
Two new species of Cassida Linnaeus, 1758 (Coleoptera: Chrysomelidae: Cassidinae) from Madagascar...
12
Accepted by G. Morse: 24 Jul. 2007; published: 17 Sept. 2007 47 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2007 · Magnolia Press Zootaxa 1586: 47–58 (2007) www.mapress.com/ zootaxa/ Two new species of Cassida Linnaeus, 1758 (Coleoptera: Chrysomelidae: Cassidinae) from Madagascar and notes on subgenera of the genus Cassida LECH BOROWIEC Department of Biodiversity and Evolutionary Taxonomy, Zoological Institute, University of Wroclaw, Przybyszewskiego 63/77, 51-148 Wroclaw, Poland. E-mail: [email protected] Abstract Cassida butterwecki sp. nov. and Cassida morondaviana sp. nov. are described from Madagascar. Both are unique spe- cies and have no close relatives in Madagascar or tropical Africa. Subgeneric division of the genus Cassida Linnaeus, 1758 is discussed. Key words: Coleoptera, Chrysomelidae, Cassidinae, Cassidini, Cassida, new species, status of subgenera, Madagascar Introduction Cassida Linnaeus, 1758 is the most speciose genus within the subfamily Cassidinae containing until now 419 nominal species. Excepting one native species from North America all have been described from the Old World, especially from the tropics and subtropics of Africa and Asia. Generally, the number of species decreases from west to the east. Africa including Madagascar and the western Palaearctic regions are richer in species than the Australian and eastern Palaearctic regions. The fauna of Madagascar is very interesting and rich with 71 described species, while in the whole of Africa south of the Sahara only 83 species occur (Borow- iec, 1999; Borowiec and wi toja ska, 2002). Only one species, Cassida dorsovittata Boh., is common in both continental Africa and Madagascar. Madagascan species represent several partially distinct groups, with no close relatives in other regions. Descriptions are given below for two new species recently studied possess- ing unique characters and unlike any Madagascan or African species groups. The study of African and Mada- gascan species showed that the diversity of the genus Cassida is very high and the subgenera proposed for Palaearctic and Oriental species are difficult to apply to many Afrotropical taxa. Both new species from Madagascar are difficult to place in any proposed subgenera. The situation has led me to review and discuss all subgeneric names proposed in the genus Cassida. Colour photos were prepared using Syncroscopy Auto-Montage Essentials software. Photos were digi- tally processed for better appearance. Measurements were taken with an ocular micrometer. Body length was measured from the anterior corner of the pronotum to the apex. Pronotal length was taken from the anterior corner to the base of the pronotum, and pronotal width was measured as the distance between the basal cor- ners. Length ratio of antennal segments was measured as a percent of length length of each segment to the length of the first segment. Male genitalia were examined but they are not diagnostic within the genus Cassida and were not figured. Spermathecae in the genus Cassida are partly diagnostic (Bordy and Doguet, 1987; Borowiec and wi toja ska, 2001), thus they were prepared and figured (Fig. 12–19). Label data is verbatim, and data from different labels is separated by double slash (//). Each type is clearly labelled with a red label.
Transcript of Two new species of Cassida Linnaeus, 1758 (Coleoptera: Chrysomelidae: Cassidinae) from Madagascar...

Accepted by G. Morse: 24 Jul. 2007; published: 17 Sept. 2007 47
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2007 · Magnolia Press
Zootaxa 1586: 47–58 (2007) www.mapress.com/zootaxa/
Two new species of Cassida Linnaeus, 1758 (Coleoptera: Chrysomelidae: Cassidinae) from Madagascar and notes on subgenera of the genus Cassida
LECH BOROWIECDepartment of Biodiversity and Evolutionary Taxonomy, Zoological Institute, University of Wrocław, Przybyszewskiego 63/77, 51-148 Wrocław, Poland. E-mail: [email protected]
Abstract
Cassida butterwecki sp. nov. and Cassida morondaviana sp. nov. are described from Madagascar. Both are unique spe-cies and have no close relatives in Madagascar or tropical Africa. Subgeneric division of the genus Cassida Linnaeus,1758 is discussed.
Key words: Coleoptera, Chrysomelidae, Cassidinae, Cassidini, Cassida, new species, status of subgenera, Madagascar
Introduction
Cassida Linnaeus, 1758 is the most speciose genus within the subfamily Cassidinae containing until now 419nominal species. Excepting one native species from North America all have been described from the OldWorld, especially from the tropics and subtropics of Africa and Asia. Generally, the number of speciesdecreases from west to the east. Africa including Madagascar and the western Palaearctic regions are richer inspecies than the Australian and eastern Palaearctic regions. The fauna of Madagascar is very interesting andrich with 71 described species, while in the whole of Africa south of the Sahara only 83 species occur (Borow-iec, 1999; Borowiec and Świętojańska, 2002). Only one species, Cassida dorsovittata Boh., is common inboth continental Africa and Madagascar. Madagascan species represent several partially distinct groups, withno close relatives in other regions. Descriptions are given below for two new species recently studied possess-ing unique characters and unlike any Madagascan or African species groups. The study of African and Mada-gascan species showed that the diversity of the genus Cassida is very high and the subgenera proposed forPalaearctic and Oriental species are difficult to apply to many Afrotropical taxa. Both new species fromMadagascar are difficult to place in any proposed subgenera. The situation has led me to review and discussall subgeneric names proposed in the genus Cassida.
Colour photos were prepared using Syncroscopy Auto-Montage Essentials software. Photos were digi-tally processed for better appearance. Measurements were taken with an ocular micrometer. Body length wasmeasured from the anterior corner of the pronotum to the apex. Pronotal length was taken from the anteriorcorner to the base of the pronotum, and pronotal width was measured as the distance between the basal cor-ners. Length ratio of antennal segments was measured as a percent of length length of each segment to thelength of the first segment. Male genitalia were examined but they are not diagnostic within the genus Cassidaand were not figured. Spermathecae in the genus Cassida are partly diagnostic (Bordy and Doguet, 1987;Borowiec and Świętojańska, 2001), thus they were prepared and figured (Fig. 12–19).
Label data is verbatim, and data from different labels is separated by double slash (//). Each type is clearlylabelled with a red label.

BOROWIEC48 · Zootaxa 1586 © 2007 Magnolia Press
For comparison with previously described species, colour photos of most Madagascan taxa are availablein Borowiec & Świętojańska (2002).
Taxonomy
Cassida butterwecki sp. nov.
EtymologyDedicated to M. Butterweck, collector of the new species.
DiagnosisA very distinct species with no close relatives in Madagascar or tropical Africa. At first glance it is more
similar to the members of the African genus Fornicocassis Spaeth, 1917 than to other Cassida species. Thegenus Fornicocassis distinctly differs in presence of deep antennal grooves while Cassida butterwecki hasventer of pronotum typical for the genus Cassida with no antennal grooves. The strongly convex elytra, thevery strong elytral punctation, and the strongly declivous explanate margin of elytra characterize the follow-ing Madagascan species: Cassida frontalis Boheman, 1856, Cassida madagascarica Borowiec, 1999, Cassidapubescens Spaeth, 1905, and Cassida rufomicans Fairmaire, 1904. C. frontalis at first glance has similarshape except beeing slightly smaller in size than C. butterwecki but differs in the strong sculpture of elytraforming irregular folds and wrinkles. The pronotal sides of C. frontalis are regularly rounded and the prono-tum is widest at or slightly behind the middle while in C. butterwecki pronotum is widest distinctly before themiddle. The remaining three congeners form a different group of species. They are two to three times largerthan C. butterwecki, with the body almost hemispherical, slope less descending than in C. butterwecki, and thepronotal sculpture forming longitudinal striation or wrinkles. They also differ in the structure of the pronotum,with broadly rounded pronotal sides, thus their pronota are regularly elliptical while in C. butterwecki thepronotum is rather subtrapezoidal.
DescriptionLength: 4.6–5.0 mm, width: 3.5–3.8 mm, length of pronotum: 1.7–1.9 mm, width of pronotum: 2.7–3.1
mm, length/width ratio: 1.26–1.35, width/length ratio of pronotum: 1.51–1.63. Body short-oval, strongly con-vex, sides and slope strongly declining (figs. 1, 2). Sexual dimorphism indistinct.
Pronotum and elytra yellowish brown, punctures of explanate margin partly marked with black, puncturesin rows partly with complete or incomplete black areola, elevated intervals with few small black spots. Thenumber of black markings vary in the type series and some specimens look predominantly pale and some pre-dominantly spotted. Clypeus yellowish brown. Prosternal process mostly yellowish brown only elevated mar-gins partly black. Alae of prosternum from brown to black. Central plate of metasternum yellowish brownwith blackish margin. Lateral plates brown to black. Whole metasternum from dark brown to black. Abdomenin the middle dark brown and gradually paler to margins. Coxae brown to partly black, trochanters brown,femora, tibiae and tarsi yellowish. Antennal segments 1–7 yellowish, 8–11 gradually infuscate, segments 10and 11 almost black.
Pronotum subtrapezoidal, with maximum width in anterior 1/4 length. Anterior margin softly curved,anterior corners rounded then sides distinctly converging posterad. Disc convex, distinctly divided into partabove head and basal and lateral part and distinctly bordered from explanate margin by a sulcus in anteriorlobes and by an impression laterally. Area above head distinctly, coarsely but shallowly punctate, distancebetween punctures mostly narrower than puncture diameter, interspaces flat and surface of the area appearsregular. Basal and lateral part elevated, coarsely punctate, punctures almost touching each other and surfaceappears slightly irregular. Explanate margin with fine punctation, punctures arranged irregularly, in some parts

Zootaxa 1586 © 2007 Magnolia Press · 49TWO NEW CASSIDA FROM MADAGASCAR
of the margin punctures agglomerate, in other parts of margin they are in a long distance thus distancesbetween punctures vary from as wide as to many times wider than puncture diameter. Honeycomb structureinvisible. Surface of pronotum slightly dull.
FIGURES 1–3. Cassida butterwecki sp. nov.: 1. Habitus dorsal; 2. Habitus lateral; 3. Tarsus.
Scutellum triangular, without or with very shallow transverse sulcus. Base of elytra only slightly widerthan pronotum, humeri angulate, strongly protruding anterad. Disc strongly convex, sides and slope stronglydeclining and disc appears slightly cylindrical. In profile disc regularly convex with top of convexity in posts-cutellar area (Fig. 2). Punctation of disc regular, very coarse, foveolate, intervals distinctly narrower thanrows, linear. Interval 2 convex, forming a linear costa surrounding postscutellar impression and forming trans-verse wrinkle at top of disc thus H-shaped top elevation is well marked. Interval 4 also slightly convex but lessconvex than interval 2, at 3/4 length forms a slightly more elevated fold. Intervals 6 mostly flat but at 3/4length also forms a short, longitudinal fold. Marginal row distinct but its punctures finer than those of disc,marginal interval well marked only below humerus, lateral transverse fold distinct, smaller transverse foldmarked also below humerus and behind the middle of marginal row. Explanate margin strongly declining,moderately broad, in the widest part approximately as wide as 1/3 width of disc. Punctation of explanate mar-gin fine and spread irregularly thus distance between punctures vary from as wide as to many times wider thanpuncture diameter, honeycomb structure invisible. Surface of elytra appears slightly dull. Apex of elytral epi-pleura with sparse, semierect to erect hair.

BOROWIEC50 · Zootaxa 1586 © 2007 Magnolia Press
Eyes large, gena obsolete. Clypeus almost as wide as long, clypeal plate flat with several small punctures,surface distinctly microreticulate, slightly shiny. Clypeal grooves fine but distinct, clearly visible on wholelength, run close to ventral margins of eyes and converging in regular triangle (Fig. 4). Labrum emarginate to2/5 length. Prosternal collar short, prosternal process moderately broad, in the middle slightly wider thanlength of trochanters, strongly expanded apically. Basal part of the prosternal process impressed along themiddle, expanded apical part in the middle with round tubercle, at sides impressed and coarsely punctate.
FIGURES 4–5. Head: 4. Cassida butterwecki sp. nov.; 5. Cassida morondaviana sp. nov.

Zootaxa 1586 © 2007 Magnolia Press · 51TWO NEW CASSIDA FROM MADAGASCAR
Antennae slim, length ratio of antennal segments: 100:58:83:71:63:42:42:38:42:42:88. Segment 3approximately 1.4 times as long as segment 2 and approximately 1.2 times as long as segment 4, segments 9and 10 almost square. Tarsi slim, last segment extending to the apical margin of marginal setae, claws dis-tinctly extending behind marginal setae (Fig. 3). Claws simple.
TypesHolotype female: “MADAGASKAR: Morondava, Kirindy forest, II/III. 1990, leg. M. Butterweck” //
“HOLOTYPE des. L. Borowiec” // “Cassida butterwecki n. sp., HOLOTYPUS, des. L. Borowiec ‘06” (pre-served in Museum für Naturkunde, Stuttgart, Germany); three paratypes: “MADAGASKAR: Morondava,Kirindy forest, II/III. 1990, leg. M. Butterweck” // “PARATYPE des. L. Borowiec” // “Cassida butterwecki n.sp., PARATYPUS, des. L. Borowiec ‘06” (preserved in Museum für Naturkunde, Stuttgart, Germany; oneparatype at the Department of Biodiversity and Evolutionary Taxonomy, Wrocław, Poland).
Cassida morondaviana sp. nov.
EtymologyNamed after its locus typicus, Morondava city in W Madagascar.
DiagnosisA very distinct species, with no close relatives in Madagascar or tropical Africa. At first glance it looks
like members of the Palaearctic subgenus Cassidulella Strand, 1928 (type species: Cassida nobilis Linnaeus,1758), especially Cassida ovalis Spaeth, 1914 from Algeria and Corsica or Cassida vittata Villers, 1789 awide-spread in the Palaearctic Region. The similarity is also emphasized by the similar structure of the sper-matheca with a C-shaped vasculum (see figures in Bordy and Doguet, 1987: figs. 15 and 16). Palaearcticmembers differ in having a less cuneiform body, finer pronotal and elytral punctation with at least the centralelytral intervals distinctly wider than the rows, interval 3 with additional irregular punctures, distinctly coar-serly punctate clypeus with deeper clypeal sulci, stouter antennae with segments 9 and 10 subquadrate, and adistinctly longer prosternal collar with the distance between the anterior margin of the fore coxal cavity andfore margin of the collar longer than half width of a coxa. From Madagascan members of the tribe Cassidini,some small species of the genus Sphenocassis Spaeth, 1911 (type species: Laccoptera humerosa Fairmaire,1898) are at first glance similar, especially S. rotundella Borowiec, 2002 and S. punctatissima (Weise, 1910)but they distinctly differ from C. morondaviana in the completely irregular elytral punctation.
DescriptionLength: male 4.4–4.7 mm, female 5.2–5.5 mm, width: male 2.6–2.9 mm, female 2.9–3.1 mm, length of
pronotum: male 1.7–1.8 mm, female 1.85–2.0 mm, width of pronotum: male 2.35–2.5 mm, female: 2.45–2.7mm, length/width ratio: male 1.62–1.69, female 1.77–1.79, width/length ratio of pronotum: 1.33–1.41, female1.29–1.37. Body cuneiform, strongly convex, sides and slope strongly declining (figs. 6, 7). Sexual dimor-phism distinct, male smaller and stouter than female.
Whole body uniformly yellowish green, including legs, only last four antennal segments infuscate.Pronotum subtrapezoidal, with maximum width in anterior 2/5 length. Anterior margin very evenly
curved, anterior corners rounded then sides strongly converging posterad, explanate margin strongly declin-ing. Disc moderately convex, indistinctly divided into part above head and basal and lateral part and indis-tinctly bordered from explanate margin. Area above head indistinctly, finely and sparsely punctate. Basal andlateral part of disc coarsely punctate, punctures almost touching each other but surface appears regular.Explanate margin finely, shallowly, and sparsely punctate, honeycomb structure distinct. Surface of pronotummoderately shiny.

BOROWIEC52 · Zootaxa 1586 © 2007 Magnolia Press
Scutellum triangular, without sulci. Base of elytra slightly wider than pronotum, humeri angulate, stronglyprotruding anterad. Disc strongly convex, sides, slope and explanate margin strongly declining. In profile discregularly convex with top of convexity almost in mid length (Fig. 8). Punctation of disc regular, coarse, inter-vals slightly narrower than rows. Punctures in rows almost touching each other. Intervals flat, completely reg-ular. Marginal row distinct, its punctures distinctly coarser than those of disc, marginal interval very narrow,well marked only below humerus, no lateral folds. Explanate margin moderately broad, in the widest partapproximately as wide as 1/3 width of disc. Punctation of explanate margin moderately coarse and spread reg-ularly, each puncture placed in centre of eye of distinct honeycomb structure. Surface of elytra appearsslightly shiny. Apex of elytral epipleura bare.
FIGURES 6–9. Cassida morondaviana sp. nov.: 6. Male habitus dorsal; 7. Female habitus dorsal; 8. Female habitus lat-eral; 9. Tarsus.
Eyes small, gena slightly shorter than 1/3 length of eye. Clypeus 1.4 times as wide as long, clypeal plateshallowly impressed in the middle, surface shiny. Clypeal grooves fine but distinct, well visible on wholelength, run in distance from ventral margins of eyes and converging in narrow triangle (Fig. 5). Labrum shal-lowly emarginate. Prosternal collar short, distance between anterior margin of fore coxal cavity and fore mar-gin of the collar shorter than half width of coxa, prosternal process moderately broad, in the middleapproximately as wide as length of trochanters, strongly expanded apically. Surface of prosternal process flat,expanded apical part with few shallow punctures.
Zootaxa 1586 © 2007 Magnolia Press · 53TWO NEW CASSIDA FROM MADAGASCAR
Antennae slim, length ratio of antennal segments: 100:55:64:50:45:41:55:45:50:55:105. Segment 3approximately 1.2 times as long as segment 2 and approximately 1.3 times as long as segment 4, segments 9and 10 elongate, 1.3–1.5 as long as wide. Tarsi moderately slim, last segment not extending to the apical mar-gin of segment 3, claws extending to apex of marginal setae (Fig. 3). Claws simple.
TypesHolotype male: “MADAGASKAR MORONDAVA, 2004.07.30 leg. W. Suppantschitsch” // “S 20° 12’
18.2’’, E 44° 21’ 44.1’’, Höhe: 20m”// “HOLOTYPE des. L. Borowiec” // “Cassida morondaviana n. sp.,HOLOTYPUS, des. L. Borowiec ‘2006” (preserved at the Department of Biodiversity and Evolutionary Tax-onomy, Wrocław, Poland); 16 paratypes: “MADAGASKAR MORONDAVA, 2004.07.30 leg. W. Suppants-chitsch” // “S 20° 12’ 18.2’’, E 44° 21’ 44.1’’, Höhe: 20m”// “PARATYPE des. L. Borowiec” // “Cassidamorondaviana n. sp., PARATYPUS, des. L. Borowiec ‘2006” (preserved at the Department of Biodiversityand Evolutionary Taxonomy, Wrocław, Poland; 10 paratypes in coll. U. Heinig, Berlin, Germany).
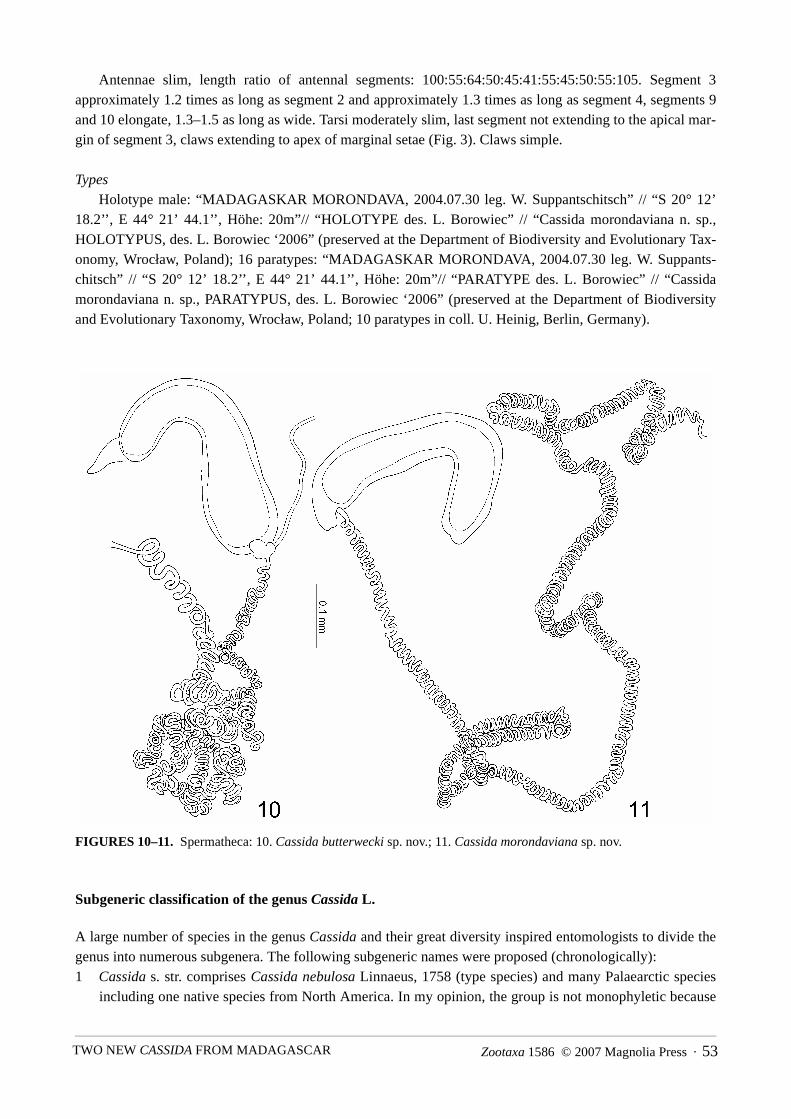
FIGURES 10–11. Spermatheca: 10. Cassida butterwecki sp. nov.; 11. Cassida morondaviana sp. nov.
Subgeneric classification of the genus Cassida L.
A large number of species in the genus Cassida and their great diversity inspired entomologists to divide thegenus into numerous subgenera. The following subgeneric names were proposed (chronologically):1 Cassida s. str. comprises Cassida nebulosa Linnaeus, 1758 (type species) and many Palaearctic species
including one native species from North America. In my opinion, the group is not monophyletic because

BOROWIEC54 · Zootaxa 1586 © 2007 Magnolia Press
at least the type species Cassida nebulosa and Cassida reitteri Weise, 1892 are probably not congenericwith most of other species. Also position of Cassida lineola Creutzer, 1799 and Cassida flaveola Thun-berg, 1794 is doubtful. The four problematic species have some details of adult morphology or immaturestages distinct from other taxa (Medvedev, 1982; Steinhausen, 2002). Other Palaearctic species belongingto the nominotypical subgenus form a coherent group and Cassida relicta Spaeth, 1927 from North Amer-ica is, without doubts, congeneric with the Palaearctic species. On the other hand, within Cassida s. str.we can create several subgroups of similar morphology and feeding preferences. See also notes underPseudocassis Steinhausen, 2002 and Betacassida Steinhausen, 2002.
2 Redtenbacher (1858) proposed the name Deloyala for Cassida seraphina Ménétries, 1836 a distinct spe-cies from Turkey and Greece. The name has been preoccupied and Hincks (1950) substituted Alledoya forDeloyala Redtenbacher not Duponchel and Chevrolat, 1843 (two years later also Gressitt (1952) proposeda new name Lasiocassis to substitute this junior homonym and his name is now the junior synonym ofAlledoya Hincks, 1950). C. hablitziae Motschulsky, 1838 from Turkey and Kazakhstan was also includedin this subgenus, a species very close to C. seraphina and C. vespertina Boheman, 1862 from the easternpart of the Palaearctic Region. In my opinion the group is artificial and C. vespertina is not closely relatedto the two western Palaearctic members. Its superficial similarity is probably the effect of evolutionaryparallelism. They have different host range, both western species feed on Chenopodiaceae, the easternspecies on Ranunculaceae and Convolvulaceae. C. seraphina and C. hablitziae form a distinct speciesgroup and C. vespertina should be placed in a different group.
3 Weise (1889) proposed the name Cassidula for Cassida nobilis Linnaeus, 1758, Cassida vittata Villers,1789 and two other species now classified in genera Ischyronota Weise, 1891 and Oxylepus Desbrochers,1884 (without designation of type species; Hincks in 1952 designated Cassida nobilis as type species).The name was preoccupied and Strand (1928) substituted Cassidulella for Cassidula Weise not de Blain-ville, 1830. Presently the subgenus comprises eight species distributed in the whole Palaearctic Region, iscoherent morphologically and represents a monophyletic group. The similarity of Madagascan Cassidamorondaviana described in this paper to Palaearctic Cassidulella is probably the result of evolutionaryparallelism.
4 Desbrochers (1891) proposed the name Pseudocassida for the common Palaearctic Cassida murraea Lin-naeus, 1768. Its structure is unique and represents a coherent monotypic group.
5 Weise (1891) proposed the name Mionycha for Cassida azurea Fabricius, 1801 from Europe. Further,three other species from the Palaearctic Region were included in the subgenus. The group is coherentmorphologically and biologically and represents a monophyletic group.
6 Weise (1891) proposed the name Odontionycha for all European species of Cassida with toothed clawswithout designation of type species. The taxon was completely artificial and species included by Weise(1891) in the group actually are spread into five subgenera or genera. Hincks (1952) designated Cassidaviridis Linnaeus, 1758 as type species and in his sense the subgenus Odontionycha comprises only thewide spread Palaearctic Cassida viridis and its four relatives from China. The group is coherent morpho-logically and biologically and represents a monophyletic group.
7 Spaeth (1912) proposed the genus Crepidaspis for Crepidaspis varicornis Spaeth, 1912 from Borneo.Later Hincks (1952) synonymized the genus Crepidaspis with the genus Thlaspidosoma Spaeth, 1901 butBorowiec (1990) rightly noted that Crepidaspis varicornis is not congeneric with Thlaspidosoma but isvery close to some Oriental and Papuan species of Cassida, especially Cassida recondita (Boheman,1862) and its relatives. He suggested also that the group is close to Oriental Taiwania Spaeth, 1913 (seenote below on the name Taiwania). The monophyly of Cassida varicornis and Cassida recondita (and itsrelatives) is unquestionable but relationships and borders between this group and other groups from theOriental Region are difficult to verify (see discussion under Taiwania, Cyclocassida and Yunocassis).Because of this difficulty Borowiec (1990) proposed provisionally the name Crepidaspis as a senior syn-

Zootaxa 1586 © 2007 Magnolia Press · 55TWO NEW CASSIDA FROM MADAGASCAR
onym for Taiwania, Cyclocassida and Yunocassis. As a result the genus Crepidaspis becomes one of thelargest subgenera containing most of tropical Cassida species.
8 Spaeth (1913) proposed the genus Taiwania for Taiwania sauteri Spaeth, 1913 from Taiwan. In severalsubsequent papers (e.g. Gressitt, 1952 and Chen et al., 1986), many Oriental species with toothed clawshave been included under the name Taiwania but in my opinion most of them are not congeneric with Tai-wania sauteri, type species for the name. According to my earlier paper (Borowiec, 1990) Taiwania isclose to Crepidaspis but relationships between both subgenera and all species included in the subgenusTaiwania are unclear. Recently, some authors (e.g. Seeno and Wilcox, 1982 or Chen et al., 1986) havetreated Taiwania as a genus with Cyclocassida Chen et Zia, 1961, Yunocassis Chen et Zia, 1961, and Cyr-tonocassis Chen et Zia, 1961 as subgenera but in my opinion Taiwania in its broad sense is a polyphyleticgroup which includes all tropical Cassida with toothed claws. In my view the name Taiwania should bereserved for Cassida sauteri and its several Oriental relatives partly classified in Taiwania and partly inYunocassis: C. achardi (Spaeth, 1926), C. appluda Spaeth, 1926, C. belli Weise, 1897, C. belliformisMaulik, 1919, C. feae Spaeth, 1904, C. inciens Spaeth, 1926, C. occursans Spaeth, 1914, C. spaethianaGressitt, 1952, and C. truncatipennis Spaeth, 1914. See also discussion under the names Yunocassis andCyclocassida.
9 Reitter in Spaeth and Reitter (1926) proposed the name Tylocentra for Cassida turcmenica Weise, 1892from Middle Asia and its relatives (without type designation; Hincks in 1952 designated Cassida turc-menica Weise as type species). Currently, the group comprises at least seven species distributed in aridparts of Palaearctic Region from Cyprus to China, and one species in NE Africa and Saudi Arabia. Thegroup is coherent morphologically and biologically. The structure of the larva is different from that whichis typical for the genus Cassida (see Medvedev and Matys, 1975; Zaitsev, 1992). Based on the unique lar-val morphology Medvedev (1982) proposed the genus Tylocentra. In my opinion this is not justified.Adults have all the characters of the genus Cassida while the larvae have the strongly reduced lateralscoli, unique characters correlated with desert and semidesert saline habitats (various stages of reductionof lateral scoli were observed in larvae of other genera from saline habitats e.g. Ischyronota Weise, 1891and Oxylepus Desbrochers, 1884). Without doubt Tylocentra is a monophyletic group within Cassida.
10 Spaeth in Spaeth and Reitter (1926) proposed the name Eremocassis for Eremocassis transcaspia Spaeth,1926 = Cassida weisei Jacobson, 1894 from Middle Asia. Borowiec (1994) based on adult morphologyand description of the larva (Zaitsev, 1992) noted that C. weisei is congeneric with Tylocentra.
11 Reitter in Spaeth and Reitter (1926) proposed the name Lordicassis for Cassida undecimnotata Gebler,1833. Presently the group comprises six closely related species distributed from NE Turkey to W China,especially in mountains and submountains. All are probably close to Pseudocassida but represents a mor-phologically and biologically coherent monophyletic group.
12 Reitter in Spaeth and Reitter (1926) proposed the name Lordiconia for Cassida canaliculata Gebler, 1833from the western Palaearctic. The species looks distinct and its relationships with other Palaearctic groupsare unclear.
13 Spaeth in Spaeth and Reitter, 1926 proposed the name Onychocassis for Cassida brevis Weise, 1884 andCassida bella Faldermann, 1837. Currently the group comprises three species distributed in the easternpart of the Mediterranean Subregion. The group looks distinct and its relationships with other Palaearcticgroups are unclear.
14 Spaeth in Hincks (1952) proposed the name Mionychella for Cassida hemisphaerica Herbst, 1799 fromEurope. At first glance the Cassida litigiosa group from Africa (Borowiec and Świętojańska, 2001) looksclose to Mionychella but this hypothesis needs verification based on structure of immatures, unknown forboth European and African taxa.
15 Chen and Zia (1961) proposed within the genus Taiwania two subgeneric names: Cyclocassida (with Tai-wania variabilis Chen and Zia, 1961 as type species) and Yunocassis (with Cassida appluda Spaeth, 1926

BOROWIEC56 · Zootaxa 1586 © 2007 Magnolia Press
as type species). In the same paper Chen and Zia (1961) proposed also a new genus Cyrtonocassis forCassida tumidicollis Chen et Zia, 1961. Later Chen et al. (1986) placed Cyrtonocassis as a subgenus ofTaiwania. The system of subgenera proposed by the Chinese authors is based on combination of only afew characters: structure of claws (simple in Cyclocassida and Yunocassis, appendiculate in Taiwania s.str. and Cyrtonocassis), structure of clypeus (narrow in Cyclocassida, broad in other subgenera with someexceptions in Taiwania s. str.), and shape of pronotum (regularly elliptical in Yunocassis, variable in othersubgenera; strongly raised pronotal disc in Cyrtonocassis and flat to moderately convex in other subgen-era). In my opinion, these characters are of less phylogenetic value and the subgenera are not coherentexcept Cyrtonocassis which probably forms a monophyletic group with two related species. Cyclo-cassida, Yunocassis and Taiwania are completely artificial. The appendiculate and simple tarsal clawsdeveloped independently in several lineages within the genus Cassida. Chen and Zia overlooked that inmany Taiwania species claws are simple but appearing appendiculate due to distally projecting sides ofthe last segment of tarsi. A good example of the artificial nature of Taiwania sensu Chen et Zia is thegroup of species close to the type species Cassida sauteri (see note under subgenus Taiwania). The grouplooks at first glance very uniform (large size, regularly elliptical pronotum, similar character of dorsal pat-tern, broad clypeus etc.) but members of the group present all types of claw structure: simple, appendicu-late, and simple but appearing appendiculate due to distally projecting sides of the last tarsal segment.Chen and Zias’ (1961) species with simple claws are placed within Yunocassis and species with appendic-ulate or apparently appendiculate claws within Taiwania s. str. On the other hand separation of Taiwaniafrom Cassida is unclear because in other geographical regions there are groups of species which havecharacters of both Taiwania and Cassida. In my opinion Taiwania is a monophyletic group within Cassidaand comprises only species close Cassida sauteri (see note under subgenus Taiwania), Yunocassis is asimple synonym of Taiwania, and Cyclocassida is paraphyletic group within the rest of species placed byChinese authors within Taiwania sensu lato.
16 Günther (1958) proposed the name Dolichocassida for Cassida (Dolichocassida) veselyi Günther, 1958from Albania and Corsica. Sassi and Borowiec (2006) after study of types synonymized Cassida veselyiwith the well known Mediterranean species Cassida pusilla Waltl, 1839. The species with two otherrelated taxa were usually classified within Cassidulella but in my opinion it is not congeneric with Cas-sidulella or any European Cassida group and Dolichocassida forms a coherent monophyletic group.
17 Steinhausen (2002) proposed the name Pseudocassis for the wide-spread Palaearctic Cassida flaveolaThunberg, 1794 based on the structure of the pupa. The species was usually classified in the nominotypi-cal subgenus. I agree with Steinhausen that this taxon is not close to most of other members of PalaearcticCassida s. str. and represents a separate line (confirmed also by different feeding preferences) but it is dif-ficult to diagnose this subgenus based on adult and larval characters. Its position within Cassida is stillunclear.
18 Steinhausen (2002) proposed the name Betacassida for the wide-spread Palaearctic Cassida nebulosaLinnaeus, 1758 based on the structure of the pupa. He overlooked that Cassida nebulosa is a type speciesfor Cassida and consequently his name is its objective synonym. I agree with Steinhausen that this taxonis not close to most other members of Palaearctic Cassida s. str. and, like Cassida flaveola, also representsa separate line (confirmed by different feeding preferences). As with the name Pseudocassis it is difficultto diagnose Cassida s. str. in its narrow sense based on adult and larval characters and I do not propose asubgeneric name for the rest of the species in the nominotypical subgenus.
Review of subgeneric names proposed in the genus Cassida L. leads me to the following conclusions:- the subgeneric names were proposed only for taxa from Palaearctic and Oriental regions. Almost one half
of the described species, especially from Africa, Madagascar and Australia, have never been classified inany proposed subgenera.

Zootaxa 1586 © 2007 Magnolia Press · 57TWO NEW CASSIDA FROM MADAGASCAR
- the status of proposed subgeneric names is not balanced. Some subgenera are only narrow groups of veryclosely related species, others are broad artificial groups of unrelated taxa.
- the subgenera were proposed based on questionable characters, some subgeneric diagnoses are based onlyon 3–5 adult characters, some partly on larval characters, and a few others were based only on pupal char-acters. None of the authors above analysed a diversity of characters in taxa from all zoogeographicregions, nor did they correlate morphological characters with biological data.
- several species from Africa, Madagascar and Australia possess unique characters and it is difficult toplace them in any previously described subgenus. On the other hand some characters in these species arecharacters proposed as diagnostic for previously described subgenera (my unpublished data).
- in my opinion, the best solution is to eliminate all proposed subgeneric names. This point of view was pre-sented in my “World Catalogue of Cassidinae” (Borowiec, 1999). Creation of narrow groups of relatedspecies based on the complex of morphological and biological characters will be more informative thansubgenera, even if many species remain as monotypic groups of uncertain position.
Acknowledgements
I would like to express my sincere thanks to U. Heinig (Berlin, Germany) and W. Schawaller (Musum fürTierkunde, Stuttgart, Germany) for the loan of the material. Thanks to Mrs H. Staszak for preparing figures ofspermathecae and to two anonymous reviewers, who greatly improved the text. This paper was supported byscientific grant of the Zoological Institute, University of Wrocław, 1018/IZ/2006.
References
Bordy, B. & Doguet, S. (1987) Contribution à la connaissance des Cassidinae de France. Étude de leur spermathèque(Coleoptera, Chrysomelidae). Nouvelle Revue d’Entomologie (N.S.), 4, 161–176.
Borowiec, L. (1990) A review of the genus Cassida L. of the Australian Region and Papuan Subregion (Coleoptera,Chrysomelidae, Cassidinae). Genus, Wrocław, 1, 1–51.
Borowiec, L. (1994) New synonyms in the Cassidinae (Coleoptera: Chrysomelidae). Genus, 5, 153–159. Borowiec, L. (1999) A world catalogue of the Cassidinae (Coleoptera: Chrysomelidae). Biologica Silesiae, Wrocław,
476 pp.Borowiec, L. & Świętojańska, J. (2001) Revision of Cassida litigiosa group from southern Africa (Coleoptera: Chry-
somelidae: Cassidinae). Annales Zoologici Warszawa, 51, 153–184.Borowiec, L. & Świętojańska, J. (2002) Cassidinae of the world — an interactive manual (Coleoptera: Chrysomelidae).
Permanent electronic publication (open in 2002): www.biol.uni.wroc.pl/cassidae/katalog%20internetowy/index.htm.
Chen, S. H. and Zia, Y. (1961) Results of the zoologico-botanical expedition to Southwest China 1955-1957 (Coleoptera,Cassidinae). Acta Entomologica Sinica, 10, 439–451.
Chen, S., Yu, P., Sun, C. & Zia, Y. (1986) Coleoptera Hispidae. In: Fauna Sinica, Insecta. Science Press, Beijing, 653 pp.+ XV pl.
Desbrochers des Loges, J. (1891) Monographie des Cassida de France suivie d’observations sur le classement desespèces de ce genre au Catalogus Coleopterorum Europae, Caucasi et Armeniae rossicae. Frelon, 1891-1892, 1, 7,1–48.
Gressitt, J. . (1952) The tortoise beetles of China (Chrysomelidae: Cassidinae). Proceedings of the California Academyof Sciences, 27, 433–592.
Günther, V., 1958. Novy druh a novy podrod celedi Cassididae (Col.). Acta Entomologica Musei Nationalis Pragae, 32,567–569.
Hincks, W.D. (1950) Some nomenclatorial notes on Chrysomelidae (Col.). No. 3, Cassidinae. Annals and Magazine ofNatural History, ser. 12, 3, 506–512.
Hincks, W.D. (1952) The genera of the Cassidinae (Coleoptera: Chrysomelidae). Transactions of the RoyalEntomological Society of London, 103, 327–358.
Medvedev, L.N. (1982) Listoyedy MNR. Opredelitel. Nauka, Moskva, 1982, 303 pp.

BOROWIEC58 · Zootaxa 1586 © 2007 Magnolia Press
Medvedev L.N. & Matys E.G. (1975) Neue paläarktische Schildkäferarten der Gattung Cassida L. (Coleoptera, Chry-somelidae). Deutsche Entomologische Zeitschrift, N.F., 22, 137–143.
Redtenbacher, L. (1858) Fauna Austriaca. Die Käfer, nach der analytischen Methode bearbeitet, 2 Aufl. Wien.Sassi, D. & Borowiec, L. (2006) Cassida inopinata, a new species from Italy and Balkan Region (Coleoptera: Chry-
somelidae: Cassidinae). Genus, Wrocław, 17, 545–560.Seeno, T.N. & Wilcox, J.A. (1982) Leaf beetle genera (Coleoptera: Chrysomelidae). Entomography, 1, 1–221.Spaeth, F. (1912) One new genus and some new species of Cassidae from Borneo, with a list of all the species at present
known from that Island. Sarawak Museum Journal, 1, 118–128. Spaeth, F. (1913) H. Sauter's Formosa-Ausbeute. Cassidinae. Annales historico-naturales Musei Nationalis Hungarici,
11, 46-48.Spaeth, F. & Reitter, E. (1926) Bestimmungs-Tabellen der europäischen Coleopteren. 95 Heft. Cassidinae der palaeark-
tischen Region. Troppau, 68 pp.Steinhausen, W. (2002) Die Puppen mitteleuropäischen Blattkäfer - Eine vorläufige Bestimmungstabelle 2. Teil. Mittei-
lungen der Münchner Entomologischen Gesselschaft, 92, 5–36. Strand, E. (1928) Nomenklatorische Bemerkungen über einige Coleopteren-Gattungen. Entomologisches Nachrichtenb-
latt, 2, 1–2. Weise, J. (1889) Neue Chrysomeliden und Coccinelliden aus dem Kaukasus. Wiener Entomologische Zeitung, 8, 259–
262. Weise, J. (1891) Bemerkungen zur Gattung Cassida. Wiener Entomologische Zeitung, 10, 203–205. Zaitsev, J.M. (1992). Preimaginal'niye stadii i osobennosti ekologii zhuka-listoeda Eremocassis weisei (Coleoptera,
Chrysomelidae). Westnik Zoologii, 1992, 2, 67–70.




















