
Fishy Behavior - Persistent effects of early-life exposure to 17α ...
69
Fishy Behavior
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of Fishy Behavior - Persistent effects of early-life exposure to 17α ...

Fishy Behavior

"The problem is not the problem.
It is your attitude towards the problem that is the problem." - Captain Jack Sparrow

Örebro studies in Biology 9 Södertörn Doctoral Dissertations 110
KRISTINA VOLKOVA
Fishy Behavior Persistent effects of early-life exposure to 17α-ethinylestradiol

© Kristina Volkova, 2015
Title: Fishy Behavior - Persistent effects of early -life exposure to 17α-ethinylestradiol
Publisher: ÖrebroUniversity 2015www.oru.se/publikationer-avhandlingar
Print: Örebro University, Repro 09/2015
ISSN 1650-8793 ISSN 1652-7399
ISBN 978-91-7529-091-1

Abstract Kristina Volkova (2015): Fishy Behavior - Persistent effects of early-life exposure to 17α-ethinylestradiol. Örebro Studies in Biology 9. The synthetic estrogen 17α-ethinylestradiol (EE2) is an endocrine disrupting chemical (EDC) of concern due to its persistent nature and widespread presence in the aquatic environment. In mammals, effects of developmental EDC exposure on reproduction and behavior not only persist to adulthood after discontinued exposure, but are also inherited by several consecutive unexposed generations. The results presented in this thesis demonstrate that non-reproductive behavior in fish is highly sensitive to the influence of EE2 during development and the effects do not appear to be restored after a long recovery period in clean water. We have shown that exposure to low doses of EE2 during development results in increased anxiety in two fish species (zebrafish and guppy) and their offspring. We have also demon-strated that the effects of EE2 on anxiety are apparent in both sexes and are transgenerationally transmitted to two consecutive generations of unex-posed offspring in the guppy. In order to investigate the possible biological mechanisms of the observed persistent effects on non-reproductive behav-ior, we also performed an RNA sequencing analysis of the whole-brain transcriptome in developmentally exposed zebrafish after remediation in clean water until adulthood. Differential expression of 33 genes in males and 62 genes in females were observed as a result of EE2 exposure, with only one gene affected in both sexes. Functional analysis revealed choles-terol biosynthesis and circadian rhythm to be the top two affected path-ways in males and females, respectively. Both pathways have previously been implicated in anxiety behavior and represent possible candidates con-necting the transcriptome alterations to the observed behavioral phenotype. The study represents an initial survey of the fish brain transcriptome by means of RNA sequencing after long-term recovery from developmental exposure to an estrogenic compound.
Keywords: Endocrine disruptors, anxiety, stress behavior, transgenerational effects, 17α-ethinylestradiol, developmental exposure, social behavior, fish. Kristina Volkova, School of Science and Technology Örebro University, SE-701 82 Örebro, Sweden, [email protected]


Abbreviations BPA: Bisphenol A DES: Diethylstilbestrol dpf: Days post fertilization EDCs: Endocrine disrupting chemicals EE2: 17α-ethinylestradiol FDR: False discovery rate MDD: Major depressive disorder mRNA: Messenger ribonucleic acid ncRNA: Non-coding RNA PAHs: Polycyclic aromatic hydrocarbons PCBs: Polychlorinated biphenyls PCR: Polymerase chain reaction qPCR: Quantitative real-time polymerase chain reaction RNA: Ribonucleic acid RNAseq: RNA sequencing analysis VTG: Vitellogenin


List of papers This thesis is based on the following four papers. Paper I. Volkova K, Reyhanian N, Kot-Wasik A, Olsén H, Porsch-Hällström I, Hallgren S. Brain circuit imprints of developmental 17α-Ethinylestradiol exposure in guppies (Poecilia reticulata): persistent effects on anxiety but not on reproductive behaviour. Gen Comp Endocrinol. 2012 Sep 1;178(2):282-90. doi: 10.1016/j.ygcen.2012.05.010. Paper II. Volkova K*, Reyhanian Caspillo N*, Porseryd T, Hallgren S, Dinnétz P, Olsén H, Porsch-Hällström I.Transgenerational effects of 17α-Ethi-nylestradiol on anxiety behavior in the guppy, Poecilia reticula. Gen Comp Endocrinol. Submitted 2015 Paper III. Volkova K, Reyhanian Caspillo N, Porseryd T, Hallgren S, Dinnétz P, Porsch-Hällström I. Developmental exposure of zebrafish (Danio rerio) to 17α-ethinylestradiol affects non-reproductive behavior and fertility as adults, and increases anxiety in unexposed progeny. Horm Behav. 2015 Jun 10;73:30-38. doi: 10.1016/j.yhbeh.2015.05.014. Paper IV. Porseryd T*, Volkova K*, Reyhanian Caspillo N, Källman T, Dinnétz P, Porsch-Hällström I. Persistent effects of developmental exposure to 17α-ethinylestradiol on the zebrafish (Danio rerio) brain transcriptome and stress behavior. Manuscript * These authors are to be considered first authors.


Table of Contents
INTRODUCTION ...................................................................................13 Endocrine disrupting chemicals................................................................. 13 17α-ethinylestradiol .................................................................................. 15 Reversible effects ...................................................................................... 16 Effects on behavior ................................................................................... 17 Transgenerational effects .......................................................................... 19
OBJECTIVES ............................................................................................20
EXPERIMENTAL ANIMALS ..................................................................21 Zebrafish (Danio rerio) ............................................................................. 21 Guppy (Poecilia reticulata) ....................................................................... 22
METHODS...............................................................................................24 Exposure ................................................................................................... 24 Behavior .................................................................................................... 25
Novel tank test ..................................................................................... 25 Shoaling test ......................................................................................... 27 Scototaxis test ....................................................................................... 29 Guppy reproductive behavior ............................................................... 30
Fertility ..................................................................................................... 31 Guppy testis histology .......................................................................... 31 Zebrafish fertilization success ............................................................... 31
Gene expression ........................................................................................ 32 RNA sequencing ................................................................................... 32 Quantitative real-time PCR .................................................................. 33
Brain aromatase activity ........................................................................... 34 Statistical analysis of behavioral tests ....................................................... 34
RESULTS: SUMMARY OF PAPERS ........................................................35 I. Brain circuit imprints of developmental 17α-Ethinylestradiol exposure in guppies (Poecilia reticulata): persistent effects on anxiety but not on reproductive behaviour. .................................................. 35 II. Transgenerational effects of 17α-Ethinylestradiol on anxiety behavior in the guppy, Poecilia reticula. ........................................... 35 III. Developmental exposure of zebrafish (Danio rerio) to 17α-ethinylestradiol affects non-reproductive behavior and fertility as adults, and increases anxiety in unexposed progeny. .................... 36 IV. Persistent effects of developmental exposure to 17α-ethinylestradiol on the zebrafish (Danio rerio) brain transcriptome and stress behavior. ................................................... 36
DISCUSSION AND CONCLUSIONS ......................................................38
ACKNOWLEDGEMENTS .......................................................................44
REFERENCES ..........................................................................................46


KRISTINA VOLKOVA Fishy Behavior 13
Introduction
Endocrine disrupting chemicals Endocrine disrupting chemicals (EDCs) have the ability to interfere with the normal function of the endocrine system of humans and wildlife. EDCs can be naturally produced compounds such as endogenous estrogens and androgens, plant phytoestrogens, but also a wide range of industrial chemicals, synthetic hormones, pharmaceuticals and pesticides (Kabir et al. 2015). By altering the hormonal and homeostatic systems of a living organism, EDCs have the ability to influence a wide range of significant biological processes such as metabolism, growth, behavior, sexual development, reproduction and fetal development. Such effects of exposure have been observed for a large number of commonly occurring compounds in a wide variety of animal species (WHO/UNEP 2013). Their widespread presence in the environment and the rapid accumulation of research pointing to adverse effects of exposure make EDCs a rising concern. Since human experiments cannot be performed, it is difficult to study the effects of EDC exposure in humans. However, several examples exist where a compound has been shown to have adverse endocrine disruptive effects on exposed humans. One such example is diethylstilbestrol (DES), a synthetic estrogen which was administered to pregnant women in the 1950’s in order to prevent miscarriage and premature births. However, DES was later found to increase the incidence of vaginal cancer in females who had been exposed to the compound during their development (Herbst et al. 1971). In addition, a recent long-term follow-up study associated developmental DES exposure to an increased risk for a wide range of reproductive complications, including infertility, spontaneous abortion and preterm delivery (Hoover et al. 2011). Males developmentally exposed to DES have been found to have a higher prevalence of urogenital abnormalities compared to unexposed males (Palmer et al. 2009; Virtanen and Adamsson 2012). Some studies have also suggested a link between increased incidence of depression and developmental DES exposure (O'Reilly et al. 2010; Pillard et al. 1993). Another human example is exposure to a group of organic compounds known as polychlorinated biphenyls (PCBs). Up until the 1970’s, PCBs were widely used in the electrical industry where they mostly functioned as

14 KRISTINA VOLKOVA Fishy Behavior
coolants, fire retardants and insulating fluids. However, the use of PCB was banned after several unfortunate PCB poisoning incidents. It was discovered that exposure to PCB caused severe health problems including Chloracne, developmental delay and speech problems (Kodavanti 2005). Also, children of mothers who had ingested PCB-contaminated cooking oil were found to be hyperactive (Kodavanti 2005). A more recent example is the plasticizer and weak estrogen mimic Bisphenol A (BPA) (Beronius et al. 2010; Vandenberg et al. 2009). The use of BPA in baby bottles was banned by the European Union in 2011, after it was found to alter synapse formation in the brain of a non-human primate (Leranth et al. 2008). In humans, associations have been reported between urinary BPA-concentration and increased risk for diabetes and cardiovascular disease (Lang et al. 2008), as well as between blood levels of BPA and ovarian dysfunction (Takeuchi et al. 2004) and miscarriage (Sugiura-Ogasawara et al. 2005). Human studies are limited to analyzing correlations, making it difficult to infer causal relationships between exposure and the effects on human health. However, a large amount of evidence is now beginning to gather from different animal models. For example, exposure experiments in rodents show that BPA interferes with the development of the prostate, mammary glands (Nagel et al. 1997), central nervous system (Ishido et al. 2004) and non-reproductive behavior such as play and maze learning (Della Seta et al. 2006; Dessì-Fulgheri et al. 2002; Farabollini et al. 1999; Farabollini et al. 2002). Risk assessment of endocrine disrupting chemicals has proven to be especially difficult. One primary reason for this is the fact that it is nearly impossible to avoid contact with endocrine disruptors in human daily life. EDCs are present in everything from food packaging to building materials and drinking water (Kabir et al. 2015). Humans all over the world are continuously exposed to a mixture of hundreds of EDCs, many of which have been detected in blood and urine samples (Beronius et al. 2010; WHO/UNEP 2013), making it impossible to find a human control group free from EDC exposure. In addition, by interacting with the endocrine system, EDCs affect a complex molecular network with an unknown amount of possible endpoints. By acting through hormones that are involved in embryonic development, EDCs also have a potential delayed on-set of effects that may not become visible until adulthood (Patisaul and Adewale 2009). By interacting with the endocrine system, EDCs are thought

KRISTINA VOLKOVA Fishy Behavior 15
to have a non-monotonic dose-response relationship, where low doses do not necessarily mean a lower level of toxicity (Gore et al. 2006; Lagarde F. 2015; Vandenberg et al. 2012; WHO/UNEP 2013). It is important to acknowledge that not only humans are exposed to EDCs on a daily basis. Endocrine disrupting chemicals make their way into the environment, affecting wildlife in both terrestrial and aquatic ecosystems. In fact, when measuring EDCs in the tissues of different species, it was discovered that concentrations in wild top predators can exceed levels known to cause effect in laboratory animal studies (WHO/UNEP 2013).
17α-ethinylestradiol 17α-ethinylestradiol (EE2) is a synthetic estrogen and main component of oral contraceptive pills. It is also regularly used on livestock to promote growth (Gadd et al. 2010; Kabir et al. 2015). EE2 is released into the environment through wastewater effluent from municipal treatment plants, hospital effluent and livestock industry (Aris et al. 2014). While wastewater effluent containing EE2 is often discharged into rivers, lakes and coastal waters, the sludge is used as fertilizer in agricultural fields (Vulliet and Cren-Olivé 2011), further contributing to the spread of EE2 in the environment. Outside of wastewater treatment plants, EE2 concentrations range from below detectable levels (<1ng/L) to as high as 200-300 ng/L (Hannah et al. 2009; Kolpin et al. 2002; Laurenson et al. 2014; Ternes et al. 1999). EE2 has also been detected in German drinking water at 0.5 ng/L (Kuch and Ballschmiter 2001). This raises cause for concern as the predicted no adverse effect level of EE2 for water-living organisms was recently decreased to 0.1 ng/L (Caldwell et al. 2012). EE2 is a potent EDC due to its high resistance to biodegradation and ability to accumulate in sediment and biota (Aris et al. 2014). Numerous experiments have demonstrated that EE2 has a toxic effect on a large number of organisms and has the ability to disturb important physiological functions (Filby et al. 2007) such as embryonic development, somatic growth (Van den Belt et al. 2003), immune function (Law et al. 2001) and stress response (Dugard et al. 2001).
In mammals, EE2 has been shown to lead to long-lasting growth retardation in exposed male rats and decreased litter size in their progeny (Vosges et al.

16 KRISTINA VOLKOVA Fishy Behavior
2008). EE2 exposure has also been shown to induce abnormalities in the reproductive organs of female rats and their offspring (Delclos et al. 2009), as well as to decrease testosterone production in the rat testis (Larcher et al. 2012). Mice developmentally exposed to EE2 through their mothers showed significant growth retardation and increased number of gonadotropin-releasing hormone neurons, demonstrating the ability of EE2 to modulate neurogenesis (Pillon et al. 2012). In addition, prenatal EE2 exposure has been shown to increase fear, anxiety and social neophobia in adult rats (Dugard et al. 2001). In fish, EE2 has been found to be >10 fold more potent than endogenous estrogen (Thorpe et al. 2003). EE2 exposure has been shown to lead to skewed sex-ratios, decreased egg and sperm production, reduced fertility, feminization of male fish, increased proportions of intersex fish and wide range of behavioral changes (Aris et al. 2014). In fact, continuously adding 5 ng/L EE2 to an experimental lake during 3 years resulted in a total collapse of a wild fish population (Kidd et al. 2007).
Reversible effects It has been demonstrated that the persistency of the effects of EDC exposure are highly dependent on species, timing of exposure, duration of exposure and concentration. The reversibility of the effects of EDC exposure also differ among different EDCs. When effects on spermatogenesis after exposure to the insecticide Lindane were compared to the effects of DES, observations showed that while mice developmentally exposed to lindane recovered, mice developmentally exposed to DES did not (Traina et al. 2003). In fish, continuous exposure to EE2 has been shown to result in complete reproductive failure and population collapse (Kidd et al. 2007; Nash et al. 2004). However, several publications describe the ability of fish to, at least partly, recover from estrogenic exposure after remediation in clean water (Baumann et al. 2014; Fenske et al. 2005; Larsen et al. 2009; Maack and Segner 2004; Schäfers et al. 2007; Van den Belt et al. 2002; Weber et al. 2003) For example, zebrafish developmentally exposed to 1, 3 or 10 ng/L EE2 during 60 or 100 days showed increased expression of the egg yolk precursor protein vitellogenin (VTG), skewed sex ratios, delay of gonad maturation and altered body size. However, after 40 days of recovery in

KRISTINA VOLKOVA Fishy Behavior 17
clean water, only body size remained significantly altered in the developmentally exposed zebrafish (Baumann et al. 2014). Other experiments indicate that the reversibility of EE2 effects on the reproductive organs and fertility depends on dose and timing of exposure (Fenske et al. 2005; Schäfers et al. 2007). In a study by Schäfers et al. a 177 day exposure to 9.3 ng/L EE2 caused inhibition of reproduction that could not be restored after a 3 month remediation period in clean water, while a shorter 75-day exposure to the same concentration only partly and reversibly impaired reproduction. The reproductive organs appear to have an ability to recover from EDC exposure after remediation in clean water, while effects on functional reproductive capacity seem to be more persistent (Hill Jr and Janz 2003). Long lasting developmental exposures are more likely to lead to irreversible effects on sexual structures and function, while short-term exposure of adults induces effects that can be reversed after discontinued exposure (McLachlan 2001). When adult zebrafish were exposed to 10 or 25 ng/L EE2 during 24 days, the observed effects on reproductive organs and VTG expression were completely reversed after recovery in clean water (Van den Belt et al. 2002). Recovery after EDC exposure has also been observed on a population level. When the collapsed fish population of the aforementioned EE2-exposed experimental lake was re-examined after 4 years of recovery, the size structure and abundance of adult fathead minnow had been restored (Blanchfield et al. 2015).
Effects on behavior The development of the reproductive system, sexual maturation and programming of later adult behavior is highly dependent on the correct timing of exposure and concentration of endogenous hormones (Bhandari et al. 2015a). Disrupting the hormonal balance at an early stage of development can therefore not only disturb gonad development but also interfere with the development of brain regions involved in adult endocrine and behavioral responses (McEwen 1987). Brain development is tightly regulated and guided not only by transcription factors but also by endogenous hormones such as gonadal steroids (Fernandez-Galaz et al. 1997), making the brain a potential target for EDCs (Gore 2008). In fact, EDCs have been shown to affect the neurodevelopmental process, brain morphology, cognitive functions, emotional reactivity and stress (Frye et al. 2012).

18 KRISTINA VOLKOVA Fishy Behavior
One well-studied endpoint of EDC exposure in mammals is reproductive behavior (Gore 2008; Patisaul and Adewale 2009). In female rats, developmental EDC exposure has been found to reduce sexual receptivity (Chung et al. 2001; Wang et al. 2002), mating behavior (Steinberg et al. 2007) and maternal behavior (Seta et al. 2005). In males, developmental exposure to EDCs resulted in deficits in sexual behavior (Jones et al. 2011), including reduced numbers of mounts (Henry and Witt 2006) and mating trials (Sager 1983).
A significant amount of studies have examined the effects of EDCs on reproductive behaviors also in fish (Söffker et al. 2012). Disturbed reproductive behavior has been observed in the three-spined stickleback (Espmark Wibe et al. 2002), goldfish (Bjerselius et al. 2001), guppy (Bayley et al. 1999; Shenoy 2014) and zebrafish (Larsen and Baatrup 2010). However, steroid hormones are believed to play a major role also in non-reproductive behavior. For example, developmental exposure to EE2 has been shown to increase fear, anxiety and social neophobia in adult rats (Dugard et al. 2001). Mice developmentally exposed to BPA displayed altered social behavior, increased anxiety, altered spatial recognition and impaired memory as adults (Della Seta et al. 2006; Ryan and Vandenbergh 2006; Wolstenholme et al. 2011; Xu et al. 2010). Increased anxiety is a well-known effect of EDC exposure in mammals (Gonçalves et al. 2010; Skinner et al. 2008; Valkusz et al. 2011), although sex, age, and timing of exposure affects the outcome (Gioiosa et al. 2013). In fish, studies examining non-reproductive behavior after developmental EDC exposure are scarce, but developmental BPA exposure has been shown to cause learning deficits in adult zebrafish (Saili et al. 2012). EDC exposure of adult fish has been shown to alter risky behavior, bottom dwelling and shoaling behavior (Bell 2004; Dzieweczynski et al. 2014; Espmark Wibe et al. 2002; Xia et al. 2010). In guppy and zebrafish males, we have previously shown that adult short term EE2 exposure increases anxiety (Hallgren et al. 2011; Reyhanian et al. 2011). We have also shown that exposure of adult zebrafish males increases shoaling intensity (Reyhanian et al. 2011). In addition, EE2 has been shown to affect aggressive behavior in zebrafish (Colman et al. 2009; Filby et al. 2012) and fathead minnow (Majewski et al. 2002).

KRISTINA VOLKOVA Fishy Behavior 19
Non-reproductive behavior such as exploratory behavior, and the ability to recover from stressors, is of high ecological significance in wild fish populations, who are the primary recipients of waterborne EDCs. Disturbances of these behaviors will therefore likely affect fitness by influencing foraging, reactions to predators and opportunities to reproduce.
Transgenerational effects Transgenerational inheritance is the transmittance of traits from one generation to several generations of progeny, meaning that phenotypical changes acquired by the parent not only persist during the life-time of that individual organism but can also be transmitted to their offspring. Transgenerational effects have been observed in a large variety of organisms, ranging from plants to mammals (Guerrero-Bosagna and Skinner 2012; Kleinmanns Julia and Schubert 2014; Skinner et al. 2010). In order to prove a transgenerational effect of EDC exposure, the experimental set up needs to include a generation that has not been exposed to the EDC in any other way than through their grandparents (Ho and Burggren 2010; McCarrey 2014). When an organism is exposed, the germ cells are also exposed to the EDC, meaning that any offspring this organism produces will have been indirectly exposed as germ cells within their parent. To meet the criteria of a transgenerational effect, the phenotype therefore needs to persist for at least two generations after initial exposure to the EDC. Transgenerational effects resulting from EDC exposure have been demonstrated in mammals on cancer, fertility and reproductive-, as well as non-reproductive behavior, such as anxiety (Anway et al. 2005; de Assis et al. 2012; Guerrero-Bosagna and Skinner 2012; Manikkam et al. 2013; Patisaul et al. 2012; Salian et al. 2009; Skinner et al. 2008; Wolstenholme et al. 2012). For example, mice exposed to BPA through their mothers showed altered social behavior that persisted also in the fourth (F4) generation (Wolstenholme et al. 2012). In rats, dietary intake of EE2 during pregnancy resulted in an increased risk of developing mammary gland cancer in daughters (F1), granddaughters (F2) and great-granddaughters (F3) of exposed females. Outcross experiments showed that the effect was transmitted through the female germline and associated with changes in the

20 KRISTINA VOLKOVA Fishy Behavior
DNA methylation machinery, as well as methylation patterns in mammary tissue (de Assis et al. 2012). Epigenetic changes, such as methylation, histone modifications and microRNA are believed to play a major role in transgenerational effects of EDCs in mammals (Anway et al. 2005; de Assis et al. 2012; Guerrero-Bosagna and Skinner 2012; Skinner et al. 2008). Epigenetic changes have also been demonstrated in studies showing transgenerational effects on the reproductive organs after exposure to BPA (Manikkam et al. 2013; Nilsson et al. 2012).
To date there are only four studies showing environmentally induced transgenerational effects in fish (Baker et al. 2014; Bhandari et al. 2015b; Corrales et al. 2014; Vignet et al. 2015). Out of these studies, only one has investigated the transgenerational effects of EE2. In this study, Medaka (Oryzias latipes) were exposed to 50 ng/L EE2 or 100 ug/L of BPA, during the first week of development. While no apparent phenotypic abnormalities were observed in the F0 or F1 generations, the F2 generation had a lower fertilization rate, and in the F3 and F4 generations there was a reduction in embryo survival compared to control animals (Bhandari et al. 2015b). No studies have yet investigated the heritability of changes in non-reproductive behavior after estrogen exposure in fish, but transgenerational effects on locomotor activity have been observed in zebrafish larvae after exposure to polycyclic aromatic hydrocarbons (PAHs) (Vignet et al. 2015).
Objectives The health risks of exposure to EDCs during development, not only for the exposed organisms but also for coming generations of progeny, are substantial and affect most organisms. At the time when this thesis work started, no information on the effects of developmental exposure on non-reproductive behavior was available in fish. The aim of this thesis was to study the existence, extent and heritability of such effects.

KRISTINA VOLKOVA Fishy Behavior 21
Experimental animals The aquatic environment is a primary recipient of a large number of EDCs. Sewage effluents have been identified as a major source of these chemicals. Waste waters from homes and industries contain natural and synthetic hormones, pharmaceuticals, pesticides, industrial chemicals, metals, and additives in personal care products. Sampling of effluents and surface waters detected over one hundred pharmaceuticals used by humans in ng/L and μg/L concentrations (Monteiro and Boxall 2010). Consequently, aquatic animals are exposed to complex mixtures of chemicals that can induce harmful effects and potentially impact populations. Despite the vulnerability of aquatic organisms, knowledge about the effects of EDCs on this group of animals is still lacking. In this thesis we examined effects of the estrogenic EDC 17α-ethinylestradiol on two different fish species.
Zebrafish (Danio rerio) The zebrafish is named after its five horizontal stripes on the side of the body, all extending onto the end of caudal fin rays. It is a freshwater fish that can be found in the tropical waters (18°C – 24°C) of Pakistan, India, Bangladesh, Myanmar and Nepal (Menon 1999). It inhabits streams, canals, ditches and ponds (Rahman 1989) where it feeds on worms and small crustaceans (Mills et al. 1989). Zebrafish can grow to 6.4 cm in length but in captivity they seldom grow larger than 4 cm (Spence et al. 2008). The lifespan of a zebrafish is around two to three years, although in ideal conditions they can live up to five years (Spence et al. 2008). The generation time for the zebrafish is approximately three to four months. In captivity, they can spawn all year around at intervals of two to three days, laying hundreds of eggs at each occasion (Spence et al. 2008). Zebrafish is a shoaling species, known to establish dominance hierarchies. Dominance has been demonstrated both during mating and foraging, and does not appear to be determined by sex. The relationship between dominance and body size remains unclear. Zebrafish generally prefer larger shoals but variation may occur with the activity level of the shoal and sex of the individual (Spence et al. 2008). Zebrafish are frequently used as a model organism in biological and medical research. Fish in general are particularly suitable for pharmacological studies as invasive treatment can be avoided by adding the compound of

22 KRISTINA VOLKOVA Fishy Behavior
interest directly to the water. The small size of zebrafish makes them relatively easy to house and allows for simpler handling compared to larger fish species. Their fast spawning time and the ability to frequently produce a large number of embryos make this species ideal for studying embryonic development. In addition, the zebrafish genome is fully sequenced, giving this species an advantage over many other fish species in terms of potential use in molecular and genetic analysis. However, while many orthologue genes have been shown to have conserved functions in fish, the genome duplication event occurring around 350 million years ago in teleost fish has given rise to paralogue genes that may have acquired novel, specialized functions (Toloza-Villalobos et al. 2015). In this thesis, the zebrafish AB strain was chosen for experiments in papers III and IV. The AB strain is an established laboratory strain commonly used in molecular analysis, and has the most studied behavioral repertoire of all zebrafish strains.
Guppy (Poecilia reticulata) The guppy is a freshwater fish that lives in the tropical waters (18°C - 28°C)(Riehl et al. 1991) of South America, mainly in Venezuela, Barbados, Trinidad, northern Brazil and the Guyanas. It has also been widely introduced to other parts of South America (Kottelat et al. 1996) and feral populations can be found in Africa (Skelton 1993). The guppy female reaches about 5 cm in length and the male about half that size (Riehl et al. 1991). Females mature at the age of 3 months and males at 2 months (Kottelat et al. 1996). The guppy is a live-bearing fish. The gestation period lasts four to six weeks, after which females give birth to 20-40 live young. The female may produce young every four weeks (Skelton 1993) and can store sperm from the male for later fertilization.

KRISTINA VOLKOVA Fishy Behavior 23
The guppy inhabits warm springs, ditches and canals (Page et al. 1991). It can live in a wide variety of habitats with dense vegetation and feeds on zooplankton, small insects and detritus (Allen 1991; Kottelat et al. 2007). Guppies will at times forage in groups in order to find food more easily. While shoaling behavior enables the individual guppy to spend less energy on antipredator behavior and spend more time feeding, by doing so the food needs to be shared with other members of the group (Magurran and Seghers 1991). Studies have shown that guppies tend to shoal in high-predation regions, but not in low-predation regions. When guppies with a high tendency to shoal were isolated from high-predation regions and relocated to predator-free environments, over time, they decreased their shoaling behavior (Magurran et al. 1992).
The guppy is becoming an increasingly popular experimental animal in biological and ecological research due to its well characterized male courtship behavior. In EDC research, this presents a great advantage as many EDCs have been found to affect reproduction and reproductive behavior.
In this thesis, the guppy was chosen as a complementary experimental animal to zebrafish because it is less inbred than the established laboratory zebrafish strains, making it more comparable to wild fish populations. Also, in contrast to zebrafish, guppies are live-bearing fish, a reproductive strategy that could potentially affect their sensitivity to developmental EDC exposure. Unfortunately, the guppy genome was not fully sequenced at the time when the experiments in this thesis were conducted, limiting the possibilities of performing molecular and genetic analysis in this species.

24 KRISTINA VOLKOVA Fishy Behavior
Methods The methods are described in detail in each paper. However, in order to give an overview on the studies performed in this thesis, general background and descriptions of the methods follow below.
Exposure In order to investigate persistent effects of EE2 in several consecutive generations, we used low EE2 concentrations and the experimental set-up of the studies presented in this thesis was designed to allow developmentally exposed fish to recover from the reversible effects of EE2. In the guppy experiment (Paper I), gravid female guppies were exposed to EE2 in a flow-through system until birth of their offspring. After birth, the developmentally exposed fry were immediately transferred to clean water, where they were raised under normal maintenance conditions until adulthood. All siblings were kept in the same aquarium until sexual maturation, at which point the males were separated from the females. In zebrafish experiments (Paper III and IV), fertilized zebrafish embryos were exposed to EE2 from 1 day until 80 days post fertilization, after which time the zebrafish were allowed to recover for 82 (Paper III) or 120 days (Paper IV) in clean water under normal maintenance conditions. The ability of zebrafish females to lay a large number of eggs made it possible to divide the fertilized eggs from each parental pair into three treatment groups (Fig 1). Eight parental pairs were used in manuscript III, and ten parental pairs were used in manuscript IV. In manuscript III, zebrafish embryos were exposed to EE2 through a semi-static system during the first 6 weeks of development. At week 6, zebrafish larvae were transferred to a flow-through system where exposure was maintained until the fish were 80 days old. In manuscript IV, zebrafish larvae were exposed to EE2 in a flow-through system from 1 day post fertilization until 80 days post fertilization. All families were kept in separate 2 L tanks throughout all experiments. Males were separated from females based on secondary sexual characteristics starting after 4 weeks of recovery. Due to the ability of EE2 to delay gonad development, the sex of all fish was continuously re-evaluated until the start of behavior studies.

KRISTINA VOLKOVA Fishy Behavior 25
Fig. 1: Experimental set-up of zebrafish exposure experiments in paper III and IV.
Behavior In the papers presented in this thesis we mainly studied the effects of EE2 on anxiety, stress and shoaling behavior. These parameters are of high ecological significance in wild fish populations, likely affecting fitness by influencing foraging, reactions to predators and opportunities to reproduce. We also investigated the effects of developmental EE2 exposure on the reproductive behavior of the guppy males.
Novel tank test The novel tank test is an adaptation of the rodent open-field test and is based on the observation that many fish species, including zebrafish and guppies, tend to dive to the bottom of the tank and stay near the bottom, when first introduced to a new environment (Egan et al. 2009; Maximino et al. 2010a). The fish, particularly the guppies, often freeze (lay immobilized) on the bottom for one or several episodes. In the first minute after the introduction to a novel environment, many fish tend to spend 75-80% of their time in the bottom half, with a steady decrease in bottom-dwelling throughout the duration of the test. By the end of a 5 or 10 minute

26 KRISTINA VOLKOVA Fishy Behavior
test, the bottom-dwelling is generally reduced to chance levels. Increased bottom-dwelling in the novel tank test is considered an indicator of higher stress and has been associated with increased cortisol levels (Egan et al. 2009; Espmark et al. 2008; Ghisleni et al. 2012). The novel tank test is a well-established test, and has a robust response to anxiogenic and anxiolytic drugs (Bencan et al. 2009; Cachat et al. 2011; Egan et al. 2009; Levin et al. 2007; Maximino et al. 2010a; Sackerman et al. 2010; Stewart et al. 2011). However, novelty appears to be the main driving force in bottom-dwelling. Fish housed in tanks that are similar to the test apparatus have been shown to present different bottom-dwelling profiles compared to fish housed in dissimilar tanks (Bencan et al. 2009; Blaser and Rosemberg 2012). Also, some differences exist in the behavioral response between different zebrafish strains (Maximino et al. 2010a). According to our observations, the color of the tank bottom and sides, and the basal stress-level of the fish when it is introduced to the tank, plays a crucial role in the novel tank test. For example, a white-colored bottom of the test tank completely inhibited the diving behavior of the zebrafish. Also, less gentle netting could increase the basal stress-level of the fish and induce extensive freezing. Both these reactions had a negative impact on the ability to detect differences between behavioral phenotypes in the test. Also, fish that were raised in tanks on transparent shelves (giving the illusion of a deeper tank), were not as reactive to the novelty of the test and displayed a less pronounced diving behavior. This supports that environmental factors are extremely important, as the novelty stimulus is conditional, as has previously been suggested (Blaser and Rosemberg 2012).
In this thesis the novel tank test was performed in a 20 x 20 x 40 cm tank filled with 15 L pre-heated pure tap water. At the right end, a transparent Plexiglas screen trapped 5 untreated male or female littermates that would later be used in the shoaling test. A black plastic sheet prevented visual contact between the shoal and the test compartment. The tank had a black bottom and was black on two sides and a horizontal midline divided the tank into top/bottom halves (Fig.2). The novel tank test was initiated by introducing a fish to the test tank by netting. Anxiety levels were assessed by recording latency to first transition across the midline into the upper half of the tank, number of total transitions across the midline into the upper half of the tank, and total time spent in the upper half of the tank. The test

KRISTINA VOLKOVA Fishy Behavior 27
session lasted for 5 minutes, after which the black plastic sheet covering the shoal was removed and the shoaling test was initiated.
Fig. 2: Experimental set-up of the novel tank test seen from the side.
Shoaling test Shoaling is an important social interaction in fish. Both zebrafish and guppies are naturally shoaling species. Increased shoaling is a predator avoidance strategy that leads to a decreased risk for both group and individual in the presence of danger, but also has a negative impact on foraging efficacy (Pitcher and Parrish 1993; Ryer and Olla 1998). Though the mechanisms behind shoaling are not well understood, this behavior is of great ecological importance to many fish species living in the wild. A decreased tendency to leave the shoal is regarded as a measure of reduced boldness (Moretz et al. 2007), which is a behavior connected to stress sensitivity in fish. Exposure to alarm pheromones (a substance secreted from the skin of a wounded fish, to warn others of danger) has been shown to increase shoal cohesion in zebrafish (Rehnberg and Smith 1988). Several different methods have been used to investigate shoaling tendency (Maximino et al. 2010a). Shoaling intensity has been shown to both increase and decrease in response to EDC exposure (Å Espmark Wibe et al. 2002; Reyhanian et al. 2011; Ward et al. 2008; Xia et al. 2010), indicating that several components are involved in regulating this behavior. Unfortunately, the shoaling test lacks the robust pharmacological validation
28 KRISTINA VOLKOVA Fishy Behavior
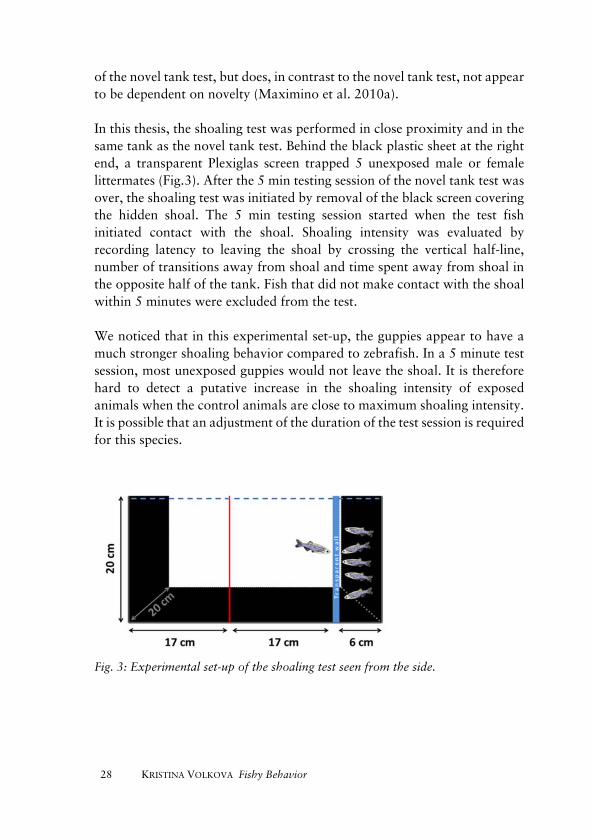
of the novel tank test, but does, in contrast to the novel tank test, not appear to be dependent on novelty (Maximino et al. 2010a). In this thesis, the shoaling test was performed in close proximity and in the same tank as the novel tank test. Behind the black plastic sheet at the right end, a transparent Plexiglas screen trapped 5 unexposed male or female littermates (Fig.3). After the 5 min testing session of the novel tank test was over, the shoaling test was initiated by removal of the black screen covering the hidden shoal. The 5 min testing session started when the test fish initiated contact with the shoal. Shoaling intensity was evaluated by recording latency to leaving the shoal by crossing the vertical half-line, number of transitions away from shoal and time spent away from shoal in the opposite half of the tank. Fish that did not make contact with the shoal within 5 minutes were excluded from the test.
We noticed that in this experimental set-up, the guppies appear to have a much stronger shoaling behavior compared to zebrafish. In a 5 minute test session, most unexposed guppies would not leave the shoal. It is therefore hard to detect a putative increase in the shoaling intensity of exposed animals when the control animals are close to maximum shoaling intensity. It is possible that an adjustment of the duration of the test session is required for this species.
Fig. 3: Experimental set-up of the shoaling test seen from the side.

KRISTINA VOLKOVA Fishy Behavior 29
Scototaxis test Many animals, including fish, show a marked preference for dark environments. The scototaxis test is based on the observation that animals usually spend the majority of the initial two-thirds of a test in the dark compartment before exploring the white area (Maximino et al. 2010a; Maximino et al. 2010b). In the scototaxis test, an increased dwelling in the dark compartment signals increased anxiety and stress. The scototaxis test has been shown to be similar to the novel tank test in detecting stress behavior (Blaser and Rosemberg 2012). However, while novelty appears to be the main controlling factor in the novel tank test, no such dependencies have been detected in the scototaxis test, as fish do not appear to be habituated to the test after repeated sessions, as in novel tank test. This could possibly make the scototaxis test a more robust test for anxiety. Unfortunately, the somewhat lacking pharmacological validation is still the main disadvantage of this test (Maximino et al. 2010a). In this thesis, the scototaxis test was performed in a 20 × 20 × 40 cm aquarium filled with pre-heated tap water up to a 10 cm level. The test tank was divided into one black and one white half. All sides of the tank were covered with plastic in the corresponding color of that half (Fig.4). The tank had two transparent central sliding doors, creating a compartment of 5 × 20 cm. No lid was used in this test as the behavior was filmed from above. The test fish was introduced into the central compartment, and after a 5 min habituation period the sliding doors were raised and the anxiety level was evaluated during 5 min as latency to first entrance into the white or black zone, number of transitions to white or black zone and total time spent in white or black zone.

30 KRISTINA VOLKOVA Fishy Behavior
Fig. 4: Experimental set-up of the scototaxis test seen from the side.
Guppy reproductive behavior The female guppy is receptive to male courtship for only 3-5 days of her ovulatory cycle, during which time she may copulate with several males (Evans et al. 2002; Liley 1972; Venkatesh et al. 1990). The female chooses her partners based on body coloring and courtship behavior. The guppy male may perform an active courtship behavior referred to as Sigmoid Display, or a sneaky mating attempt called Gonopodium Thrust, depending on the receptivity of the female. A sexually active guppy male may also perform a behavior referred to as the Gonopodium Swinging, without courting or sneaking up on a female. All the above mentioned courting behaviors of the guppy males have been studied in paper I. The behaviors were quantified during an observation period of 15 minutes, after an experimental male was introduced to a non-experimental female. The Sigmoid Display is performed as a "dance" in front of the female. In this behavior, the male arches his body into an S or C- shape while quivering intensely for up to a couple of seconds (Liley 1972). The male will either keep his unpaired fins spread (open display) or keep them folded close to the body (closed display). If the female accepts the courtship she will glide towards the male while arching her body into a C-shape, exposing her genital pore. If the male proceeds with inserting his gonopodium into the female he will make several intense leaping movements (Liley 1972).

KRISTINA VOLKOVA Fishy Behavior 31
The Gonopodium Thrust is a sneaky mating tactic performed when a male encounters an unreceptive female. With his gonopodium swung forward, the male will sneak up on the female from beneath, aiming to insert it into her genital pore in a thrusting movement. Gonopodium Swinging, where a guppy male will swing his gonopodium forward, can be performed by a male at almost any given moment, without him courting a female or engaging in sneaky mating. This behavior is thought to represent high sexual motivation.
Fertility
Guppy testis histology In paper I, the guppy testis was examined for histological changes following developmental exposure to EE2 and recovery in clean water. A normal testis should contain cysts of all spermatogenisis stages including spermatogonia, spermatocytes, spermatids, spermatozoa and Sertoli cells. The analysis aimed to identify alterations in testes morphology due to the exposure. Histology was performed by removing the testicles from guppy males developmentally exposed to EE2 after recovery in clean water. The testicles were fixed and dehydrated, embedded in Paraplast and sectioned (7 μm). The sections were mounted on poly-L-lysine coated slides and stained with Haematoxylin/Eosin. All sections were manually examined for histological changes under a light microscope.
Zebrafish fertilization success In order to assess if zebrafish developmentally exposed to EE2 had an impaired ability to reproduce, we collected data on fertilization success in paper III from the first generation (F0), after EE2 exposure and 82 days of recovery in clean water. Males and females from each treatment group were mated to an equal number of unexposed zebrafish of the opposite sex. The fish were put together in mating cages for 24 hours and the total number of eggs laid, number of fertilized eggs, hatching and survival of larvae after 6 days were recorded.

32 KRISTINA VOLKOVA Fishy Behavior
Gene expression
RNA sequencing RNA sequencing (RNA seq) is a relatively new technology used to reveal the quantity and presence of RNA in a sample at a given moment in time. By detecting all RNA present, RNA sequencing is not dependent on previous knowledge of the genome, nor does it require the selection of pre-determined genes of interest, giving it an advantage over older methods such as microarrays. In Paper IV, RNA seq analysis was performed in order to identify differentially expressed genes in zebrafish developmentally exposed to EE2, in an unbiased manner. Based on the results in the novel tank test, samples from the exposure resulting in the highest impact on anxiety were used. Hence RNA seq was performed on brains from male fish exposed to 3 ng/L and female fish exposed to 10 ng/L EE2. Whole-brain RNA was used in three biological replicates for each treatment group and sex. High-performance RNA seq was performed at Genome Infrastructure, SciLifeLab/Uppsala Genome Center according to current procedures of purification, quantification and sequencing. 40 million reads were used in the study, to increase detection capacity and discriminatory power. All bioinformatics was performed by BILS (Bioinformatics Infrastructure for Life Sciences). In short, after quality assessment reads were mapped to the zebrafish (Danio rerio) genome. The mapped reads were converted to count data, using the Ensembl annotation of the zebrafish genome sequence. Statistical modeling of gene expression was done with the library edgeR (Robinson et al. 2010; McCarthy et al. 2012) in R (R core team 2015) following the workflow suggested by the authors. All genes having expression levels lower than 1 cpm mapped reads in at least 3 different libraries were omitted from the data set. A false discovery rate (FDR) was used for identification of significantly differentially expressed genes. Genes with adjusted p<0.05 were considered as differentially expressed. A manual classification of genes and prediction of biological gene function was performed, with orthologue search in Ensembl. GO-terms were found in Zfin (for zebrafish), Entrez gene (for human and rodent orthologues) and NGNC (human orthologues). Available automatic classification systems

KRISTINA VOLKOVA Fishy Behavior 33
were not used, as we found them to have low mapping capacity in this experiment.
Due to high cost of the RNA sequencing, we could only use three samples per exposure and sex. We are well aware that the number of biological replicates are few, and this implies that great caution is needed in the interpretation of the data. It should also be better to dissect the brains into sub-divisions, as region-specific alterations cannot be discerned when using whole-brain RNA samples. We regard this study as only an initial survey, needing a lot of further confirmation.
Quantitative real-time PCR While polymerase chain reaction (PCR) is mainly used for qualitative analysis of DNA, quantitative polymerase chain reaction (qPCR) is used for quantitative analysis of mRNA gene expression. qPCR is based on the same technique as PCR with the addition of a fluorescent dye (SYBR green in paper III and IV) that binds to the newly formed double stranded DNA that is synthesized with every cycle. The amount of bound dye is measured in every cycle and is directly proportional to the amount of synthesized DNA. In Paper III, qPCR was used to study the expression of vitellogenin (VTG). The female-specific, egg yolk precursor protein VTG is normally synthesized in the female liver but not expressed in the male liver. Several studies have shown that estrogenic EDC exposure causes this gene to be expressed also in the livers of male fish. VTG is a good biomarker for ongoing EDC exposure, although it has been shown to be restored to normal levels after discontinued exposure (Baumann et al. 2014). In Paper III, the livers of 8 control males, 10 males exposed to 1.2 ng/L and 10 males exposed to 1.6 ng/L EE2, were separately analyzed with qPCR.
In Paper IV, qPCR was used to verify the findings of the RNA sequencing analysis. Four genes that were previously identified by RNA seq to be differentially expressed in the brains of zebrafish males developmentally exposed to EE2 compared to unexposed controls, were chosen and specific primers designed with Primer-BLAST. qPCR was run on brains from 8 control males and 8 males developmentally exposed to 3 ng/L EE2.

34 KRISTINA VOLKOVA Fishy Behavior
Brain aromatase activity Brain aromatase (CYP 19b) is expressed in the gonads and brain, and similar to VTG, is a common marker for ongoing exposure to estrogenic compounds. It encodes an enzyme involved in the conversion of androgens to estrogens. Higher aromatase activity is observed in females and estrogen-exposed males than in unexposed males. In Paper I we analyze brain aromatase activity in individual brains from 5, 10 and 9 males from exposure group 0, 2 and 20 ng/L EE2, respectively, using tritiated androstenedione as substrate.
Statistical analysis of behavioral tests Performing manipulative experiments to study fish behavior almost exclusively call for statistical models that can handle dependent data, as an inevitable consequence of maternal effects and the use of aquariums for rearing fish at the lab. The experimental design for the three behavioral tests: novel tank, shoaling and scototaxis tests included in this thesis have all been analyzed using mixed effects models in R 3.01. (R Core Team. 2013) and package lme4 (Bates et al 2014). The mixed effects models in our experiments use family as a random variable to cancel out maternal effects and avoid pseudo-replication due to dependences within families. In the behavioral tests we used some response variables with normally distributed and homoscedastic residuals, like several measures of time. If needed we used log transformations to normalize, and to remediate heteroscedasticity. All time variables could therefore be analyzed using Gaussian error distributions. Response variables that were measured in counts, like number of times leaving the shoal or a compartment, were analyzed assuming Poisson distribution of residuals. The estimates from all mixed effects models were tested with both Wald chi-square and with likelihood ratio tests. In no case could we find any difference in interpretation using the two significance tests for our data. All experiments that included both male and female fish were always analyzed with a full model including the interaction between sex and treatment. In the case of significant interactions we either made separate analyses for each sex or used pairwise contrasts to analyze treatment effects within sex, or sex differences within treatment group. In the first case we used Tukey’s multiple comparison where necessary. In all pairwise contrasts family-wise error rates were controlled with Holm’s method.

KRISTINA VOLKOVA Fishy Behavior 35
Results: Summary of papers
I. Brain circuit imprints of developmental 17α-Ethinylestradiol exposure in guppies (Poecilia reticulata): persistent effects on anxiety but not on reproductive behaviour. To investigate if developmental exposure to the synthetic estrogen EE2 could cause long-lasting, persistent effects on non-reproductive behavior, we exposed gravid guppy females to 0, 2 and 20 ng/L EE2, thus indirectly exposing their developing offspring. After birth, the developmentally exposed fry were transferred to clean water and allowed to recover until adulthood, without further exposure. At 6 month of age, developmentally exposed and unexposed male and female guppies were tested for anxiety and shoaling intensity. The novel tank test, assessing stress behavior detected increased anxiety in male and female guppies developmentally exposed to 20 ng/L EE2. No effect of developmental EE2 exposure was found on shoaling intensity. We also found no effect on male sexual behavior, brain aromatase activity or testis morphology, after 6 months of remediation in clean water.
II. Transgenerational effects of 17α-Ethinylestradiol on anxiety behavior in the guppy, Poecilia reticula. This study aimed to investigate whether the persistent effects on anxiety of developmental exposure to EE2 observed in paper I could be transferred to consecutive generations without further exposure. By mating the guppies developmentally exposed to 20 ng/L EE2, we produced a first generation (F1) that had only been indirectly exposed as germ cells within their parents. Stress behavior of this generation was assessed in the novel tank test, revealing increased anxiety in males but not in females. In order to investigate if the effects of EE2 on anxiety can be transgenerationally transmitted, the F1 guppies were mated, giving rise to a second generation (F2) that had not been exposed to EE2 in any other way except through their grandparents. Their stress behavior was assessed using the novel tank test and scototaxis test, revealing increased anxiety in both male and female guppies. These results show that effects of developmental EE2 exposure on anxiety are not only persistent but also transgenerationally transmitted.

36 KRISTINA VOLKOVA Fishy Behavior
III. Developmental exposure of zebrafish (Danio rerio) to 17α-ethinylestradiol affects non-reproductive behavior and fertility as adults, and increases anxiety in unexposed progeny. The aim of this study was to assess the generality of persistent effects of developmental EE2 exposure on non-reproductive behavior by exposing another fish species. Zebrafish larvae were exposed from 1 day post fertilization (dpf) until 80 dpf to measured concentrations of 0, 1.2 or 1.6 ng/L EE2. After discontinued exposure, the fish were allowed to recover in clean water for 80 days after which time fertilization success, stress behavior, shoaling intensity and hepatic VTG expression was analyzed. The results showed increased anxiety and increased shoaling intensity in both male and female zebrafish developmentally exposed to EE2. Fertilization success was decreased in both sexes, at both EE2 exposure levels, when mated with unexposed animals of the opposite sex. No effects on hepatic VTG expression were observed after remediation in clean water. To investigate if the effects on behavior were heritable, fish exposed to 1.2 ng/L were mated in order to produce an F1 generation. The fish of this generation were tested for anxiety in the novel tank test and scototaxis test, showing increased anxiety for both sexes in both tests. However, there was no significant effect on shoaling intensity in the F1 generation.
IV. Persistent effects of developmental exposure to 17α-ethinylestradiol on the zebrafish (Danio rerio) brain transcriptome and stress behavior. The main aim of this study was to to further investigate the possible mechanisms behind the persistent effects of EE2 exposure on non-reproductive behavior. Here we exposed zebrafish larvae to 0, 2.14 or 7.34 ng/L EE2 from 1 day until 80 days post fertilization. The exposed zebrafish were then allowed to recover for 120 days in clean water before behavior testing. In accordance with paper III, the novel tank test and scototaxis test detected increased anxiety in both sexes, while shoaling intensity was increased only in females. To investigate the possible mechanisms for the observed behaviors, whole-brain RNA was isolated and sent for RNA sequencing analysis from both exposed and unexposed fish. The gene expression analysis detected 33 genes in males and 62 in females to be differentially expressed in zebrafish developmentally exposed to EE2 compared to unexposed controls. The functional analysis of the

KRISTINA VOLKOVA Fishy Behavior 37
differentially expressed genes in zebrafish males exposed to 3 ng/L identified genes involved in cholesterol biosynthesis, synaptic function, immune response, DNA-repair and heme biosynthesis and biodegradation. Female zebrafish developmentally exposed to 10 ng/L EE2 showed effects on genes involved in the circadian rhythm, lipid transportation, cytoskeleton and motor proteins, immune response and heme biosynthesis and biodegradation. The top two affected pathways, cholesterol biosynthesis and circadian rhythm, have previously been related to anxiety (Wong et al. 2013; Wong et al. 2015; Rey et al. 2013).

38 KRISTINA VOLKOVA Fishy Behavior
Discussion and conclusions The results presented in this thesis demonstrate that non-reproductive behaviors in fish are highly sensitive to EE2 exposure during development, and that the disruptions caused by early-life exposure do not appear to be reversible after a long recovery in clean water. Exposure to low, environmentally relevant doses of EE2 during development led to increased anxiety as adults in two different fish species, zebrafish and guppy (Paper I, III, IV).
The concept of an event during development being able to cause a disorder observed later in life is not new. The idea was introduced by David Barker already in the 1990’s, when poor nutrition during human fetal development was shown to increase the susceptibility to develop Type 2 diabetes later in life (Hales and Barker 1992). Since then, the hypothesis of fetal origins of adult disease has been supported by numerous studies in both humans and animal models (Calkins and Devaskar 2011). Over the years, this hypothesis has become even more complex as studies emerge showing windows of sensitivity, where timing plays an important role in the outcome (Roseboom et al. 2001). For example, nutritional deprivation of women during the first half of their pregnancy resulted in significantly higher obesity rates in their sons in adulthood, while nutritional deprivation during the last trimester resulted in lower obesity rates (Ravelli et al. 1976).
In relation to EDCs, it has been shown that the timing and duration of exposure can make the difference between an effect being reversible or irreversible (Fenske et al. 2005; Schäfers et al. 2007). We chose to expose fish during a prolonged period in order not to risk missing the window of sensitivity. We were therefore not able to investigate which time point during development is crucial for cementing the observed effects on stress behavior. It cannot be excluded that a shorter and differently timed exposure to EE2 would render reversible effects on non-reproductive behavior.
Our studies indicate that zebrafish are more sensitive to developmental exposure to EE2 than guppies. In guppies, exposure to 2 ng/L EE2 did not significantly affect stress behavior (Paper I), while zebrafish developmentally exposed to 1.2 and 1.6 ng/L showed both increased anxiety and shoaling (Paper III). However, there were important differences

KRISTINA VOLKOVA Fishy Behavior 39
between the guppy and zebrafish experiments. While guppies were exposed through their mothers due to their ovoviviparity, zebrafish, being an egg-laying fish, were subjected to a direct exposure during development. Also, while the zebrafish used in papers III and IV came from the more or less homogenous laboratory strain AB, the guppies used in paper I were captured in the wild and kept by interbreeding for commercial trade before they were purchased by us. The guppies used in papers I and II are therefore less inbred than the zebrafish, leading to larger inter-individual variation. In addition, the recovery time in clean water was slightly different between experiments. While the zebrafish in paper III and IV recovered for 3 and 4 months, respectively, the guppies had a recovery time of 6 months. Any of these differences could account for the seemingly higher sensitivity of zebrafish to developmental EE2 exposure.
Despite some differences between the guppy experiment (Paper I) and zebrafish experiments (papers III, IV), all studies showed that developmental exposure to EE2 resulted in persistent effects on stress behavior, remaining after a long remediation period in clean water. While the mechanisms behind the observed persistent effects on behavior remain unknown, the RNA sequencing analysis performed on the whole-brain transcriptome of developmentally exposed zebrafish (Paper IV) could be a useful starting point for further investigations. It is challenging to interpret transcriptome data, and the study has weaknesses, as described in the Methods section. We can thus only speculate on connections between identified pathways and imprinting of anxiety behavior. This is, however, the first brain transcriptome study of fish acquiring a behavioral phenotype due to an EDC. It is also the first transcriptome study of the brain after a long remediation period following developmental EDC exposure. 94 genes were found to be differentially expressed between the developmentally exposed zebrafish and controls. However, no effects were observed on genes normally associated with the stress axis, like Corticotropin-releasing hormone (CRH), genes in the hypothalamo-pituitary-interrenal axis (HPI) and the monoamine system (Pavlidis et al. 2015). Acute, ongoing, adult EE2 exposure has previously been shown to affect such pathways (Harding et al. 2013; Wit et al. 2010). Our results thus suggest that there may be differences between the effects of acute and developmental EE2 exposure at transcriptional level, and that the anxiety behavior phenotype induced during development is not necessarily

40 KRISTINA VOLKOVA Fishy Behavior
mediated through the most obvious pathways. It is possible that differences in the expression of expected stress-associated genes would have become apparent if fish used in the RNA sequencing analysis were subjected to acute stressors in close proximity to sample extraction, or if the samples were taken before remediation in clean water.
We did however detect alterations in genes that could indirectly affect the stress response. Among those are genes involved in cholesterol biosynthesis, which was identified as the top putative pathway affected in male brains. Effects on cholesterol/lipid biosynthesis and metabolism have previously been detected in several other studies involving EE2 (Cypriani et al. 1988; Flores-Valverde et al. 2010; Hoffmann et al. 2006; Hogan et al. 2010). Furthermore, whole-brain RNA sequencing of Flouxetine exposed adult zebrafish males found cholesterol biosynthesis genes to be down-regulated, concomitant with anxiolytic behavior (Wong et al. 2013). Among these genes were the same four genes (lss, msmo1, hmgcs1 and sqlea) that we found to be upregulated in the brains of male zebrafish developmentally exposed to EE2, concomitant with increased anxiety (Paper IV). The expression of the same genes were also found to be higher in fish with high compared to low stress sensitivity (Wong et al. 2015). These findings might suggest a possible link between cholesterol biosynthesis and stress behavior phenotypes. Locally synthesized cholesterol is the precursor of many neuro-steroids that have been associated with the regulation of stress behavior in mammals (Gunn et al. 2011; Zorumski et al. 2013). Cholesterol biosynthesis in the brain also plays an important role in myelination and axonal function (Björkhem and Meaney 2004), and alterations in this pathway could thereby affect neuronal signaling. This makes cholesterol biosynthesis an important pathway to further explore in relation to EDC exposure. Another functional group found to be affected by developmental EE2 exposure in Paper IV, was circadian rhythm. In females, seven genes were upregulated including: per1a, nr1d1, dbpb and bhlhe41. Circadian rhythm genes have been shown to be responsive to EE2 in female Coho salmon (Harding et al. 2013) and different regulation of circadian rhythm genes has been associated with different personality types in zebrafish (Rey et al. 2013). Estrogen exposure alters the expression of circadian rhythm genes also in rats (Nakamura et al. 2001; Nakamura et al. 2005; Nakamura et al. 2008). In addition, estrogens have been shown to affect period length and

KRISTINA VOLKOVA Fishy Behavior 41
onset of locomotor activity (Krizo and Mintz 2014). Furthermore, in a study examining major depressive disorder (MDD) in humans, the orthologues of per1a, nr1d1, dbpb and bhlhe41 were found to have a disrupted expression pattern in MDD patients compared to healthy controls (Li et al. 2013). The same study also identified these four genes as having among the strongest cyclic patterns in the human brain. The function of these genes in the fish brain remains to be investigated, but the circadian rhythm warrants further studies concerning its influence on mood and behavior. We also identified 6 genes affected by developmental EE2 exposure with recognized functions in the synapse. The encoded proteins are in humans and rodents associated with brain development and synaptic plasticity. Also, the observed changes in genes involved in cholesterol biosynthesis, which appear to play an important role in axon myelination, might affect brain signaling. Thus, EE2 exposure during zebrafish development induces several persistent alterations in the brain that likely affect the outcome of nerve signaling in adult fish and might turn out to be important for the developmental modulations resulting in an anxiety phenotype. In papers II and III we demonstrated that the effects of EE2 on anxiety can be transmitted to untreated offspring, and in paper II we showed that such effects can also be transgenerationally transmitted to two consecutive unexposed generations. Transgenerational effects and alterations in non-reproductive behavior after developmental EDC exposure have been observed in mammals (Skinner et al. 2008; Wolstenholme et al. 2012), but not previously in fish. Studies showing that an event during fetal development can lead to effects in adulthood that also extend beyond the first generation of offspring are beginning to emerge also in humans (Painter et al. 2008).
The number of reports on transgenerational effects in fish after exposure to environmental contaminants have been increasing over the past few years, but are still only four in addition to our study. Such effects have now been demonstrated in medaka and zebrafish on fertility, larval survival, malformations and photomotor response (Baker et al. 2014; Bhandari et al. 2015b; Corrales et al. 2014; Vignet et al. 2015). Paper II is the first example of a transgenerationally transmitted behavioral phenotype in adult fish. The results presented in Paper II also extend previous findings on transgenerational effects to a third fish species, the guppy. In addition, our

42 KRISTINA VOLKOVA Fishy Behavior
results confirm the ability of EE2 to induce transgenerational effects in fish, as has previously been demonstrated on fertility in medaka (Bhandari et al. 2015b).
In mammals, epigenetic changes have been shown to accompany transgenerational effects of EDC exposure in several studies (de Assis et al. 2012; Manikkam et al. 2013; Nilsson et al. 2012; Skinner et al. 2008). Furthermore, epigenetic modifications such as DNA methylation, chromatin structure and non-coding RNAs (ncRNA) including micro-RNAs have previously been linked to phenotypic alterations (Zhang and Ho 2011). While no published study has yet investigated epigenetic transgenerational changes in fish, such studies are essential for understanding the basis of transgenerational effects. Unfortunately, we were not able to examine the epigenome of developmentally exposed fish and their offspring. However, we will be able to address one of these modifications, as changes in ncRNA expression in response to developmental EE2 exposure were observed in the RNA sequencing. The RNA sequencing analysis in paper IV detected, aside from effects on protein-encoding genes, significant differences in ncRNA expression between exposed and unexposed animals (unpublished results). In exposed females, 40% of the significantly altered genes were ncRNA. This is about 10 times more than what is expected by chance (p<0.001, odds ratio 14.55). We are currently investigating the nature of these ncRNAs as well as analyzing the ncRNA expression in males. Such observations could provide valuable information about the mechanisms that have been proposed for persistent and transgenerational effects of EDCs. Taken together, the results in this thesis show that developmental, low-dose EDC exposure leads to irreversible and transgenerationally transmittable effects on stress behavior in fish. Such effects could compromise fitness and survival of not only the exposed fish but also of their offspring, possibly leading to long-term consequences for wild fish populations. Transgenerational effects of environmental stressors should therefore be one of the essential components to be considered in ecological risk assessment. The concentrations of EE2 used in the exposure experiments are well within what has been detected in samples taken outside wastewater treatment plants, showing EE2 levels ranging from almost undetectable (<1 ng/L) to as high as 200-300 ng/L (Hannah et al. 2009; Kolpin et al. 2002; Laurenson et al. 2014; Ternes et al. 1999). While methods for more effective

KRISTINA VOLKOVA Fishy Behavior 43
removal of estrogenic EDCs from wastewater effluent exist (Barber et al. 2012; Baynes et al. 2012; Bringolf and Summerfelt 2003; Liu et al. 2009), they are seldom used due to increased costs. Studies by us and others stress the importance of eliminating environmental EDC contaminants from the aquatic environment. Heritable effects of EDCs on non-reproductive behavior not only have serious implications for the fitness of aquatic organisms, but should also be considered in the etiology of complex neurological disease in mammals and humans. The interaction between genes and environment is a well-studied phenomenon, not exclusive to any one species. Any compound with the ability to induce not only persistent but also heritable effects should therefore be of concern also in regard to fitness of all species. The results presented in this thesis thus support the application of the Precautionary Principle in EDC risk assessment and regulation.

44 KRISTINA VOLKOVA Fishy Behavior
Acknowledgements First and foremost I would like to thank my supervisor Inger Porsch Hällström for letting me be part of this project and for having the patience to let me learn things at my own pace, even when my own patience ran out. I have learnt many things from you Inger, not only about science but also about leadership and teamwork. It is these lessons I will carry with me into whatever awaits me in the future. Nasim Caspillo, my partner in crime from the very beginning. I don't know why other people complain about their postgraduate studies, but I can only assume it's because they don't have a dirty mind like yours to share an office with. You know you got something special when cleaning old fish tanks becomes a "fun day at the office". Tove “Tovson” Porseryd, thank you for your company in all those windowless rooms that are either too cold or too warm. Also, thanks for getting me hooked on coral reefs, my holidays have gotten way more expensive since I met you. Stefan Hallgren, a.k.a "The Doctor". Thank you for all the therapy sessions. Still waiting for my paycheck though. John "Bird man" Boss, the pseudo member of the IPH lab group. I don't know how you got on every group photo we ever took, but you sure deserve to be there! Thank you for all the epic adventures. Josefine "I walk so fast- I make everyone else look bad" Larsson, the other pseudo member of the IPH lab group. Thank you for being my gladiator, both at "Måndagsfysen" and at my presentations. Patrik Dinnétz, the almighty statistics-Jedi. Thank you for explaining difficult things in a way even I can understand. At least for a couple of hours. I bow to thee sensei. Anders "Mr. Gadget" Amelin, what would I have done without you? Whatever the problem, you always have a solution. You're either a genius or a wizard. Probably both. Staff at Södertörn University, unfortunately too many to mention all by name. Thank you for making Södertörn a fun place to work. A special thank you to Tiina Vinter for saving me from low blood sugar on numerous occasions, and to Linn Rabe for making my muscles hurt real bad. Gilbert Lauter, my idol. It's no secret I've always looked up to you. You do science the way it should be done. You will be a fantastic PI one day. Please call me when that happens, I want in.

KRISTINA VOLKOVA Fishy Behavior 45
Gabriele Senti, I will always be grateful to you for taking me under your wings all those years ago. In retrospect I can't believe you let an undergrad touch your stuff! I don't think I wanna know how many experiments I messed up. Don't tell me. But thank you! Mr. Anders Jonsson and Ms. Felicia Dinnétz, the teachers of EN3b, class of 2005. Thank you for being two of the greatest teachers I have ever known. You taught me chemistry and biology so well that my first year in college felt like a recap of grade school natural sciences. It was like counting pine cones and making butter all over again. It’s safe to say, you are a big reason I made it this far. My friends, even though none of you ever understood what I was working on, you always had my back. It never mattered to you if I got a PhD or not, I was always a scientist in your eyes. A special thank you to Lovisa Lindstrand, my oldest friend. I would not have been who I am today without you. I choose to believe that’s a good thing ;). My family, who probably still don't know what I actually do. Thank you for asking "How are the fish?" every time you called, and even when my answer would be "They are no longer with us ", you never gave up on me. A special thank you to Daniel Hellström, my fantastic fiancé. Thank you for putting up with me during these last few years. Despite all the evenings, weekends and holidays I missed you always supported me in what I had to do.

46 KRISTINA VOLKOVA Fishy Behavior
References
Allen, G.R., 1991. Field guide to the freshwater fishes of New Guinea. Publication, no. 9. 268 p. Christensen Research Institute, Madang, Papua New Guinea.
Anway MD, Cupp AS, Uzumcu M, Skinner MK. 2005. Epigenetic transgenerational actions of endocrine disruptors and male fertility. Sci 308:1466-1469.
Aris AZ, Shamsuddin AS, Praveena SM. 2014. Occurrence of 17α-ethynylestradiol (ee2) in the environment and effect on exposed biota: A review. Environ Int 69:104-119.
Baker TR, Peterson RE, Heideman W. 2014. Using zebrafish as a model system for studying the transgenerational effects of dioxin. Toxicol Sci 138:403-411.
Barber LB, Vajda AM, Douville C, Norris DO, Writer JH. 2012. Fish endocrine disruption responses to a major wastewater treatment facility upgrade. Environmental Science & Technology 46:2121-2131.
Bates D, Maechler M, Bolker B and Walker S. 2014.lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1-5. http://CRAN.R-project.org/package=lme4
Baumann L, Knörr S, Keiter S, Rehberger K, Volz S, Schiller V, et al. 2014. Reversibility of endocrine disruption in zebrafish (danio rerio) after discontinued exposure to the estrogen 17α-ethinylestradiol. Toxicology and Applied Pharmacology 278:230-237.
Bayley M, Nielsen JR, Baatrup E. 1999. Guppy sexual behavior as an effect biomarker of estrogen mimics. Ecotoxicol Environ Saf 43:68-73.
Baynes A, Green C, Nicol E, Beresford N, Kanda R, Henshaw A, et al. 2012. Additional treatment of wastewater reduces endocrine disruption in wild fish—a comparative study of tertiary and advanced treatments. Environmental Science & Technology 46:5565-5573.
Bell AM. 2004. An endocrine disrupter increases growth and risky behavior in threespined stickleback (gasterosteus aculeatus). Horm Behav 45:108-114.

KRISTINA VOLKOVA Fishy Behavior 47
Bencan Z, Sledge D, Levin ED. 2009. Buspirone, chlordiazepoxide and diazepam effects in a zebrafish model of anxiety. Pharmacol Biochem Behav 94:75-80.
Beronius A, Rudén C, Håkansson H, Hanberg A. 2010. Risk to all or none?: A comparative analysis of controversies in the health risk assessment of bisphenol a. Reprod Toxicol 29:132-146.
Bhandari RK, Deem SL, Holliday DK, Jandegian CM, Kassotis CD, Nagel SC, et al. 2015a. Effects of the environmental estrogenic contaminants bisphenol a and 17α-ethinyl estradiol on sexual development and adult behaviors in aquatic wildlife species. Gen Comp Endocrinol 214:195-219.
Bhandari RK, vom Saal FS, Tillitt DE. 2015b. Transgenerational effects from early developmental exposures to bisphenol a or 17[agr]-ethinylestradiol in medaka, oryzias latipes. Sci Rep 5.
Bjerselius R, Lundstedt-Enkel K, Olsen H, Mayer I, Dimberg K. 2001. Male goldfish reproductive behaviour and physiology are severely affected by exogenous exposure to 17β-estradiol. Aquat Toxicol 53:139-152.
Björkhem I, Meaney S. 2004. Brain cholesterol: Long secret life behind a barrier. Arteriosclerosis, Thrombosis, and Vascular Biology 24:806-815.
Blanchfield PJ, Kidd KA, Docker MF, Palace VP, Park BJ, Postma LD. 2015. Recovery of a wild fish population from whole-lake additions of a synthetic estrogen. Environmental Science & Technology 49:3136-3144.
Blaser RE, Rosemberg DB. 2012. Measures of anxiety in zebrafish (<italic>danio rerio</italic>): Dissociation of black/white preference and novel tank test. PLoS ONE 7:e36931.
Bringolf RB, Summerfelt RC. 2003. Reduction of estrogenic activity of municipal wastewater by aerated lagoon treatment facilities. Environ Toxicol Chem 22:77-83.
Cachat J, Stewart A, Grossman L, Gaikwad S, Kadri F, Chung KM, et al. 2011. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat Protoc 5:1786-1799.

48 KRISTINA VOLKOVA Fishy Behavior
Caldwell DJ, Mastrocco F, Anderson PD, Länge R, Sumpter JP. 2012. Predicted-no-effect concentrations for the steroid estrogens estrone, 17β-estradiol, estriol, and 17α-ethinylestradiol. Environ Toxicol Chem 31:1396-1406.
Calkins K, Devaskar SU. 2011. Fetal origins of adult disease. Current Problems in Pediatric and Adolescent Health Care 41:158-176.
Chung Y-W, Nunez AA, Clemens LG. 2001. Effects of neonatal polychlorinated biphenyl exposure on female sexual behavior. Physiol Behav 74:363-370.
Colman JR, Baldwin D, Johnson LL, Scholz NL. 2009. Effects of the synthetic estrogen, 17α-ethinylestradiol, on aggression and courtship behavior in male zebrafish (danio rerio). Aquat Toxicol 91:346-354.
Corrales J, Thornton C, White M, Willett KL. 2014. Multigenerational effects of benzo[a]pyrene exposure on survival and developmental deformities in zebrafish larvae. Aquat Toxicol 148:16-26.
Cypriani B, Tabacik C, Descomps B. 1988. Effect of estradiol and antiestrogens on cholesterol biosynthesis in hormone-dependent and -independent breast cancer cell lines. Biochimica et Biophysica Acta (BBA) - Bioenergetics 972:167-178.
de Assis S, Warri A, Cruz MI, Laja O, Tian Y, Zhang B, et al. 2012. High-fat or ethinyl-oestradiol intake during pregnancy increases mammary cancer risk in several generations of offspring. Nat Commun 3:1053.
Delclos KB, Weis CC, Bucci TJ, Olson G, Mellick P, Sadovova N, et al. 2009. Overlapping but distinct effects of genistein and ethinyl estradiol (ee2) in female sprague–dawley rats in multigenerational reproductive and chronic toxicity studies. Reprod Toxicol 27:117-132.
Della Seta D, Minder I, Belloni V, Aloisi AM, Dessì-Fulgheri F, Farabollini F. 2006. Pubertal exposure to estrogenic chemicals affects behavior in juvenile and adult male rats. Horm Behav 50:301-307.
Dessì-Fulgheri F, Porrini S, Farabollini F. 2002. Effects of perinatal exposure to bisphenol a on play behavior of female and male juvenile rats. Environ Health Perspect 110:403-407.

KRISTINA VOLKOVA Fishy Behavior 49
Dugard ML, Tremblay-Leveau H, Mellier D, Caston J. 2001. Prenatal exposure to ethinylestradiol elicits behavioral abnormalities in the rat. Brain Res Dev Brain Res 129:189-199.
Dzieweczynski TL, Campbell BA, Marks JM, Logan B. 2014. Acute exposure to 17α-ethinylestradiol alters boldness behavioral syndrome in female siamese fighting fish. Horm Behav 66:577-584.
Egan RJ, Bergner CL, Hart PC, Cachat JM, Canavello PR, Elegante MF, et al. 2009. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav Brain Res 205:38-44.
Espmark Wibe A, Rosenqvist G, Jenssen BM. 2002. Disruption of male reproductive behavior in threespine stickleback gasterosteus aculeatus exposed to 17β-estradiol. Environ Res 90:136-141.
Espmark Wibe Å, Billing A, Rosenqvist G, Jenssen BM. 2002. Butyl benzyl phthalate affects shoaling behavior and bottom-dwelling behavior in threespine stickleback. Environ Res 89:180.
Espmark ÅM, Eriksen MS, Salte R, Braastad BO, Bakken M. 2008. A note on pre-spawning maternal cortisol exposure in farmed atlantic salmon and its impact on the behaviour of offspring in response to a novel environment. Appl Anim Behav Sci 110:404-409.
Evans JP, Pitcher TE, Magurran AE. 2002. The ontogeny of courtship, colour and sperm production in male guppies. J Fish Biol 60:495-498.
Farabollini F, Porrini S, Dessı ̀ -Fulgheri F. 1999. Perinatal exposure to the estrogenic pollutant bisphenol a affects behavior in male and female rats. Pharmacol Biochem Behav 64:687-694.
Farabollini F, Porrini S, Della Seta D, Bianchi F, Dessì-Fulgheri F. 2002. Effects of perinatal exposure to bisphenol a on sociosexual behavior of female and male rats. Environ Health Perspect 110:409-414.
Fenske M, Maack G, Schäfers C, Segner H. 2005. An environmentally relevant concentration of estrogen induces arrest of male gonad development in zebrafish, danio rerio. Environ Toxicol Chem 24:1088-1098.
Fernandez-Galaz M, Parducz A, Naftolin F, Torres-Aleman I, Garcia-Segura L. 1997. Interaction of gonadal steroids and growth factors in brain sex differentiation Biomed Rev 7:67-74.

50 KRISTINA VOLKOVA Fishy Behavior
Filby AL, Thorpe KL, Maack G, Tyler CR. 2007. Gene expression profiles revealing the mechanisms of anti-androgen- and estrogen-induced feminization in fish. Aquat Toxicol 81:219-231.
Filby AL, Paull GC, Searle F, Ortiz-Zarragoitia M, Tyler CR. 2012. Environmental estrogen-induced alterations of male aggression and dominance hierarchies in fish: A mechanistic analysis. Environmental Science & Technology 46:3472-3479.
Flores-Valverde AM, Horwood J, Hill EM. 2010. Disruption of the steroid metabolome in fish caused by exposure to the environmental estrogen 17α-ethinylestradiol. Environmental Science & Technology 44:3552-3558.
Frye C, Bo E, Calamandrei G, Calzà L, Dessì-Fulgheri F, Fernández M, et al. 2012. Endocrine disrupters: A review of some sources, effects, and mechanisms of actions on behaviour and neuroendocrine systems. J Neuroendocrinol 24:144-159.
Gadd JB, Tremblay LA, Northcott GL. 2010. Steroid estrogens, conjugated estrogens and estrogenic activity in farm dairy shed effluents. Environ Pollut 158:730-736.
Ghisleni G, Capiotti KM, Da Silva RS, Oses JP, Piato ÂL, Soares V, et al. 2012. The role of crh in behavioral responses to acute restraint stress in zebrafish. Progress in Neuro-Psychopharmacology and Biological Psychiatry 36:176-182.
Gioiosa L, Parmigiani S, vom Saal FS, Palanza P. 2013. The effects of bisphenol a on emotional behavior depend upon the timing of exposure, age and gender in mice. Horm Behav 63:598-605.
Gonçalves CR, Cunha RW, Barros DM, Martínez PE. 2010. Effects of prenatal and postnatal exposure to a low dose of bisphenol a on behavior and memory in rats. Environ Toxicol Pharmacol 30:195-201.
Gore AC, Heindel JJ, Zoeller RT. 2006. Endocrine disruption for endocrinologists (and others). Endocrinology 147:s1-s3.
Gore AC. 2008. Developmental programming and endocrine disruptor effects on reproductive neuroendocrine systems. Front Neuroendocrinol 29:358-374.

KRISTINA VOLKOVA Fishy Behavior 51
Guerrero-Bosagna C, Skinner MK. 2012. Environmentally induced epigenetic transgenerational inheritance of phenotype and disease. Mol Cell Endocrinol 354:3-8.
Gunn BG, Brown AR, Lambert JJ, Belelli D. 2011. Neurosteroids and gaba(a) receptor interactions: A focus on stress. Frontiers in Neuroscience 5:131.
Hales CN, Barker DJP. 1992. Type 2 (non-insulin-dependent) diabetes mellitus: The thrifty phenotype hypothesis. Diabetologia 35:595-601.
Hallgren S, Volkova K, Reyhanian N, Olsén H, Porsch Hällström I. 2011. Anxiogenic behaviour induced by 17α-ethynylestradiol in male guppies (poecilia reticulata). Fish Physiol Biochem 37:911-918.
Hannah R, D'Aco VJ, Anderson PD, Buzby ME, Caldwell DJ, Cunningham VL, et al. 2009. Exposure assessment of 17α-ethinylestradiol in surface waters of the united states and europe. Environ Toxicol Chem 28:2725-2732.
Harding LB, Schultz IR, Goetz GW, Luckenbach JA, Young G, Goetz FW, et al. 2013. High-throughput sequencing and pathway analysis reveal alteration of the pituitary transcriptome by 17α-ethynylestradiol (ee2) in female coho salmon, oncorhynchus kisutch. Aquat Toxicol 142–143:146-163.
Henry LA, Witt DM. 2006. Effects of neonatal resveratrol exposure on adult male and female reproductive physiology and behavior. Developmental Neuroscience 28:186-195.
Herbst AL, Ulfelder H, Poskanzer DC. 1971. Adenocarcinoma of the vagina. New England Journal of Medicine 284:878-881.
Hill Jr RL, Janz DM. 2003. Developmental estrogenic exposure in zebrafish (danio rerio): I. Effects on sex ratio and breeding success. Aquat Toxicol 63:417-429.
Ho DH, Burggren WW. 2010. Epigenetics and transgenerational transfer: A physiological perspective. J Exp Biol 213:3-16.
Hoffmann JL, Torontali SP, Thomason RG, Lee DM, Brill JL, Price BB, et al. 2006. Hepatic gene expression profiling using genechips in zebrafish exposed to 17α-ethynylestradiol. Aquat Toxicol 79:233-246.

52 KRISTINA VOLKOVA Fishy Behavior
Hogan NS, Currie S, LeBlanc S, Hewitt LM, MacLatchy DL. 2010. Modulation of steroidogenesis and estrogen signalling in the estuarine killifish (fundulus heteroclitus) exposed to ethinylestradiol. Aquat Toxicol 98:148-156.
Hoover RN, Hyer M, Pfeiffer RM, Adam E, Bond B, Cheville AL, et al. 2011. Adverse health outcomes in women exposed in utero to diethylstilbestrol. New England Journal of Medicine 365:1304-1314.
Ishido M, Masuo Y, Kunimoto M, Oka S, Morita M. 2004. Bisphenol a causes hyperactivity in the rat concomitantly with impairment of tyrosine hydroxylase immunoreactivity. Journal of Neuroscience Research 76:423-433.
Jones BA, Shimell JJ, Watson NV. 2011. Pre- and postnatal bisphenol a treatment results in persistent deficits in the sexual behavior of male rats, but not female rats, in adulthood. Horm Behav 59:246-251.
Kabir ER, Rahman MS, Rahman I. 2015. A review on endocrine disruptors and their possible impacts on human health. Environ Toxicol Pharmacol 40:241-258.
Kidd KA, Blanchfield PJ, Mills KH, Palace VP, Evans RE, Lazorchak JM, et al. 2007. Collapse of a fish population after exposure to a synthetic estrogen. PNAS 104:8897-8901.
Kleinmanns Julia A, Schubert D. 2014. Polycomb and trithorax group protein-mediated control of stress responses in plants. In: Biological Chemistry, Vol. 395, 1291.
Kodavanti PRS. 2005. Neurotoxicity of persistent organic pollutants: Possible mode(s) of action and further considerations. Nonlinearity in Biology, Toxicology, Medicine 3.
Kolpin DW, Furlong ET, Meyer MTT, E.M., Zaugg SD, Barber LN, Buxton HT. 2002. Pharmaceuticals, hormones and other organic wastewater contaminants in us streams, 1999-2000. Environ Sci Technol 36:1202-1211.
Kottelat, M. and T. Whitten, 1996. Freshwater biodiversity in Asia, with special reference to fish. World Bank Tech. Pap. 343:59 p.
Kottelat, M. and J. Freyhof, 2007. Handbook of European freshwater fishes. Publications Kottelat, Cornol and Freyhof, Berlin. 646 pp.

KRISTINA VOLKOVA Fishy Behavior 53
Krizo JA, Mintz EM. 2014. Sex differences in behavioral circadian rhythms in laboratory rodents. Frontiers in Endocrinology 5:234.
Kuch HM, Ballschmiter K. 2001. Determination of endocrine-disrupting phenolic compounds and estrogens in surface and drinking water by hrgc−(nci)−ms in the picogram per liter range. Environmental Science & Technology 35:3201-3206.
Lagarde F. BC, Belcher SM., Belzunces LP., Emond C., Guerbet M. and Rousselle C. 2015. Non-monotonic dose-response relationships and endocrine disruptors: A qualitative method of assessment. Environmental Health 2015 14:13.
Lang IA, Galloway TS, Scarlett A, et al. 2008. Association of urinary bisphenol a concentration with medical disorders and laboratory abnormalities in adults. Jama 300:1303-1310.
Larcher S, Delbès G, Robaire B, Yargeau V. 2012. Degradation of 17α-ethinylestradiol by ozonation — identification of the by-products and assessment of their estrogenicity and toxicity. Environ Int 39:66-72.
Larsen MG, Bilberg K, Baatrup E. 2009. Reversibility of estrogenic sex changes in zebrafish (danio rerio). Environ Toxicol Chem 28:1783-1785.
Larsen MG, Baatrup E. 2010. Functional behavior and reproduction in androgenic sex reversed zebrafish (danio rerio). Environ Toxicol Chem 29:1828.
Laurenson J, Bloom R, Page S, Sadrieh N. 2014. Ethinyl estradiol and other human pharmaceutical estrogens in the aquatic environment: A review of recent risk assessment data. AAPS J 16:299-310.
Law W-Y, Chen W-H, Song Y-L, Dufour S, Chang C-F. 2001. Differential in vitro suppressive effects of steroids on leukocyte phagocytosis in two teleosts, tilapia and common carp. Gen Comp Endocrinol 121:163-172.
Leranth C, Hajszan T, Szigeti-Buck K, Bober J, MacLusky NJ. 2008. Bisphenol a prevents the synaptogenic response to estradiol in hippocampus and prefrontal cortex of ovariectomized nonhuman primates. Proceedings of the National Academy of Sciences 105:14187-14191.

54 KRISTINA VOLKOVA Fishy Behavior
Levin ED, Bencan Z, Cerutti DT. 2007. Anxiolytic effects of nicotine in zebrafish. Physiol Behav 90:54-58.
Li JZ, Bunney BG, Meng F, Hagenauer MH, Walsh DM, Vawter MP, et al. 2013. Circadian patterns of gene expression in the human brain and disruption in major depressive disorder. Proceedings of the National Academy of Sciences 110:9950-9955.
Liley NR. 1972. Progress in comparative endocrinologythe effects of estrogens and other steroids on the sexual behavior of the female guppy, poecilia reticulata. Gen Comp Endocrinol 3:542-552.
Liu Z-h, Kanjo Y, Mizutani S. 2009. Removal mechanisms for endocrine disrupting compounds (edcs) in wastewater treatment — physical means, biodegradation, and chemical advanced oxidation: A review. Sci Tot Environ 407:731-748.
Maack G, Segner H. 2004. Life-stage-dependent sensitivity of zebrafish (danio rerio) to estrogen exposure. Comp Biochem Physiol C Toxicol Pharmacol 139:47.
Magurran AE, Seghers BH. 1991. Variation in schooling and aggression amongst guppy (poecilia reticulata) populations in trinidad. Behaviour 118:214-234.
Magurran AE, Seghers BH, Carvalho GR, Shaw PW. 1992. Behavioural consequences of an artificial introduction of guppies (poecilia reticulata) in n. Trinidad: Evidence for the evolution of anti-predator behaviour in the wild. Proceedings of the Royal Society of London B: Biological Sciences 248:117-122.
Majewski A, Blanchfield P, Palace V, Wautier K. 2002. Waterborne 17α-ethynylestradiol affects aggressive behaviour of male fathead minnows (pimephales promelas) under artificial spawning conditions. Water Qual Res J Can 37:697 - 710.
Manikkam M, Tracey R, Guerrero-Bosagna C, Skinner MK. 2013. Plastics derived endocrine disruptors (bpa, dehp and dbp) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLoS ONE 8:e55387.
Maximino C, de Brito TM, da Silva Batista AW, Herculano AM, Morato S, Gouveia Jr A. 2010a. Measuring anxiety in zebrafish: A critical review. Behav Brain Res 214:157-171.

KRISTINA VOLKOVA Fishy Behavior 55
Maximino C, Marques de Brito T, Dias CAGdM, Gouveia A, Morato S. 2010b. Scototaxis as anxiety-like behavior in fish. Nat Protocols 5:209-216.
McCarrey JR. 2014. Distinctions between transgenerational and non-transgenerational epimutations. Mol Cell Endocrinol 398:13-23.
McCarthy DJ, Chen Y, Smyth GK (2012) Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Research 40, 4288-4297.
McEwen B. 1987. Steroid hormones and brain development: Some guidelines for understanding actions of pseudohormones and other toxic agents. Environ Health Perspect 74:177-184.
McLachlan JA. 2001. Environmental signaling: What embryos and evolution teach us about endocrine disrupting chemicals. Endocr Rev 22:319-341.
Menon, A.G.K., 1999. Check list - fresh water fishes of India. Rec. Zool. Surv. India, Misc. Publ., Occas. Pap. No. 175, 366 p.
Mills, D. and G. Vevers, 1989. The Tetra encyclopedia of freshwater tropical aquarium fishes. Tetra Press, New Jersey. 208
Monteiro SC, Boxall ABA. 2010. Occurrence and fate of human pharmaceuticals in the environment. In: Reviews of environmental contamination and toxicology, vol 202. NEW YORK:Springer, 53-154.
Moretz JA, Martins EP, Robison BD. 2007. Behavioral syndromes and the evolution of correlated behavior in zebrafish. Behav Ecol 18:556-562.
Nagel SC, vom Saal FS, Thayer KA, Dhar MG, Boechler M, Welshons WV. 1997. Relative binding affinity-serum modified access (rba-sma) assay predicts the relative in vivo bioactivity of the xenoestrogens bisphenol a and octylphenol. Environ Health Perspect 105:70-76.
Nakamura TJ, Shinohara K, Funabashi T, Kimura F. 2001. Effect of estrogen on the expression of cry1 and cry2 mrnas in the suprachiasmatic nucleus of female rats. Neuroscience Research 41:251-255.
Nakamura TJ, Moriya T, Inoue S, Shimazoe T, Watanabe S, Ebihara S, et al. 2005. Estrogen differentially regulates expression of per1 and per2

56 KRISTINA VOLKOVA Fishy Behavior
genes between central and peripheral clocks and between reproductive and nonreproductive tissues in female rats. Journal of Neuroscience Research 82:622-630.
Nakamura TJ, Sellix MT, Menaker M, Block GD. 2008. Estrogen directly modulates circadian rhythms of per2 expression in the uterus. American Journal of Physiology - Endocrinology and Metabolism 295:E1025-E1031.
Nash JP, Kime DE, Van der Ven LTM, Wester PW, Brion Fo, Maack G, et al. 2004. Long-term exposure to environmental concentrations of the pharmaceutical ethynylestradiol causes reproductive failure in fish. Environ Health Perspect 112.
Nilsson E, Larsen G, Manikkam M, Guerrero-Bosagna C, Savenkova MI, Skinner MK. 2012. Environmentally induced epigenetic transgenerational inheritance of ovarian disease. PLoS ONE 7:e36129.
O'Reilly EJ, Mirzaei F, Forman MR, Ascherio A. 2010. Diethylstilbestrol exposure in utero and depression in women. Am J Epidemiol 171:876-882.
Page, L.M. and B.M. Burr, 1991. A field guide to freshwater fishes of North America north of Mexico. Houghton Mifflin Company, Boston. 432 p.
Painter RC, Osmond C, Gluckman P, Hanson M, Phillips DIW, Roseboom TJ. 2008. Transgenerational effects of prenatal exposure to the dutch famine on neonatal adiposity and health in later life. BJOG 115:1243-1249.
Palmer JR, Herbst AL, Noller KL, Boggs DA, Troisi R, Titus-Ernstoff L, et al. 2009. Urogenital abnormalities in men exposed to diethylstilbestrol in utero: A cohort study. Environmental Health 8:1-6.
Patisaul HB, Adewale HB. 2009. Long-term effects of environmental endocrine disruptors on reproductive physiology and behavior. Frontiers in Behavioral Neuroscience 3:10.
Patisaul HB, Sullivan AW, Radford ME, Walker DM, Adewale HB, Winnik B, et al. 2012. Anxiogenic effects of developmental bisphenol a exposure are associated with gene expression changes in the juvenile rat amygdala and mitigated by soy. PLoS ONE 7:e43890.

KRISTINA VOLKOVA Fishy Behavior 57
Pavlidis M, Theodoridi A, Tsalafouta A. 2015. Neuroendocrine regulation of the stress response in adult zebrafish, danio rerio. Progress in neuro-psychopharmacology & biological psychiatry 60:121-131.
Pillard RC, Rosen LR, Meyer-Bahlburg H, Weinrich JD, Feldman JF, Gruen R, et al. 1993. Psychopathology and social functioning in men prenatally exposed to diethylstilbestrol (des). Psychosomatic Medicine 55:485-491.
Pillon D, Cadiou V, Angulo L, Duittoz AH. 2012. Maternal exposure to 17-alpha-ethinylestradiol alters embryonic development of gnrh-1 neurons in mouse. Brain Research 1433:29-37.
Pitcher TJ, Parrish JK. 1993. Functions of shoaling behaviour in teleost fish. In: Behaviour of teleost fish, Part 2 (Pitcher TJ, ed). London:Chapman and Hall, 363-439.
Pottinger TG, Carrick TR, Hughes SE, Balm PHM. 1996. Testosterone, 11-ketotestosterone, and estradiol-17β modify baseline and stress-induced interrenal and corticotropic activity in trout. Gen Comp Endocrinol 104:284-295.
Rahman, A.K.A., 1989. Freshwater fishes of Bangladesh. Zoological Society of Bangladesh. Department of Zoology, University of Dhaka. 364 p.
Ravelli G-P, Stein ZA, Susser MW. 1976. Obesity in young men after famine exposure in utero and early infancy. New England Journal of Medicine 295:349-353.
R Core Team (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/.
R Core Team. 2013. R: A language and environment for statistical computing. R Foundation for Statistical Computing. URL: http://www.R-project.org/.
Rehnberg BG, Smith RJF. 1988. The influence of alarm substance and shoal size on the behaviour of zebra danios, brachydanio rerio (cyprinidae). J Fish Biol 33:155-163.
Rey S, Boltana S, Vargas R, Roher N, MacKenzie S. 2013. Combining animal personalities with transcriptomics resolves individual variation

58 KRISTINA VOLKOVA Fishy Behavior
within a wild-type zebrafish population and identifies underpinning molecular differences in brain function. Molecular Ecology 22:6100-6115.
Reyhanian N, Volkova K, Hallgren S, Bollner T, Olsson P-E, Olsén KH, et al. 2011. 17α-ethynyl estradiol affect anxiety and shoaling behavior in adult zebra fish (danio rerio). Aquat Toxicol 105:41-48.
Riehl, R. and H.A. Baensch, 1991. Aquarien Atlas. Band. 1. Melle: Mergus, Verlag für Natur-und Heimtierkunde, Germany. 992 p.
Robinson MD, McCarthy DJ, Smyth GK (2010) edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139-140.
Roseboom TJ, van der Meulen JHP, Ravelli ACJ, Osmond C, Barker DJP, Bleker OP. 2001. Effects of prenatal exposure to the dutch famine on adult disease in later life: An overview. Mol Cell Endocrinol 185:93-98.
Ryan BC, Vandenbergh JG. 2006. Developmental exposure to environmental estrogens alters anxiety and spatial memory in female mice. Horm Behav 50:85-93.
Ryer C, Olla B. 1998. Shifting the balance between foraging and predator avoidance: The importance of food distribution for a schooling pelagic forager. Environmental Biology of Fishes 52:467-475.
Sackerman J, Donegan J, C. C, Nguyen NN, Lawless K, Long A, et al. 2010. Zebrafish behavior in novel environment: Effects of acute exposure to anxiolytic compounds and choice of danio rerio line. Int J Comp Psychol 23:43-61.
Sager DB. 1983. Effect of postnatal exposure to polychlorinated biphenyls on adult male reproductive function. Environ Res 31:76-94.
Saili KS, Corvi MM, Weber DN, Patel AU, Das SR, Przybyla J, et al. 2012. Neurodevelopmental low-dose bisphenol a exposure leads to early life-stage hyperactivity and learning deficits in adult zebrafish. Toxicology 291:83-92.
Salian S, Doshi T, Vanage G. 2009. Perinatal exposure of rats to bisphenol a affects the fertility of male offspring. Life Sci 85:742-752.

KRISTINA VOLKOVA Fishy Behavior 59
Schäfers C, Teigeler M, Wenzel A, Maack G, Fenske M, Segner H. 2007. Concentration- and time-dependent effects of the synthetic estrogen, 17α-ethinylestradiol, on reproductive capabilities of the zebrafish, danio rerio. J Toxicol Environ Health A 70:768-779.
Seta DD, Minder I, Dessì-Fulgheri F, Farabollini F. 2005. Bisphenol-a exposure during pregnancy and lactation affects maternal behavior in rats. Brain Research Bulletin 65:255-260.
Shenoy K. 2014. Prenatal exposure to low doses of atrazine affects mating behaviors in male guppies. Horm Behav 66:439-448.
Skelton, P.H., 1993. A complete guide to the freshwater fishes of southern Africa. Southern Book Publishers. 388 p.
Skinner MK, Anway MD, Savenkova MI, Gore AC, Crews D. 2008. Transgenerational epigenetic programming of the brain transcriptome and anxiety behavior. PLoS ONE 3:e3745.
Skinner MK, Manikkam M, Guerrero-Bosagna C. 2010. Epigenetic transgenerational actions of environmental factors in disease etiology. Trends in Endocrinology & Metabolism 21:214-222.
Spence R, Gerlach G, Lawrence C, Smith C. 2008. The behaviour and ecology of the zebrafish, danio rerio. Biological Reviews 83:13-34.
Steinberg RM, Juenger TE, Gore AC. 2007. The effects of prenatal pcbs on adult female paced mating reproductive behaviors in rats. Horm Behav 51:364-372.
Stewart A, Wu N, Cachat J, Hart P, Gaikwad S, Wong K, et al. 2011. Pharmacological modulation of anxiety-like phenotypes in adult zebrafish behavioral models. Progress in Neuro-Psychopharmacology and Biological Psychiatry 35:1421-1431.
Sugiura-Ogasawara M, Ozaki Y, Sonta SI, Makino T, Suzumori K. 2005. Exposure to bisphenol a is associated with recurrent miscarriage. Human Reproduction 20:2325-2329.
Söffker M, Stevens JR, Tyler CR. 2012. Comparative breeding and behavioral responses to ethinylestradiol exposure in wild and laboratory maintained zebrafish (danio rerio) populations. Environmental Science & Technology 46:11377-11383.

60 KRISTINA VOLKOVA Fishy Behavior
Takeuchi T, Tsutsumi O, Ikezuki Y, Takai Y, Taketani Y. 2004. Positive relationship between androgen and the endocrine disruptor, bisphenol a, in normal women and women with ovarian dysfunction. Endocrine Journal 51:165-169.
Ternes TA, Stumpf M, Mueller J, Haberer K, Wilken RD, Servos M. 1999. Behavior and occurrence of estrogens in municipal sewage treatment plants — i. Investigations in germany, canada and brazil. Sci Tot Environ 225:81-90.
Thorpe KL, Cummings RI, Hutchinson TH, Scholze M, Brighty G, Sumpter JP, et al. 2003. Relative potencies and combination effects of steroidal estrogens in fish. Environmental Science & Technology 37:1142-1149.
Toloza-Villalobos J, Arroyo J, Opazo J. 2015. The circadian clock of teleost fish: A comparative analysis reveals distinct fates for duplicated genes. J Mol Evol 80:57-64.
Traina ME, Rescia M, Urbani E, Mantovani A, Macrı ̀ C, Ricciardi C, et al. 2003. Long-lasting effects of lindane on mouse spermatogenesis induced by in utero exposure. Reprod Toxicol 17:25-35.
Valkusz Z, Nagyéri G, Radács M, Ocskó T, Hausinger P, László M, et al. 2011. Further analysis of behavioral and endocrine consequences of chronic exposure of male wistar rats to subtoxic doses of endocrine disruptor chlorobenzenes. Physiol Behav 103:421-430.
Van den Belt K, Wester PW, van der Ven LTM, Verheyen R, Witters H. 2002. Effects of ethynylestradiol on the reproductive physiology in zebrafish (danio rerio): Time dependency and reversibility. Environ Toxicol Chem 21:767-775.
Van den Belt K, Verheyen R, Witters H. 2003. Effects of 17α-ethynylestradiol in a partial life-cycle test with zebrafish (danio rerio): Effects on growth, gonads and female reproductive success. Sci Tot Environ 309:127-137.
Vandenberg LN, Maffini MV, Sonnenschein C, Rubin BS, Soto AM. 2009. Bisphenol-a and the great divide: A review of controversies in the field of endocrine disruption. Endocr Rev 30:75-95.

KRISTINA VOLKOVA Fishy Behavior 61
Vandenberg LN, Colborn T, Hayes TB, Heindel JJ, Jacobs DR, Lee D-H, et al. 2012. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr Rev 33:378-455.
Wang XQ, Fang J, Nunez AA, Clemens LG. 2002. Developmental exposure to polychlorinated biphenyls affects sexual behavior of rats. Physiol Behav 75:689-696.
Ward A, Duff A, Horsfall J, Currie S. 2008. Scents and scents-ability: Pollution disrupts chemical social recognition and shoaling in fish. Proc R Soc Lond B Biol Sci 275:101-105.
Weber LP, Hill RL, Jr., Janz DM. 2003. Developmental estrogenic exposure in zebrafish (danio rerio): Ii. Histological evaluation of gametogenesis and organ toxicity. Aquat Toxicol 63:431-446.
Venkatesh B, Tan CH, Lam TJ. 1990. Steroid hormone profile during gestation and parturition of the guppy (poecilia reticulata). Gen Comp Endocrinol 77:476-483.
Versonnen BJ, Roose P, Monteyne EM, Janssen CR. 2004. Estrogenic and toxic effects of methoxychlor on zebrafish (danio rerio). Environ Toxicol Chem 23:2194.
WHO/UNEP. 2013. State of the science of endocrine disrupting chemicals - 2012. ISBN: 978 92 4 150503 1.
Vignet C, Joassard L, Lyphout L, Guionnet T, Goubeau M, Le Menach K, et al. 2015. Exposures of zebrafish through diet to three environmentally relevant mixtures of pahs produce behavioral disruptions in unexposed f1 and f2 descendant. Environ Sci Pollut Res:1-13.
Virtanen HE, Adamsson A. 2012. Cryptorchidism and endocrine disrupting chemicals. Mol Cell Endocrinol 355:208-220.
Wit MD, Keil D, Ven Kvd, Vandamme S, Witters E, Coen WD. 2010. An integrated transcriptomic and proteomic approach characterizing estrogenic and metabolic effects of 17 α-ethinylestradiol in zebrafish (danio rerio). Gen Comp Endocrinol 167:190-201.
Wolstenholme JT, Taylor JA, Shetty SRJ, Edwards M, Connelly JJ, Rissman EF. 2011. Gestational exposure to low dose bisphenol a alters social behavior in juvenile mice. PLoS ONE 6:e25448.

62 KRISTINA VOLKOVA Fishy Behavior
Wolstenholme JT, Edwards M, Shetty SRJ, Gatewood JD, Taylor JA, Rissman EF, et al. 2012. Gestational exposure to bisphenol a produces transgenerational changes in behaviors and gene expression. Endocrinology 153:3828-3838.
Wong RY, Oxendine SE, Godwin J. 2013. Behavioral and neurogenomic transcriptome changes in wild-derived zebrafish with fluoxetine treatment. BMC Genomics 14:348-348.
Wong RY, Lamm MS, Godwin J. 2015. Characterizing the neurotranscriptomic states in alternative stress coping styles. BMC Genomics 16:425.
Vosges M, Braguer J-C, Combarnous Y. 2008. Long-term exposure of male rats to low-dose ethinylestradiol (ee2) in drinking water: Effects on ponderal growth and on litter size of their progeny. Reprod Toxicol 25:161-168.
Vulliet E, Cren-Olivé C. 2011. Screening of pharmaceuticals and hormones at the regional scale, in surface and groundwaters intended to human consumption. Environ Pollut 159:2929-2934.
Xia J, Niu C, Pei X. 2010. Effects of chronic exposure to nonylphenol on locomotor activity and social behavior in zebrafish (danio rerio). J Environ Sci 22:1435-1440.
Xu H, Yang J, Wang YX, Jiang Q, Chen H, Song HY. 2008. Exposure to 17 alpha-ethynylestradiol impairs reproductive functions of both male and female zebrafish (danio rerio). Aquat Toxicol 88:1-8.
Xu X-h, Zhang J, Wang Y-m, Ye Y-p, Luo Q-q. 2010. Perinatal exposure to bisphenol-a impairs learning-memory by concomitant down-regulation of n-methyl-d-aspartate receptors of hippocampus in male offspring mice. Horm Behav 58:326-333.
Zhang X, Ho S-M. 2011. Epigenetics meets endocrinology. Journal of molecular endocrinology 46:R11-R32.
Zorumski CF, Paul SM, Izumi Y, Covey DF, Mennerick S. 2013. Neurosteroids, stress and depression: Potential therapeutic opportunities. Neuroscience & Biobehavioral Reviews 37:109-122.

Södertörn Doctoral Dissertations
1. Jolanta Aidukaite, The Emergence of the Post-Socialist Welfare State: The case of the Baltic States: Estonia, Latvia and Lithuania, 2004
2. Xavier Fraudet, Politique étrangère française en mer Baltique (1871–1914): de l'exclusion à l'affirmation, 2005
3. Piotr Wawrzeniuk, Confessional Civilising in Ukraine: The Bishop Iosyf Shumliansky and the Introduction of Reforms in the Diocese of Lviv 1668–1708, 2005
4. Andrej Kotljarchuk, In the Shadows of Poland and Russia: The Grand Duchy of Lithuania and Sweden in the European Crisis of the mid-17th Century, 2006
5. Håkan Blomqvist, Nation, ras och civilisation i svensk arbetarrörelse före nazismen, 2006
6. Karin S Lindelöf, Om vi nu ska bli som Europa: Könsskapande och normalitet bland unga kvinnor i transitionens Polen, 2006
7. Andrew Stickley. On Interpersonal Violence in Russia in the Present and the Past: A Sociological Study, 2006
8. Arne Ek, Att konstruera en uppslutning kring den enda vägen: Om folk-rörelsers modernisering i skuggan av det Östeuropeiska systemskiftet, 2006
9. Agnes Ers, I mänsklighetens namn: En etnologisk studie av ett svenskt biståndsprojekt i Rumänien, 2006
10. Johnny Rodin, Rethinking Russian Federalism: The Politics of Intergovernmental Relations and Federal Reforms at the Turn of the Millennium, 2006
11. Kristian Petrov, Tillbaka till framtiden: Modernitet, postmodernitet och generationsidentitet i Gorbačevs glasnost´ och perestrojka, 2006
12. Sophie Söderholm Werkö, Patient patients?: Achieving Patient Empowerment through Active Participation, Increased Knowledge and Organisation, 2008
13. Peter Bötker, Leviatan i arkipelagen: Staten, förvaltningen och samhället. Fallet Estland, 2007

14. Matilda Dahl, States under scrutiny: International organizations, transformation and the construction of progress, 2007
15. Margrethe B. Søvik, Support, resistance and pragmatism: An examination of motivation in language policy in Kharkiv, Ukraine, 2007
16. Yulia Gradskova, Soviet People with female Bodies: Performing beauty and maternity in Soviet Russia in the mid 1930–1960s, 2007
17. Renata Ingbrant, From Her Point of View: Woman's Anti-World in the Poetry of Anna Świrszczyńska, 2007
18. Johan Eellend, Cultivating the Rural Citizen: Modernity, Agrarianism and Citizenship in Late Tsarist Estonia, 2007
19. Petra Garberding, Musik och politik i skuggan av nazismen: Kurt Atterberg och de svensk-tyska musikrelationerna, 2007
20. Aleksei Semenenko, Hamlet the Sign: Russian Translations of Hamlet and Literary Canon Formation, 2007
21. Vytautas Petronis, Constructing Lithuania: Ethnic Mapping in the Tsarist Russia, ca. 1800–1914, 2007
22. Akvile Motiejunaite, Female employment, gender roles, and attitudes: the Baltic countries in a broader context, 2008
23. Tove Lindén, Explaining Civil Society Core Activism in Post-Soviet Latvia, 2008
24. Pelle Åberg, Translating Popular Education: Civil Society Cooperation between Sweden and Estonia, 2008
25. Anders Nordström, The Interactive Dynamics of Regulation: Exploring the Council of Europe’s monitoring of Ukraine, 2008
26. Fredrik Doeser, In Search of Security After the Collapse of the Soviet Union: Foreign Policy Change in Denmark, Finland and Sweden, 1988–1993, 2008
27. Zhanna Kravchenko. Family (versus) Policy: Combining Work and Care in Russia and Sweden, 2008
28. Rein Jüriado, Learning within and between public-private partnerships, 2008
29. Elin Boalt, Ecology and evolution of tolerance in two cruciferous species, 2008
30. Lars Forsberg, Genetic Aspects of Sexual Selection and Mate Choice in Salmonids, 2008
31. Eglė Rindzevičiūtė, Constructing Soviet Cultural Policy: Cybernetics and Governance in Lithuania after World War II, 2008
32. Joakim Philipson, The Purpose of Evolution: ’struggle for existence’ in the Russian-Jewish press 1860–1900, 2008
33. Sofie Bedford, Islamic activism in Azerbaijan: Repression and mobilization in a post-Soviet context, 2009
34. Tommy Larsson Segerlind, Team Entrepreneurship: A process analysis of the venture team and the venture team roles in relation to the innovation process, 2009

35. Jenny Svensson, The Regulation of Rule-Following: Imitation and Soft Regulation in the European Union, 2009
36. Stefan Hallgren, Brain Aromatase in the guppy, Poecilia reticulate: Distribution, control and role in behavior, 2009
37. Karin Ellencrona, Functional characterization of interactions between the flavivirus NS5 protein and PDZ proteins of the mammalian host, 2009
38. Makiko Kanematsu, Saga och verklighet: Barnboksproduktion i det postsovjetiska Lettland, 2009
39. Daniel Lindvall, The Limits of the European Vision in Bosnia and Herzegovina: An Analysis of the Police Reform Negotiations, 2009
40. Charlotta Hillerdal, People in Between – Ethnicity and Material Identity: A New Approach to Deconstructed Concepts, 2009
41. Jonna Bornemark, Kunskapens gräns – gränsens vetande, 2009 42. Adolphine G. Kateka, Co-Management Challenges in the Lake Victoria
Fisheries: A Context Approach, 2010 43. René León Rosales, Vid framtidens hitersta gräns: Om pojkar och
elevpositioner i en multietnisk skola, 2010 44. Simon Larsson, Intelligensaristokrater och arkivmartyrer: Normerna för
vetenskaplig skicklighet i svensk historieforskning 1900–1945, 2010 45. Håkan Lättman, Studies on spatial and temporal distributions of
epiphytic lichens, 2010 [report] 46. Alia Jaensson, Pheromonal mediated behaviour and endocrine response
in salmonids: The impact of cypermethrin, copper, and glyphosate, 2010 47. Michael Wigerius, Roles of mammalian Scribble in polarity signaling,
virus offense and cell-fate determination, 2010 48. Anna Hedtjärn Wester, Män i kostym: Prinsar, konstnärer och
tegelbärare vid sekelskiftet 1900, 2010 49. Magnus Linnarsson, Postgång på växlande villkor: Det svenska
postväsendets organisation under stormaktstiden, 2010 50. Barbara Kunz, Kind words, cruise missiles and everything in between: A
neoclassical realist study of the use of power resources in U.S. policies towards Poland, Ukraine and Belarus 1989–2008, 2010
51. Anders Bartonek, Philosophie im Konjunktiv: Nichtidentität als Ort der Möglichkeit des Utopischen in der negativen Dialektik Theodor W. Adornos, 2010
52. Carl Cederberg, Resaying the Human: Levinas Beyond Humanism and Antihumanism, 2010
53. Johanna Ringarp, Professionens problematik: Lärarkårens kommunalisering och välfärdsstatens förvandling, 2011
54. Sofi Gerber, Öst är Väst men Väst är bäst: Östtysk identitetsformering i det förenade Tyskland, 2011
55. Susanna Sjödin Lindenskoug, Manlighetens bortre gräns: Tidelagsrättegångar i Livland åren 1685–1709, 2011

56. Dominika Polanska, The emergence of enclaves of wealth and poverty: A sociological study of residential differentiation in post-communist Poland, 2011
57. Christina Douglas, Kärlek per korrespondens: Två förlovade par under andra hälften av 1800-talet, 2011
58. Fred Saunders, The Politics of People – Not just Mangroves and Monkeys: A study of the theory and practice of community-based management of natural resources in Zanzibar, 2011
59. Anna Rosengren, Åldrandet och språket: En språkhistorisk analys av hög ålder och åldrande i Sverige cirka 1875–1975, 2011
60. Emelie Lilliefeldt, European Party Politics and Gender: Configuring Gender-Balanced Parliamentary Presence, 2011
61. Ola Svenonius, Sensitising Urban Transport Security: Surveillance and Policing in Berlin, Stockholm, and Warsaw, 2011
62. Andreas Johansson, Dissenting Democrats: Nation and Democracy in the Republic of Moldova, 2011
63. Wessam Melik, Molecular characterization of the Tick-borne encephalitis virus: Environments and replication, 2012
64. Steffen Werther, SS-Vision und Grenzland-Realität: Vom Umgang dänischer und „volksdeutscher” Nationalsozialisten in Sønderjylland mit der „großgermanischen“ Ideologie der SS, 2012
65. Peter Jakobsson, Öppenhetsindustrin, 2012 66. Kristin Ilves, Seaward Landward: Investigations on the archaeological
source value of the landing site category in the Baltic Sea region, 2012 67. Anne Kaun, Civic Experiences and Public Connection: Media and Young
People in Estonia, 2012 68. Anna Tessmann, On the Good Faith: A Fourfold Discursive Construction
of Zoroastrianism in Contemporary Russia, 2012 69. Jonas Lindström, Drömmen om den nya staden: stadsförnyelse i det
postsovjetisk Riga, 2012 70. Maria Wolrath Söderberg, Topos som meningsskapare: retorikens
topiska perspektiv på tänkande och lärande genom argumentation, 2012 71. Linus Andersson, Alternativ television: former av kritik i konstnärlig
TV-produktion, 2012 72. Håkan Lättman, Studies on spatial and temporal distributions of
epiphytic lichens, 2012 73. Fredrik Stiernstedt, Mediearbete i mediehuset: produktion i förändring
på MTG-radio, 2013 74. Jessica Moberg, Piety, Intimacy and Mobility: A Case Study of
Charismatic Christianity in Present-day Stockholm, 2013 75. Elisabeth Hemby, Historiemåleri och bilder av vardag: Tatjana
Nazarenkos konstnärskap i 1970-talets Sovjet, 2013 76. Tanya Jukkala, Suicide in Russia: A macro-sociological study, 2013

77. Maria Nyman, Resandets gränser: svenska resenärers skildringar av Ryssland under 1700-talet, 2013
78. Beate Feldmann Eellend, Visionära planer och vardagliga praktiker: postmilitära landskap i Östersjöområdet, 2013
79. Emma Lind, Genetic response to pollution in sticklebacks: natural selection in the wild, 2013
80. Anne Ross Solberg, The Mahdi wears Armani: An analysis of the Harun Yahya enterprise, 2013
81. Nikolay Zakharov, Attaining Whiteness: A Sociological Study of Race and Racialization in Russia, 2013
82. Anna Kharkina, From Kinship to Global Brand: the Discourse on Culture in Nordic Cooperation after World War II, 2013
83. Florence Fröhlig, A painful legacy of World War II: Nazi forced enlistment: Alsatian/Mosellan Prisoners of war and the Soviet Prison Camp of Tambov, 2013
84. Oskar Henriksson, Genetic connectivity of fish in the Western Indian Ocean, 2013
85. Hans Geir Aasmundsen, Pentecostalism, Globalisation and Society in Contemporary Argentina, 2013
86. Anna McWilliams, An Archaeology of the Iron Curtain: Material and Metaphor, 2013
87. Anna Danielsson, On the power of informal economies and the informal economies of power: rethinking informality, resilience and violence in Kosovo, 2014
88. Carina Guyard, Kommunikationsarbete på distans, 2014 89. Sofia Norling, Mot "väst": om vetenskap, politik och transformation i Polen
1989–2011, 2014 90. Markus Huss, Motståndets akustik: språk och (o)ljud hos Peter Weiss
1946–1960, 2014 91. Ann-Christin Randahl, Strategiska skribenter: skrivprocesser i fysik och
svenska, 2014 92. Péter Balogh, Perpetual borders: German-Polish cross-border contacts in
the Szczecin area, 2014 93. Erika Lundell, Förkroppsligad fiktion och fiktionaliserade kroppar:
levande rollspel i Östersjöregionen, 2014 94. Henriette Cederlöf, Alien Places in Late Soviet Science Fiction: The
"Unexpected Encounters" of Arkady and Boris Strugatsky as Novels and Films, 2014
95. Niklas Eriksson, Urbanism Under Sail: An archaeology of fluit ships in early modern everyday life, 2014
96. Signe Opermann, Generational Use of News Media in Estonia: Media Access, Spatial Orientations and Discursive Characteristics of the News Media, 2014

97. Liudmila Voronova, Gendering in political journalism: A comparative study of Russia and Sweden, 2014
98. Ekaterina Kalinina, Mediated Post-Soviet Nostalgia, 2014 99. Anders E. B. Blomqvist, Economic Natonalizing in the Ethnic
Borderlands of Hungary and Romania: Inclusion, Exclusion and Annihilation in Szatmár/Satu-Mare, 1867–1944, 2014
100. Ann-Judith Rabenschlag, Völkerfreundschaft nach Bedarf: Ausländische Arbeitskräfte in der Wahrnehmung von Staat und Bevölkerung der DDR, 2014
101. Yuliya Yurchuck, Ukrainian Nationalists and the Ukrainian Insurgent Army in Post-Soviet Ukraine, 2014
102. Hanna Sofia Rehnberg, Organisationer berättar: narrativitet som resurs i strategisk kommunikation, 2014
103. Jaakko Turunen, Semiotics of Politics: Dialogicality of Parliamentary Talk, 2015
104. Iveta Jurkane-Hobein, I Imagine You Here Now: Relationship Maintenance Strategies in Long-Distance Intimate Relationships, 2015
105. Katharina Wesolowski, Maybe baby? Reproductive behaviour, fertility intentions, and family policies in post-communist countries, with a special focus on Ukraine, 2015
106. Ann af Burén, Living Simultaneity: On religion among semi-secular Swedes, 2015
107. Larissa Mickwitz, En reformerad lärare: konstruktionen av en professionell och betygssättande lärare i skolpolitik och skolpraktik, 2015
108. Daniel Wojahn, Språkaktivism: diskussioner om feministiska språkförändringar i Sverige från 1960-talet till 2015, 2015
109. Hélène Edberg, Kreativt skrivande för kritiskt tänkande: en fallstudie av studenters arbete med kritisk metareflektion, 2015
110. Kristina Volkova, Fishy Behavior: Persistent effects of early-life exposure to 17α-ethinylestradiol, 2015

Publikationer i serien Örebro Studies in Biology
1. Malmgren, Jan (2001) Evolutionary Ecology of Newts.
2. Sandström, Ulf G. (2004) Biodiversity and Green Infrastructure in Urban Landscapes.
3. Olsman, Helena (2005) Bioassay analysis of dioxin-like compounds – response interactions and environmental transformation of Ah receptor agonists.
4. Comstedt, Daniel (2008) Explaining temporal variations in soil respiration rates and δ13C in coniferous forest ecosystems.
5. Boström, Björn (2008) Achieving Carbon Isotope Mass Balance in Northern Forest Soils, Soil Respiration and Fungi.
6. Rotander, Anna (2011) Monitoring persistent organic pollutants (POPs) in sub-Arctic and Arctic marine mammals, 1984–2009.
7. Nordén, Marcus (2013) Comparative avian developmental toxicity of PFAAs.
8. Westman, Ola (2013) Development of bioassays for risk assessment of PAHs and PFASs in the environment.
9. Volkova, Kristina (2015) Fishy Behavior. Persistent effects of early- life exposure to 17α -ethinylestradiol.




















