
Fish Size as a Key Factor in Copper Toxicity to Fish: Impact of water-born copper on some...
12
Basrah J. Agric. Sci. 25 (special issued 2), 2012 14 Fish Size as a Key Factor in Copper Toxicity to Fish: Impact of water-born copper on some physiological indices in rainbow trout Oncorhynchus mykiss of various size groups Nader A. Salman*, Jeffery L. Ullman, Xiao Q. Lu *Department of Fisheries, College of Agriculture, university of Basrah, Iraq; Department of biological Systems Engineering, Washington State university, USA Abstract. Rainbow trout (Oncorhynchus mykiss) of various size groups representing one month larvae (0.3-0.6 g), three months fingerlings (5-7 g) and adult big fish (300-500 g) were exposed to different concentrations of CuSO 4 .5H 2 O for previously determined exposure periods. Several physiological parameters were measured to compare adaptability or toxicity cases. Those include ventilation rate, heartbeat rate, body water contents and behavior as well as survival or mortality rate. The results showed a direct positive relationship between adaptability or tolerance and fish size. Physiological responses varied with the concentration of the exposure medium and fish size. Sever effects were noticed among rainbow trout fry at the levels of 200-400 ppb Cu. Similar responses were noticed among rainbow trout fingerlings but at higher concentrations (600-800 ppb Cu). Toxicity of Adult big fish has not been registered below the level of 800 ppb. The results were discussed in term of physiological disturbances occurred in response of copper concentrations in the exposure media which led to adaptive tolerance or fatigue mortality. The present study concluded that fishery management of pollutants in freshwater bodies must have different approaches as far as fish size is concerned. Key wards: copper toxicity, fish size, ventilation, residual effect, ion regulation. Correspondent author: [email protected] Introduction Heavy metals have long been recognized as serious pollutants of the aquatic environment. The accumulation of metals in an aquatic environment has direct impact on man and aquatic ecosystem. Copper is one of the wastes commonly discharged into inland and coastal waters in many parts of the world. Cu +2 is an important xenobiotic in aquatic ecosystems as well as a non- degradable and cumulative pollutant. Increased copper concentrations in waters have resulted from the release of municipal wastewater, power plant effluents and marine antifouling paints. Industrial operations such as mining and smelting contribute to the mobilization of metals, including copper (Cu), into watersheds. The toxicity of Cu +2 to teleosts has been extensively studied, as summarized by (22). Cu is an essential micronutrient for normal metabolic functioning of all organisms, including fish (17). However, it can be highly toxic to aquatic organisms, particularly fish at concentrations above the physiological threshold levels (5). Interest in the metals, required for metabolic activities in organisms, lies in the narrow range between their essentiality and toxicity
Transcript of Fish Size as a Key Factor in Copper Toxicity to Fish: Impact of water-born copper on some...

Basrah J. Agric. Sci. 25 (special issued 2), 2012
14
Fish Size as a Key Factor in Copper Toxicity to Fish:
Impact of water-born copper on some physiological indices in
rainbow trout Oncorhynchus mykiss of various size groups
Nader A. Salman*, Jeffery L. Ullman, Xiao Q. Lu
*Department of Fisheries, College of Agriculture, university of Basrah, Iraq; Department of biological Systems Engineering, Washington State university, USA
Abstract. Rainbow trout (Oncorhynchus mykiss) of various size groups representing one month larvae (0.3-0.6 g),
three months fingerlings (5-7 g) and adult big fish (300-500 g) were exposed to different concentrations of
CuSO4.5H2O for previously determined exposure periods. Several physiological parameters were measured to
compare adaptability or toxicity cases. Those include ventilation rate, heartbeat rate, body water contents and
behavior as well as survival or mortality rate. The results showed a direct positive relationship between adaptability
or tolerance and fish size. Physiological responses varied with the concentration of the exposure medium and fish
size. Sever effects were noticed among rainbow trout fry at the levels of 200-400 ppb Cu. Similar responses were
noticed among rainbow trout fingerlings but at higher concentrations (600-800 ppb Cu). Toxicity of Adult big fish
has not been registered below the level of 800 ppb. The results were discussed in term of physiological disturbances
occurred in response of copper concentrations in the exposure media which led to adaptive tolerance or fatigue
mortality. The present study concluded that fishery management of pollutants in freshwater bodies must have
different approaches as far as fish size is concerned.
Key wards: copper toxicity, fish size, ventilation, residual effect, ion regulation.
Correspondent author: [email protected]
Introduction
Heavy metals have long been recognized as serious pollutants of the aquatic environment. The
accumulation of metals in an aquatic environment has direct impact on man and aquatic
ecosystem. Copper is one of the wastes commonly discharged into inland and coastal waters in
many parts of the world. Cu+2
is an important xenobiotic in aquatic ecosystems as well as a non-
degradable and cumulative pollutant. Increased copper concentrations in waters have resulted
from the release of municipal wastewater, power plant effluents and marine antifouling paints.
Industrial operations such as mining and smelting contribute to the mobilization of metals,
including copper (Cu), into watersheds. The toxicity of Cu+2
to teleosts has been extensively
studied, as summarized by (22).
Cu is an essential micronutrient for normal metabolic functioning of all organisms,
including fish (17). However, it can be highly toxic to aquatic organisms, particularly fish at
concentrations above the physiological threshold levels (5). Interest in the metals, required for
metabolic activities in organisms, lies in the narrow range between their essentiality and toxicity

Basrah J. Agric. Sci. 25 (special issued 2), 2012
15
(1). Copper is toxic, so homeostatic mechanisms are required to regulate internal levels of copper
(6) (18). Generally, absorbed Cu and other essential but potentially toxic trace metals might be
bound by a variety of biomolecules for metabolic function, storage, detoxification, toxicity, or
excretion (13)(21). Chronic exposure of fish to water-borne Cu, has been shown to cause a
variety of physiological and behavioral changes including ventilation rate, residual accumulation,
water and ionic disturbance and increased fish mortality. However, these effects are not
consistent and some studies have shown that there can be minimal physiological disturbances
even when exposure to Cu were sufficient to cause some mortality in fish (25).
One important factors in the toxic effect of copper on fishes is the life history stage and
size of fish when the exposure occurs. Few studies have examined the comparative sensitivities
of the major life stages and sizes of fishes: embryo, larva, fry, fingerling and adult. Developing
fish embryos or larvae are generally considered to be the most susceptible in the fish life cycle
(7), however there are only a few studies, which discuss the effects of Cu +2
in larval stages.
Nevertheless, developing fish embryos or larvae are more sensitive to the toxicity of heavy
metals than are adult fish, and they can serve as biological indicators for determination of water
quality (27). Rainbow trout (Oncorhynchus mykiss) was found to be more sensitive to copper in
the embryonic stage than in the larval and fry stages ((11). The incipient lethal concentration
LC50 for trout, embryos exposed to copper was found to be approximately 30 times lower than
the LC50 for trout larvae and 7 times lower than the LC50 for trout adults (15).
The aim of this study is to examine the role of body size in copper toxicity test, to
determine differences in physiological responses between rainbow trout fry and fingerlings
toward sub-lethal concentration of water-born copper and to evaluate the usefulness of residual
copper accumulation, ventilation rate, water contents and body sodium as bio-indicators.
Materials and Methods
Reagents
All reagents were of analytical reagent grades purchased from Sigma-Aldrich unless otherwise
stated. Milli-Q Millipore (double deionized water) was used for dilution of standard solution and
samples. All the glassware was soaked in 10% HNO3 and was rinsed with tap water and then
Milli-Q water prior to use.
Exposure of rainbow trout to water born copper
Copper stock solution used in the toxicity tests was prepared by dissolving Cu2SO4.5H2O into a
desirable concentration using Milli Q water. Exposure of rainbow trout fry and fingerling size
was conducted in glass aquaria (28 x 72 x 42 cm) under controlled temperature (13 ± 1 oC) and
artificial aeration. Randomly chosen rainbow trout fry (40) and fingerling (20) size raised in our
fish laboratory were acclimatized to each test aquarium for 24h with aeration. Afterwards, the
fish were exposed to copper by spiking the metal with different concentrations (0, 100, 200 and
400µg/L) into the aquarium.
Food was not provided during the acclimation and in the course of exposures. Copper
concentration in water was analyzed after copper stock solution was spiked into the aquarium
and was then analyzed after the exposure experiment was completed. During exposure, rainbow
trout fish were collected and were sacrificed using a solution containing 0.1% of Tricaine
anesthetics. After scarification, the fish samples were transferred into aluminum trays and air

Basrah J. Agric. Sci. 25 (special issued 2), 2012
16
dried until a constant weight. The dry samples were grounded into fine power using a prewashed
pestle and mortar.
Digestion procedure
The sample was digested following the procedure given elsewhere (Chen et al. 2000). Briefly,
about 200 to 400 mg of finely powered sample was accurately weighed and digested with 10 ml
of concentrated HNO3 in a hot plate at 120 °C for 6 hours. The sample was then added with 2 ml
of 30% hydrogen peroxide to ensure that the sample was fully digested.
Sample was then transferred into 50 ml of volumetric flask and made up to a final volume
with Milli-Q Millipore water. Two sample blanks were taken through sample digestion with
every batch (12 samples). The digestion method was verified through digestion of the certified
meat reference material (CMRMs, 1546) purchased from National Institute of Standards and
Technology (NIST). Agreement between certified value and concentration of copper quantified
on ICP/MS in out laboratory was less than 5% (26).
Sample analysis on ICP MS
Samples were run on Agilent 7500cx ICP/MS. The instrument was warmed up at least for 40
min and then was optimized before running samples by using a tuning solution provided by the
manufactory, Agilent. The tuning solution contains 7Li,
89Y,
140Ce and
205Ti and (1 ug/L), which
cover the whole mass range, (e.g., 7Li at low mass,
89Y at medium mass and
205Ti at high mass)
in order to obtain optimum signal intensity for multi-element analysis. The operating parameters
of the instrument were adjusted for optimum sensitivity (e.g., 7Li 1000 –2000 counts, and Ti
2000 – 3000 counts), while keeping the formation of refractory oxides formed within the
ICP/MS to a minimum (that is CeO/Ce, ≤1%) to avoid excessive spectral interferences. After
optimum, semi quantitative analysis was conducted using a freshly prepared dilute acid solution
containing 2% HNO3 in order to check whether the system of instrument was relatively clear
without across contamination from previous run. Concentrations of metals in crayfish were
quantified using an external standard method.
The standard solutions were prepared by diluting the stock multi-elements standard
solution into desirable concentrations in 2% nitric acid. A blank and a QC standard were
introduced in very ten samples in order to monitor the instrument performance during entire
samples running on the instrument.
Results and Discussion
Mortality
Mortality rates in two sizes of rainbow trout exposed to different levels of water borne copper for
96h are shown in Table 1. Insignificant mortality levels of rainbow trout fry occurred in the
control and 100 ppb levels. Mortality of less than 50% was seen in fry exposed to 200 ppb.
Significant mortality was noticed in the highest copper level (400 ppb) after 48h exposure
compared with control. It increased to 97.2% after 72h and 96h.
Less than 50% mortality was shown among trout fingerlings in all treatment (0-200
ppb) except the highest concentration (400 ppb) where 87% of the fish died after 24h. No
mortalities occurred in the control group and at 100 ppb level. Mortalities occurred only during
the first 24 h exposure in concentrations of more than 100 ppb. No more mortality was recorded

Basrah J. Agric. Sci. 25 (special issued 2), 2012
17
at 48, 72 and 96h of exposure to the same copper concentration, indicating a state of adaptation
after the initial toxicity shock.
For comparison trout fingerlings showed insignificant difference in mortality rates than
those recorded in the fry stage. It appeared that the fingerlings adapted more quickly than the fry
as mortality stopped after the first shock within 24h while it continued during the following
periods in the fry test. In both only the highest concentration (about 400 ppb) caused more than
50% mortality. The present results are comparable to those reported by (8) that 10-g rainbow
trout were 2.5 times more tolerant to Cu than were 0.7-g trout. The effect of body size was also
accounted by (23) for variations in species sensitivity towards Cu-toxicity beside water
chemistry. Apparently small-sized fish are more prone to Cu toxicity because they have a high
efficiency of absorption coupled with immaturity of the excretion system as stated by (9).
According to (12) Cu is the more toxic metal for rainbow trout (Onchorchyncus mykiss) at all
stages of development. The sensitivity of different life stages of rainbow trout (based on 96-h
LC50) might be indicated in the following sequences: larvae>adult fish>eggs (15).
Table (1): Percentage mortalities of rainbow trout fry and fingerlings exposed to four levels
of water-born copper for 96h. (Date represent 20 fish per tank and 2 tanks per
treatment)
Fish size Water
Copper
conc. ppb
% Mortality
after
24h
48h
72h
96
Fry control 0 1.7 2.8 3.3
100 0 1.1 2.2 2.7
200 0 11.6 28.3 33.8
400 8.8 89.1 97.2 97.2
Fingerlings control 0 0 0 0
100 0 0 0 0
200 29 29 29 29
400 87 87 87 87
Ventilation rate
Rainbow trout fry exposed to concentrations of copper sulfate exceeding 200 ppb responded by a
decrease in their ventilation activity. Values of 73-78 movement per minute in high Cu
concentrations are significantly lower than those recorded in control fish (127 -131 per min). A
progressive decrease in the ventilation rate was recorded in all fish as the experiment proceeds to
the final exposure period of 96h. Levels of Cu have also raised and ventilation rate decreased in
all treatments to values ranging from 79.2 to 91.8 per minute (Fig. 1).
Data of Table (2) showed ventilation rate of trout fingerlings which were exposed to
higher Cu concentrations after adaptation period on lower copper media. It seems that the
adapting process masked the breathing stress exhibited by fish suddenly transferred to water with
certain level of copper. In general values of ventilation rate were lower in fingerlings than those
recorded in the fry size. This may indicate better handling of the exposure stress. With elevated

Basrah J. Agric. Sci. 25 (special issued 2), 2012
18
levels of copper in the water media, the primary method of uptake of the metal is via the gills.
According to (28) Copper diffuses passively across the gill epithelia into the bloodstream. Such
mechanism can disturb the ordinary ventilation movements.
Table (2): Ventilation of trout fingerlings exposed to high copper levels after 2 weeks
adaptation
Adapted Cu
medium
ppb
Exposed Cu
medium
ppb
Ventilation
rate per min.
100 400 93
200 400 102
100 600 93
200 600 80
100 800 104
Figure (1): Ventilation rate per minute in rainbow trout fry exposed to different levels of
copper sulfate for 24, 48, 72 and 96 hours
Residual Copper
Slight insignificant increase in the residual copper in the tissues of rainbow trout fry was
noticed. The data in Table 3 revealed a significant increase in fish exposed to 100 and 200 ppb of
copper after 96h reaching 2.8 and 3.1 mg/kg dry wt. respectively compared with the control
treatment. This indicates a progressive accumulation of copper in the whole body of fry fish.

Basrah J. Agric. Sci. 25 (special issued 2), 2012
19
In the fingerling exposure trial, two ascending lines are obvious upon examining the
residual copper data in rainbow trout fingerlings as appeared in Table 3. One is the significant
increase in the body copper contents of fingerlings exposed to nearly 200 and 400 ppb Cu. This
increment was not significant during the first 48h of exposure. However another ascending trend
which is more related to time of exposure appeared after 72 and 96h. Such significant (p<0.05)
trend has pointed out the importance of calculating the multiple regression equation for the
relationship between Cu concentration in water and exposure time with the residual copper
contents. As seen in the three dimensional graph in Fig. 2, the equation is:
Water Cu concentration = 111.794+33.565 Residual Cu. + 0.266Time
Table (3): Residual copper concentration (mg/kg dry wt. ± SD n=3) in the whole body of
rainbow trout fingerlings and fry exposed to various levels of CuSO4.5H2O for 96h
period
Time
interval (h)
Exposed Cu
(PPb)
Fingerling body
Cu
(mg. kg-1
dry wt.)
Fry body Cu
(mg. kg-1
dry wt.)
24 control 1.8 ± 0.5 1.8 ± 0.5
100 1.5 ± 0.5 1.6 ± 0.4
200 1.6 ± 0.7 2.0 ± 0.3
400 2.4* 2.0 ± 0.4
48 control 4.1 ± 0.6 1.5 ± 0.3
100 4.0 ± 0.3 1.9 ± 0.4
200 5.1 ± 0.4 1.8 ± 0.1
400 5.6 ± 0.5 -
72 control 3.7 ± 0.6 2.1 ± 0.2
100 4.4 ± 1.1 2.7 ± 0.7
200 5.3 ± 0.1 2.0 ± 0.4
400 5.2 ± 2.2 -
96 Control 5.1 ± 1.0 1.8 ± 1.0
100 5.6 ± 0.9 2.8 ± 0.9
200 10.1 ±6.0 3.1 ± 6.0
400 8.2 ± 3.6 -
*A sample of one fish only
The effect of water-borne copper on the accumulation of copper in the fish body was
more obvious in the fingerlings more than the fry. Residual copper values recorded in the bigger
size fingerlings are nearly 4-5 times those recorded in the smaller-size fry. The above
observations raised the question of the importance of fish size as a limiting factor in copper
toxicity to fish. In this respect, (10) observed in juvenile rainbow trout (Oncorhynchus mykiss)

Basrah J. Agric. Sci. 25 (special issued 2), 2012
20
during exposure to sublethal waterborne Cu (40 µg/L) for 21 days, a time-dependent
accumulation of Cu in the gill, liver, plasma, and carcass, with significant difference in Cu-
exposed fish relative to the controls being evident by day 7. Their data suggest that rainbow trout
can withstand significant above-background Cu accumulation in hepatic putative metal-sensitive
compartments without chronic toxic effects at the organism level.
Traditionally copper residues in liver have been used as an indicator of fish exposure to
copper (2). Once copper has entered the bloodstream it is transported to the liver which is the
primary site of accumulation of the metal within fish (22). For xenobiotic residue level, in
particular at the site of action, will be reflected by adverse effects, but for essential elements,
including copper, it is much harder to determine the level, which results in adverse effects.
Lowest observed effect concentration is the lowest concentration exposure to which results in
adverse effects such as residue level and structural or physiological changes. If any of these
changes can be related to exposure concentration or time and show enough specificity it can be
used as a biomarker.
Figure (2). Multiple regression diagram for the relationship between Cu concentration in
water and exposure time with the residual copper contents in rainbow trout
fingerlings
Basrah J. Agric. Sci. 25 (special issued 2), 2012
21
Water and Ionic Balance
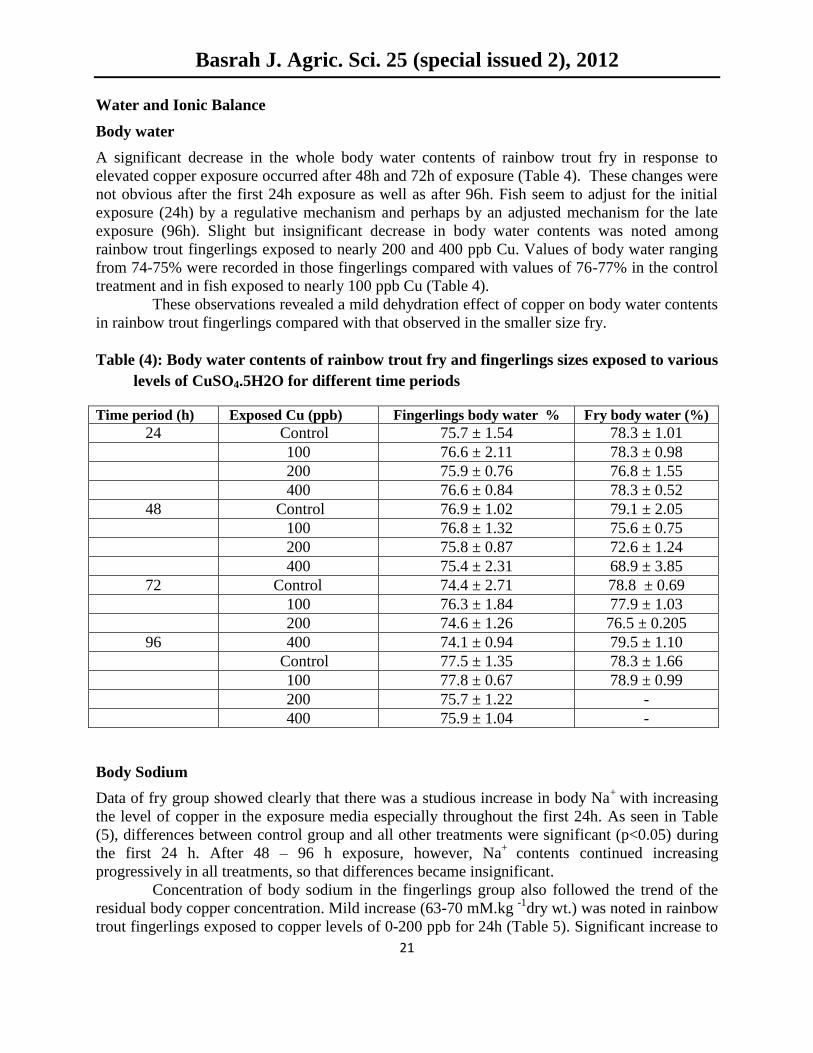
Body water
A significant decrease in the whole body water contents of rainbow trout fry in response to
elevated copper exposure occurred after 48h and 72h of exposure (Table 4). These changes were
not obvious after the first 24h exposure as well as after 96h. Fish seem to adjust for the initial
exposure (24h) by a regulative mechanism and perhaps by an adjusted mechanism for the late
exposure (96h). Slight but insignificant decrease in body water contents was noted among
rainbow trout fingerlings exposed to nearly 200 and 400 ppb Cu. Values of body water ranging
from 74-75% were recorded in those fingerlings compared with values of 76-77% in the control
treatment and in fish exposed to nearly 100 ppb Cu (Table 4).
These observations revealed a mild dehydration effect of copper on body water contents
in rainbow trout fingerlings compared with that observed in the smaller size fry.
Table (4): Body water contents of rainbow trout fry and fingerlings sizes exposed to various
levels of CuSO4.5H2O for different time periods
Time period (h) Exposed Cu (ppb) Fingerlings body water % Fry body water (%)
24 Control 75.7 ± 1.54 78.3 ± 1.01
100 76.6 ± 2.11 78.3 ± 0.98
200 75.9 ± 0.76 76.8 ± 1.55
400 76.6 ± 0.84 78.3 ± 0.52
48 Control 76.9 ± 1.02 79.1 ± 2.05
100 76.8 ± 1.32 75.6 ± 0.75
200 75.8 ± 0.87 72.6 ± 1.24
400 75.4 ± 2.31 68.9 ± 3.85
72 Control 74.4 ± 2.71 78.8 ± 0.69
100 76.3 ± 1.84 77.9 ± 1.03
200 74.6 ± 1.26 76.5 ± 0.205
96 400 74.1 ± 0.94 79.5 ± 1.10
Control 77.5 ± 1.35 78.3 ± 1.66
100 77.8 ± 0.67 78.9 ± 0.99
200 75.7 ± 1.22 -
400 75.9 ± 1.04 -
Body Sodium
Data of fry group showed clearly that there was a studious increase in body Na+
with increasing
the level of copper in the exposure media especially throughout the first 24h. As seen in Table
(5), differences between control group and all other treatments were significant (p<0.05) during
the first 24 h. After 48 – 96 h exposure, however, Na+
contents continued increasing
progressively in all treatments, so that differences became insignificant.
Concentration of body sodium in the fingerlings group also followed the trend of the
residual body copper concentration. Mild increase (63-70 mM.kg -1
dry wt.) was noted in rainbow
trout fingerlings exposed to copper levels of 0-200 ppb for 24h (Table 5). Significant increase to

Basrah J. Agric. Sci. 25 (special issued 2), 2012
22
97 mM.kg -1
dry wt., however, was recorded in fish exposed to 400 ppb water-born copper. After
48-96h exposure at the same copper levels, concentrations of body sodium showed remarkable
increments to levels reaching about 200 mM.kg -1
dry wt. along with minor differences between
treatments due to exposure level. For comparison between the two size groups (fry and
fingerlings), levels of body sodium in the fingerlings significantly exceed those recorded in the
fry in all treatments (Fig. 3). This might be due to more effective Na+ excretion mechanism
occurred in fry or less Na+
influx in response to elevated copper exposure compared with the
fingerlings group. A possible additional strategy for fish to compensate for Cu-induced branchial
Na+ losses is to increase uptake to meet their Na
+ requirement through branchial uptake. Similar
comparable observations were recorded by (24) and (29).
Table (5): Whole body sodium (mM.kg -1
dry wt. ± SE ) in rainbow trout fry and fingerlings
exposed to various levels of water-borne copper for 24-96h.
Time period
(h)
Water Cu
(ppb)
Na+ mM.kg
-1dry
wt. Fry
Fingerlings
24 control 35.1 ± 3.8 a 63.0 ± 17.1
a
100 40.6 ± 5.8 a 67.1 ± 23.0
a
200 75.0 ± 6.5 b 70.3 ± 28.5
a
400 76.2 ± 4.4 b 97.3 ± 30.4
b
48 Control 96.2 ± 3.2 191.0 ± 39.2
100 101.6 ± 2.1 218.3 ± 15.5
200 98.5 ± 2.1 229.0 ± 21.8
400 - 212.9 ± 40.5
72 Control 144.0 ± 3.0 176.6 ± 42.3
100 120.5 ± 6.0 205.9 ± 47.2
200 136.8 ± 5.1 239.0 ± 4.6
400 - 129.8 ± 57.3
96 Control 163.7 ± 1.3 228.6 ± 15.0
100 168.4 ± 10.3 245.5 ± 20.0
200 159.4 ± 12.2 232.2 ± 17.7
400 - 204.5 ± 16.8

Basrah J. Agric. Sci. 25 (special issued 2), 2012
23
Fry Fingerlings
Figure (3): Three dimensional histogram of the relationship between body sodium and
exposed copper media and exposure time in rainbow trout fry and fingerlings
It has been reported that, the primary mechanisms of Cu toxicity to freshwater fish is an
osmoregulatory disturbance result from the combined effects of a decrease in Na+ influx and an
increase in Na+ efflux, which yield a net reduction of plasma and whole body Na+ (14). On
contrary, in the present results a progressive increase in body sodium was seen in both fry and
fingerlings exposed to 0-400 ppb Cu. This has also been reported in tilapia larvae as K+
increased by 23% and Na+ increased by 26% due to exposure to 30 µg/L Cu
+2 for 72 h (30).
Copper exposure stress can be counteracted by different defense modes such as physiological
compensation of ions (19) (16). Previous studies found this phenomenon in rainbow trout
(Oncorhynchus mykiss). Exposure to Cu+2
caused disruption of the Na+
and Ca+2
balance, which
became evident within two days of exposure and subsequently recovered to control levels (16).
Stress caused a brief increase in diffusive Na+ efflux and a brief declines in plasma Na
+ and Cl
-
concentrations, and this disturbance was rapidly compensated for by a three-fold increase in Na+
and Cl- influx in rainbow trout (19).
It seems that mechanisms of copper toxicity are directly or indirectly associated with
Na+ metabolism. The interaction between sodium and copper was previously reported as Cu can
affect the epithelial Na+ channel (2) (4) (20). Accordingly, interference with Na
+ homeostasis is
the fundamental mechanism of acute waterborne Cu toxicity (14). In the present study, after
prolonged sublethal exposure to waterborne Cu, a partial or complete recovery of ionoregulatory
function may occurred. This could be related, in part to an increase in Na+/K
+-ATPase activity as
stated by (16).
Acknowledgement
The sponsorship of International Institute of Education (IIE) and SRF-Iraq program to the
fellowship of the first author in Washington State University, Pullman is highly appreciated.

Basrah J. Agric. Sci. 25 (special issued 2), 2012
24
References
1. Fatoki, O.S., Lujiz N. Ogunfowokan, A.O. 2002. Trace metals Pollution in Umtatma river.
Water South Africa, 28: 83–89.
2. Grosell, M., Wood, C.M. 2002. Copper uptake across rainbow trout gills: mechanisms of
apical entry. J. Exp. Biol. 205: 1179–1188.
3. Handy, R. D. 1996. Dietary exposure to toxic metals in fish. In Toxicology of Aquatic
Pollution: Physiological, Cellular and Molecular Approaches (Taylor, E. W., ed.), pp.
29–60. Cambridge: Cambridge University Press.
4. Handy, R.D., Eddy, F.B., and Baines, H. 2002. Sodium-dependent copper uptake across
epithelia: a review of rationale with experimental evidence from gill and intestine.
Biochim. Biophys. Acta, 1566: 104–115.
5. Handy, R. D. 1996. Dietary exposure to toxic metals in fish. In Toxicology of Aquatic
Pollution: Physiological, Cellular and Molecular Approaches (Taylor, E. W., ed.), pp.
29–60. Cambridge: Cambridge University Press.
6. Harris E.D. 1991 Copper transport: an overview. Proc. Soc. Exp. Biol. Med. 196:130–140
7. Heath A.G. 1995. Water and pollution and fish physiology. CRC Press, Florida, p 245.
8. Howarth, R.S., Sprague, J.B. 1978. Copper lethality to rainbow trout in waters of various
hardness and pH. Water Res. 12:455-462.
9. Kamunde C, Grosell M, Higgs D, Wood C.M. 2002. Copper metabolism in actively growing
rainbow trout (oncorhynchus mykiss): copper uptake. J Exp Biol 205:279–290.
10. Kamunde, C and MacPhail, R. 2008. Bioaccumulation and hepatic speciation of copper in
rainbow trout (Oncorhynchus mykiss) during chronic waterborne copper exposure. Arch
Environ Contam Toxicol (2008) 54:493–503.
11. Kazlauskiene, N. 2004. Classification and Possible Consequences of the Sublethal Effects of
Heavy Metal Model Mixture in Fish at Early Ontogenesis. Ed. M. Anke et al. 22.
Workshop 2004. Macro and Trace Elements; Mengen- and Spurenelemente. SCHU
BERT-Verlag, Leipzig. 1. Auflage. pp 1172-1177.
12. Kazlauskienė, N. Vosylienė, M. Z. 2008. Characteristic Features of the Effect of Cu and Zn
Mixtures on Rainbow Trout Oncorhynchus mykiss in Ontogenesis. Polish J. of Environ.
Stud. 17, (2): 291-293.
13. Klaassen C.D, Liu J, Choudhuri S. 1999. Metallothionein: an intracellular protein to protect
against cadmium toxicity. Annu Rev Pharmacol Toxicol 39:267–294
14. Laurén, D.J., and McDonald, D.G. 1987. Acclimation to copper by rainbow trout, Salmo
gairdneri: physiology. Can. J. Fish. Aquat. Sci. 44: 99–104.
15. Marciul Ioniene D., Montvydiene, D. Kazlauskiene, N. Svecevicius, G. 2002. Comparative
analysis of the sensitivity of test-organisms of different phylogenetic level and life
stages to heavy metals. Environ. & Chem. Physics. 24 (2): 73.

Basrah J. Agric. Sci. 25 (special issued 2), 2012
25
16. McGeer, J.C., Szebedinszky, C., McDonald, D.G., and Wood, C.M. 2000. Effects of
sublethal exposure to waterborne Cu, Cd or Zn in rainbow trout. 1: Ionoregulatory
disturbance and metabolic costs. Aquat. Toxicol. 50: 231–243.
17. Ogino, C., Yang, G.Y., 1980. Requirements of carp and rainbow trout for dietary manganese
and copper. Bull. Jpn. Soc. Sci. Fish. 46, 455–458.
18. Pena, M. M. O., Lee, J. & Thiele, D. J. 1999. A delicate balance: homeostatic control of
copper uptake and distribution. Journal of Nutrition 129, 1251–1260.
19. Postlethwaite E. K, McDonald D.G. 1995. Mechanisms of Na+ and Cl
- regulation in
freshwater adapted rainbow trout (Oncorhynchus mykiss) during exercise and stress. J
Exp. Biol. 198:295–304.
20. Pyle, G.G., Kamunde, C.N., McDonald, D.G., and Wood, C.M. 2003. Dietary sodium
inhibits aqueous copper uptake in rainbow trout, Oncorhynchus mykiss. J. Exp. Biol.
206: 609–618.
21. Rainbow P.S. 2002. Trace metal concentrations in aquatic invertebrates: why and so what?
Environ Pollut 120:71–80
22. Sorensen EM (1991) Cadmium. In: Metal poisoning in fish. CRC Press, Boca Raton, Florida,
pp 175–234
23. Spear, P.A., Pierce, R.C. 1979. Copper in the aquatic environment: Chemistry, distribution
and toxicology.NRCC 16454. National Research Council of Canada, Ottawa, ON.
24. Sullivan G.V., Fryer J.N., Perry S.F. 1995. Immunolocalization of proton pump (H1-ATPase)
in pavement cells of rainbow trout gill. J Exp Biol 198:2619–2629.
25. Taylor, L.N., J.C. McGeer, C.M. Wood and D.G. McDonald, 2000. The physiological effects
of chronic cooper exposure to rainbow trout (Oncorhynchus mykiss) in hard and soft
water. An evaluation of chronic indicators. Environ. Toxicol., 9: 2298–2308.
26. Ullman et al., Toxic metals of As, Cd, Pb and essential metals of Co, Cr, Cu, Fe, Mn, Ni, Se,
V and Zn measured in cattle, goat, and chicken collected from As affected districts in
Bangladesh. Un-published results.
27. Westernhagen H.V. 1988. Sublethal effects of pollutants on fish eggs and larvae. In: Hoar
WS, Randall DJ (eds) Fish physiology, volume 11. The physiology of developing fish.
Part A; eggs and larvae. Academic Press, San Diego, California, pp 253–346.
28. Wilson R.W., Taylor E.W. 1993. The physiological response of freshwater rainbow trout,
Oncorhynchus mykiss, during acutely lethal copper exposure. J Comp Physiol B
163:38–47.
29. Wood C.M. 2001. Toxic responses of the gill. In: Schlenck D, Benson WH (Eds) Target
organ toxicity in marine and freshwater teleosts. Vol. 1. Organs. Taylor & Francis,
London, pp 1–89.
30. Wu S. M., Jong, K. J, Kuo, S. Y. 2003. Effects of copper sulfate on ion balance and growth
in tilapia larvae (Oreochromis mossambicus). Arch. Environ. Contam. Toxicol. 45,
357–363



















