
RELATIVE ABUNDANCE AND THE SPECIES-SPECIFIC REINFORCEMENT OF MALE MATING PREFERENCE IN THE...
17
2639 q 2005 The Society for the Study of Evolution. All rights reserved. Evolution, 59(12), 2005, pp. 2639–2655 RELATIVE ABUNDANCE AND THE SPECIES-SPECIFIC REINFORCEMENT OF MALE MATING PREFERENCE IN THE CHRYSOCHUS (COLEOPTERA: CHRYSOMELIDAE) HYBRID ZONE MERRILL A. PETERSON, 1,2 BARBARA M. HONCHAK, 1,3 STEFANIE E. LOCKE, 1,4 TIMOTHY E. BEEMAN, 1 JESSICA MENDOZA, 1 JABIN GREEN, 1 KATI J. BUCKINGHAM, 1 MICHAEL A. WHITE, 1,5 AND KIRSTEN J. MONSEN 1,6 1 Biology Department, Western Washington University, Bellingham, Washington 98225 2 E-mail: [email protected] Abstract. Most studies of reinforcement have focused on the evolution of either female choice or male mating cues, following the long-held view in sexual selection theory that mating mistakes are typically more costly for females than for males. However, factors such as conspecific sperm precedence can buffer females against the cost of mating mistakes, suggesting that in some hybrid zones mating mistakes may be more costly for males than for females. Thus, the historical bias in reinforcement research may underestimate its frequency. In this study, we present evidence that reinforcement has driven the evolution of male choice in a hybrid zone between the highly promiscuous leaf beetles Chrysochus cobaltinus and C. auratus, the hybrids of which have extremely low fitness. In addition, there is evidence for male choice in these beetles and that male mating mistakes may be costly, due to reduced opportunities to mate with conspecific females. The present study combines laboratory and field methods to quantify the strength of sexual isolation, test the hypothesis of reproductive character displacement, and assess the link between relative abundance and the strength of selection against hybridization. We document that, while sexual isolation is weak, it is sufficient to produce positive assortative mating. In addition, reproductive character displacement was only detected in the relatively rare species. The strong postzygotic barriers in this system are sufficient to generate the bimodality that characterizes this hybrid zone, but the weak sexual isolation is not, calling into question whether strong prezygotic isolation is necessary for the maintenance of bimodality. Growing evidence that the cost of mating mistakes is sufficient to shape the evolution of male mate choice suggests that the reinforcement of male mate choice may prove to be a widespread occurrence. Key words. Bimodal hybrid zone, hybridization, reproductive character displacement, sexual isolation, speciation. Received March 3, 2005. Accepted September 24, 2005. The evolution of barriers to genetic exchange is widely considered to be central to the origin of species and thus has been the primary focus of speciation research (Barton 1996; Harrison 1998; Howard 1998; Howard et al. 1998). The roots of this focus on reproductive barriers trace to the biological species concept (Mayr 1963), which emphasizes the role of barriers to reproduction in defining species boundaries (Coy- ne 1992). Although species concepts continue to be debated (Mayden 1997; Harrison 1998; Hey 2001), an advantage of the biological species concept is that it points to understand- ing the evolution of these reproductive barriers as the key to understanding speciation (Coyne 1992). One of the most fre- quently studied and controversial theories on the evolution of premating barriers has been the theory of reinforcement. Simply put, this theory predicts that selection against hy- bridization (e.g., as a result of low hybrid fitness) should favor the evolution of prezygotic barriers between hybrid- izing groups (Dobzhansky 1940). An expected result of re- inforcement is reproductive character displacement of mating cues and/or mate preferences, giving rise to an increase in assortative mating by individuals from sympatric populations 3 Present address: Department of Biological Sciences, Northern Arizona University, PO Box 5640, Flagstaff, Arizona 86011-5640. 4 Present address: Department of Biology, West Chester Uni- versity of Pennsylvania, West Chester, Pennsylvania 19383. 5 Present address: Laboratory of Genetics, 425-G Henry Mall, University of Wisconsin, Madison, Wisconsin 53706. 6 Present address: Department of Biology and Molecular Biology, Montclair State University, Upper Montclair, New Jersey 07043. of hybridizing species, compared to those from allopatric populations (Howard 1993; but see Lemmon et al. 2004). Although reinforcement theory provides an intuitively ap- pealing mechanism by which natural selection can play a role in the speciation process, its support has often been challenged. Some (e.g., Butlin 1987, 1989) suggested that reinforcement should be viewed strictly as a process that leads to the evolution of prezygotic barriers when postzy- gotic barriers are not complete, and they argued that many purported examples of reinforcement do not meet this strict definition. Others (Howard 1993; Liou and Price 1994) de- fined it more broadly, arguing that there is no fundamental difference between partial and complete postzygotic barri- ers. Consensus on this issue appears to be growing, in light of theory indicating that reproductive character displace- ment can evolve similarly under both complete postzygotic barriers and strong, but incomplete postzygotic barriers (Liou and Price 1994; Butlin 1995). Even under its broader definition, reinforcement has often been challenged both with theoretical arguments against its plausibility and with assertions that its empirical support is weak and/or open to other interpretations (see reviews in Howard 1993; Noor 1999; Servedio and Noor 2003). However, a recent resur- gence of theoretical and empirical investigations of rein- forcement, spurred by reviews suggesting that reproductive character displacement is a common phenomenon (Coyne and Orr 1989; Howard 1993), has strengthened the case for reinforcement as a process that can lead to the evolution of heightened premating barriers. Specifically, many new the- oretical studies (Liou and Price 1994; Kelly and Noor 1996;
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of RELATIVE ABUNDANCE AND THE SPECIES-SPECIFIC REINFORCEMENT OF MALE MATING PREFERENCE IN THE...

2639
q 2005 The Society for the Study of Evolution. All rights reserved.
Evolution, 59(12), 2005, pp. 2639–2655
RELATIVE ABUNDANCE AND THE SPECIES-SPECIFIC REINFORCEMENT OF MALEMATING PREFERENCE IN THE CHRYSOCHUS (COLEOPTERA: CHRYSOMELIDAE)
HYBRID ZONE
MERRILL A. PETERSON,1,2 BARBARA M. HONCHAK,1,3 STEFANIE E. LOCKE,1,4 TIMOTHY E. BEEMAN,1
JESSICA MENDOZA,1 JABIN GREEN,1 KATI J. BUCKINGHAM,1 MICHAEL A. WHITE,1,5 AND KIRSTEN J. MONSEN1,6
1Biology Department, Western Washington University, Bellingham, Washington 982252E-mail: [email protected]
Abstract. Most studies of reinforcement have focused on the evolution of either female choice or male mating cues,following the long-held view in sexual selection theory that mating mistakes are typically more costly for femalesthan for males. However, factors such as conspecific sperm precedence can buffer females against the cost of matingmistakes, suggesting that in some hybrid zones mating mistakes may be more costly for males than for females. Thus,the historical bias in reinforcement research may underestimate its frequency. In this study, we present evidence thatreinforcement has driven the evolution of male choice in a hybrid zone between the highly promiscuous leaf beetlesChrysochus cobaltinus and C. auratus, the hybrids of which have extremely low fitness. In addition, there is evidencefor male choice in these beetles and that male mating mistakes may be costly, due to reduced opportunities to matewith conspecific females. The present study combines laboratory and field methods to quantify the strength of sexualisolation, test the hypothesis of reproductive character displacement, and assess the link between relative abundanceand the strength of selection against hybridization. We document that, while sexual isolation is weak, it is sufficientto produce positive assortative mating. In addition, reproductive character displacement was only detected in therelatively rare species. The strong postzygotic barriers in this system are sufficient to generate the bimodality thatcharacterizes this hybrid zone, but the weak sexual isolation is not, calling into question whether strong prezygoticisolation is necessary for the maintenance of bimodality. Growing evidence that the cost of mating mistakes is sufficientto shape the evolution of male mate choice suggests that the reinforcement of male mate choice may prove to be awidespread occurrence.
Key words. Bimodal hybrid zone, hybridization, reproductive character displacement, sexual isolation, speciation.
Received March 3, 2005. Accepted September 24, 2005.
The evolution of barriers to genetic exchange is widelyconsidered to be central to the origin of species and thus hasbeen the primary focus of speciation research (Barton 1996;Harrison 1998; Howard 1998; Howard et al. 1998). The rootsof this focus on reproductive barriers trace to the biologicalspecies concept (Mayr 1963), which emphasizes the role ofbarriers to reproduction in defining species boundaries (Coy-ne 1992). Although species concepts continue to be debated(Mayden 1997; Harrison 1998; Hey 2001), an advantage ofthe biological species concept is that it points to understand-ing the evolution of these reproductive barriers as the key tounderstanding speciation (Coyne 1992). One of the most fre-quently studied and controversial theories on the evolutionof premating barriers has been the theory of reinforcement.Simply put, this theory predicts that selection against hy-bridization (e.g., as a result of low hybrid fitness) shouldfavor the evolution of prezygotic barriers between hybrid-izing groups (Dobzhansky 1940). An expected result of re-inforcement is reproductive character displacement of matingcues and/or mate preferences, giving rise to an increase inassortative mating by individuals from sympatric populations
3 Present address: Department of Biological Sciences, NorthernArizona University, PO Box 5640, Flagstaff, Arizona 86011-5640.
4 Present address: Department of Biology, West Chester Uni-versity of Pennsylvania, West Chester, Pennsylvania 19383.
5 Present address: Laboratory of Genetics, 425-G Henry Mall,University of Wisconsin, Madison, Wisconsin 53706.
6 Present address: Department of Biology and Molecular Biology,Montclair State University, Upper Montclair, New Jersey 07043.
of hybridizing species, compared to those from allopatricpopulations (Howard 1993; but see Lemmon et al. 2004).
Although reinforcement theory provides an intuitively ap-pealing mechanism by which natural selection can play arole in the speciation process, its support has often beenchallenged. Some (e.g., Butlin 1987, 1989) suggested thatreinforcement should be viewed strictly as a process thatleads to the evolution of prezygotic barriers when postzy-gotic barriers are not complete, and they argued that manypurported examples of reinforcement do not meet this strictdefinition. Others (Howard 1993; Liou and Price 1994) de-fined it more broadly, arguing that there is no fundamentaldifference between partial and complete postzygotic barri-ers. Consensus on this issue appears to be growing, in lightof theory indicating that reproductive character displace-ment can evolve similarly under both complete postzygoticbarriers and strong, but incomplete postzygotic barriers(Liou and Price 1994; Butlin 1995). Even under its broaderdefinition, reinforcement has often been challenged bothwith theoretical arguments against its plausibility and withassertions that its empirical support is weak and/or open toother interpretations (see reviews in Howard 1993; Noor1999; Servedio and Noor 2003). However, a recent resur-gence of theoretical and empirical investigations of rein-forcement, spurred by reviews suggesting that reproductivecharacter displacement is a common phenomenon (Coyneand Orr 1989; Howard 1993), has strengthened the case forreinforcement as a process that can lead to the evolution ofheightened premating barriers. Specifically, many new the-oretical studies (Liou and Price 1994; Kelly and Noor 1996;

2640 MERRILL A. PETERSON ET AL.
Servedio and Kirkpatrick 1997; Cain et al. 1999; Kirkpatrickand Servedio 1999) have demonstrated that reinforcementcan occur under a much broader range of conditions thanhad been previously thought. In addition, examples fromseveral new systems studied during the past decade (Noor1995a; Sætre et al. 1997; Rundle and Schluter 1998; Higgieet al. 2000; Gabor and Ryan 2001; Jiggins et al. 2001; Hobeland Gerhardt 2003; Nosil et al. 2003) have strengthened theempirical evidence for reinforcement (reviewed in Noor1999; Servedio and Noor 2003).
In general, these new empirical examples have relied onestimates of sexual isolaton from the laboratory and have notverified that similar patterns of sexual isolation occur undernatural conditions (but see Sætre et al. 1997). There is littlea priori reason to expect that laboratory studies of assortativemating will produce results that correspond to mating deci-sions made under natural conditions, because mating behav-iors are likely to vary with environmental conditions (Noor1999). In spite of this shortcoming, the growing number ofempirical examples consistent with reinforcement, coupledwith theoretical results indicating that it can occur under lessrestrictive conditions than previously assumed, has led long-standing critics to acknowledge that it is not as implausibleas they had previously argued (Butlin and Tregenza 1997).Reinforcement advocates have gone even further, suggestingthat its existence is now well established and that it is timeto begin efforts to refine our understanding of reinforcement’sfrequency and importance (Servedio and Noor 2003).
A significant problem currently compromising our abilityto understand the generality of reinforcement stems from ageneral bias in how the phenomenon is studied. The vastmajority of reinforcement studies to date (reviewed in How-ard 1993; Noor 1999; Servedio and Noor 2003) focused onthe evolution of either female preferences or male matingsignals. This bias undoubtedly traces to the premise fromsexual selection theory (e.g., Trivers 1972; Jennions andPetrie 1997) that, in general, mating mistakes are more cost-ly for females than for males. Nonetheless, it may also bethe case that heterospecific matings can be sufficiently cost-ly for males to drive the reinforcement of male mate pref-erences. For example, heterospecific matings, like matingswith conspecific females, might diminish the amounts ofsperm or accessory substances in subsequent ejaculates(Bonduriansky 2001; Wedell et al. 2002) or reduce subse-quent opportunities for mating if males spend considerabletime and effort finding, copulating with, and/or guardingfemales (Dickinson 1995; Marshall et al. 2002; Martin andHosken 2004) or if they suffer injury during copulation(Sota and Kubota 1998). Thus, if heterospecific matingsyield either no offspring or offspring of reduced fitness, itstands to reason that reinforcing selection may be sufficientto drive the evolution of increased mate discrimination bymales. Recent examples of the reproductive character dis-placement of male mating preferences (Gabor and Ryan2001; Jiggins et al. 2001; Albert and Schluter 2004) supportthis assertion (for an exception see Nosil et al. 2003). How-ever, given the strong bias toward studying reinforcementof either female choice or male mating signals, it remainsunclear whether this is a common phenomenon or underwhich conditions it generally occurs.
The Chrysochus Hybrid Zone
In this paper, we provide evidence that reinforcement hasshaped the evolution of male mate choice in a hybrid zoneinvolving the chrysomelid beetles Chrysochus auratus and C.cobaltinus. These sister species are herbivorous beetles thatfeed on plants in the Apocynaceae (including Asclepiadaceae;Backlund et al. 2000; see also Dobler and Farrell 1999).Chrysochus cobaltinus eats Apocynum cannabinum and A. an-drosaemifolium, as well as Asclepias speciosa and A. erio-carpa (Sady 1994; Dickinson 1995; Dobler and Farrell 1999).Chrysochus auratus has a narrower diet, feeding exclusivelyon A. cannabinum and A. androsaemifolium (Dussourd andEisner 1987; Williams 1992; Dobler and Farrell 1999).Adults feed on leaves of their host plants, and mating occurson the host plants (Weiss and West 1921; Dickinson 1995).Larvae of both species attack the roots of the adult host plants(Weiss and West 1921).
In low-lying areas within the Yakima River valley ofWashington state, C. cobaltinus and C. auratus form a 75-km-wide hybrid zone (Peterson et al. 2001, 2005). This hy-brid zone apparently formed as a result of secondary contact(Peterson et al. 2005). Chrysochus auratus has a primarilyeastern North American distribution, spanning the easternUnited States and adjacent southern Canada, west to theRocky Mountains, whereas C. cobaltinus is found exclusivelyin western North America, from southern California to south-ern British Columbia, east to Utah and western Montana(Dobler and Farrell 1999; Peterson et al. 2001). The onlyknown area of sympatry between the two species occurs insouth-central Washington (Peterson et al. 2001). Availableevidence indicates that the Chrysochus hybrid zone is of post-Pleistocene origin. Specifically, during the late Pleistocene,most of central Washington, including the Yakima River val-ley, was repeatedly inundated and ravaged by flooding fromglacial Lake Missoula (Bretz 1969; Benito and O’Connor2003; Clague et al. 2003). It is extremely unlikely that anyChrysochus populations in the region could have survivedthese repeated floods, which ended approximately 13,000years ago and have been described as the largest known fresh-water floods on the planet (Benito 1997). The current dis-tribution of C. auratus (Peterson et al. 2001) suggests that itexpanded into south-central Washington via Alberta and Brit-ish Columbia after the retreat of the Pleistocene glaciers.
Unlike in many hybrid zones, mate choices in the Chry-sochus hybrid zone are unusually easy to observe in the field(Peterson et al. 2001). This advantage is due in part to thefact that Chrysochus adults are highly polygamous; individ-uals average one mating per day (Dickinson 1995), such thatsome males and females mate more than 50 in their lives(Dickinson 1997). In addition, males exhibit postcopulatorymate guarding in which males ride the backs of females. Thus,although copulation usually lasts less than 20 min, matingpairs remain together for an average of 1.7 h (Dickinson1995), facilitating observations of mate choice. There is littleevidence that female choice is important in courtship; uponencountering a female, a male mounts the female from therear and begins attempting to copulate. Females may lowertheir abdomen in an apparent attempt to prevent copulation,but this behavior is largely ineffective; once males begin

2641REINFORCEMENT IN A BEETLE HYBRID ZONE
attempting to copulate with a female, they typically persistuntil they are successful (M. A. Peterson, pers. obs.). Furtherevidence for male choice comes from observations of malesattempting to copulate with dead females. When presentedwith dead conspecific and heterospecific females, Chrysochusmales are significantly more likely to copulate with the con-specific female (M. A. Peterson, S. Dobler, E. Larson, D.Juarez, T. Schlarbaum, K. Monsen, and W. Francke, unpubl.data).
In patches where the two species have similar abundance,hybridization is frequent and F1 hybrids represent up to 15%of the individuals (Peterson et al. 2001). Hybrids have in-termediate color and antennal morphology to C. auratus andC. cobaltinus (Peterson et al. 2001). As a result, identificationbased on phenotype is highly accurate; greater than 97% ofmore than 1000 hybrid zone individuals identified on thebasis of color and morphology were correctly identified, asconfirmed with multilocus species-specific markers (Petersonet al. 2005). The fact that sympatric populations are stronglybimodal (sensu Jiggins and Mallet 2000), with few or no F2or backcross individuals, facilitates accurate identificationusing color and morphology (Peterson et al. 2005). Both di-rect and indirect evidence indicate that there may be sub-stantial fitness costs to hybridization, due to the extremelylow fitness of F1 hybrids. First, experiments with hybrid fe-males reveal that they are effectively sterile (Peterson et al.2005). In addition, extensive allozyme (Peterson et al. 2005)and mitochondrial DNA (K. Monsen, B. Honchak, S. Locke,and M. Peterson, unpubl. data) genotyping of beetles fromsites spanning the hybrid zone has demonstrated that intro-gression is extremely weak, indicating that successful repro-duction by hybrids in the field is exceedingly rare. Theseresults suggest that this hybrid zone is a tension zone (Bartonand Hewitt 1985), in which endogenous selection againsthybrids maintains the hybrid zone in the face of gene flow(e.g., Barton 1980; Szymura and Barton 1986; Hewitt 1990;Jiggins et al. 1996).
Although hybrids have extremely low fitness, individualsin sympatric populations may not be under strong selectionto avoid mating with heterospecific individuals. This linkbetween hybrid fitness and the strength of selection againsthybridization may depend on the frequency with which in-dividuals encounter heterospecific potential mates. For ex-ample, it has been argued that if one species is relativelyrare, that species should be under stronger selection againsthybridization (Howard 1993; Noor 1995a). Although, to ourknowledge, the relationship between relative abundance andthe probability of hybridization has been seldom explicitlyexamined, available evidence suggests that such a relation-ship does exist (e.g., Avise and Saunders 1984; Rieseberg etal. 1998; Ribi and Oertli 2002). Furthermore, for at least threeinsect systems, species-specific differences in abundance areconsistent with species-specific differences in reproductivecharacter displacement (Waage 1979; Noor 1995a; Nosil etal. 2003).
Conspecific sperm precedence (Howard 1999) may bufferfemales against mating mistakes, substantially weakeningthe strength of selection against hybridization (Marshall etal. 2002). For males, however, the cost of time and energyspent courting, copulating, and guarding a heterospecific
mate cannot be reduced once that commitment has beenmade. In the case of Chrysochus, Dickinson (1995) foundthat postcopulatory guarding reduces the probability thatmales will copulate with another mate within 2 h by 41%.Thus, any mating mistake by a Chrysochus male shouldreduce his opportunities to mate with conspecific females.What remains unclear is whether males may suffer furtherreductions in their success in siring conspecific progeny (theonly progeny that will enhance their fitness) as a result ofenergy spent courting, copulating, and guarding females(e.g., Martin and Hosken 2004), or as a result of reducedsperm availability for those matings (e.g., Carroll 1994;Simmons 2001; Lewis 2004). Regardless, the lost matingopportunities resulting from time spent with heterospecificmates would suggest that the frequency with which malesmate with heterospecific females should be positively cor-related with the strength of selection against hybridizationexperienced by those males.
In this paper, we take advantage of the ease with whichthe mate choices of these beetles can be observed in thelaboratory and field to assess the strength of assortative mat-ing and sexual isolation and to determine whether male matechoice exhibits a pattern of reproductive character displace-ment. This is one of the few examples of the reinforcementof male mate choice, and it is one of the only examples ofreinforcement in which measures of sexual isolation (sensuSpieth and Ringo 1983) in the laboratory and field can becompared. Furthermore, our results provide insight into theconditions under which reinforcement of male mate choiceis likely to occur, in that species-specific differences in theprobability of hybridization, mediated by relative abundance,appear to underlie species-specific differences in the rein-forcement of male mate choice.
MATERIALS AND METHODS
Assortative Mating and Sexual Isolation
Assortative mating and sexual isolation in nature
To assess the strength of sexual isolation between C. cob-altinus and C. auratus in the field, we conducted a survey ofmating and single beetles in a 60 3 50-m patch of A. can-nabinum near the center of the hybrid zone (site 17 in Fig.1). This site (9.2 km WNW of Mabton, WA) contains largepopulations of both parental species and has been our focalhybrid zone site in other studies (Peterson et al. 2001, 2005).On 1 July 2000, we noted the identity of all mating and singleindividuals encountered within 2 m of two 60-m paralleltransects at the site. For all pairs (either actively engaged incopulation or with the male in the quiescent mounted posturetypical of postcopulatory mate guarding), we noted the iden-tity of the male and the female.
We compared the observed frequencies of the differentcombinations of male and female types with frequencies ex-pected under random mating, using a G-test of independence,to determine if beetles at this site exhibit positive assortativemating. As reported previously (Peterson et al. 2005), C.auratus, C. cobaltinus, and hybrids accounted for 56.6%,26.8%, and 16.7% of the beetles, respectively, at the site on1 July 2000. For our analyses of assortative mating, we as-

2642 MERRILL A. PETERSON ET AL.
FIG. 1. Map of the hybrid zone in Washington state, indicating the location of populations surveyed in summer 2000. Filled circlesindicate pure Chrysochus cobaltinus populations, open circles indicate pure C. auratus populations, and shaded circles indicate mixedpopulations. Sites 1–22 are hybrid zone sites that were surveyed for relative abundance and the probability of hybridization in summer2000. Sites A1 and A2 are the allopatric sources for C. cobaltinus and C. auratus used in the reproductive character displacement, andsite 17 is the sympatric source for that experiment, as well as the focal hybrid zone site for the other aspects of this study.
sumed a 50:50 sex ratio for nonmating individuals, based onmultiple surveys at this and other sites that have consistentlyfound a 50:50 sex ratio (M. A. Peterson, K. Monsen, L.Tantalo, and C. Fabry, unpubl. data). We omitted matingcombinations involving hybrid beetles, both to restrict ouranalysis to assortative mating between the parental speciesand to simplify comparisons with laboratory experiments (seebelow).
Although the initial G-tests allowed us to determine if therewas a pattern of positive assortative mating between C. au-ratus and C. cobaltinus at this site, those tests do not revealwhether sexual isolation is a contributor to such a pattern.Rolan-Alvarez and Caballero (2000) developed a pair sexualisolation index (PSI) that explicitly quantifies the strength ofsexual isolation between different combinations of males andfemales. PSI factors out the effects of mating propensity orsexual selection that are known to confound other measuresof sexual isolation (Spieth and Ringo 1983; Gilbert andStarmer 1985; Marın 1997; Rolan-Alvarez and Caballero2000). For each pair type, PSI is calculated as:
(ij )(N )PSI 5 ,i, j (total pairs with i males)(total pairs with j females)
(1)
where i represents male identity (auratus or cobaltinus), jrepresents female identity (auratus or cobaltinus), ij repre-
sents total copulations (m 3 f) of a particular pair type, andN is the total copulations observed.
Values of PSI can range from zero to infinity, with PSI .1 indicating an excess of pairs of a particular type comparedto random expectations, and PSI , 1 indicating a deficit ofpairs of that type. The pattern characteristic of sexual iso-lation would be one in which PSI . 1 for both conspecificcombinations, but PSI , 1 for both directions of heteros-pecific crosses (Cruz et al. 2004). We obtained bootstrappedestimates of PSI, its standard deviation, and statistical sig-nificance (PSI ± 1) between all pairwise combinations of C.auratus, C. cobaltinus, and hybrid males and females for thefield sample, using the BASIC program described by Cruzet al. (2004). For these analyses, we assumed that nonmatingindividuals were in a 50:50 sex ratio, and performed 5000resamplings to obtain bootstrapped estimates of PSI. We thenused a sequential Bonferroni test (Rice 1989) to adjust thetablewide alpha level. Note that because PSI estimates arenot statistically independent, the P-values from this testshould be interpreted with caution (Rolan-Alvarez and Ca-ballero 2000). In spite of statistical nonindependence, PSI isa useful index, particularly when used in concert with otherindices of overall sexual isolation (e.g. IPSI) that do not sufferfrom statistical nonindependence (E. Rolan-Alvarez, pers.comm.; see Comparison of Field- and Laboratory-BasedMeasures of Sexual Isolation for a further discussion of IPSI).Moreover, we were mainly interested in the overall pattern

2643REINFORCEMENT IN A BEETLE HYBRID ZONE
of PSI estimates for the laboratory and field, rather than thestatistical significance of any specific PSI estimate.
Assortative mating and sexual isolation in the laboratory
To control for factors other than sexual isolation that maycontribute to assortative mating in the field and to determineif laboratory-based measures of sexual isolation are repre-sentative of mate choices in nature, we performed a no-choicemating experiment in the laboratory during the summer of2002. We used a no-choice design because sexual isolationestimates from male-choice, female-choice, and multiple-choice experiments are potentially influenced by additionalfactors, such as sex- and species-specific differences in mat-ing propensity and intrasexual interactions (Arnold et al.1996; Marın 1997). For this experiment, we collected malesand females (presumably nonvirgins) of each species fromthe same focal site at which the mating surveys had beenconducted in 2000. We sexed each beetle under a dissectingmicroscope by gently opening the genitalic aperture. Thisprocedure allows us to unambiguously sex beetles, with noapparent negative effects. For this experiment, all individualswere maintained individually in the laboratory on fresh cut-tings of A. cannabinum in a 45-ml plastic tube.
Females of each species (86 C. auratus and 85 C. cobal-tinus) were paired first with a male of one species and sub-sequently with a male of the other species. In case the orderin which females encountered males of each type influencedthe probability that pairs would form, half of the females ofeach species were paired with the conspecific male first, andthe other half were initially paired with the heterospecificmale. Each male was assigned to be paired only once witha female of each species. Thus, males and females each en-countered one conspecific individual and one heterospecificindividual in this experiment. Our use of individuals in twotrials each should not increase the likelihood of detectingdifferences between treatments, as each individual encoun-tered both treatments. The only exception was that maleswere used in only one trial if they died between trials orappeared to lack vigor before use in the second trial. In eitherof those events, they were replaced with a vigorous male.We only had to replace eight males during the experiment.All pairs were observed in a constant temperature room(288C, similar to typical daytime conditions in the field duringthe reproductive season) for 3 h, or until copulation wasinitiated, whichever came first. Copulating pairs were sepa-rated immediately and isolated, to ensure that they did notcopulate again. The amount of time before copulation beganwas noted for each pair.
To estimate the strength of sexual isolation from this ex-periment, we obtained bootstrapped estimates (5000 random-izations) of PSI for each pairwise combination of males andfemales. The only difference between this analysis and theanalysis for the field data is that, because the number of malesand females of each species (the mating pool) in the labo-ratory was known rather than estimated, only the frequenciesof mating pairs (instead of mating pairs and nonmating in-dividuals) were resampled in the bootstrap analysis (E. Ro-lan-Alvarez, pers. comm.). As with the field data, we used asequential Bonferroni test (Rice 1989) to adjust tablewide
alpha levels. The mating trials in the no-choice experimentwere allowed to run for up to 3 h, but we were uncertain asto whether such an experiment artificially prolonged the con-tact between individual males and females, relative to thetypical duration of time for such interactions in the field. Ifso, it is possible that our results would be biased, simplybecause pairs that would be unlikely to mate in nature matedonly because they were kept in extended contact with eachother. To determine if the duration of the experiment influ-enced the results, we compared PSI estimates from analysesusing only those copulations within the first 0.5 h of eachtrial with PSI estimates from analyses based on copulationsoccurring during the entire 3 h. In addition to our analysesbased on the numbers of copulating versus noncopulatingpairs, we also used analysis of variance (ANOVA) to deter-mine if female species, male species, or their interaction in-fluenced the duration of time to begin copulation for differentpair combinations. In particular, we were interested in wheth-er the time to copulation was greater for heterospecific cross-es than conspecific crosses.
Comparison of field- and laboratory-based measures ofsexual isolation
To compare the laboratory and field estimates of sexualisolation, we compared overall patterns of PSI. In addition,we estimated the joint isolation between pure parental morphsin the field, using IPSI, a modified joint isolation statistic(Merrel 1950) developed by Rolan-Alvarez and Caballero(2000). This statistic is a modification of the widely usedjoint isolation statistic, I (Merrel 1950). IPSI is substantiallyless prone to bias (Rolan-Alvarez and Caballero 2000; E.Rolan-Alvarez, pers. comm.) than is I (the standard jointisolation statistic of Merrel [1950], which is influenced bythe effects of mating propensity and the number of individ-uals of each type in the mating pool; Gilbert and Starmer1985). IPSI is calculated as:
(PSI 1 PSI ) 2 (PSI 1 PSI )CC AA AC CAI 5 , (2)PSI (PSI 1 PSI 1 PSI 1 PSI )CC AA AC CA
where PSICC is the PSI estimate for male and female C. cob-altinus, PSIAC is the PSI estimate for male C. auratus withfemale C. cobaltinus, and so forth. Values of IPSI can theo-retically range from 21 to 1, with IPSI . 0 indicating sexualisolation between species.
Because the laboratory studies of sexual isolation did notinclude hybrid individuals, we again restricted our analysesof the field data to only those pairs in which both individualswere of the parental phenotype. We obtained bootstrappedpairwise estimates of PSI for each pairwise combination ofparental males and females in both the field and the labo-ratory. These PSI estimates were used to estimate IPSI and itssignificance, using the BASIC program described by Cruz etal. (2004), enabling us to compare the overall strength ofsexual isolation in the laboratory and the field.
Reproductive Character Displacement
To test the hypothesis of reproductive character displace-ment, we performed a mate choice experiment to comparethe mate preferences of allopatric and sympatric male Chry-

2644 MERRILL A. PETERSON ET AL.
sochus that were presented with a simultaneous choice of onefemale of each species. All choice tests were conducted inthe laboratory over a 3-week period beginning in mid-July2001. Presumably nonvirgin beetles used in these experi-ments were collected in the field during June and July 2001and kept individually in the laboratory in 45-ml plastic tubeswith fresh cuttings of A. cannabinum. For each species, weobtained individuals from a sympatric location as well asfrom a location outside of the hybrid zone. The sympatricsource was the same focal hybrid zone site used for both thefield survey and the no-choice experiment. The allopatricsites (5.3-km SSE of Ellensburg, WA, for C. cobaltinus, siteA1 in Fig. 1; and 9.5 km SSE of Othello, WA, for C. auratus,site A2 in Fig. 1), were chosen to be approximately the samedistance from the focal hybrid zone site (90 km and 102.5km, respectively).
For the trials in the mating experiments, a single malebeetle and a female of each species were placed in the same45-ml clear plastic tube in a constant temperature room (306 1.58C). Each male was used in two trials, one trial featuringa choice of allopatric females of each species and the otherinvolving a choice of sympatric females of each species. Theorder of trials was determined at random for each male, andfemales from source populations were assigned to males atrandom. Because males from both allopatric and sympatricpopulations were used in this experiment, all pairwise com-binations of allopatric and sympatric males and females wererepresented in the design. All trials were observed until thefirst copulation. Upon the initiation of copulation, the identityof the female was noted and the three beetles were separatedinto individual tubes. For most groups of three beetles, thefirst copulation occurred on the first day of observation, butin some cases, several consecutive trials were required beforethe first copulation. In these cases, the beetles were kept intheir individual tubes between observations. Males that failedto copulate under one or both of their choices were consideredto be refractory to mating and were omitted from the analysesfor those choices in which they were refractory.
To test the hypothesis of reproductive character displace-ment, we used Fisher’s exact tests in making the followingspecific comparisons: allopatric males presented with sym-patric versus allopatric pairs of females, sympatric males pre-sented with sympatric versus allopatric pairs of females, al-lopatric males versus sympatric males presented with allo-patric pairs of females, and allopatric males versus sympatricmales presented with sympatric pairs of females. We used aBonferroni correction to adjust the alpha level for multiplecomparisons among trials involving males of each species.These comparisons allowed us to test the hypothesis thatpositive assortative mating is greater for males from the sym-patric site than from the allopatric site, as well as the hy-pothesis that positive assortative mating is greater for testsinvolving sympatric females than for tests involving allo-patric females.
Relative Abundance and the Probability ofHeterospecific Mating
To clarify the relationship between relative abundance, theprobability of hybridization, and the pattern of reproductive
character displacement of male mate choice described in thispaper, we adopted a multipronged approach. First, for anexplicit exploration of the relationship between relative abun-dance and the probability of hybridization, we performed amultiple choice experiment in the laboratory, in which wevaried the relative abundance of C. auratus and C. cobaltinus.In addition, to determine if the relationship seen in this ex-periment also exists in nature, we combined data from fieldsurveys of relative abundance and the frequency of heter-ospecific mating at sites spanning the hybrid zone. Finally,to determine the temporal stability of the relative abundanceof the two species at the focal hybrid zone site, we augmentedexisting surveys of relative abundance taken in 1999 (Peter-son et al. 2001) and 2000 (Peterson et al. 2005) with addi-tional surveys in both 2001 and 2004, giving us relative abun-dance data from this site for four years of a six-year period.These data, taken together with our analyses of the relation-ship between relative abundance and the probability of hy-bridization, provide a context in which to interpret any spe-cies-specific differences in reproductive character displace-ment in beetles from the focal hybrid zone site.
Multiple-choice experiment
Under the multiple-choice experiment, conducted in 2004,we enclosed 100 beetles (50:50 sex ratio for each species)from the focal hybrid zone site in metal, screened cages (30cm long 3 30 cm wide 3 60 cm high) in a 288C constant-temperature room, and observed mate choices over a periodof three days for each group of 100 beetles. Each cage con-tained large stalks of host plant (A. cannabinum). We variedthe relative abundance of the two species, with C. cobaltinusrepresenting 10%, 30%, 50%, 70%, and 90% of the individ-uals. For each trial, we haphazardly placed the 100 beetlesin the cage at approximately 0900 h on the first day for thattrial. During a 5-min period following an initial 1.5-h accli-mation period in the constant-temperature room, we countedthe number of copulating pairs of each type. We repeatedthese observations at 1.5-h intervals for a total of five ob-servations per day for three consecutive days. Prior to be-ginning the trials each day, we replaced any dead or dyingindividuals with vigorous individuals of the same sex andspecies. Our rationale for using a 1.5-h interval between ob-servations is that copulation typically lasts less than 20 min(Dickinson 1995), making it highly unlikely that a singlecopulation event would span two observations. Thus, the cop-ulations seen at different observation times would be inde-pendent copulations, much like different copulations ob-served at different times within a natural population.
Because of the large number of beetles required, we per-formed the observations using a single group of 100 beetlesper relative abundance treatment. Although the focal site hasrelatively large populations of both species, obtaining enoughbeetles to run independent replicates for each relative abun-dance treatment without severely impacting the source pop-ulation would have been impossible. Instead, individualsused in each trial were randomly drawn from a pool of beetles(293 C. auratus males, 248 C. auratus females, 113 C. cob-altinus males, and 99 C. cobaltinus females) collected in thefield on 22 July 2004. However, to assess whether single

2645REINFORCEMENT IN A BEETLE HYBRID ZONE
FIG. 2. Frequencies of mating pairs of Chrysochus auratus (A) andC. cobaltinus (C) observed in the field (shaded bars) and expectedfrequencies under random mating (white bars). The frequencies ofdifferent mating pair types differed from expectations under randommating (G3 5 10.60, P , 0.025) in a manner typical of positiveassortative mating.
TABLE 1. Pair sexual isolation (PSI) between pairwise combina-tions of Chrysochus at the focal hybrid zone site. Bootstrappedestimates of standard deviations are in parentheses. See Materialsand Methods for interpretation of PSI estimates.
Males
Females
C. auratus Hybrids C. cobaltinus
C. auratus 1.27*(0.07) 0.67(0.21) 0.58*(0.12)Hybrids 0.90 (0.19) 1.53(0.52) 0.98 (0.26)C. cobaltinus 0.68*(0.09) 1.18(0.30) 1.59*(0.18)
* P , 0.05 after sequential Bonferroni adjustment.
groups of 100 beetles produced representative results for aparticular relative abundance, we ran three replicate trials ata 50:50 ratio of the two species. For each of the 15 obser-vations for each group of 100 beetles, we determined theproportion of copulating individuals of each sex and speciesthat were with a heterospecific mate. To compare the threereplicates at a 50:50 relative abundance, we ran a separateANOVA using only those three replicate groups, using an-gular-transformed probabilities of mating with a heterospe-cific individual (Sokal and Rohlf 1981). Subsequently, weperformed ANOVA to determine the effects of relative abun-dance, sex, species, and their interactions on the angular-transformed probability of heterospecific mating.
Correlation between relative abundance and the probabilityof hybridization in the field
In early to mid July 2000, we surveyed relative abundanceand the frequency of heterospecific versus conspecific pair-ings at 22 sites spanning the hybrid zone (Fig. 1). For themost part, these surveys involved exhaustive searches ofsmall patches of dogbane, but for the large focal hybrid zonepatch, we used transect counts, as reported previously (Pe-terson et al. 2005). The results of these surveys allow us toquantify the relationship between relative abundance and theprobability of hybridization. We report only those data fromsurveys during which we observed at least 10 pairs at a site.Under this criterion, we obtained sufficient data from sevendifferent sites (sites 1, 2, 3, 14, 17, 18, 22 in Fig. 1). Todetermine the influence of relative abundance on the prob-ability of hybridization, we performed linear regression, us-ing the angular-transformed proportion of all paired individ-uals that were with a heterospecific mate. As before, we ex-cluded pairings with hybrid individuals from these analyses.Because of the small number of datapoints for each-sex-byspecies combination, we pooled all of the sex- and species-specific estimates of heterospecific mating probability into asingle regression analysis. Thus, sites that were included in
the analysis could contribute up to four datapoints for theregression (one datapoint for each sex of each species). Thispooling better enabled us to assess the general relationshipbetween abundance and the probability of hybridization.However, pooling also prevented us from teasing apart theinfluence of species, sex, and their interaction on this rela-tionship in natural populations.
Relative abundance and the probability of hybridization atthe focal hybrid zone site
To supplement our earlier surveys of relative abundanceat the focal site, taken in 1999 (Peterson et al. 2001) and2000 (Peterson et al. 2005), we conducted additional surveysin 2001 and 2004 at the same site. In 2001, we estimated therelative abundance of C. auratus and C. cobaltinus from 319teneral adults and pupae that we collected from undergroundpupation chambers in early June. On 22 July 2004, we as-sessed the relative abundance of adults on host plants, usingthe same survey methods as employed previously (Petersonet al. 2005). To estimate the relative probabilities of hybrid-ization for C. auratus and C. cobaltinus at the focal site, wedetermined the proportion of paired individuals that wereengaging in either copulation or postcopulatory contact withheterospecific beetles on 1 July 2000 and 9 July 2000 (forsurvey procedure see Peterson et al. 2005), augmenting re-sults from 1999 (Peterson et al. 2001). For each sex, wecompared the frequency of C. auratus and C. cobaltinuspaired with conspecific versus heterospecific individuals, us-ing a G-test of independence.
RESULTS
Assortative Mating and Sexual Isolation
Of the 1824 beetles encountered during the 1 July 2000survey, there were 179 pairs, accounting for 19.6% of theindividuals. The relative frequencies of different mating com-binations differed from what would be expected under anassumption of random mating (G3 5 10.75, P , 0.025, Fig.2), based on the relative abundance of C. auratus, C. cob-altinus, and F1 hybrids encountered during that survey (re-ported in Peterson et al. 2005). Examination of the nonran-dom mating pattern reveals a deficit of heterospecific pairsof both types and an excess of homospecific pairs, particularlythose involving C. cobaltinus. Such a result is indicative ofpositive assortative mating. Values of PSI for heterospecificpairs were less than one, while PSI for conspecific pairs ex-ceeded one (Table 1), a result that was repeated if F1 hybrids

2646 MERRILL A. PETERSON ET AL.
TABLE 2. Proportion of pairs in no-choice experiment mating with-in 0.5 h and within 3 h. The sample size (N) indicates the numberof trials for each treatment.
Treatment 0.5 h 3 h N
Chrysochus auratus male 3C. auratus female 0.26 0.57 86
C. auratus male 3 C. cobaltinus female 0.15 0.40 85C. cobaltinus male 3 C. auratus female 0.11 0.35 86C. cobaltinus male 3 C. cobaltinus female 0.38 0.62 85
TABLE 3. Pair sexual isolation (PSI) estimates for pairwise com-binations of Chrysochus in the no-choice experiment in the labo-ratory, based on matings occurring within 0.5 h. Bootstrapped es-timates of standard deviations are in parentheses.
Males
Females
C. auratus C. cobaltinus
C. auratus 1.55*(0.18) 0.63*(0.11)C. cobaltinus 0.54*(0.13) 1.32*(0.11)
* P , 0.05 after sequential Bonferroni adjustment.
FIG. 3. Mean time to copulation (6SE) for the four pairwise com-binations of Chrysochus auratus and C. cobaltinus individuals in 3-h laboratory no-choice trials. White bars represent C. auratus fe-males and shaded bars represent C. cobaltinus females. Pairs thatdid not copulate during the 3-h trial were omitted. Mean time tocopulation was significantly influenced by female species (F1,162 59.463, P 5 0.002) and a male species by female species interaction(F1,162 5 3.935, P 5 0.049) but not by male species (F1,162 5 0.574,P 5 0.45).
were omitted from the analysis (not shown). This result isconsistent with the hypothesis that positive assortative matingat this site is explained at least in part by sexual isolationbetween C. auratus and C. cobaltinus (Cruz et al. 2004).
In the laboratory no-choice experiment, the order in whichfemales were presented conspecific and heterospecific malesdid not affect the outcome (C. auratus females: G1 5 0.24,P . 0.25, C. cobaltinus females: G1 5 0.63, P . 0.25). Thus,for all subsequent analyses, we pooled all females of a givenspecies. It has been suggested that sexual isolation indicesare compromised in experiments in which more than 50% ofthe potential matings occur (Casares et al. 1998). Becausethis was the case for both conspecific pair types after 3 h,but was not the case for the first 0.5 h of the mating trials(Table 2), we report the results of our analyses of the datafor the first 0.5 hour of each trial. We repeated all analysesusing the results over the entire 3 h of each trial, and for allof the results reported below, we found no qualitative dif-ferences from the results for the first 0.5 h.
As we found in the field, there was a strong pattern ofsexual isolation in the laboratory, indicated by the fact thatthe proportion of heterospecific pairs mating in the no-choiceexperiment was lower than that of conspecific pairs (Table2). PSI analyses quantified this difference formally; PSI val-ues from the no-choice experiment exceeded one for con-specific pairs and were less than one for heterospecific pairs(Table 3), as expected for sexual isolation (Cruz et al. 2004).The similarity between field and laboratory measures of sex-ual isolation was also revealed by comparisons of the overallstrength of sexual isolation (IPSI). Specifically, these esti-mates were quite similar for the no-choice experiment IPSI 50.424 (SD 5 0.107) and the field IPSI 5 0.393 (SD 5 0.087).The fact that both were greater than zero (P , 0.001) indi-cated that C. auratus and C. cobaltinus exhibit significantsexual isolation in both the laboratory and the field.
An interesting result of the no-choice experiment was that,for pairs involving a C. cobaltinus male, the probability ofcopulation in the first 0.5 h if the female was conspecific was3.58 times greater than if the female was heterospecific (Table2). In contrast, for pairs involving a C. auratus male, theconspecific mating probability within the first 0.5 h was only1.67 times that of the heterospecific mating probability. Giv-en the apparent lack of female choice in this system, thisdifference suggests that C. cobaltinus males are more dis-criminating than C. auratus males.
Analyses of time to copulation (restricted to only thosepairs that copulated) revealed a significant effect of femalespecies, with copulation occurring more slowly in trials in-volving C. auratus females than those with C. cobaltinus
females (F1,162 5 9.463, P 5 0.002; Fig. 3). There was noeffect of male species (F1,162 5 0.574, P 5 0.45), but therewas a significant interaction between female species and malespecies (F1,162 5 3.935, P 5 0.049), due to the apparentlygreater mate discrimination by C. cobaltinus males than byC. auratus males (Fig. 3). Specifically, for trials involvingC. cobaltinus males, the time to copulation was substantiallylonger if the female was heterospecific than if she was con-specific. In contrast, the difference between the two treat-ments involving C. auratus males was minimal (Fig. 3). Thisdifference between times to copulation for trials with C. au-ratus versus C. cobaltinus males mirrored the results for thespecies-specific probabilities of heterospecific copulation bymales, again indicating that C. cobaltinus males are morediscriminating than C. auratus males.
Reproductive Character Displacement
As predicted from reinforcement theory, the mating ex-periments revealed a pattern consistent with reproductivecharacter displacement. When given a choice of conspecificand heterospecific females from the hybrid zone, sympatricmale C. cobaltinus were significantly more likely to choosethe conspecific female than were males from the allopatricpopulation (P , 0.01 after sequential Bonferroni adjustment;
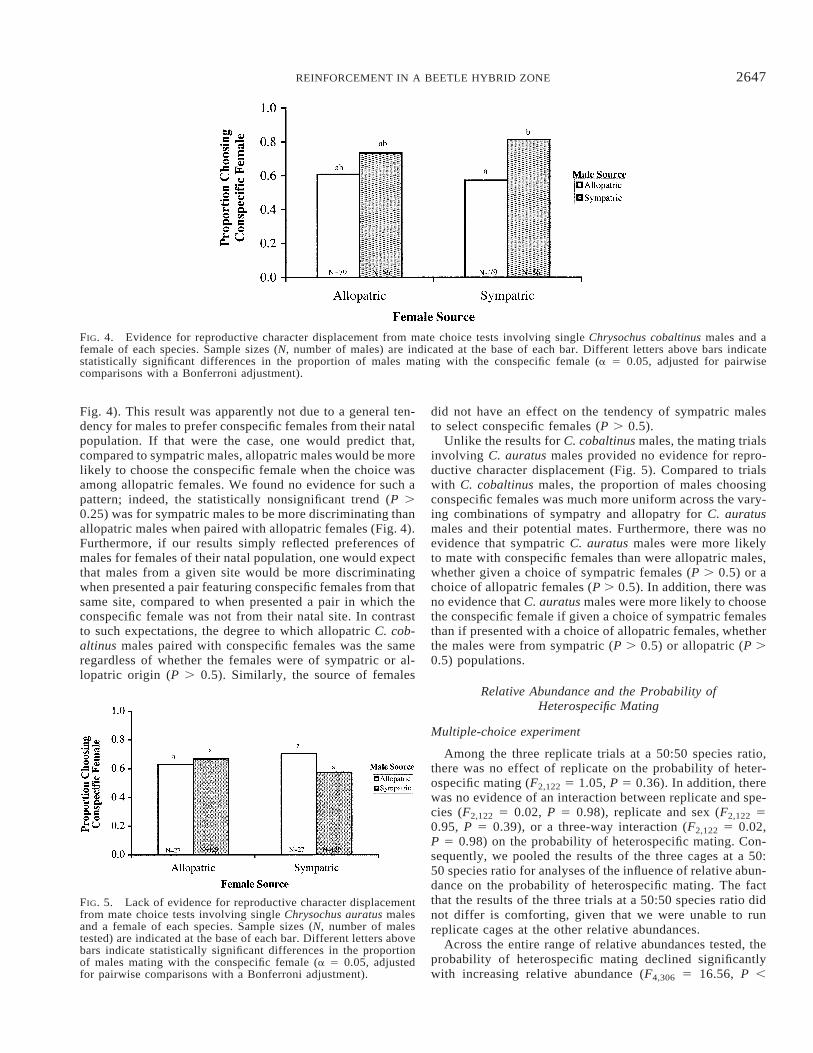
2647REINFORCEMENT IN A BEETLE HYBRID ZONE
FIG. 4. Evidence for reproductive character displacement from mate choice tests involving single Chrysochus cobaltinus males and afemale of each species. Sample sizes (N, number of males) are indicated at the base of each bar. Different letters above bars indicatestatistically significant differences in the proportion of males mating with the conspecific female (a 5 0.05, adjusted for pairwisecomparisons with a Bonferroni adjustment).
FIG. 5. Lack of evidence for reproductive character displacementfrom mate choice tests involving single Chrysochus auratus malesand a female of each species. Sample sizes (N, number of malestested) are indicated at the base of each bar. Different letters abovebars indicate statistically significant differences in the proportionof males mating with the conspecific female (a 5 0.05, adjustedfor pairwise comparisons with a Bonferroni adjustment).
Fig. 4). This result was apparently not due to a general ten-dency for males to prefer conspecific females from their natalpopulation. If that were the case, one would predict that,compared to sympatric males, allopatric males would be morelikely to choose the conspecific female when the choice wasamong allopatric females. We found no evidence for such apattern; indeed, the statistically nonsignificant trend (P .0.25) was for sympatric males to be more discriminating thanallopatric males when paired with allopatric females (Fig. 4).Furthermore, if our results simply reflected preferences ofmales for females of their natal population, one would expectthat males from a given site would be more discriminatingwhen presented a pair featuring conspecific females from thatsame site, compared to when presented a pair in which theconspecific female was not from their natal site. In contrastto such expectations, the degree to which allopatric C. cob-altinus males paired with conspecific females was the sameregardless of whether the females were of sympatric or al-lopatric origin (P . 0.5). Similarly, the source of females
did not have an effect on the tendency of sympatric malesto select conspecific females (P . 0.5).
Unlike the results for C. cobaltinus males, the mating trialsinvolving C. auratus males provided no evidence for repro-ductive character displacement (Fig. 5). Compared to trialswith C. cobaltinus males, the proportion of males choosingconspecific females was much more uniform across the vary-ing combinations of sympatry and allopatry for C. auratusmales and their potential mates. Furthermore, there was noevidence that sympatric C. auratus males were more likelyto mate with conspecific females than were allopatric males,whether given a choice of sympatric females (P . 0.5) or achoice of allopatric females (P . 0.5). In addition, there wasno evidence that C. auratus males were more likely to choosethe conspecific female if given a choice of sympatric femalesthan if presented with a choice of allopatric females, whetherthe males were from sympatric (P . 0.5) or allopatric (P .0.5) populations.
Relative Abundance and the Probability ofHeterospecific Mating
Multiple-choice experiment
Among the three replicate trials at a 50:50 species ratio,there was no effect of replicate on the probability of heter-ospecific mating (F2,122 5 1.05, P 5 0.36). In addition, therewas no evidence of an interaction between replicate and spe-cies (F2,122 5 0.02, P 5 0.98), replicate and sex (F2,122 50.95, P 5 0.39), or a three-way interaction (F2,122 5 0.02,P 5 0.98) on the probability of heterospecific mating. Con-sequently, we pooled the results of the three cages at a 50:50 species ratio for analyses of the influence of relative abun-dance on the probability of heterospecific mating. The factthat the results of the three trials at a 50:50 species ratio didnot differ is comforting, given that we were unable to runreplicate cages at the other relative abundances.
Across the entire range of relative abundances tested, theprobability of heterospecific mating declined significantlywith increasing relative abundance (F4,306 5 16.56, P ,

2648 MERRILL A. PETERSON ET AL.
FIG. 6. The relationship between relative abundance and the mean probability (6SE) that copulating individuals were paired withheterospecific mates. Probabilities of such mating mistakes are shown for Chrysochus cobaltinus males (filled circles), C. cobaltinusfemales (open circles), C. auratus males (filled squares), and C. auratus females (open squares). Datapoints are slightly offset at eachrelative abundance value for clarity. In addition, expectations for random copulation are indicated by the triangles and dashed line.
TABLE 4. Relative abundance of Chrysochus auratus, C. cobaltin-us, and hybrids at 22 hybrid zone sites surveyed in July 2000.Almost all individuals categorized as hybrids are F1 hybrids (Pe-terson et al. 2005).
Site
Relative abundance (prop. of total)
C. auratus C. cobaltinus HybridsEstimated total
no. beetles
1 0.02 0.90 0.08 1332 0.01 0.89 0.10 1393 0.54 0.23 0.22 3614 0.00 0.50 0.50 25 0.17 0.67 0.17 126 0.00 0.98 0.02 887 0.00 0.96 0.04 258 0.00 0.98 0.02 1639 0.00 0.99 0.01 137
10 0.00 0.89 0.11 911 0.33 0.67 0.00 312 0.25 0.61 0.14 5113 0.47 0.42 0.11 9114 0.98 0.01 0.01 18815 0.50 0.50 0.00 216 0.85 0.06 0.09 3317 0.57 0.27 0.17 .10,00018 0.32 0.55 0.13 21419 0.67 0.10 0.23 5220 0.56 0.22 0.22 921 0.23 0.69 0.08 1322 0.96 0.01 0.03 152
0.001; Fig. 6). Again, there was no effect of the sex (F1,3065 0.66, P 5 0.42) or species (F1,306 5 0.07, P 5 0.79) ofbeetle on the probability of hybridization. Furthermore, therewere no significant interactions between sex and relative
abundance (F4,306 5 0.98, P 5 0.42); species and relativeabundance (F4,306 5 0.561, P 5 0.691); or sex, species, andrelative abundance (F4,306 5 0.22, P 5 0.93). Particularlywhen relative abundance was at least 0.3, the species- andsex-specific probability of heterospecific copulation was lessthan that expected under random mating (Fig. 6), consistentwith the pattern of sexual isolation reported above for boththe laboratory and the field.
Correlation between relative abundance and the probabilityof hybridization in the field
Our surveys at sites spanning the hybrid zone in July 2000showed that the focal hybrid zone site harbored substantiallymore beetles than any other site (Table 4). Indeed, our tran-sect counts at that site produce 1824 beetles (Peterson et al.2005) using a method that sampled approximately 8% of thetotal area of the site. Thus, this site almost certainly has morethan 10,000 beetles. In contrast, no other site surveyed con-tained more than 400 beetles (Table 4). The unusually largenumber of beetles at the focal site is due to its having botha large patch of host plants and high densities of beetles.While other patches surveyed had either many host plants orsimilarly high densities, none featured both. It is noteworthy,however, that for allopatric populations of both species, wehave encountered occasional patches featuring populationsizes that approach those seen in our focal hybrid zone site(M. A. Peterson, K. Monsen, E. Larson, M. Brassil, D. Juarez,K. Buckingham, L. Tantalo, and J. Holland, unpubl. data).In general, sites in the northwest portion of the hybrid zonetended to be dominated by C. cobaltinus, whereas most of

2649REINFORCEMENT IN A BEETLE HYBRID ZONE
TABLE 5. Relative abundance of Chrysochus auratus, C. cobaltin-us, and hybrids on four dates at the focal hybrid zone site.
Date
Proportion of total
C. auratus C. cobaltinus Hybrids N
21 July 1999* 0.59 0.29 0.12 6991 July 2000 0.57 0.27 0.17 18243 and 5 June 2001 0.46 0.28 0.25 319
22 July 2004 0.63 0.20 0.17 928
* Source: Peterson et al. (2001).
TABLE 6. Proportion of copulations by males and females of eachChrysochus species with conspecific, heterospecific, and hybridmates at the focal hybrid zone site on three dates. The sample size(N) indicates the number of paired individuals of each type en-countered on each date.
Identity
Proportion copulating with
Conspecificmate
Heterospecificmate Hybrid mate N
C. auratus males21 July 1999* 0.55 0.29 0.14 77
1 July 2000 0.75 0.17 0.08 859 July 2000 0.70 0.16 0.14 189
C. auratus females21 July 1999* 0.55 0.29 0.14 77
1 July 2000 0.60 0.24 0.16 1069 July 2000 0.63 0.24 0.14 211
C. cobaltinus males21 July 1999* 0.31 0.56 0.13 39
1 July 2000 0.45 0.40 0.15 629 July 2000 0.35 0.51 0.14 99
C. cobaltinus females21 July 1999* 0.31 0.56 0.13 39
1 July 2000 0.53 0.26 0.17 539 July 2000 0.43 0.38 0.10 82
* In 1999, the probability of hybridization was assessed for each species,but sex-specific probabilities were not distinguished, so the values reportedare species-specific values.
the sites in the vicinity of the focal hybrid zone site andeastward were dominated by C. auratus (Table 4, Fig. 1).
Across sites, there was a significant negative relationshipbetween relative abundance and the probability of hybrid-ization; arcsine(proportion hybridizing)0.5 5 1.40 2 1.34(relative abundance) (P , 0.001, 5 0.85), mirroring the2Radjgeneral pattern seen in the multiple-choice experiment (Fig.6). At sites in which a species is scarce, individuals of thatspecies nearly always hybridize, whereas the abundant spe-cies at such sites hybridizes extremely infrequently.
Relative abundance and the probability of hybridization atthe focal hybrid zone site
At the focal hybrid zone site, C. auratus consistently out-numbered C. cobaltinus on all four sampling dates, with theratio of C. auratus:C. cobaltinus ranging from 1.6:1 to 3.3:1 (Table 5). As expected from the above-described relation-ship between relative abundance and the probability of hy-bridization, the two species also differed in the probabilityof heterospecific mating at this site. Specifically, on all threedates on which we assessed hybridization frequencies, themajority of C. auratus males and females were paired withconspecific individuals, while only 16–29% were paired withheterospecific individuals (Table 6). In contrast, individualsof the relatively scarce C. cobaltinus were typically pairedwith conspecific mates less than 50% of the time, and onaverage were paired with heterospecific mates as often aswith conspecific mates (Table 6). As expected from the rel-ative abundance of the two species, the ratio of males matingwith heterospecific versus conspecific females was muchgreater for C. cobaltinus males than for C. auratus males (G15 12.82, P , 0.001; G1 5 39.58, P , 0.001 for 1 and 9 July2000, respectively). Although less extreme, this differencewas mirrored in C. auratus and C. cobaltinus females; theratio of females mating with heterospecific versus conspecificmales was significantly greater for C. cobaltinus than C. au-ratus on 9 July (G1 5 8.10, P , 0.005), but not on 1 July(G1 5 0.37, P . 0.25). We are unable to make similar com-parisons using the 1999 data, because we did not separatethe sex-specific mating frequencies in that year.
DISCUSSION
Sexual Isolation and Bimodality in the ChrysochusHybrid Zone
By using a combination of laboratory and field studies, wefound strong evidence for positive assortative mating in theChrysochus hybrid zone and demonstrated that sexual iso-
lation plays a prominent role in generating the pattern ofassortative mating. First, our results from the field indicatedsignificant deviations from random mating that were consis-tent with positive assortative mating. Measures of pairwisesexual isolation (PSI) conformed to what would be expectedif sexual isolation were a factor contributing to that assor-tative mating. Subsequent experiments in the laboratory un-der controlled conditions corroborated this evidence of sexualisolation between the two parentals.
Compared to the essentially complete postzygotic isolationbetween C. auratus and C. cobaltinus (Peterson et al. 2005),the level of premating isolation in this system is relativelyweak. Based on the observed frequency of heterospecific cop-ulation at this site, there should be an abundance of recom-binant genotypes after only a few generations (Peterson etal. 2005). In other words, in short order, there should be achange from a bimodal distribution of genotypes (Jiggins andMallet 2000) to a unimodal distribution of genotypes typicalof a hybrid swarm. While it is possible that postmating pre-zygotic barriers such as conspecific sperm precedence (How-ard 1999) also exist in this system, such barriers are notnecessary to explain the bimodality of the Chrysochus hybridzone, given the evidence for strong postzygotic barriers inthis system. Indeed, we argue that postzygotic barriers, notweak premating barriers or possible postmating-prezygoticbarriers, maintain the bimodal structure of this hybrid zone.Our view of the maintenance of bimodality in the Chrysochushybrid zone contrasts with the argument developed by Jigginsand Mallet (2000). In their review, they found that bimodalhybrid zones typically featured strong premating or post-mating-prezygotic barriers, unlike unimodal hybrid zones.Thus, they argued that postzygotic barriers themselves may

2650 MERRILL A. PETERSON ET AL.
be insufficient to maintain bimodality in the absence of pre-zygotic barriers. However, in the Chrysochus hybrid zone,because the postzygotic barriers are essentially complete,they are sufficient to maintain bimodality. In the absence ofthose postzygotic barriers, it is unlikely that the weak sexualisolation in this hybrid zone could prevent the evolution ofunimodality (Peterson et al. 2005).
Relative Abundance and the Species-Specificityof Reinforcement
We documented a pattern of reproductive character dis-placement in beetles from the Chrysochus hybrid zone. Theseresults, taken together with evidence of extremely low hybridfitness (Peterson et al. 2005), strongly suggest that reinforce-ment has played a role in the evolution of male mating pref-erence in this system. Regardless of how reinforcement isdefined, it appears that the term is appropriate for the Chry-sochus hybrid zone, even in its strict sense, because we haveevidence of rare successful reproduction by hybrids at somehybrid zone sites (Peterson et al. 2005).
The pattern of reproductive character displacement in thissystem is one in which C. cobaltinus males from the hybridzone are more likely to mate with a conspecific female whengiven a choice of hybrid zone females than are C. cobaltinusmales from outside of the hybrid zone. Although the re-sponses of hybrid zone and allopatric C. cobaltinus males toallopatric females showed a similar pattern (indicating thatour evidence for reproductive character displacement was notsimply due to a general preference by males for females fromtheir natal population), the difference was reduced and non-significant. Given that precopulatory female choice appar-ently plays a minimal role in this system, our interpretationof these results is that hybrid-zone C. cobaltinus males aremore discriminating than allopatric males and that hybrid-zone females are somewhat more distinguishable than allo-patric females. In contrast to the pattern of reproductive char-acter displacement in the mating preferences of C. cobaltinusmales, there was no evidence for reproductive character dis-placement in C. auratus males. The species-specificity of thereinforcement of male mate choice is consistent with resultsfrom the no-choice experiment, which demonstrated that C.cobaltinus males from the hybrid zone are more discrimi-nating than are C. auratus males from the same location.
It is notable that we were able to detect reproductive char-acter displacement of male choice even though sexual iso-lation in the hybrid zone is rather weak. This result shouldbe encouraging to others working in systems with weak sex-ual isolation, in that it suggests that one does not need in-ordinately large sample sizes in such cases to detect differ-ences in the mate preferences of allopatric and sympatricpopulations.
Species-specific reinforcement has been documented for atleast two other hybrid zones. In one case, male sailfin mollies(Poecilia latipinna) show increasing discrimination in areasof overlap with a parthenogenetic molly species (Poeciliaformosa), whose females must copulate with heterospecificmales to initiate egg development (Gabor and Ryan 2001).Because heterospecific copulation is advantageous (indeednecessary) for P. formosa females, that species has not
evolved a tendency to avoid those copulations in areas ofoverlap, in contrast to P. latipinna. Similarly, in a pair ofhybridizing toad species, Spea bombifrons and Spea multi-plicata, the cost of hybridization differs for females of thetwo species, due to the fact that hybrid fitness depends onthe direction of the heterospecific cross (Pfennig and Si-movich 2002). In both of these systems, the species speci-ficity of reinforcement results from the fact that hybridizationis advantageous for one species and disadvantageous for theother.
In contrast, for the Chrysochus hybrid zone, the uniformlylow fitness of hybrids suggests that hybridization should bedisadvantageous for both species. For this system, the reasonfor a species-specific pattern of reinforcement apparently isinherent differences in relative abundance, which in turncause differences in the strength of selection against hybrid-ization. In particular, we documented both in the multiple-choice experiment and in the field that the probability of suchmating mistakes is indeed inversely related to relative abun-dance. At the focal hybrid zone site that was our source forsympatric beetles in the reproductive character displacementexperiment, C. auratus has been the numerically dominantspecies in all of our surveys spanning a six-year period. Fur-thermore, the proportion of C. cobaltinus individuals matingwith heterospecific individuals at this site is also consistentlygreater than for C. auratus. Since mate guarding by Chry-sochus males substantially reduces their probability of re-mating (Dickinson 1995), mating mistakes are almost cer-tainly costly for males. Thus, based on their relative abun-dance, it appears that C. cobaltinus males are under strongerselection against hybridization than C. auratus males, ex-plaining why C. cobaltinus males exhibited reproductivecharacter displacement, whereas C. auratus males did not.
We established both that sexual isolation results in a pat-tern of positive assortative mating and that reinforcement hasdriven increased mate discrimination by C. cobaltinus males.However, observations of frequent heterospecific mating byC. cobaltinus males at the focal hybrid zone site raise thequestion of whether C. cobaltinus males are indeed less likelyto engage in heterospecific mating in multiple-choice sce-narios, after taking into account the effect of relative abun-dance. If this were the case, one might expect the residualsfor C. cobaltinus males in the relationship in Figure 6 to begenerally negative. In fact, although C. cobaltinus males havethe lowest mean residuals, they are not significantly negative,nor are they significantly lower than the residuals for C. au-ratus males or for females of either species. This patternsuggests a possible inconsistency between our evidence forthe reproductive character displacement of C. cobaltinus malemate preference and the frequency with which those malesengage in heterospecific matings in both the laboratory andthe field. The most plausible explanation for this apparentinconsistency is that the enhancement of sexual isolation viathe reproductive character displacement of C. cobaltinus malemate preference is too subtle to have a noticeable effect onmate choices in multiple-choice scenarios, at least with thesample sizes employed in this study. This explanation is plau-sible, particularly given that even in the hybrid zone, sexualisolation is fairly weak. Thus, it appears that reinforcementin this system has not yet driven the evolution of male mate

2651REINFORCEMENT IN A BEETLE HYBRID ZONE
preferences that are sufficiently strong to leave a strikingsignature on patterns of mate choice in the field.
Other Reasons for the Lack of Reproductive CharacterDisplacement in Chrysochus auratus
In addition to the apparent influence of relative abundanceon the species specificity of selection against hybridization,there are several potential explanations for the lack of re-productive character displacement in C. auratus that warrantdiscussion. One such explanation is that our ability to detectreproductive character displacement in C. auratus was com-promised by the relatively small sample size for males ofthat species. We view this as unlikely, because in the trialsin which C. auratus males had a choice of allopatric females,the proportions of allopatric and sympatric males mating withthe conspecific female were nearly identical. Furthermore,for trials involving sympatric females, the nonsignificant dif-ference between allopatric and sympatric males was in theopposite direction of the pattern predicted under reinforce-ment.
It may also be that the ability of sympatric C. auratus torespond to selection against hybridization is lower than thatof C. cobaltinus. For instance, theory has often demonstratedthat gene flow from naive allopatric populations may counterthe effects of selection against hybridization (Sanderson1989; Liou and Price 1994; Servedio and Kirkpatrick 1997).A comparative analysis of population genetic structure in C.auratus and C. cobaltinus (Dobler and Farrell 1999) foundsignificant population-genetic structure in C. cobaltinus butremarkable homogeneity in C. auratus. Differences amongthe two species in levels of gene flow may thus underlie thespecies-specific pattern of reproductive character displace-ment. In addition to among-population homogeneity, Doblerand Farrell (1999) documented comparatively little geneticvariation within populations of C. auratus compared to C.cobaltinus. This lack of variation in C. auratus may be dueto bottlenecks associated with postglacial range expansion,as has been documented in other species (Hewitt 1996; Ber-natchez and Wilson 1998). Thus, even if the strength of se-lection against hybridization acting on the two species werethe same, that selection would not lead to reproductive char-acter displacement in C. auratus if the species lacked suffi-cient genetic variation to respond to such selection. To ourknowledge, no studies have examined the role of recent de-mographic history in the process of reinforcement withinzones of secondary contact, and we view this as a potentiallyfruitful avenue of inquiry for future reinforcement research.
The possibility that C. auratus males may have a reducedability to respond to selection against hybridization shouldnot be viewed as undermining our previous arguments thatC. auratus males face weaker selection against hybridizationdue to their numerical dominance. Instead, it is likely thatsuch explanations are complementary; C. auratus males ap-pear to face weaker selection against hybridization than C.cobaltinus males, but may also have a reduced ability torespond to that selection. Ultimately, such a combination offactors may provide the best explanation for the species spec-ificity of reproductive character displacement.
One additional explanation for the lack of reproductive
character displacement in C. auratus male mating preferencefollows recent theory demonstrating, somewhat counterin-tuitively, that reinforcement may increase sexual isolation ina hybrid zone without producing a pattern of reproductivecharacter displacement. Specifically, simulations by Lemmonet al. (2004) showed that when selection against hybrids isimposed on the middle of a cline across which exists a gradualgradient in mate preference, that selection may increase thestrength of mating discrimination in the vicinity of the hybridzone, but not enough to match the extremes in mate pref-erence at the ends of the cline. Such a situation almost cer-tainly does not apply to the Chrysochus hybrid zone, whichapparently formed as a result of secondary contact followingapproximately 2–3 million years of allopatric divergence(Dobler and Farrell 1999; Peterson et al. 2001). Thus, uponthe initial formation of this hybrid zone, it is likely not onlythat the gradient in mate preference from one side of thehybrid zone to the other was abrupt, but also that selectionagainst hybridization was imposed immediately due to thenear sterility of hybrids.
Why Might Hybridization Frequency Not Correlate withSelection against Hybridization?
There are several reasons why the strength of selectionagainst hybridization for Chrysochus males at the focal hybridzone site might not be correlated with the frequency of hy-bridization at that site. Each of these reasons could undermineour argument that the species specificity of reproductive char-acter displacement reflects the species-specific action of re-inforcement that is due to differences in relative abundance.However, we view each of these possibilities as unlikely.
First, the species-specific pattern of selection against hy-bridization has probably differed among sites within the hy-brid zone due to spatial variation in relative abundance. Ifso, gene flow from nearby populations may have influencedthe evolution of male choice in our focal hybrid zone site,reducing the importance of relative abundance at that site.However, relative abundance in the sites closest to the focalsite was generally similar to that in the focal site. Further-more, the focal site harbored substantially larger beetle pop-ulations than any of the other sites in the hybrid zone. Thus,it is far more likely that the evolution of male choice in thefocal population has influenced the evolution of male choicein nearby populations than vice versa.
Hybridization frequency also might not correlate with se-lection against hybridization if male reproductive success(total progeny sired) is not correlated with lifetime matingsuccess (total matings). In other words, male reproductivesuccess may be maximized after a few matings, such that anyadditional matings (even with conspecific females) would notenhance fitness. However, it would be difficult to understandwhy males mate daily throughout their lives (Dickinson 1995)if this were the case. Furthermore, although males may ex-perience diminishing fitness returns with each subsequentmating (Wedell et al. 2002; Lewis 2004), males do generallyachieve greater lifetime reproductive success with additionalcopulations (Bateman 1948; Parish and Coulson 1998; Joneset al. 2002; Lewis 2004). Thus, there is little reason to believe

2652 MERRILL A. PETERSON ET AL.
that Chrysochus male reproductive success is independent oflifetime mating success.
There may be no cost of heterospecific matings by malesif such matings do not reduce the reproductive success thata male can achieve via conspecific matings. For instance,Chrysochus males may vary in their choosiness within a pop-ulation. As a result, indiscriminate males may mate moreoften than choosy males, even if some of the matings by theindiscriminate males are with heterospecific females. As longas the total number of offspring sired via conspecific copu-lations by indiscriminate males is at least as great as that forchoosy males, the indiscriminate strategy would not be dis-advantageous. However, given that copulation and ejacula-tion are often costly for males (Carroll 1994; Wedell et al.2002; Lewis 2004), it is likely that males would nonethelessbenefit by saving their ejaculate for conspecific females. Fur-thermore, because the time spent in copulation and postcop-ulatory mate guarding by Chrysochus males substantially re-duces their subsequent opportunities to mate (Dickinson1995), mating mistakes by Chrysochus males would reducethe number of times that they can mate with conspecific fe-males. This seems particularly true for C. cobaltinus malesat the focal hybrid zone site, because these males copulatewith heterospecific females approximately half of the time.Thus, we feel that hybridization frequency does serve as agood proxy for the relative strength of selection against hy-bridization faced by males, a conclusion that is supported bythe species-specific patterns of reproductive character dis-placement in male mate preference. In sum, it appears thatthe reproductive character displacement of male C. cobaltinusmating preference has resulted from selection against hy-bridization that stems from its relatively high probability ofencountering and mating with heterospecific females and theinherent cost of such mating mistakes for those males.
Nonreinforcement Explanations for ReproductiveCharacter Displacement
Our argument that the pattern of reproductive characterdisplacement in the Chrysochus hybrid zone is due to rein-forcement also hinges on our ability to rule out other possibleexplanations of that pattern. Several causes of a pattern ofreproductive character displacement that are distinct fromreinforcement have been proposed (Butlin 1987; Noor 1999;Servedio 2001). One such explanation comes from theoryshowing that direct selection on either mating preferences orthe traits that influence those preferences can have a strongereffect than reinforcement (Servedio 2001, 2004). For ex-ample, if hybrid zones result in ecological character displace-ment, such that one or both species occupy niches that aredifferent from those occupied in allopatry, those novel nichescould impose direct selection on mating preferences (e.g.,Albert and Schluter 2004). In such a case, certain types ofmating signals may be more effective in the niche used insympatry than in the niche used in allopatry. As a result, apattern mimicking reproductive character displacement dueto reinforcement could emerge (Servedio 2001). Althoughthis mechanism is intuitively appealing, we have no evidencethat ecological character displacement has evolved in theChrysochus hybrid zone. The two species exploit the same
host plant in both sympatry and allopatry and appear to oc-cupy the same niche in both situations. It is noteworthy thatin the hybrid zone, a second host plant used by C. cobaltinusin the region, A. speciosa, is used by only a small proportionof C. cobaltinus individuals (M. A. Peterson, pers. obs.).Thus, it appears unlikely that direct selection on mating pref-erences, resulting from ecological character displacement,could play a prominent role in the evolution of prematingbarriers in this system.
Another form of direct selection on mate preferences mightoccur if heterospecific copulation results in injury to males(e.g., Sota and Kubota 1998; Servedio 2001). Although sucha scenario differs from the classical model of reinforcement,in which selection is imposed via low hybrid fitness, it wasrecently suggested that it should be treated as a form ofreinforcement (Servedio and Noor 2003). Furthermore, wehave never witnessed copulation-related injuries to males, inspite of observing hundreds of heterospecific matings in thelaboratory.
An additional explanation that has been proposed for pat-terns of reproductive character displacement is the differ-ential fusion of populations in areas of overlap (Noor 1999).Specifically, populations that already had divergent matingsystems upon contact might persist as distinct sympatric en-tities, while those that lacked such divergent mating systemswould fuse. Again, this is unlikely for the Chrysochus hybridzone, because the near sterility of hybrids (Peterson et al.2005) is apparently sufficient to prevent fusion.
Finally, Butlin (1987) has argued that a pattern of repro-ductive character displacement could simply be due to clinalvariation in mating systems, such that the area of overlapcoincides with extremes along a clinal gradient. Ruling outthis explanation will require assessing male mate preferencesand female mating cues from numerous allopatric and sym-patric populations. However, as has been argued previously(Noor 1995b), given the evidence for selection against hy-bridization, the more parsimonious explanation for a patternof reproductive character displacement is reinforcement.
Reproductive Character Displacement in Male MatingPreference and the Bias in Reinforcement Research
Most empirical studies of reinforcement have focused onthe evolution of either female choice or male mating cues(e.g., reviews in Howard 1993; Gabor and Ryan 2001). Thisbias reflects the long-standing view in sexual selection theorythat mating mistakes are typically more costly to femalesthan to males (Trivers 1972). Indeed, this view of the relativecosts of mating mistakes has resulted in relatively little re-search being done on the evolution of male mate choice ingeneral (Bonduriansky 2001). However, recent research onconspecific sperm precedence (reviewed in Howard 1999) hasindicated a mechanism by which females may lower the costof mating mistakes in hybrid zones; if conspecific spermprecedence is strong, females may be able to mate with het-erospecific males with impunity, as long as they have alsomated with at least one conspecific male (Marshall et al.2002). Furthermore, there is growing evidence that selectionon male mate preference may be a common occurrence, par-ticularly in systems in which there is either partial or com-

2653REINFORCEMENT IN A BEETLE HYBRID ZONE
plete sex role reversal or in which there is substantial vari-ation among females in their quality as mates (Bonduriansky2001). Such variation in female quality is likely to exist insystems in which females enhance their fitness with multiplematings or differ in fecundity due to such factors as bodysize and first- or last-male sperm precedence (Bonduriansky2001). If male choice is under selection in such systems andconspecific sperm precedence buffers females against matingmistakes, selection against hybridization may be greater formales than for females (Marshall et al. 2002). Indeed, if poly-androus females gain material benefits (e.g., nutrients) oroviposition stimulants from heterospecific matings, selectionmay favor indiscriminate mating by females, while also fa-voring males that only mate with conspecific females (Mar-shall et al. 2002).
The results from the Chrysochus hybrid zone, taken to-gether with other recent examples of hybrid zones featuringreproductive character displacement in male choice (Gaborand Ryan 2001; Jiggins et al. 2001; Albert and Schluter 2004)indicate that the evolution of male choice may frequentlyplay a role in speciation. Furthermore, our results suggestthat the cost of mating mistakes need not be very strong forreinforcement to drive the evolution of male choice. If weare to understand the generality of reinforcement and theconditions that favor it (as advocated by Howard 1993; Ser-vedio and Noor 2003), it is critical that we undertake morestudies of the reinforcement of male choice to resolve theconditions under which it is most likely to occur.
ACKNOWLEDGMENTS
We thank L. Tantalo and C. Fabry for assistance with thefield surveys; M. Morris, T. McFarland, and S. Schwartz forhelp with running mating trials and maintaining beetles inthe laboratory; and C. Yoon for helpful discussion throughoutthis project. We are also grateful to E. Rolan-Alvarez forproviding a copy of his sexual isolation program. In addition,we appreciate the thoughtful comments of S. Dobler, R. Har-rison, D. Howard, P. Nosil, E. Rolan-Alvarez, and two anon-ymous reviewers, whose efforts helped improve an earlierversion of this manuscript. This research was funded by aProject Development Award from Western Washington Uni-versity, Biology Department research funds, and a grant(DEB-0212652) from the National Science Foundation. Dur-ing the preparation of this paper, MAP was supported by anAlexander von Humboldt Foundation Research Fellowship.MAP expresses his appreciation to S. Dobler for providinga stimulating and productive environment for working on thispaper during his sabbatical at Hamburg University.
LITERATURE CITED
Albert, A. Y. K., and D. Schluter. 2004. Reproductive characterdisplacement of male stickleback mate preference: reinforce-ment or direct selection? Evolution 58:1099–1107.
Arnold, S. J., P. A. Verrell, and S. G. Tilley. 1996. The evolutionof asymmetry in sexual isolation: a model and test case. Evo-lution 50:1024–1033.
Avise, J. C., and N. C. Saunders. 1984. Hybridization and intro-gression among species of sunfish (Lepomis): analysis by mi-tochondrial DNA and allozyme markers. Genetics 108:237–255.
Backlund, M., B. Oxelman, and B. Bremer. 2000. Phylogeneticrelationships within the Gentiales based on NDHF and RBCL
sequences, with particular reference to the Loganiaceae. Am. J.Bot. 7:1029–1043.
Barton, N. H. 1980. The fitness of hybrids between two chromo-somal races of the grasshopper Podisma pedestris. Heredity 45:47–59.
———. 1996. Speciation: more than the sum of the parts. Curr.Biol. 6:1244–1246.
Barton, N. H., and G. M. Hewitt. 1985. Analysis of hybrid zones.Annu. Rev. Ecol. Syst. 16:113–148.
Bateman, A. J. 1948. Intrasexual selection in Drosophila. Heredity2:349–368.
Benito, G. 1997. Energy expenditure and geomorphic work of thecataclysmic Missoula flooding in the Columbia River Gorge,USA. Earth Surf. Proc. Landf. 22:457–472.
Benito, G., and J. E. O’Connor. 2003. Number and size of last-glacial Missoula floods in the Columbia River valley betweenthe Pasco Basin, Washington, and Portland, Oregon. Geol. Soc.Am. Bull. 115:624–638.
Bernatchez, L., and C. C. Wilson. 1998. Comparative phylogeog-raphy of Nearctic and Palearctic fishes. Mol. Ecol. 7:431–452.
Bonduriansky, R. 2001. The evolution of male mate choice in in-sects: a synthesis of ideas and evidence. Biol. Rev. 76:305–339.
Bretz, J. H. 1969. The Lake Missoula floods and the ChanneledScabland. J. Geol. 77:505–543.
Butlin, R. 1987. Speciation by reinforcement. Trends Ecol. Evol.2:8–13.
———. 1989. Reinforcement of premating isolation. Pp. 158–179in D. Otte and J. A. Endler, eds. Speciation and its consequences.Sinauer Associates, Sunderland, MA.
———. 1995. Reinforcement: an idea evolving. Trends Ecol. Evol.10:432–434.
Butlin, R. K., and T. Tregenza. 1997. Is speciation no accident?Nature 387:551–552.
Cain, M. L., V. Andreasen, and D. J. Howard. 1999. Reinforcingselection is effective under a relatively broad set of conditionsin a mosaic hybrid zone. Evolution 53:1343–1353.
Carroll, A. L. 1994. Interactions between body-size and matinghistory influence the reproductive success of males of a tortricidmoth, Zeiraphera canadensis. Can. J. Zool. 72:2124–2132.
Casares, P., M. C. Carracedo, B. del Rio, R. Pineiro, L. Garcia-Florez, and A. R. Barros. 1998. Disentangling the effects ofmating propensity and mating choice in Drosophila. Evolution52:126–133.
Clague, J. J., R. Barendregt, R. J. Enkin, and F. F. Foit Jr. 2003.Paleomagnetic and tephra evidence for tens of Missoula floodsin southern Washington. Geology 31:247–250.
Coyne, J. A. 1992. Genetics and speciation. Nature 355:511–515.Coyne, J. A., and H. A. Orr. 1989. Patterns of speciation in Dro-
sophila. Evolution 43:362–381.Cruz, R., M. Carballo, P. Conde-Padın, and E. Rolan-Alvarez. 2004.
Testing alternative models for sexual isolation in natural pop-ulations of Littorina saxatilis: Indirect support for by-productecological speciation? J. Evol. Biol. 17:288–293.
Dickinson, J. L. 1995. Trade-offs between postcopulatory ridingand mate location in the blue milkweed beetle. Behav. Ecol. 6:280–286.
———. 1997. Multiple mating, sperm competition, and cryptic fe-male choice in the leaf beetles (Coleoptera: Chrysomelidae). Pp.164–183 in J. C. Choe and B. J. Crespi, eds. The evolution ofmating systems in insects and arachnids. Cambridge Univ. Press,Cambridge, U.K.
Dobler, S., and B. D. Farrell. 1999. Host use evolution in Chry-sochus milkweed beetles: evidence from behaviour, populationgenetics and phylogeny. Mol. Ecol. 8:1297–1307.
Dobzhansky, T. 1940. Speciation as a stage in evolutionary diver-gence. Am. Nat. 74:312–321.
Dussourd, D. E., and T. Eisner. 1987. Vein-cutting behavior: insectcounterploy to the latex defense of plants. Science 237:898–901.
Gabor, C. R., and M. J. Ryan. 2001. Geographical variation inreproductive character displacement in mate choice by male sail-fin mollies. Proc. R. Soc. Lond. B 268:1063–1070.
Gilbert, D. G., and W. T. Starmer. 1985. Statistics of sexual iso-lation. Evolution 39:1380–1383.

2654 MERRILL A. PETERSON ET AL.
Harrison, R. G. 1998. Linking evolutionary pattern and process: therelevance of species concepts for the study of speciation. Pp.19–31 in D. J. Howard and S. B. Berlocher, eds. Endless forms:species and speciation. Oxford Univ. Press, Oxford, U.K.
Hewitt, G. 1990. Divergence and speciation as viewed from aninsect hybrid zone. Can. J. Zool. 68:1701–1715.
———. 1996. Some genetic consequences of ice ages, and theirrole in divergence and speciation. Biol. J. Linn. Soc. 58:247–276.
———. 2000. The genetic legacy of the Quaternary ice ages. Nature405:907–913.
Hey, J. 2001. The mind of the species problem. Trends Ecol. Evol.16:326–329.
Higgie, M., S. Chenoweth, and M. W. Blows. 2000. Natural selec-tion and the reinforcement of mate recognition. Science 290:519–521.
Hobel, G., and H. C. Gerhardt. 2003. Reproductive character dis-placement in the acoustic communication system of green treefrogs (Hyla cinerea). Evolution 57:894–904.
Howard, D. J. 1993. Reinforcement: origin, dynamics, and fate ofan evolutionary hypothesis. Pp. 46–69 in R. G. Harrison, ed.Hybrid zones and the evolutionary process. Oxford Univ. Press,Oxford, U.K.
———. 1998. Unanswered questions and future directions in thestudy of speciation. Pp. 439–448 in D. J. Howard and S. B.Berlocher, eds. Endless forms: species and speciation. OxfordUniv. Press, Oxford, U.K.
———. 1999. Conspecific sperm and pollen precedence and spe-ciation. Annu. Rev. Ecol. Syst. 30:109–132.
Howard, D. J., M. Reece, P. G. Gregory, J. Chu, and M. Cain. 1998.The evolution of barriers to fertilization between closely relatedorganisms. Pp. 279–288 in D. J. Howard and S. B. Berlocher,eds. Endless forms: species and speciation. Oxford Univ. Press,Oxford, U.K.
Jennions, M. D., and M. Petrie. 1997. Variation in mate choice andmating preferences: a review of causes and consequences. Biol.Rev. 72:283–327.
Jiggins, C. D., and J. Mallet. 2000. Bimodal hybrid zones and spe-ciation. Trends Ecol. Evol. 15:250–255.
Jiggins, C. D., W. O. McMillan, W. Neukirchen, and J. Mallet.1996. What can hybrid zones tell us about speciation? The caseof Heliconius erato and H. himera (Lepidoptera: Nymphalidae).Biol. J. Linn. Soc. 59:221–242.
Jiggins, C. D., R. E. Naisbitt, R. L. Coe, and J. Mallet. 2001. Re-productive isolation caused by colour pattern mimicry. Nature411:302–305.
Jones, A. G., J. R. Arguello, and S. J. Arnold. 2002. Validation ofBateman’s principles: a genetic study of sexual selection andmating patterns in the rough-skinned newt. Proc. R. Soc. Lond.B 269:2533–2539.
Kelly, J. K., and M. A. F. Noor. 1996. Speciation by reinforcement:a model derived from studies of Drosophila. Genetics 143:1485–1497.
Kirkpatrick, M., and M. R. Servedio. 1999. The reinforcement ofmating preferences on an island. Genetics 151:865–884.
Klicka, J., and R. M. Zink. 1999. Pleistocene effects on NorthAmerican songbird evolution. Proc. R. Soc. Lond. B 266:695–700.
Lemmon, A. R., C. Smadja, and M. Kirkpatrick. 2004. Reproductivecharacter displacement is not the only possible outcome of re-inforcement. J. Evol. Biol. 17:177–183.
Lewis, S. M. 2004. Multiple mating and repeated copulations: ef-fects on male reproductive success in red flour beetles. Anim.Behav. 67:799–804.
Liou, L. W., and R. D. Price. 1994. Speciation by reinforcementof premating isolation. Evolution 48:1451–1459.
Marın, I. 1997. Sexual isolation in Drosophila. III. Estimating iso-lation using male-choice experiments. J. Theor. Biol. 188:515–524.
Marshall, J. L., M. L. Arnold, and D. J. Howard. 2002. Reinforce-ment: the road not taken. Trends Ecol. Evol. 17:558–563.
Martin, O. Y., and D. J. Hosken. 2004. Copulation reduces male
but not female longevity in Saltella sphondylii (Diptera: Sepsi-dae). J. Evol. Biol. 17:357–362.
Mayden, R. L. 1997. A hierarchy of species concepts: the denoue-ment in the saga of the species problem. Pp. 381–424 in M. F.Claridge, H. A. Dawah, and M. R. Wilson, eds. Species: theunits of biodiversity. Chapman and Hall, New York.
Mayr, E. 1963. Animal species and evolution. Belknap Press, Cam-bridge, MA.
Merrel, D. J. 1950. Measurement of sexual isolation and selectivemating. Evolution 4:326–331.
Noor, M. A. 1995a. Speciation driven by natural selection in Dro-sophila. Nature 375:674–675.
———. 1995b. Reinforcement in speciation. Trends Ecol. Evol. 10:492.
———. 1999. Reinforcement and other consequences of sympatry.Heredity 83:503–508.
Nosil, P., B. J. Crespi, and C. P. Sandoval. 2003. Reproductiveisolation driven by the combined effects of ecological adaptationand reinforcement. Proc. R. Soc. Lond. B 270:1911–1918.
Parish, D. M. B., and J. C. Coulson. 1998. Parental investment,reproductive success and polygyny in the lapwing, Vanellus va-nellus. Anim. Behav. 56:1161–1167.
Peterson, M. A., S. Dobler, J. Holland, L. Tantalo, and S. Locke.2001. Behavioral, molecular, and morphological evidence for ahybrid zone between Chrysochus auratus and C. cobaltinus (Co-leoptera: Chrysomelidae). Ann. Entomol. Soc. Am. 94:1–9.
Peterson, M. A., K. J. Monsen, H. Pedersen, T. McFarland, and J.Bearden. 2005. Direct and indirect analysis of the fitness ofChrysochus (Coleoptera: Chrysomelidae) hybrids. Biol. J. Linn.Soc. 84:273–286.
Pfennig, K. S., and M. A. Simovich. 2002. Differential selectionto avoid hybridization in two toad species. Evolution 56:1840–1848.
Ribi, G., and S. Oertli. 2000. Frequency of interspecific matingsand of hybrid offspring in sympatric populations of Viviparusater and V. contectus (Mollusca: Prosobranchia). Biol. J. Linn.Soc. 71:133–143.
Rice, W. R. 1989. Analyzing tables of statistical tests. Evolution43:223–225.
Rieseberg, L. H., S. J. E. Baird, and A. M. Desrochers. 1998. Pat-terns of mating in wild sunflower hybrid zones. Evolution 52:713–726.
Rolan-Alvarez, E., and A. Caballero. 2000. Estimating sexual se-lection and sexual isolation effects from mating frequencies.Evolution 54:30–36.
Rundle, H. D., and D. Schluter. 1998. Reinforcement of sticklebackmate preferences: sympatry breeds contempt. Evolution 52:200–208.
Sady, M. B. 1994. Survey of the blue milkweed beetle, Chrysochuscobaltinus LeConte (Coleoptera: Chrysomelidae), in western Ne-vada. Coleop. Bull. 48:299.
Sætre, G.-P., T. Moum, S. Bures, T. Kral, M. Adamjan, and J.Moreno. 1997. A sexually selected character displacement inflycatchers reinforces premating isolation. Nature 387:589–592.
Sanderson, N. 1989. Can gene flow prevent reinforcement? Evo-lution 43:1223–1235.
Servedio, M. R. 2001. Beyond reinforcement: the evolution of pre-mating isolation by direct selection on preferences and post-mating, prezygotic incompatibilities. Evolution 55:1909–1920.
———. 2004. The evolution of premating isolation: local adapta-tion and natural and sexual selection against hybrids. Evolution58:913–924.
Servedio, M. R., and M. Kirkpatrick. 1997. The effects of geneflow on reinforcement. Evolution 51:1764–1772.
Servedio, M. R., and M. A. F. Noor. 2003. The role of reinforcementin speciation: theory and data. Annu. Rev. Ecol. Evol. Syst. 34:339–364.
Simmons, L. W. 2001. Sperm competition and its evolutionary con-sequences in the insects. Princeton Univ. Press, Princeton, NJ.
Sokal, R. R., and F. J. Rohlf. 1981. Biometry. 2nd ed. Freeman,New York.
Sota, T., and K. Kubota. 1998. Genital lock-and-key as a selectiveagent against hybridization. Evolution 52:1507–1513.

2655REINFORCEMENT IN A BEETLE HYBRID ZONE
Spieth, H. T., and J. M. Ringo. 1983. Mating behavior and sexualisolation in Drosophila. Pp. 315–379 in M. Ashburner, H. L.Carson, and J. N. Thompson Jr., eds. The genetics and biologyof Drosophila. Vol. 3C. Academic Press, New York.
Szymura, J. M., and N. H. Barton. 1986. Genetic analysis of a hybridzone between the fire-bellied toads, Bombina bombina and B.variegata, near Cracow in southern Poland. Evolution 40:1141–1159.
Trivers, R. L. 1972. Parental investment and sexual selection. Pp.136–179 in B. Campbell, ed. Sexual selection and the descentof man, 1871–1971. Aldine Publishing Co., Chicago.
Waage, J. 1979. Reproductive character displacement in Calopteryx(Odonata: Calopterygidae). Evolution 33:104–116.
Wedell, N., M. J. G. Gage, and G. A. Parker. 2002. Sperm com-petition, male prudence and sperm-limited females. Trends Ecol.Evol. 17:313–320.
Weiss, H. B., and E. West. 1921. Notes on insects of the spreadingdogbane, Apocynum androsaemifolium L., with a description ofa new dogbane midge by Dr. E.P. Felt. Can. Entomol. 53:146–152.
Williams, C. E. 1992. Movement of the dogbane beetle, Chrysochusauratus (Coleoptera: Chrysomelidae) in a patchy environment.Banisteria 1:8–10.
Corresponding Editor: R. Harrison




















