
Morphological evidence of species differentiation within Lepus capensis Linnaeus, 1758 (Leporidae,...
13
ORIGINAL INVESTIGATION Morphological evidence of species differentiation within Lepus capensis Linnaeus, 1758 (Leporidae, Lagomorpha) in Cape Province, South Africa Fernando Palacios a, , Chiara Angelone a,b , Germa´n Alonso c , Santiago Reig d a Departamento de Biodiversidad y Biologı´a Evolutiva, Museo Nacional de Ciencias Naturales (CSIC), C/Jose´Gutie´rrez Abascal 2, 28006 Madrid, Spain b Dipartimento di Scienze Geologiche, Universita´ Roma Tre, Largo San Leonardo Murialdo 1, 00146 Rome, Italy c Departamento de Ecologı´a, Universidad Complutense, Facultad de Ciencias Biolo´gicas, C/Jose´Antonio Novais 2, Ciudad Universitaria, 28040 Madrid, Spain d Unidad de Medicina Experimental, Laboratorio de Imagen, Hospital General Universitario Gregorio Maran˜ o´n, C/Dr. Esquerdo 46, 28007 Madrid, Spain Received 10 October 2007; accepted 25 October 2007 Abstract A morphological study was carried out of the hares (Lepus) from Cape Province previously assigned to the subspecies L. capensis capensis, L. c. centralis, and L. c. grantii. The purpose of the study was to characterize the species L. capensis Linnaeus, 1758. In doing so, it was possible to distinguish two populations which we consider different species, as each shows homogeneous cranial, dental and pelage features. One of them, defined as L. capensis, is distributed near Cape Town not far from the coast, between Lambert’s Bay and Cape Agulhas. The other species, defined as L. centralis, which includes L. grantii as a synonym, is distributed in central and western Cape Province. L. capensis and L. centralis have a parapatric distribution, with a small area of sympatry in a contact zone in Compagnies Drift area, near Lambert’s Bay. With respect to cranial differences between the two species, L. capensis has a stronger maxilla and more robust dental series, while L. centralis has larger tympanic bullae. Among dental characters, L. centralis usually has a deeper groove and more abundant cement than L. capensis in the first upper incisor, and its internal lobe is squared, while in L. capensis is rounded. As for pelage color and pattern, L. capensis shows a more extended white ventral area than L. centralis. Our results are of interest for further research on taxonomic problems regarding Old World hare populations in which L. capensis is concerned. r 2007 Deutsche Gesellschaft fu¨r Sa¨ugetierkunde. Published by Elsevier GmbH. All rights reserved. Keywords: Lepus capensis; L. centralis; Taxonomy; Syntypes; Museum collections Introduction Lepus capensis Linnaeus, 1758 is the hare species for which the largest area of distribution has been recognized. It has been described as ranging from south to north Africa, from Middle East to eastern Asia, and in the Iberian peninsula (Ellerman and Morrison-Scott 1951; Ellerman et al. 1953). Petter (1959, 1961) considered L. europaeus as a synonym of L. capensis, thus expanding the range of L. capensis to the European area of the brown hare. Since these revisions, L. capensis has been a widely accepted taxon in publications about taxonomy, biology, ARTICLE IN PRESS www.elsevier.de/mambio 1616-5047/$ - see front matter r 2007 Deutsche Gesellschaft fu¨r Sa¨ugetierkunde. Published by Elsevier GmbH. All rights reserved. doi:10.1016/j.mambio.2007.10.013 Mamm. biol. 73 (2008) 358–370 Corresponding author. Tel.: +34 91 4111328; fax: +34 91 5645078. E-mail address: [email protected] (F. Palacios).
Transcript of Morphological evidence of species differentiation within Lepus capensis Linnaeus, 1758 (Leporidae,...

ARTICLE IN PRESS
1616-5047/$ - see front m
doi:10.1016/j.mambio.20
�Corresponding autho
E-mail address: ferna
www.elsevier.de/mambio
ORIGINAL INVESTIGATION
Morphological evidence of species differentiation within Lepus capensisLinnaeus, 1758 (Leporidae, Lagomorpha) in Cape Province, South Africa
Fernando Palaciosa,�, Chiara Angelonea,b, German Alonsoc, Santiago Reigd
aDepartamento de Biodiversidad y Biologıa Evolutiva, Museo Nacional de Ciencias Naturales (CSIC), C/Jose Gutierrez Abascal 2,
28006 Madrid, SpainbDipartimento di Scienze Geologiche, Universita Roma Tre, Largo San Leonardo Murialdo 1, 00146 Rome, ItalycDepartamento de Ecologıa, Universidad Complutense, Facultad de Ciencias Biologicas, C/Jose Antonio Novais 2,
Ciudad Universitaria, 28040 Madrid, SpaindUnidad de Medicina Experimental, Laboratorio de Imagen, Hospital General Universitario Gregorio Maranon, C/Dr. Esquerdo 46,
28007 Madrid, Spain
Received 10 October 2007; accepted 25 October 2007
Abstract
A morphological study was carried out of the hares (Lepus) from Cape Province previously assigned to thesubspecies L. capensis capensis, L. c. centralis, and L. c. grantii. The purpose of the study was to characterize the speciesL. capensis Linnaeus, 1758. In doing so, it was possible to distinguish two populations which we consider differentspecies, as each shows homogeneous cranial, dental and pelage features. One of them, defined as L. capensis, isdistributed near Cape Town not far from the coast, between Lambert’s Bay and Cape Agulhas. The other species,defined as L. centralis, which includes L. grantii as a synonym, is distributed in central and western Cape Province.L. capensis and L. centralis have a parapatric distribution, with a small area of sympatry in a contact zone inCompagnies Drift area, near Lambert’s Bay. With respect to cranial differences between the two species, L. capensis
has a stronger maxilla and more robust dental series, while L. centralis has larger tympanic bullae. Among dentalcharacters, L. centralis usually has a deeper groove and more abundant cement than L. capensis in the first upperincisor, and its internal lobe is squared, while in L. capensis is rounded. As for pelage color and pattern, L. capensis
shows a more extended white ventral area than L. centralis. Our results are of interest for further research ontaxonomic problems regarding Old World hare populations in which L. capensis is concerned.r 2007 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
Keywords: Lepus capensis; L. centralis; Taxonomy; Syntypes; Museum collections
Introduction
Lepus capensis Linnaeus, 1758 is the hare species forwhich the largest area of distribution has beenrecognized. It has been described as ranging from south
atter r 2007 Deutsche Gesellschaft fur Saugetierku
07.10.013
r. Tel.: +34 91 4111328; fax: +34 91 5645078.
[email protected] (F. Palacios).
to north Africa, from Middle East to eastern Asia, andin the Iberian peninsula (Ellerman and Morrison-Scott1951; Ellerman et al. 1953). Petter (1959, 1961)considered L. europaeus as a synonym of L. capensis,thus expanding the range of L. capensis to the Europeanarea of the brown hare.
Since these revisions, L. capensis has been a widelyaccepted taxon in publications about taxonomy, biology,
nde. Published by Elsevier GmbH. All rights reserved.
Mamm. biol. 73 (2008) 358–370

ARTICLE IN PRESSF. Palacios et al. / Mamm. biol. 73 (2008) 358–370 359
ecology and genetics of Old World hares and in regionaland country faunas. Nevertheless, L. capensis remains,at a taxonomical level, one of the most poorlycharacterized species, not only in the case of the nominalsubspecies (L. capensis capensis), but also with respect toother populations assigned to the species.
The scanty data available regarding the nominalsubspecies were provided by Roberts (1951) who, in hisbook on South African mammals, included a shortdescription, with data on body and skull dimensions,and pelage color and pattern of several specimens fromEendekuil, Lambert’s Bay and Vredendal. However, thislast locality has to be considered doubtful, becauseRoberts (1951) assigned the two subadult specimensfrom Vredendal of the Transvaal Museum to L. capensis
grantii when describing the body and skull measure-ments, and to L. capensis capensis when describingpelage pattern. Roberts (1951) also postulated theL. capensis capensis distribution area as covering aterritory comprised between Oudtshoorn and OlifantsRiver (SW Cape Province). It is important to note thatRoberts (1951) described some morphological andchorological differences between different populationsof L. capensis, although he did not assign them ataxonomic rank above the subspecies level. Robinsonand Dippenaar (1987), in their morphometric analysesof lagomorphs from South Africa, confirmed thatno specific differences could be recognized withinL. capensis populations. It is worth to remark thatRoberts (1951) and Robinson and Dippenaar (1987)only studied the scanty specimens of L. capensis capensis
from the Transvaal Museum (Pretoria, South Africa),and did not take into account in their analyses the largeseries preserved in the Amathole Museum (KingWilliam’s Town) and the old collection kept in theSouth African Museum (Cape Town). Petter (1959),who studied the collections of the London and Parismuseums, did not include in his revision specimens fromSW Cape Province and accepted most of the Africanforms listed by Ellerman et al. (1953) under L. capensis.
An analysis of the morphological characters definingL. capensis capensis has never been performed, in spiteof the compelling need of comparing the nominalsubspecies with other L. capensis populations and withother species of the genus Lepus. The lack of a precisecharacterization of L. capensis capensis is also a result ofthe absence of holotype material. Moreover, thedescription of several African hare populations assubspecies of L. capensis led to the acceptance of a highmorphological variability within this species and to therecognition of L. capensis in regions so far away fromSouth Africa as Europe or Asia.
Hoffmann (1998), discussing the taxonomic status ofL. capensis, mentioned that ‘‘there is a state of confusionaround this species which affects to different taxon pairsin eleven geographic contact zones’’. This confusion is due
in part to the emergence and development of criteria todistinguish some local populations within the polytypicL. capensis as valid species, as well as to the failure of theclassic taxonomic criteria set up in general revisions.
As for the validity of classic taxonomic criteria forLepus, the presence of a simple groove in the first upperincisors used by Petter (1959, 1961) to recognizeL. capensis in the three Old World continents hasrevealed to be uneffective. There is evidence thatL. capensis is not the only species showing this feature.Successive studies (Palacios 1996; Palacios and LopezMartınez 1980) showed that hares from Europe(L. castroviejoi, L. granatensis, L. europaeus andL. corsicanus) also are characterized by a simple incisorgroove with a moderate cement filling. The presence ofcement in the incisor groove, used by Robinson (1986) todistinguish L. capensis from L. europaeus, based onForsyth Major’s (1898) statement that L. europaeus lackscement in the incisor groove, also must be considered aninvalid discriminant character. In fact, our data show thatabout 60% of L. europaeus individuals from differentEuropean populations have cement in the incisor groove.
Today, the distribution of L. capensis is considered toinclude Southern, Northern and Eastern Africa, theMiddle East, and Western Asia (Flux and Angermann1990; Hoffmann 1993). Its European distribution isexclusively limited to Sardinia (Amori et al. 1996).
In the light of the confusion over range, distribution,and species limits, it becomes clear that there is apressing need to clarify and define the taxonomy ofAfrican, Asian, and European hares nominally identi-fied as L. capensis. Such an investigation has a specialweight and priority in Cape Province, South Africa,wherein is located the type locality of L. capensis.
The purpose of this paper is the morphologicalcharacterization of Lepus capensis capensis. An examina-tion and definition of the morphological variation in thetype locality would enable its identification fromphenotypic traits. This is necessary in order to comparepopulations putatively belonging to the nominal sub-species with other populations attributed to the samespecies in Africa, Europe and Asia; this study also will bea starting point for further systematic studies of Lepus.
The study area was limited to western Cape Province.In this area are also included the distributional ranges oftwo additional subspecies of L. capensis: L. c. grantii
and L. c. centralis. These subspecies were recognized byRoberts (1951), but have remained poorly defined froma morphological perspective.
Material and methods
General aspects
The studied specimens are held in most cases under the
species name L. capensis in the collections of the Transvaal

ARTICLE IN PRESSF. Palacios et al. / Mamm. biol. 73 (2008) 358–370360
Museum (TM), Pretoria; Amathole Museum, formerly Kaf-
frarian Museum (KM), King Williams Town; National
Museum (NMB), Bloemfontein; South African Museum
(ZM), Cape Town; Natural History Museum, formerly British
Museum of Natural History (BMNH), London; Museum
National d’Histoire Naturelle (MNHN), Paris; Naturhistor-
isches Museum (NMW), Vienna; National Museum of
Natural History, formerly United States National Museum
(USNM), Washington, DC; Carnegie Museum of Natural
History (CMNH), Pittsburgh.
The morphological study was based on skull measurements,
dental characters in occlusal and crossed sections, and
pelage design of adult specimens of relative age IV. Only a
few specimens of relative age between III and IV have been
added to complete the samples. Specimens of relative ages I, II
and III have been studied only for identification. The latter
two age classes, composed of subadults, are abundant in the
studied collections but have not been included in the analyses
to avoid population range overlap in cranial and dental
measurements and in morphological character states. Only
two juvenile specimens of relative age I have been used to
demonstrate the existence of species differences also in young
animals.
Relative age estimations have been made on the
basis of cranial ossification and development (Cabon-
Raczynska 1964; Palacios 1989, 1996), and considering as
adults the specimens showing the following features: sutures
between basioccipitale-exooccipitale and exooccipitale-supraoc-
cipitale ossified; interparietal bones ossified; sutura sagittalis
ossified in its posterior portion; osseous coating of the
mandibular capitulum knitted and polished; well-developed,
non-porous supraorbital bones; well developed, angular-
shaped temporal tubercle; premaxillaries and corpus mandibu-
lae not porous.
The studied specimens’ provenance is western Cape
Province. Some specimens are from the surroundings of
Cape Town, between Saldanha Bay and Cape Agulhas,
and other from the Upper Karroo, Lower Karroo, Little
Namaqualand and Bushman Land (see Roberts 1951).
In particular, the analysis of the collections of the South
African Museum and Amathole Museum allowed us to
substantially increase the amount of L. capensis material from
the Cape of Good Hope and surrounding area with respect to
previous studies (Roberts 1951; Robinson and Dippenaar
1987).
Collection numbers, localities, and relative age of the
studied specimens are listed in Table 1. The complete list of
the examined specimens that includes juveniles, subadults, and
adults may be requested to the authors.
The dental nomenclature used in this study follows
Palacios and Lopez Martınez (1980). The study of teeth
morphology variation was based on occlusal surface
enamel drawings except for the first upper incisor, where
both sectioned and occlusal surface enamel drawings were
used indistinctly. There was considerable consistency in
the incisor enamel pattern in a sample of 19 specimens in
terms of the studied character states in both occlusal surface
and cross-section enamel drawings. Drawings were carried
out using a camera lucida and refracted light on the teeth
surface which was perpendicularly oriented to their own
longitudinal axes.
Measuring protocol
We used a set of 44 cranial, 7 mandibular, 5 dental, and 3
foramina measurements (see Table 2). Description of these
measurements follows Cabon-Raczynska (1964) and Palacios
(1989, 1996). To avoid transcription errors while collecting
data, measurements were recorded to the nearest 0.01mm
using a digital caliper connected to a laptop computer using
the automatic data capturing software EASYCAL (Marcus
and Reig 1991).
A measurement error test was performed to assess the
precision of the morphometric characters. Repeatability was
calculated by measuring a series of 10 skulls from a single
locality three times and estimating the variance among skulls
and within repeats (repeatability ¼ s2 among/s2 within+s2
among; Baley and Byrnes 1990). Repeatability was over 92%
in all measurements.
Data analysis
Sample statistics were calculated separately for each group,
pooling together specimens from both sexes. To assess the
significance of potential differences between species in mean
values, Student’s T test was performed on each variable. The
univariate statistical tests were performed to illustrate the main
taxonomic differences between the two species using a wide
variety of variables. Thus, no correction for multiple
comparisons (i.e. Bonferroni) was performed, under the
assumption that the analysis was rather exploratory, and not
intended to confirm or discard the taxonomic differentiation
between the two species. The main conclusion about morpho-
metric differentiation between the two species is drawn from
the multivariate analysis that follows.
Principal component analysis (PCA) was performed to
transform a set of linear measurements into a set factors that
joined those measurements having a similar and independent
variability patterns. The principal components (PC) extracted
served to describe the main patterns of variability within the
whole sample without any a priori hypothesis about the species
assignment of each specimen. Factors were extracted from the
variance–covariance matrix of log-transformed values (Marcus
1990) using PROC PRINCOMP (SAS). Extracted PC were not
rotated to ensure that they represented the main axis of
allometric variability in the data. A subset of 10 traits was
selected for the PCA. This reduction in the number of variables
was aimed to get a better ratio in the number of variables relative
to the number of skulls measured for multivariate analysis, while
providing an adequate depiction of skull morphology. Selection
of the 10 morphometric characters was made according to three
basic criteria: depiction of overall skull dimensions (i.e. trying to
produce an even coverage of skull shape), power to discriminate
among populations or species, and maximum repeatability.
To test for differences between the two species in the
multivariate space of the 10 variables used in the PCA, a
MANOVA was performed using the log-transformed vari-
ables. Prior to the MANOVA analyses, the assumption of
homogeneity of variance–covariance matrices across groups
was tested to p40.05 using PROC DISCRIM (SAS). All the
data analyses were performed using the software developed by
SAS Institute Inc. (1989).

ARTICLE IN PRESS
Table 1. Adult specimens used for the study of skull morphology and univariate and multivariate analyses of skull dimensions and
for the study of teeth morphology
Species Relative age Collection number Locality Skull Teeth
L. capensis IV KM 10486 Compagnies Drift X X
KM 10488 Compagnies Drift X X
KM 10490 Compagnies Drift X X
KM 10493 Compagnies Drift X
KM 10504 Compagnies Drift X
KM 10505 Compagnies Drift X X
KM 10512 Compagnies Drift X X
KM 13040 Gouna, E Calvinia X
KM 30196 Darling X X
ZM 7212 Stellenbosch X X
ZM 13673a Stellenbosch X X
ZM 13673b Stellenbosch X X
ZM 35556 Hopefield, Malmesbury X X
III–IV KM 10487 Compagnies Drift X
KM 10494 Compagnies Drift X
KM 10495 Compagnies Drift X X
KM 10498 Compagnies Drift X
KM 10507 Compagnies Drift X
TM 8859 Eendekuil X
TM 8861 Eendekuil X
TM 41457 Langebaan X
L. centralis IV KM 10482 Eenriet X X
KM 10491 Compagnies Drift X X
KM 10496 Compagnies Drift X X
KM 10503 Compagnies Drift X
KM 10510 Compagnies Drift X
KM 11501 Deelfontein X
KM 11503 Deelfontein X
KM 13020 Calvinia X
KM 13022 Calvinia X
KM 13027 Gouna, E Calvinia X
KM 14850 Sutherland X
KM 14852 Sutherland X
KM 18226 Beaufort West X
KM 29028 W Williston X
KM 29032 Fraserburg X X
ZM 19131 Ezelfontein X X
TM 6022 Calvinia X
TM 27292 Farm Gonas, W Carnarvon X
TM 27293 Farm Gonas, W Carnarvon X X
TM 27296 Farm Gonas, W Carnarvon X
TM 28049 Bitterfontein X
TM 28963 Springbok X
TM 28979 Hutchinson X
TM 28980 Hutchinson X X
TM 28981 Hutchinson X X
TM 28982 Hutchinson X
TM 28983 Hutchinson X
TM 28984 Hutchinson X
TM 32452 Farm Vissenvil, Kenhardt X
TM 37999 Deelfontein X
BMNH 4.2.3.109 Port Nolloth X
BMNH 2.9.1.100 Deelfontein X X
III–IV KM 10483 Eenriet X
F. Palacios et al. / Mamm. biol. 73 (2008) 358–370 361
ARTICLE IN PRESS
Table 1. (continued )
Species Relative age Collection number Locality Skull Teeth
KM 10485 Ezelfontein X
KM 13041 E Calvinia X
KM 23246 Victoria West X
KM 23247 Victoria West X
ZM 8576 Namaqualand X
TM 2156 Lamberts Bay X
TM 7308 Vredendal X
Table 2. Skull measurements and their abbreviations.
Abbreviation Measurement
AH Alveolar height, between external margin of
alveoli dentales and the top of bulla alveolaris
maxillae
ANW Anterior nasal width
BCL Brain-case length (Acrocranium—
intersection of frontal suture with the line
joining the two anterior borders of incisura
postorbitalis)
BCW Brain case width
BL Basal length (Prosthion–Basion)
CBL Condylobasal length (Prosthion—posterior
plane of the occipital condyle)
CSL Coronary suture length
DFZ Diameter of foramen zygomaticus
DLFP Diameter of lateral foramen palatinum
DPFP Diameter of posterior foramen palatinum
DIRIMS Distance between first upper incisor
posterior end and sutura incisivomaxillaris
EDMA External diameter of meatus acusticus
ENL External nasal length
FIL Frontoincisive (facial) length (Prosthion—
intersection of frontal suture with the line
joining the two anterior borders of incisura
postorbitalis)
FSL Frontal suture length
FTL Facial tubercle length
FUISW First upper incisor section width
FUISL First upper incisor section length
HBC Height of the brain case between the
basisphenoideum and ossa parietalia
HCMM1 Height of corpus mandibulae at lower first
molar level
HDFM Horizontal diameter of foramen magnum
HPF Height between processus palatinus and the
nasal processes of frontal bone
IFL Foramen incisivum length
IFW Incisive foramina width
INL Internal nasal length
LCTRL Lower cheek-tooth row length
LMRML Length of ramus mandibulae
LP3SL Lower third premolar section length
LP3SW Lower third premolar section width
MCL Mandible capitulum length
MLAP Mandible length, taken from the angular
process
Table 2. (continued )
Abbreviation Measurement
MLCP Mandible length, taken from the capitulum
MH Mandible height between inc. varosum and
the capitulum
MMPH Height of processus muscularis mandibulae,
between the anterior margin of processus
articularis and the tip of the process,
perpendicular to the margin
NPW Narrowing of the palatine bridge
ONL Occipitonasal length (anterior nasal tip—
Acrocranion)
PL Palatal length
PNW Posterior nasal width
PPW Postpalatal width
POW Preorbital width
PZW Posterior zygomatic width
RIC Radius of first upper incisor curvature
RW Rostral width
SFW Smallest frontal width
SOOL Length of medial part of pars parietalis of
squama ossis occipitalis
SOOW Width of medial part of pars parietalis of
squama ossis occipitalis
SSL Sagittal suture length
SSOL Sagittal suture ossification length
TBL Tympanic bullae length
TBW Tympanic bullae width
TL Total skull length (Prosthion–Acrocranion)
TW Tympanic width
UCTRL Upper cheek–tooth row length
UDL Upper diastema length
UP4SL Upper fourth premolar section length
VDFM Vertical diameter of foramen magnum
WSP Width between supraorbital processes
WFT Width between facial tubercles
ZAL Zygomatic arch length
F. Palacios et al. / Mamm. biol. 73 (2008) 358–370362
Results
Two hare populations were differentiated on the basisof their morphological features. Specimens in thesepopulations are in both cases hares showing simpleincisor grooves, previously identified as characteristic ofLepus capensis. The distinction between the two

ARTICLE IN PRESSF. Palacios et al. / Mamm. biol. 73 (2008) 358–370 363
populations is further supported by the homogeneity ofthe morphological characters within their respectiveareas of distribution and by the presence in somelocalities of specimens of the two populations. This factreflects the existence of partial sympatry in the limit oftheir areas of distribution, although due to the scarcityof available specimens it has not been possible todetermine if this situation also exists in additionalcontact areas.
The results enabled us to establish that the twopopulations belong to distinct species. One of them fitsin the distribution area of L. capensis capensis as definedby Roberts (1951). For this reason it is designated fromhere on as L. capensis. The second population fits withinthe areas of distribution of L. capensis centralis Thomas,1903 and L. capensis grantii Thomas and Schwann, 1904and is designated henceforth as L. centralis as that namehas taxonomic priority.
Fig. 2. Skull posterior view of adult specimens (relative age
IV) of L. capensis KM 10490 Compagnies Drift (left) and L.
centralis KM 28963 Springbok (right).
Skull morphology
Adult individuals of the two species show markedlydifferent skull morphologies (Figs. 1 and 2). Forinstance, the maxillary bone and the masticatoryapparatus are more robust in L. capensis than inL. centralis. Other differences can be noticed in thepalatine bridge, where the lamina horizontalis is wider in
Fig. 1. Skull dorsal and ventral views of adult specimens (relative
(above) and L. centralis (below). L. capensis specimens: ZM 35556 H
specimens: KM 18226 Beaufort West (adult); KM 13021 Calvinia (
L. capensis. This latter species also shows more massivefacial tubercles.
Lepus centralis is characterized by a more gracile skullin its rostral part, due to the narrowness of thepremaxillary region and a lesser development of thefacial tubercles. On the other hand, L. centralis shows agreater development of the posterior part of the skullwith respect to L. capensis, due to larger dimensions ofthe tympanic bullae and of the squama ossis occipitalis
(Fig. 2) as well as a larger width of the braincase.Lepus capensis also shows more developed nasal
bones lying in a more advanced and higher position
age IV) and juvenile specimens (relative age I) of L. capensis
opefield (adult); KM 18322 Bredasdorp (juvenile). L. centralis
juvenile).

ARTICLE IN PRESSF. Palacios et al. / Mamm. biol. 73 (2008) 358–370364
with respect to the premaxillary bone if compared toL. centralis. As far as this feature is concerned,L. capensis is more similar to the L. victoriae–L. saxatilis
group than to L. centralis, even if in L. capensis the nasalprotrusion never reaches that of the specimens of theabove mentioned group. The shape of the nasal bonesalso distinguishes L. capensis and L. centralis: theposterior border is squared in the former and roundedin the latter. Moreover, in L. capensis the external edgeof the parietal bone is rounded, while in L. centralis it isstraight and parallel to the internal edge.
The distinctive cranial characters of both species areevident since the first ontogenetic stages, as shown bythe young individuals of relative age I of both species(Fig. 1). It is worth remarking that the L. centralis
specimen shows much more developed tympanic bullae
in spite of its lesser size than the L. capensis specimen.The difference in the posterior contour of nasal bones isalso evident in Figure 1.
Skull dimensions
Univariate analysis
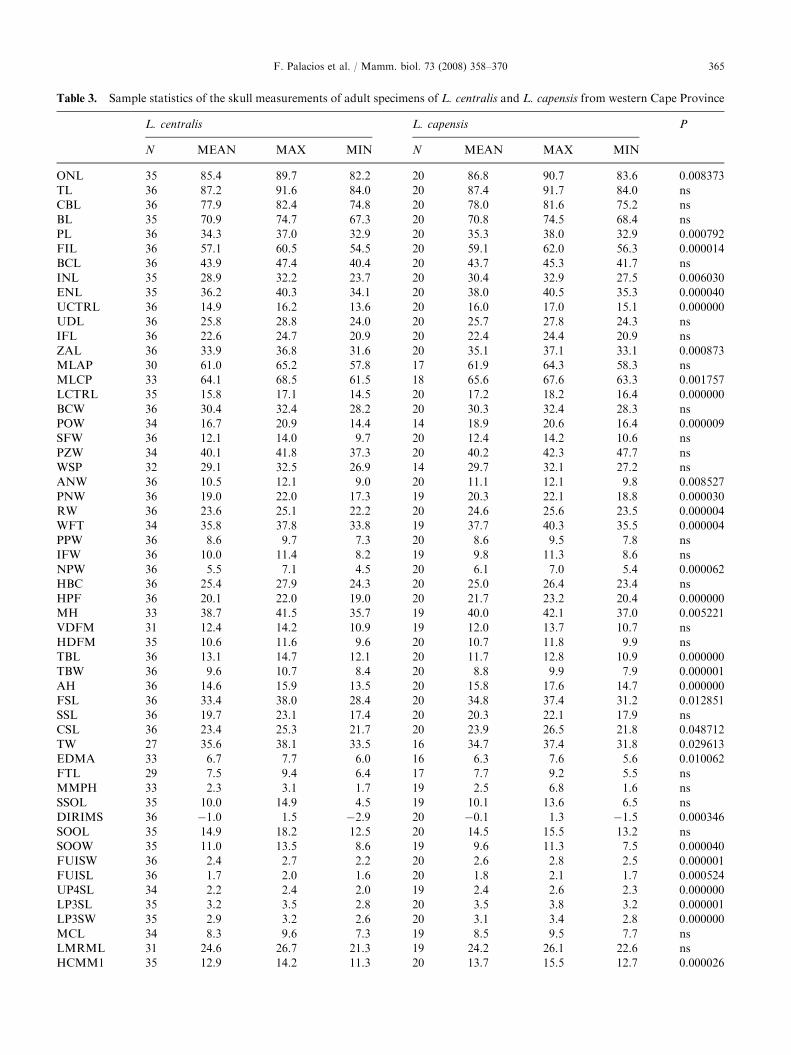
The results of the univariate statistical analysisperformed on cranial measurements of adult specimensof both populations are reported in Table 3. In spite ofthe general size of the skull being similar in the twospecies (see TL and PZW values), L. capensis dimen-sional values are significantly higher than those ofL. centralis in the following cranial measurements:HPF, AH, UCTRL, LCTRL, RW, WFT, POW, ENL,PNW, NPW, HCMM1, and in particular in dentalmeasurements UP4SL, LP3SW and FUISW. On theother hand, L. centralis shows significantly higher valuesof skull measurements FIL, TBL, TBW, DIRIMS andSOOW than L. capensis. Moreover, the ratio ONL/TLis higher in L. capensis due to its more prominentnasal border with respect to the premaxillary anterioredge. These results confirm the morphological observa-tions highlighted in the section on skull morphology(Figs. 1 and 2).
Multivariate analysis
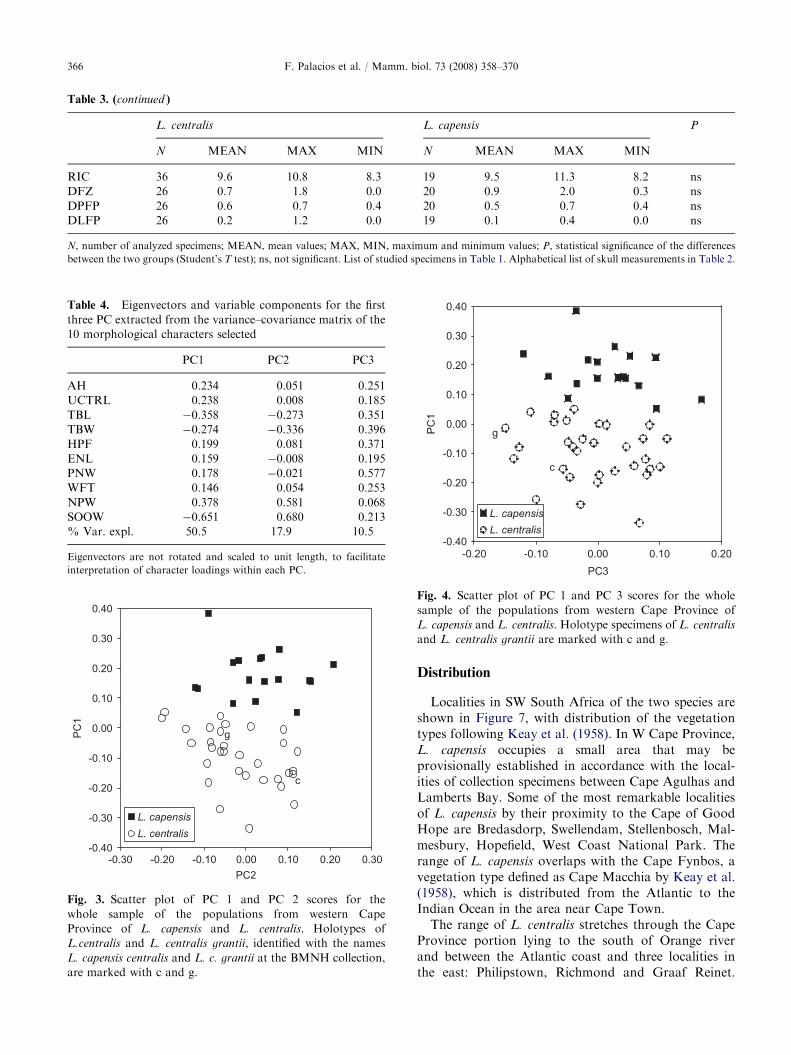
A PCA of skull measurements was performed.Although five PC were necessary to explain 90% ofthe variance, we focused on the description of the firstthree, which seem to convey the maximum informationabout morphometric differences between species. Thefirst three principal components explain 79%(50,5+17,9+10,5%) of the analyzed species variability(Table 4). The first PC describes the most evidentdifferences between the two species and can be inter-preted as the result of the combined variation of SOOWand in a lesser extent of TBL and TBW, as well as thatof NPW, AH and UCTRL in the opposite direction.
The second PC describes the combined variation ofSOOW and NPW (positive), and of TBL and TBW(negative). The third component describes variation ofgeneral size, particularly in PNW, HPF, TBW, TBL,AH and WFT. Bivariate plots of PC 1 versus PC 2 orPC 3 show a marked separation of the two species alongthese PC axes (Figs. 3 and 4). Thus, most of thevariability (80%) explaining differences between the twospecies is due to shape rather than size. This translatesinto a higher probability that such variables have ataxonomic rather than ecological significance (i.e. sizegradients related with climate or environmental factors).The MANOVA test performed on the set of 10 variablesindicated highly significant differences between the twospecies (F(10,38) ¼ 30.6, po0.00001).
Dental morphology
Data on the character states allowing the discrimina-tion between L. centralis and L. capensis are shown inFigure 5 and Table 5. The most important differencesbetween the two species are concentrated in the firstupper incisor, where L. centralis shows a narrow,squared I1/ internal lobe, slightly more prominent thanthe external one; its posterior contour is flat or slightlyconcave, and the incisor groove is very deep andcompletely filled with cement. L. capensis shows a wide,rounded and more prominent I1/ internal lobe; theposterior contour of the I1/ is generally concave and theincisor groove usually is more shallow and has littlecement.
A higher degree of crenulation and complexity in theenamel of P2/, P3/ and P/3 entrants is also noticeable inL. capensis, especially in the number of loops in theanterior side of the P3/ hypoflexus. In P/3 morphology,it also stands out that the length and width of theocclusal section in L. centralis are approximately thesame and the internal contour is straight, while inL. capensis the tooth section is longer and its internalcontour is rounded.
Pelage pattern
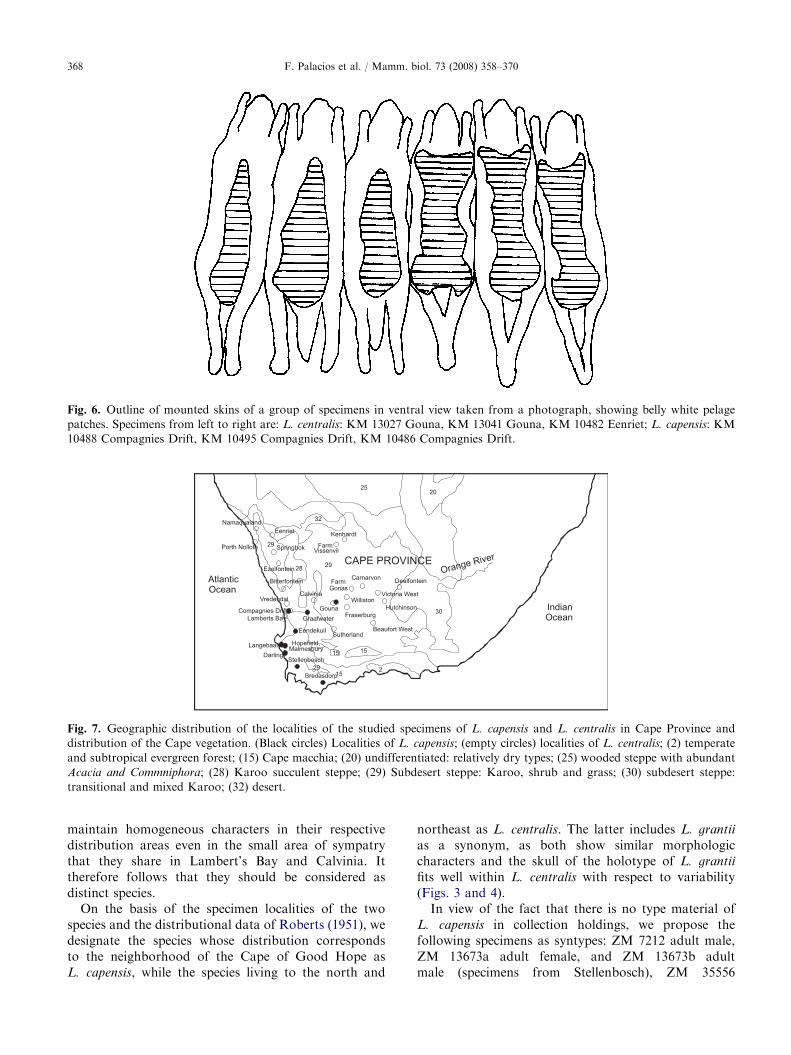
White pelage is more extended in the underparts ofL. capensis than in L. centralis (Fig. 6). In the former,the white pelage reaches the posterior part of the breastand the base of the anterior limbs, while in L. centralis itis reduced to a patch along the centre of the belly,separated from breast and limbs by the transitionalvinaceous-pinkish pelage of the sides. These resultsagree with the pelage description of the subspecies ofL. capensis capensis and L. capensis centralis by Roberts(1951). Nevertheless, the study of pelage color deservesfurther investigation as it is necessary to determine theextent of potential intrapopulation variability.
ARTICLE IN PRESS
Table 3. Sample statistics of the skull measurements of adult specimens of L. centralis and L. capensis from western Cape Province
L. centralis L. capensis P
N MEAN MAX MIN N MEAN MAX MIN
ONL 35 85.4 89.7 82.2 20 86.8 90.7 83.6 0.008373
TL 36 87.2 91.6 84.0 20 87.4 91.7 84.0 ns
CBL 36 77.9 82.4 74.8 20 78.0 81.6 75.2 ns
BL 35 70.9 74.7 67.3 20 70.8 74.5 68.4 ns
PL 36 34.3 37.0 32.9 20 35.3 38.0 32.9 0.000792
FIL 36 57.1 60.5 54.5 20 59.1 62.0 56.3 0.000014
BCL 36 43.9 47.4 40.4 20 43.7 45.3 41.7 ns
INL 35 28.9 32.2 23.7 20 30.4 32.9 27.5 0.006030
ENL 35 36.2 40.3 34.1 20 38.0 40.5 35.3 0.000040
UCTRL 36 14.9 16.2 13.6 20 16.0 17.0 15.1 0.000000
UDL 36 25.8 28.8 24.0 20 25.7 27.8 24.3 ns
IFL 36 22.6 24.7 20.9 20 22.4 24.4 20.9 ns
ZAL 36 33.9 36.8 31.6 20 35.1 37.1 33.1 0.000873
MLAP 30 61.0 65.2 57.8 17 61.9 64.3 58.3 ns
MLCP 33 64.1 68.5 61.5 18 65.6 67.6 63.3 0.001757
LCTRL 35 15.8 17.1 14.5 20 17.2 18.2 16.4 0.000000
BCW 36 30.4 32.4 28.2 20 30.3 32.4 28.3 ns
POW 34 16.7 20.9 14.4 14 18.9 20.6 16.4 0.000009
SFW 36 12.1 14.0 9.7 20 12.4 14.2 10.6 ns
PZW 34 40.1 41.8 37.3 20 40.2 42.3 47.7 ns
WSP 32 29.1 32.5 26.9 14 29.7 32.1 27.2 ns
ANW 36 10.5 12.1 9.0 20 11.1 12.1 9.8 0.008527
PNW 36 19.0 22.0 17.3 19 20.3 22.1 18.8 0.000030
RW 36 23.6 25.1 22.2 20 24.6 25.6 23.5 0.000004
WFT 34 35.8 37.8 33.8 19 37.7 40.3 35.5 0.000004
PPW 36 8.6 9.7 7.3 20 8.6 9.5 7.8 ns
IFW 36 10.0 11.4 8.2 19 9.8 11.3 8.6 ns
NPW 36 5.5 7.1 4.5 20 6.1 7.0 5.4 0.000062
HBC 36 25.4 27.9 24.3 20 25.0 26.4 23.4 ns
HPF 36 20.1 22.0 19.0 20 21.7 23.2 20.4 0.000000
MH 33 38.7 41.5 35.7 19 40.0 42.1 37.0 0.005221
VDFM 31 12.4 14.2 10.9 19 12.0 13.7 10.7 ns
HDFM 35 10.6 11.6 9.6 20 10.7 11.8 9.9 ns
TBL 36 13.1 14.7 12.1 20 11.7 12.8 10.9 0.000000
TBW 36 9.6 10.7 8.4 20 8.8 9.9 7.9 0.000001
AH 36 14.6 15.9 13.5 20 15.8 17.6 14.7 0.000000
FSL 36 33.4 38.0 28.4 20 34.8 37.4 31.2 0.012851
SSL 36 19.7 23.1 17.4 20 20.3 22.1 17.9 ns
CSL 36 23.4 25.3 21.7 20 23.9 26.5 21.8 0.048712
TW 27 35.6 38.1 33.5 16 34.7 37.4 31.8 0.029613
EDMA 33 6.7 7.7 6.0 16 6.3 7.6 5.6 0.010062
FTL 29 7.5 9.4 6.4 17 7.7 9.2 5.5 ns
MMPH 33 2.3 3.1 1.7 19 2.5 6.8 1.6 ns
SSOL 35 10.0 14.9 4.5 19 10.1 13.6 6.5 ns
DIRIMS 36 �1.0 1.5 �2.9 20 �0.1 1.3 �1.5 0.000346
SOOL 35 14.9 18.2 12.5 20 14.5 15.5 13.2 ns
SOOW 35 11.0 13.5 8.6 19 9.6 11.3 7.5 0.000040
FUISW 36 2.4 2.7 2.2 20 2.6 2.8 2.5 0.000001
FUISL 36 1.7 2.0 1.6 20 1.8 2.1 1.7 0.000524
UP4SL 34 2.2 2.4 2.0 19 2.4 2.6 2.3 0.000000
LP3SL 35 3.2 3.5 2.8 20 3.5 3.8 3.2 0.000001
LP3SW 35 2.9 3.2 2.6 20 3.1 3.4 2.8 0.000000
MCL 34 8.3 9.6 7.3 19 8.5 9.5 7.7 ns
LMRML 31 24.6 26.7 21.3 19 24.2 26.1 22.6 ns
HCMM1 35 12.9 14.2 11.3 20 13.7 15.5 12.7 0.000026
F. Palacios et al. / Mamm. biol. 73 (2008) 358–370 365
ARTICLE IN PRESS
Table 3. (continued )
L. centralis L. capensis P
N MEAN MAX MIN N MEAN MAX MIN
RIC 36 9.6 10.8 8.3 19 9.5 11.3 8.2 ns
DFZ 26 0.7 1.8 0.0 20 0.9 2.0 0.3 ns
DPFP 26 0.6 0.7 0.4 20 0.5 0.7 0.4 ns
DLFP 26 0.2 1.2 0.0 19 0.1 0.4 0.0 ns
N, number of analyzed specimens; MEAN, mean values; MAX, MIN, maximum and minimum values; P, statistical significance of the differences
between the two groups (Student’s T test); ns, not significant. List of studied specimens in Table 1. Alphabetical list of skull measurements in Table 2.
Table 4. Eigenvectors and variable components for the first
three PC extracted from the variance–covariance matrix of the
10 morphological characters selected
PC1 PC2 PC3
AH 0.234 0.051 0.251
UCTRL 0.238 0.008 0.185
TBL �0.358 �0.273 0.351
TBW �0.274 �0.336 0.396
HPF 0.199 0.081 0.371
ENL 0.159 �0.008 0.195
PNW 0.178 �0.021 0.577
WFT 0.146 0.054 0.253
NPW 0.378 0.581 0.068
SOOW �0.651 0.680 0.213
% Var. expl. 50.5 17.9 10.5
Eigenvectors are not rotated and scaled to unit length, to facilitate
interpretation of character loadings within each PC.
-0.40
-0.30
-0.20
-0.10
0.00
0.10
0.20
0.30
0.40
-0.30 -0.20 -0.10 0.00 0.10 0.20 0.30
PC2
PC
1
L. capensis
L. centralis
g
c
Fig. 3. Scatter plot of PC 1 and PC 2 scores for the
whole sample of the populations from western Cape
Province of L. capensis and L. centralis. Holotypes of
L.centralis and L. centralis grantii, identified with the names
L. capensis centralis and L. c. grantii at the BMNH collection,
are marked with c and g.
-0.40
-0.30
-0.20
-0.10
0.00
0.10
0.20
0.30
0.40
-0.20 -0.10 0.00 0.10 0.20
PC3
PC
1
L. capensis
L. centralis
g
c
Fig. 4. Scatter plot of PC 1 and PC 3 scores for the whole
sample of the populations from western Cape Province of
L. capensis and L. centralis. Holotype specimens of L. centralis
and L. centralis grantii are marked with c and g.
F. Palacios et al. / Mamm. biol. 73 (2008) 358–370366
Distribution
Localities in SW South Africa of the two species areshown in Figure 7, with distribution of the vegetationtypes following Keay et al. (1958). In W Cape Province,L. capensis occupies a small area that may beprovisionally established in accordance with the local-ities of collection specimens between Cape Agulhas andLamberts Bay. Some of the most remarkable localitiesof L. capensis by their proximity to the Cape of GoodHope are Bredasdorp, Swellendam, Stellenbosch, Mal-mesbury, Hopefield, West Coast National Park. Therange of L. capensis overlaps with the Cape Fynbos, avegetation type defined as Cape Macchia by Keay et al.(1958), which is distributed from the Atlantic to theIndian Ocean in the area near Cape Town.
The range of L. centralis stretches through the CapeProvince portion lying to the south of Orange riverand between the Atlantic coast and three localities inthe east: Philipstown, Richmond and Graaf Reinet.

ARTICLE IN PRESS
L. c
a pe n
sis
P2/I1/
1 mmL
.ce n
t ral
is
P3/ P/3
Fig. 5. Comparison of teeth morphologies in L. capensis (above) and L. centralis (below): I1/, upper first incisor; P2/, upper second
premolar; P3/, upper third premolar, P/3; lower third premolar. See text and Tables 1 and 5 for further details. Drawings of L.
centralis correspond to specimen KM 10491 Compagnies Drift; those of L. capensis to ZM 13673a Stellenbosch, except for P/3
which corresponds to KM 10504 Compagnies Drift.
Table 5. Comparison of some selected characters of occlusal and crossed dental sections in L. capensis and L. centralis
Dental
element
Character L. centralis L. capensis
I1/ General shape Somewhat squared Somewhat rectangular
Internal lobe shape Narrow, squared, partially higher than
external one
Broad, rounded, fully elevated
Posterior contour Usually flat Slightly concave
Groove depth Very deep Deep
Cement Very abundant Not abundant
P2/ Crenulation depth on paraflexus
anterior edge
Absent or very shallow undulation Shallow to medium depth
Loop number on paraflexus
anterior edge
0–1 Usually 1 or more
P3/ Hypercones relative thickness Mesial hypercone approximately as thick as
distal hypercone
Mesial hypercone thicker than
distal hypercone
Loop density on hypoflexus
anterior edge
Usually less than 7 Usually 7 or more
P/3 Lingual anteroconid outline Smooth Wavy
Paraflexid depth (when present) Deep Very deep
Crenulation posterior edge
hypoflexid
Smooth or slightly undulated Usually slightly undulated
Lingual contour Straight Rounded
I1/, upper first incisor; P2/, upper second premolar; P3/, upper third premolar; P/3, lower third premolar. List of studied specimens in Table 1.
F. Palacios et al. / Mamm. biol. 73 (2008) 358–370 367
The southern localities of L. centralis near the range ofL. capensis are, from east to west, Beaufort West,Sutherland, Kruidfontein, Calvinia and Lamberts Bay.In the latter two localities (specifically in CompagniesDrift and Gouna), sympatry between the two species hasbeen detected. However, it is noticeable that nocollection examined holds L. centralis specimens fromthe L. capensis exclusive area, between Cape Agulhasand Cape Columbine, suggesting that the two speciesalso have distinct areas of parapatry.
The range of L. centralis overlaps with vegetationtypes defined by Keay et al. (1958) as Subdesert Steppe(Karoo shrub and grass) and Karoo Succulent Steppe.
Discussion
The two hare populations differ distinctly in theircranial and dental size and morphology, pelage colorpattern, and geographical distribution. Moreover, they
ARTICLE IN PRESS
Fig. 6. Outline of mounted skins of a group of specimens in ventral view taken from a photograph, showing belly white pelage
patches. Specimens from left to right are: L. centralis: KM 13027 Gouna, KM 13041 Gouna, KM 10482 Eenriet; L. capensis: KM
10488 Compagnies Drift, KM 10495 Compagnies Drift, KM 10486 Compagnies Drift.
CAPE PROVINCE
AtlanticOcean
IndianOcean
Orange River
29
15 15
152
29
2928
32
30
Kenhardt
Williston
FarmVissenvil
FarmGonas
CarnarvonDeelfontein
Victoria West
Hutchinson
Beaufort West
FraserburgGouna
Sutherland
Porth Nolloth
Namaqualand
Eenriet
Springbok
Ezelfontein
Bitterfontein
VredendalCalvinia
Compagnies Drift
Lamberts Bay Graafwater
Langebaan
Darling
Hopefield,Malmesbury
Stellenbosch
Bredasdorp
Eendekuil
2520
Fig. 7. Geographic distribution of the localities of the studied specimens of L. capensis and L. centralis in Cape Province and
distribution of the Cape vegetation. (Black circles) Localities of L. capensis; (empty circles) localities of L. centralis; (2) temperate
and subtropical evergreen forest; (15) Cape macchia; (20) undifferentiated: relatively dry types; (25) wooded steppe with abundant
Acacia and Commniphora; (28) Karoo succulent steppe; (29) Subdesert steppe: Karoo, shrub and grass; (30) subdesert steppe:
transitional and mixed Karoo; (32) desert.
F. Palacios et al. / Mamm. biol. 73 (2008) 358–370368
maintain homogeneous characters in their respectivedistribution areas even in the small area of sympatrythat they share in Lambert’s Bay and Calvinia. Ittherefore follows that they should be considered asdistinct species.
On the basis of the specimen localities of the twospecies and the distributional data of Roberts (1951), wedesignate the species whose distribution correspondsto the neighborhood of the Cape of Good Hope asL. capensis, while the species living to the north and
northeast as L. centralis. The latter includes L. grantii
as a synonym, as both show similar morphologiccharacters and the skull of the holotype of L. grantii
fits well within L. centralis with respect to variability(Figs. 3 and 4).
In view of the fact that there is no type material ofL. capensis in collection holdings, we propose thefollowing specimens as syntypes: ZM 7212 adult male,ZM 13673a adult female, and ZM 13673b adultmale (specimens from Stellenbosch), ZM 35556

ARTICLE IN PRESSF. Palacios et al. / Mamm. biol. 73 (2008) 358–370 369
adult (Hopefield, Malmesbury), KM 18322 juvenile(Bredasdorp), and KM 30196 adult female (Darling).
This work constitutes a morphological characteriza-tion of L. capensis in the area in which the species’type locality is putatively included, such that theconcrete characters which define L. capensis canhenceforth be used comparatively in analysis of otherspecies and populations. Until now, the results ofprevious taxonomic work, both the classical studies(Roberts 1951) and the recent morphometric studies(Robinson and Dippenaar 1987), included all themedium sized hares from South Africa showing a simplegroove in the first upper incisor in the speciesL. capensis, thus accepting an excess of morphologicvariability in a single species. This consequently led tomorphologic range overlap.
In Europe, where the scarce knowledge on haretaxonomy has been similar in the past decades, somehare populations referred to L. capensis by Petter (1961)have been recently recognized as independent species. Inparticular: L. granatensis (Iberian peninsula), L. corsi-
canus (Italian peninsula and Sicily), and L. europaeus
(widespread in continental Europe) have been consid-ered valid species on the basis of morphology andchorology (Palacios 1983, 1989, 1996), in concordantcombination with biochemical (Bonhomme et al. 1986)and molecular analyses (Perez-Suarez et al. 1994;Pierpaoli et al. 1999; Estonba et al. 2006).
Further research will determine whether the dis-tribution of L. capensis spreads to the east of theCape of Good Hope, and whether it is bigger thanthe small area that surrounds the type locality. It mayalso be possible to determine whether other haremetapopulations from Southern Africa belong toL. capensis.
L. centralis is provisionally recognized as a fullspecies, so that the appropriate distinction can beestablished with L. capensis in the study area. Newinvestigations taking into account material from neigh-boring areas will allow the recognition of a definitivename for that population considering synonyms andpriorities. Future research should also encompassmolecular studies to shed further light on the character-ization of L. capensis in the type locality that can be usedcomparatively in taxonomic analyses of other speciesand populations.
Acknowledgements
We thank the curators, collection managers andtechnicians charged of the studied mammal collectionsfor their help: Lloyd Wingate, Fred Kigozi, AmatholeMuseum (King William’s Town); Stephan du Toit, NicoAvenant, Johan Eksteen, National Museum (Bloemfon-tein); Liz Herzholdt, Naas Rautenbach, Gary Bronner,
Teresa Kearney, Duncan MacFadyen, Transvaal Mu-seum (Pretoria); Michael Cluver, Denise Hammerton,Melissa Stander, South African Museum (Cape Town);Paulina Jenkins, Richard Harbord, Natural HistoryMuseum (London); Francis Petter, Michel Tranier,Jacques Cuisin, Museum National d’Histoire Naturelle(Paris); Friederike Spitzenberger, NaturhistorischesMuseum (Vienna); Michael Carleton, National Museumof Natural History (Washington); and Duane Schlitter,Steve Williams, Carnegie Museum of Natural History(Pittsburgh). We also thank Rainer Hutterer for theabstract translation to German. The comments of LuisRuedas, the anonymous reviewer, and the editor werevery helpful to improve the manuscript and areacknowledged. Financial support is greatly acknowl-edged from the Consejerıa de Educacion, Community ofMadrid, and the Direccion General de Investigacion,Secretarıa de Estado de Polıtica Cientıfica y Tecnologicaof Spain.
Zusammenfassung
Morphologische Hinweise auf artliche Differenzierung
innerhalb des Taxons Lepus capensis Linnaeus, 1758
(Leporidae, Lagomorpha) in der Kapprovinz Sudafrikas
Die Hasen (Lepus) der Kap-Provinz, die bisher denUnterarten L. capensis capensis, L. c. centralis, und L. c.
grantii zugerechnet werden, wurden morphologischuntersucht. Das Ziel der Studie war, die Art L. capensis
Linnaeus, 1758 naher zu charakterisieren. Dabei konn-ten zwei distinkte Populationen unterschieden werden,die wir als verschiedene Arten ansehen, da innerhalbbeider Populationen homogene Schadel-, Zahn- undFellmerkmale existieren. Die eine Art, die wir alsL. capensis definieren, kommt bei Kapstadt in Kusten-nahe, zwischen Lambert’s Bay und Cape Agulhas, vor.Die andere Art, die wir als L. centralis definieren (mitgrantii als Synonym), ist in der zentralen und westlichenKap-Provinz verbreitet. L. capensis und L. centralis sindparapatrisch verbreitet mit einem kleinen Sympatrie-Areal in der Kontaktzone im Compagnies Drift Gebietnahe Lambert’s Bay. Differenzen im Schadelbau beiderArten sind bei L. capensis durch eine kraftigere Maxillaund robustere Zahnreihen gegeben, wahrend L. centralis
großere Ohrblasen aufweist. Der erste obere Schneide-zahn von L. centralis ist in der Regel tiefer gefurcht undhat mehr Zement als bei L. capensis, auch ist der innereLobus bei der erstgenannten Art quadratisch und beiL. capensis abgerundet. In der Fellfarbung istL. capensis durch ein ausgedehnteres weißes Bauchfellvon L. centralis unterschieden. Unsere Resultate sindrelevant fur alle weiteren taxonomischen Untersuch-ungen an Hasen der Alten Welt, sofern Populationenvon L. capensis betroffen sind.

ARTICLE IN PRESSF. Palacios et al. / Mamm. biol. 73 (2008) 358–370370
References
Amori, G., Angelici, F.M., Prigioni, C., Vigna Taglianti, A.,
1996. The mammal fauna of Italy: a review. Hystrix 8,
3–7.
Baley, R.C., Byrnes, J., 1990. A new old method for assessing
measurement error in both univariate and multivariate
morphometric studies. Syst. Zool. 39, 124–130.
Bonhomme, F., Fernandez, J., Palacios, F., Catalan, J.,
Machordon, A., 1986. Caracterisation biochimique du
complexe d’especes du genre Lepus en Espagne. Mammalia
50, 495–506.
Cabon-Raczynska, K., 1964. Studies on the European hare.
III. Morphological variation of the skull. Acta Theriol. 9,
249–285.
Ellerman, J.R., Morrison-Scott, T.C.S., 1951. Checklist of
Palaeartic and Indian Mammals (1758 to 1946). Trustees of
The British Museum (National History), London.
Ellerman, J.R., Morrison-Scott, T.C.S., Hayman, R.W., 1953.
Southern African Mammals (1758–1951): A Reclassifica-
tion. Trustees of The British Museum (National History),
London.
Estonba, A., Solıs, A., Iriondo, M., Sanz-Martın, M.J., Perez-
Suarez, G., Markov, G., Palacios, F., 2006. The genetic
distinctiveness of the three Iberian hare species: Lepus
europaeus, L. granatensis, and L. castroviejoi. Mamm. Biol.
71, 52–59.
Flux, J.E.C., Angermann, R., 1990. The hares and jackrabbits.
In: Chapman, J.A., Flux, J.E.C. (Eds.), Rabbits, Hares and
Pikas. Status Survey and Conservation Action Plan. IUCN,
Gland, Switzerland, pp. 61–94.
Forsyth Major, C.I., 1898. On fossil and recent Lagomorpha.
Trans. Linn. Soc. Lond. 7, 433–520.
Hoffmann, R.S., 1993. Order Lagomorpha. In: Wilson, D.E.,
Reeder, D.M. (Eds.), Mammal Species of the World. A
Taxonomic and Geographic Reference. Smithsonian In-
stitution Press, Washington and London, pp. 807–827.
Hoffmann, R.S., 1998. The trouble with Lepus capensis. Euro-
American Mammal Congress, Santiago de Compostela,
p. 94 (abstract).
Keay, R.W.J., Aubreville, A., Duvigneaud, P., Hoyle, A.C.,
Mendonca, F.A., Pichi-Sermolli, R.E.G., 1958. Vegetation
Map of Africa South of the Tropic of Cancer. Oxford
University Press, London.
Marcus, L.F., 1990. Traditional morphometrics. In: Rohlf,
F.J., Bookstein, F.L. (Eds.), Proceedings of the Michigan
Morphometrics Workshop. Special Publication #2, Uni-
versity of Michigan Museum of Zoology, Ann Arbor.
Marcus, L.F., Reig, S., 1991. Data capturing software for
digital calipers. In: Proceedings of the 71st Annual Meeting
of the American Society of Mammalogists, Manhattan, KS
(abstract).
Palacios, F., 1983. On the taxonomic status of the genus Lepus
in Spain. Acta Zool. Fennica 174, 27–30.
Palacios, F., 1989. Biometric and morphologic features of the
species of the genus Lepus in Spain. Mammalia 73,
227–264.
Palacios, F., 1996. Systematics of the indigenous hares of Italy
traditionally identified as Lepus europaeus Pallas, 1778
(Mammalia, Leporidae). Bonn. Zool. Beitr. 46, 59–91.
Palacios, F., Lopez Martınez, N., 1980. Morfologıa dentaria
de las liebres europeas (Lagomorpha, Leporidae). Donana,
Acta Vert. 7, 61–81.
Perez-Suarez, G., Palacios, F., Boursot, P., 1994. Speciation
and paraphyly in western mediterranean hares (Lepus
castroviejoi, L. granatensis and L. capensis) revealed by
mytochondrial DNA phylogeny. Biochem. Genet. 32,
423–436.
Petter, F., 1959. Elements d’une revision des Lievres africains
du sous-genre Lepus. Mammalia 23, 41–67.
Petter, F., 1961. Elements d’une revision des Lievres europeens
et asiatiques du sous-genre Lepus. Zeitschr. f. Saugetierk.
26, 1–11.
Pierpaoli, M., Riga, F., Trocchi, V., Randi, E., 1999. Species
distinction and evolutionary relationships of the Italian
hare Lepus corsicanus as described by mitochondrial DNA
sequencing. Mol. Ecol. 8, 1805–1817.
Roberts, A., 1951. The Mammals of South Africa. Trustees of
the Mammals of South Africa Book Fund, Johannesburg.
Robinson, T.J., 1986. Incisor morphology as an aid in the
systematics of the South African Leporidae (Mammalia,
Lagomorpha). S. Afr. J. Zool. 21, 297–302.
Robinson, T.J., Dippenaar, N.J., 1987. Morphometrics of the
South African Leporidae. II. Lepus Linnaeus, 1758 and
Bunolagus Thomas, 1929. Ann. Transvaal Mus. 34,
379–404.
SAS Institute Inc., 1989. SAS Users Guide. Statistics. V6.3.
SAS Institute Inc., Cary, NC.




















