
FIRST REPORT OF IMMATURES, GENITALIA AND MATERNAL CARE IN EUGENYSA COLUMBIANA (BOHEMAN) (COLEOPTERA:...
18
50 The Coleopterists Bulletin, 56(1):50–67. 2002. FIRST REPORT OF IMMATURES,GENITALIA AND MATERNAL CARE IN EUGENYSA COLUMBIANA (BOHEMAN) (COLEOPTERA:CHRYSOMELIDAE: CASSIDINAE:EUGENYSINI) CAROLINE S. CHABOO Department of Entomology, Comstock Hall Cornell University, Ithaca, NY 14853, U.S.A. Abstract Illustrated descriptions of larvae, pupae and adult genitalia are provided for Eugenysa columbiana (Boheman) in Costa Rica. These represent the first such descriptions for the tribe Eugenysini. Females are reported as exhibiting guarding of larvae and pupae. Male presence with the female and larval group is also described. The distribution range is extended to Peru, and host plant data is recorded. A new character from the spermatheca is presented and its potential usefulness in deducing eugenysine relationships is dis- cussed. Resu ´ men Se presentan descripciones ilustradas de larvas y pupas y genitalia de adultos para Eugenysa columbiana (Boheman) en Costa Rica que constituyen el primer registo para la tribu. Se reporta el cuidado maternal de larvas y pupas. Se describe tambie ´n la pre- sencı ´a del macho con la hembra y el grupo de larvas. El rango de distribucio ´ n se extienda a ` Peru y se presentan datos de la planta hospedejera. Se presenta un nuevo caracter de la spermateca y se discute uso el potencial para clarificar las relaciones filogeneticas de los escarabajos eugenysines. Eugenysa columbiana (Boheman) is a member of the Neotropical tribe Eu- genysini which consists of 33 species in three genera, Agenysa Spaeth (8 spe- cies), Eugenysa Chevrolat (19 species), and Miocalaspis Weise (6 species) (Blackwelder 1946; Seeno and Wilcox 1982; Borowiec 1999). The tribe ex- tends from Argentina and Brazil to Mexico with one species, Agenysa guia- niensis (Boheman), also recorded on the Caribbean island of Curac ¸ao (Boro- wiec 1999). Viana (1968) revised the Eugenysini, with good keys and descrip- tions for adults of the species. Descriptions of genitalia and immatures were not included then and are still undescribed. Eugenysines are among the largest and showiest cassidines yet their natural history is poorly known. Host plants for only two species have been recorded: an Ipomoea sp. (Convolvulaceae) for Agenysa boliviana Spaeth (Viana 1968) and Mikania guaco Bonpland (Asteraceae) for Eugenysa coscaroni Viana (Windsor and Choe 1994). Maternal care has been reported in E. coscaroni (Windsor and Choe 1994). This remarkable behavior is a rare phenomenon in the Chrysomelidae; among the approximately 40,000 species in the family (Lawrence 1982), only 22 spe- cies in 8 genera from the subfamilies Chrysomelinae and Cassidinae have been reported as maternal care providers (Table 1). Knowledge of these species ranges from fairly detailed observations (Windsor 1987; Choe 1989; Frieiro- Costa 1995; Kudo and Ishibashi 1995; Kudo et al. 1995), to brief notes (Monte 1932; Preston-Mafham 1993) or just a single photograph (O’Toole and Pres- ton-Mafham 1985). In all these species, females show a stereotypical behavior
-
Upload
un-lincoln -
Category
Documents
-
view
0 -
download
0
Transcript of FIRST REPORT OF IMMATURES, GENITALIA AND MATERNAL CARE IN EUGENYSA COLUMBIANA (BOHEMAN) (COLEOPTERA:...

50
The Coleopterists Bulletin, 56(1):50–67. 2002.
FIRST REPORT OF IMMATURES, GENITALIA AND MATERNAL CARE IN
EUGENYSA COLUMBIANA (BOHEMAN) (COLEOPTERA: CHRYSOMELIDAE:CASSIDINAE: EUGENYSINI)
CAROLINE S. CHABOO
Department of Entomology, Comstock HallCornell University, Ithaca, NY 14853, U.S.A.
Abstract
Illustrated descriptions of larvae, pupae and adult genitalia are provided for Eugenysacolumbiana (Boheman) in Costa Rica. These represent the first such descriptions for thetribe Eugenysini. Females are reported as exhibiting guarding of larvae and pupae. Malepresence with the female and larval group is also described. The distribution range isextended to Peru, and host plant data is recorded. A new character from the spermathecais presented and its potential usefulness in deducing eugenysine relationships is dis-cussed.
Resumen
Se presentan descripciones ilustradas de larvas y pupas y genitalia de adultos paraEugenysa columbiana (Boheman) en Costa Rica que constituyen el primer registo parala tribu. Se reporta el cuidado maternal de larvas y pupas. Se describe tambien la pre-sencıa del macho con la hembra y el grupo de larvas. El rango de distribucion se extiendaa Peru y se presentan datos de la planta hospedejera. Se presenta un nuevo caracter dela spermateca y se discute uso el potencial para clarificar las relaciones filogeneticas delos escarabajos eugenysines.
Eugenysa columbiana (Boheman) is a member of the Neotropical tribe Eu-genysini which consists of 33 species in three genera, Agenysa Spaeth (8 spe-cies), Eugenysa Chevrolat (19 species), and Miocalaspis Weise (6 species)(Blackwelder 1946; Seeno and Wilcox 1982; Borowiec 1999). The tribe ex-tends from Argentina and Brazil to Mexico with one species, Agenysa guia-niensis (Boheman), also recorded on the Caribbean island of Curacao (Boro-wiec 1999). Viana (1968) revised the Eugenysini, with good keys and descrip-tions for adults of the species. Descriptions of genitalia and immatures werenot included then and are still undescribed.
Eugenysines are among the largest and showiest cassidines yet their naturalhistory is poorly known. Host plants for only two species have been recorded:an Ipomoea sp. (Convolvulaceae) for Agenysa boliviana Spaeth (Viana 1968)and Mikania guaco Bonpland (Asteraceae) for Eugenysa coscaroni Viana(Windsor and Choe 1994).
Maternal care has been reported in E. coscaroni (Windsor and Choe 1994).This remarkable behavior is a rare phenomenon in the Chrysomelidae; amongthe approximately 40,000 species in the family (Lawrence 1982), only 22 spe-cies in 8 genera from the subfamilies Chrysomelinae and Cassidinae have beenreported as maternal care providers (Table 1). Knowledge of these speciesranges from fairly detailed observations (Windsor 1987; Choe 1989; Frieiro-Costa 1995; Kudo and Ishibashi 1995; Kudo et al. 1995), to brief notes (Monte1932; Preston-Mafham 1993) or just a single photograph (O’Toole and Pres-ton-Mafham 1985). In all these species, females show a stereotypical behavior
51THE COLEOPTERISTS BULLETIN 56(1), 2002
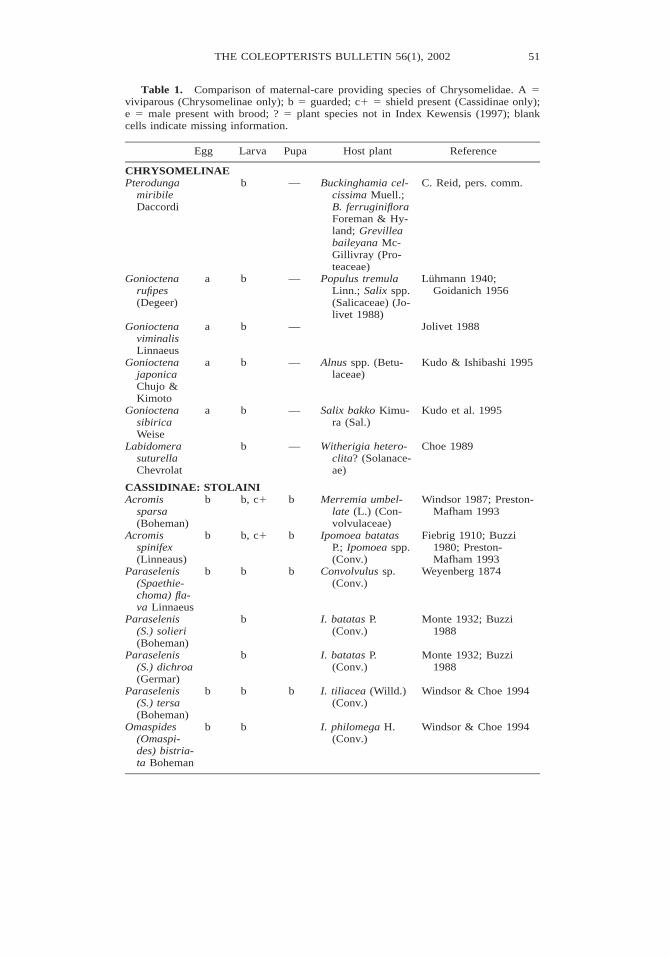
Table 1. Comparison of maternal-care providing species of Chrysomelidae. A 5viviparous (Chrysomelinae only); b 5 guarded; c1 5 shield present (Cassidinae only);e 5 male present with brood; ? 5 plant species not in Index Kewensis (1997); blankcells indicate missing information.
Egg Larva Pupa Host plant Reference
CHRYSOMELINAEPterodunga
miribileDaccordi
b — Buckinghamia cel-cissima Muell.;B. ferruginifloraForeman & Hy-land; Grevilleabaileyana Mc-Gillivray (Pro-teaceae)
C. Reid, pers. comm.
Gonioctenarufipes(Degeer)
a b — Populus tremulaLinn.; Salix spp.(Salicaceae) (Jo-livet 1988)
Luhmann 1940;Goidanich 1956
GonioctenaviminalisLinnaeus
a b — Jolivet 1988
GonioctenajaponicaChujo &Kimoto
a b — Alnus spp. (Betu-laceae)
Kudo & Ishibashi 1995
GonioctenasibiricaWeise
a b — Salix bakko Kimu-ra (Sal.)
Kudo et al. 1995
LabidomerasuturellaChevrolat
b — Witherigia hetero-clita? (Solanace-ae)
Choe 1989
CASSIDINAE: STOLAINIAcromis
sparsa(Boheman)
b b, c1 b Merremia umbel-late (L.) (Con-volvulaceae)
Windsor 1987; Preston-Mafham 1993
Acromisspinifex(Linneaus)
b b, c1 b Ipomoea batatasP.; Ipomoea spp.(Conv.)
Fiebrig 1910; Buzzi1980; Preston-Mafham 1993
Paraselenis(Spaethie-choma) fla-va Linnaeus
b b b Convolvulus sp.(Conv.)
Weyenberg 1874
Paraselenis(S.) solieri(Boheman)
b I. batatas P.(Conv.)
Monte 1932; Buzzi1988
Paraselenis(S.) dichroa(Germar)
b I. batatas P.(Conv.)
Monte 1932; Buzzi1988
Paraselenis(S.) tersa(Boheman)
b b b I. tiliacea (Willd.)(Conv.)
Windsor & Choe 1994
Omaspides(Omaspi-des) bistria-ta Boheman
b b I. philomega H.(Conv.)
Windsor & Choe 1994

52 THE COLEOPTERISTS BULLETIN 56(1), 2002
Table 1. Continued.
Egg Larva Pupa Host plant Reference
Omaspides(O.) con-vexicollisSpaeth b b b
I. philomega H.(Conv.) Rodriguez 1994a
Omaspides(O.) nigro-lineata(Boheman) b
I. batatas P.(Conv.) Monte 1932; Buzzi 1988
Omaspides(O.) pallidi-pennis(Boheman) b b, c1 b
Passiflora sp. Pas-sifloraceae)(von Lengerken1939); I. albaL. (Buzzi 1994)(Conv.)
Ohaus 1899–1900; CostaLima 1914, 1955; vonLengerken 1939
Omaspides(O.) trico-lorata(Boheman) b b b I. alba L. (Conv.)
Frieiro-Costa 1995; Friei-ro-Costa & Vasconcel-los-Neto 2000
Omaspides(P.) sobrina(Boheman) b, c1 b Jolivet 1988
Omaspides sp. b
O’Toole & Preston-Mafham 1985; O’Toole1995
Cyrtonotathalassina(Boheman) b
I. batatas P.(Conv.) (CostaLima, 1936)
Ohaus 1899–1900; vonLengerken 1939
CASSIDINAE: EUGENYSINIEugenysa
columbiana(Boheman) b, c1, e b
Mikania sp. (As-teraceae) Chaboo, this paper
EugenysacoscaroniViana b b, c1, e b
M. guaco Bonpl.(Aster.) Windsor & Choe 1994
of sitting on groups of eggs or larvae, and responding to intruders or to objectsintroduced by the observer. General ideas about the origins of maternal carein beetles (Brandmayr 1992) and specifically in chrysomelids (Windsor andChoe 1994) have been discussed.
Windsor and Choe (1994) also reported that males of E. coscaroni weresometimes present with the larval group but they concluded that male presencewas probably related to mate guarding. The possibility of paternal care hasarisen from an illustration that shows a male Acromis spinifex (Linnaeus) sit-ting on an egg mass (von Lengerken 1939); this image has been reproducedin the literature (Linsenmaier 1972; Jolivet 1997) thus perpetuating the ideaof male involvement in brood care (Jolivet 1997; Buzzi 1998).
This paper reports the first observations of maternal care in E. columbianafrom Costa Rica. The presence of adult males with females and larval groupsis also reported. Distributional data, a range extension to Peru, and the firsthost plant record are provided for the species. Illustrated descriptions of the

53THE COLEOPTERISTS BULLETIN 56(1), 2002
reproductive system, larvae and pupae are provided, representing the first suchdescriptions in the tribe Eugenysini. The implications of morphology and be-havior in E. columbiana are discussed.
Materials and Methods
Aggregations of E. columbiana were observed at two sites in southern CostaRica, the Las Cruces Biological Station and near the Rio Penas Blancas, duringJune 2000. Specimens were collected and reared, under license number A-002-00 from the Ministerio del Ambiente y Energia (MINAE). Voucher adult andlarval specimens of E. columbiana have been deposited in the collections ofCUIC and INBio; dissected vouchers are deposited in the AMNH collection.
Insect identifications were made by comparisons with previously identifiedspecimens from the collections listed below but types were not examined. Plantidentification was by L. D. Gomez (Wilson Botanical Garden). Cassidine spe-cies names follow Borowiec (1999), and plant species names are in accordancewith the Index Kewensis (1997). Larval age was determined by counting headcapsules in the exuvio-fecal shield. Locality data are stated as on specimenlabels.
Genitalic study in this species involved both mature and teneral adults thatwere recently killed and preserved in alcohol. Specimens were dissected indistilled water; potassium hydroxide (KOH) was avoided as it almost instantlydestroys the spermathecal muscles and, very quickly thereafter, the softer partsof the female reproductive system. Comparative genitalic studies involved old-er museum specimens of nine additional eugenysine species: A. caedemadens(Lichtenstein), A. guianiensis, A. connectens (Baly), A. parellina (Boheman),E. colossa (Boheman), E. coscaroni, E. grossa (Linnaeus), E. martae Boro-wiec, E. unicolor Borowiec and Dabrowska, and M. gentilis (Erichson). Sper-mathecal terminology follows Samuelson (1966).
Specimen examinations and illustrations were made with a Wild M5A mi-croscope and a Zeiss compound microscope, both with camera lucida attach-ments. The aedeagus, spermatheca, larvae and pupae were photographed witha Nikon D1 Digital camera with an Infinity K2 lens attachment and Micropticsfiber optic flash unit. Measurements were taken with a digital stage micro-meter; length was measured along the dorsal midline, and width across themetanotum.
The following collections and individuals provided specimens for this study:AMNH, American Museum of Natural History, L. Herman and S. Lodhi;BMNH, British Museum of Natural History, M. Brendell and S. Shute; CSCC,Caroline S. Chaboo Collection; CUIC, Cornell University Insect Collection,J. K. Liebherr and R. Hoebeke; INBio, Instituto Nacional de Biodiversidad,A. Solıs; MCZC, Museum of Comparative Zoology Collection, P. Perkins;MMUE, University of Manchester Collection, C. Johnson and P. Rispin; UCR,University of Costa Rica, P. Hanson.
Field Observations
On June 6, 2000, in the vicinity of the Rio Penas Blancas in southern CostaRica, I found two cassidines species, Stolas lebasii (Boheman) (Cassidinae:Stolaini) (one adult) and E. columbiana, on a woody vine that was wrappedaround a small tree. Leaves of the vine were well-chewed, with large holesalong margins and within the leaf surface (Fig. 1). The vine was later identifiedas a Mikania sp. (Asteraceae).

54 THE COLEOPTERISTS BULLETIN 56(1), 2002
Fig. 1. Leaf of Mikania sp. showing feeding damage by E. columbiana.
Fig. 2. Adult of E. columbiana attending pupal group on Mikania sp. in Costa Rica.
The specimens of E. columbiana consisted of 6 tightly-clustered larvalbroods (16 to 111 individuals) and 12 adults. There were two egg clusters, allwith the upper portions broken, probably from hatching. Each cluster of oval-shaped whitish eggs encircled the stem, and individual eggs were attached tothe stem at one end; this arrangement corresponds to the egg mass of E.coscaroni (Windsor et al. 1992). Larvae fell into two distinct size classes, andeach group consisted of only one size class. No solitary larvae were observedoutside groups or actively feeding.
Six adults were in mating pairs that remained in copula for the entire periodof observation. In addition there were two singletons, and two pairs of adultsassociated with the two largest larval broods. In one group, one adult sat atoplarvae at one end while the other adult sat at the opposite end with its forelegson larvae. In the second group, each adult sat opposite each other on the vine,both with the posterior part of their bodies covering larvae.
At one point, the sole S. lebasii individual stepped on the elytra of an adultE. columbiana, spent about 10 min touching with the antenna and legs, andeventually moved on. Later it walked over one larval group and was quicklydislodged into the bushes below by the repeated and rapid flexing movementsof exuvio-fecal shield. Three muscoid flies landed on one larval group butwere unable to settle because individual larvae, along with their neighbors,quickly flexed their shields. I disturbed the vine by first lightly shaking andthen by more violent shaking. Larvae responded with the reflex abdominalmovements (a behavior called cycloalexy), single adults dropped-off into thebushes (this behavior is called thanatosis and is the usual reflex of cassidineswhen disturbed). Adults attending larvae did not move.

55THE COLEOPTERISTS BULLETIN 56(1), 2002
Two larval groups were collected, one untended group of smaller-sized lar-vae (111 4th instar individuals), and one group of larger-sized larvae (42 5th
instar individuals) along with its two attendant adults. When handled, bothadults and larvae exuded an orange-brown liquid with a faint odor from themouth. The adults were not obviously sexually dimorphic, however later dis-sections revealed them to be male and female.
On June 11, 2000 at the Las Cruces Biological Station I observed anothergroup of E. columbiana, a single adult on top of a group of 32 pupae (Fig.2). When lightly touched with a pencil the pupae flexed their bodies, and theadult moved about but remained with the group.
Rearing experiments. Eight large-sized larvae were maintained in a Tup-perware y container with a moist cotton ball for one week. They continuedeating Mikania leaves thus confirming that the vine is a host plant. In thecontainer, larvae spread out for feeding but regrouped when not active. Fecalshields were quickly flexed over the abdomen whenever I brought a forcepswithin a few cm of the group. These specimens died before molting into pupae.
Three pupae were collected and maintained in a Tupperwarey container,with a piece of the vine and a moist cotton ball. Like pupae in the field, theseindividuals responded to light touch with a reflex jerking motion during ninedays of observation, then were unresponsive for two days prior to adults eclo-sion. Newly-emerged adults were very mobile.
Descriptions
Eugenysa columbiana (Boheman)
Calaspidea columbiana Boheman, 1850:212 (original description); Boheman,1856:35 (catalog); Boheman, 1862:91 (catalog); Gemminger and Harold,1876:3629 (list); Wagener, 1877:53 (list); Spaeth, 1905:91 (description);Spaeth, 1914:31 (catalog); Spaeth, 1937:85 (description).
Calaspidea columbina: Champion, 1893:135 (localities; misspelling); Boro-wiec, 1999:79 (cited as misspelling).
Eugenysa columbiana: Blackwelder, 1946:737 (checklist); Viana, 1968:21 (de-scription); Wilcox, 1975:150 (checklist); Windsor and Choe, 1994:113(distribution); Borowiec, 1996:184 (list); Borowiec, 1999:79 (catalog).
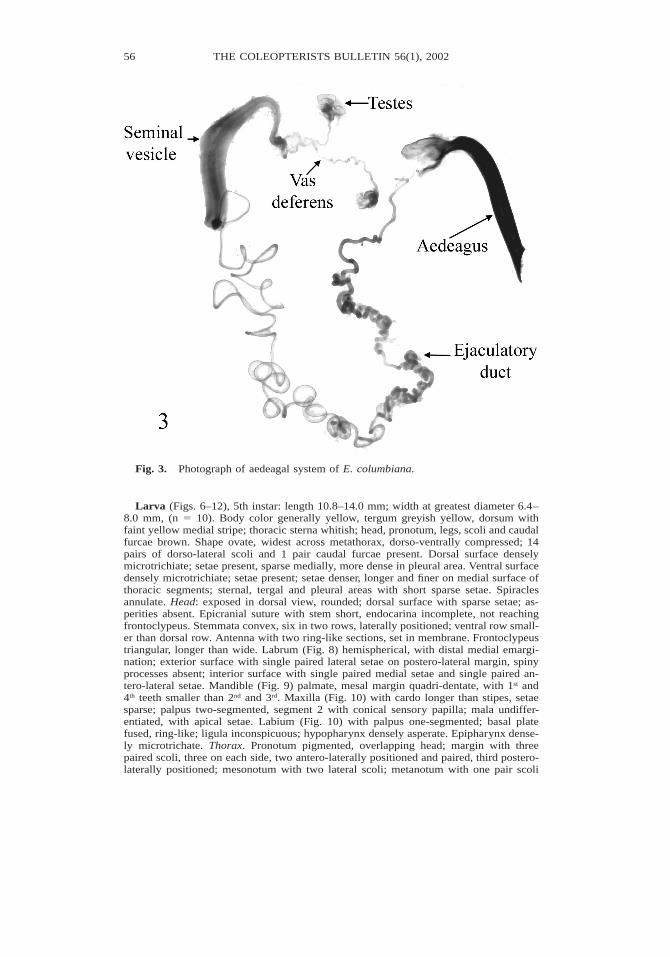
Description. Male genitalia (Fig. 3). Median lobe forming 908 angle with basal piece;ejaculatory guides present; tegmen with manubrium (5 basal piece) as long as arms (5lateral lobes); lateral plates slightly projected from rim of apical foramen. Ejaculatoryduct very long, about 8 times longer than aedeagus; junction of duct and seminal vesiclewith sclerotized protuberant knob; seminal vesicle wide distally and tapered proximally,proximal portion deflexed and tapered into vas deferens; duct coiled, coils loose proxi-mally and tight distally.
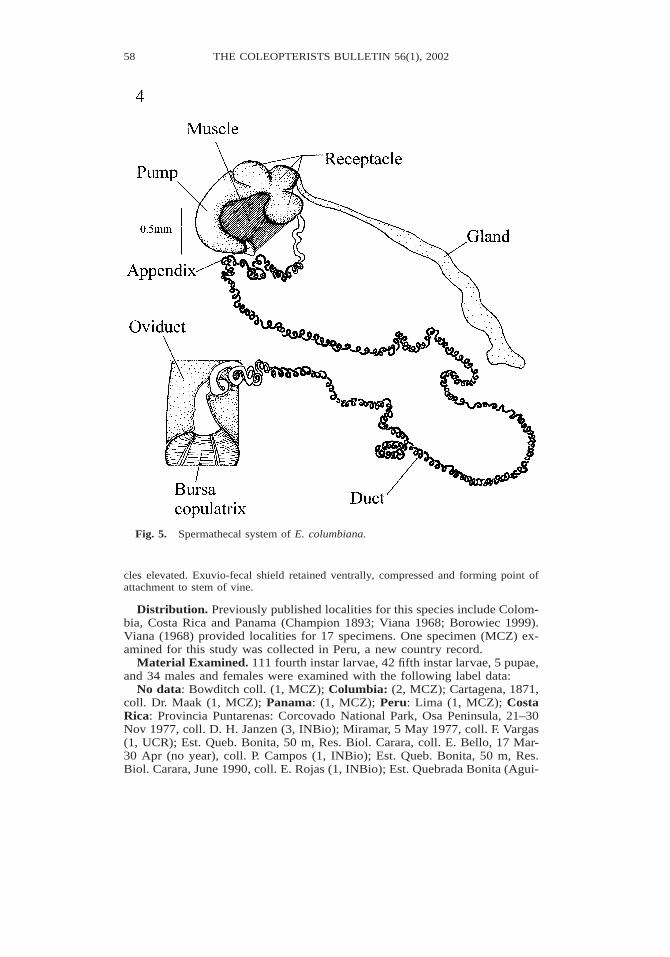
Female Genitalia Ovipositor. spiculum lightly sclerotized, with emarginate apex, andwidened distally; palpi shorter than T8. Reproductive tract. Whitish; each ovary with11–15 ovarioles; calyx, lateral oviducts and common oviduct enclosed in layer of string-like tissue; bursa copulatrix rounded; oviduct section distal to bursa copulatrix withpaired lateral lobes and short paired mesal projections; mesal projections yellowish.Spermatheca (Figs. 4–5). Well-sclerotized, falcate, with muscle on inner margin; pumpwith diameter smaller than in receptacle, deflexed and apically narrowed; appendix pre-sent. Receptacle complex, with three chambers distinguishable externally. Gland present,elongate and basally positioned; spermathecal duct and gland entry into receptacle cham-bers clearly separated, with spermathecal duct entering chamber 1 and gland enteringchamber 2. Duct very long, more than 15 times spermathecal length, entirely tightlycoiled.
56 THE COLEOPTERISTS BULLETIN 56(1), 2002
Fig. 3. Photograph of aedeagal system of E. columbiana.
Larva (Figs. 6–12), 5th instar: length 10.8–14.0 mm; width at greatest diameter 6.4–8.0 mm, (n 5 10). Body color generally yellow, tergum greyish yellow, dorsum withfaint yellow medial stripe; thoracic sterna whitish; head, pronotum, legs, scoli and caudalfurcae brown. Shape ovate, widest across metathorax, dorso-ventrally compressed; 14pairs of dorso-lateral scoli and 1 pair caudal furcae present. Dorsal surface denselymicrotrichiate; setae present, sparse medially, more dense in pleural area. Ventral surfacedensely microtrichiate; setae present; setae denser, longer and finer on medial surface ofthoracic segments; sternal, tergal and pleural areas with short sparse setae. Spiraclesannulate. Head: exposed in dorsal view, rounded; dorsal surface with sparse setae; as-perities absent. Epicranial suture with stem short, endocarina incomplete, not reachingfrontoclypeus. Stemmata convex, six in two rows, laterally positioned; ventral row small-er than dorsal row. Antenna with two ring-like sections, set in membrane. Frontoclypeustriangular, longer than wide. Labrum (Fig. 8) hemispherical, with distal medial emargi-nation; exterior surface with single paired lateral setae on postero-lateral margin, spinyprocesses absent; interior surface with single paired medial setae and single paired an-tero-lateral setae. Mandible (Fig. 9) palmate, mesal margin quadri-dentate, with 1st and4th teeth smaller than 2nd and 3rd. Maxilla (Fig. 10) with cardo longer than stipes, setaesparse; palpus two-segmented, segment 2 with conical sensory papilla; mala undiffer-entiated, with apical setae. Labium (Fig. 10) with palpus one-segmented; basal platefused, ring-like; ligula inconspicuous; hypopharynx densely asperate. Epipharynx dense-ly microtrichate. Thorax. Pronotum pigmented, overlapping head; margin with threepaired scoli, three on each side, two antero-laterally positioned and paired, third postero-laterally positioned; mesonotum with two lateral scoli; metanotum with one pair scoli

57THE COLEOPTERISTS BULLETIN 56(1), 2002
Fig. 4. Photograph of spermatheca of E. columbiana (spermathecal muscle removedby KOH).
on postero-lateral angles. Pronotal anterior scoli and mesonotal anterior scoli shortest.Spiracles annulate, prominent, projecting, without setae on inner margin. Legs (Fig. 11)four-segmented, with sparse long setation; tarsungulus curved. Abdomen. Ten-segmented;segments 1–8 with paired lateral scoli; 1–7 with paired annulate spiracles in dorsal laterallines. Lateral scoli (Fig. 12) curved, tapered; scoli on segments 1–2 anteriorly directed;scoli on segments 6–7 latero-posteriorly directed; scoli on segments 8 posteriorly directedand not visible in dorsal view. Scoli on segments 6–8 long, almost twice longer thanabdominal scoli. Caudal furca (Fig. 13) paired, long, unbranched, tapered, dorsally pro-jected; surface wrinkled, setae present, asperities absent; basal plate narrow, sclerotized.Thoracic and abdominal scoli with surface slightly wrinkled and setose; setation distinct,with basal portion long and bristle portion stiff and barb-like. Shield. Present, well-developed up to 7 mm in height, composed of feces and exuviae with four intact headcapsules.
Larva (Fig. 14), 4th instar: length 6.3–8.7 mm (n 5 10). Body color light yellow,without grey cast. Scoli number and arrangement, setation and trichiation pattern, andfecal shield with characteristics similar to 5th instar larvae. Shield (Fig. 15): present,well-developed, composed of feces and exuviae with three intact head capsules.
Pupa (Figs. 16–17), length 13.0 mm (n 5 1). Color dark brown to black, dorsumwith narrow medial cream-colored stripe extending from anterior pronotal margin to 1st
abdominal tergum. Integument smooth, devoid of setation or trichiation. Head hiddenby pronotum, not visible in dorsal view. Pronotum with well-developed explanate mar-gin, anterior edge continuous; dorsal surface feebly wrinkled, without dorsal or lateralprojections. Abdominal segments 1–3 with margins laterally extended into short lobes;other segments with rounded margins; abdominal segment 9 without urogomphi. Spira-
58 THE COLEOPTERISTS BULLETIN 56(1), 2002
Fig. 5. Spermathecal system of E. columbiana.
cles elevated. Exuvio-fecal shield retained ventrally, compressed and forming point ofattachment to stem of vine.
Distribution. Previously published localities for this species include Colom-bia, Costa Rica and Panama (Champion 1893; Viana 1968; Borowiec 1999).Viana (1968) provided localities for 17 specimens. One specimen (MCZ) ex-amined for this study was collected in Peru, a new country record.
Material Examined. 111 fourth instar larvae, 42 fifth instar larvae, 5 pupae,and 34 males and females were examined with the following label data:
No data: Bowditch coll. (1, MCZ); Columbia: (2, MCZ); Cartagena, 1871,coll. Dr. Maak (1, MCZ); Panama: (1, MCZ); Peru: Lima (1, MCZ); CostaRica: Provincia Puntarenas: Corcovado National Park, Osa Peninsula, 21–30Nov 1977, coll. D. H. Janzen (3, INBio); Miramar, 5 May 1977, coll. F. Vargas(1, UCR); Est. Queb. Bonita, 50 m, Res. Biol. Carara, coll. E. Bello, 17 Mar-30 Apr (no year), coll. P. Campos (1, INBio); Est. Queb. Bonita, 50 m, Res.Biol. Carara, June 1990, coll. E. Rojas (1, INBio); Est. Quebrada Bonita (Agui-

59THE COLEOPTERISTS BULLETIN 56(1), 2002
Figs. 6–13. Larva of E. columbiana, 5th instar. 6) Ventral view, scale bar 5 1 mm;7) Dorsal view, scale bar 5 1 mm; 8) Labrum, ventral view, scale bar 5 0.25 mm; 9)Mandible, dorsal view, scale bar 5 0.25 mm; 10) Maxilla and labium, ventral view (halfshown), scale bar 5 0.25 mm; 11) Proleg, laterosternal view, scale bar 5 0.5 mm; 12)First abdominal scoli, dorsal view, scale bar 5 0.5mm; 13) Caudal furca, posterior view,scale bar 5 0.5 mm.
rre), R. B. Carara, A. C. Pacifico Central, 100 m, Oct 1994, coll. R. Guzman(1, INBio); Est. Quebrada Bonita, R. B. Carara, 100 m, Feb 1995, coll. R.Guzman (1, INBio); La Palma Coopeunioro, Alfrededores del Albergue Unio-ro, 250 m, 13–22 Mar 1995, coll. L. Angula (2, INBio); Cerro de Oro, ACO-SA, 270 m, 14–18 Mar 1995, coll. M. Madrigal (1, INBio); 300 m, 4–5 Nov1995, coll A. Azofeifa (2, INBio); Est. Agujas, Sendero Zamia, 300 m, 26–30 Nov 1995, coll. A. Azofeifa (1, INBio); Est. Agujas, Sendero Zamia, RioAgujas, 300 m, 9–8 Mar 1996, coll. A. Azofeifa (1, INBio); Est. Rio Bonito,2.3 Km al O. del Cerro la Gamba, 110 m, 10–30 Apr 1996, coll. E. Fletes (1,INBio); San Vito, Las Cruces Biol. Stn., ex. Mikania sp., 12 June 2000, coll.C. S. Chaboo (3 pupae; CSCC); Provincia Cartago: Parque del Este, Tres Rios,11 July 1990, coll. J. Segura (1, UCR); Provincia San Jose: Parque del Este,June 1990, coll. J. Seguro (1, UCR); Rio Penas Blancas, north bank, ex. Mi-

60 THE COLEOPTERISTS BULLETIN 56(1), 2002
Figs. 14–15. Larva of E. columbiana. 14) 4th instar; 15) Exuvio-fecal shield.
Figs. 16–17. Pupa of E. columbiana. 16) Dorsal view; 17) Ventral view.

61THE COLEOPTERISTS BULLETIN 56(1), 2002
kania sp., 6.VI.2000, colls. C.S. Chaboo and H. Rowell (7 adults, 153 larvae;AMNH, CSCC, CUIC, INBio).
Remarks. The testes varied in size and texture, from soft small roundedstructures to large, stiff structures. This variation may be related to maturityof particular individuals. I was unable to determine if the endophallus surfaceis spiny or not.
The 4th instar larva is similar to the 5th instar larva in the number andarrangement of scoli, furcae, setation and trichiation. The only obvious dis-tinctions are in color and size. The shield has a pyramidal shape composed oflayers of exuvial skins. Each exuviae was folded over once, with head capsulesneatly stacked posteriorly, and the older caudal furcal integument enclosingthe more recent one. The head capsules thus provide a convenient count fordetermining the larval instar.
Discussion
These observations of E. columbiana, supported by larval and adult repro-ductive morphology, suggest a complex defense system that is consistent withreports for other chrysomelids (summarized in Jolivet 1997), and an unusualmating system. Several levels of defense are discernable: grouping of eggs,larvae and pupae, and reflex behaviors like thanatosis and cycloalexy can bedefensive (Vasconcellos-Neto and Jolivet 1989; Damman and Cappuccino1996); larval scoli and trichiation can provide a physical armor for mechanicaldefense; setation can increase detection of intruders (Eisner et al. 1967); andred and black adult coloration may provide some aposematic protection. Thefecal shield, host plant choice and maternal care suggest more sophisticateddefensive strategies.
Cassidine fecal shields have well-demonstrated defensive functions (Eisneret al. 1967; Olmstead and Denno 1992, 1993; Olmstead 1994; Gomez et al.1999; Eisner 2000), although Muller and Hilker (1999) have found contrarydata. Vencl and Morton (1999) hypothesised that in species with maternal carethe larvae have reduced or temporary fecal shields. However both E. coscaroniand E. columbiana have well-developed shields, constructed of feces and ex-uvial skins, and are obvious contradictions to this hypothesis.
Host plant choice might also provide a chemical defense strategy. Could theliquid regurgitated upon handling be a plant-derivative with repellent effects,as has been demonstrated for many chrysomelids (Pasteels et al. 1998) ormight the fecal shield incorporate plant compounds, as in other chrysomelidspecies (Gomez 1997; Pasteels et al. 1998; Vencl and Morton 1998; Gomezet al. 1999; Muller and Hilker 1999)? Interestingly, Mikania is now reportedas a host plant for five cassidine species, E. columbiana, E. coscaroni, S.lebasii, Echoma clypeata (Panzer) (5 Echoma marginata (Linneaus)), andEchoma anaglyptoides Borowiec (Windsor and Choe 1994; Windsor et al.1995). The plant recurs in the traditional folk medicines of diverse indigenouspeoples (Bishayee 1994; Lentz et al. 1998; Facey et al. 1999). Consequentsystematic explorations have revealed chemicals that are new (e.g., mikanoli-dae, dihydromikanolides, melampolides) (Ruengler et al. 2001) or rare (e.g.,sesquiterpinoids, nerolidol derivatives) (Diaz et al. 1992) and which show in-secticidal, anti-bacterial, anti-protozoal, and anti-inflammatory properties (Fie-ro 1999; Muelas et al. 2000). It is highly intriguing if these beetle species areexploiting this unusual chemistry.
Prolonged presence of males with the female and larval brood is now re-

62 THE COLEOPTERISTS BULLETIN 56(1), 2002
corded for two eugenysines, E. coscaroni (Windsor and Choe 1994) and E.columbiana. Male presence may be a function of territoriality, which wouldincrease mating opportunities, or of mate guarding which would assure pater-nity (Reid and Roitberg 1994). Paternal guarding of the larval brood is unlikelysimply because of its rarity in insects but if it does indeed occur, this wouldrepresent another level of sophistication in this species’ defense system.
Cassidine reproductive structures have been documented in several works(Spett and Lewitt 1925, 1926; Sanderson 1948; Varma 1955; Pajni et al. 1987;Bordy and Doguet 1987; Mann and Crowson 1983, 1996; Mann 1988; Verma1985, 1988, 1996). In these studies, where a few genera and species wereexamined, and in my doctoral dissertation research involving more than 300species from all cassidine tribes, the system of E. columbiana is unusual inthree aspects. First, the ejaculatory duct of this species is exceptionally longand tightly folded into numerous coils. Second, the spermathecal duct in thefemale is also exceptionally long and coiled, as predicted for such correlatedreproductive structures (Suzuki 1989). Third, the spermathecae has three re-ceptacle chambers. This discovery in E. columbiana led me to sample othereugenysines species, and it appears that the three-chambered receptacle is morewidespread within the tribe. Its function and its potential as a phylogenetically-informative character needs further exploration.
The only clues we have about the physiological mechanisms operating inthe cassidine spermatheca and the aedeagus come from the elegant studies ofV. Rodriguez (1994b, 1995) with Chelymorpha cribraria (Fabricius) (5 Chely-morpha alternans Boheman). In that species the flagellum penetrates far upthe spermathecal duct, even reaching into the spermatheca in one case (Ro-driguez 1994b; Eberhard 1996:6–7, 352). Should this mechanism apply toother cassidines, the flagellum may navigate the very long and highly coiledspermathecal duct in E. columbiana.
All the morphological and behavioral features discussed here fit with knowndata on the remarkable biology of cassidines; each aspect presents an inter-esting avenue of research. Among species exhibiting maternal care (Table 1),comparative data for particular aspects is quite patchy. Given that eight cas-sidine species have been reported as maternal care providers only in the last12 years, then it is highly probable that more species with this unusual behaviorawait discovery. In order to clarify the nature of parental involvement andother complex behaviors in E. columbiana and other cassidine species, an inter-disciplinary approach to observations and experimentation is necessary.
Acknowledgments
I thank the curators and collections for providing specimens for study. Fieldwork was funded by a Rawlins Grant from the Department of Entomology,Cornell University and a field study grant from the American Museum ofNatural History. F. Campos (OTS, San Jose), facilitated my collection andstudy permits in Costa Rica and H. Cochran (INBio) facilitated my exportpermit. I thank H. Rowell for the field excursion, J. and R. Figueroa Sandi forhelp in the field, M. Schlaeper for taking field photos and L. D. Gomez forthe plant identification and his many kindnesses during my stay at the WilsonBotanical Garden. J. Pruski (U.S. National Herbarium) provided informationon Mikania while N. Upton (Green Umbrella, nature documentary production)discussed his observations of A. sparsa. T. Nguyen and C. Johnson generouslyprovided technical help and F. Merino, D. Silva and J. Stark provided German

63THE COLEOPTERISTS BULLETIN 56(1), 2002
and Spanish translations. Discussion with C. Johnson, K. Olmstead, C. Reid,H. Rowell and N. Upton, and comments from A. Berkov, L. Herman and twoanonymous reviewers greatly improved this manuscript. These institutions andindividuals are gratefully acknowledged for their support of my research.
Literature Cited
Baly, J. S. 1869. Characters of some undescribed species of Phytophaga belonging tothe families Cassididae and Hispidae. Transactions of the Entomological Societyof London 1869:83–90.
Bishayee, A., and M. Chatterjee. 1994. Anticarcinogenic biological response of Mi-kania cordata: reflections in hepatic biotransformation systems. Cancer letters.81(2):193–200.
Blackwelder, R. E. 1946. Checklist of the coleopterous insects of Mexico, CentralAmerica, The West Indies, and South America. Part 4. U.S. National Museum,Bulletin 185. Smithsonian Institution, Washington, D.C. 763 pp.
Boheman, C. H. 1850. Monographia Cassididarum. Holmiae. Vol. 1, 452 pp.Boheman, C. H. 1854. Monographia Cassididarum. Holmiae. Vol. 2, 506 pp.Boheman, C. H. 1856. Catalogue of coleopterous insects in the collection of the British
Museum. Part 9. Cassididae. British Museum, London. 225 pp.Boheman, C. H. 1862. Monographia Cassididarum. Holmiae. Vol. 4, 504 pp.Bondar, G. 1930. Insetos daninhos e moletias da batata doce no Brasil. O Campo, Rio
de Janeiro, 1(11):33–36.Bontems, C. 1988. Localization of spermatozoa inside viviparous and oviparous females
of Chrysomelinae (pp. 299–316). In: Biology of the Chrysomelidae (P. Jolivet,E. Petitpierre, and T. H. Hsiao, editors). Kluwer Academic Publisher, Dordrecht.615 pp.
Bordy, B., and S. Doguet. 1987. Contribution a la connaissance des Cassidinae deFrance Etude de leur spermatheque (Coleoptera: Chrysomelidae). Nouvelle RevueEntomologie (N.S.) 4(2):161–176.
Borowiec, L. 1987. A new species of Eugenysa Chevrolat from Colombia (Coleoptera:Chrysomelidae: Cassidinae). Polskie Pismo Entomologiczne 56:883–885.
Borowiec, L. 1996. Faunistic records of Neotropical Cassidinae (Coleoptera: Chryso-melidae). Polskie Pismo Entomologiczne 65:119–251.
Borowiec, L. 1999. A world catalogue of the Cassidinae (Coleoptera: Chrysomelidae).Biologica Silesiae, Wroclaw. 476 pp.
Borowiec, L., and A. Dabrowska. 1997. Two new species of Eugenysa Chevrolat, 1837from Ecuador and Peru (Coleoptera: Chrysomelidae: Cassidinae). Genus 8:673–678.
Brandmayr, P. 1992. Short review of the presocial evolution in Coleoptera. EthologyEcology and Evolution. Special Issue 2:7–16.
Buzzi, Z. J. 1980. Estagios imaturos de Acromis nebulosa (Boheman, 1854) (Coleoptera:Chrysomelidae: Cassidinae). Dusenia 12(2):63–67.
Buzzi, Z. J. 1988. Biology of neotropical Cassidinae (pp. 559–580). In: Biology ofChrysomelidae (P. H. Jolivet, M. L. Cox and E. Petitpierre, editors). Kluwer Ac-ademic Publishers, Dordrecht. 615 pp.
Champion, G. C. 1893. Fam. Cassididae. Biologia Centrali-Americana, Coleoptera, Vol.VI, part 2:125–232.
Chevrolat, L. A. A. 1837. In: Dejean, P. F. M. A., Catalogue des coleopteres de lacollection de M. le comte Dejean. Ed. 3, revue, corrigee et augmentee. Paris, livr.5:385–503.
Choe, J. C. 1989. Maternal care in Labidomera suturella Chevrolat (Coleoptera: Chryso-melidae: Chrysomelinae) from Costa Rica. Psyche 96:63–67.
Costa Lima, A. da. 1914. Nota relativa ao cassidideo Omoplata pallidipennis (Dejean).Memorias do Instituto Oswaldo Cruz 6:112–145.
Costa Lima, A. M. 1936. Terceiro catalogo dos insetos que vivem nas plantas do Brasilseus parasitos e predadores. Rio de Janeiro. 460 pp.

64 THE COLEOPTERISTS BULLETIN 56(1), 2002
Costa Lima, A. M. 1955. Insectos do Brasil, 9(29). Coleoptera, parte 3:221–237.Damman, H., and N. Cappuccino. 1996. Two forms of egg defence in a chrysomelid
beetle: egg clumping and excrement cover. Ecological Entomology 16:163–167.Diaz, J. G., V. L. Goedken, and W. Herz. 1992. Sesquiterpene lactones and other
constituents of Mikania rimachii and Mikania microptera. Phytochemistry 31:597–608.
Eberhard, W. G. 1996. Female control: sexual selection by cryptic female choice.Monographs in Behavior and Ecology. Princeton University Press, Princeton. 502pp.
Eisner, T., E. van Tassel, and J. E. Carrel. 1967. Defensive use of a ‘fecal shield’ bya beetle larva. Science 158:1471–1473.
Erichson, G. F. 1847. Conspectus Insectorum Coleopterorum, quae in Republica Perua-na observata sunt, auctore. Archiv fur Naturgeschichte 13(2):67–185.
Fabricius, J. C. 1775. Systema Entomologiae, sistens Insectorum classes, ordines, gen-era, species, adiectis synonymis, locis, descriptionibus, observationibus. Flens-burgi et Lipsiae. 832 pp.
Facey, C. P., K. O. Pascoe, R. B. Porter, and A. D. Jones. Investigation of plantsused in Jamaican folk medicine for anti-bacterial activity. Journal of Pharmacyand Pharmocology 51:1455–1460.
Fiebrig, K. 1910. Cassiden und Cryptocephaliden Paraguays. Ihre Entwicklungsstadienund Schutzvorrichtungen. Zoologisches Jahrbuch, Supplement 12, Heft 2:161–264.
Fiero, I. M., A. C. B. da Silva, L. C. da Silva, R. S. de Moura, and F. C. Barja.1999. Studies on the anti-allergic activity of Mikania glomerata. Journal of Eth-nopharmacology 66(1):19–24.
Frieiro-Costa, F. 1995. Biologia de populacoes e etologia de Omaspides tricolotata(Boheman, 1854) (Coleoptera: Chrysomelidae: Cassidinae) na Serra do Japi, Jun-diaı-SP. Ph.D. Thesis, Universidade Estadual de Campinas, Sao Paulo, Brasil.
Frieiro-Costa, F. A., and J. Vasconcellos-Neto. 2000. Biological and ecological studieson Omaspides tricolorata (Coleoptera: Chrysomelidae: Cassidinae). (pp. 247).Abstract Books. XXI International Congress of Entomology, Brazil, 1047 pp.
Gemminger, M., and E. von Harold. 1876. Catalogus coleopterum hucusque descrip-torum synonymicus et systematicus. Chrysomelidae (II), Languriidae, Erotylidae,Endomychidae, Coccinellidae, Corylophidae, Platypsyllide, Acredit Index Gene-rum universalis. 12:3479–3822. Monachii: E.H. Gummi, Paris and London.
Goidanich, A. 1956. Gregarismi od individualismi larvali e cure materne nei crisomelidi(Col., Chrysomelidae). Memorie della societa entomologica italiana 35:151–183.
Gomez, N. E. 1997. The fecal shields of larva of tortoise beetles (Cassidinae: Chryso-melidae): a role in chemical defense using plant-derived secondary compounds.Ph.D. Dissertation, Technischen Universitat Carolo-Wilhemina zu Braunschweig.124 pp.
Gomez, N. E., L. Witte, and T. Hartmann. 1999. Chemical defense in larval tortoisebeetles: essential oil composition of fecal shields of Eurypedus nigrosignata andfoliage of its host plant, Cordia curassavica. Journal of Chemical Ecology 25(5):1007–1027.
Index Kewensis. 1997. CD-ROM, Version 2.0. Oxford University Press, Royal BotanicGardens, Kew.
Jolivet, P. 1988. Les soins parentaux chez les Chrysomelides. L’Entomologiste 44(2):93–101.
Jolivet, P. 1997. Biologie des coleopteres chrysomelides. Societe novelles des editionboubee, Paris. 279 pp.
Kudo, S., and E. Ishibashi. 1995. Notes on maternal care in the ovoviviparous leafbeetle Gonioctena japonica (Coleoptera: Chrysomelidae). Canadian Entomologist127:275–276.
Kudo, S., E. Ishibashi, and S. Makino. 1995. Reproductive and subsocial behaviourin the ovoviviparous leaf beetle Gonioctena sibirica (Coleoptera: Chrysomelidae).Ecological Entomology 20:367–373.

65THE COLEOPTERISTS BULLETIN 56(1), 2002
Lawrence, J. F. 1982. Coleoptera. In: Synopses and classification of living organismsS. P. Parker, editor. McGraw Hill, New York. Vol. 2:482–553.
Lentz, D. L., A. M. Clark, C. D. Hufford, B. Meurer-Grimes, C. M. Passreiter, J.Cordero, O. Ibrahim, and A. L. Okunade. 1998. Antimicrobial properties ofHonduran medicinal plants. Journal of Ethnopharmacology 63(3):253–263.
Lichtenstein, A. A. H. 1796. Catalogus musei zoologici ditissimi Hamburgi. Section 3,Insecta 13. Schniebes, Hamburg, 224 pp.
Linnaeus, C. 1758. Systema naturae. 10th ed. Holmiae IV: 824 pp.Linnaeus, C. 1763. Amoenitates Academicae seu Dissertationes variae Physicae, Me-
dicae Botanicae antehac seorsim editae, nunc collectae et auctae cum tabulis ae-neis. Volumen sectum. Holmiae. 486 pp.
Linsenmaier, W. 1972. Insects of the world. McGraw-Hill Book Company, New York,392 pp.
Luhman, M. 1940. Beitrage zur biologie des Chrysomelidae. 7. Beobachtungen anPhytodecta rufipes Fbr. Entomologische Blatter 36:8–11.
Mann, J. S. 1988. Male genitalia of Chrysomelidae (Coleoptera). I. Subfamilies Cassi-dinae and Hispinae. Journal of Animal Physiology 35(2):123–130.
Mann, J. S., and R. A. Crowson. 1983. On the internal male reproductive organs andtheir taxonomic significance in leaf beetles (Coleoptera: Chrysomelidae). Ento-mologia Generalis 9(1–2):75–99.
Mann, J. S., and R. A. Crowson. 1996. Internal sac structure and phylogeny of Chryso-melidae (pp. 291–316). In: (P. H. A. Jolivet, and M. L. Cox, editors). Chryso-melidae biology. SPB Academic Publishing, Amsterdam. Vol. 1: 444 pp.
Monte, O. 1932. Alguns Cassidideos, pragas da batata doce. Boletim de AgriculturaZootechnica e veterinaria, Belo Horizonte 5(4):43–46.
Muelas, S. S., J. J. Nogal, D. R. A. Martinez, J. A. Escario, F. A. R. Martinez, andB. A. Gomez. 2000. In vitro screening of American plant extracts on Trypano-soma cruzi and Trichomonas vaginalis. Journal of Ethnopharmacology 71(1–2):101–107.
Muller, C., and M. Hilker. 1999. Unexpected reactions of a generalist predator towardsdefensive devices of cassidine larvae (Coleoptera, Chrysomelidae). Ocecologia118:166–172.
Ohaus, F. 1899. Bericht uber eine entomologische Reise nach Centralbrasilien. Stettinerentomologische Zeitung 60:204–245.
Ohaus, F. 1900. Bericht uber eine entomologische Reise nach Centralbrasilien. Stettinerentomologische Zeitung 61(3–4):164–191.
Olmstead, K. L. 1994. Waste products as chrysomelid defenses (pp. 311–318). In: Novelaspects of the biology of Chrysomelidae (P. H. Jolivet, M. L. Cox, and E. Petit-pierre, editors). Kluwer Academic Publishers, Dordrecht. 582 pp.
Olmstead, K. L., and R. F. Denno. 1992. Cost of shield defence for tortoise beetles(Coleoptera: Chrysomelidae). Ecological Entomology 17:237–243.
Olmstead, K. L., and R. F. Denno. 1993. Effectiveness of tortoise beetle larval shieldsagainst different predator species. Ecology 74:1394–1409.
O’Toole, C. 1995. Alien Empire: an exploration of the lives of insects. Harper CollinsPublishers, New York. 224 pp.
O’Toole, C., and K. Preston-Mafham. 1985. Insects in Camera: a photographic essayon behaviour. Oxford University Press, Oxford. 154 pp.
Pajni, H. R., S. S. Gandhi, and S. R. Singla. 1987. Studies on the ovipositor of someindian Chrysomelidae (Coleoptera). Research Bulletin (Science) of the PanjabUniversity 38(1–2):87–96.
Panzer, G. W. 1798. Voets beschreibungen und abbildungen hartschaaligter insecten,Coleoptera Linn. Aus dem original getreu ubersetzt, mit der in selbigem fehlendensynonymie und bestandigem commentar versehen von D. Georg Wolfgang FranzPanzer. Vierter Theil, Erlangen. 112 pp.
Pasteels, J. M., J. C. Braekman, and D. Daloze. 1998. Chemical defense in Chryso-melidae (pp. 233–252). In: Biology of Chrysomelidae (P. H. Jolivet, M. L. Cox,and E. Petitpierre, editors). Kluwer Academic Publishers, Dordrecht. 615 pp.

66 THE COLEOPTERISTS BULLETIN 56(1), 2002
Preston-Mafham, R., and K. 1993. Encyclopedia of land invertebrate behaviour. MITPress, Cambridge. 320 pp.
Reid, M. L., and B. D. Roitberg. 1994. Benefits of prolonged male residence withmates and brood in pine engravers (Coleoptera: Scolytidae). Oikos 70(1):140–148.
Rodriguez, V. 1994a. Sexual behavior in Omaspides convexicollis Spaeth and O. bis-triata Boheman, with notes on maternal care of eggs and young. ColeopteristsBulletin 48:140–144.
Rodriguez, V. 1994b. Function of the spermathecal muscle in Chelymorpha alternansBoheman (Coleoptera: Chrysomelidae: Cassidinae). Physiological Ecology 19:198–202.
Rodriguez, V. 1995. Relation of flagellum length to reproductive success in male Chely-morpha alternans Boheman. Coleopterists Bulletin 49:201–205.
Ruenglar, P., V. Brecht, C. G. Tamayo, and I. Merfort. 2001. Germacranolides fromMikania guaco. Phytochemistry 56:475–489.
Sanderson, M. W. 1948. Larval, pupal, and adult stages of North American Physonota(Chrysomelidae). Annals of the Entomological Society of America 41:468–477.
Samuelson, G. A. 1966. Alticinae of New Guinea II. Amphimeloides (Coleoptera:Chrysomelidae). Pacific Insects 8:403–445.
Seeno, T. N., and J. A. Wilcox. 1982. Leaf beetle genera. Entomography 1:1–221.Spaeth, F. 1905. Beschreibung neuer Cassididen nebst synonymischen Bemerkungen.
Verhandlungen der kaiserlich-koniglichen zoologisch-botanischen Gesellschaft inWien 55:79–118.
Spaeth, F. 1914. Chrysomelidae, 16. Cassidinae (116 pp.). In: Coleopterum Catalogusauspiciis et auxilio (S. Schenkling, editor). W. Junk, Berlin, Pars. 62.
Spaeth, F. 1937. Mitteilungen uber Cassidinen mit besonderer Berucksichtigung derSammlung des Naturkundemuseums der Stadt Stett. (Col.). Stettiner Entomolo-gische Zeitung 98:79–95.
Spett, G., and M. Lewitt. 1925. Zur kenntnis des inneren Geschlechtsapparates derBlattkafer (Coleoptera: Chrysomelidae). Wiener Entomologische Zeitung 42(1–3):39–44.
Spett, G., and M. Lewitt. 1926. Versuch einer verwertung des receptaculum seminisals systematisches merkmal bei den Chrysomeliden. Archiv fur naturgeschichteAb. A(6):96–142.
Suzuki, K. 1989. Character correlation in the internal reproductive systems of the familyChrysomelidae. Entomography 6:511–518.
Vasconcellos-Neto, J., and P. Jolivet. 1989. Ring defense strategy (cycloalexy) amongbrazilian chrysomelid larvae. Entomography 6:347–354.
Vencl, F. V., and T. C. Morton. 1999. Macroevolutionary aspects of larval shielddefences (pp. 217–238). In: Advances in Chrysomelidae biology (M. L. Cox,editor). Backhuys Publishers, Leiden. Vol. 1: 671 pp.
Varma, B. K. 1955. Taxonomic value of spermathecal capsules as subfamily charactersamong the Chrysomelidae (Coleoptera). Indian Journal of Entomology 17(1):189–192.
Verma, K. K. 1985. Male reproductive organs as taxonomic characters for a broadclassification of Chrysomelidae. Journal of Natural History 9:249–255.
Verma, K. K. 1988. Male reproductive organs as taxonomic characters for a broadclassification of Chrysomelidae. Entomography 3:485–487.
Verma, K. K. 1996. Inter-subfamily relations among Chrysomelidae (Coleoptera) assuggested by organization of the male genital system (pp. 317–351). In: Chryso-melidae biology (P. H. A. Jolivet and M. L. Cox, editors). SPB Academic Pub-lishing, Amsterdam. Vol. 1: 444 pp.
Viana, M. J. 1968. Revision sistematica de las ‘‘Eugenysini’’ Spaeth con nuevas especiesy catalogo bibliografico de la tribus (Coleopt., Chrysomelidae, Cassidinae). Re-vista del museo argentino de ciencias naturales ‘‘Bernardino Rivadavia’’ 3(1):1–106.

67THE COLEOPTERISTS BULLETIN 56(1), 2002
von Lengerken, H. 1939. Die brutfursorge- und brutpflegeinstinkte der kafer. Akade-mische verlagsgesellschaft. Geest and Portig, Leipzig. 386 pp.
Wagener, B. 1877. Cassididae. Mitteilungen der Munchener Entomologischen Vereins1:49–79.
Weise, J. 1899. Einige neue Cassidinen-Gattungen und Arten. Archiv fur Naturgeschich-te 65(1):268–273.
Weyenbergh, H. 1874. Histoire attendrissante de l’amour maternal de ‘Omoplata flavaL.’ Periodico Zoolojico 1:47–52.
Wilcox, J. A. 1975. Family 129. Chrysomelidae. In: Checklist of the beetles of Northand Central America and the West Indies, North America Beetle Fauna Series (R.H. Arnett, Jr., editor). Flora and Fauna Publications, Gainesville. 166 pp.
Windsor, D. M. 1987. Natural history of a subsocial tortoise beetle, Acromis sparsaBoheman (Chrysomelidae, Cassidinae) in Panama. Psyche 94(1–2):127–150.
Windsor, D. M., and J. C. Choe. 1994. Origins of parental care in chrysomelid beetles(pp. 111–117). In: Novel aspects of the biology of Chrysomelidae (P. H. Jolivet,M. L. Cox, and E. Petipierre, editors). Kluwer Academic Publishers, Dordrecht.582 pp.
Windsor, D. M., E. G. Riley, and H. P. Stockwell. 1992. An introduction to the biologyand systematics of Panamanian tortoise beetles (Coleoptera: Chrysomelidae: Cas-sidinae) (pp. 372–391). In: Insects of Panama and Mesoamerica: selected studies(D. Quintero and A. Aiello, editors). Oxford University Press, Oxford. 692 pp.
Windsor, D. M., M. Valverde de Macedo, and A. de Toledo Siquiera-Campos. 1995.Flower feeding by species of Echoma Chevrolat (Coleoptera: Chrysomelidae: Cas-sidinae) on Mikania (Asteraceae) in Panama and Brazil. Coleoperists Bulletin 49:101–108.
(Received 1 September 2000; accepted 28 May 2001)




















