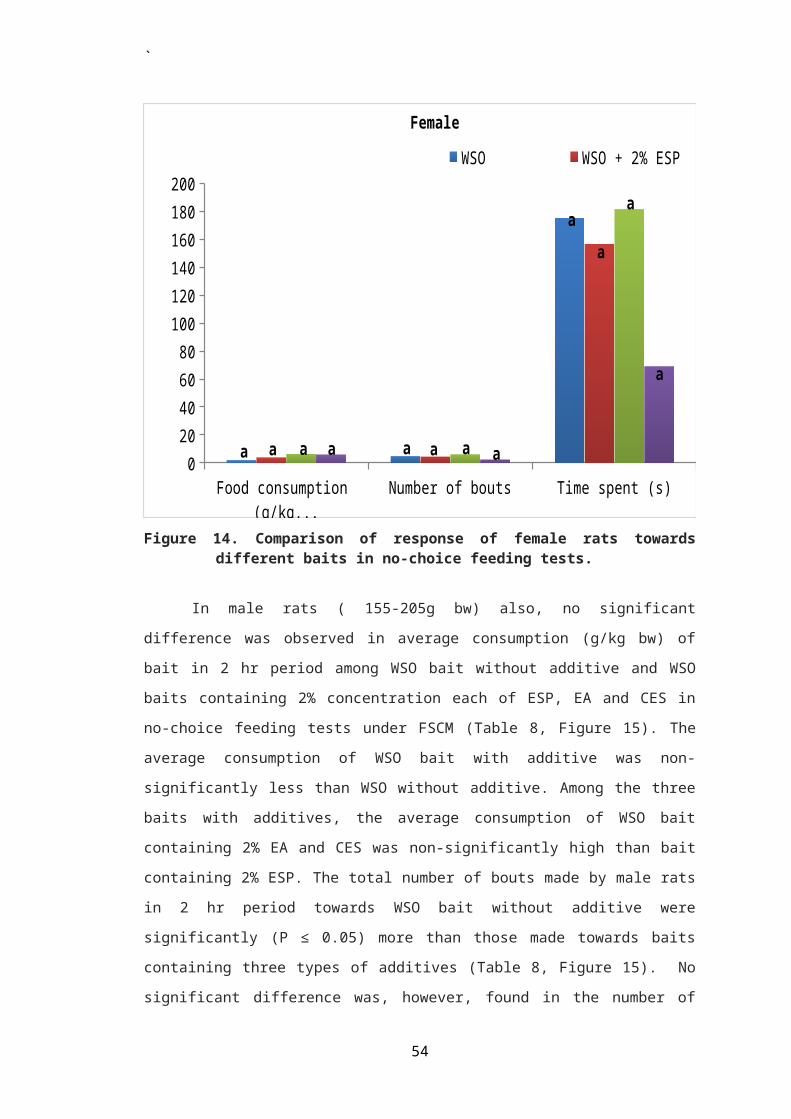
`
Food consumption (g/kg...
Number of bouts Time spent (s)0
20406080
100120140160180200
FemaleWSO WSO + 2% ESP
a a a a a
a
aa
a
a a a
Figure 14. Comparison of response of female rats towardsdifferent baits in no-choice feeding tests.
In male rats ( 155-205g bw) also, no significant
difference was observed in average consumption (g/kg bw) of
bait in 2 hr period among WSO bait without additive and WSO
baits containing 2% concentration each of ESP, EA and CES in
no-choice feeding tests under FSCM (Table 8, Figure 15). The
average consumption of WSO bait with additive was non-
significantly less than WSO without additive. Among the three
baits with additives, the average consumption of WSO bait
containing 2% EA and CES was non-significantly high than bait
containing 2% ESP. The total number of bouts made by male rats
in 2 hr period towards WSO bait without additive were
significantly (P ≤ 0.05) more than those made towards baits
containing three types of additives (Table 8, Figure 15). No
significant difference was, however, found in the number of
54

`
bouts among the WSO baits containing 2% ESP, EA and CES. Total
time spent (s) in bouting in 2 hr period a day by male rats
was also significantly (P ≤ 0.05) more in WSO without any
additive than baits containing three additives (Table 8,
Figure 15). No significant difference was, however, found in
the time spent among the WSO baits containing 2% ESP, EA and
CES. The time spent for bouting in the baits containing 2% ESP
and EA was non-significantly more than that spent in bait
containing 2% CES.
Table 8:Acceptance of bait containing different additives bymale R. rattus as recorded under FSCM
Parameters WSO bait WSO +2% ESP WSO + 2% EA WSO + 2% CES
Foodconsumptio
n(g/kg bw)
7.28±1.82a 2.61±0.65a 6.04±1.51a 6.22±1.55a
Number ofbouts made
12.55±4.35a 4.10 ±1.22b 4.40±0.56b 2.80±0.19b
Time spenttowardsbouting(s)
689.45±350.08a
172.45±77.12b
163.35±42.76b
99.05±14.30b
-Values are Mean ± SE, Values with different superscripts (a-b) in a row differ significantly at P ≤ 0.05.
55
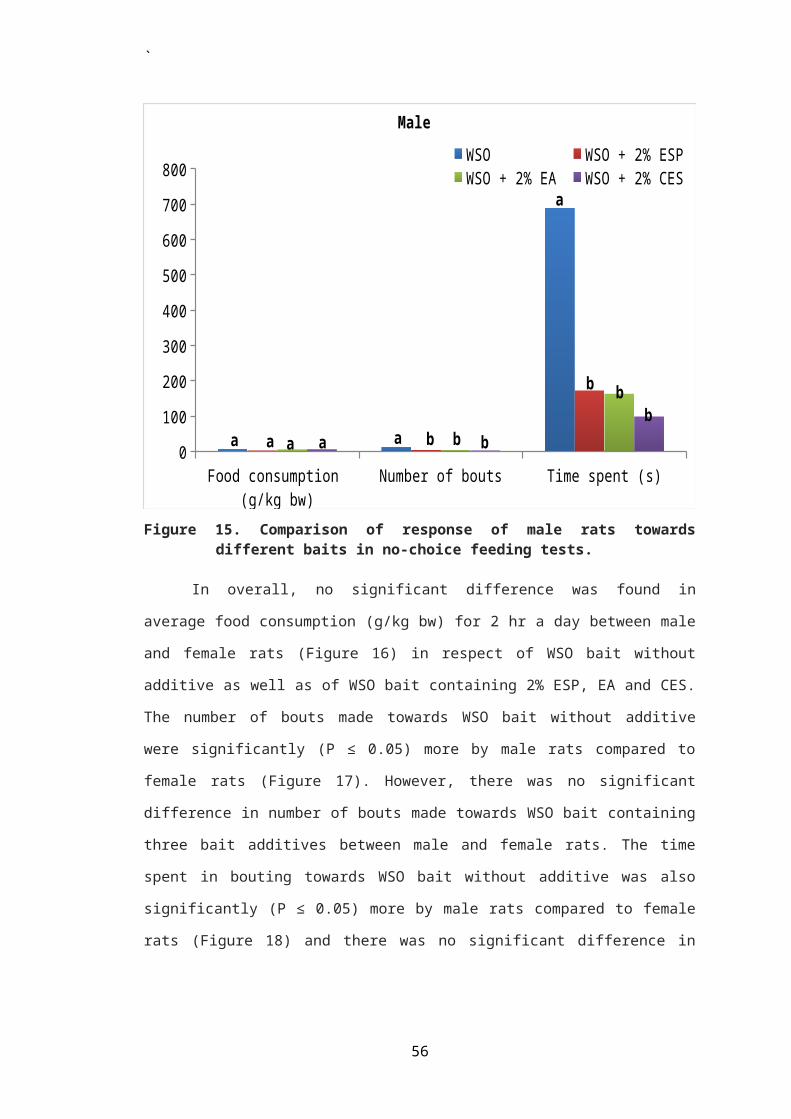
`
Food consumption (g/kg bw)
Number of bouts Time spent (s)0
100
200300
400
500
600
700800
MaleWSO WSO + 2% ESPWSO + 2% EA WSO + 2% CES
a a a a a b b bb
bb
a
Figure 15. Comparison of response of male rats towardsdifferent baits in no-choice feeding tests.
In overall, no significant difference was found in
average food consumption (g/kg bw) for 2 hr a day between male
and female rats (Figure 16) in respect of WSO bait without
additive as well as of WSO bait containing 2% ESP, EA and CES.
The number of bouts made towards WSO bait without additive
were significantly (P ≤ 0.05) more by male rats compared to
female rats (Figure 17). However, there was no significant
difference in number of bouts made towards WSO bait containing
three bait additives between male and female rats. The time
spent in bouting towards WSO bait without additive was also
significantly (P ≤ 0.05) more by male rats compared to female
rats (Figure 18) and there was no significant difference in
56

`
time spent in bouting towards WSO bait containing three bait
additives between male and female rats.
These results under FSCM thus indicate almost similar
consumptions of WSO bait without additive and those with
additives in no-choice tests. Among baits with different
additives, however, the bait consumption, number of bouts made
and the time spent were non-significantly more towards the
bait containing 2% EA than baits containing 2% ESP and CES
which may again be due to high nutritional value of EA as
compared to ESP and CES. The results of no-choice feeding
tests under FSCM also indicate sex specific differences in
behaviour of R. rattus regarding consumption of WSO bait
containing poultry egg components as bait additives, number of
bouts made as well as the time spent.
WSO WSO + 2% ESP WSO + 2% EA WSO + 2% CES0
1
2
3
4
5
6
7
8FemaleMale
Food
con
sume
d (g
/kg
bw)
a
a
a
a
a a a a
57

`
Figure 16. Comparison of total consumption of different baitsin 2 hr period between male and female rats in no-choice feeding tests.
WSO WSO + 2% ESP WSO + 2% EA WSO + 2% CES0
2
4
6
8
10
12
14 FemaleMale
Numb
er o
f bo
uts
a
b
a a a
a
a a
Figure 17. Comparison of total number of bouts made towardsdifferent baits in 2 hr period between male and femalerats in no-choice feeding tests.
58
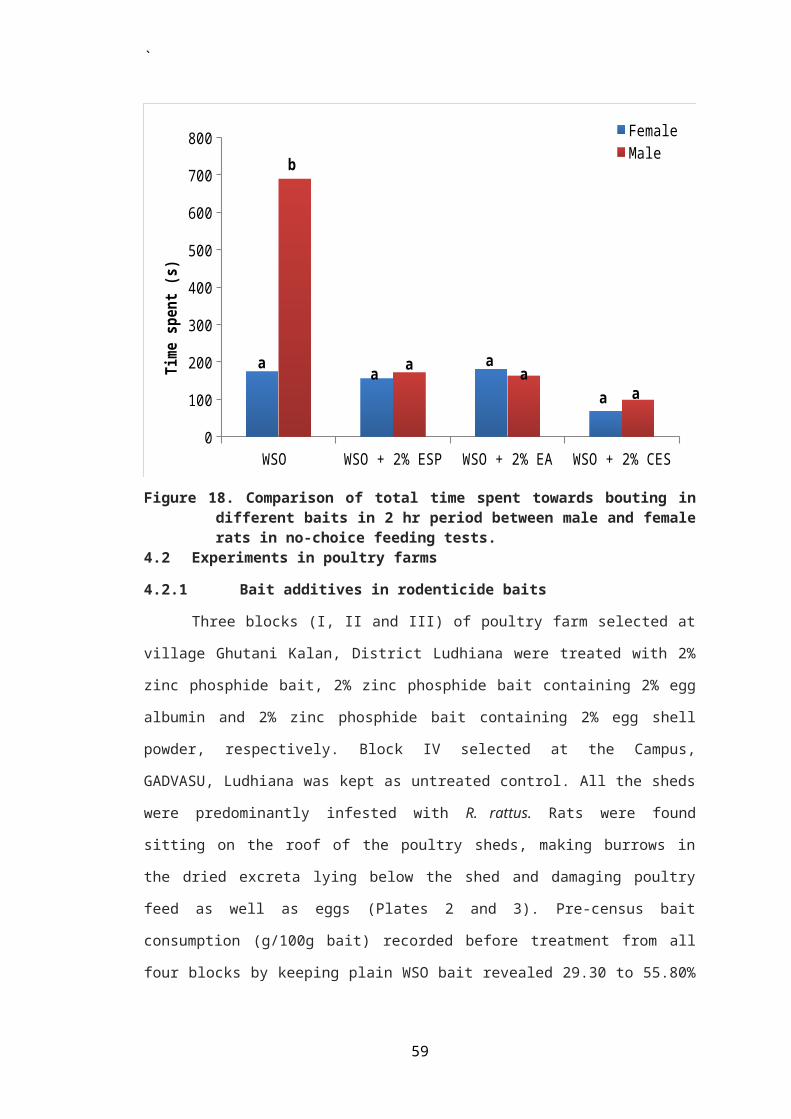
`
WSO WSO + 2% ESP WSO + 2% EA WSO + 2% CES0
100
200
300
400
500
600
700
800 FemaleMale
Time
spe
nt (
s)
a
b
a a aa
a a
Figure 18. Comparison of total time spent towards bouting indifferent baits in 2 hr period between male and femalerats in no-choice feeding tests.
4.2 Experiments in poultry farms
4.2.1 Bait additives in rodenticide baits
Three blocks (I, II and III) of poultry farm selected at
village Ghutani Kalan, District Ludhiana were treated with 2%
zinc phosphide bait, 2% zinc phosphide bait containing 2% egg
albumin and 2% zinc phosphide bait containing 2% egg shell
powder, respectively. Block IV selected at the Campus,
GADVASU, Ludhiana was kept as untreated control. All the sheds
were predominantly infested with R. rattus. Rats were found
sitting on the roof of the poultry sheds, making burrows in
the dried excreta lying below the shed and damaging poultry
feed as well as eggs (Plates 2 and 3). Pre-census bait
consumption (g/100g bait) recorded before treatment from all
four blocks by keeping plain WSO bait revealed 29.30 to 55.80%
59

`
consumption (Table 9). No significant difference was found in
pre-census bait consumption among the four treated and
untreated blocks indicating almost similar level of rodent
activity in all the blocks (Figure 19). The consumption of
treatment bait was found to be 26.95, 68.20 and 68.65% in
blocks I, II and III, respectively (Table 9). The consumption
of 2% zinc phosphide bait containing 2% egg albumin and 2% egg
shell powder was found to be significantly (P ≤ 0.05) higher
than that of 2% zinc phosphide bait alone (Figure 19).
Comparatively less consumption of zinc phosphide bait without
additive in block I may be due to neophobia behaviour shown by
rats leading to bait aversion towards highly toxic rodenticide
bait. Record of post census bait consumption after one week of
treatment from all the blocks revealed a decrease in bait
consumption from that observed during pre census in all the
three treated blocks, whereas, an increase in bait consumption
from that pre-census bait consumption was observed in
untreated block.
Table 9. Census and treatment bait consumption in differentblocks of poultry farms
Block Treatment Bait consumption (g/100g bait)
Pre-census Treatment Post-census
I ZnP 29.30±11.89a 26.95±13.99a 23.75±10.82
II ZnP + EA 55.60±7.74a 68.20±8.86b 42.20±3.19
III ZnP + ESP 55.80±13.42a 68.65±13.95b 45.70±8.56
IV Control 33.40±8.70a - 38.15±4.65
Values are Mean±SE, ZnP = 2% Zinc phosphide, EA = 2% Egg albumin, ESP = 2% Egg shell powder, Values with different
60

`
superscripts (a-b) in a column differ significantly at P ≤ 0.05.
Pre-census Treatment0
10
20
30
40
50
60
70
80ZnP ZnP + EA ZnP + ESP UT
Bait
con
sump
tion
(g/
100g
ba
it)
a
aa
a a
bb
Figure 19. Comparison of bait consumptions during census andtreatment in different poultry blocks.
The reduction in rodent activity in the three treated
blocks was found to vary from 28.13 to 44.41% with respect to
untreated block and 19.23 to 42.69% with respect to same shed
(Table 10). There was no significant difference in reduction in
rodent activity between the three treated blocks (Figure 20),
though apparently, there was a higher decrease in rodent
activity in block III treated with 2% zinc phosphide bait
containing 2% ESP as additive. Results in poultry farm therefore
reveal higher efficacy of 2% zinc phosphide containing 2% ESP.
Less efficacy of rodenticide treatment in poultry farm during
present studies may be due to the availability of other food
items such as poultry feed, eggs, chicks etc in the poultry farm
which may further have led to reduced acceptance of rodenticide
bait. Present results, however, suggest the potential of EA and
ESP in increasing the acceptance of 2% zinc phosphide bait.
61
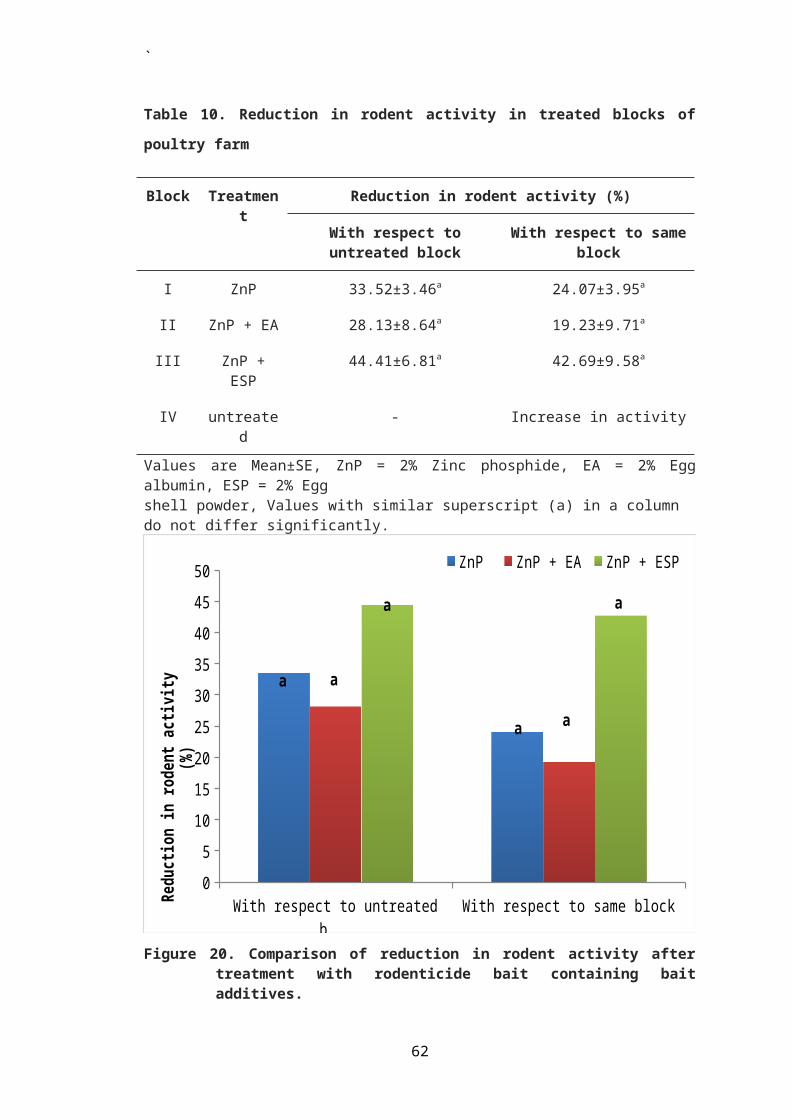
`
Table 10. Reduction in rodent activity in treated blocks of
poultry farm
Block Treatment
Reduction in rodent activity (%)
With respect tountreated block
With respect to sameblock
I ZnP 33.52±3.46a 24.07±3.95a
II ZnP + EA 28.13±8.64a 19.23±9.71a
III ZnP +ESP
44.41±6.81a 42.69±9.58a
IV untreated
- Increase in activity
Values are Mean±SE, ZnP = 2% Zinc phosphide, EA = 2% Eggalbumin, ESP = 2% Egg shell powder, Values with similar superscript (a) in a column do not differ significantly.
With respect to untreated b...
With respect to same block05
101520253035404550 ZnP ZnP + EA ZnP + ESP
Redu
ctio
n in
rod
ent
acti
vity
(%
)
a a
a
a a
a
Figure 20. Comparison of reduction in rodent activity aftertreatment with rodenticide bait containing baitadditives.
62

`
The literature also reveals the use of cereals in whole-
some or cracked form mixed with additives such as vegetable
oil, egg shell, egg yolk, minced meat, sugar and flavours
(Shafi et al 1990, Pervez et al 2003). Khan et al (2000) reported
preference for poison bait containing egg shells by rodents in
rice crop fields. Pervez et al (2005) found highest potential of
egg mixed brodifacoum bait in enhancing acceptance by field
rodents in wheat crop. However, the addition of egg shell at
2% did not show a significant increase in relative consumption
for the cereal base by B. bengalensis (Naeem et al 2011). Rather
they observed an inverse trend indicating decreased preference
for plain millet based bait mixed with 5% egg shell. Shafi et al
(1993) also observed that B. bengalensis show a high preference
toward minced meat bait than egg shell and egg yolk mixed
bait. Mushtaq et al (2013) also did not found any increase in
acceptance of cereal base containing egg shell by Hystrix indica.
However, Pervez et al (1999 and 2003) found an additive effect of
2% egg shell in enhancement of bait preference. These
contradictory results may primarily be attributed to the
nature of bait material used and due to the different species
tested.
4.2.2 Bait additives in trapsRodent trapping with wooden single rat traps was carried
out in sheds of poultry farm located in the campus of GADVASU,Ludhiana. Each trap was baited with a piece of chapattismeared with 2% (by weight) poultry ESP and 2% EA. For thefirst time, trapping was carried out from all the sheds byalternatively placing traps containing chapatti pieces withoutadditive and those containing chapatti pieces smeared with 2%
63

`
egg shell powder. Results revealed total trap index of R. rattus
to be 11.60 (6.84 for male and 4.76 for female rats) in trapsbaited with chapatti piece smeared with ESP and 13.38 (6.84for male and 6.54 for female rats) in traps baited withchapatti piece without any additive (Table 11). No significantdifference was found in total trap index as well as that ofmale and female rats separately between the traps baited withchapatti pieces with and without ESP (Figure 21). Also nosignificant difference was found in the trap index of trapsbaited with chapatti pieces smeared with and without ESPbetween the two sexes (Figure 22). Table 11. Comparison of trap index of rats trapped by smearing
chapatti pieces with and without egg shell powder asadditive
Additive used onchapatti
Trap index(no. of animals trapped/100 traps/day)
Male Female Total
Egg shell powder 6.84 ±2.04 a 4.76 ±1.87 a 11.60± 3.69 a
Untreated 6.84 ±1.22 a 6.54 ±2.87 a 13.38± 3.61 a
Values are Mean ±SE, Values with similar superscript (a) in acolumn and in a row for two sexes do not differ significantly.
64

`
Male Female Total0
2
4
6
8
10
12
14
16Egg shell powder
Trap
ind
ex
a aa
a
aa
Figure 21. Comparison of trap index of rats trapped bysmearing chapatti pieces with and without egg shellpowder as additive.
Egg shell powder Untreated0
1
2
3
4
5
6
7
8 MaleFemale
Trap
ind
ex
a
a
a
a
Figure 22. Comparison of trap index of male and female ratstrapped by smearing chapatti pieces with and withoutegg shell powder as additive.
65

`
For the second time, trapping was carried out from all
the sheds by alternatively placing traps containing chapatti
pieces without additive and those containing chapatti pieces
smeared with 2% EA. Results revealed total trap index of R.
rattus to be 5.35 (3.27 for male and 2.08 for female rats) in
traps baited with chapatti piece smeared with EA and 7.13
(2.08 for male and 5.05 for female rats) in traps baited with
chapatti piece without any additive (Table 12). No significant
difference was found in total trap index as well as that of
male and female rats separately between the traps baited with
chapatti pieces with and without EA (Figure 23). Also no
significant difference was found in the trap index of traps
baited with chapatti pieces smeared with and without EA
between the two sexes (Figure 24).
Table 12. Comparison of trap index of rats trapped by smearingchapatti pieces with and without egg albumin asadditive
Additive used onchapatti
Trap index(no. of animals trapped/100 traps/day)
Male Female Total
Egg albumin 3.27 ±1.06 a 2.08 ±0.78 a 5.35 ±1.68 a
Untreated 2.08 ±1.29 a 5.05 ±1.42 a 7.13 ±2.59 a
Values are Mean ±SE, Values with similar superscript (a) in acolumn and in a row for two sexes do not differ significantly.
66

`
Male Female Total0
1
2
3
4
5
6
7
8 Egg albuminUntreated
Trap
ind
ex
aa
a
aa
a
Figure 23. Comparison of trap index of rats trapped bysmearing chapatti pieces with and without egg albuminas additive.
Egg albumin Untreated0
1
2
3
4
5
6 Male
Trap
ind
ex
a
aa
a
67

`
Figure 24. Comparison of trap index of male and female ratstrapped by smearing chapatti pieces with and withoutegg albumin as additive.
For the third time, trapping was carried out from all the
sheds by alternatively placing traps containing chapatti
pieces smeared with 2% ESP and 2% EA. Results revealed total
trap index of R. rattus to be 13.69 (6.25 for male and 7.44 for
female rats) in traps baited with chapatti piece smeared with
ESP and 12.79 (8.03 for male and 4.76 for female rats) in
traps baited with chapatti piece smeared with EA (Table 13).
No significant difference was found in total trap index as
well as that of male and female rats separately between the
traps baited with chapatti pieces smeared with ESP and EA
(Figure 25). Also no significant difference was found in the
trap index of traps baited with chapatti pieces smeared with
ESP and EA between the two sexes (Figure 26).
Table 13. Comparison of trap index of rats trapped by smearingchapatti pieces with egg shell powder and egg albuminas additives
Additive used onchapatti
Trap index(no. of animals trapped/100 traps/day)
Male Female Total
Egg shell powder 6.25 ±2.01 a 7.44 ±2.61 a 13.69± 4.12 a
Egg albumin 8.03 ±2.22 a 4.76 ±1.55 a 12.79± 2.03 a
Values are Mean ±SE, Values with similar superscript (a) in acolumn and in a row for two sexes do not differ significantly.
68

`
Male Female Total0
2
4
6
8
10
12
14
16 Egg shell ...
Trap
ind
ex
aa a
a
a
a
Figure 25. Comparison of trap index of rats trapped bysmearing chapatti pieces with egg shell powder and eggalbumin as additives.
Egg shell powder Egg albumin0
1
2
3
4
5
6
7
8
9 Male Female
Trap
ind
ex
a
a a
a
Figure 26. Comparison of trap index of male and female ratstrapped by smearing chapatti pieces with egg shellpowder and egg albumin as additives.
69

`
In overall, the results of poultry trapping experiments
revealed no significant effect of poultry egg components i.e.
ESP and EA in trapping of rats by single catch rat traps. This
may be due to the reason that rats might not have come to know
about the additive smeared on chapatti piece before they
pulled it from the string of the trap and got trapped. Due to
this very reason there was no significant difference in trap
index between traps baited with chapatti piece smeared with
and without additive and also between the traps baited with
chapatti smeared with ESP and EA.
During present studies, results of laboratory experiments
in bi-choice revealed significantly (P ≤ 0.05) higher
consumption of WSO bait containing all the three
concentrations (2, 5 and 10%) of EA and 2 and 5% ESP by female
rats and that of bait containing 5 and 10% EA by male rats. In
no-choice experiments, the bait consumption, number of bouts
made and the time spent in WSO bait containing 2 % EA by
female rats was non-significantly high than that containing 2%
ESP and CES. In male rats, the results were, however,
inconclusive. In poultry farm also significantly higher
consumption of 2% zinc phosphide bait containing 2% EA and 2%
ESP leading to upto 44.41% reduction in rodent activity was
observed compared to 33.52% reduction in rodent activity with
only 2% zinc phosphide treatment.
We may therefore assume that use of 2% egg albumin and
egg shell powder in cereal based bait may enhance bait intake
as compared to plain bait. This combination may yield
significant control of R. rattus if used in 2% zinc phosphide
bait.
70

CHAPTER – V
SUMMARY
Present study on evaluation of poultry components i.e.
egg shell powder (ESP), egg albumin (EA) and crushed egg
shells (CES) as bait additives for increasing the acceptance
of cereal based bait by house rat, Rattus rattus was carried out
in bi-choice and no-choice laboratory feeding tests. The
effective concentrations of these components were then tested
in poultry farms to evaluate the acceptance and efficacy of 2%
zinc phosphide bait without additive and that containing 2%
ESP and EA. The effect of these poultry components was also
studied on rodent trapping.
In bi-choice laboratory experiments, no significant
difference was observed in mean daily consumption (g/100g bw)
of WSO bait during pre- and post-treatment periods among all
the treated and untreated groups of rats of both sexes for all
the bait additives tested. During treatment with different
concentrations of ESP, significantly (P ≤ 0.05) higher mean
daily consumption (g/100g bw) of WSO bait containing 2 and 5%
ESP from that of WSO bait alone was observed in female rats.
There was no significant difference in consumption between WSO
bait without additive and that containing 10% ESP by female
rats. Also there was no significant difference in mean daily
consumption of WSO bait containing egg shell powder among the
three treated groups. Percent acceptance of WSO bait with
additive over WSO bait alone was also not found to differ
significantly in female rats among the three treated groups.
The percent acceptance was found to be 67.21, 58.04 and
49.18%, respectively in rats of groups treated with 2, 5 and
10% ESP.

`
In male rat, there was no significant difference observed
in mean daily consumption of WSO bait containing bait additive
and that without any additive at all the concentrations of ESP
tested. Also there was no significant difference in mean daily
consumption of WSO bait containing ESP among the three treated
groups. Percent acceptance of WSO bait with additive over WSO
bait alone was also not found to differ significantly among
the three treated groups. The percent acceptance was found to
be 35.19, 41.20 and 47.32%, respectively in rats of groups
treated with 2, 5 and 10% ESP.
Percent acceptance of WSO bait containing ESP was
generally low in male rats as compared to female rats. This
difference in acceptance of bait between male and female rats
was found to be statistically significant (P ≤ 0.05) when rats
were exposed to WSO bait containing 2% ESP in choice with WSO
bait without additive.
During treatment with different concentrations of EA,
significantly (P ≤ 0.05) higher mean daily consumption of WSO
bait containing 2, 5 and 10% EA from that of WSO without
additive was observed in female rats. There was no significant
difference in mean daily consumption of WSO bait containing EA
among the three treated groups. Percent acceptance of WSO bait
with additive over WSO bait alone was also not found to differ
significantly in female rats among the three treated groups.
The percent acceptance was found to be 67.45, 69.13 and 65%,
respectively in rats of groups treated with 2, 5 and 10% EA.
In male rat, significantly (P ≤ 0.05) higher mean daily
consumption of WSO bait containing 5 and 10% EA was found from
that of WSO bait alone. There was no significant difference
between the consumption of WSO bait without additive and that
72

`
containing 2% EA. Also there was no significant difference in
mean daily consumption of WSO bait containing EA among the
three treated groups. Percent acceptance of WSO bait with
additive over WSO bait alone was found to be significantly (P
≤ 0.05) high in rats of groups treated with 5% (70.63%) and
10% (76.22%) EA as compared to rats of group treated with 2%
EA (57.12%).
Percent acceptance of WSO bait containing EA by male rats
as compared to female rats was generally low at 2% EA and high
at 5 and 10% EA. This difference in acceptance of bait between
male and female rats was, however, found to be statistically
non-significant at all the concentrations tested.
During treatment with different concentrations of CES, no
significant difference in mean daily consumption between WSO
bait containing 2, 5 and 10% CES and WSO bait without additive
was observed in both male and female rats. The consumption of
two kinds of baits was almost similar. Also there was no
significant difference in mean daily consumption of WSO bait
containing CES among the three treated groups. Percent
acceptance of WSO bait with additive was also not found to
differ significantly among the three treated groups. The
percent acceptance was found to be 55.34, 54.25 and 53.62%,
respectively in female rats and 62.39, 55.35 and 58.94%,
respectively in male rats of groups treated with 2, 5 and 10%
CES.
Percent acceptance of WSO bait containing CES by male
rats as compared to female rats was generally high at all the
three concentrations of CES tested. This difference in
acceptance of bait between male and female rats was, however,
73

`
not found to differ significantly at any of the concentrations
tested.
In female rats, the percent acceptance of WSO bait
containing 2% ESP, EA and CES was not found to differ
significantly. However, the percent acceptance of WSO bait
containing 5% EA was significantly (P ≤ 0.05) high from WSO
bait containing 5% CES and non-significantly high from WSO
bait containing 5% ESP. The percent acceptance of WSO bait
containing 10% EA was significantly (P ≤ 0.05) high from WSO
bait containing 10% ESP and non-significantly high from WSO
bait containing 10% CES. There was no significant difference
in percent acceptance of WSO bait containing ESP and CES at
all the concentrations tested.
In male rats, percent acceptance of WSO bait containing
EA and CES was found to be significantly (P ≤ 0.05) high from
that of WSO bait containing ESP at all the concentrations
tested. At 5 and 10% concentrations, percent acceptance of WSO
bait containing EA was comparatively more than WSO bait
containing similar concentrations of CES. Statistically, there
was no significant difference in percent acceptance of WSO
bait containing EA and CES. Present results thus indicate sex
specific differences in acceptance of cereal based bait
containing different poultry egg components as bait additives
in bi-choice. So from present studies it can be concluded that
addition of EA in WSO bait increases the acceptance of bait by
both male and female rats. This may be due to high nutritional
value of EA than ESP and CES.
In no-choice experiments under food scale consumption
monitor, no significant difference was observed in average
consumption (g/kg bw) of bait for 2 hr period by female rats
74

`
among WSO bait without additive and WSO baits containing 2%
concentration each of ESP, EA and CES. The average consumption
of WSO bait with all the three types of additives was non-
significantly high than WSO bait without additive. Among the
three treated baits, the average consumption of bait
containing 2% EA was non-significantly high than other two
bait additives. The total number of bouts made by female rats
in 2 hr period towards WSO bait without additive and those
with three bait additives were also not found to differ
significantly. The number of bouts by female rats were highest
towards bait containing 2% EA and least towards bait
containing 2% CES. Total time spent (s) in bouting in 2 hr
period by female rats was also maximum in bait containing 2%
EA and least in bait containing 2% CES. The total time spent
by rats in 2 hr among WSO bait without additive and baits
containing three additives was also not found to differ
significantly.
In male rats also, no significant difference was observed
in average consumption (g/kg bw) of bait for 2 hr period among
WSO bait without additive and WSO baits containing 2%
concentration each of ESP, EA and CES in no-choice feeding
tests under FSCM. The average consumption of WSO bait with
additive was non-significantly less than WSO without additive.
Among the three baits with additives, the average consumption
of WSO bait containing 2% ESP and EA was non-significantly
high than bait containing 2% ESP. The total number of bouts
made by male rats in 2 hr period towards WSO bait without
additive were significantly (P ≤ 0.05) more than those made
towards baits containing three types of additives. No
significant difference was, however, found in the number of
75

`
bouts among the WSO baits containing 2% ESP, EA and CES. Total
time spent (s) in bouting in 2 hr period a day by male rats
was also significantly (P ≤ 0.05) more in WSO without any
additive than baits containing three additives. No significant
difference was found in the time spent among the WSO baits
containing 2% ESP, EA and CES. The time spent for bouting in
the baits containing 2% ESP and EA was non-significantly more
than that spent in bait containing 2% CES.
In overall, no significant difference was found in
average food consumption (g/kg bw) for 2 hr a day between male
and female rats in respect of WSO bait without additive as
well as of WSO bait containing 2% ESP, EA and CES. The number
of bouts made towards WSO bait without additive were
significantly (P ≤ 0.05) more by male rats compared to female
rats. However, there was no significant difference in number
of bouts made towards WSO bait containing three bait additives
between male and female rats. The time spent in bouting
towards WSO bait without additive was also significantly (P ≤
0.05) more by male rats compared to female rats and there was
no significant difference in time spent in bouting towards WSO
bait containing three bait additives between male and female
rats.
These results in no-choice indicate almost similar
consumptions of WSO bait without additive and those with
additives. Among baits with different additives, however, the
bait consumption, number of bouts made and the time spent were
non-significantly more towards the bait containing 2% EA than
baits containing 2% ESP and CES by female rats. The results on
acceptance of baits by male rats are thus inconclusive. The
results of no-choice feeding tests thus indicate sex specific
76

`
differences in behaviour of R. rattus towards WSO bait containing
poultry egg components as bait additives.
Treatment of three blocks (I, II and III) of poultry farm
at village Ghutani Kalan, District Ludhiana with 2% zinc
phosphide bait without additive and that containing 2% EA and
ESP, respectively revealed 26.95, 68.20 and 68.65% consumption
of treated bait in blocks I, II and III, respectively.
Consumption of treated baits was significantly (P ≤ 0.05) high
in blocks II and III as compared to that in block I. No
significant difference was found in pre-census bait
consumption among the four treated and untreated blocks
indicating almost similar level of rodent activity in all the
blocks. Percent reduction in rodent activity in blocks treated
with 2% zinc phosphide without additive and those containing
2% EA (block II) and 2% ESP (block III) was found reduced by
33.52, 28.13 and 44.41%, respectively with respect to
untreated block and by 24.07, 19.23 and 42.69%, respectively
with respect to same block. However, there was no significant
difference in reduction in rodent activity between the blocks
I, II and III. In block I, significantly less acceptance of 2%
zinc phosphide without additive may be due to neophobia
behaviour shown by rats towards highly toxic rodenticide
leading to subsequent bait aversion. The results suggest the
potential of 2% EA and ESP in increasing the acceptance of 2%
zinc phosphide bait.
Rodent trapping with wooden single rat traps was carried
out in sheds of poultry farm located in the campus of GADVASU,
Ludhiana. Each trap was baited with a piece of chapatti
smeared by weight with 2% ESP and EA. For the first time,
trapping was carried out from all the sheds by alternatively
77

`
placing traps containing chapatti pieces without additive and
those containing chapatti pieces smeared with 2% ESP. Results
revealed total trap index of R. rattus to be 11.60 in traps
baited with ESP and 13.38 in traps baited with chapatti piece
without any additive. No significant difference was found in
total trap index as well as that of male and female rats
between the traps baited with chapatti pieces with and without
ESP. Also no significant difference was found in the trap
index between the two sexes.
For the second time, trapping was carried out by
alternatively placing traps containing chapatti pieces without
additive and those containing chapatti pieces smeared with 2%
EA. Results revealed total trap index of R. rattus to be 5.35 in
traps baited with EA and 7.13 in traps baited with chapatti
piece without any additive. No significant difference was
found in total trap index as well as that of male and female
rats between the traps baited with chapatti pieces with and
without egg albumin. Also no significant difference was found
in the trap index between the two sexes.
For the third time, trapping was carried out from all the
sheds by alternatively placing traps containing chapatti
pieces smeared with 2% ESP and 2% EA. Results revealed total
trap index of R. rattus to be 13.69 in traps baited with ESP and
12.79 in traps baited with EA. No significant difference was
found in total trap index as well as that of male and female
rats between the traps baited with chapatti pieces smeared
with ESP and EA. Also no significant difference was found in
the trap index of traps between the two sexes.
In overall, the results of poultry trapping experiments
revealed no significant effect of poultry egg components i.e.
78

`
ESP and EA in trapping of rats by single catch rat traps. This
may be due to the reason that rats might not have come to know
about the additive smeared on chapatti piece before they
pulled it from the string of the trap and got trapped.
During present studies, results of laboratory experiments
in bi-choice revealed significantly (P ≤ 0.05) higher
consumption of WSO bait containing all the three
concentrations of EA and 2 and 5% ESP by female rats and that
of bait containing 5 and 10% EA by male rats. In no-choice
experiments, the consumption, number of bouts made and the
time spent in WSO bait containing 2% EA by female rats was
non-significantly high than that containing 2% ESP and CES. In
poultry farm, significantly higher consumption of 2% zinc
phosphide bait containing 2% EA and 2% ESP leading to upto
44.41% reduction in rodent activity was observed compared to
33.52% reduction in rodent activity with only 2% zinc
phosphide treatment.
We may therefore assume that use of 2% egg albumin and
egg shell powder in cereal based bait may enhance bait intake
as compared to plain bait. This combination may yield
significant control of R. rattus if used in 2% zinc phosphide
bait.
79

REFERENCES
Abbas S M (2003) Field evaluation of cereal grain bait materials against thelesser bandicoot rat (Bandicota bengalensis) in wheat crop. M.Sc. thesis,Arid Agriculture University, Rawalpindi, Pakistan.
Ahmed E, Hussain 1 and Brooks J E (1995) Losses of stored fooddue to rat at grain markets in Pakistan. Int J Biodeter Biodegr17: 125-33.
Ahmad M S, Munir S and Khan A A (1994) Laboratory evaluationof some vegetable oils as bait enhancers against roof rat,Rattus rattus. Pakistan J Zool 26(2): 93–97.
Ahmad N, Chopra G and Sood M L (1984) Assessment of rodentdamage in poultry farms. Poult Guide 21(3): 57-60.
Ahmad N, Parshad V R and Malhi C S (1992) Control of roof rat(Rattus rattus L.) in poultry farm. Indian Poult. Rev 23: 11–13.
Alam M S, Main M X and Hasanuzzaman A T M (2007) Evaluation ofdifferent bait materials for trapping rodents. Bangladesh JAgri Environ 3(2): 31-36.
Amusa N A, Iken J E and Fyenuwo J O (2005) The incidences offield diseases and vertebrate pest on popcorn (Zea mayseverta) varieties cultivated in forest agro-ecologies inNigeria. World J Agric Sci 1(2): 173-77.
Aplin K P, Chesser T and Have J (2003) Evolutionary biology ofthe genus Rattus: profile of an archetypal rodent pest. In:Singleton G R, Hinds L A, Krebs C J and Spratt D M (ed)Rats, Mice and People: Rodent Biology and Management. pp 564. No. 96,ACIAR Monograph, Canberra, Australia.
Askham and Leonard R (1987) The rat: Its Biology and Control.Cooperative Extension Bulletin EB1377, Washington StateUniversity, Pullman, WA.
Babbar B K, Singla N and Kaur R (2009) Sugar as additive inzinc phosphide bait for improving acceptance by Rattusrattus. Rodent Newsl 33(1-2): 5-6.
Barnett S A (1956) Behaviour components in the feeding of wildand laboratory rats. Behaviour 1: 24-43.
Barnett S A (1958) Experiments on “neophobia” in wild andlaboratory rats. British J Psychol 49: 195-201.

`
Barnett S A (1966) The feeding of rodents. Proc Indian Rodent Symp.pp 113-23. Calcutta.
Barnett S A, Dickson, R G, Marples T G and Radha E (1978)Sequences of feeding, sampling and exploration by wild andlaboratory rats. Behav Process 3: 29-43.
Barnett S A and Prakash I (1975) Rodents of Economic Importance inIndia. pp 1-175. Arnold Heinmann, New Delhi.
Bean N J, Galef B G and Mason J R (1988) The effect of carbondisulfide on food consumption by house mice. J Wildl Manage52: 502-07.
Berdoy M (1994) Making decisions in the wild: constraints,conflicts and communication in foraging rats. In: Galef JrB G and Valsechhi P (ed) Behavioural Aspects of Feeding. pp 289-313. Ettore Majorana International Life Science Series,Vol 12. Harwood Academic Publishers, Chur.
Berdoy M and MacDonald D W (1991) Factors affecting feeding inwild rat. Acta Oecol 12: 261-79.
Bhardwaj D (1983) Rodent pests and their control in poultryfarms of Jodhpur. Ann Arid Zone 22(4): 337-41.
Bhardwaj D and Khan J A (1979) Effect of texture of food onbait-shy behaviour in wild rats (Rattus rattus L.) Appl Anim Ethol5: 361-67.
Bhardwaj D and Prakash I (1979) Intensity of poison aversionand bait-shyness in Rattus rattus due to exposure to RH-787. ZAngew Zool 66: 427-36.
Bhardwaj D and Prakash I (1982) Mitigation of poison aversionin Rattus rattus through acclimatisation. Indian J Exp Biol 20:396-98.
Biggins D E and Kosoy M Y (2001) Influences of introducedplague on North American mammals: implications fromecology of plague in Asia. J Mammal 82: 906-16.
Borah R K and Bora D K (2012) Incidence and damage of Bandicotabengalensis in rice based cropping system in upperBrahmaputra valley of Assam. Ann Plant Prot Sci 20: 380-82.
Brooks J E, Ahmed E and Hussain I (1987) Rat Population and StoredFood Losses at a Pakistan Grain Market. Technical Report No. 12, pp9. National Agriculture Research Centre, Islamabad.
81

`
Brooks J E and LaVoie G K (1990) Rodent control will reducepost-harvest food losses: Grain storage and Handling.Agribusiness Worldwide, pp 11-17.
Buckle A P (1994) Rodent control methods: chemical. In: BuckleA P and Smith R H (ed) Rodent Pests and their Control. pp 127-60.CAB International, Bristol, UK.
Buckle A P and Smith R H (1994) Rodent Pests and their Control. pp405. CAB International, Bristol, UK.
Bull J O (1972) The influence of attractants and repellents onthe feeding behaviour of Rattus norvegicus. Proc 5th Vertebr PestConf. pp 154–59. University of California, Davis,California, USA.
Bullard R (1985) Isolation and characterization of naturalproducts that attract or repel wild vertebrates. In: AcreeT and Soderlund D (ed) Semiochemistry: Flavours and Pheromones.pp. 65-93. Walter de Gruyter & Co, New York, USA.
Bullard R W and Shumake S A (1977) Food-base flavour additiveimproves bait acceptance by rice field rats. J Wildl Manage41: 290–97.
Canby T Y (1977) The rat: Lapdog of the devil. National Geographic152(1): 60-87.
Cao D, Hart K and Pimentel D (2002) Post harvest food losses(vertebrates). In: Pimentel D (ed) Encyclopaedia of PestManagement. pp 648-49. CRC Press.
Chattopadhyay D, Madhu N R and Manna C K (2010) Burrowingpattern and damage caused by the rodent pest, Indian houserat (Rattus rattus) in the Gangetic and Non Gangetic plain ofWest Bengal, India. Univ J Zool Rajshahi Univ 28: 57-60.
Chitty D and. Kempson D A (1949) Prebaiting small mammals anda new design of live trap. Ecology 30: 536–42.
Chopra G (1992) Poultry farms. In: Prakash I and Ghosh P K(ed) Rodents in Indian Agriculture. pp 309-30. ScientificPublishers, Jodhpur, India.
Chopra G, Ahmad N and Sood M L (1984) Food consumption andpreference behaviour of Rattus meltada (Gray). J Res Punjab AgricUniv 21(4): 561-69.
82

`
Chopra G and Dhindsa J S (1987) Rodent pest management inpoultry farms. Poult Advisor 20(3): 49-60.
Chopra G, Kandhwal S, Kumar P and Rai D (2008a) Opinion surveyof rodent menace and control in commercial poultryfacilities. Pestology 32(2): 61-65.
Chopra G, Kandhwal S, Kumar P and Rai D (2008b) Pattern ofrodent distribution and infestation in commercial poultryfacilities in Haryana. Environ Ecol 26(3): 1006-09.
Chopra G, Kandhwal S, Kumar P, Rai D and Gupta S K (2009) Foodpreference behaviour of Rattus rattus L. infesting poultryfarms. J Exp Zool India 12(1): 107-111.
Chopra G and Sabharwal A K (1992) Rodent distribution andextent of damage to paddy in rice-belt of Haryana. J Haryanastudies 24: 84-89.
Clark D A (1982) Foraging behaviour of a vertebrate omnivore(Rattus rattus): meal structure, sampling and diet breadth.Ecology 63: 763-72.
Collier G and Bolles R (1968) Some determinants of intake ofsucrose solutions. J Comp Physiol Psychol 65: 379–83.
Conover M R, Pitt C, Kessler K K, DuBow T J and Sanborn W A(1995) Review of human injuries, illnesses and economiclosses caused by wildlife in the United States. Wildl Soc Bull23: 407-14.
Cowan P E (1977) Systematic patrolling and orderly behaviourof rats during recovery from deprivation. Anim Behav 25:171-84.
Cowan D P, Quy R J and Lambert M S (2003) Ecologicalperspectives on the management of commensal rodents. In:Singleton G R, Hinds L A, Krebs C J and Spratt D M (ed)Rats, Mice and People: Rodent Biology and Management. pp 433-39.ACIAR Monograph No. 96, Canberra, Australia.
Cuong L Q, Chien H U, Han L U, Duc V H and Singleton G R(2002) Relationship between rodent damage and yield lossin rice in Mekong Delta. In: Singleton G R, Hinds L A,Krebs C J and Spratt D M (ed) Rats, Mice and People: RodentBiology and Management. pp 217-19. ACIAR Monograph No. 96,Canberra, Australia.
83

`
Drickamer L (1997) Responses to odors of dominant andsubordinate house mice in live traps and responses toodors in live traps by dominant and subordinate males. JChem Ecol 23: 2493-2506.
Elizalde G and Sclafani A (1990) Fat appetite in rats: flavourpreferences conditioned by nutritive and non-nutritive oilemulsions. Appetite 15: 189–97.
Ellerman J R (1961) The Fauna of India including Pakistan, Burma and Ceylon:Mammalia. 2nd edn, Vol 3 (Rodentia), Parts 1 & 2, pp 884.Government of India, Delhi.
Fayenuwo J O, Olakojo S A, Akande M, Amusa N A and Olujimi O A(2007) Comparative evaluation of vertebrate pest damage onsome newly developed quality protein maize (QPM) varietiesin South western Nigeria. Afr J Agri Res 2(11): 592-95.
Field R J (1971) The use of sexual attractant pheromones to increase theacceptability of the male chemosteri1ant 3-chloro-l, 2 propanediol by wildNorway rats (Rattus norvegicus). Thesis, Michigan StateUniversity.
Fiedler L A (1988) Rodent problems in Africa. In: Prakash I(ed) Rodent Pest Management. pp 35-65. CRC Press, Inc BocaRaton, Florida, USA.
Fiedler L A, Fall M W and Reidinger Jr R F (1982) Developmentand evaluation of methods to reduce rat damage to coconutin the Philippines. Proc Vertebr Pest Conf 10: 73-79.
Fitzwater W D and Prakash I (1989) Handbook of Vertebrate PestControl. pp 103. Indian Council of Agricultural Research,New Delhi.
Flynn F W, Schulkin J and Havens M (1993) Sex differences insalt preference and taste reactivity in rats. Brain Res Bull32: 91–95.
Frank C L (1988) Diet selection by a heteromyid rodent: roleof net metabolic water production. Ecology 69:1943-51.
Fuller M R, Millspaugh J J, Church K E and Kenward R E (2005)Wildlife radio telemetry. In: Braun C E (ed) Techniques forWildlife Investigations and Management. pp 377-417. 6th edn. TheWildlife Society, Bethesda, MD.
Galef B G Jr, Kennett D J and Stein M (1985) Demonstratorinfluence on observer diet preference effects of simple
84

`
exposure and the presence of demonstrator. Anim learning Behav13: 25-30.
Galef B G Jr, Mason J R, Pretti G and Bean N J (1988) Carbondisulfide: a semiochemical mediating socially-induced dietchoice in rats. Physiol Behav 42: 119-24.
Galef B G Jr and Wigmor S W (1983) Transfer of informationconcerning distant foods: A laboratory investigation ofthe ‘information-centre’ hypothesis. Anim Bahav 31: 748-58.
Galef B G Jr and Whiskin E E (2001) Interaction of social andindividual learning in food preferences of Norway rats.Anim Behav 62: 41-46.
Gebauer U, Dubbel V and Friedrich E (1992) Control of smallrodent pests with spring traps. Allgemeine Forst Zeitschrift 47:408-09.
Gogoi P P and Borah R K (2013) Incidence of lesser bandicootrat, Bandicota bengalensis (Gray) in rice ecosystem in theupper Brahmaputra valley. Indian J Entomol 75(1): 19-22.
Gomez Villafane I E, Bilenca D N, Cavia R, Mino M H, CittadinoE A and Busch M (2001) Environmental factors associatedwith rodent infestations in Argentine poultry farms. BritishPoult Sci 42: 300–07.
Gora S, Sabhlok V P and Pasahan S C (1995) The populationstructure, damage and control measures of rodent in thepoultry farms. Rodent Newsl 19(4): 5.
Gratz N G (1994) Rodents as carriers of disease. In: Buckle AP and Smith R H (ed) Rodent Pests and their Control. pp 85-110.CAB International, Bristol, UK.
Gratz N G (1997) The burden of rodent-borne diseases in AfricaSouth of the Sahara. Belgian J Zool 127: 71–84.
Greaves J H and Rowe F P (1969) Responses of confined rodentpopulations to an ultrasound generator. J Wildl Manage33(2): 409-17.
Hasanuzzaman A T M, Alam M S and Bazzaz M M (2009) Comparativeefficiency of some indigenous traps to capture rats in thewheat field of Bangladesh. J Agric Rural Dev 7(1&2): 121-25.
85

`
Hausmann I (1932) The behaviour of albino rats in choosingfood and stimulants. J Comp Psychol 13: 279-309.
Henderson C F and Tilton E W (1955) Tests with acaricidesagainst brown wheat mite. J Econ Entomol 48: 157-161.
Howard W E (1988) Areas of further research. In: Prakash I(ed) Rodent Pest Management. pp 451-58. CRC Press, Inc BocaRaton, Florida, USA.
Howard W E and Marsh R E (1970) Olfaction in rodent control.Proc 4th Vertebr Pest Conf. pp 64-70. Sacramento, California.
Howard W E, Marsh R E, Palmateer S D (1972) Rat acceptance ofdifferent sugar concentrates in water baits. Int Pest Contr11: 17–20.
Hugh-Jones M E, Hubbert W T and Hagstad H V (1995) Zoonoses:Recognition, Control and Prevention. pp 369. Ames, Iowa StateUniversity Press.
Hussain I and Iqbal M A (2002) Occurrence of rodent filth ingrain commodities sampled from ration shop, Rawalpindi.Pakistan J Zool 34: 239-42.
Hussain I, Ahmed S B and Hassin D (2006) Importance of rodentsin commercial poultry farming and their control measures.Poult Planner 2: 16-20.
Imaizumi M, Takeda M, Suzuki A, Sawano S and Fushiki T (2001)Preference for high-fat food in mice: fried potatoescompared with boiled potatoes. Appetite 36(3): 237–38.
Islam Z and Karim A N M R (1995). Rat control by trapping indeepwater rice. Int J Pest Manage 41: 229–33.
Jackson W B (1965) Feeding patterns in domestic rodents. PestContr 33: 12-15.
Jackson W B (1977) Evaluation of rodent depredation to cropand stored products. EPPO Bull 7: 439.
Jackson W B (1990) Rats and mice. In: Mallis A (ed) Handbook ofPest Control. pp 9-85. 7th edn, Chapter l. Franzak and Foster,Cleveland, USA.
Jacob J, Ylonen H, Runcie M J, Jones D A and Singleton G R(2003) What affects bait uptake by house mice inAustralian grain fields? J Wildl Manage 67: 341–51.
86

`
Kamal N Q and Hossain M M (2003) Comparison of different baitsto attract rats to traps in rice fields in Bangladesh. In:Singleton G R, Hinds L A, Krebs C J and Spratt D M (eds)Rats, Mice and People: Rodent Biology and Management. pp 281-83.ACIAR Monograph No. 96, Canberra, Australia.
Karasawa K and Muto S (1978) Taste preference and aversion forsourness in rats. Japanese J Nutrition 36: 181–86.
Kassa H and Jackson W B (1984) Bait acceptance andchemosterilant efficacy of alpha-chlorohydrin in theNorway rats, Rattus norvegicus. Int Pest Contr 26: 7.
Kaur H and Parshad V R (2005) Laboratory and field evaluationof three odorant compounds for improving attraction of thelesser bandicoot rat, Bandicota bengalensis (Gray) to 0.0375%coumatetralyl bait. Int BiodeterBiodegr 56(3): 135-42.
Kaur G and Parshad V R (2006a) Evaluation of appropriatemethod for application of conspecific urine in plain andpoison bait for acceptance for the control of Bandicotabengalensis (Gray and Hardwicke) in the fields. Environ Ecol24(4): 747-53.
Kaur G and Parshad V R (2006b) Effect of conspecific urine forimproving the efficacy in trapping of rodents. Environ Ecol24(4): 765-67.
Kaur G, Kocher D K, Sandhu H S and Brar R S (2008) Sexspecific value of oral LD50 of cholecalciferol (vitaminD3) against house rat (Rattus rattus). Toxicol Int 15: 143-44.
Khan J A (1974) Laboratory experiments on the food preferencesof the black rat Rattus rattus (Linn). Zool J Linn Soc 54: 1667-84.
Khan J A, Sohail A and Yazdani S (2000) Significance ofadditives to enhance poison baits acceptance against fieldrats in rice paddy in central Punjab, Pakistan. Int J Agric Biol2: 74-76.
Kandhwal S (2009) Evaluation of bait carrier for Rattus Rattus L.infesting commercial poultry facilities in India: A steptowards sustainable poultry management. Int J Arts Sci 3(1): 50-60.
87

`
Khatri A K and Veda O P (1984) Note on the incidence of houserat, Rattus rattus (Gray) in poultry farms in Jabalpur. PoultAdviser 17(3): 47.
Khokhar A R, Pervez A, Rizvi W A and Ismail S (1999) Rodentidentification and its control in a plantation forest atPasni, Balochistan. Pakistan J Zoo1 31: 65-69.
Koehler A E, Tobin M E and Sugihara R T (1994) Effects of CS2
starch xanthate on consumption by rats. Proc Vertebr Pest Conf16: 113-17.
Kok A D, Daniel M, Parker D M and Bark N P (2013) Rules ofattraction: The role of bait in small mammal sampling athigh altitude in South Africa. Afr Zool 48(1): 84-95.
Kolody N, Brosvic G M, Pak D and Loeffler S (1993) Tastepreference behavior in Long-Evans rats and Egyptian spinymice. Bull Psychonomic Soc 31: 307–10.
Krebs C J and Boonstra B (1984) Trappability estimates formark-recapture data. Canadian J Zool 62: 2440-44.
Kumar D and Gangwar S K (2001) Laboratory evaluation ofvarious bait foods against Bandicota bengalensis (Gray) inmid-altitude hills of North-East region Indian J Plant Prot 29:101-05.
Kumar V P and Khan J A (1978) Food preference of the gerbils,Tatera indica Hardwicke. Zool J Linn Soc 64: 51-58.
Kumari S and Prakash I (1980) Role of conspecific urine inenhancing food consumption and masking of shynessbehaviour in the desert gerbil, M. hurriane (Jerdon). Indian JExp Biol 18: 730-32.
Kumari S and Prakash I (1988) Relative efficacy of male andfemale conspecific urine in masking shyness behaviour inIndian gerbil, Tatera indica. Indian Acad Sci (Anim Sci) 93: 431-36.
Laland K N and Plotkin H C (1991) Excretory depositssurrounding food sites facilitate social learning of foodpreferences in Norway rats. Anim Behav 41: 997-1005.
Lund M (1994) Commensal rodents. In: Buckle A P and Smith R H(ed) Rodent Pests and their Control. pp 23-43. CAB International,Bristol, UK.
88

`
Lund M (2008) Rodent behaviour in relation to baitingtechniques. EPPO Bull 18: 185-193.
Macdonald D W, Mathews F and Berdoy M (1999) The behaviour andecology of Rattus norvegicus: from opportunism to kamikazetendencies. In: Singleton G R, Hinds L A, Leirs H andZhang Z (ed) Ecologically-based Rodent Management. pp 49-80.ACIAR Monograph No 59, Canberra, Australia.
Malhi C S and Kaur K (1995) Food preference behaviour of thefive-striped squirrel, Funambulus pennanti Wroughton. BehavProcess 34: 55-66.
Malhi C S, Parshad V R and Ahmad N (1991) Integrated approachof rodent control and its economics in poultry farm. IndianPoult Rev 22: 21–26.
Malhi C S and Sheikher C (1989) Some social and ecologicalconsideration for rodent control in District Pauri ofGarhwal Himalayas. Trop Pest Manage 35(1): 67-69.
Marsh R E (1986) Principles and techniques of formulatingeffective rodent baits: present and future. In: HelmyMohammed A H, Zagloul T M, Salit A M and Zakaria M (ed)Proc 2nd Symp Recent Adv Rodent Contr. pp 211–23. Ministry ofHealth, Kuwait.
Marsh R E (1988) Bait additives as a means of improvingacceptance by rodents. EPPO Bull 18: 195-202.
Marsh R E (1994) Roof rats. pp 125-33. In: Prevention and Control ofWildlife Damage. Great Plains Agricultural Council WildlifeResources Committee and University of Nebraska Instituteof Agriculture and Natural Resources.
Mathur M, Jain A P, Kashyap N and Parveen F (1992) Studies onbait preference and acceptance of flocoumfen in Rattus rattusinfesting poultry farms and godowns. In: Borrecco J E andMarsh R E (ed) Proc 15th Vertebr Pest Conf. pp 178-81. Universityof California, Davis.
Matschke G H, Fagerstone K A, Halstead N D, LaVoie G K andOtis D L (1982) Population reduction of Richardson’sground squirrels with zinc phosphide. J Wildl Manage 46: 674-77.
89

`
McFadden I (1984) Composition and Presentation of Baits and their Acceptanceby Kiore (Rattus exulans). Technical Report No. 7, New ZealandWildlife Service.
Meehan A (1984) Rats and Mice. pp 383. Rentokil Limited, EastGrinstead, UK.
Meerburg G and Kijlstra W A (2008) The ethics of rodentcontrol. Pest Manage Sci 64(12): 1205-11.
Meerburg G M, Singleton G R and Kijlstra A (2009) Rodent-bornediseases and their risks for public health. Critical RevMicrobiol 35: 221–70.
Munjal K (2000) Investigations on the ecology and control of house rat, Rattusrattus L. Ph.D. thesis, Kurukshetra University, Kurukshetra.
Mushtaq M, Hussain I, Mian A, Munir S, Ahmed I and Khan AA(2013). Field evaluation of some bait additives againstIndian crested porcupine (Hystrix indica) (Rodentia:Hystricidae). Integr Zool 8: 285-92.
Naeem M, Ahmad I, Hussain I and Ahmedani M S (2011)Performance of taste enhancers mixed with cereal bases andevaluation of the most preferred bait composition forBandicota bengalensis (Gray). Afr J Biotech 10(19): 3938-44.
Naganuma K, Takashi S and Ya S I (1973) Studies on the foodhabits of rats: feeding preferences of Norway rats on someexperienced diet. Japaneese J Sanit Zool 24: 23-26.
Neelanarayanan P, Nagarajan R and Kanakasabai R (1995) Tanjorebow trap: an eco-friendly tool for rodent management.Rodent Newsl 19: 8–11.
Ohara I and Naim M (1977) Effects of monosodium glutamate oneating and drinking behavior in rats. Physiol Behav 19: 627–34.
Owan P E (1978) Poison and bait shyness in two species ofgerbil, Meriones hurrianae and Tatera indica. Zeitschrift furangewandte Zoologie 65: 57-68.
Owan P E, Srihara K and Sridhara S (1979) A preliminary studyon bait shyness in Bandicota bengalensis towards zincphosphide. Curr Sci 48: 85-86.
90

`
Pai H H, Hong Y J and Wang C H (2005) A community-basedsurveillance on determinants of rodent infestation. ToxicolRev 24(4): 259-69.
Palis F G, Singleton G, Sumalde Z and Hossain M (2007) Thesocial and cultural dimensions of rodent pest management.Integr Zool 2: 174-83.
Pank L F (1976) Effects of bait formulations on toxicantlosses and efficacy. Proc 7th Vertebr Pest Conf. pp 196–202.University of California, Davis, USA.
Parshad V R (1999a) Rodent control in India. Integ Pest Manage Rev4: 97-126.
Parshad V R (1999b) Rodent Management in Poultry Farms. TechnicalBulletin No. 8, pp 11. All India Coordinated ResearchProject on Rodent Control, Central Arid Zone ResearchInstitute, Jodhpur, Rajasthan.
Parshad V R, Ahmad N and Chopra G (1987) Deterioration ofpoultry farm environment by commensal rodents and theircontrol. Int Biodeter 23: 29-46.
Parshad V R, Ahmad N and Malhi S (1991) Control of Rattus rattusin poultry farms. Indian Poult Rev 22: 37-40.
Parshad V R and Jindal S (1991). Multi-choice food preferencebehaviour of the Indian mole rat, Bandicota Bengalenseis. BehavProcess 24: 59-70.
Parshad V R, Kaur M and Singla N (2006) Species-specificdifferences in live and kill trapping of rodents in fodderberseem (Trifolium alexandrium) and maize (Zea mays) fields.Indian J Ecol 33(1): 40-44.
Pathak A and Saxena Y (1995) A study on the selectivepreference of food additives by Mus musculus (albino). UttarPradesh J Zool 15(2): 141-43.
Pervez A (2007) Laboratory evaluation of some additive poisonbaits for controlling commensal and field rodents. Pakistan JZool 39(1): 35-41.
Pervez A, Ahmad S M, Ahmad S and Rizvi S W A (1999) Thesignificance of additives to enhance poison baitacceptance against rodents damaging paddy in lower Sindh(Pakistan). Pakistan J Zool 31(3): 207-10.
91

`
Pervez A, Ahmed S M, Khan A A and Lathiya S B (2005)Comparative field efficacy of some additive formulatedbaits against rodent pests of wheat crop in Sindh,Pakistan. Pakistan J Zool 37(4): 269-74.
Pervez A, Ahmad S M and Rizvi S W A (2003) Evaluation of someadditives to improve poison bait acceptance in the lesserbandicoot rat, Bandicota bengalensis. Pakistan J Zool 35: 109-13.
Pervez A, Rizvi S W A and Ahmad S M (2000) Laboratoryevaluation of some additives as enhancers for baitacceptance in short-tailed mole rat, Nesokia indica. Pakistan JZool 32(4): 351-54.
Pradhan M S and Talmale S S (2011) A checklist of valid Indianrodent taxa (Mammalia: Rodentia): Online version.Zoological Survey of India, Jabalpur, Madhya Pradesh,India.http://zsi.gov.in/checklist/Valid_Indian_Rodents.pdf.
Prakash I (1976) Rodent Pest Management: Principles and Practices. pp 1-28. Monograph No. 4, Central Arid Zone Research Institute,Jodhpur, Rajasthan.
Prakash I (1988) Rodent Pest Management. pp 486. CRC Press, IncBoca Raton, Florida, USA.
Prakash I, Adavani R, Soni B K and Rana B D (1980) Evaluationof the bait base for the control of common house rat Rattusrattus rufescens (Gray). Z Angew Zool 67: 211-23.
Prakash I, Fitzwater W D and Jain A P (1970) Toxic chemicalsand baits for the control of two gerbils, Meriones hurrianaeJordon and Tatera indica Hardwicke. J Bombay Nat Hist Soc 66: 500-09.
Prakash I and Ghosh P (1992) Rodents in Indian Agriculture. Vol I,Scientific Publishers, Jodhpur, India.
Prakash I and Jain A P (1971) Bait shyness of two gerbils,Tatera indica indica Hardwicke and Meriones hurrianae Jerdon. AnnAppl Biol 69: 169-72.
Prakash I and Mathur R P (1987) Management of Rodent Pests. pp 133.Indian Council of Agricultural Research, New Delhi.
Prakash I and Mathur R P (1988) Rodent problems in Asia. In:Prakash I (ed) Rodent Pest Management. pp 67-84. CRC Press, IncBoca Raton, Florida, USA.
92

`
Prakash I and Mathur R P (1992) Acute rodenticide. In: PrakashI and Ghosh P K (ed) Rodent in Indian Agriculture. pp 497-516.Scientific Publishers, Jodhpur, Rajasthan.
Prakash I, Rana B D and Jain A P (1975) Bait-shyness in threespecies of Rattus. Z Angew Zool 62: 89-97.
Prakash S, Kumar S, Veer V, Gopalan N, Purnanand, Pandey K Sand Rao K M (2003) Laboratory evaluation of fourrodenticides admixed in a cereal-based bait againstcommensal rat, Rattus rattus (L.) (Rodentia: Muridae:Murinae). J Stored Prod Res 39: 141-47.
Prescott C V, El-Amin M and Smith R H (1992) Calciferols andbait shyness in laboratory rats. In: Borrecco J E andMarsh R E (ed) Proc 15th Vertebr Pest Conf. pp 218-33. Universityof California, Davis.
Ramalingaswami V (1994) Resurgence of the plague. Sci Reporter 31:15–17.
Ramirez I (1993) Food deprivation reduces rats’ oilpreference. Appetite 21: 53–67.
Rana B D, Jain A P and Soni B K (1992) Bait preferences. In:Prakash I and Gosh P K (ed) Rodents in Indian Agriculture. pp419-26. Scientific publishers, Jodhpur, Rajasthan.
Rana S A, Rana N, Iqbal M Z and Shahbaz F (2006) Seasonaldemographic variation in bandicoot rats, Bandicotabengalensis (Gray) in irrigated crops. Pakistan J Agric Sci 43: 1-2.
Reidinger Jr R F (1997) Recent studies on flavour aversionlearning in wildlife damage management. In: Mason J R(ed) Repellents in Wildlife Management: Proceedings of a symposium. pp101-20. National Wildlife Research Centre, Colorado StateUniversity, Fort Collins, CO, USA.
Reidinger Jr R F and Libay J L (1981) Size and weight ofcoconuts damaged by rats. Philippine Agriculturist 64(3): 281-86.
Reidinger Jr R F and Mason J R (1983) Exploitablecharacteristics of neophobia and food aversions forimprovements in rodent and bird control. In: Kaukeiner D E(ed) Proc Vertebr Pest Contr Manage Materials: 4th Symp. pp 20–39.American Society for Testing and Materials, Philadelphia,USA.
93

`
Riley A L and Clarke C M (1977) Conditioned taste aversions: Abibliography. In: Barker L M, Best M R and Domjan M (ed)Learning Mechanisms in Food Selection. pp 593-616. Waco, Texas.
Robards G E and Saunders G (1998) Food preferences of housemice (Mus domesticus) and their implications for controlstrategies. Wildl Res 25: 595-601.
Roberts T J (1977) The Mammals of Pakistan. pp 271-75. EarnestBenn Ltd, London.
Robertson H, Saul E and Tiraa A (1998) Rat control onRarotonga: some lessons for mainland islands in NewZealand. Ecol Manage 6: 1-12.
Roonwal M L (1987) Recent advances in rodentology in India. RecZool Surv India. 105: 1-126.
Rowe F P (1961) The toxicity and acceptability of the sodiumsalt of pindone, an anti-coagulant rodenticide to thehouse mouse (Mus musculus L.). J Hyg 59: 335-41.
Rowe F P (1973) Aspects of mouse behaviour related to control.Mammal Rev 2: 58–63.
Rowe F P, Bradfield A and Redfern R (1974) Food preferences ofwild house mice, Mus musculus. J Hyg 73(3): 473-78.
Rzoska J (1953) Bait shyness, a study in rat behaviour. British JAnim Behav 1: 128-35.
Salmon T and Marsh R (1989) California ground squirreltrapping influenced by anal-gland odors. J Mammal 70: 428-31.
Santra K B and Manna C K (2008) Studies of some aspects ofrodent ecology in the four districts of the Gangetic plainof West Bengal, India. Univ J Zool Rajshahi Univ 27: 85-90.
Saxena Y, Jhalani S and Aziz Q (1995) Laboratory studies onsome new additives as attractants against Mus musculus. UttarPradesh J Zool 15: 89–90.
Saxena Y and Mathur A (1995) Bait-shyness and poison-aversion:a study in Mus musculus albino. Uttar Pradesh J Zool 15: 11–14.
Schisla R M, Hinchen J D and Hammann W C (1970) Newrodenticide, ‘MR-100’, containing a taste enhancer. Nature228: 1229–30.
94

`
Selvaraj R and Archunan G (2002) Role of male scent glands inimproving poison bait acceptance in female rats, Rattusnorvegicus. Indian J Exp Biol 40: 53-57.
Shafi M M (1991) Some approaches to enhance poison bait acceptance incommensal and field rodents of economics importance in Pakistan. Ph.D.thesis, University of Karachi, Karachi.
Shafi M M, Ahmad S, Ahmad S M and Pervez A (1991) Tasteadditives improve poison bait acceptance in Indian gerbil,Tatera indica. J Plant Prot Tropics 8: 77-82.
Shafi M M, Ahmad S and Pervez A (1992a) The influence of someadditives on poison acceptance in short tailed mole rat,Nesokia indica. Int J Trop Agri 10: 54-59.
Shafi M M, Ahmad S, Pervez A and Ahmad S M (1992b) Tasteenhancers improve poison bait acceptance in field rodentdamaging wheat crop. Trop Pest Manage 38: 214-17.
Shafi M M, Ahmad S M, Pervez A and Ahmad S (1992c) Enhancementof poison bait acceptance through taste additives in Rattusnorvegicus. J Stored Prod Res 28(4): 239-43.
Shafi M M, Pervez A, Ahmad A and Ahmed S M (1990) Role ofsome taste additives to enhance poison bait acceptance inthe black rat, Rattus rattus L. Trop Pest Manage 36(4): 371-374.
Shafi M M, Pervez A, Ahmad S and Ahmad S M (1993) Someapproaches to enhance poison bait acceptance in the lesserbandicoot rat, Bandicota bengalensis. Trop Sci 33: 350-58.
Shepherd D S and Inglis I R (1993) Toxic bait aversions indifferent rat strains exposed to an acute rodenticide. JWildl Manage 57: 640-47.
Shimizu T, Takayanagi M A and Kusano T (1980) Taste preferenceand licking of thallous nitrate by rats and mice. J Pestic Sci5: 403-06.
Shumake S A (1978) Food preference behaviour in birds andmammals. In: Bullard R W (ed) Flavour Chemistry of Animal Foods. AC S Symposium Series No. 67.
Shumake S A, Hakim A A and Gaddis S E (2002) Carbon disulphideeffects on pre-baited vs. non-pre-baited rats exposed tolow dosage zinc phosphide rodenticide bait. Crop Prot 21:545-50.
95

`
Siddiqui J A and Khan J A (1982) Food preferences of the softfurred field rat, Rattus meltada pallidior Ryley. Z Angew Zool69: 107-19.
Singh R (2003) Scientists bring new twist to ‘death bychocolate’ with chocolate flavoured mousetrap.www2.warwick.ac.uk/about/warwickmagazine3/newsinbrief.
Singla L D, Singla N, Parshad V R, Juyal P D and Sood N K(2008a) Rodents as reservoir of parasites in India. IntegrZool 3: 21-26.
Singla L D, Singla N, Parshad V R, Sandhu B and Singh J (2003)Occurrence and pathomorphological observations on Cysticercusfasciolaris in bandicoot rats in India. In: Singleton G R,Hinds L A, Krebs C J and Spratt D M (eds) Rats, Mice andPeople: Rodent biology and management. pp 57-59. ACIAR MonographNo. 96, Canberra, Australia.
Singla N and Babbar B K (2010) Rodent damage andinfestation in wheat and rice crop fields: Districtwise analysis in Punjab State. Indian J Ecol 37: 184-88.
Singla N and Babbar B K (2012) Critical timings of rodenticidebait application for controlling rodents in sugarcanecrop sown in Punjab, India. Sugar Tech 14(1): 76-82.
Singla N, Babbar B K and Parshad V R (2008b) Effect offeeding crude cotton seed oil containing gossypol onfertility of male Rattus rattus and Bandicota bengalensis. PestManage Econ Zool 16(2): 197-205.
Singla N and Garg M (2013) Effect of crude cottonseed oilcontaining gossypol on fertility of male and estrouscycle of female Bandicota bengalensis Gray and Hardwicke. JAppl Anim Res 41: 156-65.
Singla N, Kaur G, Babbar B K and Sandhu B S (2013) Potentialof triptolide in reproductive management of house rat,Rattus rattus (Linnaeus). Integr Zool 8: 260-76.
Singla N, Kaur R and Parshad V R (2010) Efficacy of sodiumchloride as a taste additive for improvement in baitacceptance by rodents. Ann Arid Zone 48(2): 165-67.
Singla N and Mittal M (2012) Management of post control rodentpopulation rebuildup in sugarcane. J Res Punjab Agric Univ 49:60-64.
96

`
Singla N and Parshad V R (2000) Management of post controlrodent population rebuild-up with alpha-chlorohydrin inrice fields. Pestology 24: 36-41.
Singla N and Parshad V R (2009) Laboratory acceptance andefficacy of alpha-chlorohydrin against the Indian gerbil,Tatera indica Hardwicke. Int Pest Contr 51(1): 36, 37-40.
Singla N and Parshad V R (2010) Efficacy of acute andanticoagulant rodenticide baiting in sugarcanefields of Punjab, India. Int J Pest Manage 56: 201-10.
Singla N, Singla L D, Gupta K and Sood N K (2012) Pathologicalalterations in natural cases of Capillaria hepatica infectionalone and in concurrence with Cysticercus fasciolaris inBandicota bengalensis. J Parasitic Dis 17: 16-20.
Singleton G R (2003) Impacts of Rodents on Rice Production in Asia. IRRIDiscussion Paper Series No. 45. International RiceResearch Institute, Los Baños, Philippines.
Singleton G R, Leirs H, Hinds L A and Zhang Z (1999)Ecologically-based management of rodent pests –reevaluating our approach to an old problem. In: SingletonG R, Hinds L A, Leirs H and Zhang Z (ed) Ecologically-basedManagement of Rodent Pests. pp 17-29. Australian Centre forInternational Agricultural Research, Canberra, Australia.
Singleton G R and Petch D A (1994) A review of the Biology andManagement of Rodent Pests in Southeast Asia. ACIAR Technical ReportNo. 30, pp 65. Australian Centre for InternationalAgricultural Research, Canberra, Australia.
Sivaprakasam C and Durairaj G (1995) Laboratory evaluation ofbait base for the control of Indian field mouse, Musbooduga (Gray). Indian J Exp Biol 33: 497-99.
Smith R H (1994) Rodent control methods: non-chemical and non-lethal chemical. In: Buckle A P and Smith R H (ed) RodentPests and their Control. pp 109–25.CAB International, Bristol,UK.
Smythe W R (1976) Criteria for rodent bait selection. Proc 7th
Vertebr Pest Conf. pp 211-14.
Soni B K and Rana B D (1988) Infestation pattern and controlof rodents in poultry farms. Pestology 9(9): 5-7.
97

`
Soni G R and Prakash I (1987) Effect of conspecific urineodour on behaviour of soft-furred field rat, Rattus meltada.Behav Process 14: 175-82.
Sood M L and Gill R P S (1980) Bait shyness and poisonaversion in Bandicota bengalensis using Rh-787 as rodenticide.J Bombay Nat Hist Soc 77: 319-20.
Sridhara S and Krishnamurthy T R (1992) Population dynamics ofRattus rattus in poultry and implications for control. In:Borecco J E and Marsh R E (ed) Proc 15th Vertebr Pest Conf.University of California, Davis.
Sridhara S and Tripathi R S (2005) Distribution of Rodents in IndiaAgriculture. pp 136. Central Arid Zone Research Institute,Jodhpur.
Srihari K and Chakravarthy A K (1992) Cardamom. In: Prakash Iand Ghosh P K (ed) Rodents in Indian Agriculture. pp 289–308.Scientific Publishers, Jodhpur, India.
Srivastava V and Srivastava R C (1985) Trapping rodents withglue. Indian J Agric Sci 55: 385–86.
Stenseth N C (1977) On the importance of spatiotemporalheterogenecity for the population dynamics of rodentstowards a theoretical foundation of rodent control. Oikos29(3): 545-52.
Sterner R T, Ramey C A, Edge W D, Manning T, Wolff J O andFagerstone K A (1996) Efficacy of zinc phosphide baits tocontrol voles in alfalfa-an enclosure study. Crop Prot 15:727-34.
Steven R B (2008) Ecological rat management. Int Pest Contr 50:27.
Stoddart D M (1982) Demonstration of olfactory discriminationby short tailed vole Microtus agrestis. Anim Behav 30: 293-94.
Strupp B and Levitsky D A (1984) Social transmission of foodpreferences in adult hooded rats (Rattus norvegicus). J CompPsychol 98: 257-66.
Sullivan L M (2002) Roof Rat Control around Homes and other Structures.Cooperative Extension, University of Arizona.
Szocs G, Toth M, Karpati Z, Zhu J, Lofstedt C, Plass E andFrancke W (2004) Identification of polyenic hydrocarbons
98

`
from the northern winter moth, Operophtera fagata, anddevelopment of a species specific lure for pheromonetraps. Chemecology 14: 53-58.
Tabuchi E, Ono T, Uwano T, Takashima Y and Kawasaki M (1991)Rat preference for food related odors. Brain Res Bull 27: 387-91.
Tanaka R (1963) On the problem of the trap response types ofsmall mammal populations. Res Population Ecol 5:139-46.
Taylor K A (1984) Vertebrate pest problems in Africa. Proc ConfOrg Pract Vertebr Pest Contr. pp 662. Imperial ChemicalIndustries PLC, Surrey, England.
Timm R M (1994) House mice. pp B31–B46. In: Hygnstrom S E,Timm R M and Larson G E (ed) Prevention and Control of WildlifeDamage. Vol 1. University of Nebraska CooperativeExtension, Lincoln.
Tobin M E (1992) Rodent damage in Hawaiian macadamia orchards.Proc Vertebr Pest Conf 15: 272-76.
Tobin M E and Fall M W (2006) Pest control: rodents inagricultural sciences. Encyclopedia of Life Support System (EOLSS),Eolss publishers, Oxford, UK. http://www.eolss.net.
Tobin M E, Matschke G H, Susihara R T, McCann G R, Koehler A Eand Andrews K J (1993) Laboratory Efficacy of Cholecalciferol againstField Rodents. Denver Wildlife Research Report No. 11-55-002.pp 13. United States Department of Agriculture, Animal andPlant Health Inspection Service.
Tobin M E and Richmond M E (1993) Vole Management in Fruit Orchards.Biological Report 5. pp 18. United States Fish andWildlife Service Technical Report Series.
Tobin M E, Sugihara R T and Ota A K (1990) Rodent damage toHawaiian sugarcane. Proc Vertebr Pest Conf 14: 120-23.
Tobin M E, Sugihara R T and Koehler A E (1997) Bait placementand acceptance by rats in macadamia orchards. Crop Prot 16:507-10.
Turner C E J (1986) Structural and litter pests. Poult Sci 65(4):644-48.
Vander Wall S B, Beck M J, Briggs J S, Roth J K, Thayer T C,Hollander J L and Armstrong J M (2003) Interspecific
99

`
variation in the olfactory abilities of granivorousrodents. J Mammal 84: 487-96.
Wagner M W (1971a) Comparative rodent preferences forartificial sweeteners. J Comp Physiol Psych 75: 483-90.
Wagner M W (1971b) Species variability and rodent aversionsfor three sugars. Psychol Rep 29: 799-804.
Watkins R W, Whiterow A, Bull D S and Cowan D P (1999) The useof familiar odors to reduce the impact of containerneophobia on the control of Norway rats (Rattus norvegicus).In: Johnson R E, Muller-Schwarze D and Sorensen P W (ed)Advances in Chemical Signals in Vertebrates. pp 655-61. PlenumPress, NY, USA.
Watson J S and Perry J S (1954). Control of Rats and Mice II. In:Chitty D (ed) Clarendon Press, Oxford.
Weihong J, Veitch C R and Craig J L (1999) An evaluation ofthe efficiency of rodent trapping methods: the effect oftrap arrangement, cover type and bait. New Zealand J Ecol 23:45–51.
White J, Wilson J and Horskins K (1997) The role of adjacenthabitats in rodent damage levels in Australian macadamiaorchard systems. Crop Prot 16: 727-32.
Wiener J G and Smith M H (1972) Relative efficiencies of foursmall mammals traps. J Mammal 53: 868-73.
Williams J M (1974) The effect of artificial rat damage oncoconut yields in Fiji. PANS 20: 275-82.
Wines M (2004) For sniffing out land mines, a platoon oftwitching noses. The New York Times, 18 May, 2004.
Witmer G, Burke P and Jojola S (2008) An evaluation ofeffectiveness of potential Norway rat attractants. In:Timm R M, Madon M B and Davic C A (ed) Proc 23rd Vertebr PestConf. pp 35-38. San Diego, California.
Witmer G, Fall M and Fiedler L (1995) Rodent control, researchneeds, and technology transfer. In: Bissonette J (ed)Integrating People and Wildlife for a Sustainable Future. pp 693-97. TheWildlife Society, Bethesda, MD.
100

`
Witmer G, Snow N P and Burke P W (2010) Potential attractantsfor detecting and removing invading Gambian giant pouchedrats (Cricetomys gambianus). Pest Manage Sci 66: 412-16.
Wood B J (1994) Rodents in agriculture and forestry. In:Buckle A P and Smith R H (ed) Rodent Pests and their Control. pp45-83. CAB International, Bristol, UK.
Yamaguchi K (1995) The taste sensitivity preference for fivebasic tasting substances. Nippon Kasei Gakkaishi 46: 1037-45.
Ylonen H, Mappes T and Viitala J (1990) Different demographyof friends and strangers: an experiment on impact ofkinship and familiarity in Clethrionomys glareolus (Schreb).Oecologia 83: 333-37.
Young P T (1946) Studies of food preferences appetite anddietery habit. J Comp Psychol 39: 139-76.
101




















