
Biomarkers of environmental stress in gills of Pinna nobilis (Linnaeus 1758) from Balearic Island
Upload
independentCategory
view
4download
0
30 Accepted by P. Cordeiro-Estrela: 15 Jan. 2011; published: 15 Mar. 2011
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2011 · Magnolia Press
Zootaxa 2791: 30–40 (2011) www.mapress.com/zootaxa/ Article
Designation of a neotype for the Brazilian porcupine, Coendou prehensilis (Linnaeus, 1758)
YURI L. R. LEITE1,3, VILACIO CALDARA JÚNIOR1, ANA CAROLINA LOSS1, LEONORA PIRES COSTA1,ÉVERTON R. A. MELO2, JOSÉ RAMON GADELHA2 & ANTONIO ROSSANO M. PONTES2
1Universidade Federal do Espírito Santo, Centro de Ciências Humanas e Naturais, Departamento de Ciências Biológicas, Labo-ratório de Mastozoologia e Biogeografia. Av. Marechal Campos 1468, Maruípe, 29.043-900, Vitória, ES, Brazil.2Universidade Federal de Pernambuco, Centro de Ciências Biológicas, Departamento de Zoologia, Laboratório de Estudo e Conser-vação da Natureza. Rua Prof. Moraes Rego 1235, Cidade Universitária, 50.740-620, Recife, PE, Brazil.3Corresponding author. E-mail: [email protected]
Abstract
The Brazilian porcupine was one of several species described and illustrated by the 17th-century naturalist Georg Marc-grave, whose text was among the primary references upon which Linnaeus based his Hystrix prehensilis. As currently un-derstood, Coendou prehensilis is a wide-ranging polytypic taxon that has never been revised and may represent a complexof closely related species. Given that no name-bearing type specimen of C. prehensilis is believed to be extant, and in orderto avoid ambiguous application of this name, we designate a specimen collected at the type locality in Pernambuco, north-eastern Brazil, as the neotype for Hystrix prehensilis Linnaeus. The geographic distribution of mitochondrial DNA hap-lotypes suggests that specimens from west-central Brazil previously identified as “Coendou prehensilis” belong to adifferent species, but only a comprehensive taxonomic revision of the genus will shed light on species limits and the geo-graphic ranges of C. prehensilis and other congeneric forms.
Key words: Erethizontidae, Mammalia, Neotropics, Rodentia, taxonomy
Resumo
O ouriço-cacheiro (Coendou prehensilis) é uma das muitas espécies descritas e ilustradas pelo naturalista do século 17Georg Marcgrave, cujo texto foi uma das referências primárias utilizadas por Linnaeus para embasar seu Hystrix prehen-silis. Da forma como é atualmente compreendido, Coendou prehensilis apresenta distribuição ampla, é um táxon politípicoque nunca foi revisado e pode representar um complexo de espécies filogeneticamente próximas. Como se acredita quenão exista um espécime tipo associado ao nome C. prehensilis, nós designamos um espécime coletado na localidade tipoem Pernambuco, nordeste do Brasil como o neótipo de Hystrix prehensilis Linnaeus, para evitar aplicação ambígua dessenome. A distribuição geográfica de haplótipos de DNA mitocondrial sugere que espécimes do centro oeste do Brasil iden-tificados previamente como “Coendou prehensilis” pertencem à uma espécie diferente, mas somente uma revisão ta-xonômica abrangente do gênero poderá esclarecer os limites específicos e a distribuição geográfica de C. prehensilis eoutras formas desse mesmo gênero.
Introduction
Georg Marcgrave (1610–1644) became one of the most important pre-Linnaean naturalists in the New World,mainly because his work was among the primary references upon which Linnaeus (1758) based some of his bino-mial scientific names in the tenth edition of his Systema Naturae (Hershkovitz 1987, Vanzolini 1996). Marcgravearrived in Brazil in 1638 to join other scientists and artists invited by Johan Maurits, count of Nassau-Siegen, thengovernor of a successful colony established by the Dutch government in the northeastern part of the country. Marc-grave explored northeastern Brazil; particularly those regions in the present states of Pernambuco, Paraíba, and Rio

Zootaxa 2791 © 2011 Magnolia Press · 31NEOTYPE FOR COENDOU PREHENSILIS
Grande do Norte (Gudger 1912). He carefully documented the natural history of this country until 1643, when hewas sent to Angola and died shortly after in 1644 (Boeseman et al. 1990). His manuscript Historiae rerum natu-ralium Brasiliae was published four years after his death (Marcgrave 1648), as the second part of a monumentalvolume called Historia Naturalis Brasiliae, organized by Willem Piso. On page 233 of this volume, Marcgravedescribed in detail the Brazilian porcupine or Cuandu brasiliensibus (Appendix 1) and provided a small woodcutillustration showing an animal in side view, perching on a tree branch (Fig. 1).
In the section describing porcupines, Linnaeus (1758: 57) gave only a short description of his Hystrix prehens-ilis: “H. pedibus tetradactylis, cauda elongata prehensili seminuda” (short-spined porcupine, with tetradactylousfeet, and long half-naked prehensile tail). His description is based on a number of early references, including notonly Marcgrave’s, but also authors referring to two other species of porcupines, distinct from Cuandu brasiliensi-bus. Considering that Hystrix prehensilis was a composite species and given the lack of a type specimen, Husson(1978) selected the specimen figured by Marcgrave (1648) (Fig. 1) as the lectotype of Hystrix prehensilis Linnaeus,1758. Unfortunately, no name-bearing specimen of C. prehensilis or any other animal described by Marcgrave(1648) is believed to be extant. In addition, many of these early descriptions from Marcgrave (1648) and their oftenrather poor woodcut illustrations are unreliable for a positive identification of the species (Boeseman et al. 1990).Therefore, the nomenclatural confusion that Husson (1978) wanted to prevent still persists.
FIGURE 1. Woodcut illustration of Cuandu brasiliensibus from Marcgrave (1648).
The original type locality of Hystrix prehensilis was given as “Asia, America meridionali” (Linnaeus 1758),and later fixed to “Pernambuco” (Thomas 1911). Some authors regard the large porcupines currently known asCoendou prehensilis as a widespread species, ranging from Venezuela across Brazil to northern Argentina (e.g.,Cabrera 1961; Eisenberg & Redford 1999; Woods & Kilpatrick 2005). Nevertheless, these animals have not beentaxonomically revised and may represent a complex of closely related species (Voss & da Silva 2001). As currentlyunderstood, Coendou prehensilis is wide ranging and polytypic, and has eight junior synonyms (Woods and Kilpat-rick 2005): Coendou longicaudatus Daudin in Lacépède, Hystrix cuandu Desmarest, Cercolabes platycentrotusBrandt, Cercolabes boliviensis Gray, Cercolabes tricolor Gray, Hystrix brandtii Jentink, Coendou centralisThomas, and Coendou sanctaemartae Allen. Recent authors have already applied some of these names to subspe-cies (e.g., C. prehensilis boliviensis, Anderson 1997) or full species (e.g., C. sanctaemartae, Alberico et al. 1999).
Coendou prehensilis is one of the oldest and most widely applied names for Neotropical porcupines. In order toavoid ambiguous application of this name as we resolve the taxonomic chaos and uncover the real diversity of NewWord porcupines (e.g., Voss & da Silva 2001), a neotype is necessary to define this taxon objectively under theconditions specified in Article 75 of the International Code of Zoological Nomenclature (ICZN 1999). Therefore,in this paper we designate, describe and illustrate a specimen of C. prehensilis collected in Pernambuco, where

LEITE ET AL.32 · Zootaxa 2791 © 2011 Magnolia Press
Marcgrave’s porcupine is assumed to have been collected (Thomas 1911). In addition, we compared DNAsequence of the mitochondrial cytochrome b gene of the neotype to published sequences from other erethizontids.
Material and methods
Morphometric data. We took four external measurements in millimeters—head and body length (HB: from tip ofnose to inflection point of tail), tail length (TL: from anus to tip of tail), hindfoot length with claw (HF), ear lengthfrom notch (EL)—and weight in grams (W). In addition, we measured some quills, vibrissae, hairs and spinoushairs from different regions of the body. Eighteen cranial measurements were taken using a digital caliper andrecorded to the nearest 0.01 millimeter, following Voss & da Silva (2001): condylo-incisive length (CIL); length ofdiastema (LD); length of incisive foramen (LIF); breadth of incisive foramen (BIF); maxillary toothrow length(MTR); length of molars (LM); breadth fourth pre-molar (BP4); breadth of first molar (BM1); anterior palatalbreadth (APB); posterior palatal breadth (PPB); posterior zygomatic breadth (PZB); height of infraorbital foramen(HIF); zygomatic length (ZL); length of nasals (LN); breadth of nasal aperture (BNA); breadth of braincase (BB);depth of incisor (DI); breadth of incisor tips (BIT).
Molecular data and phylogenetic analyses. We extracted total genomic DNA from liver tissue preserved in95% ethanol, using salt protocol and proteinase K (Bruford et al. 1992). Primers MVZ05 and MVZ16 (Smith &Patton 1993) were used in the polymerase chain reaction (PCR) to amplify the initial 801 base pairs (bp) of themitochondrial cytochrome b (cyt b) gene. PCR product was purified using ExoSAP enzymes (GE Healthcare Life-Sciences). In the cycle-sequencing reactions, we used primers MVZ05 and MVZ127 (Smith & Patton 1993, Leite& Patton 2002) for the light strand, and MVZ04 and MVZ16 (Smith & Patton 1993) for the heavy strand, with Big-Dye Terminator 3.1 (Applied Biosystems, Inc.). Sequences were read on an ABI-310 capillary automatedsequencer (Applied Biosystems, Inc.) and aligned using Clustal W (Thompson et al. 1994), as implemented inMEGA version 4.0 (Tamura et al. 2007). Genetic distances were estimated in MEGA, using Kimura's 2-parametermodel (K2P). Additional sequences were obtained from GenBank (http://www.ncbi.nlm.nih.gov/Genbank/),including those from the outgroups Erethizon dorsatum and Chaetomys subspinosus. The cyt b sequence generatedin the present study was stored in GenBank under the accession number HM462243.
TABLE 1. Specimens used in the phylogenetic analyses of cytochrome b, specimen number, GenBank accession number,locality and reference data. Numbered localities are plotted on the map (Figure 4).
In addition to our specimen of Coendou prehensilis, we used published DNA sequences of Coendou, Sphiggu-rus, Erethizon, and Chaetomys available from Genbank (Table 1). We examined the hierarchical relationships
Species Specimennumber
GenBank accession
Locality Locality num-ber on the map (Fig. 4)
Reference
C. prehensilis MN 73383 HM462243 Usina Trapiche, Penambuco 1 this study
“C. prehensilis” Manso 138 AF411582 UHE Manso, Mato Grosso 2 Bonvicino et al. 2002
“C. prehensilis” Manso 212 AF411581 UHE Manso, Mato Grosso 2 Bonvicino et al. 2002
“C. prehensilis” Manso 849 AF411584 UHE Manso, Mato Grosso 2 Bonvicino et al. 2002
“C. bicolor” MNFS 1016 U34851 Fazenda Santa Fé, Acre 3 Lara et al. 1996
“C. bicolor” MNFS 439 U34852 Eirunepé, Amazonas 4 Lara et al. 1996
S. melanurus UFPB 3001 AF411583 São João da Baliza, Roraima 5 Bonvicino et al. 2002
S. villosus CIT 1326 EU544661 UHE Rosal, Espírito Santo 6 Vilela et al. 2009
S. villosus MN 46937 AF411580 Sumidouro, Rio de Janeiro 7 Bonvicino et al. 2002
S. villosus MN 46938 AF407277 Rio das Ostras, Rio de Janeiro 8 Bonvicino et al. 2002
S. villosus CIT 2113 EU544662 Biritiba Mirim, São Paulo 9 Vilela et al. 2009
E. dorsatum – FJ357428 – – Vilela et al. 2009
C. subspinosus – EU544660 Salvador, Bahia – Vilela et al. 2009

Zootaxa 2791 © 2011 Magnolia Press · 33NEOTYPE FOR COENDOU PREHENSILIS
among cyt b haplotypes using maximum parsimony (MP) and maximum likelihood (ML) in PAUP* 4.0b10 (Swof-ford 2002). Heuristic searches were conducted using the tree-bisection-reconnection (TBR) algorithm via randomstepwise addition with 100 replicates. Node support was estimated by bootstrap analyses with 500 replicates. Weused jModelTest 0.1.1 (Posada 2008) to select the evolutionary model that best fits our data for ML analysis, usingthe Akaike Information Criterion (AIC) (Akaike 1974). The Tamura-Nei model was selected, with gamma distribu-tion shape parameter of 0.259 (TrN+G), and empirical base frequencies of: A = 0.3064, C = 0.2773, G = 0.1148,and T = 0.3015.
Results
Coendou prehensilis (Linnaeus 1758)Figs. 2, 3.
Neotype. Flat skin, skull of an adult male housed at Museu Nacional, Universidade Federal do Rio de Janeiro, Bra-zil (MN 73383), collected by Antonio Rossano Mendes Pontes (field number ARMP 63) on 12 May 2009. Tissuesample fixed in ethanol is housed in the Animal Tissue Collection at Universidade Federal do Espírito Santo,Vitória, Brazil (UFES-CTA 1797).
Type locality. Mata Xanguá, Usina Trapiche, municipality of Sirinhaém, state of Pernambuco, Brazil,8°38'50"S 35°10'15"W, elevation 100 m (Fig. 4). This area is in the Pernambuco Endemism Center (herein CEPE),the biogeographical unit of the Brazilian Atlantic forest located north of the São Francisco River in northeasternBrazil, which encompasses the states of Rio Grande do Norte, Paraíba, Pernambuco and Alagoas. The CEPE com-prises an important endemism center in South America, housing numerous endemic species (sensu Prance 1982,1987; Silva & Casteletti 2003), suggesting it is a hotspot within a hotspot, the Pernambuco refugium (Punde et al.2008; Carnaval & Moritz 2009), a world priority for biodiversity conservation, and one of the most importanthotspot on the planet (Myers et al. 2000). Usina Trapiche mill is one of the various scenarios of the 2% of what wasleft of the entire CEPE. It comprises an isolated archipelago of 5,202 ha of hundreds of small, irregular, impactedand isolated Atlantic forest fragments surrounded by a sugar-cane matrix. Mata Xanguá is the largest forest frag-ment (469.76 ha) of Usina Trapiche mill and is highly irregular shaped (shape index = 2.88; calculated followingLaurance & Yensen 1991), and formed by lowland evergreen tropical rainforest (Oliveira & Fontes 2000).
Distribution. According to Cabrera (1961), the distribution of the nominotypical subspecies is eastern Venezu-ela, the Guianas, and eastern Brazil. Given that specimens from Surinam described by Husson (1978) match ourdescription of the neotype (see below), we believe this is a good approximation of the geographic range of C. pre-hensilis. Woods & Kilpatrick (2005) extended the distribution of C. prehensilis westward to central Brazil, northernArgentina, eastern Paraguay, and Bolivia, but current data do not allow us to establish the western limits of its dis-tribution, especially given that similar species probably occur in west-central Brazil (see below). Only a compre-hensive taxonomic revision of the genus will shed light on the geographic range of C. prehensilis and congeners.
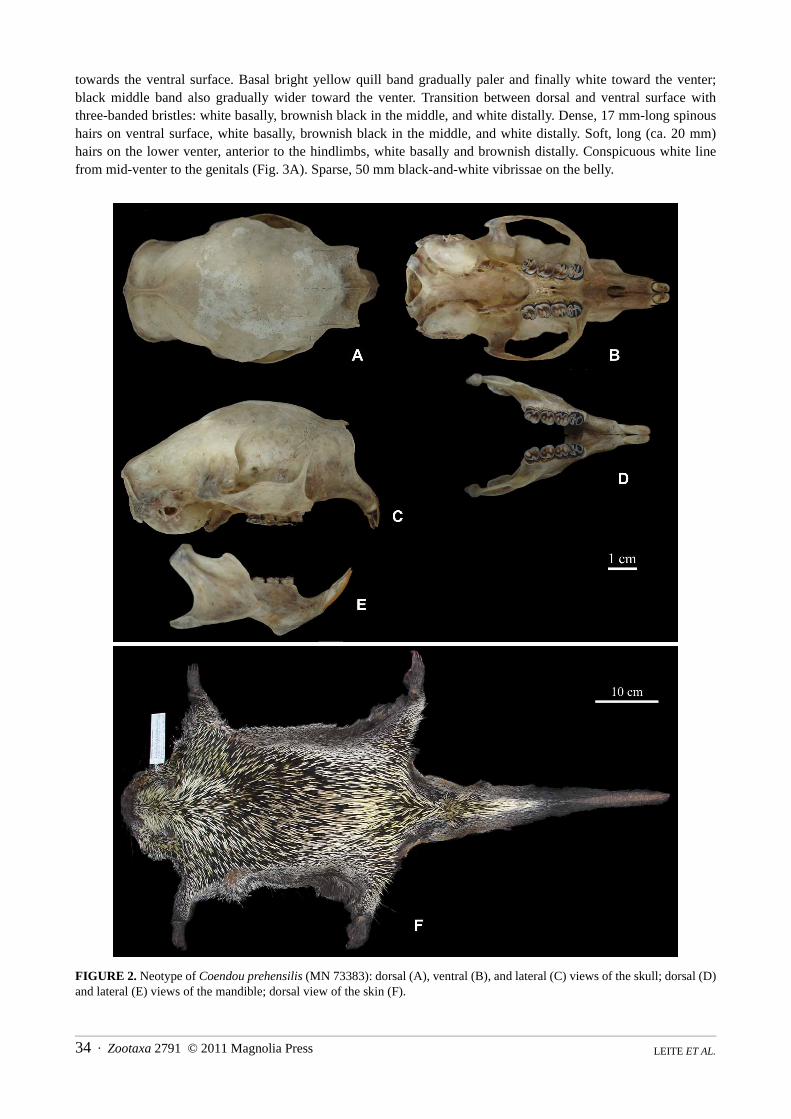
Description of the neotype. External—Body measurements: HB = 460 mm, TL = 400 mm, HF = 76 mm, EL= 28 mm. Weight = 2,720 grams. Tricolored quills on the top of head yellowish white basally, black in the middle,and white or whitish yellow distally. Thin, black and sparse hairs concealed under 5–20 mm long quills. Quillsshort (2–12 mm) around the eyes and muzzle, slightly longer (19 mm) on cheeks, whitish basally and distally, andblack in the middle (Fig. 3C). Coarse hairs around nose short and brownish; slightly longer pelage on the chin.Mystacial vibrissae long and black, some extending well beyond the shoulder (120 mm). Subocular and supraor-bital vibrissae 48–95 mm; genal vibrissae 44–82 mm; shorter submental vibrissae present. Bulbous muzzle verylarge (ca. 50 x 50 x 30 mm), soft and pinkish in the live animal. Conspicuously rounded ears nearly naked, coveredwith minute hairs; very short (10 mm) quills (either completely white, or black basally and white distally) aroundthe tragus.
Dorsal body surface densely covered with long quills (37–83 mm), gradually longer from shoulder to rump;with thin and sparse black hairs concealed underneath. Three patterns of quill bands on the shoulders: bicoloredquills (white or whitish yellow base and black tip); tricolored quills (white or whitish yellow base, blackish middle,and whitish tip); and four-banded quills (same as previous, with an additional brownish tip). Quills on mid-dorsumbright yellow basally, black in the middle, and white to pale yellow distally, with different color proportions oneach quill. Quills on rump tend to be dirty yellow distally. Quills gradually shorter and thinner from mid-dorsum
LEITE ET AL.34 · Zootaxa 2791 © 2011 Magnolia Press
towards the ventral surface. Basal bright yellow quill band gradually paler and finally white toward the venter;black middle band also gradually wider toward the venter. Transition between dorsal and ventral surface withthree-banded bristles: white basally, brownish black in the middle, and white distally. Dense, 17 mm-long spinoushairs on ventral surface, white basally, brownish black in the middle, and white distally. Soft, long (ca. 20 mm)hairs on the lower venter, anterior to the hindlimbs, white basally and brownish distally. Conspicuous white linefrom mid-venter to the genitals (Fig. 3A). Sparse, 50 mm black-and-white vibrissae on the belly.
FIGURE 2. Neotype of Coendou prehensilis (MN 73383): dorsal (A), ventral (B), and lateral (C) views of the skull; dorsal (D)and lateral (E) views of the mandible; dorsal view of the skin (F).

Zootaxa 2791 © 2011 Magnolia Press · 35NEOTYPE FOR COENDOU PREHENSILIS
FIGURE 3. Details of the neotype of Coendou prehensilis (MN 73383) before specimen preparation: ventral view (A), tail (B),and face (C). Scale bar = 50 mm.
Fore- and hindlimbs densely covered with coarse hairs, black or brown basally and whitish distally; hairsbecome progressively stiffer toward the dorsal surface. Small (<20 mm) quills along the outer margin of the fore-limbs become shorter and whitish basally toward the feet. Hindlimb quills concentrated near the rump; quills brightyellow basally, brownish black in the middle, and white distally. Postcranial vibrissae present, predominantly blackbasally and whitish distally, 20–32 mm long on the forelimbs and 30–52 mm on the hindlimbs. Plantar surface yel-lowish brown in the live animal; claws long, curved inward.
Tail dorsally prehensile and slightly shorter than head-and-body. Caudal quills bright yellow basally, black inthe middle, and white to pale yellow distally; distal band wider than others. Proximal one-third of tail covered dor-sally with longer quills (ca. 50 mm), covered laterally with bicolored bristles (brownish black basally and whitedistally, stiffer and darker toward the ventral surface), and covered ventrally with brownish bristles. Midde of tailcovered dorsally with shorter quills (ca. 35 mm), laterally with bristles darker than those on the proximal third. Dis-tal third of tail naked and calloused dorsally, covered laterally and ventrally with relatively sparse black bristles(Fig. 3B).
Skull—Frontal and nasal sinuses inflated dorsally (Fig. 1), from mid-nasals to fronto-parietal suture; parietalsslightly inflated. Postorbital ridge well developed and slightly inflated; left and right temporalis scars are closelyapproximated for about 16 mm along the dorsal midline of the posterior braincase, but do not form a definitive sag-ittal crest; supraoccipital ridge well developed. Rostrum short and tall; nasal aperture heart-shaped in frontal view.Nasals short, anterior margins convex and longer medially. Naso-frontal suture U-shaped and anterior to the pos-torbital ridge. Gnathic process well developed; zygomatic arches curved and widest across the jugals in dorsalview. Lacrimal-sphenoid suture nearly touches sphenofrontal foramen. Jugal tall, tapering posteriorly in lateralview. Incisive foramina oval, contained completely by the premaxillae and adjacent to premaxillarysuture. Longitudinal maxillary ridge extending from anteromedian edge of upper premolar to premaxillary suture.Anterior margin of mesopterygoid fossa extends to the middle of the third molars; bony roof of the fossa perforatedby sphenopalatine fenestrae. Auditory bullae large and bean-shaped, constricted posteriorly and contacting paroc-cipital processes. Basoccipital width subequal to auditory bulla width. Distinct, inward-extending ridge on roof ofexternal auditory meatus. Depression between occipital condyle and paroccipital process U-shaped. Cranial mea-surements in millimeters: CIL = 86.27; LD = 23.31; LIF = 5.11; BIF = 3.11; MTR = 19.11; LM = 14.50; BP4 =4.86; BM1 = 5.09; APB = 6.46; PPB = 8.82; PZB = 49.78; HIF = 12.92; ZL = 31.27; LN = 28.53; BNA = 21.98;BB = 35.29; DI = 4.18; BIT = 7.42.
Dentition—Dental formula: I 1/1, C 0/0, P 1/1, M 3/3. Anterior surface of incisors orange-yellow and remain-ing surfaces whitish. Maxillary toothrows nearly parallel and showing typical erethizontid occlusal surface (Fig. 1).Ancestral pentalophodont morphology (Vucetich & Verzi 1994) on maxillary teeth, where anteroloph, protoloph,mesoloph and metaloph link to posteroloph.

LEITE ET AL.36 · Zootaxa 2791 © 2011 Magnolia Press
Comparisons. There is no other large porcupine with tricolored quills within the range of C. prehensilis. Thistaxon and C. bicolor Tschudi are the two largest species of Neotropical porcupines, measuring from about 380 to560 mm of head-and-body length and weighting from to 3 to 5 kg (Emmons & Feer 1997). The neotype of C. pre-hensilis is on the lower range of these values, measuring 460 mm (HB) and weighing 2.7 kg. Despite its overallsmall size, this specimen has heavily worn molars, indicative of an old adult individual according to Voss & Anger-mann (1997). Coendou bicolor has typically bicolored spines, yellow-white basally and black distally (Emmons &Feer 1997).
The pelage of the neotype matches Marcgrave’s overall description (Appendix 1): the dorsal surface is coveredwith yellowish-based quills that are blackish medially and whitish distally. The tail is slightly longer than head-and-body in Marcgrave’s description, but the neotype has slightly longer head-and-body. Among the specimensfrom Surinam, housed at the Rijksmuseum van Natuurlijke Historie, Leiden (RMNH), and described by Husson(1978), one had longer head-and-body (RMNH 18299: HB = 487 mm, TL = 471 mm) and another had longer tail(RMNH 21902: HB = 460 mm, TL = 480 mm). In general, the pelage of the neotype agrees with Husson’s (1978)description of Surinamese material, albeit the dorsal quills are shorter (20 mm on the head and 83 mm on the rump)on the former than the latter (40 mm on the head and 100 mm on the rump). The detailed description of the tailgiven by Husson (1978) matches very well the pattern we found on the neotype (Fig. 3B).
FIGURE 4. Map showing the type locality of Coendou prehensilis (star) and other localities where specimens of erethizontidswith cytochrome b sequences available were collected: “Coendou prehensilis” (black square), “Coendou bicolor” (black cir-cles), Sphiggurus melanurus (open square), and S. villosus (open circles). See Table 1 for the list of specimens and localities.
The cranial morphology of the neotype is also very similar to Husson’s (1978) description of Surinamese mate-rial. The two main distinctions are: the posterior margin of the incisive foramen is anterior to the suture between thepremaxillary and the maxillary in the neotype, and at this suture on Surinamese specimens; anterior margin ofmesopterygoid fossa reaches mid-M3 on the neotype and between M2 and M3 on Surinamese specimens.
Phylogenetic analyses of DNA sequences. The strict consensus of the six most-parsimonious trees recoveredby our analyses (length = 361 steps) showed a topology very similar to the ML tree (-Ln = 2654.6), but higherbootstrap support for nodes in general (Fig. 5). Only MP confirmed the monophyly of the three “C. prehensilis”sequences from Bonvicino et al. (2002), but with no bootstrap support. These sequences grouped with specimensidentified as “C. bicolor” in Genbank, diverging only 0.6% from them (Fig. 5). Together, they formed a well sup-ported clade (100% MP and 90% ML bootstrap) that probably represents a single species, considering that up to

Zootaxa 2791 © 2011 Magnolia Press · 37NEOTYPE FOR COENDOU PREHENSILIS
2,000 km separate some of them (Fig. 4). The sequence from the neotype of C. prehensilis diverged 4.9% on aver-age from a sister-group comprising haplotypes from specimens identified as “C. bicolor” and “C. prehensilis” byBonvicino et al. (2002). Coendou and Sphiggurus formed two reciprocally monophyletic groups, well supported inMP analysis (99% and 92%, respectively), showing 10.9% genetic divergence between them (Fig. 5). The recipro-cal monophyly of Coendou and Sphiggurus had low support in the ML analysis (76% and 66%, respectively).
FIGURE 5. Parsimony tree based on cytochrome b sequences from Neotropical porcupines of the genera Coendou and Sphig-gurus (Erethizontidae). Numbers above branches correspond to parsimony and likelihood bootstrap support values > 50%;numbers below branches indicate the average pairwise genetic distances (Kimura 2-parameter) among specimens. Erethizondorsatum and Chaetomys subspinosus were used as outgroups.
Discussion
One of the main challenges in the taxonomy of Neotropical mammals in general, and porcupines in particular, isthe uncertain application of many old names, especially when no type material is available (Voss & Angermann1997). The fixation of broad Linnaean type localities (Thomas 1911) and the clarification of the taxonomic historyof hystricognath rodents (Tate 1935) were important steps toward resolving the confusing taxonomy of erethizon-tids, but still insufficient to allow proper species identification.
As pointed out above, specimens from western Brazilian Amazonia identified as “C. bicolor” by Lara et al.(1996) and those from Mato Grosso identified as “C. prehensilis” by Bonvicino et al. (2002) are likely conspecific.Judging by the geographic distribution of DNA haplotypes, we hypothesize that they are not conspecific with trueC. prehensilis as represented by the neotype from northeastern Brazil described here, which differs by 4.9%sequence divergence from them. This value is relatively high for intraspecific cyt b divergences (average = 2.49%)and it is within the range found in sister-species of mammals (4–11%) (Bradley & Baker 2001). Isolation by dis-tance would be a good explanation for high intraspecific divergence, given that the collecting sites in Mato Grossoand Pernambuco are 2,400 km apart (Fig. 4). However, we do not observe this pattern within samples identified as“C. bicolor” from distant localities: one of the western Amazonia samples (MNFS 1016) is from Acre, which isalso 2,000 km apart from Mato Grosso (Fig. 4), but they differ by only 0.6%.
Assuming that samples from Mato Grosso (Manso 138, 212, 849) and Amazonia (MNFS 439, 1016) representa single species distinct from C. prehensilis, we suspect that names currently considered junior synonyms of C. pre-hensilis are more likely to apply to this species than C. bicolor, with type locality in east-central Peru. Cercolabes

LEITE ET AL.38 · Zootaxa 2791 © 2011 Magnolia Press
platycentrotus Brandt, 1835 is one of the oldest names available, but the type locality is unknown (“America aus-tralis?” in the original description). It should be noted that Brant (1835) did not mention G. H. Langsdorff as thecollector of the specimen, so the type locality f C. platycentrotus cannot be limited to Brazil based on his paper.Cercolabes boliviensis Gray, 1850 is based on a specimen from Bolivia previously examined by Waterhouse(1848), who had restricted the collecting locality to Santa Cruz de la Sierra. Coendou centralis Thomas, 1903 wascollected during the Percy Sladen Expedition to Mato Grosso, Brazil. For instance, specimens from Mato Grossoreported by Bonvicino et al. (2002) were collected at the Manso hydropower area, which is just 50 km north ofChapada dos Guimarães, the type locality of C. centralis. Of course this hypothesis must be tested with propercomparisons of museum specimens, including types, and evaluation of the old literature.
Name-bearing types are the international standards of reference that provide objectivity in zoological nomen-clature (ICZN 1999) and the designation of a name-bearing type is a necessary step to define Coendou prehensilisobjectively. As pointed out by Ellerman (1940): “The genus [Coendou] is in need of revision; it appears to be in amore chaotic state even than is usual among these Neotropical Rodents”. This statement is still current seventyyears later, despite some significant advances in recent years (e.g., Voss & Angerman 1997, Voss & da Silva 2001).
Acknowledgements
We thank A. Gentry and C. O. Azevedo for very productive discussions regarding Linnaean types and zoologicalnomenclature, and R. S. Voss for suggesting the description of the neotype and providing valuable comments to themanuscript. We are grateful to Salgado and Trapiche mills (within which the forest fragments stretch) for theircooperation, for having granted some seed money essential for the accomplishment of this study, and also for hav-ing provided the necessary infrastructure for the project. In 2008 we received the recognition and financial supportfrom Conservation International in Brazil, which has sponsored this study, both field inventories and molecularanalysis through the Critical Ecosystem Partnership Fund (CEPF). Conselho Nacional de Desenvolvimento Cientí-fico e Tecnológico (CNPq) granted an undergraduate scholarship to JRG, and fellowship support to ARMP andYLRL. Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) provided a graduate scholarshipto ACL. J. A. de Oliveira (MN) kindly catalogued the neotype and provided museum number, and D. Astúaallowed access to the mammal collection at UFPE. Instituto Brasileiro do Meio Ambiente e Recursos NaturaisRenováveis (IBAMA) granted collecting permits to the authors.
References
Akaike, H. (1974) A new look at the statistical model identification. IEEE Transactions on Automatic Control, 19, 716–723.Alberico, M.S., Rojas-Díaz, V. & Moreno, J.G. (1999) Aporte sobre la taxonomía y distribución de los puercoespines (Roden-
tia: Erethizontidae) en Colombia. Revista da la Academia Colombiana de Ciencias Exactas Fisica, 23 (suplemento espe-cial), 595–612.
Anderson, S. (1997) Mammals of Bolivia, taxonomy and distribution. Bulletin of the American Museum of Natural History,231, 1–652.
Boeseman, M., Holthuis, L.B., Hoogmoed, M.S. & Smeenk, C. (1990) Seventeenth century drawings of Brazilian animals inLeningrad. Zoologische Verhandelingen Leiden, 267, 1–189.
Bonvicino, C.R., Penna-Firme, V. & Braggio, E. (2002) Molecular and karyotypic evidence of the taxonomic status of Coendouand Sphiggurus (Rodentia: Hystricognathi). Journal of Mammalogy, 83(4), 1071–1076.
Bradley, R.D. & Baker, R.J. (2001) A test of the genetic species concept: cytochrome-b sequences and mammals. Journal ofMammalogy, 82(4), 960–973.
Brandt, J.F. (1835) Mammalium rodentium exoticorum novorum vel minus rite cognitorum musei academici zoologici descrip-tiones et icones. Mémoires de L’Académie Impériale des Sciences de Saint Petersbourg, ser. 6 (tome III), pt. 2, 358–442 +17 pls.
Bruford, M.W., Hanotte, O., Brookfield, J.F.Y. & Burke, T. (1992) Single-locus and DNA fingerprinting. In: Hoelzel, A.R.(Ed), Molecular genetic analyses of populations. A Pratical Approach. IRL Press, Oxford, pp. 225–269.
Cabrera, A. (1961) Catálogo de los mamíferos de América del Sur. Revista del Museo Argentino de Ciencias Naturales Bernar-dino Rivadavia, 4, 309–732.
Carnaval, A.C. & Moritz, C. (2009) Predicting diversity within hotspots to enhance conservation. Science, 323, 785–789.

Zootaxa 2791 © 2011 Magnolia Press · 39NEOTYPE FOR COENDOU PREHENSILIS
Eisenberg, J.F. & Redford, K.H. (1999) Mammals of the Neotropics, 3. The central Neotropics. University of Chicago Press,Chicago, 609 pp.
Ellerman, J.R. (1940) The families and genera of living rodents. Vol. 1. Rodents other than Muridae. Trustees of the BritishMuseum (Natural History), London, 689 pp.
Emmons, L.H. & Feer, F. (1997) Neotropical rainforest mammals: a field guide. 2nd ed., University of Chicago Press, Chicago.Gray, J.E. (1850) On the species of Cercolabes confounded under the name of C. prehensilis. Annals and Magazine of Natural
History, vol. 5, 2nd series, 380–381.Gudger, E.W. (1912) George Marcgrave, the first student of American natural history. The Popular Science Monthly, 81(15),
250–274.Hershkovitz, P. (1987) A history of the recent mammalogy of the neotropical region from 1492 to 1850. Fieldiana: Zoology,
n.s., 39, 11–98.Husson, A.M. (1978) The mammals of Suriname. E. J. Brill, Leiden, 569 pp., 160 pls.ICZN [International Commission on Zoological Nomenclature] (1999) International Code of Zoological Nomenclature. 4th
Edition. The International Trust for Zoological Nomenclature, London, 306 pp. Available from http://www.nhm.ac.uk/hosted-sites/iczn/code/ (accessed 4 June 2010).
Lara, M.C., Patton, J.L. & Silva, M.N.F. (1996) The simultaneous diversification of South American echimyid rodents(Hystricognathi) based on complete cytochrome b sequences. Molecular Phylogenetics and Evolution, 5, 403–413.
Laurance, W.F. & Yensen, E. (1991) Predicting the impacts of edge effects in fragmented habitats. Biological Conservation, 55,77–92.
Leite, Y.L.R. & Patton, J.L. (2002) Evolution of South American spiny rats (Rodentia, Echimyidae): the star-phylogenyhypothesis revisited. Molecular Phylogenetics and Evolution, 25, 455–464.
Linnaeus, C. (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus,differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Laurentii Salvii, Holmiae [= Stockholm], ii+824 pp.Available from: http://gdz.sub.uni-goettingen.de/dms/load/img/?IDDOC=265100 (accessed on 10 April 2010).
Marcgrave, G. (1648) Historiae rerum naturalium Brasiliae, libri octo. Cum appendice de Tapuyis, et Chilensibus. Ioannes deLaet, Antwerpianus, In ordinem digessit & annotationes addidit, & varia ab auctore omissa supplevit & illustravit. In:Piso, W. & Macgrave, G. Historia naturalis Brasiliae: in qua non tantum plantae et animalia, sed et indigenarum morbi,ingenia et mores describuntur et iconibus supra quingentas illustrantur. Lugdun. Batavorum [Leiden]: Apud FranciscumHackium, et Amstelodami: Apud Lud. Elzevirium. Available from http://www.botanicus.org/item/31753000818648(accessed on 10 April 2010).
Myers, N., Mittermeier, R.A., Mittermeier, C.G., Fonseca, G.A.B. & Kent, J. (2000) Biodiversity hotspots for conservation pri-orities. Nature, 403, 853–858.
Oliveira, A.T. & Fontes, M.A.L. (2000) Patterns of floristic differentiation among Atlantic forests in southeastern Brazil andthe influence of climate. Biotropica, 32, 793–810.
Posada, D. (2008) jModelTest: Phylogenetic Model Averaging. Molecular Biology and Evolution, 25, 1253–1256.Prance, G.T. (1982) Forest refuges: evidences from woody angiosperms. In: Whitmore, T.C. & Prance, G.T. (Eds), Biological
diversification in the tropics. Columbia University Press, New York, pp. 137–158.Prance, G.T. (1987) Biogeography of neotropical plants. In: Prance, G.T. (Ed), Biogeography and quaternary history in tropical
America. Oxford Monographs on Biogeography. Clarendon Press, Oxford, pp. 175–196.Punde, S., Godbole, A. & Sarnaik, J. (2008) Searching for hotspots within a hotspot – using global prioritization schemes at a
regional level. An example from the Northern Western Ghats. Paper presented at the annual meeting of the InternationalCongress for Conservation Biology, Convention Center, Chattanooga, TN, July 10, 2008. Available from http://www.alla-cademic.com/meta/p237444_index.html (accessed 6 June 2010).
Silva, J.M.C. & Casteletti, C.H.M. (2003) Status of the biodiversity of the Atlantic forest of Brazil. In: Galindo-Leal, C. &Câmara, I.G. (Eds), The Atlantic forest of south America: biodiversity status, threats, and outlook. CABS and Island Press,Washington, DC, pp. 43–59.
Smith, M.F. & Patton, J.L. (1993) Diversification of South American muroid rodents: evidence from mitochondrial DNAsequence data for the Akodontine tribe. Biological Journal of the Linnean Society, 50, 149–177.
Swofford, D.L. (2000) PAUP*: Phylogenetic Analysis Using Parsimony (*and other methods), Version 4.0b. Sinauer Associ-ates, Sunderland.
Tamura, K., Dudley, J., Nei, M. & Kumar, S. (2007) MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) softwareversion 4.0. Molecular Biology and Evolution, 24, 1596–1599.
Tate, G.H.H. (1935) The taxonomy of the genera of Neotropical hystricoid rodents. Bulletin of the American Museum of Natu-ral History, 68, 295–447.
Thomas, O. (1903) On the mammals collected by Mr. A. Robert at Chapada, Matto Grosso (Percy Sladen Expedition to CentralBrazil). Proceedings of the Zoological Society of London, 1903, vol. 2, 232–243.
Thomas, O. (1911) The mammals of the tenth edition of Linnaeus; an attempt to fix the types of the genera and the exact basesand localities of the species. Proceedings of the Zoological Society of London, 1911, 120–158.
Thompson, J.D., Higgins, D.G & Gibson, T.J. (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research,22(22), 4673–4680.

LEITE ET AL.40 · Zootaxa 2791 © 2011 Magnolia Press
Vanzolini, P.E. (1996) A contribuição zoológica dos primeiros naturalistas viajantes no Brasil. Revista da Universidade de SãoPaulo, 30, 190–238.
Voss, R.S. & Angermann, R. (1997) Revisionary notes on Neotropical porcupines (Rodentia: Erethizontidae). 1. Type materialdescribed by Olfers (1818) and Kuhl (1820) in the Berlin Zoological Museum. American Museum Novitates, 3214, 1–42.
Voss, R.S. & da Silva, M.N.F. (2001) Revisionary notes on Neotropical porcupines (Rodentia: Erethizontidae). 2. A review ofthe Coendou vestitus group with descriptions of two new species from Amazonia. American Museum Novitates, 3351, 1–36.
Vucetich, M.G. & Verzi, D.H. (1994) Las homologías en los diseños oclusales de los roedores Caviomorpha: un modelo alter-nativo. Mastozoologia Neotropical, 1(1), 61–72.
Waterhouse, G.R. (1848) The natural history of the Mammalia. Vol. 2, Hippolyte Bailliere, London, 500 pp. + 21 pls.Woods, C.A. & Kilpatrick, C.W. (2005) Suborder Hystricognathi. In: Wilson, D.E. & Reeder, D.M. (Eds), Mammal species of
the world: a taxonomic and geographic reference. 3rd ed., Johns Hopkins University Press, Baltimore, pp. 1538–1600.
APPENDIX 1. Translated description from Marcgrave’s Historiae rerum naturalium Brasiliae, page 233.
CUANDU Brasiliensibus: Ouriço cacheiro in Portuguese; eenysere Vercken in Flemish. It is of the size of a large monkey; andthe body is covered with quills, three or four inches long, but it has no hair. These quills are yellowish (like white feathers) fromthe base to nearly the middle, and the rest is black, the tip is white; sometimes, the color is reddish brown instead of black insome of them. The quills are sharp as needles and the animal can eject them. The body length from the occiput to the beginningof the tail is one Rhynland foot. The tail is one foot and five inches long, and it has quills on the first half; the distal part isnaked with some small bristles, like pig’s tail. The head is about three inches and a half long and is also covered with quills,which are smooth near the mouth and nostrils. The ears are small and almost hidden beneath the quills; the mouth is very low,near the neck, and is similar to the mouth of the hare, with two teeth, which are visible from the outside. The nostrils are obvi-ous and the muzzle is broad and almost square; the eyes are round, prominent, shining like carbuncle. There is a lot of hair, fourinches long, around the nostrils and mouth, like cat's or hare's whiskers. The feet are like monkey feet, but with only four fin-gers; the thumb is missing, and there is only the place as if it had been amputated. The front feet are smaller than the hind feet;the length of the foreleg is about four inches, and the hind legs are a little longer. The legs have quills, but not the feet. Duringthe day, it usually sleeps; it wanders at night, breathing as if gasping for air; it likes to eat chicken. It makes a pig-like sound iii.It climbs trees, but slowly, because it has no thumb; it grasps the tree with the tail to avoid falling when descending; it is notcapable of jumping, such is its fear of falling. The meat of this animal is good and delicious; I often ate it and country peoplelike it very much.
Copyright © 2022 FDOKUMEN




















