
Increases in cAMP, MAPK Activity, and CREB Phosphorylation during REM Sleep: Implications for REM...
9
Behavioral/Cognitive Increases in cAMP, MAPK Activity, and CREB Phosphorylation during REM Sleep: Implications for REM Sleep and Memory Consolidation Jie Luo, Trongha X. Phan, Yimei Yang, Michael G. Garelick, and Daniel R. Storm Department of Pharmacology, School of Medicine, University of Washington, Seattle, Washington 98195 The cyclic adenosine monophosphate (cAMP), mitogen-activated protein kinase (MAPK), and cAMP response element-binding protein (CREB) transcriptional pathway is required for consolidation of hippocampus-dependent memory. In mice, this pathway undergoes a circadian oscillation required for memory persistence that reaches a peak during the daytime. Because mice exhibit polyphasic sleep patterns during the day, this suggested the interesting possibility that cAMP, MAPK activity, and CREB phosphorylation may be elevated during sleep. Here, we report that cAMP, phospho-p44/42 MAPK, and phospho-CREB are higher in rapid eye movement (REM) sleep compared with awake mice but are not elevated in non-REM sleep. This peak of activity during REM sleep does not occur in mice lacking calmodulin-stimulated adenylyl cyclases, a mouse strain that learns but cannot consolidate hippocampus-dependent memory. We conclude that a preferential increase in cAMP, MAPK activity, and CREB phosphorylation during REM sleep may contribute to hippocampus-dependent memory consolidation. Introduction Sleep is an indispensable physiological state that naturally occurs in a wide variety of species (Campbell and Tobler, 1984; Cirelli and Tononi, 2008; Siegel, 2008). It is characterized by the revers- ible loss of consciousness and reduced motor activity. In terres- trial mammals and birds, sleep is further separated into rapid eye movement (REM) sleep and non-REM (NREM) sleep based on distinctive EEG/EMG measurements (Lesku et al., 2006; Allada and Siegel, 2008; Siegel, 2009). REM sleep is evidenced by rapid, low-voltage waves (4 – 8 Hz) in parallel with muscle atonia and rapid eye/whisker movements (Aserinsky and Kleitman, 1953). NREM sleep, however, is composed of stage 1, 2 and slow-wave sleep (SWS) featuring low-frequency, large-amplitude waves (0.5– 4 Hz) (Rechtschaffen and Kales, 1968). Sleep has been strongly implicated in the off-line reprocessing of recently acquired memory (Stickgold, 1998; Walker and Stick- gold, 2004; Stickgold, 2005; Marshall and Born, 2007; Diekel- mann and Born, 2010; Hernandez and Abel, 2011). However, the mechanistic relationship between sleep and memory consolida- tion is undefined. In rodents, neuronal firings recapitulating those evoked by previous awake behavior have been detected in both REM sleep and NREM sleep (Wilson and McNaughton, 1994; Skaggs and McNaughton, 1996; Shen et al., 1998; Poe et al., 2000; Louie and Wilson, 2001; Ji and Wilson, 2007). These find- ings suggest the intriguing hypothesis that molecular cascades activated during memory acquisition may be recruited during subsequent sleep to promote memory consolidation. Consolidation of hippocampus-dependent memory depends upon activation of the calmodulin-stimulated adenylyl cyclases, mitogen-activated protein kinase (MAPK), and cAMP response element-binding protein (CREB)-mediated transcriptional path- way (Bourtchuladze et al., 1994; Wu et al., 1995; Atkins et al., 1998; Blum et al., 1999; Wong et al., 1999; Athos et al., 2002; Pittenger et al., 2002; Sindreu et al., 2007). However, how long- term memory can endure for periods far exceeding the lifetimes of synaptic proteins produced during memory acquisition re- mains to be determined. Recent studies have revealed that the cAMP/MAPK/CREB pathway undergoes a circadian oscillation in area CA1 of the mouse hippocampus with peak activity during the daytime at ZT4 (zeitgeber time 4, 12:00 P.M.). Disruption of this signaling oscillation days after hippocampus-dependent memory is consolidated impairs the persistence of memory (Eckel-Mahan et al., 2008; Phan et al., 2011). This has led to the idea that hippocampus-dependent memories are maintained over extended periods of time by periodic reactivation of this memory consolidation pathway during the circadian cycle. Because the cAMP/MAPK/CREB transcriptional pathway is maximal during the daytime when mice exhibit periods of wake- fulness, REM sleep and NREM sleep, a critical question is whether or not this signaling pathway is activated during sleep. Received Oct. 25, 2012; revised March 1, 2013; accepted March 7, 2013. Author contributions: J.L., T.X.P., and D.R.S. designed research; J.L., Y.Y., and M.G.G. performed research; J.L. analyzed data; J.L. and D.R.S. wrote the paper. This work was supported by National Institutes of Health Grants NS 20498 and MH 073601. We thank members of the D.R.S. laboratory for critical reading of the manuscript and the W. M. Keck Center for Advanced Studies in Neural Signaling for imaging assistance. The authors declare no competing financial interests. Correspondence should be addressed to Dr. Daniel R. Storm, Department of Pharmacology, Mail Box 357750, Health Sciences Building, University of Washington, Seattle, WA 98195-7750. E-mail: [email protected]. T. X. Phan’s present address: Picower Institute for Learning and Memory, Massachusetts Institute of Technology, 77 Massachusetts Avenue, Cambridge, MA 02139. M. G. Garelick’s present address: Center for Neural Science, New York University, 4 Washington Place West, New York, NY 10003. DOI:10.1523/JNEUROSCI.5018-12.2013 Copyright © 2013 the authors 0270-6474/13/336460-09$15.00/0 6460 • The Journal of Neuroscience, April 10, 2013 • 33(15):6460 – 6468
Transcript of Increases in cAMP, MAPK Activity, and CREB Phosphorylation during REM Sleep: Implications for REM...

Behavioral/Cognitive
Increases in cAMP, MAPK Activity, and CREBPhosphorylation during REM Sleep: Implications for REMSleep and Memory Consolidation
Jie Luo, Trongha X. Phan, Yimei Yang, Michael G. Garelick, and Daniel R. StormDepartment of Pharmacology, School of Medicine, University of Washington, Seattle, Washington 98195
The cyclic adenosine monophosphate (cAMP), mitogen-activated protein kinase (MAPK), and cAMP response element-binding protein(CREB) transcriptional pathway is required for consolidation of hippocampus-dependent memory. In mice, this pathway undergoes acircadian oscillation required for memory persistence that reaches a peak during the daytime. Because mice exhibit polyphasic sleeppatterns during the day, this suggested the interesting possibility that cAMP, MAPK activity, and CREB phosphorylation may be elevatedduring sleep. Here, we report that cAMP, phospho-p44/42 MAPK, and phospho-CREB are higher in rapid eye movement (REM) sleepcompared with awake mice but are not elevated in non-REM sleep. This peak of activity during REM sleep does not occur in mice lackingcalmodulin-stimulated adenylyl cyclases, a mouse strain that learns but cannot consolidate hippocampus-dependent memory. Weconclude that a preferential increase in cAMP, MAPK activity, and CREB phosphorylation during REM sleep may contribute tohippocampus-dependent memory consolidation.
IntroductionSleep is an indispensable physiological state that naturally occursin a wide variety of species (Campbell and Tobler, 1984; Cirelliand Tononi, 2008; Siegel, 2008). It is characterized by the revers-ible loss of consciousness and reduced motor activity. In terres-trial mammals and birds, sleep is further separated into rapid eyemovement (REM) sleep and non-REM (NREM) sleep based ondistinctive EEG/EMG measurements (Lesku et al., 2006; Alladaand Siegel, 2008; Siegel, 2009). REM sleep is evidenced by rapid,low-voltage � waves (4 – 8 Hz) in parallel with muscle atonia andrapid eye/whisker movements (Aserinsky and Kleitman, 1953).NREM sleep, however, is composed of stage 1, 2 and slow-wavesleep (SWS) featuring low-frequency, large-amplitude � waves(0.5– 4 Hz) (Rechtschaffen and Kales, 1968).
Sleep has been strongly implicated in the off-line reprocessingof recently acquired memory (Stickgold, 1998; Walker and Stick-gold, 2004; Stickgold, 2005; Marshall and Born, 2007; Diekel-mann and Born, 2010; Hernandez and Abel, 2011). However, the
mechanistic relationship between sleep and memory consolida-tion is undefined. In rodents, neuronal firings recapitulatingthose evoked by previous awake behavior have been detected inboth REM sleep and NREM sleep (Wilson and McNaughton,1994; Skaggs and McNaughton, 1996; Shen et al., 1998; Poe et al.,2000; Louie and Wilson, 2001; Ji and Wilson, 2007). These find-ings suggest the intriguing hypothesis that molecular cascadesactivated during memory acquisition may be recruited duringsubsequent sleep to promote memory consolidation.
Consolidation of hippocampus-dependent memory dependsupon activation of the calmodulin-stimulated adenylyl cyclases,mitogen-activated protein kinase (MAPK), and cAMP responseelement-binding protein (CREB)-mediated transcriptional path-way (Bourtchuladze et al., 1994; Wu et al., 1995; Atkins et al.,1998; Blum et al., 1999; Wong et al., 1999; Athos et al., 2002;Pittenger et al., 2002; Sindreu et al., 2007). However, how long-term memory can endure for periods far exceeding the lifetimesof synaptic proteins produced during memory acquisition re-mains to be determined. Recent studies have revealed that thecAMP/MAPK/CREB pathway undergoes a circadian oscillationin area CA1 of the mouse hippocampus with peak activity duringthe daytime at ZT4 (zeitgeber time 4, 12:00 P.M.). Disruption ofthis signaling oscillation days after hippocampus-dependentmemory is consolidated impairs the persistence of memory(Eckel-Mahan et al., 2008; Phan et al., 2011). This has led to theidea that hippocampus-dependent memories are maintainedover extended periods of time by periodic reactivation of thismemory consolidation pathway during the circadian cycle.
Because the cAMP/MAPK/CREB transcriptional pathway ismaximal during the daytime when mice exhibit periods of wake-fulness, REM sleep and NREM sleep, a critical question iswhether or not this signaling pathway is activated during sleep.
Received Oct. 25, 2012; revised March 1, 2013; accepted March 7, 2013.Author contributions: J.L., T.X.P., and D.R.S. designed research; J.L., Y.Y., and M.G.G. performed research; J.L.
analyzed data; J.L. and D.R.S. wrote the paper.This work was supported by National Institutes of Health Grants NS 20498 and MH 073601. We thank members
of the D.R.S. laboratory for critical reading of the manuscript and the W. M. Keck Center for Advanced Studies inNeural Signaling for imaging assistance.
The authors declare no competing financial interests.Correspondence should be addressed to Dr. Daniel R. Storm, Department of Pharmacology, Mail Box 357750,
Health Sciences Building, University of Washington, Seattle, WA 98195-7750. E-mail: [email protected]. X. Phan’s present address: Picower Institute for Learning and Memory, Massachusetts Institute of Technology,
77 Massachusetts Avenue, Cambridge, MA 02139.M. G. Garelick’s present address: Center for Neural Science, New York University, 4 Washington Place West,
New York, NY 10003.DOI:10.1523/JNEUROSCI.5018-12.2013
Copyright © 2013 the authors 0270-6474/13/336460-09$15.00/0
6460 • The Journal of Neuroscience, April 10, 2013 • 33(15):6460 – 6468

Here, we report that cAMP as well as MAPK activity and CREBphosphorylation are significantly higher in REM sleep comparedwith awake mice but not higher in NREM sleep. These increasesdo not occur in memory-deficient mice lacking calmodulin-stimulated adenylyl cyclases. Our results support the hypothesisthat the activation of the cAMP/MAPK/CREB transcriptionalpathway during REM sleep contributes to memory consolidationand are consistent with electrophysiological studies showing re-play activity in the hippocampus during REM sleep (Poe et al.,2000; Louie and Wilson, 2001).
Materials and MethodsMice. Adult (3– 6 months of age) male C57BL/6J wild-type (WT) andtype 1 and type 8 adenylyl cyclases (AC1 and AC8) double-knock-out(DKO) mice were used in the experiments. DKO mice were generated aspreviously described and backcross bred into C57BL/6J background formore than nine generations (Wong et al., 1999). Mice were entrained ina 12 h light/12 h dark cycle with lights on at 8:00 A.M. (ZT0) at least 7 dbefore the start of the experiment. Animal procedures were performedunder protocols approved by the Institutional Animal Care and UseCommittee of the University of Washington and conform to NationalInstitutes of Health guidelines.
EEG implantations and recordings. Each mouse was implanted with anEEG headmount following the manufacturer’s instructions (PinnacleTechnology). Briefly, mice were anesthetized with a ketamine/xylazinemixture and aligned in a stereotaxic apparatus. After exposing the skullsurface, headmounts were centered along the sagittal suture with thefront edge 3.5 mm anterior to bregma. Headmounts were then securedwith four stainless steel screws (also functioned as EEG electrodes) andcoated in acrylic for insulation. When positioned properly, all fourscrews (two anterior: AP 3.0 mm, ML �1.75 mm; two posterior: AP �4.0mm, ML �1.75 mm, relative to bregma) should sit on the cerebral regionof an adult mouse (20 –30 g). Two EMG wires were inserted bilaterallyinto the trapezius muscles to monitor neck activity. Mice were allowed5 d to recover in their home cages before tethering to the commutatorsusing 100 � preamplifiers in circular recording cages. Mice were adaptedto the recording apparatus for additional 5 d and EEG/EMG signals werecollected using Sirenia software (Pinnacle Technology). Captured datawere scored off-line manually to distinguish waking, REM sleep, andNREM sleep based on standard criteria including: (1) the amplitude andfrequency of EEG; (2) the amplitude and frequency of EMG; and (3) theregularity of EEG/EMG. On the experimental day, mice were monitoredbased on both behavioral activities and EEG/EMG patterns. Mice werekilled after 10 min in waking or NREM sleep or after 1 min in REM sleepbetween 12:00 P.M. (ZT4) and 2:00 P.M. (ZT6) to avoid variations re-sulted from circadian changes.
Passive avoidance. The training chamber was a shuttle box that wasequally divided into lit and dark compartments by a metal partition witha trap door (Coulbourn Instruments). During training, mice were al-lowed to explore the lit side freely for 1 min before the trap door wasopened. Immediately after mice entered the dark side, the door wasclosed and a mild foot shock (0.7 mA, 2 s) was delivered. Mice were keptin the dark side for 20 s and then returned to their home cages. Duringtesting, mice were put back into the lit side, and the time mice spentbefore crossing over into the dark side (crossover latency) was recorded.The cutoff value for crossover latency was set to 5 min.
Contextual fear conditioning. The contextual training and testing wereperformed in a contextual arena with clear sidewalls and a metal shock floor(Coulbourn Instruments). During training, mice were allowed to explorethe chamber freely for 2 min before receiving a mild foot shock (0.7 mA, 2 s)from the metal grid. Mice were kept in the chamber for additional 1 min andreturned to their home cages. During testing, mice were placed back into thetraining chamber without shock for 2 min. Freezing behavior, defined bybody immobility except for respiration, was scored every 10 s over a 2 minperiod. Data were presented as a percentage of time in freezing over totalsampling period.
Immunohistochemistry. Mice were cervical dislocated and brains wereremoved quickly (within 2 min) and immersed in 4% paraformalde-
hyde/50 mM NaF/1 mM Na3VO4 in 1 � PBS, pH 7.4, overnight at 4°C,with one change of fixative after 3 h. Brains were cryoprotected in 30%sucrose, frozen in 2-methylbutane/dry ice bath, and cut into serial coro-nal sections (20 �m) in a cryostat (Leica). Sections were pretreated aspreviously described (Sindreu et al., 2007) and first stained with rabbitanti-phospho-p44/42 MAPK [also referred as extracellular signal-regulated kinase 1 and 2 (ERK1/2), 1:10,000, Cell Signaling Technology].The signal was amplified by deposition of cyanine-3 tyramide complexesusing Tyramide Signal Amplification System (PerkinElmer Life and An-alytical Sciences). Sections were blocked again and incubated with rabbitanti-phospho-CREB (1:100, Cell Signaling Technology). Sections werethen incubated with AlexaFluor-488-conjugated secondary antibody(Invitrogen), counterstained in DAPI (Invitrogen), and mounted usingAqua-Poly/Mount (Polysciences). Absence of cross-reactivity betweenthe first primary antibody and the second secondary antibody was con-firmed in experiments in which the second primary antibody was omit-ted (data not shown).
Imaging and analysis. Sections were imaged on either a Nikon EclipseE400 Microscope or a Zeiss 510 Meta laser scanning confocal micro-scope. Density filter, pinhole, detector gain, and offset were kept constantfor experimental comparisons. Hippocampal sections spanning between�1.70 mm and �2.18 mm relative to bregma were examined in thestudy. For phospho-ERK1/2 (pERK)-positive cells, four hippocampalsections per mouse were analyzed using 20� objective lens. Cell countsper subregion were averaged across sections and then across animals toyield group means. For phospho-CREB (pCREB) fluorescent intensityanalysis, individual images were taken under 10� objective lens andmontages were constructed and analyzed using Fiji software (NationalInstitutes of Health). Area, mean gray value, and integrated intensitywere measured and compared among three groups.
Western analysis. Mice were cervical dislocated, and hippocampi werequickly dissected out and homogenized with 50 strokes of Kontes “B”pestle in 200 �l of lysis buffer [10 mM HEPES, pH 7.5, 150 mM NaCl, 2.5mM EDTA, pH 8.0, 2.5 mM EGTA, pH 8.0, 50 mM NaF, 5 mM DTT, 1.5%NP-40, 10% glycerol, 1 mM PMSF, 1 mM Na3VO4, 1 � phosphataseinhibitor mixture 2 and 3 (Sigma), and one tablet/10 Complete MiniEDTA-free protease inhibitor (Roche)]. Homogenates were spun at14,000 � g for 10 min, and supernatant was retained as the whole-cellsoluble fraction. Samples were mixed with equal amounts of Laemmlisample buffer (Bio-Rad), heated at 95°C for 10 min, and loaded onto4 –15% Tris-HCl polyacrylamide gels (Bio-Rad) for SDS-PAGE anal-ysis. Proteins were then transferred onto PVDF membranes (Milli-pore) and blocked with 5% BSA in 1 � TBS with 0.05% Tween 20.Blots were incubated with rabbit anti-phospho-(Ser/Thr) protein ki-nase A (PKA) substrates (referred as pPKA-s, 1:1000, Cell SignalingTechnology) overnight at 4°C. Blots were then incubated with alka-line phosphatase-conjugated secondary antibody (Sigma) and devel-oped using CDP-Star substrate (Applied Biosystems). After strippingin 25 mM glycine/1% SDS, pH 2.0, for 40 min at room temperature,blots were blocked again and incubated with mouse anti-actin (1:5000, Millipore). Protein bands were visualized with ChemiDocXRS� system and analyzed using Image Lab software (Bio-Rad). Op-tical density of bands immunopositive for pPKA-s was normalized tothat of corresponding actin bands.
Measurement of cAMP. Hippocampal homogenates were prepared asdescribed above, except that 0.5 mM 3-isobutyl-1-methylxanthine wasalso included in the lysis buffer. Total cAMP activity was measured usingAmersham cAMP Biotrak Enzymeimmunoassay System according to themanufacturer’s instructions (GE Healthcare). The results were normal-ized by total protein amounts which were determined using Pierce BCAProtein Assay Kit (Thermo Scientific).
Statistical analysis. Results are expressed as mean � SEM. Behavioraldata from passive avoidance and contextual fear conditioning studieswere analyzed by two-way ANOVA. All other data were analyzed byone-way ANOVA. Bonferroni’s multiple-comparison tests were fol-lowed for post hoc analysis. Significance was set at p � 0.05.
Luo et al. • cAMP/MAPK/CREB Signaling Elevates during REM Sleep J. Neurosci., April 10, 2013 • 33(15):6460 – 6468 • 6461

ResultsDetermination of mouse sleeparchitecture using EEG/EMGrecordings
Prefabricated headmounts were im-planted on top of the mouse skull to cap-ture cortical EEG and neck EMG activitiesas described in Materials and Methods.Signals were then amplified, filtered, andtransmitted to the recording units fordocumentation and analysis. Waking,REM sleep and NREM sleep were identi-fied based on standard criteria as previ-ously described (Tobler et al., 1997;Crocker and Sehgal, 2010). Waking wascharacterized by high-frequency, low-amplitude EEG signals in combinationwith chaotic EMG activity (Fig. 1A). REMsleep, however, exhibited regular and fastEEG spikes in low amplitude paralleled bythe lack of EMG activity (Fig. 1B). NREMsleep was distinguished by low-frequency,high-amplitude EEG waves and attenu-ated muscle activity (Fig. 1C). Similar toadult humans (Carskadon and Dement,2011), a cycle of waking-NREM sleep-REM sleep was also detected in mice.
To examine the general sleep patternsof WT mice, we performed EEG/EMG re-cordings for five consecutive days. Thedata were then visually scored, and thepercentages of time spent in total sleep,REM sleep, and NREM sleep were aver-aged across days and animals to yieldgroup means. We observed that micespent 44.3 � 0.5% of the recording time in total sleep over a 24 hperiod: 3.8 � 0.1% in REM sleep and 40.5 � 0.4% in NREM sleep(Fig. 2A–C). In the presence of light (ZT0-ZT12), mice spent67.2 � 1.3% of the recording time in total sleep (Fig. 2A). Thetime spent in NREM sleep was �10 times greater than that inREM sleep (NREM, 60.9 � 1.1%; REM, 6.2 � 0.3%; Fig. 2B,C).In the absence of light (ZT12-ZT24), mice spent much less timein sleep (21.4 � 1.3%; Fig. 2A). However, the time in NREM sleepincreased to 15 times as many as that in REM sleep (NREM,20.1 � 1.3%; REM, 1.3 � 0.2%; Fig. 2B,C). When further ana-lyzed in 4 h intervals, we detected parallel oscillations of timespent in total sleep, REM sleep, and NREM sleep over a 24 hperiod (Fig. 2D–F). The preponderance of sleep during the lightphase corresponds to normal sleep patterns of mice under 12:12light/dark entrained conditions (Veasey et al., 2000).
Activation of MAPK and CREB during REM sleepWe previously discovered that MAPK activity and CREB phos-phorylation underwent circadian oscillations with peak activitiesbetween ZT4 and ZT8 (Eckel-Mahan et al., 2008). In the presentstudy, we hypothesized that the molecular components involvedin memory consolidation may be reactivated during specificphases of sleep within this period. Therefore, experiments wereperformed between ZT4 and ZT6 to monitor changes in MAPKactivity during waking and sleep. The behavioral states of micewere identified based on their physical activities and EEG/EMGsignals. Awake mice demonstrated full awareness to the environ-ment and high levels of EMG activity. Mice in sleep lay motion-
less with closed eyes and sometimes with head tucked under thebody. Because NREM sleep usually stays long (19.7 � 1.3 min)whereas REM sleep is transitory (3.1 � 0.1 min), mice were al-lowed 10 min in waking or NREM sleep but 1 min in REM sleepbefore cervical dislocation. MAPK activity was determined byimmunostaining hippocampal sections for dually phosphory-lated, activated MAPK (pERK) (Atkins et al., 1998; Sindreu et al.,2007), and cells immunopositive for pERK were counted in thepyramidal layer of the CA1 and the granular layer of the dentategyrus (DG). We detected pERK labeling in both cell bodies andapical dendrites (Fig. 3A). In the CA1, there were 3 times morepERK� cells in mice during REM sleep compared with awakemice (REM, 73 � 5; waking, 23 � 5; one-way ANOVA, F(2,16) �41.05, p � 0.0001; Fig. 3B). No significant difference was detectedbetween NREM sleep and waking (NREM, 16 � 5; waking, 23 �5; Fig. 3B). In the DG, a significant increase in pERK� cells,although to a lesser extent, was also detected in mice during REMsleep compared with awake mice (REM, 9 � 1; waking, 5 � 1;one-way ANOVA, F(2,16) � 8.944, p � 0.0025; Fig. 3C). More-over, the numbers of pERK� cells in mice during REM sleep werehigher than those during NREM sleep in the CA1 (p � 0.001) andDG (p � 0.01).
MAPK activation of CREB-mediated transcription is stronglyimplicated in memory consolidation and is activated duringtraining for hippocampus-dependent memory (Bourtchuladze etal., 1994; Wu et al., 1995; Atkins et al., 1998; Impey et al., 1998b;Wong et al., 1999; Athos et al., 2002; Pittenger et al., 2002;Sindreu et al., 2007). To examine changes in CREB activity be-
Figure 1. Representative EEG/EMG signals of waking, REM sleep, and NREM sleep in WT mice. A, EEG/EMG data from mice inwaking. B, EEG/EMG data from mice in REM sleep. C, EEG/EMG data from mice in NREM sleep. Note the fast, low-amplitude EEGtraces in parallel with a complete loss of EMG activity during REM sleep, contrasting to high-amplitude EEG slow waves duringNREM sleep and high levels of EMG activity during waking.
6462 • J. Neurosci., April 10, 2013 • 33(15):6460 – 6468 Luo et al. • cAMP/MAPK/CREB Signaling Elevates during REM Sleep
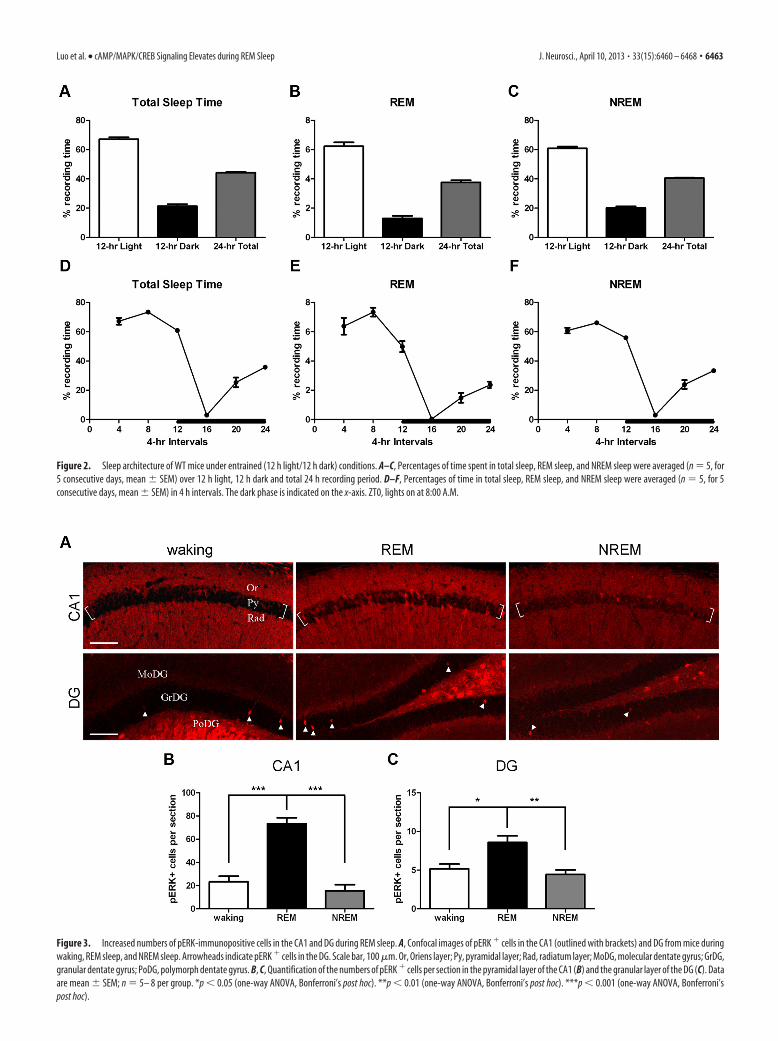
Figure 3. Increased numbers of pERK-immunopositive cells in the CA1 and DG during REM sleep. A, Confocal images of pERK � cells in the CA1 (outlined with brackets) and DG from mice duringwaking, REM sleep, and NREM sleep. Arrowheads indicate pERK � cells in the DG. Scale bar, 100 �m. Or, Oriens layer; Py, pyramidal layer; Rad, radiatum layer; MoDG, molecular dentate gyrus; GrDG,granular dentate gyrus; PoDG, polymorph dentate gyrus. B, C, Quantification of the numbers of pERK � cells per section in the pyramidal layer of the CA1 (B) and the granular layer of the DG (C). Dataare mean � SEM; n � 5– 8 per group. *p � 0.05 (one-way ANOVA, Bonferroni’s post hoc). **p � 0.01 (one-way ANOVA, Bonferroni’s post hoc). ***p � 0.001 (one-way ANOVA, Bonferroni’spost hoc).
Figure 2. Sleep architecture of WT mice under entrained (12 h light/12 h dark) conditions. A–C, Percentages of time spent in total sleep, REM sleep, and NREM sleep were averaged (n � 5, for5 consecutive days, mean � SEM) over 12 h light, 12 h dark and total 24 h recording period. D–F, Percentages of time in total sleep, REM sleep, and NREM sleep were averaged (n � 5, for 5consecutive days, mean � SEM) in 4 h intervals. The dark phase is indicated on the x-axis. ZT0, lights on at 8:00 A.M.
Luo et al. • cAMP/MAPK/CREB Signaling Elevates during REM Sleep J. Neurosci., April 10, 2013 • 33(15):6460 – 6468 • 6463

tween waking and sleep, we immunostained hippocampal sec-tions for CREB phosphorylated at Ser-133 (pCREB). Because weobserved a large number of cells immunopositive for pCREB inthe CA1 and DG (Fig. 4A), we compared total fluorescent inten-sity within the subregions among three behavioral states. Theintensity of pCREB immunofluorescence was significantly higherin REM sleep compared with awake mice in the CA1 (one-wayANOVA, F(2,16) � 5.117, p � 0.0191; Fig. 4B) and DG (one-wayANOVA, F(2,16) � 4.109, p � 0.0363; Fig. 4C), but not in NREMsleep relative to awake animals. Together, these data indicate thatMAPK activity and CREB phosphorylation increase during REMsleep in the CA1 pyramidal layer and DG granular layer of thehippocampus compared with awake mice.
Hippocampal cAMP/PKA signaling increases duringREM sleepcAMP is required for activation and nuclear translocation ofMAPK as well as subsequent CREB phosphorylation duringmemory formation (Impey et al., 1998b; Sindreu et al., 2007). Wetherefore measured cAMP levels and PKA activity between wak-ing and sleep. The average hippocampal cAMP of all mice exam-ined (including waking, REM, and NREM groups) between ZT4and ZT6 was 30.9 � 2.7 pmol/mg of protein. Within this timeframe, however, the cAMP level was greatest in REM sleep(41.0 � 4.1 pmol/mg of protein), significantly higher than that inawake animals or NREM sleep (one-way ANOVA, F(2,14) �6.155, p � 0.0121; Fig. 5A).
To generally monitor cAMP-dependent PKA activity duringsleep, we examined the levels of downstream substrates phos-phorylated by PKA using Western analysis. An antibody thatdetects proteins phosphorylated at a conserved PKA phosphory-lation motif was used, and the immunoreactivity was determined
by the optical intensity of the bands. Of an array of bands de-tected, we analyzed three (labeled 1–3) in sizes ranging between50 and 100 kDa for PKA activity during waking and sleep (Fig.5B). An increase in pPKA-s labeling during REM sleep comparedwith awake animals was detected in all three bands analyzed, twoof which were significantly higher (band 2, one-way ANOVA,F(2,14) � 11.62, p � 0.0011; band 3, one-way ANOVA, F(2,14) �15.68, p � 0.0003; Fig. 5C). It was also evident that pPKA-s in-tensity was greatly enhanced during REM sleep relative to NREMsleep (band 1, p � 0.01; band 2, p � 0.01; band 3, p � 0.001).However, PKA activity between NREM sleep and waking was notstatistically different. In addition, although the specific PKA sub-strates showing higher phosphorylation during REM sleep werenot characterized here, CREB phosphorylation was significantlyhigher during REM sleep (Fig. 4). Collectively, these data indicatethat hippocampal cAMP/PKA signaling is higher during REMsleep than in animals that are awake.
Calmodulin-stimulated adenylyl cyclases are required formemory consolidation and activation of the cAMP/MAPK/CREB transcriptional pathway during REM sleepWe reasoned that the increase in the cAMP/MAPK/CREB signal-ing cascade during REM sleep might contribute to memory con-solidation. However, because REM sleep in mice is very brief(3.1 � 0.1 min) and occurs at a low frequency (3.8 � 0.1% of totalrecording time), it is impossible to block the pathway exclusivelyduring REM sleep using chemical inhibitors. Instead, we chose atransgenic mouse model showing deficits in memory consolida-tion and examined the activation of the cAMP/MAPK/CREBpathway during sleep. DKO mice have impairments in long-termpotentiation (LTP) at the Schaffer collateral-CA1 synapses(Wong et al., 1999; Zhang et al., 2011), as well as in long-term
Figure 4. Elevated pCREB immunoreactivity in the CA1 and DG during REM sleep. A, Immunofluorescent images of pCREB in the CA1 (outlined with brackets) and DG from mice during waking,REM sleep, and NREM sleep. Scale bar, 200 �m. Or, Oriens layer; Py, pyramidal layer; Rad, radiatum layer; MoDG, molecular dentate gyrus; GrDG, granular dentate gyrus; PoDG, polymorph dentategyrus. B, C, Quantification of the relative fluorescent intensity of pCREB in the pyramidal layer of the CA1 (B) and the granular layer of the DG (C). Data are mean � SEM; n � 5– 8 per group. *p �0.05 (one-way ANOVA, Bonferroni’s post hoc).
6464 • J. Neurosci., April 10, 2013 • 33(15):6460 – 6468 Luo et al. • cAMP/MAPK/CREB Signaling Elevates during REM Sleep

memory for several hippocampus-dependent tasks (Wong et al.,1999; Wang et al., 2004; Zhang et al., 2011). When subjected topassive avoidance test, both WT and DKO mice exhibited in-creased crossover latency at comparable levels 3 h after training(two-way ANOVA, genotype, F(1,24) � 1.37, p � 0.2537; trial,F(1,24) � 49.36, p � 0.0001; interaction, F(1,24) � 2.04, p � 0.1660;Fig. 6A), suggesting that DKO mice have normal learning andshort-term memory for passive avoidance task. However, whentested for long-term passive avoidance memory, DKO miceshowed significantly shorter crossover latency compared with WTmice 24 h after training (two-way ANOVA, genotype, F(1,32) �25.47, p � 0.0001; trial, F(1,32) � 64.82, p � 0.0001; interaction,F(1,32) � 25.98, p � 0.0001; Fig. 6B). In another test where mem-ory for contextual fear was evaluated, DKO mice learned to asso-ciate the contextual chamber with aversive foot shock anddemonstrated elevated freezing behavior similar to that of WTmice 3 h after training (two-way ANOVA, genotype, F(1,24) �0.48, p � 0.4956; trial, F(1,24) � 70.89, p � 0.0001; interaction,F(1,24) � 0.21, p � 0.6486; Fig. 6C). However, DKO mice spentsignificantly less time in freezing than WT mice 24 h and 8 d after train-ing (two-way ANOVA, genotype, F(1,21) � 51.77, p � 0.0001; trial,F(2,21) � 30.15, p � 0.0001; interaction, F(2,21) � 14.42, p �0.0001; Fig. 6D). These results indicate that consolidation ofhippocampus-dependent memory is compromised in DKO mice
and that calmodulin-stimulated adenylyl cyclases are indispens-able for memory consolidation.
MAPK activity, CREB phosphorylation, and cAMP/PKA sig-naling during sleep and awake periods were analyzed in DKOmice. Quantification of pERK� cells revealed no significant dif-ferences among waking, REM sleep, and NREM sleep in thepyramidal layer of the CA1 (one-way ANOVA, F(2,17) � 3.476,p � 0.0543; Fig. 7A). In addition, the total fluorescent intensity ofpCREB in the CA1 remained relatively constant among threebehavioral states (one-way ANOVA, F(2,17) � 1.119, p � 0.3495;Fig. 7B), suggesting that REM sleep-associated CREB activationwas abolished in DKO mice. We also monitored cAMP levels andPKA activity in the hippocampus of DKO mice. No differences incAMP levels (Fig. 7C) or PKA activity (Fig. 7D,E) were detectedbetween waking and REM sleep. However, in DKO mice, cAMPand PKA activity was higher in NREM sleep compared withawake mice (cAMP, one-way ANOVA, F(2,18) � 4.659, p �0.0234; band 1, one-way ANOVA, F(2,18) � 6.950, p � 0.0058;band 2, one-way ANOVA, F(2,18) � 10.38, p � 0.0010; band 3,one-way ANOVA, F(2,18) � 18.98, p � 0.0001; Fig. 7C–E). We donot know why there is an increase in cAMP during NREM sleep inDKO mice. However, there are at least 7 different adenylyl cycla-ses expressed in the hippocampus with different regulatory prop-erties (Hanoune and Defer, 2001). The fact that one can see adifferential increase in cAMP during REM sleep but not NREMsleep in WT mice clearly indicates that there are different mech-anisms for activation of adenylyl cyclases in REM and NREMsleep. The higher levels of cAMP and PKA activity during NREMsleep may result from compensation because of the loss ofcalmodulin-stimulated adenylyl cyclases. Nevertheless, the in-creases in pERK, pCREB, and cAMP/PKA during REM sleep rel-ative to awake animals observed in WT mice are lost in DKOmice, a memory-deficient mouse strain. Collectively, our datasuggest that calmodulin-stimulated adenylyl cyclases are re-quired for activation of the cAMP/MAPK/CREB pathway duringREM sleep and that the preferential activation of this pathwayduring REM sleep may contribute to memory consolidation.
DiscussionThe cAMP, MAPK, and CREB-mediated transcription is acti-vated during the formation of hippocampus-dependent long-term memory (Atkins et al., 1998; Blum et al., 1999; Wong et al.,1999; Pittenger et al., 2002; Sindreu et al., 2007), an event thatrequires de novo transcription and translation (Davis and Squire,1984; Silva et al., 1998; Impey et al., 1999). However, training-induced increases in cAMP, MAPK activity, and CREB phos-phorylation are transient (Bernabeu et al., 1997; Sindreu et al.,2007), and the newly synthesized synaptic proteins have limitedlifetimes. Therefore, it is not clear how hippocampus-dependentmemories are maintained over extended periods of time. Werecently discovered that the cAMP/MAPK/CREB transcriptionalpathway in the mouse hippocampus is repeatedly activated dur-ing the circadian cycle with a maximum during the daytime(Eckel-Mahan et al., 2008) that depends upon the suprachias-matic nucleus (SCN) (Phan et al., 2011). Disruption of this oscil-lation after acquisition and consolidation impairs the persistenceof memory. Because mice are nocturnal and exhibit polyphasicsleep patterns during the daytime, it raised the interesting possi-bility that this memory consolidation pathway may be activatedduring sleep. The objective of this study was to monitor thecAMP/MAPK/CREB transcriptional pathway during variousstages of sleep.
Figure 5. cAMP/PKA signaling increases during REM sleep. A, cAMP levels of hippocampitaken from mice during waking, REM sleep, and NREM sleep. Data are mean � SEM; n � 5 or6 per group. *p � 0.05 (one-way ANOVA, Bonferroni’s post hoc). B, Images of immunoblotsprobed with pPKA-s and actin. Blots show individually run samples. pPKA-s bands labeled 1–3were analyzed. The ladder is indicated on the right. C, Summary of the relative levels of threepPKA-s bands (1–3). Data are mean � SEM; n � 5 or 6 per group. *p � 0.05 (one-way ANOVA,Bonferroni’s post hoc). **p � 0.01 (one-way ANOVA, Bonferroni’s post hoc). ***p � 0.001(one-way ANOVA, Bonferroni’s post hoc).
Luo et al. • cAMP/MAPK/CREB Signaling Elevates during REM Sleep J. Neurosci., April 10, 2013 • 33(15):6460 – 6468 • 6465

In this study, we took advantage of theEEG/EMG recording system and moni-tored cAMP, MAPK activity, and CREBphosphorylation in the hippocampus ofmice during REM sleep, NREM sleep, andwhen mice were awake. MAPK activation,measured as pERK, has been implicated inconsolidation of hippocampus-dependentmemory (Kelly et al., 2003; Kelleher et al.,2004). When comparing MAPK activityamong waking, REM sleep, and NREMsleep, we detected much higher pERK lev-els in the CA1 and DG during REM sleepcompared with waking but not whencomparing NREM sleep with waking. Wealso examined the levels of phospho-p38MAPK, which oscillates out of phase rela-tive to pERK in the SCN (Obrietan et al.,1998; Pizzio et al., 2003), in sleeping andawake animals. However, no differenceswere detected among different behavioralstates (data not shown).
Because MAPK mediates phosphory-lation and transactivation of CREB dur-ing memory formation (Xing et al.,1996; Impey et al., 1998b; Sindreu et al.,2007), and more importantly, CREB-mediated transcription is required for consol-idation of hippocampus-dependent memory(Bourtchuladze et al., 1994; Athos et al.,2002; Pittenger et al., 2002), we examinedCREB phosphorylation in REM and
Figure 6. DKO mice exhibit impairments in consolidating hippocampus-dependent memory. A, WT (open bars) and DKO(closed bars) mice were trained in the passive avoidance test, and crossover latency was measured 3 h after training. Data aremean � SEM; n � 7 per group. B, WT and DKO mice were tested for passive avoidance memory 24 h after training. Data aremean � SEM; n � 8 –10 per group. ***p � 0.001 (two-way ANOVA, Bonferroni’s post hoc). C, WT and DKO mice were trained forcontextual learning, and freezing percentage was measured 3 h after training. Data are mean � SEM; n � 7 per group. D, WT andDKO mice were tested for contextual fear memory 24 h and 8 d after training. Data are mean � SEM; n � 4 or 5 per group. **p �0.01 (two-way ANOVA, Bonferroni’s post hoc). ***p � 0.001 (two-way ANOVA, Bonferroni’s post hoc).
Figure 7. The cAMP/MAPK/CREB activation is attenuated during REM sleep in DKO mice. A, Quantification of the numbers of pERK � cells per section in the CA1 of DKO mice during waking, REMsleep, and NREM sleep. Data are mean � SEM; n � 6 or 7 per group. B, Analysis of the relative fluorescent intensity of pCREB in the CA1 of DKO mice in three behavioral states. Data are mean � SEM;n � 6 – 8 per group. C, Hippocampal cAMP from DKO mice in waking, REM sleep, and NREM sleep. Data are mean � SEM; n � 7 per group. *p � 0.05 (one-way ANOVA, Bonferroni’s post hoc).D, Images of immunoblots probed with pPKA-s and actin. Blots show individually run samples. pPKA-s bands labeled 1–3 were analyzed. The ladder is indicated on the right. E, Summary of therelative levels of three pPKA-s bands (1–3). Data are mean � SEM; n � 7 per group. *p � 0.05 (one-way ANOVA, Bonferroni’s post hoc). **p � 0.01 (one-way ANOVA, Bonferroni’s post hoc).***p � 0.001 (one-way ANOVA, Bonferroni’s post hoc).
6466 • J. Neurosci., April 10, 2013 • 33(15):6460 – 6468 Luo et al. • cAMP/MAPK/CREB Signaling Elevates during REM Sleep

NREM sleep. We used a phosphopeptide-specific antibody forCREB phosphorylated at Ser-133 (Impey et al., 1998a) and dis-covered that pCREB levels were significantly higher in REM sleepcompared with waking but not when comparing NREM sleepwith waking.
MAPK is activated by cAMP in the hippocampus (Impey et al.,1998b; Roberson et al., 1999). We next examined cAMP levelsand discovered that cAMP was also higher during REM sleepcompared with awake animals. In addition, to assess cAMP-dependent PKA activity between waking and sleep, we used anantibody that detects the downstream substrates sharing a con-served PKA phosphorylation motif as a general indicator for PKAactivation (Sindreu et al., 2007). We detected elevated levels oftwo PKA substrates ranging from 50 to 75 kDa during REM sleep.One potential candidate of these substrates is STEP61, the onlyisoform of the striatal-enriched protein tyrosine phosphatase(STEP) family that is expressed in the hippocampus (Boulangeret al., 1995). STEP is phosphorylated directly by PKA (Paul et al.,2000). In addition, the activity of STEP is also regulated by PKA-activated DARPP-32 (dopamine and cyclic adenosine 3�,5�-monophosphate-regulated phosphoprotein, 32 kDa), whichblocks the inhibitory activity of protein phosphatase-1 and there-fore retains STEP in phosphorylated state (Greengard et al., 1999;Fitzpatrick and Lombroso, 2011). The resulting phosphorylationof STEP prevents it from interacting with and inactivating pERKby dephosphorylation at the regulatory tyrosine residue (Paul etal., 2003). To evaluate the levels of phospho-STEP61 during wak-ing and sleep, we processed hippocampal lysates for Westernanalysis and observed an increase in phospho-STEP61 duringREM sleep compared with waking or NREM sleep. However,these differences were not statistically significant (data notshown). Nevertheless, our data have shown that CREB, an im-portant downstream target of the cAMP/PKA pathway, is acti-vated during REM sleep (Fig. 4). Together, our results indicatethat cAMP, MAPK activity, and CREB phosphorylation are sig-nificantly higher in REM sleep, but not in NREM sleep, comparedwith awake mice.
The fact that the cAMP/MAPK/CREB transcriptional path-way is elevated during REM sleep suggests the intriguing hypoth-esis that the activation of this pathway during REM sleep maycontribute to memory consolidation. To test this hypothesis, onehas to demonstrate that blocking the activation of this pathwayduring REM sleep compromises memory consolidation. How-ever, it is impossible to disrupt this pathway specifically duringthe brief periods of REM sleep with chemical inhibitors. Wetherefore used DKO transgenic mice as a tool to blockcalmodulin-stimulated cAMP increases during REM sleep. Thereis no difference between the sleep architecture of DKO and WTmice (J.L. and D.R.S., unpublished observations). However,DKO mice exhibit impairments in consolidating hippocampus-dependent memory and no increases in cAMP, MAPK activity, orCREB phosphorylation during REM sleep. These data supportthe hypothesis that memory consolidation depends uponcAMP signals generated by calmodulin-stimulated adenylylcyclases during REM sleep that activate MAPK and CREB-mediated transcription.
There is considerable evidence supporting the idea that mem-ory consolidation may occur during REM sleep in rodents. Train-ing leads to an increase in subsequent REM sleep, the onset timeand duration of which vary depending upon task complexity,animal species, and training protocols (Smith, 1996; Peigneuxet al., 2001; Rauchs et al., 2005). In addition, depriving REM sleepafter training interferes with spatial reference memory in maze
learning (Smith and Rose, 1996; Youngblood et al., 1997; Smith etal., 1998). Furthermore, neuronal activities reflecting prior wak-ing experience are reproduced in a temporal order during subse-quent REM sleep (Poe et al., 2000; Louie and Wilson, 2001).However, hippocampal ensemble discharges seen in the earlierlearning episodes are also detected during SWS (Wilson and Mc-Naughton, 1994; Skaggs and McNaughton, 1996; Shen et al.,1998; Ji and Wilson, 2007). These results suggest that REM sleepand SWS may play complementary roles in consolidating mem-ory obtained from preceding wakefulness. In conclusion, ourdata indicate that the activation of the cAMP/MAPK/CREB tran-scriptional pathway during REM sleep may play an importantrole in consolidation of hippocampus-dependent memory.
ReferencesAllada R, Siegel JM (2008) Unearthing the phylogenetic roots of sleep. Curr
Biol 18:R670 –R679. CrossRef MedlineAserinsky E, Kleitman N (1953) Regularly occurring periods of eye motility,
and concomitant phenomena, during sleep. Science 118:273–274.CrossRef Medline
Athos J, Impey S, Pineda VV, Chen X, Storm DR (2002) Hippocampal CRE-mediated gene expression is required for contextual memory formation.Nat Neurosci 5:1119 –1120. CrossRef Medline
Atkins CM, Selcher JC, Petraitis JJ, Trzaskos JM, Sweatt JD (1998) TheMAPK cascade is required for mammalian associative learning. Nat Neu-rosci 1:602– 609. CrossRef Medline
Bernabeu R, Cammarota M, Izquierdo I, Medina JH (1997) Involvement ofhippocampal AMPA glutamate receptor changes and the cAMP/proteinkinase A/CREB-P signalling pathway in memory consolidation of anavoidance task in rats. Braz J Med Biol Res 30:961–965. CrossRef Medline
Blum S, Moore AN, Adams F, Dash PK (1999) A mitogen-activated proteinkinase cascade in the CA1/CA2 subfield of the dorsal hippocampus isessential for long-term spatial memory. J Neurosci 19:3535–3544.Medline
Boulanger LM, Lombroso PJ, Raghunathan A, During MJ, Wahle P, NaegeleJR (1995) Cellular and molecular characterization of a brain-enrichedprotein tyrosine phosphatase. J Neurosci 15:1532–1544. Medline
Bourtchuladze R, Frenguelli B, Blendy J, Cioffi D, Schutz G, Silva AJ (1994)Deficient long-term memory in mice with a targeted mutation of thecAMP-responsive element-binding protein. Cell 79:59 – 68. CrossRefMedline
Campbell SS, Tobler I (1984) Animal sleep: a review of sleep duration acrossphylogeny. Neurosci Biobehav Rev 8:269 –300. CrossRef Medline
Carskadon MA, Dement WC (2011) Monitoring and staging human sleep.In: Principles and practice of sleep medicine, Ed 5 (Kryger TR, DementWC, eds), pp 16 –26. St. Louis: Elsevier Saunders.
Cirelli C, Tononi G (2008) Is sleep essential? PLoS Biol 6:e216. CrossRefMedline
Crocker A, Sehgal A (2010) Genetic analysis of sleep. Genes Dev 24:1220 –1235. CrossRef Medline
Davis HP, Squire LR (1984) Protein synthesis and memory: a review. Psy-chol Bull 96:518 –559. CrossRef Medline
Diekelmann S, Born J (2010) The memory function of sleep. Nat Rev Neu-rosci 11:114 –126. CrossRef Medline
Eckel-Mahan KL, Phan T, Han S, Wang H, Chan GC, Scheiner ZS, Storm DR(2008) Circadian oscillation of hippocampal MAPK activity and cAMP:implications for memory persistence. Nat Neurosci 11:1074 –1082.CrossRef Medline
Fitzpatrick CJ, Lombroso PJ (2011) The role of striatal-enriched proteintyrosine phosphatase (STEP) in cognition. Front Neuroanat 5:47.CrossRef Medline
Greengard P, Allen PB, Nairn AC (1999) Beyond the dopamine receptor:the DARPP-32/protein phosphatase-1 cascade. Neuron 23:435– 447.CrossRef Medline
Hanoune J, Defer N (2001) Regulation and role of adenylyl cyclase iso-forms. Annu Rev Pharmacol Toxicol 41:145–174. CrossRef Medline
Hernandez PJ, Abel T (2011) A molecular basis for interactions betweensleep and memory. Sleep Med Clin 6:71– 84. CrossRef Medline
Impey S, Smith DM, Obrietan K, Donahue R, Wade C, Storm DR (1998a)Stimulation of cAMP response element (CRE)-mediated transcriptionduring contextual learning. Nat Neurosci 1:595– 601. CrossRef Medline
Luo et al. • cAMP/MAPK/CREB Signaling Elevates during REM Sleep J. Neurosci., April 10, 2013 • 33(15):6460 – 6468 • 6467

Impey S, Obrietan K, Wong ST, Poser S, Yano S, Wayman G, Deloulme JC,Chan G, Storm DR (1998b) Cross talk between ERK and PKA is re-quired for Ca 2� stimulation of CREB-dependent transcription and ERKnuclear translocation. Neuron 21:869 – 883. CrossRef Medline
Impey S, Obrietan K, Storm DR (1999) Making new connections: role ofERK/MAP kinase signaling in neuronal plasticity. Neuron 23:11–14.CrossRef Medline
Ji D, Wilson MA (2007) Coordinated memory replay in the visual cortexand hippocampus during sleep. Nat Neurosci 10:100 –107. CrossRefMedline
Kelleher RJ 3rd, Govindarajan A, Jung HY, Kang H, Tonegawa S (2004)Translational control by MAPK signaling in long-term synaptic plasticityand memory. Cell 116:467– 479. CrossRef Medline
Kelly A, Laroche S, Davis S (2003) Activation of mitogen-activated proteinkinase/extracellular signal-regulated kinase in hippocampal circuitry isrequired for consolidation and reconsolidation of recognition memory.J Neurosci 23:5354 –5360. Medline
Lesku JA, Rattenborg NC, Amlaner CJ Jr (2006) The evolution of sleep: aphylogenetic approach. In: Sleep: a comprehensive handbook (Lee-Chiong T, ed), pp 49 – 61. Hoboken, New Jersey: Wiley.
Louie K, Wilson MA (2001) Temporally structured replay of awake hip-pocampal ensemble activity during rapid eye movement sleep. Neuron29:145–156. CrossRef Medline
Marshall L, Born J (2007) The contribution of sleep to hippocampus-dependent memory consolidation. Trends Cogn Sci 11:442– 450.CrossRef Medline
Obrietan K, Impey S, Storm DR (1998) Light and circadian rhythmicityregulate MAP kinase activation in the suprachiasmatic nuclei. Nat Neu-rosci 1:693–700. CrossRef Medline
Paul S, Snyder GL, Yokakura H, Picciotto MR, Nairn AC, Lombroso PJ(2000) The dopamine/D1 receptor mediates the phosphorylation andinactivation of the protein tyrosine phosphatase STEP via a PKA-dependent pathway. J Neurosci 20:5630 –5638. Medline
Paul S, Nairn AC, Wang P, Lombroso PJ (2003) NMDA-mediated activa-tion of the tyrosine phosphatase STEP regulates the duration of ERKsignaling. Nat Neurosci 6:34 – 42. CrossRef Medline
Peigneux P, Laureys S, Delbeuck X, Maquet P (2001) Sleeping brain, learningbrain: the role of sleep for memory systems. Neuroreport 12:A111–A124.CrossRef Medline
Phan TX, Phan TH, Chan GC, Sindreu CB, Eckel-Mahan KL, Storm DR(2011) The diurnal oscillation of MAP (mitogen-activated protein) ki-nase and adenylyl cyclase activities in the hippocampus depends on thesuprachiasmatic nucleus. J Neurosci 31:10640 –10647. CrossRef Medline
Pittenger C, Huang YY, Paletzki RF, Bourtchouladze R, Scanlin H, VronskayaS, Kandel ER (2002) Reversible inhibition of CREB/ATF transcriptionfactors in region CA1 of the dorsal hippocampus disrupts hippocampus-dependent spatial memory. Neuron 34:447– 462. CrossRef Medline
Pizzio GA, Hainich EC, Ferreyra GA, Coso OA, Golombek DA (2003) Cir-cadian and photic regulation of ERK, JNK and p38 in the hamster SCN.Neuroreport 14:1417–1419. CrossRef Medline
Poe GR, Nitz DA, McNaughton BL, Barnes CA (2000) Experience-dependent phase-reversal of hippocampal neuron firing during REMsleep. Brain Res 855:176 –180. CrossRef Medline
Rauchs G, Desgranges B, Foret J, Eustache F (2005) The relationships be-tween memory systems and sleep stages. J Sleep Res 14:123–140. CrossRefMedline
Rechtschaffen A, Kales A (eds) (1968) A manual of standardized terminol-ogy, techniques and scoring system for sleep stages of human subjects.Bethesda, Maryland: U.S. National Institute of Neurological Diseases andBlindness, Neurological Information Network.
Roberson ED, English JD, Adams JP, Selcher JC, Kondratick C, Sweatt JD(1999) The mitogen-activated protein kinase cascade couples PKA andPKC to cAMP response element binding protein phosphorylation in areaCA1 of hippocampus. J Neurosci 19:4337– 4348. Medline
Shen J, Kudrimoti HS, McNaughton BL, Barnes CA (1998) Reactivation ofneuronal ensembles in hippocampal dentate gyrus during sleep after spa-tial experience. J Sleep Res 7 [Suppl 1]:6 –16.
Siegel JM (2008) Do all animals sleep? Trends Neurosci 31:208 –213.CrossRef Medline
Siegel JM (2009) Sleep viewed as a state of adaptive inactivity. Nat Rev Neu-rosci 10:747–753. CrossRef Medline
Silva AJ, Kogan JH, Frankland PW, Kida S (1998) CREB and memory. AnnuRev Neurosci 21:127–148. CrossRef Medline
Sindreu CB, Scheiner ZS, Storm DR (2007) Ca 2� stimulated adenylyl cycla-ses regulate ERK-dependent activation of MSK1 during fear condition-ing. Neuron 53:79 – 89. CrossRef Medline
Skaggs WE, McNaughton BL (1996) Replay of neuronal firing sequences inrat hippocampus during sleep following spatial experience. Science 271:1870 –1873. CrossRef Medline
Smith C (1996) Sleep states, memory processes and synaptic plasticity.Behav Brain Res 78:49 –56. CrossRef Medline
Smith C, Rose GM (1996) Evidence for a paradoxical sleep window for placelearning in the Morris water maze. Physiol Behav 59:93–97. CrossRefMedline
Smith CT, Conway JM, Rose GM (1998) Brief paradoxical sleep deprivationimpairs reference, but not working, memory in the radial arm maze task.Neurobiol Learn Mem 69:211–217. CrossRef Medline
Stickgold R (1998) Sleep: off-line memory reprocessing. Trends Cogn Sci2:484 – 492. CrossRef Medline
Stickgold R (2005) Sleep-dependent memory consolidation. Nature 437:1272–1278. CrossRef Medline
Tobler I, Deboer T, Fischer M (1997) Sleep and sleep regulation in normaland prion protein-deficient mice. J Neurosci 17:1869 –1879. Medline
Veasey SC, Valladares O, Fenik P, Kapfhamer D, Sanford L, Benington J,Bucan M (2000) An automated system for recording and analysis ofsleep in mice. Sleep 23:1025–1040. Medline
Walker MP, Stickgold R (2004) Sleep-dependent learning and memoryconsolidation. Neuron 44:121–133. CrossRef Medline
Wang H, Ferguson GD, Pineda VV, Cundiff PE, Storm DR (2004) Overex-pression of type-1 adenylyl cyclase in mouse forebrain enhances recogni-tion memory and LTP. Nat Neurosci 7:635– 642. CrossRef Medline
Wilson MA, McNaughton BL (1994) Reactivation of hippocampal ensem-ble memories during sleep. Science 265:676 – 679. CrossRef Medline
Wong ST, Athos J, Figueroa XA, Pineda VV, Schaefer ML, Chavkin CC,Muglia LJ, Storm DR (1999) Calcium-stimulated adenylyl cyclase activ-ity is critical for hippocampus-dependent long-term memory and latephase LTP. Neuron 23:787–798. CrossRef Medline
Wu ZL, Thomas SA, Villacres EC, Xia Z, Simmons ML, Chavkin C, PalmiterRD, Storm DR (1995) Altered behavior and long-term potentiation intype I adenylyl cyclase mutant mice. Proc Natl Acad Sci U S A 92:220 –224. CrossRef Medline
Xing J, Ginty DD, Greenberg ME (1996) Coupling of the RAS-MAPK path-way to gene activation by RSK2, a growth factor-regulated CREB kinase.Science 273:959 –963. CrossRef Medline
Youngblood BD, Zhou J, Smagin GN, Ryan DH, Harris RB (1997) Sleepdeprivation by the “flower pot” technique and spatial reference memory.Physiol Behav 61:249 –256. CrossRef Medline
Zhang M, Storm DR, Wang H (2011) Bidirectional synaptic plasticity andspatial memory flexibility require Ca 2�-stimulated adenylyl cyclases.J Neurosci 31:10174 –10183. CrossRef Medline
6468 • J. Neurosci., April 10, 2013 • 33(15):6460 – 6468 Luo et al. • cAMP/MAPK/CREB Signaling Elevates during REM Sleep




















