
Cardiac changes during arousals from non-REM sleep in healthy volunteers

lable at ScienceDirect
Sleep Medicine Reviews xxx (2011) 1e14
Contents lists avai
Sleep Medicine Reviews
journal homepage: www.elsevier .com/locate /smrv
HISTORICAL REVIEW
Forerunners of REM sleep
Hartmut Schulz a,*, Piero Salzarulo b
aDepartment of Educational Science and Psychology, Free University Berlin, Berlin, GermanybDepartment of Psychology, University of Florence, Florence, Italy
a r t i c l e i n f o
Article history:Received 12 April 2011Received in revised form27 May 2011Accepted 28 May 2011Available online xxx
Keywords:Sleep statesREM sleepRapid eye movementsHistory of sleep science
Abbreviations: EEG, electroencephalogram; KWNREM, non rapid eye movement; REM, rapid eye mo* Corresponding author. Tel.: þ49 361 22 532 07; f
E-mail address: [email protected] (H. Schul
1087-0792/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.smrv.2011.05.006
Please cite this article in press as: Schuj.smrv.2011.05.006
s u m m a r y
The development of sleep research can be divided into two main periods. The first one was initiated in1863 by the first systematic measurement of the depth of sleep, the second in 1953 by the discovery ofrecurrent episodes of rapid eye movements in sleep. The main methodological procedure in the first ofthese two periods was the measurement of a single physiological variable, while beginning with long-term measurements of the electroencephalogram (EEG) in sleep, multi-channel, polygraphic recordingbecame the method of choice for sleep studies. Although rhythmic changes in the ultradian frequencyrange of one to 2 h were observed early in many variables during sleep (movements, autonomic func-tions, penile erections), the recognition of the existence of two different states of sleep (rapid eyemovement (REM) and non-rapid eye movement (NREM sleep)) was contingent upon a ‘synthetic’ view,which focus on the coalescence of multiple variables. The dual concept of sleep organization evolvedstepwise in parallel to the rapid growth of neurophysiological knowledge and techniques in the first halfof the 20th century, culminating in the discovery of REM sleep.
� 2011 Elsevier Ltd. All rights reserved.
Introduction
Mammalian sleep is composed of two different states of sleep,REM and NREM sleep, which recur in alternating bouts or cycles.The starting point for this conception of sleep was the recognitionof recurrent episodes of rapid eye movements by Aserinsky andKleitman in 1953.1 They found, in addition, that respiratory rate andbody motility are increased at times when rapid eye movementsoccur. With these indicators at hand, they immediately probed fora possible relationship between the occurrence of rapid eyemovements and dreaming. The assumption was confirmed andthe authors concluded: “the fact that these eye movements, EEGpattern, and autonomic nervous system activity are significantlyrelated and do not occur randomly suggests that these physiolog-ical phenomena, and probably dreaming, are likely all manifesta-tions of a peculiar level of cortical activity which is encounterednormally during sleep.”1, p. 274 Two years later the observationswere confirmed in larger samples of adults2 and infants.3 Anothertwo years later Dement and Kleitman4 demonstrated that rapid eyemovements occurred selectively in what they called stage 1 sleep,
I, Kaiser Wilhelm Institute;vement.ax: þ49 361 22 532 08.z).
All rights reserved.
lz H, Salzarulo P, Forerunn
which was defined by an absolute lack of spindle activity. A finaland decisive step to establish sleep with rapid eye movements asa peculiar sleep state, different from other states of sleep, was madewhen Jouvet et al.5 observed periodic abolition of tonic neckmuscleactivity in decorticated and mesencephalic cats together withmiotic pupils and movements of the vibrissae. In normal cats thisparadoxical state of sleep, as Jouvet preferred to call the state ofREM sleep, was associated with high frequency, low voltage corticalactivity, and with an increased arousal threshold to either auditoryor reticular stimulation.5 A loss of muscle tone during phasesof REM sleep was confirmed also in human sleepers.6, p. 292, 7 Thediscovery of REM sleep by Aserinsky and Kleitman in humans, andof the corresponding state of paradoxical sleep in cats by Jouvetet al. represents a turning point in sleep research. The recognitionof REM sleep as a singular sleep state fully supported the concept ofsleep as an actively regulated physiological process and thusbecame the starting point for systematic studies on all aspects ofsleep8,9 and its disorders.10
Why had the complex structure of sleep with two functionallydifferent kinds of sleep remained unrecognized for so long,although sleep, and the associated process of dreaming, were ofprimary interest since ever? In the following, we will trace thedevelopment of reasoning on sleep and observations on the phys-iology of sleep with special reference to those observations witha hunch for what was established later as REM sleep. We will alsodiscuss some conditions which prevented an earlier recognition of
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e142
the structure of sleep as it is known today. The present review islimited mainly to the European and US literature. The main reasonfor neglecting literature from other regions is our linguistic inabilityto search for relevant documents in the respective languages.
The idea that there may be two different kinds of sleep waspresent in the literature since ever. By observing sleeping animalsand human subjects, quiet, deep and dreamless sleep was separatedfrom more restless sleep with dreams.11, 9, p. 212 The first type ofsleep was located early in the night (primo somno in Latin) the lattertype later in night sleep. The French medical doctor Jean Fernel inhis treatise on physiology12 made a difference between a state ofdeep sleep (sommeil profond) without dream remembrance, anda medium state of sleep, accompanied by dreaming (sommeil pro-pice aux songes).13 In 1852 Lélut wrote: “A distinction which seemsto arise by itself and which is universally recognized, is thedistinction of complete sleep, being free of dreams, and incompletesleep, so to say mixed with dreams, or more briefly the distinctionof sleep and dream”14, p. 10 [Appendix 1]. In the same vein Lié-beault15 distinguished between light and deep sleep. Deep sleepcould appear in two forms, either with a slowing of ideas, an arrestof sensory and motor functions, or with ideas which are strongenough to activate the motor system and part of the sensory system,resulting in somnambulism, however, without memory of dreams.In light dreaming sleep, on the other hand, the sensory systems arenot shut but damped, and the muscles relaxed. In this state dreamsare never translated into muscular actions since the ideas are lessclear, changing and thus they have less energy. This form of sleepregularly leads to memory of dreams, especially those whichoccurred before awakening15, p. 25 [Appendix 2]. Kohlschütter, whohad performed first experiments on the depth of sleep a few yearsearlier,16 supposed that dreaming is a special state which may differfrom the general slowing of circulation of the brain (“Hirncircula-tion”) during sleep and may rather resemble that during wakeful-ness17, p. 46 [Appendix 3]. Studies in the last two decades, usingimaging techniques, have shown indeed that cerebral blood flow inREM sleep is always higher than in corresponding NREM sleep,18,19
and changes in flow and regional differences in blood supply may beassociated with dreaming.20
MacWilliam, an Aberdeen physiologist, also recognized twodifferent “conditions“ of sleep with “strikingly different associationsas far as nervous, circulatory, respiratory, and other functions areconcerned: 1) undisturbed or sound sleep, attended by lowering ofblood pressure, heart and respiratory rates, etc., and 2) disturbedsleep, modified by reflex excitations, dreams, nightmare, etc.,sometimes accompanied by extensive rises of blood pressure(hitherto not recognized), increased heart action, changes in respi-ration, and various reflex effects”.21, p. 1197 In a paragraph devoted tothe Dreaming State of the Dog MacWilliam described phenomenawhichhehadobserved in thedog, adding towhathehadobserved inhumans. In dogs “during dreams of hunting, etc., with the familiarmovements of toes and paws, tail and ears, biting action, series ofsubdued barks, etc.” he gave a quite precise account of physiologicalprocesses, as far as this was possible by a keen eye and hands-ontesting. “The heart is often rapid and irregular with inhibitorypauses, bouts of acceleration, etc., while a violent cardiac impulsemay be perceived; respiration is frequently hurried and irregular,with grasps, etc. The knee-jerk may be increased” e as Lombardnoted in man during a dream of active movement. It has not beenfound practicable to get actualmeasurements of blood pressure thatare satisfactory, for disappearance of the changes present in thedreaming state is very quickwhen the animal awakes. But the fingeron an artery has sometimes given unequivocal evidence of a riseof blood pressure”.21, p. 1198 From these observations he concluded“. that the foregoing facts must be taken as profoundly modifyingthe simple conception of night as the time of rest, and sleep as
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
a condition in which quiescence prevails and recuperative changesgo on, restoring the bodily and mental capacities which havebecome more or less reduced at the end of the hours of work andwakefulness - . This conception, while true as regards undis-turbed or sound sleep, has to be qualified by the consideration thatnight and sleep are occasionally the season of acute reflex andemotional disturbances which, in the peculiar conditions present,induce very pronounced effects on the circulatory system,throwing a formidable strain upon its weak points, whether thesebe cardiac, with susceptibility to fibrillation or anginal pain, etc., orarterial, with risk of rupture”.21, p. 1199 He envisaged not only “twodifferent conditions of sleep” but became aware also of specificregulatory properties of this active state of sleep (cf. Parmeggiani22),and their potential clinical consequences,which had towait another40 years to become a topic of sleep medicine.
In summary, the concept of two different sorts of sleep wasrepeatedly mentioned in the ancient literature on sleep, enrichedwith more detailed descriptions from the 19th century on. Twomain differences were assumed between the two states, a) thepresence or absence of dreams, and b) whether sleep was disturbedor undisturbed. It was also general knowledge that quiet anddreamless sleep prevails early in the night, while more restlesssleep and dreaming dominates later. The two states correspondroughly to the later division of sleep into deep NREM sleep and REMsleep. However, hints to the cyclic alternation of the two sleepstates appear for the first time only late in the 19th century.
Grading the depth of sleep
Psychophysics is a 19th century concept, although the termwasused occasionally already earlier. The aim of psychophysics is tofind lawful relationships between sensory stimulation and sensa-tion. The theory of psychophysics was formulated by GustavFechner in Elemente der Psychophysik, published in 1860.23 One ofFechner’s students, Ernst Kohlschütter, suggested to measure thedepth of sleep, or firmness (Festigkeit) of sleep, as he called it, byapplying graded acoustic stimuli to arouse and wake up humansleepers.23, vol. 2, p.440 This technique allowed for the first time tostudy the awakening threshold systematically. The result wasa monophasic depth of sleep curve with a rapid increase of thethreshold, reaching a peakwithin the first hour of sleep, followed bya steady decrease for the remaining hours of sleep. Technical issuesand treatment of the observational data in this pioneering investi-gation have been intensively discussed byothers.24 Later sleep depthcurves unanimously confirmed the initial steep rise of the awak-ening threshold, however, they revealed additional local minimaandmaximaof the threshold later in sleep,with inter-peak distancesof 1e3 h. This became especially evident, when curves were estab-lished not by averaging across subjects, but by using single subjectdata (Fig. 1).25e30 Michelson26,27 pointed out that the fluctuationsin the depth of sleep were not caused by external disturbances butresulted from the interaction of internal antagonistic processes, onecatabolic, the other anabolic in nature. He reasoned: “Also duringthe day two different kinds of physiological processes are contin-uously active in parallel in our organism, one which leads toconsumption and one which leads to re-use of the nourishingmaterial in our tissues. Depending upon the special conditions,consumption temporarily outweighs over replacement, and viceversa. . Remember that not only the two antagonistic states ofwakefulness and sleep in the life of our organism are periodicfunctions, but also the vegetative processes of our body in general,circulation, respiration, nutrition, etc. display remarkable period-icity. This in mind, we may get a better understanding of the shownvariations in the depth of sleep, even if its causal relationship withbodily functions is still unknown at present”27, p. 100 [Appendix 4].
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

Fig. 1. a. Depth of sleep curve (subject: O. Mönninghoff). Mean intensity of the acoustic stimuli needed to wake up the sleeper (y-axis) at different times of the night (x-axis). Dottedsections represent missing values. (Redrawn from 25, Table 2, column a) b. Depth of sleep curve (subject: E. Michelson) of night sleep (M. I) and afternoon sleep (M. II, dotted curve).Mean intensity of the acoustic stimuli, needed to wake up the sleeper (y-axis) at different times of night time or daytime sleep (x-axis). (From 27, Fig 1). c. Depth of sleep curve(subject: P. Köhler). Mean intensity of the acoustic stimuli needed to wake up the sleeper (y-axis) at different times of the night (x-axis). The curve was established in imitation ofMichelson’s procedure (1899), however, Köhler appreciated that his experimental setting was much more crude. (From 30, p. 419).
H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e14 3
Ultradian rhythms in sleep
Rhythmic fluctuations in the ultradian frequency range of one to2 h were seen not only as variations of the depth of sleep but also indifferent physiological measures. Howell31 (1897), who studiedvolume changes of the arm during sleep by means of plethysmo-graphic recordings, found three different sorts of changes in sleep,a) a general increase of volume during sleep, presumably caused bya vasodilatation in the arm vessels, b) short-lasting variations involume in connection with either external stimuli, or internalevents such as sighs or body movements, and c) slow variationswith “roughly speaking, an hourly period, the extreme limits ofduration varying perhaps between half an hour and an hour anda half”31, p. 320 (Fig. 2). Howell noted the hourly variations in all ofhis records and regarded them as a characteristic feature of sleep.His tentative interpretation was that the rhythmic increases anddecreases in vasomotor tone may reflect changes in the activity ofthe vasomotor center, controlling the vessels of the skin.
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
Ultradian rhythmic variations in sleep has also been observed inthe temporal distribution of body movements32 (Fig. 3). Compa-rable variations in distribution of body movements were found byZickgraf33 in seismographic recordings from the sleeper’s bed. Inmost cases, periods of body quiescence were associated witha reduction in heart rate (Fig. 4). Short rhythmical componentswere also seen in the electrodermogram, and in the alveolar CO2curve during sleep.34 Fig. 5 shows coordinated regulation ofdifferent physiological functions, namely respiration, electricalactivity of the skin, and movement activity, suggesting rhythmicvariations in the central regulation of these functions. Ultradianvariations in motility during sleep was observed not only in adultsbut also in young infants.35,36, p. 31
A cycle of penile erections
Of special interest is a report on recurrent episodes of penileerections (erectio membri virilis) during sleep by Ohlmeyer et al.37
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

Fig. 2. Plethysmographic record of hand and lower part of forearm. A fall in the curve indicates dilation, a rise constriction. Phases with predominantly dilation alternate withphases of increasing constriction, suggesting ultradian rhythmicity. The short, rapid rises in constriction were associated with external stimuli (noise) or sighs and movements. Adetailed account of all events can be found in the original publication. (From 31, Fig. 2). � The Rockefeller University Press; with permission.
H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e144
The authors documented in five young subjects an erection cyclewith a mean duration of 85.4 min, and a mean active phase of25.3 min. Individual mean periods varied from 79.7 to 102.2 min(Fig. 6). Measurements were repeated in two subjects in differentyears.37, Table 1 To test the stability of the observed periodicity, thetemporal distribution of erections in the tracings was comparedwith a fixed grid (mean value: 85.4 min, variation: � 15%). Thecomparison resulted in a 79% hit rate between observed events andfixed grid intervals, suggesting continuity of the observedphenomenon during sleep.
In addition, the authors showed that spontaneous awakeningfrom sleep, on which subjects made notes, were significantlyassociated with the erection cycle. Finally, when they reanalyzedintervals between minima and maxima of arousability fromMichelson’s experiments,27 they calculated a mean period of82.5 min which was close to the period of the erection cycle, sup-porting their assumption that penile erections and recurrentdispositions to awaken are temporally associated.
A second paper from this group,38 which was submitted onlytwo month later, appeared in print only 1947, necessitated by a haltin the publication of the journal Pflüger’s Archiv of Physiologyaround the end of World War II. The aim of the second seriesof studies was twofold. First, erections were recorded underconditions of delayed and extended sleep. Sleep episodes startedbetween 02:00 h. and 05:00 h. and ended between between10:30 h. and 16:30 h. The results showed that the erection cyclewasnot restricted to night time sleep but continued during daytime
Fig. 3. Time course of relative frequency of movements in sleep. Data from a single subject. Alater in the night. This trend is superimposed by marked differences in relative motility, sh
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
sleepwithout change in periodicity. Second, in a series of long-termrecordings, ranging from 11 to 26 consecutive sleep nights, thehypothesis was tested that erections in sleep where part of anongoing biological rhythm which is continuous during sleep andwakefulness. The authors found support for their hypothesis by thefact that erections in consecutive nights did not occur at the sameclock time but shifted from night to night by a mean value of11.6 min. The findings that the erection cycle a) was ongoing duringdaytime sleep, and b) displayed probably long-term stabilityqualifies the rhythm as an early example of the basic rest activitycycle (BRAC), a concept proposed later by Kleitman.39,40
Halverson,41 who had performed long-term observation duringdaytime hours on penile tumescence inmale infants between 3 and20 weeks of age, reported that when tumescence occurred duringsleep, it “frequently awakened the infant”41, p. 533 corresponding towhat Ohlmeyer and Brilmayer38 had later observed in adults. About20 years later it was Charles Fisher and coworkers who confirmedthe “cycle of penile erections synchronous with dreaming (REM)sleep”, as the title of their first paper on the phenomenon states.42
Finally, Rothfeld43 presented two patients with narcolepticsymptoms who got erections when they became extremely sleepyor when fighting sleep (Rothfeld, cases 2 and 3). Since sleep innarcoleptic patients frequently begins with an episode of REMsleep, the reported erections could be an early sign of REM sleep inthese patients. This assumption is in agreement with the finding byScharf et al.44 that short REM latencies in daytime naps areaccompanied by penile erection.
decrease of motility in the first hour after retiring is followed by an increase of motilityowing an ultradian rhythm with peaks at intervals of about 1.5 h (From32, Fig. 9).
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/
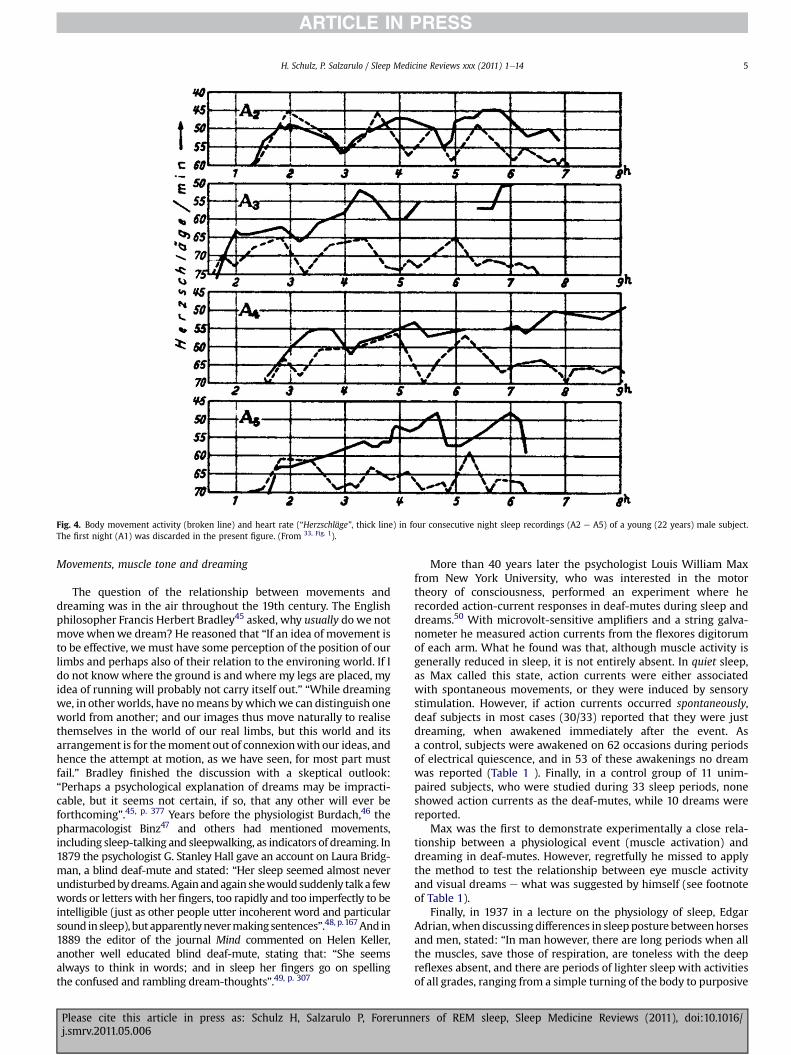
Fig. 4. Body movement activity (broken line) and heart rate (“Herzschläge”, thick line) in four consecutive night sleep recordings (A2 e A5) of a young (22 years) male subject.The first night (A1) was discarded in the present figure. (From 33, Fig. 1).
H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e14 5
Movements, muscle tone and dreaming
The question of the relationship between movements anddreaming was in the air throughout the 19th century. The Englishphilosopher Francis Herbert Bradley45 asked, why usually dowe notmove whenwe dream? He reasoned that “If an idea of movement isto be effective, we must have some perception of the position of ourlimbs and perhaps also of their relation to the environing world. If Ido not knowwhere the ground is and where my legs are placed, myidea of running will probably not carry itself out.” “While dreamingwe, in otherworlds, have nomeans bywhichwe can distinguish oneworld from another; and our images thus move naturally to realisethemselves in the world of our real limbs, but this world and itsarrangement is for themoment out of connexionwith our ideas, andhence the attempt at motion, as we have seen, for most part mustfail.” Bradley finished the discussion with a skeptical outlook:“Perhaps a psychological explanation of dreams may be impracti-cable, but it seems not certain, if so, that any other will ever beforthcoming”.45, p. 377 Years before the physiologist Burdach,46 thepharmacologist Binz47 and others had mentioned movements,including sleep-talking and sleepwalking, as indicators of dreaming. In1879 the psychologist G. Stanley Hall gave an account on Laura Bridg-man, a blind deaf-mute and stated: “Her sleep seemed almost neverundisturbedbydreams.Againandagain shewould suddenly talka fewwords or letters with her fingers, too rapidly and too imperfectly to beintelligible (just as other people utter incoherent word and particularsound insleep), butapparentlynevermakingsentences”.48, p.167And in1889 the editor of the journal Mind commented on Helen Keller,another well educated blind deaf-mute, stating that: “She seemsalways to think in words; and in sleep her fingers go on spellingthe confused and rambling dream-thoughts”.49, p. 307
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
More than 40 years later the psychologist Louis William Maxfrom New York University, who was interested in the motortheory of consciousness, performed an experiment where herecorded action-current responses in deaf-mutes during sleep anddreams.50 With microvolt-sensitive amplifiers and a string galva-nometer he measured action currents from the flexores digitorumof each arm. What he found was that, although muscle activity isgenerally reduced in sleep, it is not entirely absent. In quiet sleep,as Max called this state, action currents were either associatedwith spontaneous movements, or they were induced by sensorystimulation. However, if action currents occurred spontaneously,deaf subjects in most cases (30/33) reported that they were justdreaming, when awakened immediately after the event. Asa control, subjects were awakened on 62 occasions during periodsof electrical quiescence, and in 53 of these awakenings no dreamwas reported (Table 1 ). Finally, in a control group of 11 unim-paired subjects, who were studied during 33 sleep periods, noneshowed action currents as the deaf-mutes, while 10 dreams werereported.
Max was the first to demonstrate experimentally a close rela-tionship between a physiological event (muscle activation) anddreaming in deaf-mutes. However, regretfully he missed to applythe method to test the relationship between eye muscle activityand visual dreams e what was suggested by himself (see footnoteof Table 1).
Finally, in 1937 in a lecture on the physiology of sleep, EdgarAdrian,whendiscussingdifferences in sleepposture betweenhorsesand men, stated: “In man however, there are long periods when allthe muscles, save those of respiration, are toneless with the deepreflexes absent, and there are periods of lighter sleep with activitiesof all grades, ranging from a simple turning of the body to purposive
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

Fig. 5. Body movement activity (“Seismograph”/actography; upper insert),electrodermal activity from the arm (“Arm” EDG) and leg (“Bein” EDG; middle insert),and CO2 curve (“Kurve”,lower insert). Recording time from 08:00 h to 04:00 h(From 34, Fig. 1. With kind permission from Springer Science þ Business Media).
Table 1Relationship between finger action currents and dream reports in deaf-mutesubjects. For details see Max.50
Action current
Present Absent
Dream report Present 30 9a
Absent 3 53
a Max noted: “I do not regard all of these 9 cases as negative, however; in 5 of themthe dream content was primarily visual in nature, and the seat of activity may havebeen in the eye-muscles which at the time were not in the electrical circuit”.50, p. 481
And he added: “A third amplifier, connected to the eye-muscles, might conceivablyhave registered ocular activity during these visual dreams”.50, p. 482.
H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e146
arm movements or the complex pattern of speech”.51, p. 238
Muscle atonia was later recognized as a distinctive feature of REMsleep.52,53
One of the first accounts of a state of active sleep in a younginfant probably comes from Wilhelm Zipperling, a spa doctor, whogot interested in this topic when he became aware in his ownpreterm born child of motor phenomena which affected especiallythe area of the eye and facial muscles.54 His report was on a boywho was born in the 7th post conceptional month. From day 17 on,when the child was breast fed, he observed for the first timedifferent conspicuous behaviors, on which he gave a detailedaccount: “At first there are abrupt and rhythmic horizontaldeflections of both eyes to one side, followed by a much slowerreturn to the resting position, approximately 12e20 times within
Fig. 6. From top to bottom five recordings of erections in night sleep. Duration of the recordipenis, which was connected via an electrical circuit with a kymograph. Erections of a certaideflection in the tracing. The number of episodes with erection varies between three and f
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
¼ e ½ minute. In one attack the deflections are directed to theright, at other time to the left, following no rule. At the same time,the sucking movements become weaker and soon stop completely.Breathing becomes irregular and breaths are separated by varyingpauses. The breaths are jerky, more deeply and faster and occa-sionally inspiration rushes, so that three inspirations follow directlyone another. However, these irregularities of the respiration arevery small and escape easily the superficial observation, since itnever comes to cyanosis, and there was never the slightest sign ofshortness of breath.
The globes move now from their horizontal position upward,either outward or inwards, preferably in the direction of the earlierdeviations of the eyes. At the same time blepharospasm sets in,a rapid sequence of 4e6 eye closure followed by some seconds withlids open and globes remaining in an upward position. Occasion-ally, for fractions of a second there is contraction of the wholem. orbicularis oculi. Meanwhile twitching in the area of the mouthsets in. The corner of the mouth on one side e the same side towhich the globes deflected originally e will be pulled sidewardsand upwards, several times abruptly, then slowly, tonically forseveral seconds, frequently bilateral. In rare cases thewhole patternis accompanied by jerky short backward movements of the head.Otherwise any movements of the remaining body are alwaysmissing. Such an attack, which begins abruptly with the eyesymptoms, associated by those of the facialis, andwhich now eitherbecomes detached or arises at the same time, with completeimmobility of the remaining body, takes to in the maximum caseten full minutes. The attack stops just as suddenly, as it came,muscles involved return into their resting position, the childballoons, takes a deep breath, makes coordinated movements withthe arms and legs and cries in most cases. After a pause of ½ to3 min comes another, usually weaker attack and so on three to fourtimes and still more, until in the maximum case a full hour, onthe average 10e20 min have passed, before everything fadedaway”54, p. 31-32 [Appendix 8].
ngs was up to 6 h per night. Erections were measured by means of a contact ring on then strength resulted in an interruption of the circuit, which is indicated as a downwardour in each tracing. (From 37. With kind permission from Springer Science þ Business Media)
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e14 7
After three months the events became rare, although they wereobserved occasionally with unabated violence. A consultation ofProf. Rietschel of the Dresden nursery came to the result that suchevents are benign and not rare in preterm infants. The observedsymptoms are described as Stäupchen in the older Germanpediatricliterature, or shuttering attacks or benign neonatal sleep myoclonus,which has to be separated from neonatal epileptic seizures. Duringa visit in the Dresden gynecological hospital Zipperling studied 42newborns (age: 1e10 days) and observed symptoms of shutteringattacks (Stäupchen) in 13 of them. Themothers were not alarmed bythe symptoms, and in the literature one can find the term “Ilbambino gioca coi angeli” (“The child playswith the angels.”) for thefacial expression and gestures of the young infants in this state.55
Eye movements in sleep
There are trivial and non-trivial reasons which may have pre-vented the observation of eye movements during sleep for longtime. Trivial reasons are that eye lids normally are closed in sleep,and in most cases the sleeper is lying in the dark. In addition, therewas no reason to expect movements of the eyeballs during sleep,and if there should be some, an observer would most probablyassume that the sleeper is either not yet fully asleep, or that he justwoke up.56
The main non-trivial reason not to become aware of eyemovements in sleep, others than those at sleep onset, before 19531
results from the widespread assumption that the eyes wouldremain in a fixed position during sleep. Most observers, armedwitha candle or flashlight, when carefully opening the eye lids found theeyeballs directed slightly upward, and rotated either inward oroutward. One of the first observers was the Scottish anatomist SirCharles Bell, and the phenomenon bears his name.57 In the secondhalf of the 19th century, physiologists started to investigate ingreater detail the mechanisms of eye closure, the size and reactionof the pupil,58, vol. I, Table 9.7, p. 446 as well as the position of the eyesduring sleep. Slotopolsky,59 who reviewed the literature on theposition of the eyes during sleep, adding own observations inchildren and adults, concluded that the general assumption of Bell’sphenomenon during sleep is not justified. He suggested that inmany cases this specific position of the eyes was probably inducedby manipulations of the observer, while opening the eye lids of thesleeper.
It is of interest that Berger and Loewy60 mentioned, witha reference to Duval,61 that dreams are sufficient to provokedilatation of the pupil, which is otherwise constricted duringsleep60, p. 392 [Appendix 9].
Raehlmann and Witkowski62,63 were the first to describe slowmovements of the eyes at the onset of sleep, which was confirmedby Siemens.64 In addition, occasional deviations in the position ofthe eyeballs during drowsiness and in sleep were recognized.62e65
Witkowski,66, p. 508 described eye movements in sleep as extremelyslow and for the most part not coordinated, frequently evenrestricted to one side. Piéron,67, p. 138 confirmed observations ofRaehlmann and Witkowski62 and Polimanti68 on atypical andindependent eye movements [Appendix 10]. Polimanti himself,who observed Bell’s phenomenon at sleep onset, added that “Indeep sleep the globes do not always remain in this position butmake frequently slow or rapid movements in all directions, whichcan be attributed to dreams”68, p. 510 [Appendix 11]. The presenceof slow eye movements in sleep, which were preferentially unco-ordinated, was confirmed in young infants69 and in adults.70
Plotke71 and Slotopolsky59 emphasized that eye movements insleep are far more sluggish in adults than in infants. Later, Fuchsand Wu56 mentioned “rotating movements of the eyes duringsleep“ in Chinese persons with physiologic logophthalmus,
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
preventing full closure of the eyes in sleep. However, to our bestknowledge there are no definite reports on rapid eye movementsduring sleep before 1953.1
Concerning an association between eye movements anddreaming, there is only few mentioning in the early literature. SirFrancis Galton denied any relationship between dreaming and eyemovements. Whenwriting on Visualised Numerals he stated that “Itis with them as it is with all of us in dreams, the imagery is beforeand around, but our eyes during sleep are turned inwards andupwards”.72, p. 87 A few years later, however, a relationship betweendreaming and eye movements was suggested by George TrumbullLadd, professor of moral philosophy at Yale university, who per-formed systematic self-observations on the relationship betweenretinal schemata and visual dream-images at sleep onset.73 Heassumed that the upward and inward position of the eyeballsbehind the closed lids “is probably most favourable to the disap-pearance from consciousness of all disturbing visual images”. Heproceeds by assuming that “Perhaps in deep and dreamless sleep.this position of the eyeballs is maintained unchanged. But I aminclined also to believe that, in somewhat vivid visual dreams,the eyeballs move gently in their sockets, taking variouspositions induced by the retinal phantasms as they control thedreams”.73, p. 304 Many years later Jacobson,74 who was interestedin the relationship between eye movements and mental activity,suggested an association between eye movements and dreamingwhen he wrote: “When a person dreams, he is tense in somelocality. Most often his eyes are active. Watch the sleeper whoseeyes move under his closed lids, but be quiet as you do so. Awakenhim in some unobtrusive way, . Dreams are quickly forgotten asa rule, but if you and the sleeper go at the matter seriously, you arelikely to find after awakening him that he has seen something ina dream. You are less likely to get a report of some visual picture ifyou watch him closely and arouse him at a timewhen his eyes havebeen completely relaxed or approximately so”.74, p. 144 However,in both cases most probably the authors referred to slow eyemovements at sleep onset and not to rapid eye movements, whichoccur later in sleep, usually for the first timewith a latency of about1 h after the onset of sleep. Dreamlike mental activity in associationwith eye movements early in sleep, as found by Ladd73 andJacobson,74 was much later confirmed by systematic studies.75
There are a few reports of eye movements in sleeping infantsand children. As already mentioned, some observers found morevivid movements of the globes in infants than in adults. Thepediatrician Wilhelm Preyer, who made careful records on thephysiological and mental development of his own son, confirmedthe appearance of eye movements in sleep: “While during sleep fora very long time, still to day 60, he often moved the eyes asym-metrically, and the lids on both sides.While the eyes were half openand snoring was ongoing. At the age of three month uncoordinatedeye movements were no longer observed. However, after that timeI have looked at the sleeping child only occasionally”60, p. 23
[Appendix 12]. Most interesting is his observation of a differenttype of lateral impulsive eyemovements, which occurred at the endof sleep, before awakening: “The eye movements before openingthe eye with awakening, especially lateral movements, are impul-sive. I saw these movements, which cannot be induced by light,even in adulthood”69, p. 133 [Appendix 13].
Of special interest is a study by Denisova and Figurin onPeriodic phenomena during sleep in infants, which was presented in1925 at the Third All-Russian Congress of Pediatrics.35 The authorsperformed in young infants continuous recordings of the respi-ration rate, in combination with systematic observations of bodymovements as well as lid and eye movements. The data clearlyshowed recurrent bouts of lid, eye, and body movements,concomitant with increases in respiratory rate during sleep
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

Fig. 7. Recording of respiration and documentation of movements of the body, the lids and the eyes in three infants, age 2 months (upper insert), 9 months 19 days (middle insert),and 1 year 19 days (lower insert). (Adapted from 35).
c Interestingly, the definition of stage B included “rolling of the eyes“. Thus, it issurprising, that Loomis et al. did not say anything on eye movements during Bphases later in sleep, since eye movements were presumably picked up by thefrontal electrode. This can be inferred from the statement that “Each time the eye iswinked a single continuous potential excursion lasting 1/10 s occurs from the frontelectrode only. The change shows no high frequency muscle potentials and prob-ably represents a rotation of the eyeball whose retina is at a different potential fromthe cornea”83, p. 259
H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e148
(Fig. 7). The mean recurrence time of the events was 50 min. Thisvalue is compatible with the mean duration of the NREM-REMcycle in young infants.76
Slow eye movements in “light sleep” were also described by deToni77 in 214 children, 3 days to 12 years old. The eye movements,which he observed, were pendular, mostly horizontal, witha frequency of 4e6 per minute. These eye movements, which werenot linked to changes of respiratory or cardiac rhythms, were rare in‘deep sleep’. In ‘deep sleep’ occasionally short periods with“irregular” eye movements occurred, with pauses between them.However, this observation was not followed up by the author, whohad concentrated on the study of eye movements during the firsthalf of night sleep. In summary, while many authors confirmed theoccurrence of slow eye movement in sleep, a few also mentionedexplicitly rapid eye movement,68,69,77 and some suggested a rela-tionship between eye movements and dreaming.68,73,74 Neverthe-less, it remains unclear whether the irregular eye movement ofdeep sleep actually correspond to rapid eye movements of REMsleep or not.
Early EEG sleep research: from 1935 to 1953
Only a few years after the development of electroencephalog-raphy by Hans Berger,78 professor of psychiatry in Jena, the banker-scientist Alfred L. Loomis together with E. Newton Harvey,a Princeton professor, and Garret A. Hobart were the first to applythe new technique to study sleep by long-term recordings inhumans. The results were published as an impressive series ofpapers between 1935 and 1939. While first recordings were per-formed with a massive 8 foot long drum polygraph, later the groupdeveloped a recording system equipped with 6 EEG amplifiers,allowing recordings from six different brain regions, each paired
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
with a frequency filter channel, and one channel for a time signal.79
The Loomis group subdivided sleep into five different states,running from the drowsy state Awith alphawaves to the deep sleepstate E with random (slow) waves. The intermediate states B, C, andD were characterized by low voltage (B), spindles (C), and bya combination of spindles and random waves (D).80 The sleepstages were found to be “fundamentally alike in the alpha and thenon-alpha types of individual”.79, p. 424 The time course of sleep wasdescribed by these researchers as “a continual shift of statesupward and downward sometimes associated with recognizedstimuli, sometimes without any external stimulus, but probably asa result of internal stimuli”80, p. 143 emphasizing the role of internalstimuli as cause of state shifts. The graphical representation of thetime course of sleep states shows clearly ultradian cyclicity witha cycle duration of about 1.5 h (Fig. 8)
Concerning dreams, the authors concluded from their studies:“We are now inclined to believe that dreams are not likely to beassociated with any unusual pattern of electrical potential but witha state of sleep”.80, p. 142 It was the B state which they found asso-ciated with dreaming, based on occasional dream reports aftereither spontaneous or deliberate awakenings.c The B state in theirnomenclature, which was defined as “A quite straight record, withno alpha rhythm and only low voltage changes of potential.” similar
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

Fig. 8. Sleep profiles of four individuals. The upper two were classified as non-alpha type, the lower two as alpha type. Sleep states (AeE) on the y-axis, hours of the night on thex-axis. The crosses represent major movements. In the first tracing N indicates noise and D a dream report. (From 80. With permission).
d After the medical exam Rudolf Klaue (1912-1970) hold the position of a physi-cian at the clinical department of the KWI, where he passed habilitation, workingon Parkinson’s disease. Later he was at the neurological university clinics ofFrankfurt, where he became lecturer (“Privatdozent”), and Würzburg. His finalposition was director of the psychiatric clinic Karl-Bonhoeffer-Heilstätten in Berlin-Wittenau (1956-1970). All his later publications were on neurological diseases,none on EEG.
H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e14 9
to the electroencephalographic characteristics of stage 1 and REMsleep, as defined later.81 Davis et al.82 concluded from their studiesthat dreams do not occur exclusively in the B state but also in the Cstate, which is characterized by spindles.
At the same time the relationship between sleep stages anddreamswas studied also by other groups. Helen Blake et al.84,85 whohad developed a quantified method to define sleep stages, and alsoused a different terminology, observed “in all subjects a uniformgeneral correlation between potentials and dreams”.85, p. 52
However, dream reporting was not restricted to the null state: “Wehave shown that a subject abruptly awakened, almost at any timeduring the night, can recall having dreamed; the longer the imme-diately preceding period with no alpha waves, the less is the recall,andwhen this period is about aminute (especially if deltawaves arepresent), there is no trace of a dream’s having been in progress. Thislargely excludes the possibility that, in the other states, the dreamran its course as a flashwhile the subject was actually in the processof waking; presumably dream consciousness, like wakingconsciousness, blurs and fades progressively as the activity of thecortical neurones falls to lower and lower levels”.85, p. 58
Blake et al.85 havedepicted their conceptof the time course of thedifferent EEG features schematically (Fig. 9). The figure emphasizesthe slow, decreasing phase of sleep, followed by a final increasingphase, basically a monophasic time course of sleep, superimposedby minor fluctuations of higher frequency. Delta waves, which areprominent in thefirst half of sleep, are followedbya singlenull phasebefore the EEG pattern transits into pre-awakening intermittentalpha, and finally into full awakening with alpha waves.
Summarizing the results from these early EEG studies, thereappeared a lawful relationship between dream reporting and sleepstate, with highest probability if the EEG is flat, and lowest prob-ability if the EEG is of high amplitude. However, already at this timeit was under discussionwhether dreaming is restricted to a specificsleep state or rather is associated with a broader array of stages,a discussion which is still ongoing in present time.86,87
At the same time, when Loomis and coworkers analyzed sleep inhumans with electroencephalographic techniques, Alois E. Korn-müller studied the relationship between the cytoarchitecture of the
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
brain and electrical potentials in animals at the Kaiser WilhelmInstitute (KWI) for Brain Research in Berlin. Rudolf Klaued, amedicalstudent who prepared his dissertation in Kornmüller’s laboratory,used the EEG technique to measure cortical activity during normalsleep and during drug-induced narcosis in the cat. He found out thatnormal sleep of the animals was characterized by two main sleepphases, which he called state I and II.88 State I, which occurred first,was therefore observed in all sleep recordings. The EEG potentialswere more irregular than during waking and the amplitudeincreased, to become twice as high as during waking. State IIdeveloped out of state I, but not always. In this state the amplitude ofthe EEG potentials was reduced to only half of the normal wakingEEG. State II can transit into state I at any time (Fig. 10).
State II was recognized by Klaue as deep sleep88, p. 515
[Appendix 14]. This assumption was also supported by the observa-tion, that after an awakeningout of state II therewas a latency of up to30 s until the normal waking EEG was installed, while after awak-enings out of state I the normal waking EEG returned immediately.
A third characteristic difference between the two states was themuscle tone. While muscle activity was present but arrested(“Gliederkatalepsie”) in state I, the muscles were found to be atonicin state II. Thus, the essential characteristics of REM sleep state inthe cat, which were later studied in detail5,6,9 were already recog-nized in this early study.
An additional feature of REM sleep in the cat, namely a state-dependent increase of phasic muscle activity was first recognizedby Derbyshire et al.89 who wrote: “In two of the three cats whichwewere able to observe without anesthesia, we found during sleepoccasional groups of large waves ., larger than those recorded inthe waking state. At other times when sleep was apparently less
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

Fig. 9. Schematic representation of EEG potentials during night sleep. Alpha waves (black heavy line) in per cent presence, spindles (dashes) in per cent presence and delta waves(dots) in extent of predominance. Oral temperature (black thin line). Blake et al. differed between six stages of sleep, which correspond to the depth of sleep. The stages were basedon quantitative analysis of alpha, delta and 14 Hz spindle activity in the EEG and were defined as 1. Alpha, 2. Alpha þ Delta, 3. Delta, 4. Null (or low voltage), 5. Intermittent alpha,and 6. Alpha (low intensity). Sleep stages along the x-axis. The arrow indicates sleep onset. The schema suggests a monophasic time course of body temperature and sleep,superimposed by shorter fluctuations in the relative presence of alpha and delta potentials during sleep. (From 85 Used with permission.)
H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e1410
tranquil, judging by twitching of the vibrissae, there were onlysmall rapid waves, as in the alert waking state”.89, p. 582
EEG patterns, characteristic for the two different states of sleep,were also recognized in early clinical sleep recordings. In 1939 Rudolf
Fig. 10. Upper insert: Schematic representation of a cat brain. Position of recording electrodestate I; d, transition from state I into state II, marked by an arrow. (From 88, Fig. 4).
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
Janzen, a visiting neurologist at the KWI for Brain Research in Berlinpublished a paper onphysiological sleep and sleep attacks inpatientswith narcolepsy.90 EEG recordings were performed repeatedly inthese patients, and in one patient the EEGwas recorded continuously
s marked by crosses. Lower insert: EEG potentials. Tracing a, waking; b, falling asleep; c,
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e14 11
for 24 h. Fig.11 shows seven consecutive 10 s EEG tracings from sleeponset. While the first 6 tracings show a waxing and waning EEGpattern, which is typical for the sleep onset phase, “stripe 6 was fol-lowed by a longer time of an uninterrupted state of reduced voltageproduction, and an absence of the spontaneous variations of the stateawake. Stripe 7 represents a segment of this phase. The patientwas indeep sleep and could not be awakened by faint external stimuli. Thus,while stripes 1-6 represent the bioelectrical equivalent of the sleeponset phase, stripe 7 shows deep sleep”90, p. 99 [Appendix 15]. WhatJanzen described here (cf. Fig.11, traces in segment 7) is an equivalentof a sleep onset REM sleep phase, a phenomenonwhich is frequentlyobserved in narcoleptic patients.91
The political development in Germany in the late thirties, fol-lowed by World War II, led to an abrupt halt of most of thesepromising research activities.92,93
Fig. 11. EEG recordings in a female narcoleptic patient (age: 32 years). Each of the 7 continuosites. The first 6 tracings show the typical waxing andwaning EEG patterns of sleep onset. BegiAt this time the patient was in deep sleep and could not be awakened by faint external stim
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
In 1950 L. R. Tskipuridze, a Georgian neurophysiologist, pub-lished a paper on the sleep EEG of intact cats.94 He observed thathigh-voltage slow waves, which were interpreted as inhibitory innature, dominated during deep sleep. However, beside deep sleephe observed a state of restless sleep with a different pattern ofelectrical activity: “During restless sleep the pattern of electricalactivity varies from region to region, but occasionally in both areasthe slow electrical potentials disappear and instead them appearthe fast ones”.94, p. 222; engl. transl. 2001, p. 11
The alternation of two kinds of sleep, one with a high-voltageEEG and spindles, the other with a low voltage EEG was confirmedin young infants by Kirchhoff and Fröhlich.95 They studied 41 younginfants (age range: 2e15 months) for 3e6 h each with the aim torecord the EEG during one or more sleep phases. While the basicsequence of potential changes in the young infants was found to be
us inserts shows bipolar EEG tracings from frontal, precentral and postcentral recordingnningwith insert 7 a longer uninterrupted state of reduced voltage production followed.uli. (From 90, Fig 2, With kind permission from Springer Science & Business Media).
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

Research agenda
1. After a phase of rapid growth, sleep research and medi-
cine have reached amature state. Studying the history of
the field shows how strongly the understanding of sleep
phenomena at a given time is depending on actual
theoretical conceptions, and technical and methodolog-
ical prerequisites.
2. The relationship between physiological and psycholog-
icalprocesses insleep,especiallydreaming, isapersistent
yetunresolvedissue,whichneedsnewresearchprotocols
for clarification.
Practice points
1. Experimental sleep research began in the second half of
the 19th century in parallel with the development of
physiology and psychology as experimental sciences.
2. Repeatedly measured depth of sleep and continuously
measured single physiological and behavioral variables
showed that sleep is not homogenous but undergoes
systematic variations in the range of 1e2 h (ultradian
variations).
3. The complex structure of sleep became evident with the
use of the electroencephalogram as the primary mea-
surement instrument of sleep research in the thirties of
the last century. The potential shifts and patterns of the
EEG allowed for the first time the definition of sleep
stages.
4. Two different kinds of sleep were described already
before the discovery of REM sleep. However, the coa-
lescence of changes in different physiological systems
was clearly recognized only when recurrent phaseswith
rapid eye movements were first discovered in 1953.
H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e1412
similar to what had been described by Loomis et al.80 and Blake andGerard84 in adults, main differences were found in the duration ofthe different EEG stages. The disintegration of the waking EEG atsleep onset was more rapid in infants, the stage with delta waveswas shorter, and the null phase with a nearly isoelectric EEGappeared earlier andwas quite short. The sleep stages with spindlesand delta waves occurred repeatedly, suggesting “several periodicfluctuations of the depth of sleep”95, p. 353 [Appendix 16]. Theseresults were confirmed and greatly extended later, after therecognition of REM sleep.96,97
The EEG proved from the beginning to be the most efficientmethod to represent the fluctuations and distribution of corticalactivity during sleep. The method was also attractive since brainpotentialsof sleepcorrelatedwith the reactivityof thebrain to sensorystimuli, and thus served as an estimate for the depth of sleep.84
Conclusion
The objective of the present review was to put the beginning of“modern” sleep research in perspective with those earlier observa-tions andfindingswhich suggested that sleep is not homogenousbutrather composed of quiet, deep sleep and restless sleep, frequentlyassociated with dreaming. The idea that movements are associatedwith dreaming is so pervasive that it was present in the ancientphilosophical approaches and became later a topic of experimentalstudies on sleep. Early observations on animals and babies wereamong the most important contributions to show the complexity ofsleep. Neurologists offered detailed descriptions of particular eventsoccurring during sleep and proposed a sleep semiology.98
The development of sleep research as an experimental science canbe subdivided into twomajor periods. The first one, lasted from 1863to1953, the secondone from1953untilpresent times.Thefirstperiod,with a duration of 90 years, started with Kohlschütter’s systematicmeasurements of the depth of sleep by the application of gradedacoustic stimuli to wake up the sleeping subject.16 The method wasbased on principles of psychophysics, assuming a lawful relationshipbetween intensity of sensation and stimulus strength. The results ofthe first experiment showed systematic variations in the depth ofsleep and the temporal distribution thereof. On the background of therapid development of physiology as an experimental science in thesecond half of the 19th century, paralleled by the invention ofmeasurement instruments and recording devices, physiologicalfunctions were measured not only during wakefulness but increas-ingly also during sleep. Many of the physiological and behavioralvariables displayed systematic variations in the course of sleep, sug-gesting that sleep is not homogenous. However, until themid-thirtiesof the last century, the general procedure was to measure in anexperiment a single variable,which represents the outputof a specificphysiological system. A major change in the measurement approachevolved in the decade between 1930 and 1940when the newmethodof electroencephalography enabled researchers to measure brainactivity online. Now, for the first time, polygraphic recordings ofmultiple EEG channels became feasible99 and themethodwasappliedsoon also in sleeping subjects.100 This essential expansion in scopeand technical development79 initiated a transition from a purelydescriptive to a ‘synthetic’ approach in sleep research, resulting in theconcept of different sleep stages or states, which allowed for the firsttime to depict the time course of sleep in detail.80,84 This promisingdevelopment in sleep research came soon thereafter to an abrupt haltat the eve of World War II, when some of the pioneering researchgroups were dissolved. As a consequence the further development ofthepsychophysiologyof sleep sloweddownandonly scattered resultson this topic were published between 1940 and 1953.
A breakthrough in the understanding of the structure of sleepwas initiated by the discovery of episodes with rapid eye
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
movements in sleep and their close association with dreaming.1
Far-reaching research activities gathered momentum by the newand fully unexpected finding. Next steps were the demonstrationthat rapid eye movements in sleep are linked exclusively toa certain EEG-defined stage of sleep4 and the recognition of muscleatonia as another hallmark of the new type of sleep.5 An essentialpresupposition for the recognition of two different states of sleepwas the transition from a descriptive attitude when measuringphysiological functions in sleep to a ‘synthetic’ view which focusedon the coalescence of several variables. This new approach enabledthe concept of ‘state’,101 followed by the study of specific neuronalmodalities which regulate the two alternating states of sleep, REMand NREM sleep.22 The progress in sleep research became theindispensable prerequisite for the succeeding development of sleepmedicine as a new medical discipline.
Conflicts of interest
The authors confirm that they have no conflicts of interest.
Acknowledgment
We thank Claude Gottesmann for continuous support during thepreparation of the manuscript.
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e14 13
Appendix. Supplementary material
Supplementary data related to this article can be found online atdoi:10.1016/j.smrv.2011.05.006.
References
1. Aserinsky E, Kleitman N. Regularly occurring periods of eye motility, andconcomitant phenomena, during sleep. Science 1953;118:273e4.
2. Aserinsky E, Kleitman N. Two types of ocular motility occurring in sleep.J Appl Physiol 1955;8:1e10.
3. Aserinsky E, Kleitman N. A motility cycle in sleeping infants as manifestedby ocular and gross bodily activity. J Appl Physiol 1955;8:11e8.
4. Dement WC, Kleitman N. Cyclic variations in EEG during sleep and theirrelation to eye movements, body motility, and dreaming. ElectroencephalClin Neurophysiol 1957;9:673e90.
5. Jouvet M, Michel F, Courjon J. Sur la mise en jeu de deux méchanismes àexpression électro-encéphalographique différente au cours du sommeilphysiologique chez le chat. C R Acad Sci 1959;248:3043e5.
6. Dement WC. The occurrence of low voltage fast electroencephalogrampatterns during behavioral sleep in the cat. Electroencephal Clin Neuro-physiol 1958;10:291e6.
7. Berger RJ. Tonus of extrinsic laryngeal muscles during sleep and dreaming.Science 1961;134:840.
8. Parmeggiani PL. A retrospective assessment of sleep research in theWesternworld during the late 19th and early 20th centuries. In: Liu SY, Inoué S,editors. Sleep: ancient and modern. SSTLPH: Shanghai; 1995. p. 89e99.
*9. Gottesmann C. The golden age of rapid eye movement sleep discoveries. I.Lucretius e 1964. Prog Neurobiol 2001;65:211e87.
10. Dement WC. History of sleep physiology and medicine. In: Kryger MH,Roth T, Dement WC, editors. Principles and practice of sleep medicine. 4th ed.Philadelphia: Elsevier, Saunders; 2005. p. 1e12.
11. Lucrece TC. De rerum natura; 1900. Trans. by C. Baily. Oxonii, E. Top-ographeo Clarendoniano.
12. Fernel J. Universa medicina; 1554. French translation 1655.13. Leroux V, Pigne C. Introduction. Camenae, no. 5; 2008.14. Lélut L- F. Mémoire sur le sommeil, les songes et le somnambulisme. Orléans:
Coignet Darnault; 1852.15. Liebeault A-A. Le sommeil provoqué et les états analogues. Paris: Octave Doin;
1889.16. Kohlschütter E. Messungen der Festigkeit des Schlafes. Z Ration Med
1863;17:209e53.17. Kohlschütter E. Mechanik des Schlafes. Z Ration Med 1869;34:42e8.18. Madsen PL, Holm S, Vorstrup S, Friberg L, Lassen NA, Wildschiøtz G. Human
regional cerebral blood flow during rapid-eye-movement sleep. J CerebBlood Flow Metab 1991;11:502e7.
19. Zoccoli G, Walker AM, Lenzi P, Franzini C. The cerebral circulation duringsleep: regulation mechanisms and functional implications. Sleep Med Rev2002;6:443e55.
20. Maquet P, Péters J-M, Aerts J, Delfiore G, Degueldre C, Luxen A, et al.Functional neuroanatomy of human rapid-eye-movement sleep anddreaming. Nature 1996;383:163e6.
*21. MacWilliam JA. Some applications of physiology to medicine: iii.dbloodpressure and heart action in sleep and dreams: their relation to haemor-rhages, angina, and sudden death. Br Med J 1923;2:1196e200.
22. Parmeggiani PL. Systemic Homeostasis and Poikilostasis in Sleep. Is REM Sleepa Physiological Paradox?. London: Imperial College Press; 2011.
23. Fechner G Th. Elemente der Psychophysik, 2 vol. Leipzig: Breitkopf undHärtel; 1860.
24. Basner M. Arousal threshold determination in 1862: Kohlschütter’smeasurements on the firmness of sleep. Sleep Med 2010;11:417e22.
25. Mönninghoff O, Piesbergen F. Messungen über die Tiefe des Schlafes. Z Biol1883;19:114e28.
26. Michelson E. Über die Tiefe des Schlafes. Dorpat; 1891.*27. Michelson E. Untersuchungen über die Tiefe des Schlafes. Kraepelins psychol
Arbeiten 1899;2:84e117.28. De Sanctis S, Neyroz U. Experimental investigation on the depth of sleep.
Psychol Rev 1902;9:254e82.29. Hacker F. Systematische Traumbeobachtungen mit besonderer
Berücksichtigung der Gedanken. Arch Psychol 1911;21:1e131.30. Köhler P. Ein Beitrag zur Traumpsychologie. Arch Ges Psychol
1912;23:415e83.31. Howell WH. A contribution to the physiology of sleep based upon
plethysmographic experiments. J Exp Med 1897;2:313e45.32. Johnson HM. Sleep. In: Valentine WL, editor. Readings in Experimental
Psychology. New York: Harper; 1931. p. 241e91.33. Zickgraf H. Untersuchungen über die Zusammenhänge zwischen den
Schlafbewegungen und der Tiefe des normalen Schlafes. Z Klin Med1953;152:96e117.
* The most important references are denoted by an asterisk.
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
34. Regelsberger H. Über vegetative Korrelationen im Schlafe des Menschen.Z Ges Neurol Psychiat 1942;174:727e39.
*35. (Russ.) Denisova MP, Figurin NL. Periodic phenomena in the sleep of chil-dren. Nov Refl Fiziol Nerv Syst 1926;2:338e45.
36. Wada T. Experimental study of hunger in its relation to activity. Arch Psy-chol 1922;57:1e65.
*37. Ohlmeyer P, Brilmayer H, Hüllstrung H. Periodische Vorgänge im Schlaf.Pflügers Arch ges Physiol 1944;248:559e60.
38. Ohlmeyer P, Brilmayer H. Periodische Vorgänge im Schlaf. II. Mitteilung.Pflügers Arch Ges Physiol 1947;249:50e5.
39. Kleitman N. Sleep and Wakefulness. 2nd ed. Chicago: Univ. of Chicago Press;1963.
40. Kleitman N. Basic rest-activity cycle e 22 years later. Sleep 1982;5:311e7.
41. Halverson HM. Genital and sphincter behavior of the male infant. J GenetPsychol 1940;56:95e136.
42. Fisher C, Gross J, Zuch J. Cycle of penile erections synchronous withdreaming (REM) sleep. Arch Gen Psychiat 1965;12:29e45.
43. Rothfeld J. Über Orgasmolepsie und über sexuelle Erregungen bei narko-leptischen Schlafzuständen nebst Beziehungen zur Narkolepsiefrage. Z gesNeurol Psychiat 1932;138:705e19.
44. Scharf MB, Hemsath R, Lysaght RJ, DeMarchis M, Zipkin J. Penile tumes-cence in temporally normal and pathological rapid eye movement sleep.J Urol 1983;130:909e11.
45. Bradley FH. On the failure of movement in dream. Mind 1894;3:373e7.46. Burdach KF. Der Mensch nach den verschiedenen Seiten seiner Natur. Stutt-
gart: P. Balz’sche Buchhandlung; 1837.47. Binz C. Über den Traum. Bonn: Adolph Marcus; 1878.48. Stanley Hall G. Laura Bridgman. Mind 1879;4:149e72.49. Anonym. The blind-deaf-mute Helen Keller. Mind 1889;14:305e9.*50. Max LW. An experimental study of the motor theory of consciousness. III.
Action-current responses in deaf-mutes during sleep, sensory stimulationand dreams. J Comp Psychol 1935;19:469e86.
51. Adrian ED. The physiology of sleep. Irish J Med Sci 1937;12:237e48.52. Jouvet M. Recherches sur les structures nerveuses et les mécanismes
responsables des différentes phases du sommeil physiologique. Arch ItalBiol 1962;100:125e206.
53. Giaquinto S, Pompeiano O, Somogyi I. Reflex activity of extensor and flexormuscles following muscular afferent excitation during sleep and wake-fulness. Experientia 1963;19:481e2.
*54. Zipperling W. Über eine besondere Form motorischer Reizzustände beiNeugeborenen (sog. “Stäupchen”). Z Kinderheilk 1912;5:31e40.
55. Klotz. Bemerkungen zur Arbeit Zipperlings über die sog. “Stäupchen”.Zeitschr Kinderheilk 1913;9:85e6.
56. Fuchs A, Wu FC. Sleep with half-open eyes. (physiologic lagophthalmus).Am J Ophthal 1948;31:717e20.
57. Stokes G. Observations on the closure of the eye in sleep. Dublin J Med Sci1840;18:70e5.
58. Loewenfeld IE. The Pupil. Anatomy, Physiology, and Clinical Applications, vols2. Boston, Oxford: Butterworth, Heinemann; 1999.
59. Slotopolsky B. Untersuchungen über das Bellsche Phänomen und ver-wandte Probleme. Z ges Neurol Psychiat 1930;125:252e326.
60. Berger E, Loewy R. L’état des yeux pendant le sommeil et la théorie dusommeil. J Anat Physiol (Paris) 1898;34:364e418.
61. Duval MM, Sommeil. Article in: nouveau dictionnaire de médecine de chir-urgie pratiques, illustré de figures intercalées dans le texte, sous la direction duDr Jaccoud. Paris: JB Baillière; 1882. Tome 33 p.262e88.
62. Raehlmann E, Witkowski L. Ueber atypische Augenbewegungen. ArchPhysiol; 1877:454e71.
63. Raehlmann E, Witkowski L. Ueber das Verhalten der Pupillen während desSchlafes nebst Bemerkungen zur Innervation der Iris. Arch Anat Physiol1878;108-11:119e21.
64. Siemens F. Zur Lehre vom epileptischen Schlaf und vom Schlaf überhaupt.Arch Psychiat Nervenkrankh 1879;9:72e82.
65. Sander W. Über die Beziehungen der Augen zum wachen und schlafendenZustande des Gehirns und über ihre Veränderungen bei Krankheiten. ArchPsychiat 1879;9:129e46.
66. Witkowski L. Ueber einige Bewegungserscheinungen an den Augen. ArchPsychiat Nervenkrankh 1881;11:507e16.
67. Piéron H. Le problème physiologique du sommeil. Paris: Masson & Cie; 1913.68. Polimanti O. Sur le phénomène de Charles Bell. Nouv Iconograph Salpêtrière
1907;20:508e18.69. Preyer W. Die Seele des Kindes. 1st ed., 1882. Leipzig: Grieben; 1900.
5th ed.70. Gernert R. Das Verhalten der Augen im Schlaf. Berlin: G Schade; 1898.71. Plotke L. Verhalten der Augen im Schlafe. Arch Psychiat 1880;10:205e20.72. Galton F. Visualised numerals. J Anthropol Instit 1881;10:85e102.73. Ladd GT. Contribution to the psychology of visual dreams. Mind
1892;1:299e304.74. Jacobson E. You can sleep well. the a b c’s of restful sleep for the average
person. New York: Whittlesey House; 1938.75. Foulkes D, Vogel G. Mental activity at sleep onset. J Abnorm Psychol
1965;70:231e43.76. Stern E, Parmelee AH, Akiyama Y, Schultz MA, Wenner WH. Sleep char-
acteristics in infants. Pediatrics 1969;43:65e70.
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/

H. Schulz, P. Salzarulo / Sleep Medicine Reviews xxx (2011) 1e1414
77. De Toni G. I movimenti pendolari dei bulbi oculari dei bambini durante ilsonno fisiologico, ed in alcuni stati morbosi. Pediatria 1933;41:489e98.
78. Berger H. Über das Elektrenkephalogramm des Menschen. Arch PsychiatNervenkr 1929;87:527e70.
79. Loomis AL, Harvey EN, Hobart GA. Distribution of disturbance patterns inthe human electroencephalogram, with special reference to sleep.J Neurophysiol 1938;1:413e30.
*80. Loomis AL, Harvey EN, Hobart GA. Cerebral states during sleep as studiedby human brain potentials. J Exp Psychol 1937;21:127e44.
81. Rechtschaffen A, Kales AA, editors. Manual of standardized terminology,techniques, and scoring system for sleep stages in human subjects. LosAngeles: University of California, Brain Information Service; 1968.
82. Davis H, Davis PA, Loomis AL, Harvey EN, Hobart GA. Human brainpotentials during the onset of sleep. J Neurophysiol 1938;1:24e38.
83. Loomis AL, Harvey EN, Hobart G. Electrical potentials of the human brain.J Exp Psychol 1936;19:249e79.
84. Blake H, Gerard RW. Brain potentials during sleep. Am J Physiol1937;119:692e703.
*85. Blake H, Gerard RW, Kleitman N. Factors influencing brain potentials duringsleep. J Neurophysiol 1939;2:48e60.
86. Salzarulo P, Cipolli C, Lairy GC, Pêcheux M. L’étude psychophysiologique del’activité mentale du sommeil. Evol Psychiatr 1973;38:33e70.
87. Solms M. Dreaming and REM sleep are controlled by different brainmechanisms. Behav Brain Sci 2000;23:843e50.
*88. Klaue R. Die bioelektrische Tätigkeit der Großhirnrinde im normalen Schlafund in der Narkose durch Schlafmittel. J Psychol Neurol 1937;47:510e31.
89. Derbyshire AJ, Rempel B, Forbes A, Lambert EF. The effects of anesthetics onaction potentials in the cerebral cortex of the cat. Am J Physiol1936;116:577e96.
Please cite this article in press as: Schulz H, Salzarulo P, Forerunnj.smrv.2011.05.006
90. Janzen R. Hirnbioelektrische Untersuchungen über den physiologischenSchlaf und den Schlafanfall bei Kranken mit genuiner Narkolepsie. Dtsch ZNervenheilk 1939;149:93e106.
91. Rechtschaffen A, Wolpert E, Dement WC, Mitchell SA, Fisher C. Nocturnalsleep of narcoleptics. EEG Clin Neurophysiol 1963;15:599e609.
92. Borck C. Hirnströme. Eine Kulturgeschichte der Elektroenzephalographie.Göttingen: Wallstein; 2005.
93. Conant J. Tuxedo Park. A Wall Street Tycoon and the Secret Palace of Sciencethat Changed the Course of World War II. New York: Simon & Schuster; 2002.
94. Tskipuridze LR. The electrical activity of the cerebral cortex in an intact catduring natural sleep. Georgian J Neurosci; 2001:1e12. 1-1[English trans-lation of the original paper, published in Russian in: JIEATEMBCTBOALAEEMJJ HAUL GPUIJHCLOK CCP 1950; 8:209e23].
95. Kirchhoff HW, Fröhlich B. Electroencephalographische Untersuchungenüber den Schlaf der Säuglinge. Arch Psychiat Nervenkr 1952;189:341e54.
96. Dreyfus-Brisac C, Fischgold H, Samson D, Sainte-Anne Dargassies S,Monod N, Blanc C. Veille, sommeil, réactivité sensorielle chez le prématuré,le nouveau-né et le nourrisson. Electroencephal Clin Neurophysiol1956;8(suppl. 6):417e40.
97. Roffwarg HP, Dement WC, Fischer C. Preliminary obsrvations of the sleep-dream patterns in neonates, infants, children and adults. In: Harms E, editor.Problems of sleep and dreams in children. Pergamon press; 1964. p. 60e72.
98. Lhermitte J. Le Sommeil. Paris: Armand Colin; 1931.99. Kreezer G. The electro-encephalogram and its use in psychology. Am J
Psychol 1938;51:737e59.100. Loomis AL, Harvey EN, Hobart GA. Electrical potentials of the human brain.
Science 1935;81:597e8. 82:198e200.101. Prechtl HFR. The behavioural states of the newborn infant (a review). Brain
Res 1974;76:185e212.
ers of REM sleep, Sleep Medicine Reviews (2011), doi:10.1016/
Copyright © 2022 FDOKUMEN




















