
Keys for the identification of families and genera of Atlantic shallow water ascidians

INTRODUCTION
The spermathecae of leaf beetles in the subfamilyCassidinae have been little studied. The opinion thattheir structure is uniform and do not offer diagnostic orphylogenetic characters, has been prevailing for manyyears. Recent studies on spermathecae of some speciesin the tribe Stolaini have indicated a large diversity ofspermathecal structure (Rodriguez 1994a, b; Borowiecand Skuza 2004; D. Windsor letter inf.). Some positiveresults were also obtained in usage of structure of sper-matheca to taxonomic studies in the genus Cassida L.on both species and subgenus level (Bordy and Doguet1987; Borowiec and Świętojańska 2001). These positiveresults stimulated the senior author to compare thespermathecae of various taxa within the subfamily.
This paper contains the results of a study on thestructure of spermathecae within selected genera ofthe tribes Eugenysini and Stolaini which are character-ized by subsocial behaviour, especially maternal care.The study also includes taxa characterized by variousdegree of sexual dimorphism from absent to the high-est observed in the subfamily. The following questionsmotivated our studys:
– are sexual dimorphism and spermathecal com-plexity correlated?
– is spermathecal structure constant within partic-ular genera and subgenera?
– is the structure of the spermatheca diagnostic forspecies?
– how variable is the structure of the spermathecawithin species?
THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI
(COLEOPTERA: CHRYSOMELIDAE: CASSIDINAE) AND ITS TAXONOMIC SIGNIFICANCE
AA NN NN AA LL EE SS ZZ OO OO LL OO GG II CC II ((WWaarrsszzaawwaa)),, 22000077,, 5577((33)):: 446633--447799
LECH BOROWIEC1 and SABINA OPALIŃSKA
Department of Biodiversity and Evolutionary Taxonomy, University of Wrocław,Przybyszewskiego 63/77, Wrocław, Poland; 1e-mail: [email protected]
Abstract.— Spermathecae of 43 nominal and one unidentified species of Cassidinaebelonging to the tribes Eugenysini (two genera) and Stolaini (four genera) have beenstudied and figured. Spermathecae of particular genera are characterized by a set ofconstant and distinct characters and in many cases they are also diagnostic for species. Nocorrelation was observed between the degree of sexual dimorphism within the species andthe complexity of spermathecal structure. The distinctive structure of the spermatheca inthe monotypical subgenus Pseudomoplata Spaeth, 1952 of the genus Echoma Chevrolat,1837 suggests that the taxon should be excluded from the genus Echoma. The spermathecaof Paraselenis marginipennis (Spaeth, 1907), the only member of the subgenusPseudechoma Spaeth, 1913 of the genus Paraselenis Spaeth, 1913, is more similar to thespermathecae of some members of the genus Omaspides Chevrolat, 1837 than to any othermember of Paraselenis and Pseudechoma and should probably be transferred to thegenus Omaspides.
Key words.— morphology, spermathecae, Coleoptera, Chrysomelidae, Cassidinae, Stolaini,Eugenysini, Neotropics.

MATHERIAL AND METHODS
We examined spermathecae taken from 54 speci-mens of tortoise beetles belonging to 43 species andfour genera of the tribes Stolaini (Acromis Chevrolat,1837; Echoma Chevrolat, 1837; Omaspides Chevrolat,1837; Paraselenis Spaeth, 1913) and Eugenysini (Eu-genysa Chevrolat, 1837; Agenysa Spaeth, 1905). Altho-ugh Borowiec (1995) synonymized Eugenysini, Stolainiand Omocerini under the group name Mesomphaliini weused tribal names Eugenysini and Stolaini in old sensebecause some recent molecular and morphological phy-logenetic analyses do not support a monophyletic Sto-laini (Hsiao and Windsor 1999; Chaboo 2005).
All specimens were borrowed from the collection of: DBET – Department of Biodiversity and Evolutionary
Taxonomy, University of Wrocław, Poland;IRSN – Institut Royal des Sciences Naturelles, Brux-
elles, Belgium.
List of examined specimens with label data is giv-en below:
1. AAccrroommiiss ssppaarrssaa (Boheman, 1854): “Coll. R. I. Sc.N. B, Brazil, amz coll. P. J. Roelofs”, [coll. IRSN];
2. AAccrroommiiss ssppaarrssaa (Boheman, 1854): “Coll. R. I. Sc.N. B, Brazil, Teffe (Ega), Amazones, M de Mathan,1 er Trimestre 1879”, [coll. IRSN];
3. AAccrroommiixx ssppiinniiffeexx (Linnaeus, 1763): “Coll. R. I. Sc.N. B, Paraguay: Asuncion, XII-1936, Le Moult ven-dit”, [coll. IRSN];
4. Accrroommiixx ssppiinniiffeexx (Linnaeus, 1763): “Coll. R. I. Sc.N. B, Paraguay: Asuncion, XII-1936, Le Moult ven-dit ”, [coll. IRSN];
5. AAggeennyyssaa ccaaeeddeemmaaddeennss (Lichteinstein, 1795):“Guyane française, Bas Maroni, Le Moult vendit“,[coll. DBET];
6. AAggeennyyssaa ccoonnnneecctteennss (Baly, 1869): “Peru: Reg.Iquitos, Mishuyaey, I-1937, Le Moult vendit”, [coll.DBET];
7. AAggeennyyssaa ccoonnnneecctteennss (Baly, 1869): “Equador,Napo, Tena, Alto Cindi, 400 m, z, JUN 1996, Ameri-no”, [coll. DBET];
8. AAggeennyyssaa ccrraassssiiccoorrnniiss Spaeth, 1905: “Equador,Napo, Santa Clara, 800 m, A. Jasiński”, [coll. DBET];
9. AAggeennyyssaa gguuiiaanniieennssiiss (Boheman, 1856): “Ve-nezuela”; “66/4/8”; “Klages coll’n, Exot Coleopt.,CMA r 2278”, [coll. DBET];
10. AAggeennyyssaa ppeerruuvviiaannaa Spaeth, 1915: “Peru, Chan-chamayo, 1967”, [coll. DBET];
11. EEcchhoommaa (s.str.) aannaaggllyyppttooiiddeess Borowiec, 1998:“Decembre”; “Guyane française, Nouveau Chant-ier, Le Moult vendit”, [coll. DBET];
12. EEcchhoommaa (s.str.) bbiippllaaggoossaa (Boheman, 1854):“Coll. R. I. Sc. N. B, Brazil, Rondonia, s. Breuningvendit”, [coll. DBET];
13. EEcchhoommaa (s.str.) bboonnffiillssii (Boheman, 1854): “Ecua-dor, Pichincha, Puerto Quito, 28-V-1983, Lg. Er-nesto Martinez”, [coll. DBET];
14. EEcchhoommaa (s.str.) ccllyyppeeaattaa (Panzer, 1798): “Coll. R.I. Sc. N. B, Guyane française, Pabiacabo, Riviére deKourou, Le Moult vendit, JUIN”, [coll. IRSN];
15. EEcchhoommaa (s.str.) ccllyyppeeaattaa (Panzer, 1798): “Coll. R.I. Sc. N. B, Guyane française, Juillet, Guyane fran-çaise, Pabiacabo, Riviére de Kourou, coll. Le Moult,Juillet”, [coll. IRSN];
16. EEcchhoommaa (s.str.) ccllyyppeeaattaa (Panzer, 1798): “Omo-plata flavipennis”, [coll. DBET];
17. EEcchhoommaa (PPsseeuuddoommooppllaattaa) wweeyyeennbbeerrgghhii (Dohrn,1878): “Argentine, Prov. Tucuman“; “illegible“;”weyenbgi, +, D., det. Spaeth”, [coll. DBET];
18. EEuuggeennyyssaa ggrroossssaa (Fabricius, 1798): “Coll. R. I. Sc.N. B, Guyane française, Cayenne, Coll. Madon”,[coll. IRSN];
19. EEuuggeennyyssaa vveennoossaa (Fabricius, 1798): “Coll. R. I. Sc.N. B, Guyane française, Cayenne, Coll. Madon”,[coll. IRSN];
20. OOmmaassppiiddeess (s.str.) aabbbbrreevviiaattaa Baly, 1872: “Peru,Chanchamayo, 1967”, [coll. DBET];
21. OOmmaassppiiddeess (s.str.) aauugguussttaa Boheman, 1856:“Peru, Chanchamayo, Hoffmanns”; “Omaspides au-gusta”, [coll. DBET];
22. OOmmaassppiiddeess (s.str.) bbaassiilliiccaa Boheman, 1856:“Brazil”, [coll. DBET];
23. OOmmaassppiiddeess ((PPaarroommaassppiiddeess)) bbrruunnnneeoossiiggnnaattaa(Boheman, 1854): “ Brazil, Minas, R. Pére David,coll. P. Jésuites”, [coll. DBET];
24. OOmmaassppiiddeess (s.str.) ccllaatthhrraattaa (Linnaeus, 1758):“Coll. R. I. Sc. N. B, Guyane française, Bas Maroni,Le Moult vendit”, [coll. IRSN];
25. OOmmaassppiiddeess (s.str.) ccllaatthhrraattaa (Linnaeus, 1758):“Coll. R. I. Sc. N. B, Guyane française, Bas Maroni,Le Moult vendit”, [coll. IRNS];
26. OOmmaassppiiddeess (s.str.) nniittiiddiiccoolllliiss Spaeth, 1937:“Coll. R. I. Sc. N. B, Columbien (Fusagasuga), E.Pehlke S.”; “Paraty., nitidi – collis., Spaeth det.”,[coll. DBET];
27. OOmmaassppiiddeess (s.str.) ppaalllliiddiippeennnniiss (Boheman,1854): “Coll. R. I. Sc. N. B, Brazil: Sta Catarina,Itapiranga, IX-1956, S. Breuning vendit”, [coll.IRSN];
28. OOmmaassppiiddeess (s.str.) ppaalllliiddiippeennnniiss (Boheman,1854): “Coll. R. I. Sc. N. B, Brazil: Sta Catarina, Itapi-ranga, X-1952, S. Breuning vendit”, [coll. IRSN];
29. OOmmaassppiiddeess (s.str.) sspp.. pprrooppee ppaalllliiddiippeennnniiss (Bo-heman, 1854): “Coll. R. I. Sc. N. B, Brazil, Minas, R.Pére David, coll. P. Jésuites”, [coll. IRSN];
30. OOmmaassppiiddeess (s.str.) ppuullcchheellllaa Baly, 1859: “S.Paulo de Olivenca, Brazil, S. Klages”, [coll. DBET];
31. OOmmaassppiiddeess (PPaarreecchhoommaa) sseemmiilliinneeaattaa (Bohe-man, 1854): “Taguara, Brazil, Acc. No. 2906”, [coll.DBET];
464 L. BOROWIEC and S. OPALIŃSKA

32. OOmmaassppiiddeess (PPaarroommaassppiiddeess) ssoobbrriinnaa (Boheman,1854): “Itatiaya, Ohaüs, 26”, [coll. DBET];
33. OOmmaassppiiddeess (PPaarroommaassppiiddeess) ssqquuaalliiddaa (Bohe-man, 1854): ”Bocaina, leg. Kessel, +”, [coll. DBET];
34. OOmmaassppiiddeess (s.str.) ttrriicchhrrooaa (Boheman, 1854):“Rio de Janeiro, Coll. Keâel”, [coll. DBET];
35. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) aauulliiccaa (Bohe-man, 1854): “Sud Brazil: Mendes, 90 Km de Rio Jan,Le Moult vendit”, [coll. DBET];
36. PPaarraasseelleenniiss (s.str.) aaxxiillllaarriiss (Sahlberg, 1823):“Rio Jan”, [coll. DBET];
37. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) ddeecciippiieennss (Bohe-man, 1854): “Brazil, Minas, R. Pére David, coll. P.Jésuites ”, [coll. DBET];
38. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) ddiicchhrrooaa (Germar,1824): “Argentina: Territoire des Missions, coll. E.Wagner, Le Moult vendit”, [coll. DBET];
39. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) ffiilliiaa (Spaeth,1907): “Paraguay: Rio Paraguay, San Antonio, XI-1936, Le Moult vendit”, [coll. DBET];
40. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) ffllaavvaa (Linnaeus,1758): “Hungarian Soil- Zool. Exp., Paraguay: VillaHayes, 50 km N from Asuncion, 1.I.1966”, [coll.DBET];
41. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) hhyyaalliinnaa (Bohe-man, 1854): ”Brazil, M. Berl.”, [coll. DBET];
42. PPaarraasseelleenniiss (PPsseeuuddeecchhoommaa) mmaarrggiinniippeennnniiss(Spaeth, 1907): “Archidona (Equad.) R., HaenschS.”, [coll. DBET];
43. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) nniiggrrooppuunnccttaattaa(Borowiec, 2003): “Patatype, des. L. Borowiec”;“Bolivia, C. Tello, Caranavi, prov. La Paz, dp., T., 2001”; “Paraselenis (Spaeth.) nigropunctata n.sp., Paratypus, des. L. Borowiec 2003”, [coll.DBET];
44. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) nnoorrmmaalliiss (Ger-mar, 1824): ”Coll. R. I. Sc. N. B, Brazil, Rio Janeiro,coll. P. Madon, Le Moult vendit”, [coll. IRSN];
45. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) nnuuppttaa (Boheman,1854): “Coll. R. I. Sc. N. B, Brazil, Teffe (Ega), Ama-zones, M de Mathan, 1 er Trimestre 1879”, [coll.IRSN];
46. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) nnuuppttaa (Boheman,1854): “Coll. R. I. Sc. N. B, Brazil, Teffe (Ega), Ama-zones, M de Mathan, 1 er Trimestre 1879 ”, [coll.IRSN];
47. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) nnuuppttaa (Boheman,1854): “Santarem., June, 1919., S. M. Klages., Acc.6324 “, [coll. DBET];
48. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) ppuunnccttiiccoolllliiss(Spaeth, 1907): “Bahia”; “Echoma puncticollis”,[coll. DBET];
49. PPaarraasseelleenniiss ssaallttaaeennssiiss Borowiec, 2002: “Coll. R.I. Sc. N. B, Argentina: Rio Salado, Chaco de Santia-go del Estero- coll. E. Wagner”, [coll. IRSN];
50. PPaarraasseelleenniiss ssaallttaaeennssiiss Borowiec, 2002: “Coll. R.
I. Sc. N. B, Argentina: Rio Salado, Chaco de Santia-go del Estero- coll. E. Wagner”, [coll. IRSN];
51. PPaarraasseelleenniiss ssaallttaaeennssiiss Borowiec, 2002: “Coll. R.I. Sc. N. B, Argentina: Rio Salado, Chaco de Santia-go del Estero- coll. E. Wagner”, [coll. IRSN];
52. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) ssccaappuulloossaa (Bohe-man, 1854): “Paraguay, leg. Fiebrig”, [coll. DBET];
53. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) ssoolliieerrii (Bohe-man, 1854): “Guyane française, les roches deKourou, Le Moult vendit “, [coll. DBET];
54. PPaarraasseelleenniiss (SSppaaeetthhiieecchhoommaa) tteerrssaa (Bohe-man, 1854): ”Panama: Canal Zone, Achiote, Road, 10 km SW Gatun, 12 June, 76:, E. G. Riley”, [coll.DBET].
Dried specimens were placed in a wet chamber for24 hours before preparation; then the abdomen wasremoved and placed in 10% KOH solution for the next24 hours. After maceration abdomina were placed indistilled water for 30 minutes. The spermathecae wasremoved from the bursa copulatrix under a stereomi-croscope and placed on a microscope slides in a drop ofdistilled water. Figures were made using Nikon EclipseE 600 microscope equipped with a drawing tube under10× or 20× magnification. After examination sperma-thecae and abdomina were attached to a white cardusing a drop of glue and then pinned under the speci-men.
Spermathecal character names were adopted fromthe classical paper by Rodriguez (1994 a). Generalstructure of spermatheca in Cassidinae were present-ed in previous paper on spermatheca of the genusChelymorpha Chevr. (Borowiec and Skuza 2004). Thealternative character states are given for each charac-ter (in parenthesis abbreviations used in tables):
– length of vasculum without velum;– shape of vasculum:
a) hook-shaped (h), b) saccus-shaped (s), c) kidney-shaped (k);
– velum:a) present, b) absent;
– shape of ampulla:a) elongate (e), b) bubble-shaped (b), c) weakly dif-
ferentiated from vasculum, feebly separate (fs), d) absent;
– length of ampulla/length of vasculum ratio (A/V ratio):a) below 1/3 (< 1/3), b) approximately 1/3 (1/3),
c) between 1/2 and 1/3 (> 1/3 <1/2), d) approx-imately 1/2 (1/2), e) above 1/2 (>1/2);
– spiralization of ductus:a) febble, b) strong, c) absent;
– number of windings:a) absent, b) below 50 (< 50), c) above 50 (> 50);
– length of ductus;– length of ductus/length of vasculum ratio (D/V ratio).
THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI 465

RESULTS
Spermathecal characters for particular genera aresummarized as follows:
Genus EEuuggeennyyssaa(Table 1)
Vasculum kidney-shaped, length 1.9–2.3 mm, velumabsent; ampulla bubble-shaped not longer than 1/3length of vasculum, ductus strongly spiralized withmore than 50 windings and length 5.8–6.1 mm, D/Vratio 2.6–3.3:1.
Genus AAggeennyyssaa(Table 2)
Vasculum saccus-shaped, length 1.3–1.4 mm, velumpresent; ampulla weakly differentiated from vasculum,ductus strongly spiralized with more than 50 windingsand length 3.0–4.8 mm, D/V ratio 3:1–3.5:1.
Genus AAccrroommiiss(Table 3)
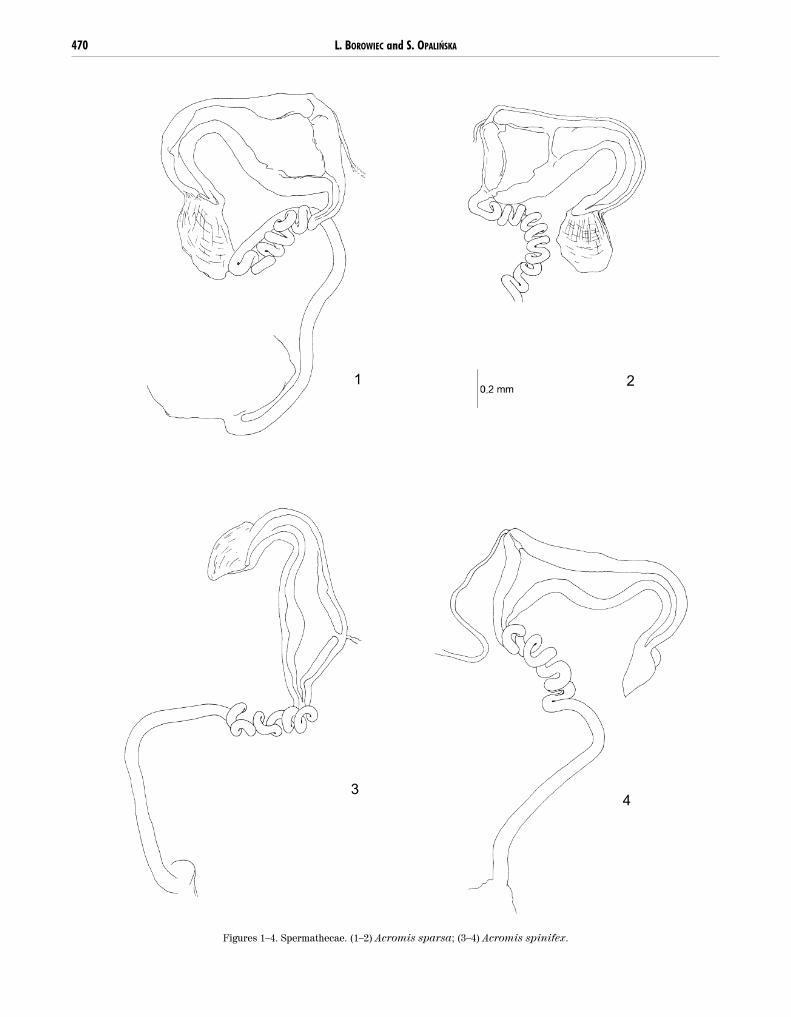
Vasculum saccus- or hook-shaped, length 1.2–1.4mm, velum present; ampulla weakly differentiated,ductus feebly spiralized with less than 50 windings andlength 1.9–2.6mm, D/V ratio 1.6:1–1.9:1.
Genus EEcchhoommaa(Table 4)
Vasculum kidney-shaped (except one species withhook-shaped), length 0.7–1.05 mm, velum absent(except one species); ampulla elongate, A/V ratio to 1/2,ductus more or less spiralized but usually with lessthan 50 windings and length 1.2–5.7 mm, D/V ratio vari-able from 1:1 to 6:1.
Genus OOmmaassppiiddeess group 1 of nominotypical subgenus(Table 5)
Vasculum hook-shaped, length 1.3–1.5 mm, velumpresent; ampulla elongate, longer than half length ofvasculum, ductus not or very feebly spiralized withlength 0.92–2.47 mm, D/V ratio 0.6–2:1.
Genus OOmmaassppiiddeess group 2 of nominotypical subgenus and subgeus PPaarreecchhoommaa
(Table 6)
Vasculum hook-shaped (in one species saccus-shaped), length 1.2–1.5 mm, velum present; ampullaelongate (in one species absent), not longer than 1/3length of vasculum (except one species), ductus feeblyspiralized with less than 50 windings and length 2.0–3.2mm, D/V ratio 2.0–2.5:1.
Genus OOmmaassppiiddeess subgenus PPaarroommaassppiiddeess(Table 7)
Vasculum small, hook-shaped, length 0.8–1.0 mm,velum present; ampulla weakly differentiated, notlonger than 1/3 length of vasculum, ductus feebly spi-ralized with less than 50 windings and length 1.0–1.9mm, D/V ratio 1.1–2.0:1.
Genus PPaarraasseelleenniiss(Table 8)
Vasculum small, hook-shaped, length 1.0–1.3 mm,velum present; ampulla elongate or weakly differentiat-ed, usually not longer than 1/3 length of vasculum, duc-tus feeble spiralized with less than 50 windings andlength 1.0–2.2 mm, D/V ratio 1.1–2.0:1.
DISCUSSION
The spermathecae in the analysed genera ofEugenysini and Stolaini are diversely shaped and rep-resent several morphological groups correlating inpart with external characters. Thus, spermathecaeappear to offer many characters of both diagnostic andphylogenetic value. Within species the structure of thespermatheca is quite constant, with variability notedmainly in the compactness of the spiral of ductus, thenumber of windings of the ampulla and the ductus, andthe spatial position of the ampulla in relation to thevasculum. We did not observe any correlation betweenthe degree of sexual dimorphism in adults and the com-plexity of spermathecal structure. In genera containingtaxa with both indistinct and very distinct sexual di-morphism (Paraselenis, Omaspides), spermathecaewere similarly shaped although some differences were
466 L. BOROWIEC and S. OPALIŃSKA
Table 1. Characteristics of spermathecae in the genus Eugenysa (no. of specimen corresponding with no. in list of examined specimens).
Vasculum Ampulla Ductus
No. Species Length (mm) Shape Velum A/V ratio Shape Spiral. Number of Length (mm) D/V ratiowindings
18. E. grossa 1.97 k absent < 1/3 b strong < 50 6.1 3:119. E. venosa 2.26 k absent 1/3 b strong > 50 5.8 3:1

THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI 467
Table 2. Characteristics of spermathecae in the genus Agenysa (no. of specimen corresponding with no. in list of examined specimens).
Vasculum Ampulla Ductus
No. Species Length (mm) Shape Velum A/V ratio Shape Spiral. Number of Length (mm) D/V ratiowindings
5. A. caedemad. 1.15 s present < 1/3 fs strong > 50 4.78 4:16. A. connect. 1.18 s present < 1/3 fs strong > 50 3.71 4:17. A. connect. 1.13 s present < 1/3 fs strong > 50 3.20 3:18. A. crassicor. 1.42 s present < 1/3 fs strong > 50 – –9. A. guianensis 1.24 s present < 1/3 fs strong > 50 3.03 3:1
10. A. peruviana 1.30 s present < 1/3 fs – – – –
Table 3. Characteristics of spermathecae in the genus Acromis (no. of specimen corresponding with no. in list of examined specimens).
Vasculum Ampulla Ductus
No. Species Length (mm) Shape Velum A/V ratio Shape Spiral. Number of Length (mm) D/V ratiowindings
1. A. sparsa 1.38 s present – absent feeble < 50 2.60 2:12. A. sparsa 1.38 s present – absent – – – –3. A.spinifex 1.24 h present – absent feeble < 50 1.95 1:14. A. spinifex 1.45 h present – absent feeble < 50 1.97 1:1
Table 4. Characteristics of spermathecae in the genus Echoma (no. of specimen corresponding with no. in list of examined specimens).
Vasculum Ampulla Ductus
No. Species Length (mm) Shape Velum A/V ratio Shape Spiral. Number of Length (mm) D/V ratiowindings
11. E. anaglypto. 0.93 k absent 1/2 e feeble < 50 1.55 2:112. E. biplagosa 0.83 k absent < 1/3 e strong < 50 3.10 3:113. E. bonfilsi 0.89 k absent < 1/3 e strong < 50 5,75 6:114. E. clypeata 0.90 k absent < 1/3 e strong – – –15. E. clypeata 0.72 k absent > 1/3 < 1/2 e strong > 50 4.79 5:116. E. clypeata 0.75 k absent < 1/3 e strong < 50 4.76 5:117. E. weyenber. 1.05 h present > 1/3 < 1/2 e strong < 50 1.29 1:1
Table 5. Characteristics of spermathecae in the subgenus Omaspides s. str., subgroup 1 (no. of specimen corresponding with no. in list of examined specimens).
Vasculum Ampulla Ductus
No. Species Length (mm) Shape Velum A/V ratio Shape Spiral. Number of Length (mm) D/V ratiowindings
21. O. augusta 1.20 h present < 1/3 e feeble < 50 2.47 2:122. O. basilica 1.42 h present > 1/2 e absent 0 1.10 1:126. O. nitidicollis 1.38 h present > 1/2 e absent 0 1.11 1:130. O. pulchella 1.55 h present > 1/2 e absent 0 0.92 1:1

468 L. BOROWIEC and S. OPALIŃSKA
Table 6. Characteristics of spermathecae in the genus Omaspides, subgroups 2–4 (no. of specimen corresponding with no. in list of examined specimens).
Vasculum Ampulla Ductus
No. Species Length (mm) Shape Velum A/V ratio Shape Spiral. Number of Length (mm) D/V ratiowindings
20. O. abbreviata 1.24 h present 1/3 e feeble < 50 2.58 2:124. O. clathrata 1.21 h present 1/3 e feeble < 50 2.91 3:125. O. clathrata 1.48 h present < 1/3 e feeble < 50 3.25 3:131. O. semilineata 1.90 s present – absent feeble < 50 2.00 1:127. O. pallidipennis 0.93 h present < 1/3 fs feeble < 50 1,90 2:128. O. pallidipennis 0.80 h present < 1/3 fs feeble – – –29. O. pallidipennis 0.89 h present < 1/3 fs feeble < 50 1,00 1:134. O. trichroa 1.05 h present < 1/3 fs feeble < 50 1,33 1:1
Table 7. Characteristics of spermathecae in the subgenus Paromaspides (no. of specimen corresponding with no. in list of examined specmens).
Vasculum Ampulla Ductus
No. Species Length (mm) Shape Velum A/V ratio Shape Spiral. Number of Length (mm) D/V ratiowindings
23. O. brunneosig. 0.85 h present > 1/3 < 1/2 e/fs feeble < 50 – –32. O. sobrina 0.88 h present < 1/3 e/fs feeble < 50 1.45 2:133. O. squalida 0.75 h present > 1/3 < 1/2 e/fs feeble < 50 – –
Table 8. Characteristics of spermathecae in the genus Paraselenis (no. of specimen corresponding with no. in list of examined specimens).
Vasculum Ampulla Ductus
No. Species Length (mm) Shape Velum A/V ratio Shape Spiral. Number of Length (mm) D/V ratiowindings
35. P. aulica 1.22 h present < 1/3 e/fs feeble < 50 1.75 1:136. P. axillaris 1.10 h present < 1/3 e/fs feeble < 50 1.20 1:137. P. decipiens 0.73 h present < 1/3 e/fs feeble < 50 1.98 2:138. P. dichroa 1.00 h present < 1/3 e feeble < 50 1.30 1:139. P. filia 0.78 h present > 1/3 < 1/2 e feeble < 50 1.22 1:140. P. flava 1.20 h present < 1/3 e feeble < 50 1.56 1:141. P. hyalina 1.09 h present < 1/3 e feeble < 50 1.00 1:143. P. nigropunctata 0.90 h present < 1/3 e/fs feeble < 50 1.01 1:144. P. normalis 1.03 h present < 1/3 e/fs feeble < 50 1.74 2:145. P. nupta 1.12 h present 1/3 e – – – –46. P. nupta 1.16 h present < 1/3 e feeble < 50 2.25 2:147. P. nupta 0.92 h present 1/3 e feeble < 50 2.05 2:148. P. puncticollis 1.10 h present > 1/3 < 1/2 e feeble < 50 2.03 2:149. P. saltaensis 1.05 h present < 1/3 e/fs feeble < 50 1.60 1:150. P. saltaensis 1.24 h present < 1/3 e feeble < 50 1.63 1:151. P. saltaensis 1.18 h present < 1/3 e/fs feeble < 50 1,56 1:152. P. scapulosa 1.31 h present < 1/3 e feeble < 50 1.76 1:153. P. solieri 1.02 h present > 1/3 < 1/2 e feeble < 50 1.86 2:154. P. tersa 1.10 h present 1/3 e feeble < 50 1.26 1:142. P. marginipen. 1.11 h present > 1/2 e feeble < 50 2.25 2:1

observed between taxa classified in distinct subgenera(see discussion below). On the other hand, spermathe-cae of superficially similar species are often very simi-lar and their diagnostic value is of little use. The char-acteristics of spermathecae in particular genera are asfollows:
Tribe Eugenysini
Genus Agenysa (Figs 5–10): spermathecae simpleand varying little. In all species vasculum saccus-shaped with velum present, ampulla very small, ductusvery thin, long and strongly spiraled but windingsslightly loose. Spermathecal characteristics of speciesare too small to use as diagnostic characters.
Genus Eugenysa (Figs 18, 19): the genus morpho-logically is very similar to Agenysa but spermathecaeare completely distinct, vasculum is kidney-shapedwithout velum, ampulla large, bubble-shaped, ductusas long as in Agenysa but thicker and more loosely spi-raled.
Generally, spermathecae of both studied genera aredistinctive but are of little value in diagnosing species.Spermathecae in the genus Agenysa are more similarto the spermathecae of specialized genera of the tribeCassidini (our unpublished data) while the generalstructure of spermathecae of the genus Eugenysa aresimilar to spermathecae of some genera of the tribeStolaini. The unusual features of spermatheca of thegenus Eugenysa were also observed by Chaboo (2002)for Eugenysa columbiana (Boheman, 1850) and itappears that especially large bubble-shaped ampulla iswidespread within the genus.
Tribe Stolaini
Genus Acromis (Figs 1–4): spermathecae of bothstudied species and spermatheca of Acromis venosaErichson, 1847 figured by Chaboo (2001) are morpho-logically constant and distinct from other genera of thetribe. They are also characteristic for species and canbe use as diagnostic characters although species of thegenus Acromis are well distinguished by externalcharacters.
Genus Echoma: in nominotypical subgenus (Figs11–16) spermathecae are quite similarly shaped butwith diagnostic characters (shape of vasculum andampulla, length and degree of spiralization of ductus).The spermatheca of Echoma weyenberghi, a memberof the subgenus Pseudomoplata Spaeth, 1952, is completely distinct (Fig. 17). It is also distinct fromspermathecae of other studied genera, suggesting thatthe taxon should be excluded from the genus Echomaas a distinct monotypic genus.
Genus Omaspides (Figs 20–27, 29–32, 52–54):large and heterogenous genus Omaspides is dividedinto three subgenera: Omaspides s. str., ParechomaSpaeth, 1913, and Paromaspides Spaeth, 1913. Firsttwo subgenera are characterized by the absence ofsexual dimorphism, third subgenus is distinctly dimor-phic. Spermathecae within this genus are very diverse.Consequently, we have grouped taxa of this genus intofour subgroups. The first one contains four species ofnominotypical subgenus with ductus without or withonly a few windings (Figs 20–23). The second subgroupcontains two species of nominotypical subgenus andOmaspides semilineata of the subgenus Pare-choma. All are characterized by ductus divided intotwo distinct parts: basal with windings and distal notspiralized (Figs 24–27). Members of the nominotypicalsubgenus have vasculum hook-shaped, while the onlymember of the subgenus Parechoma has it saccus-shaped. The different vasculum shape in O. semilin-eata confirms its distinct subgeneric placement. Thethird subgroup contains three species of the nomino-typical subgenus. At first glance their spermathecaeare similar to the second subgroup but the vasculum isbroader basally and ampulla distinctly marked (Figs29–32). The fourth subgroup contains only members ofthe subgenus Paromaspides. Their spermathecae arewell characterized by small size (it is correlated withthe size of beetles; the subgenus comprises the small-est species of the genus Omaspides), well markedampulla, and spiralized part of ductus distinctly longerthan simple distal part (Figs 52–54). These character-istics amply confirm the distinct subgeneric placementof Paromaspides.
Generally, spermathecae in the genus Omaspidesare the most diverse in all studied genera, they charac-terize subgenera well, and they can be diagnostic formany species. Only in this genus are spermathecae inspecies without sexual diorphism more diverse than inspecies with sexual dimorphism. However, the signifi-cance of this difference may be affected by the smallnumber of taxa compared: 9 in the group withoutdimorphism and only three in the group with dimor-phism.
Genus Paraselenis (Figs 28, 33–51): the genus isdivided into three subgenera. Spermathecae of thenominotypical subgenus and the most speciose sub-genus, Spaethiechoma Hincks, 1952 are similar andquite uniform (Figs 33–51). Differences between spe-cies are not distinct and, due to infraspecific variation,spermathecal characters are not diagnostic. The mostdistinctive spermatheca is that of Paraselenis mar-ginipennis (Spaeth, 1907), the only member of thesubgenus Pseudechoma Spaeth, 1913. Its spermathe-ca is more similar to those of the second subgroup ofthe genus Omaspides than to any member of thegenus Paraselenis (Fig. 28). Externally Paraselenis
THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI 469
470 L. BOROWIEC and S. OPALIŃSKA
Figures 1–4. Spermathecae. (1–2) Acromis sparsa; (3–4) Acromis spinifex.
1
43
2

THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI 471
5 10 9
8 7
6
Figures 5–10. Spermathecae. (5) Agenysa caedemadens; (6–7) Agenysa connectens; (8) Agenysa crassicornis; (9) Agenysa guianiensis; (10) Agenysa peruviana.

472 L. BOROWIEC and S. OPALIŃSKA
Figures 11–17. Spermathecae. (11) Echoma anaglyptoides; (12) Echoma biplagosa; (13) Echoma bonfilsi; (14–16) Echoma clypeata; (17) Echoma weyenberghi.
1617
12
13
15 1114

THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI 473
18
19
Figures 18–19. Spermathecae. (18) Eugenysa grossa; (19) Eugenysa venosa.

474 L. BOROWIEC and S. OPALIŃSKA
Figures 20–23. Spermathecae: (20) Omaspides augusta; (21) Omaspides basilica; (22) Omaspides nitidicollis; (23) Omaspides pulchella.
20
2122
23

THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI 475
24 2528
27 26
Figures 24–28. Spermathecae. (24) Omaspides abbreviata; (25–26) Omaspides clathrata; (27) Omaspides semilineata; (28) Paraselenismarginipennis.

476 L. BOROWIEC and S. OPALIŃSKA
29 30
31 32
Figures 29–32. Spermathecae. (29–30) Omaspides pallidipennis; (31) Omaspides sp. prope pallidipennis; (32) Omaspides trichroa.

THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI 477
33 3735 36
43 3934 38
42 4140
Figures 33–43. Spermathecae. (33) Paraselenis axillaris; (34) Paraselenis decipiens; (35) Paraselenis filia; (36) Paraselenis flava; (37) Paraselenis hyalina; (38) Paraselenis nigropunctata; (39) Paraselenis nupta; (40) Paraselenis puncticollis; (41) Paraselenis
scapulosa; (42) Paraselenis solieri; (43) Paraselenis tersa.

478 L. BOROWIEC and S. OPALIŃSKA
49 45 44 51
4850 46 47
Figures 44–51. Spermathecae. (44) Paraselenis aulica; (45) Paraselenis dichroa; (46) Paraselenis normalis; (47–48) Paraselenis nupta; (49–51) Paraselenis saltaensis.

THE STRUCTURE OF THE SPERMATHECAE OF SELECTED GENERA OF STOLAINI AND EUGENYSINI 479
52 53 54
Figures 52–54. Spermathecae. (52) Omaspides brunneosignata; (53) Omaspides sobrina; (54) Omaspides squalida.
marginipennis is also similar to the members of thenominotypical subgenus of Omaspides and differsonly in dimorphic structure of antennae, while inOmaspides s. str., antennae are not dimorphic, but thedimorphism is not as strong as in most species of thegenus Paraselenis. In our opinion the subgenusPseudechoma should be transferred from Parasele-nis to Omaspides.
ACKNOWLEDGEMENTS
We would like to express our sincere thanks toanonymous reviewers for correction of English andcritical comments of the manuscript. The project wassponsored by Institute of Zoology, University of Wro-cław, grant no. 1018/DS./2006.
REFERENCES
Bordy, B. and S. Doguet. 1987. Contribution à la connaissancedes Cassidinae de France. Etude de leur spermatheque(Coleoptera, Chrysomelidae). Nouvelle Revue d’Entomolo-gie (N.S.), 4: 161–176.
Borowiec, L. 1995. Tribal classification of the cassidoid His-pinae (Coleoptera: Chrysomelidae). In: J. Pakaluk, S. A.Ślipiński, Biology, Phylogeny, and Classification ofColeoptera: Papers Celebrating the 80th Birthday of RoyA. Crowson, Warszawa, 541–558.
Borowiec, L. and M. Skuza. 2004. The structure of spermathe-ca in the genus Chelymorpha Chevrolat, 1837 (Coleo-ptera: Chrysomelidae: Cassidinae) and its taxonomic sig-nificance. Annales Zoologici, 54(2): 439–451.
Borowiec, L. and J. Świętojańska. 2001. Revision of Cassidalitigiosa group from southern Africa (Coleoptera: Chry-somelidae: Cassidinae). Annales Zoologici, 51(1): 153–184.
Chaboo, C. S. 2001. Revision and phylogenetic analysis ofAcromis Chevrolat (Coleoptera: Chrysomelidae: Cassidi-nae: Eugenysini). Coleoptersts Bulletin, 56: 50–67.
Chaboo, C. S. 2002. First report of immatures, genitalia andmaternal care in Eugenysa columbiana (Boheman)(Coleoptera: Chrysomelidae: Cassidinae: Eugenysini).Coleopterists Bulletin, 56: 75–102.
Chaboo, C. S. 2005. Biology and phylogeny of the CassidinaeGyllenhal sensu lato (tortoise and leaf-mining beetles)(Coleoptera: Chrysomelidae). Ph.D. Dissertation, CornellUniversity, 497 pp.
Hsiao, T. H. and D. M. Windsor. 1999. Historical and biologicalrelationships among Hispinae inferred from 12S MTDNAsequence data. In: M. L. Cox, Advances in ChrysomelidaeBiology 1, Backhuys Publishers, Leiden, 39–50.
Rodriguez, V. 1994a. Function of the spermathecal muscle inChelymorpha alternans Boheman (Coleoptera: Chry-somelidae; Cassidinae). Physiological Entomology (1994),19: 198–202.
Rodriguez, V., 1994 b. Sexual behavior in Omaspides convex-icollis Spaeth and O. bistriata Boheman (Coleoptera:Chrysomelidae: Cassidinae), with notes on maternal careof eggs and young. Coleopterists Bulletin, 48: 140–144.
Received: February 20, 2007Accepted: May 21, 2007
Copyright © 2022 FDOKUMEN




















