
Principais coleções brasileiras de Diptera: histórico e situação atual

UNIVERSIDADE FEDERAL DO ESPÍRITO SANTO
CENTRO DE CIÊNCIAS HUMANAS E NATURAIS
DEPARTAMENTO DE CIÊNCIAS BIOLÓGICAS
JOÃO PAULO MAIRES HOPPE
DISTRIBUIÇÃO ATUAL E POTENCIAL DE Vampyrum
spectrum (Linnaeus, 1758) (Chiroptera, Phyllostomidae)
VITÓRIA
2013

II
JOÃO PAULO MAIRES HOPPE
DISTRIBUIÇÃO ATUAL E POTENCIAL DE Vampyrum
spectrum (Linnaeus, 1758) (Chiroptera, Phyllostomidae)
Monografia apresentada ao Departamento de
Ciências Biológicas do Centro de Ciências
Humanas e Naturais da Universidade Federal do
Espírito Santo como requisito parcial para
obtenção do título de Bacharel em Ciências
Biológicas.
Orientador: Dr. Albert David Ditchfield
VITÓRIA
2013

III
JOÃO PAULO MAIRES HOPPE
DISTRIBUIÇÃO ATUAL E POTENCIAL DE Vampyrum
spectrum (Linnaeus, 1758) (Chiroptera, Phyllostomidae)
Monografia apresentada ao Departamento de Ciências Biológicas do Centro de
Ciências Humanas e Naturais da Universidade Federal do Espírito Santo como requisito
parcial para a obtenção do título de Bacharel em Ciências Biológicas.
Aprovada em ____ de ________________ de 20____
COMISSÃO EXAMINADORA
______________________________________________
Prof. Dr. Albert David Ditchfield
Universidade Federal do Espírito Santo (UFES)
Orientador
______________________________________________
Profa. Dr
a. Leonora Pires Costa
Universidade Federal do Espírito Santo (UFES)
______________________________________________
Prof. Dr. Vilácio Caldara Junior
Instituto Federal do Espírito Santo - Santa Teresa (IFES)

IV
SUMÁRIO AGRADECIMENTOS ..................................................................................................... V RESUMO ..................................................................................................................... VIII LISTA DE FIGURAS ...................................................................................................... X LISTA DE TABELAS ................................................................................................... XI 1 INTRODUÇÃO ........................................................................................ 1
2 MATERIAL & MÉTODOS .................................................................... 3 2.1 Banco de Dados ................................................................................... 3 2.1.1 Dados de Ocorrência Geográfica ............................................................ 3
2.1.2 Dados climáticos e de relevo .................................................................. 3
2.2 Modelagem Ecológica ......................................................................... 3
2.2.1 Estatística Pré-Modelagem ..................................................................... 3
2.2.2 Parâmetros dos Algoritmos ..................................................................... 4
2.3 Validação ............................................................................................. 5
2.3.1 Threshold ................................................................................................ 5
2.3.2 Pontos aleatórios ..................................................................................... 6
2.3.3 Matriz de confusão .................................................................................. 6
2.4 Projeção Combinada ........................................................................... 7
2.5 Similaridade ........................................................................................ 7 2.6 Mapas de Distribuição Ambiental ....................................................... 7
3 RESULTADOS ......................................................................................... 8 3.1 Distribuição Atual ............................................................................... 8 3.2 Distribuição Potencial ......................................................................... 8
3.2.1 Variáveis Escolhidas ............................................................................... 8
3.2.2 Modelos de Distribuição de Espécies ..................................................... 9
4 DISCUSSÃO ........................................................................................... 18 4.1 Distribuição Atual ............................................................................. 18
4.2 Modelagem Ecológica ....................................................................... 20 4.2.1 Validação dos Modelos ......................................................................... 20
4.2.2 Algoritmos: Definições, Operação e Estado da Arte ............................ 22
4.2.3 Modelos de Distribuição de Espécies (MDEs) ..................................... 25
4.2.4 Variáveis Ecogeográficas ..................................................................... 27
4.2.5 Áreas Prioritárias para Inventários ....................................................... 29
4.2.6 Ecologia ................................................................................................ 32
4.2.7 Inferências Biogeográficas.................................................................... 32
5 CONCLUSÕES ....................................................................................... 34
6 REFERÊNCIAS BIBLIOGRÁFICAS ................................................. 35 7 ANEXOS ................................................................................................. 41 7.1 Anexo 1: Pontos de Ocorrência de Vampyrum spectrum .................. 41 7.2 Anexo2: Lista de Instituições ............................................................ 45
7.3 Anexo 3: Fontes dos Pontos de Ocorrência ...................................... 46 7.4 Anexo 4: Variáveis Ecogeográficas .................................................. 49

V
AGRADECIMENTOS
Este trabalho não representa apenas a conclusão do curso de Ciências Biológicas,
mas também o fim de um longo caminho percorrido desde o início de 2008. Caminho
este que não poderia ser percorrido sozinho.
Agradeço à Universidade Federal do Espírito Santo (UFES) e o Laboratório de
Estudos em Quirópteros (LABEQ) pelo espaço e oportunidades concedidas para
desenvolvimento de meu trabalho. Ao Conselho Nacional de Pesquisa e
Desenvolvimento Tecnológico (CNPq), agradeço pela bolsa de iniciação científica. Sem
os auxílios e recursos fornecidos, esse trabalho não seria possível.
Agradeço aos membros da banca, Leonora Pires Costa e Vilácio Caldara Junior,
por aceitarem meu convite e contribuírem com este trabalho.
Sou grato a todos os professores que tive durante o curso. Os ensinamentos de
cada um de vocês fará parte de minha vida para sempre, mesmo que em determinados
momentos seja o que não fazer! Dentre os bons, no entanto, alguns são excelentes.
Gostaria de agradecer à professora Maria do Carmo Batitucci e ao professor Clayton
Perônico, pelo estímulo que me deram quando mais precisava. Também gostaria de
agradecer ao professor Alexandre Pires Aguiar, pelas oportunidades que me concedeu, e
pelas longas conversas pelos corredores e em sua sala. Aprendi muito com você.
Agradeço também ao professor Francisco Barreto, meu co-orientador. Obrigado
pelo espaço concedido no Simulacrum, pelas conversas e risos, e pela oportunidade.
Junto de Flávia Machado, vocês foram os responsáveis pela minha paixão recém-
descoberta pela Estatística!
Para Albert David Ditchfield, preciso agradecer por várias razões. Agradeço por
me acolher no LABEQ quando cheguei à UFES, careca e confuso, e por me receber
novamente após peregrinação pela Zoologia! Ainda mais que as três cadeiras que passei
sob sua tutela, sou grato especialmente ao que foi transmitido fora da sala de aula: de
você aprendi desde artes marciais medievais até filosofia da ciência, passando por
praticamente todas as áreas do conhecimento. Muito obrigado por tudo!
Agradeço os colegas de laboratórios. Sim, “laboratórios”: Adriana, Anazélia,
Bernardo, Marcus, Fernanda e Weverson no LAPIS; Aos colegas do LABEQ, Amanda,
Ianna, Luane, Bruna e Vinícius (o segundo maior biólogo da UFES!). Carlos, você foi
único: seus campos foram lendários! Day, Fran e Mariana, fiquem tranquilas: não

VI
esqueci as “parasitas”! Patrícia, Julia, Mirella, Raiane, Jordana e Fernanda (de novo!),
obrigado pela excelente companhia no Simulacrum.
Aos amigos do LGA, Eduardo Loyola, Marina Monjardim e Yuri Marins. O apoio
e amizade de vocês fez toda a diferença. Um abraço também aos parceiros da
entomologia, Bruno Cancian e Thales Brotto (o maior biólogo da UFES!), foi bom
trabalhar com vocês!
Arthur Machado, obrigado pela atenção e recursos. O Alpha shape não seria o
mesmo sem você. Ainda no LBCV, sou grato à Luana Centoducatte, pelos auxílios no
ArcGIS; Danielle Oliveira, pelos artigos difíceis de conseguir; e Gabriel Santos, pelos
bate-papos estatísticos e troca de reclamações da vida.
Sou grato aos meus pais, Paulo e Marta, à minha irmã, Paloma, e ao mais novo
membro da família, o pequeno Arthur. Obrigado pelo apoio em todos os momentos, e
por me escutarem horas sem fim, enquanto tentava transmitir meu fascínio pelas
ciências. Em cada momento em que fui chamado, aos gritos de “venha ver esse bicho!”,
tenham certeza de que um sorriso passou pelo meu rosto. Agradeço também à Randriely
Merscher, minha namorada, pelo carinho, amor e amizade.
Essa vitória é de todos nós!

VII
The Road goes ever on and on
Down from the door where it began.
Now far ahead the Road has gone,
And I must follow, if I can,
Pursuiting with eager feet,
Until it joins some larger way
Where many paths and errands meet.
And wither then? I cannot say.
Bilbo Baggins
The Lord of the Rings: The Fellowship of the Ring

VIII
RESUMO
Vampyrum spectrum (Linnaeus, 1758) se distingue por ser o único representante do
gênero monotípico Vampyrum Rafinesque, 1758, e por ser o maior morcego do
Ocidente, na subordem Vespertilioniformes. É um predador de topo de cadeia, raro em
inventários devido a sua baixa densidade populacional e relativa dependência por
florestas primárias. A espécie é classificada como Quase Ameaçada pela Lista
Vermelha da IUCN, com tendência de redução populacional no futuro. Apesar disso, o
conhecimento da distribuição da espécie é limitado a levantamentos locais. Neste
contexto, a Modelagem Ecológica pode ser uma ferramenta de auxílio eficaz, utilizando
pontos de ocorrência e dados ecogeográficos para elucidar os efeitos das variáveis
ambientais, padrões de distribuição, e regiões de distribuição potencial elevada para a
espécie. Neste trabalho, foi estudada a distribuição atual de Vampyrum spectrum; e
cinco algoritmos somente-Presença foram utilizados para (1) Determinar a distribuição
potencial, e (2) indicar as Áreas Protegidas, no Brasil, com maior probabilidade de
ocorrência. Os pontos de ocorrência foram compilados da literatura, indexadores online,
e do levantamento de Ditchfield em coleções de museus de cinco países. As variáveis
ambientais foram obtidas no WorldClim e U.S. Geological Survey. Os dados foram
tratados na plataforma R para eliminação de variáveis correlacionadas. Foram utilizados
cinco algoritmos: Maxent, GARP, SVM, ENFA e RF, com dois thresholds, e os
resultados destes foram combinados em três ensembles distintos. A validação foi por
meio da matriz de confusão e teste binomial de duas proporções. Foi computada a
similaridade dos modelos de acordo com os valores obtidos em uma Análise de
Componentes Principais. Foram levantados 126 pontos geográficos únicos para a
espécie, em 17 países da América Latina. As variáveis ambientais mais relacionadas à
distribuição foram relacionadas a valores mínimos de temperatura e pluviosidade. Os
algoritmos apresentaram grau variado de similaridade, exceto o RF. Os dados ampliam
o limite sul de distribuição da espécie em quase 6º, além de ocorrências na Caatinga,
bioma com condições diversas das previstas. A técnica do Alpha Shape para extensão
de ocorrência, utilizada no trabalho, mostra resultados mais conservadores do que os
utilizados de forma clássica na literatura (como localidades marginais e Polígono
Mínimo Convexo). Os Modelos de Distribuição de Espécies apontam regiões com
probabilidades de ocorrência (inferidas da adequabilidade ambiental alta) elevadas no
Brasil, especialmente na região Norte. São listadas 35 Áreas Protegidas nestes locais,
obtidos via IBGE, em condições favoráveis, mas sem registros de ocorrência para a

IX
espécie. Os dados ecológicos levantados pelo trabalho não são totalmente explicados
com o conhecimento atual, especialmente a ocorrência na Caatinga brasileira. São
levantadas três hipóteses: (1) capacidade migratória ainda desconhecida; (2)
classificação errônea das regiões intermediárias devido à disparidade entre os dados; (3)
populações relictuais. Mais estudos são necessários para elucidar este cenário. As
análises deste estudo também dão suporte para duas teorias na Biogeografia do Brasil: a
hipótese da Hiléia Baiana, e a existência de dois corredores distintos ligando a Floresta
Amazônica e a Mata Atlântica.

X
LISTA DE FIGURAS
Figura 1: Matriz de confusão ............................................................................................ 6
Figura 2: Alpha Shape de extensão de ocorrência de Vampyrum spectrum. .................... 8
Figura 3: MDE utilizando Maxent, threshold MTP. ...................................................... 10
Figura 4: MDE utilizando Maxent, threshold MTC. ...................................................... 10
Figura 5: MDE utilizando GARP, threshold MTP. ........................................................ 11
Figura 6: MDE utilizando GARP, threshold MTC. ....................................................... 11
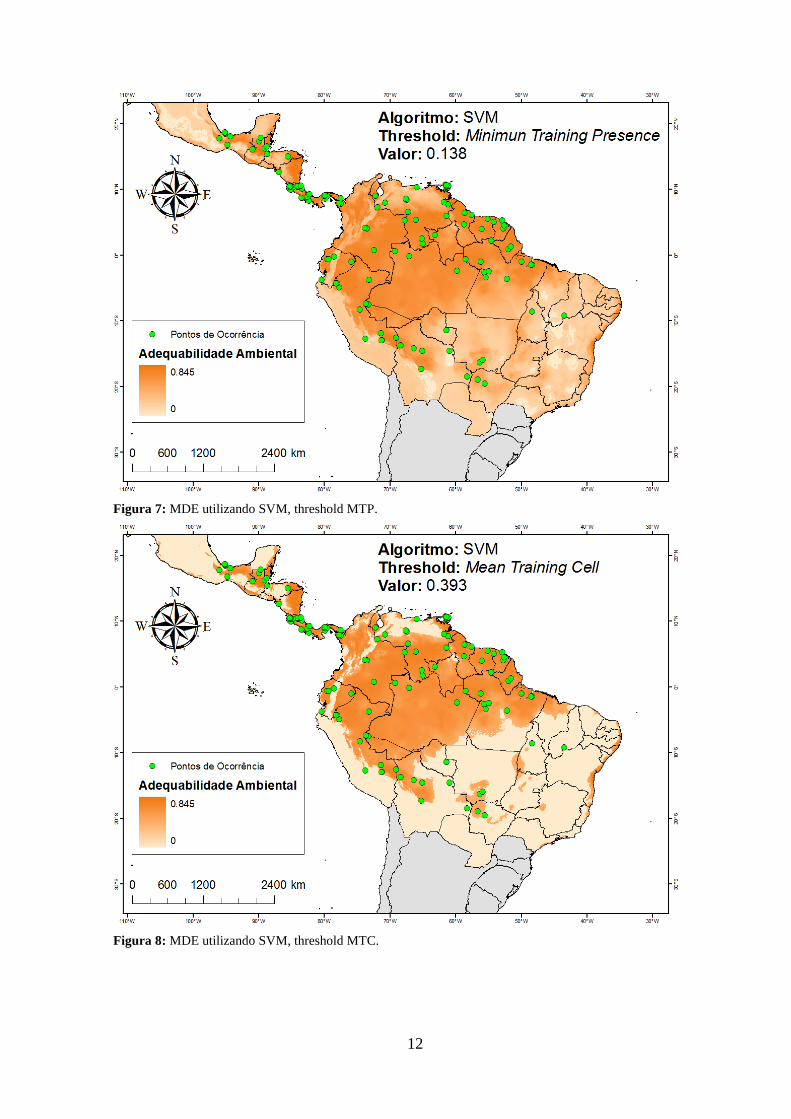
Figura 7: MDE utilizando SVM, threshold MTP. .......................................................... 12
Figura 8: MDE utilizando SVM, threshold MTC. ......................................................... 12
Figura 9: MDE utilizando ENFA, threshold MTP. ........................................................ 13
Figura 10: MDE utilizando ENFA, threshold MTC. ...................................................... 13
Figura 11: MDE utilizando RF, threshold MTP. ............................................................ 14
Figura 12: MDE utilizando RF, threshold MTC. ........................................................... 14
Figura 13: MDE combinando todos os algoritmos, com threshold MTP. ...................... 15
Figura 14: MDE combinando todos os algoritmos, com threshold MTC. ..................... 15
Figura 15: MDE combinando os algoritmos com TSS > 0.5, e p-valor no teste binomial
< 0.05. .......................................................................................................... 16
Figura 16: Similaridade dos modelos de acordo com os valores dos dois primeiros
Componentes Principais. ............................................................................. 17
Figura 17: Extensão de ocorrência de Vampyrum spectrum, de acordo com Navarro &
Wilson (1982). ............................................................................................. 18
Figura 18: Extensão de ocorrência de Vampyrum spectrum, de acordo com Williams &
Genoways (2007). ........................................................................................ 19
Figura 19: Hiperplano e vetores suporte para SVM. Obtido de Tarca et al., 2007. ....... 23
Figura 20: Esquematização da Marginalidade de uma espécie (δS), em relação a Área de
Estudo (δG). Obtido de Hirzel et al., 2002. .................................................. 24
Figura 21: Áreas Prioritárias para Inventários de Vampyrum spectrum no Brasil. ........ 31

XI
LISTA DE TABELAS
Tabela 1: Valores de Area Under Curve (AUC), teste binomial e parâmetros da matriz
de confusão, relativos aos algoritmos e três projeções combinadas
(Ensembles). .................................................................................................. 9
Tabela 2: Áreas Protegidas no Brasil em áreas de adequabilidade ambiental alta.
Valores extraídos do Ensemble II (para explicação, ver texto). ESEC =
Estação Ecológica, PARNA = Parque Nacional, PARNES = Parque
Estadual, REEXT = Reserva Extrativista, REBIO = Reserva Biológica,
REFNA = Reserva Florestal, RPPN = Reserva Particular do Patrimônio
Nacional. ...................................................................................................... 29

1
1 INTRODUÇÃO
Vampyrum spectrum (Linnaeus, 1758) é o único representante do gênero
monotípico Vampyrum Rafinesque, 1815. É o maior dos morcegos do Ocidente e da
subordem Vespertilioniformes (Antebraço: 96 – 114 mm, Comprimento do crânio: 49 –
54 mm). Distingue-se também dos demais Phyllostomidae por não possuir cauda,
característica que compartilha com Chrotopterus (Williams & Genoways, 2007). É um
predador de topo de cadeia, se alimentando principalmente de pequenos roedores,
pássaros e pequenos morcegos, complementando a dieta com insetos e ainda frutas,
numa frequência reduzida (Bonato et al., 2004; Nogueira et al., 2007). Apesar de
qualitativamente esta dieta não diferir de outros morcegos de grande porte, McNab
(1971) e Fleming et al. (1972), através de uma “matriz de nicho” bidimensional,
concluíram que a espécie pertence a um nicho distinto. Bonato et al. (2004) demonstram
a mesma conclusão através de Análise de Componentes Principais (PCA), comparando
três espécies de morcegos carnívoros de porte médio à grande.
Usualmente, morcegos neotropicais não seguem a associação inversamente
proporcional entre massa corporal e abundância encontrada em outros mamíferos (e.g.
Damuth, 1981; Robinson & Redford, 1986). Isto, porém, não se aplica aos grandes
morcegos carnívoros: quanto maior sua massa corporal, mais raro eles se tornam (Arita,
1993). A espécie é considerada Quase Ameaçada pela Lista Vermelha da IUCN de
2008, com tendência para redução populacional no futuro, devido à sua dependência de
florestas primárias, raridade e baixa densidade populacional (Aguirre et al., 2008).
Apesar disso, também é encontrada em campos, pântanos e bordas de florestas tropicais
(Handley, 1976). Apesar desse conhecimento, a literatura científica é escassa em termos
de mapeamento biogeográfico para a espécie, com a possível exceção de levantamentos
locais (e.g. Sousa et al., 2011, para o Brasil; Acosta & Azurduy, 2006, para a Bolívia).
Neste contexto, o conhecimento da distribuição das espécies e seus padrões são
imprescindíveis para o monitoramento da biodiversidade. Porém, tal conhecimento é
limitado, principalmente nas regiões tropicais (Anderson et al., 2003). O uso de
algoritmos ecológicos pode auxiliar nessa questão, lidando com o conceito de nicho
ecológico (De Marco Jr. & Siqueira, 2009). Tais algoritmos fazem parte do campo de
“Machine Learning”, termo referente a uma série de tópicos lidando com a criação e
validação de algoritmos que realizam reconhecimento de padrões, classificação e
predição, a partir de dados existentes (Tarca et al., 2007). Os primeiros métodos para a

2
Modelagem Ecológica precisavam de dados de presença e ausência da espécie em
localidades amostradas, tornando a amostragem de Presença/Ausência crucial para todo
o procedimento. Entretanto, dados de ausência são muito difíceis de obter precisamente.
Uma localidade pode receber o rótulo de “ausente” quando: (1) a espécie não pôde ser
detectada ainda que presente (a cobra Coronella austriaca precisa de 34 amostragens
sem sucesso de captura em um determinado local para assumir, com intervalo de 95%
de confiança, que é ausente (Kéry, 2000 apud Hirzel et al., 2002)); (2) a espécie está
ausente por razões históricas, mesmo que o local seja adequado para sua ocorrência (e.g.
barreiras geográficas, extinções locais); (3) o local é verdadeiramente não adequado
para a espécie. Somente a última causa é relevante para as análises, e falsas ausências
(casos 1 e 2) exercem influência considerável (Hirzel et al. 2002). A Análise Fatorial de
Nicho Ecológico (ENFA), proposto por Hirzel e colaboradores (2002), foi um dos
primeiros algoritmos voltados à Modelagem Ecológica a eliminarem o requisito de
dados de Ausência, trabalhando somente com dados de Presença.
Esse desenvolvimento permitiu o uso de dados de coleções e museus, em sua
essência dados de Presença, e onde a categorização de Ausência é praticamente
impossível. Desde então, os métodos mais populares na Modelagem Ecológica
trabalham apenas com dados de Presença. Também torna possível trabalhar com
espécies cuja ausência pode ser difícil de comprovar mesmo em condições de campo,
por raridade, baixa densidade populacional, sucesso de amostragem e captura baixo,
e/ou outros fatores.
Essas características tornam os morcegos candidatos ideais para a aplicação da
Modelagem Ecológica. Entretanto, os estudos de modelagem de morcegos são de
número reduzido (mas veja Yom-Tov & Kadmon, 1998; Henry et al., 2010). Mantilla-
Meluk et al. (2009) conduziram um estudo incluindo Vampyrum spectrum, mas de
abrangência limitada à Colômbia.
Neste estudo, conduzi uma revisão da literatura para determinar a distribuição
atual de Vampyrum spectrum e verificar as variáveis ecogeográficas mais importantes
para sua distribuição. Além disso, utilizei cinco algoritmos somente-Presença de
Modelagem Ecológica para: (1) Determinar sua distribuição potencial, além de uma
combinação dos modelos mais eficientes; (2) Indicar as Áreas Protegidas, no Brasil,
com grandes probabilidades de ocorrência onde ela ainda não foi amostrada.

3
2 Material & Métodos
2.1 Banco de Dados
2.1.1 Dados de Ocorrência Geográfica
Os pontos de ocorrência foram coletados através da pesquisa nos indexadores
Global Biodiversity Information Facility (GBIF; www.gbif.com), Mammal Networked
Information System (MaNIS; http://manisnet.org), na literatura científica, e através de
consultas em coleções de museus na Alemanha, Brasil, Canadá, Estados Unidos e
Inglaterra pelo Prof. Dr. Albert David Ditchfield (Anexos 1, 2 e 3).
Para obter a extensão de ocorrência de Vampyrum spectrum, foi utilizado o
algoritmo alpha shape (Edelsbrunner et al., 1983; Akkiraju et al., 1995), por meio do
pacote alphahull (Pateiro-Lopez & Rodriguez-Casal, 2011), disponível na plataforma R
(2013). O mapa gerado foi utilizado para comparação com os Modelos de Distribuição.
2.1.2 Dados climáticos e de relevo
Foram tomadas as 19 variáveis bioclimáticas disponíveis no WorldClim (Hijmans
et al., 2005), compostas por camadas ambientais de temperatura, precipitação e uma
combinação destas, com extremos máximos e mínimos, sazonalidade e médias anuais,
mensais e diárias. Também foram tomadas duas variáveis de relevo: Inclinação do
terreno e Modelo de Elevação Digital, no U.S. Geological Survey (USGS, 2001) (Anexo
4).
Todas as variáveis continham abrangência global, com resolução de 0,0417º (i.e.
cada pixel tem aproximadamente 5 km de aresta). As camadas foram então restringidas
à área de interesse para o estudo, incluindo México, América Central e América do Sul
(exceto a região sul do Brasil, Uruguai, Chile e Argentina).
2.2 Modelagem Ecológica
2.2.1 Estatística Pré-Modelagem
Os valores ambientais de cada uma das camadas ambientais foram extraídos por
meio da plataforma R (2013), utilizando os pacotes maptools (Lewin-Koh et al., 2012) e
SDMTools (VanDerWal et al., 2012). Os dados obtidos foram então submetidos a uma
Análise de Componentes Principais (PCA), de forma a reduzir o número de camadas
ambientais, eliminando aquelas correlacionadas. Em seguida, foi feita uma matriz de
distância das coordenadas geográficas e das variáveis ambientais para o teste de Mantel,

4
visando verificar quais camadas possuem influência significativa para a distribuição da
espécie.
2.2.2 Parâmetros dos Algoritmos
MAXENT: O algoritmo de Máxima Entropia prediz a probabilidade de
ocorrência das espécies encontrando a distribuição de máxima entropia, sujeita a uma
série de restrições que representam o conhecimento incompleto da distribuição do grupo
(Phillips et al., 2006).
Foram escolhidos os seguintes parâmetros para o estudo: Cinco iterações de 5.000
repetições cada, das quais foi computada uma média. O tipo de replicação escolhido foi
o Crossvalidate.
GARP: Acrônimo para Genetic Algorithm for Rule Set Production, o GARP
trabalha criando um conjunto de regras de inferência lógica que indicam a presença ou
ausência de uma espécie em uma região. Essas regras são selecionadas a partir de um
processo iterativo de seleção, validação e teste (Stockwell & Noble, 1992; Anderson et
al., 2003). Atualmente, existem diversas versões do GARP disponíveis. Foi utilizado o
“GARP with Best Subsets – New openModeller Implementation”, versão 3.0.4,
disponível no openModeller 1.3.0 (Muñoz et al., 2009).
Foram escolhidos os seguintes parâmetros: Cinco baterias com 200 modelos,
limite de convergência de 0.0001, erro de omissão extrínseco de 0%, 50% de erro de
comissão, e máximo de 400 iterações. Os resultados foram combinados em um
ensemble do GARP.
SVM: Os Support Vector Machines foram desenvolvidos por Cortes & Vapnik
(1995), constituindo uma nova metodologia para algoritmos “Machine Learning”. Os
SVM, a partir de um conjunto inicial de dados, predizem para cada input uma de duas
classes possíveis, formando o output (especificamente, são classificadores binários não
probabilísticos). A decisão de classificação é tomada buscando o hiperplano que
maximiza as margens de distância entre as duas classes de dados (Guo et al., 2005). Em
sua implementação original, os SVM são lineares, mas podem ser transformados em
SVM não lineares aplicando uma função kernel não linear, modificando o Espaço
Original em um Espaço Projetado. Dados que não podiam ser separados linearmente no
Espaço Original podem então ser separados linearmente no Espaço Projetado. Uma das
funções kernel mais utilizadas é a “radial basis function” (RBF). Desse modo, o

5
hiperplano pode ser traçado no Espaço Projetado para dados que não poderiam então ser
separados linearmente no Espaço Original (Tarca et al., 2007).
O algoritmo está disponível no openModeller 1.3.0 na versão 0.4. Foram
escolhidos os seguintes parâmetros: SVM de duas classes, Kernel RBF, Gamma 1/k
(onde k é igual ao número de camadas ambientais utilizadas para treinar o algoritmo),
Número de Pseudo Ausências igual ao número de presenças. Foram feitas cinco baterias
de testes, posteriormente combinados em um ensemble do SVM.
ENFA: A Análise Fatorial de Nicho Ecológico (de “Ecological-Niche Factor
Analysis”) é um algoritmo desenvolvido por Hirzel et al. (2002), modificando a Análise
de Componentes Principais (PCA) para lidar com o problema de ausência. O método
parte do princípio de que as espécies estão distribuídas de forma não aleatória devido as
variáveis ecogeográficas, i.e., uma espécie ocorre preferencialmente na faixa ótima de
ocorrência das células amostradas, em vez de distribuídas por toda sua extensão. Logo,
os valores da média e variância dos dados observados e do conjunto total podem diferir.
O algoritmo originalmente era disponibilizado no software Biomapper, mas está
descontinuado. Foi utilizada a versão 0.1.3, disponível no openModeller 1.3.0, com os
seguintes parâmetros: 10.000 pontos de background, cinco tentativas para criar o
modelo, método de descarte de componentes foi “broken stick method”. Foram feitas
cinco baterias de testes, posteriormente foram combinados em um ensemble do ENFA.
RANDOM FORESTS: O último algoritmo analisado neste trabalho, o Random
Forests (RF) faz parte dos métodos baseados em árvores (Breiman, 2001). O princípio
desses métodos consiste em particionar o conjunto de dados em dois subconjuntos
baseado na divisão ótima dentre todas as divisões possíveis de acordo com algum
critério definido a priori (Knudby et al., 2010).
O RF está disponível na versão 0.2 no openModeller 1.3.0. Foram escolhidos os
seguintes parâmetros: 300 Árvores, Número de variáveis por árvore igual a √k (onde k é
igual ao número de camadas utilizadas para treinar o algoritmo). Foram feitas cinco
baterias de testes, posteriormente combinados em um ensemble do RF.
2.3 Validação
2.3.1 Threshold
Foram escolhidos dois limites para a análise dos modelos, relativos aos valores de
presença no treino destes.

6
O “Minimum Training Presence” (a terminologia é do Maxent 3.3.3e. No
openModeller 1.3.0, o “Lowest Prediction” é equivalente) que representa o menor valor
de adequabilidade ambiental em um ponto de ocorrência; e o “Mean Training Cell”
(MTC), tomado da média dos valores de adequabilidade de cada célula do modelo.
2.3.2 Pontos aleatórios
Foram gerados aleatoriamente pontos de presença aleatórios em ambiente SIG, na
proporção 10:1 com os pontos de ocorrência reais, para alimentar a matriz de confusão
(ver embaixo).
2.3.3 Matriz de confusão
A matriz de confusão é uma tabela que permite a visualização do desempenho de
um algoritmo, especificamente se o algoritmo está confundindo as duas classes, daí seu
nome (Figura 1).
“A” corresponde ao número
de verdadeiros positivos, pontos de
ocorrência reais que foram
previstos pelo modelo, enquanto
“D” representam o número de
verdadeiros negativos, pontos
gerados aleatoriamente que foram
corretamente aferidos como
ausências pelo modelo. Os erros de
comissão e omissão correspondem
a “B” e “C” na matriz,
respectivamente (ver seção 4.2.1.
Validação dos Modelos).
A partir desses valores, podemos calcular a: Sensibilidade, uma medida de quanto
o modelo acertou corretamente os pontos verdadeiros; Especificidade, o quanto acertou
corretamente os pontos aleatórios; Acurácia, o quanto acertou corretamente os pontos
verdadeiros e aleatórios; e TSS (“True Skill Statistic”), uma medida alternativa à
acurácia que é independente da prevalência (Allouche et al., 2006).
Figura 1: Matriz de confusão

7
Também foi realizado o teste binomial de proporções, utilizando os valores de
Verdadeiros Positivos (“A”) e de todos os registros (126), em relação à área prevista
como favorável pelo modelo e a área total utilizada.
2.4 Projeção Combinada
É uma estratégia para aumentar a confiabilidade dos modelos gerados,
combinando diversos algoritmos. É conhecida como “ensemble forecasting” na
literatura anglófona. Trata-se da geração de um modelo de consenso que indica quais
áreas de adequabilidade ambiental alta foram indicadas para a espécie.
Foram gerados três ensembles neste trabalho: (I) com os modelos gerados por
meio do threshold MTP; (II) com os modelos gerados por meio do threshold MTC; e
(III) com os modelos que obtiveram de p-valor no teste binomial abaixo de 0,05, e TSS
superior a 0,5.
Os mesmos procedimentos estatísticos de validação foram utilizados para avaliar
os ensembles, com o threshold sendo computado via média aritmética dos thresholds
dos modelos utilizados para a combinação.
2.5 Similaridade
Para calcular a similaridade entre os modelos gerados neste estudo, foi feita uma
PCA com os vetores de adequabilidade de cada um. A PCA foi feita por meio do pacote
FactoMineR (Husson et al., 2013), com auxílio de funções gráficas do pacote Maptools.
2.6 Mapas de Distribuição Ambiental
Para cada algoritmo (com threshold MTP e MTC), e as três projeções
combinadas, foi feito um mapa de distribuição potencial, levando em conta a
adequabilidade ambiental, em escala contínua; a divisão geopolítica dos estados
brasileiros e os pontos de ocorrência de Vampyrum spectrum levantados para este
estudo, refletindo sua distribuição atual.
Os shapes de Biomas foram obtidos por meio do IBGE (www.ibge.gov.br). Os
shapes de Áreas Protegidas foram obtidos no GISMAPS (www.gismaps.com.br).
Todos os mapas foram confeccionados em ambiente SIG.

8
3 RESULTADOS
3.1 Distribuição Atual
Foram coletados 145 pontos de ocorrência georreferenciados para Vampyrum
spectrum. Destes, 126 são pontos únicos geograficamente (i.e. não há outro com latitude
e longitude igual), distribuídos em 17 países: Brasil (25); Venezuela (16); Costa Rica,
Peru e Trinidad & Tobago (12); Panamá (10); México (9); Bolívia (6); Guiana Francesa
(5); Equador (4); Guiana, Colômbia e Suriname (3); Belize e Guatemala (2); Honduras e
Nicarágua (1). A distribuição esperada pelo α-shape está na Figura 2.
Figura 2: Alpha Shape de extensão de ocorrência de Vampyrum spectrum.
As referências, coordenadas geográficas e instituições dos pontos utilizados pode
ser conferida nos Anexos 1, 2 e 3.
3.2 Distribuição Potencial
3.2.1 Variáveis Escolhidas
Foram escolhidas oito variáveis ecogeográficas para confeccionar os modelos de
distribuição potencial, quatro relativas à temperatura: BIO4 (Sazonalidade da
Temperatura), BIO6 (Temperatura mínima do mês mais frio), BIO7 (Variação anual da
temperatura), BIO11 (Temperatura média no trimestre mais frio); e quatro relativas à
precipitação: BIO12 (Precipitação anual), BIO13 (Precipitação no mês mais úmido),
BIO16 (Precipitação no mês mais úmido), BIO18 (Precipitação no trimestre mais frio).

9
3.2.2 Modelos de Distribuição de Espécies
No total, foram gerados 13 modelos de distribuição potencial para Vampyrum
spectrum, dois para cada algoritmo, além de três projeções combinadas. Exceto o SVM
(MTP), todos os modelos diferem do acaso pelo teste binomial. Todos os modelos
apresentaram valores de MTP menores que MTC, exceto o Random Forests (ver Tabela
1).
Tabela 1: Valores de Area Under Curve (AUC), teste binomial e parâmetros da matriz de confusão,
relativos aos algoritmos e três projeções combinadas (Ensembles).
Threshold
Algoritmo AUC Tipo Valor Sensibilidade Especificidade Acurácia TSS Binomial
Maxent 0.787 ± 0.044 MTP 0.061 0.954 0.716 0.738 0.669 < 0.0001
MTC 0.211 0.913 0.429 0.465 0.332 < 0.0001
GARP 0.712 ± 0.015 MTP 0.180 0.968 0.723 0.745 0.691 < 0.0001
MTC 0.603 0.913 0.591 0.620 0.504 < 0.0001
ENFA 0.642 ± 0.004 MTP 0.010 0.968 0.819 0.833 0.787 < 0.0001
MTC 0.349 0.651 0.408 0.430 0.059 < 0.0001
RF 0.870 ± 0.014 MTP 0.734 0.881 0.591 0.618 0.472 < 0.0001
MTC 0.729 0.897 0.604 0.630 0.630 < 0.0001
SVM 0.796 ± 0.009 MTP 0.138 0.984 0.966 0.967 0.950 0.091
MTC 0.393 0.897 0.455 0.495 0.351 < 0.0001
Ensemble I 0.761 ± 0.087 - 0.225 0.976 0.686 0.712 0.662 < 0.0001
Ensemble II 0.761 ± 0.087 - 0.457 0.873 0.353 0.400 0.226 < 0.0001
Ensemble III 0.744 ± 0.087 - 0.317 0.936 0.591 0.623 0.528 < 0.0001
Para o Ensemble III, foram utilizados os seguintes modelos: Maxent (MTP),
GARP (MTP e MTC), ENFA (MTP) e RF (MTC). Os mapas de distribuição potencial
para cada um dos modelos podem ser visualizados nas Figuras 3 a 15.
Uma vez que, quanto menor o threshold utilizado, maior a área prevista como
favorável, os modelos apresentaram redução de área favorável com a mudança de
threshold (exceto o Random Forests). Para Maxent, GARP, SVM e ENFA, as reduções
de área foram respectivamente: 39,9%, 19,1%, 52,8% e 47,9%. O Random Forests
apresentou aumento de 2%.

10
Figura 3: MDE utilizando Maxent, threshold MTP.
Figura 4: MDE utilizando Maxent, threshold MTC.

11
Figura 5: MDE utilizando GARP, threshold MTP.
Figura 6: MDE utilizando GARP, threshold MTC.
12
Figura 7: MDE utilizando SVM, threshold MTP.
Figura 8: MDE utilizando SVM, threshold MTC.

13
Figura 9: MDE utilizando ENFA, threshold MTP.
Figura 10: MDE utilizando ENFA, threshold MTC.

14
Figura 11: MDE utilizando RF, threshold MTP.
Figura 12: MDE utilizando RF, threshold MTC.

15
Figura 13: MDE combinando todos os algoritmos, com threshold MTP.
Figura 14: MDE combinando todos os algoritmos, com threshold MTC.

16
Figura 15: MDE combinando os algoritmos com TSS > 0.5, e p-valor no teste binomial < 0.05.
A PCA de similaridade dos modelos apresenta uma primeira componente
responsável por 67,4% dos dados, enquanto a segunda componente explica mais
16,12%. ENFA, Maxent e SVM apresentam padrões semelhantes de similaridade.
GARP mostra padrões mais dissimilares, enquanto o Random Forests mostra
comportamento bem distinto dos demais. As combinações I e III possuem padrões
similares aos modelos ENFA, Maxent e SVM, enquanto a combinação II apresenta
padrões intermediários entre o Random Forests e os demais modelos (Figura 16).

17
Figura 16: Similaridade dos modelos de acordo com os valores dos dois primeiros Componentes
Principais.

18
4 DISCUSSÃO
4.1 Distribuição Atual
Os dados obtidos neste estudo mostram que Vampyrum spectrum encontra-se
distribuído por quase toda a América Central e boa parte do continente sul-americano,
desde o sul do México até o Centro-Oeste brasileiro e centro da Bolívia. No Brasil,
ocorre em praticamente todos os estados da região Norte (exceto Maranhão), na região
Centro-Oeste e no Nordeste (Figura 2). Os dados permitem ampliar a distribuição da
espécie até a latitude 19,5º S, superior ao limite estabelecido por Navarro & Wilson
(1982) de 14º S (Figura 17).
Para o Brasil, Sousa et
al. (2011) relataram a primeira
ocorrência para a espécie no
bioma Cerrado, no estado do
Mato Grosso. Os autores
também fazem uma revisão da
distribuição no Brasil, mas
nem todos os dados podem ser
confirmados mediante consulta
na literatura científica. Em
específico, os autores citam
uma ocorrência no Acre
(Peracchi et al., 2010) que não especifica as coordenadas geográficas (nem fornece um
mapa para atribuição, mesmo que de forma aproximada), e outra no Roraima (Taddei &
Reis, 1980), onde não consta menção à Vampyrum spectrum. Desta forma, a localidade
nova relatada pelos autores, no município de Nova Xavantina (14º 38’ 19’’ S, 52º 21’
39’’ W) não foi utilizada, assim como uma comunicação pessoal de Guimarães para o
estado do Tocantins, citada no mesmo artigo. A eliminação desses dados não prejudicou
os modelos, uma vez que as áreas de ocorrências dos mesmos são favoráveis em todos
os modelos, mesmo naqueles mais restritivos.
Vampyrum spectrum mostra uma distribuição concentrada principalmente na
Floresta Amazônica. Porém, é também reportada a ocorrência no Pantanal e Cerrados
brasileiros, assim como a Caatinga. Vieira (1955) registrou a primeira ocorrência no
Pantanal, aproximadamente 50 anos de intervalo entre a ocorrência seguinte, reportada
Figura 17: Extensão de ocorrência de Vampyrum spectrum, de
acordo com Navarro & Wilson (1982).

19
por Silveira e colaboradores em 2011. Os resultados indicam que a espécie possui
distribuição superior à prevista tradicionalmente na literatura. Aliada à baixa densidade
da espécie e raridade nos inventários, isso sugere um grau de vício amostral,
corroborado pelos dados de Bernard et al. (2010), que mostram uma predominância na
faixa de 0º a 5º S nos estados do Pará e Amazonas.
O método do alpha shape, utilizado para prever a extensão de distribuição de
Vampyrum spectrum, cria uma função para explicar os dados de ocorrência, ajustando
um envelope de distribuição em volta dos registros. O procedimento descreve os dados
de maneira mais satisfatória que outras metodologias utilizadas na literatura, como o
Polígono Mínimo Convexo (Hirsch & Chiarello, 2011). O parâmetro crítico é o valor de
alfa: à medida que alfa aumenta, o shape tende a ter a mesma área que o “Convex Hull”
(definido como o menor polígono envoltório em que nenhum ângulo interno excede
180o); em contraste, quando alfa tende a zero, cada ponto tende a se tornar um polígono
isolado, e a área do alpha shape tende à zero também. A escolha do valor de alfa, então,
possui um fator de subjetividade.
Comparando a
extensão esperada pelo
alpha shape com outras
disponíveis na literatura, é
visível a redução da área da
espécie. As diferenças se
devem principalmente as
diferença das metodologias
utilizadas. A extensão
fornecida por Navarro &
Wilson (1982) é baseada na
longitude máxima à oeste
(no México) e na latitude no extremo sul (Figura 17), enquanto a extensão de Williams
& Genoways (2007) é baseada nas localidades marginais (Figura 18).
Figura 18: Extensão de ocorrência de Vampyrum spectrum, de
acordo com Williams & Genoways (2007).

20
4.2 Modelagem Ecológica
4.2.1 Validação dos Modelos
DEFINIÇÃO: A validação de um modelo, também conhecida como “teste
externo”, consiste na confrontação dos resultados obtidos pelo algoritmo com aqueles
que não foram utilizados para a produção do modelo. Sua contraparte, a verificação (ou
“teste interno”), utiliza geralmente todos os dados tanto para o treino do modelo, quanto
para o teste, enquanto na validação os dados do teste nunca são conhecidos pelo modelo
(Giannini et al., 2012). A validação não corresponde a uma verdade absoluta em termos
de avaliação de modelos, para a Modelagem Ecológica. De fato, é uma área ainda em
evolução, com metodologias estatísticas não totalmente resolvidas na literatura
(Peterson et al., 2008; Jiménez-Valverde, 2012), e diferentes formas foram propostas
(e.g., Pearce & Ferrier, 2000; Lobo et al., 2008), mas todas seguindo o mesmo
princípio: para avaliar o modelo, é preciso quantificar os erros que podem acontecer no
processo, a saber, os erros de omissão e comissão.
Erro De Omissão: O erro de omissão, também chamado de “Erro de
Subprevisão”, representa a fração de pontos de presença reais, usados para o treino do
modelo, mas que por algum motivo estão fora da área de adequabilidade ambiental
prevista pelo algoritmo. O motivo pode ser um erro verdadeiro, como classificação
taxonômica errônea e/ou erro de georeferenciamento. Mas também pode representar um
outlier para o algoritmo, um ponto com informação ambiental muito fora do padrão dos
demais pontos utilizados para treinar o modelo. De fato, quando há incerteza dos dados,
é preferível deixar uma margem de segurança (algo entre 5 a 10% dos dados) ao redor
da omissão para que o algoritmo possa trabalhar melhor, do contrário uma área de maior
proporção deverá ser prevista para acondicionar esses pontos (Phillips et al., 2006;
Giannini et al., 2012).
Erro De Comissão: Ou “Erro de Sobreprevisão”. O erro de comissão é o caso
específico de não saber determinar corretamente uma ausência, discutido por Hirzel et
al. (2002). Consiste na previsão de ocorrência da espécie em uma localidade onde não
há registro. Pode ser dividido em erro de comissão aparente (quando a ausência na
localidade é por falta de esforço amostral, ou ela não ocorre no momento devido à
fatores históricos); e erro de comissão verdadeiro (quando o local é realmente
inabitável).

21
MÉTODO: A validação exige que confrontemos os dados utilizados para treinar
o modelo com um conjunto independente de dados. Idealmente, tal conjunto se daria
com a coleta de novos dados em campo, ou levantamento na literatura. Entretanto, esta
abordagem nem sempre é possível.
Uma alternativa é a divisão aleatória entre dados de treino e teste dos pontos de
ocorrência. Porém, a eficiência deste método nem sempre é eficaz, pois na verificação o
conjunto inicial de dados é dividido várias vezes, em várias iterações. Logo, todos os
dados são utilizados para a verificação, o que fere a premissa da novidade dos dados de
teste para a validação.
Outra opção, mais eficaz, é a geração de pontos aleatórios. Teoricamente,
qualquer gerador de números aleatórios pode atuar nesse quesito, mas existem
ferramentas especializadas para tal, e.g. a função “OM_PSEUDO” no openModeller
1.3.0, ou criação em ambiente SIG.
THRESHOLD: As análises mais tradicionais da capacidade discriminatória de
um modelo dependem da adoção de um limite de corte arbitrário (o threshold). No
estado da arte da Modelagem Ecológica a escolha do threshold ainda é arbitrária, mas
um subconjunto de opções pode ser delimitado do total. Liu et al. (2005) é a melhor
revisão do assunto na literatura.
A finalidade da Modelagem Ecológica é um bom norteador para a escolha do
threshold. Uma vez que Vampyrum spectrum é uma espécie pouco coletada, um
threshold mais baixo (que aumenta o erro de sobreprevisão) é desejável, pois aumenta a
área de probabilidade de ocorrência da espécie. O menor valor de corte arbitrário
relativo aos dados do modelo de distribuição de espécies é o “Minimum Training
Presence”.
O segundo threshold utilizado neste estudo, o “Mean Training Cell”, foi calculado
a partir da média de adequabilidade ambiental de todas as células com valores
informativos (i.e. retirando os valores “NO DATA”) de cada modelo, em contraste com
o “Minimum Training Presence” que leva em conta os valores apenas dos pontos de
ocorrência. Procedeu-se dessa forma devido ao desempenho péssimo dos modelos
quando a média era calculada dos pontos de ocorrência. Um teste com o Maxent gerou
valores de TSS de -0,410. O TSS utilizando todas as células foi de 0,332.

22
A diferença entre usar valores das células e valores dos pontos também gerou uma
peculiaridade que aparenta ser um erro superficialmente: o threshold mínimo foi
superior ao threshold médio para o Random Forests.
AUC: Sigla para “Area Under Curve”, a AUC é a área sobre a curva ROC (de
“receiver operating characteristic”). A curva ROC é calculada através de funções
derivadas de parâmetros da matriz de confusão, plotando a Sensibilidade e
Especificidade do modelo à medida que o threshold se altera. A área sobre a curva
fornece um número que tradicionalmente é utilizado para estimar a acurácia dos
modelos.
Em um estudo investigativo, Lobo et al. (2008) demonstraram que a AUC é
potencialmente enganosa, e não constitui um parâmetro confiável para avaliar e
comparar os modelos. Neste trabalho, os valores de AUC para os modelos foram
fornecidos para fins descritivos apenas, e não foram utilizados como critérios de
desempenho para os modelos.
4.2.2 Algoritmos: Definições, Operação e Estado da Arte
MAXENT: O Maxent é um algoritmo que trabalho com o Princípio de Máxima
Entropia, ligado à Teoria da Informação. De forma sucinta, o algoritmo busca a
distribuição de probabilidades de ocorrência, dado um conjunto inicial de dados, que
maximiza a função de máxima entropia. Tal função é a distribuição de dados mais
adequada para descrever os dados iniciais, segundo tal princípio. Mas, uma vez que os
dados iniciais não representam o conhecimento total do grupo (i.e., ele é incompleto), o
algoritmo é sujeito a uma série de restrições que representam essa lacuna (Phillips et al.,
2006). Ainda que o Princípio seja conhecido a muito tempo, sua aplicação na Ecologia
(e Modelagem Ecológica) é recente e de ascensão rápida. Elith et al. (2006), em um
estudo comparativo com outros métodos, demonstrou que o Maxent é superior a todos
eles, o que explica a prevalência do método na literatura.
Entretanto, essas asserções foram desafiadas com o tempo, variando de situações
onde outros algoritmos superam o Maxent (e.g. Terribile et al., 2010), ou mesmo
demonstrando a equivalência do algoritmo com os GLM (de “Generalized Linear
Models”, ou Modelos Lineares Generalizados), metodologia mais antiga (Renner &
Warton, 2013). O último é notável devido à colocação no artigo de apresentação do
Maxent, ao citar um de seus pontos fracos: “It is not as mature a statistical method as
GLM or GAM” (Phillips et al., 2006).

23
Mesmo sofrendo golpes na sua constituição original, o Maxent continua sendo
uma ferramenta poderosa para elucidar padrões de diversidade e distribuição.
GARP: O GARP é também um algoritmo da família “Machine Learning”,
tradicionalmente aplicado em situações onde a busca exaustiva por todas as
possibilidades, dado um conjunto inicial de dados, é muito extensa para ser realizada
num espaço temporal razoável (Stockwell & Noble, 1992). É utilizado na Modelagem
Ecológica com resultados favoráveis em uma gama ampla de situações, como potencial
de espécies invasoras, localização de áreas de ocorrência favoráveis para direcionar
esforços de coleta, elucidar padrões de distribuição, entre outros (Anderson et al.,
2003).
Várias implementações do GARP estão disponíveis atualmente, variando em
detalhes técnicos de seu funcionamento, principalmente no que diz respeito à criação e
manipulação de regras de inferência lógica para explicar a presença ou ausência das
espécies no local analisado. A mais recente encontra-se disponível no openModeller
1.3.0, que até o momento de escrita não possui GUI (i.e. é operada por linhas de
comando no prompt do sistema operacional).
SVM: Os Support Vector
Machines são classificadores
binários não probabilísticos,
dividindo um conjunto inicial
de dados em duas classes
possíveis a partir da
maximização da distância entre
ambas. Para tal, o algoritmo
busca um hiperplano, uma
função intermediária entre os
vetores suporte, funções
marginais à distribuição dos
pontos das duas classes, que dão nome ao método (Figura 19) (Guo et al., 2005; Tarca
et al., 2007).
Atualmente, os SVM suportam funções não lineares para classificar o conjunto
inicial, mediante aplicação de funções kernel não lineares, modificando o Espaço
Original dos dados em um Espaço Projetado de dimensões infinitas. Desse modo, dados
Figura 19: Hiperplano e vetores suporte para SVM. Obtido de
Tarca et al., 2007.

24
que outrora não poderiam ser bem classificados podem agora serem corretamente
explicados pelo algoritmo. Existe uma diversidade de funções kernel para a projeção,
com destaque para as funções exponencial e Radial Basis Function, que estão
programadas na versão do SVM disponível no openModeller 1.3.0.
Estudos anteriores (e.g. Barreto, 2008) mostravam um desempenho geral pior para
o SVM em relação a outros algoritmos. Tal comportamento não foi visualizado neste
trabalho, pelo contrário: o SVM alcançou índices superiores a todos os outros modelos
de distribuição.
ENFA: A Análise Fatorial de Nicho Ecológico foi uma das primeiras técnicas a
lidar com o problema da Ausência na Modelagem Ecológica, i.e., a dificuldade em
aferir corretamente o rótulo de “Ausente” para uma dada espécie em uma dada
localidade. Para tal, Hirzel et al. (2002) criam novos conceitos para explicar a
distribuição das espécies, e modificam a Análise de Componentes Principais (PCA)
para explicar essa distribuição.
Os conceitos criados são os de “Marginalidade”: distância entre as médias dos
dados observados e do conjunto total, corrigidos pelo desvio padrão; e
“Especialização”, razão entre os desvios padrão do conjunto total e dos dados
observados da espécie (Figura 20).
Figura 20: Esquematização da Marginalidade de uma espécie (δS), em relação a Área de Estudo (δG).
Obtido de Hirzel et al., 2002.

25
Hutchinson (1957) define o nicho ecológico como o hipervolume de n-dimensões
ecológicas onde a espécie consegue manter populações viáveis. Os autores então unem
seu conceito ao de Hutchinson, equivalendo o “nicho ecológico” do último com o
subconjunto de células no espaço ecogeográfico onde a espécie provavelmente ocorre.
Esse nicho multivariado é quantificado pela Marginalidade e Especialização. Na PCA,
onde os eixos são escolhidos para maximizar a variância da distribuição. A ENFA, em
contraste, escolhe o primeiro eixo para maximizar a Marginalidade da espécie, e os
eixos seguintes para maximizar a Especialização (Hirzel et al., 2002).
O algoritmo originalmente era disponibilizado no software Biomapper, mas está
descontinuado. Há uma função “enfa” no pacote adehabitat para a plataforma R
(Calenge, 2007), e também uma versão codificada no openModeller 1.3.0.
RANDOM FORESTS: O Random Forests pertence a uma família de algoritmos
“Machine Learning” diferente dos demais utilizados neste trabalho: a dos métodos
baseados em árvores (Breiman, 2001). O funcionamento é similar ao dos métodos
evolutivos do GARP, no que diz respeito à definição de conjuntos de regras para
explicar o conjunto de dados inicial. O diferencial dos métodos baseados em árvore é
que o conjunto inicial dos dados é dividido em dois subconjuntos ótimos por algum
critério definido a priori (um subconjunto, escolhido ao acaso, das variáveis utilizadas)
(Knudby et al., 2010). O processo continua até que (1) o critério seja atingido; ou (2),
cada partição tenha apenas uma observação, gerando então uma Árvore. O processo
então é repetido até o número desejado de Árvores ser alcançado. Na etapa final, cada
uma das Árvores concede um ”voto” para o modelo, e aquele que receber mais votos
dentre todas as Árvores torna-se o Modelo Ecológico (Grossmann et al., 2010).
O algoritmo encontra-se disponível no pacote randomForest (Liaw & Wiener,
2002) para a plataforma R, e codificado no openModeller 1.3.0.
4.2.3 Modelos de Distribuição de Espécies (MDEs)
De forma geral, os MDEs apresentaram variabilidade e propriedades em comum
uns com os outros, exceto os derivados do Random Forests (Figura 16). Enquanto as
distâncias entre os MDEs oriundos dos thresholds MTP e MTC foram aproximadamente
às mesmas para cada algoritmo, o Random Forests não apresentou diferenças
significativas. As distâncias entre os MDEs de cada algoritmo foram variadas: ENFA,
Maxent e SVM ficaram mais próximos, imediatamente próximos do GARP, enquanto o
Random Forests se distanciou de forma acentuada de todos os demais. A discrepância

26
pode ser explicada pelos métodos diferentes que cada algoritmo utiliza ao lidar com o
conjunto inicial de dados, mas somente isto não esclarece completamente a
dissimilaridade do Random Forests com os demais algoritmos. É possível que a causa
não resida na estrutura dos algoritmos baseados em árvores (ou no Random Forests em
si), mas na implementação deste no openModeller 1.3.0. GARP, SVM e ENFA são os
algoritmos implementados no openModeller mais utilizados, particularmente o GARP,
com quatro implementações. Em contraste, o Random Forests está numa versão inicial,
ainda não muito testada, provavelmente necessitando de reparos em sua constituição.
Seja como for, os resultados deste trabalho evidenciam que a implementação atual do
Random Forests não é favorável para a Modelagem Ecológica.
Todos os MDEs apresentaram variações na área classificada como favorável com
a mudança do threshold (de 2 a 52,8%). Tal resultado é esperado devido às propriedades
intrínsecas da matriz de confusão: mantendo os demais valores constantes, thresholds
menores apresentam áreas favoráveis maiores. Um modo intuito pra compreensão do
threshold é pensar nele como o inverso da constante alfa (de rejeição ou não da hipótese
nula): valores reduzidos de alfa tornam os modelos mais restritivos, diminuindo a
probabilidade do modelo cometer erros verdadeiros, mas aumentando a frequência dos
falsos negativos. Da mesma forma, valores elevados de alfa relaxam os modelos,
diminuindo a taxa de falsos negativos, porém ao custo do aumento da frequência dos
falsos positivos. Os thresholds funcionam da mesma maneira, porém os valores
elevados restringem o modelo; e valores reduzidos, relaxam-no.
Exceto o Random Forests, todos os MDEs apresentaram aumento do valor do
threshold quando o MTP era trocado pelo MTC, resultando em áreas favoráveis maiores
e, consequentemente, em parâmetros de validação superiores para MDEs utilizando
threshold MTP (ver Tabela 1). Porém, um MDE confeccionado de um threshold deveras
reduzido terá potencialmente toda a sua área de estudo classificada como favorável. Se
isto é desejável ou não, depende do foco de estudo. Para este, que visa direcionar
esforços de coleta, modelos mais restritivos garantem maiores índices de adequabilidade
ambiental e, portanto, aumentam as probabilidades de ocorrência da espécie. Um MDE
com parâmetros de validação elevados, mas que apresenta toda sua área de estudo como
favorável, não cumpriria seu intuito de indicar áreas para coleta. Portanto, o Ensemble II
(Figura 14), média dos MDEs com threshold MTC, foi o utilizado para uma das

27
finalidades deste estudo: indicar áreas para esforços de coleta no Brasil (ver seção 4.2.5.
Áreas Prioritárias para Inventários).
4.2.4 Variáveis Ecogeográficas
As variáveis ecogeográficas, referidas na literatura especializada como
“Superfícies Climáticas” (ou “Dados Climáticos Espacialmente Interpolados dispostos
em Rede”), são utilizadas em diversas áreas, incluindo Ciências do Meio Ambiente,
Agrônomas e Biológicas. Neste trabalho, as camadas utilizadas são oriundas do estudo
de Hijmans et al. (2005), que combinaram dados climáticos de vários bancos de dados,
de modo a criar camadas de alcance global e resolução espacial fina.
Basicamente, as camadas relacionam dados de temperatura, precipitação e
umidade, com alcances mínimos, médios e máximos. Desta forma, são ferramentas
poderosas para a Modelagem Ecológica, mas precisam de um tratamento antes de ser
aplicadas.
Primeiro, o alcance das camadas deve ser restringido para o local de estudo. Os
algoritmos são sensíveis em maior ou menor grau ao tamanho do local avaliado em
relação à área de ocorrência da espécie. O ENFA, em particular, é especialmente
sensível, produzindo alterações notáveis com mudanças de escala no estudo, devido à
Marginalização e Especialização variar com a média e desvio padrão deste. A Validação
também é sensível a variações na área de estudo, especialmente com a metodologia de
criação de pontos aleatórios utilizada: com uma área cada vez maior, os locais abaixo do
ponto de corte arbitrário tornam-se mais numerosos, afetando a Sensibilidade, Acurácia
e TSS para baixo de modo artificial.
Em seguida, é preciso lidar com o problema da correlação entre as variáveis
ecogeográficas. Ainda que os dados do WorldClim sejam derivados de um grande
número de banco de dados, a forma em que foram confeccionados acaba trazendo uma
grande correlação entre as camadas. É desejável reduzir o número de variáveis no
estudo, de modo a evitar artefatos estatísticos. Os testes de PCA e Mantel auxiliam
nessa demanda.
PCA: A Análise de Componentes Principais é um procedimento matemático que,
através de uma série de transformações nos dados, transforma uma série de variáveis
correlacionadas em um conjunto de valores lineares não correlacionados, chamados de
Componentes Principais (Pearson, 1901). É similar a outras técnicas estatísticas (como

28
Análise de Fatores, Análise Canônica Discriminante), mas diferencia-se principalmente
devido ao fato de ser puramente descritiva (i.e. não testa hipóteses acerca da
distribuição dos dados).
Uma PCA gera tantas Componentes Principais quantas variáveis foram utilizadas
no conjunto inicial de dados. A Primeira Componente Principal é aquela que explica a
maior variância dos dados, e assim por diante, até toda a variância dos dados ser
explicada. Dessa forma, é possível eliminar o ruído de outras variáveis, escolhendo
aquelas que mais contribuem para cada Componente Principal.
TESTE DE MANTEL: O teste de Mantel é uma alternativa eficaz para lidar com
três problemas inerentes ao campo da Modelagem Ecológica: (1) as camadas
ecogeográficas possuem forte correlação, tornando a formulação de inferências causais
difícil, mesmo que a correlação com a distribuição da espécie seja demonstrada; (2) as
variáveis possuem um grau característico de autocorrelação, o que pode limitar sua
influência na distribuição da espécie em escalas de referência particulares; (3) a própria
distribuição da espécie exibe um grau de agregação (autocorrelação), uma vez que os
pontos não são independentes uns dos outros, mas dependentes devido a fatores como
dispersão.
O teste evita esses problemas ao trabalhar com matrizes de dissimilaridade,
calculando a correlação entre as células da matriz por meio de permutações com
alterações aleatórias. A hipótese nula é de que os valores de correlação de cada permuta
não irão variar de forma significativa, mesmo com alterações na estrutura das matrizes,
indicando que não há relação de causalidade. Neste caso, a pergunta é “amostras
próximas uma das outras possuem composição similar?”.
Portanto, submeter o conjunto de dados das variáveis ecogeográficas (extraídos
dos pontos de ocorrência) a uma PCA e testes de Mantel possibilita ver quais são as
variáveis com maior correlação com os pontos de ocorrência, logo quais são as
responsáveis pela distribuição da espécie.
As variáveis ecogeográficas que mais contribuíram para explicar a distribuição de
Vampyrum spectrum foram BIO4, BIO6, BIO7, BIO11, BIO12, BIO13, BIO16 e
BIO18. As quatro primeiras são relacionadas à temperatura: Variação anual (7),
Sazonalidade (4), Mínima no mês mais frio (6), e Mínima no trimestre mais frio (11).
Tradicionalmente, V. spectrum é associado a ambientes Neotropicais, em regiões

29
florestadas (Navarro & Wilson, 1982). As análises dão suporte a essa afirmação,
mostrando que temperaturas mais baixas limitam a distribuição da espécie. As demais
variáveis são relacionadas à precipitação: no mês mais úmido (13), trimestre mais
úmido (16), trimestre mais frio (18) e precipitação anual (11). Valores baixos para essas
variáveis são limitantes para a distribuição de Vampyrum spectrum, o que também dá
suporte para a afirmação de Navarro & Wilson (1982).
Entretanto, as análises não são limitadas para ambientes florestais. Os registros
recentes no Pantanal mato-grossense e Caatinga do Piauí mostram que a espécie
também ocorre em locais diversos de florestas em clímax.
4.2.5 Áreas Prioritárias para Inventários
No Brasil, as áreas com maior adequabilidade ambiental para Vampyrum
spectrum estão concentradas na região Norte do Brasil, principalmente na Amazônia e
na região do Cerrado do Maranhão. Há algumas regiões de alta adequabilidade
ambiental, ao sul do Mato Grosso e norte do Mato Grosso do Sul, assim como no
Cerrado do Tocantins (Figura 21).
Nessas regiões são encontradas 35 Áreas Protegidas (Tabela 2), entre Reservas
Biológicas, Estações Ecológicas e RPPNs, onde não há registro para Vampyrum
spectrum. No Amazonas e Pará, várias destas áreas já foram inventariadas, com graus
variados de esforço de coleta (Bernard et al., 2010).Devido a sua raridade e baixa
densidade populacional, é possível que a espécie ocorra nestes locais, mas ainda assim
não foi detectada. Nos demais estados, tal padrão não é verificado. Essas regiões são de
grande interesse não somente para a ecologia de V. spectrum, mas também para todos os
Chiroptera, por possuírem encontros de biomas em seus territórios (i.e. Maranhão com
Caatinga e Floresta Amazônica; Mato Grosso com Cerrado, Floresta Amazônica e
Pantanal).
A amostragem nestes locais pouco inventariados, assim como o monitoramento
nos locais onde o processo já foi feito, podem aprimorar os conhecimentos da
quiropterofauna brasileira.
Tabela 2: Áreas Protegidas no Brasil em áreas de adequabilidade ambiental alta. Valores extraídos do
Ensemble II (para explicação, ver texto). ESEC = Estação Ecológica, PARNA = Parque Nacional,
PARNES = Parque Estadual, REEXT = Reserva Extrativista, REBIO = Reserva Biológica, REFNA =
Reserva Florestal, RPPN = Reserva Particular do Patrimônio Nacional.
Área Protegida Estado Adequabilidade
PARNA da Serra do Divisor Acre 0.812 ESEC Rio Acre Acre 0.628

30
Área Protegida Estado Adequabilidade
REEXT Chico Mendes Acre 0.622 RPPN Santuário Ecológico Amazônia Viva Acre 0.614 REEXT Alto Juruá Acre 0.538 REEXT Rio Cajari Amapá 0.793 RPPN Seringal Triunfo Amapá 0.726 REBIO do Lago Piratuba Amapá 0.625 ESEC do Jari Amapá - Pará 0.802 REBIO do Uatumã Amazonas 0.807 ESEC de Anavilhanas Amazonas 0.726 ESEC de Jutaí-Solimões Amazonas 0.703 ESEC de Mamirauá Amazonas 0.703 PARNA do Jaú Amazonas 0.695
PARNES Serra do Aracá Amazonas - Roraima 0.714 RPPN Fazenda São Francisco Maranhão 0.542 PARNA dos Lençóis Maranhenses Maranhão 0.507 PARNA da Chapada dos Guimarães Mato Grosso 0.531 RPPN Estância Ecológica SESC - Pantanal Mato Grosso 0.525 ESEC de Taiamã Mato Grosso 0.488 RPPN Fazenda Estância Dorochê Mato Grosso 0.461 PARNA do Pantanal Mato Grossense Mato Grosso 0.458 RPPN Fazenda Santa Helena Mato Grosso do Sul 0.518 PARNA da Amazônia Pará 0.791 REBIO do Rio Trombetas Pará 0.731 REFNA do Tumucumaque Pará 0.611
ESEC Samuel Rondônia 0.574 REEXT Rio Preto/Jacundá Rondônia 0.523 REEXT Rio Jacy-Paraná Rondônia 0.488 PARNA do Viruá Roraima 0.751 ESEC do Niquiá Roraima 0.702 PARNA Serra da Mocidade Roraima 0.689 ESEC de Caracaraí Roraima 0.688 REFNA do Parima Roraima 0.653 PARNA do Araguaia Tocantins 0.507

31
Figura 21: Áreas Prioritárias para Inventários de Vampyrum spectrum no Brasil.

32
4.2.6 Ecologia
Handley (1976) lista uma variedade de ambientes de captura de Vampyrum
spectrum que inclui bancos de rios, pântanos, sopé de montes e florestas clímax. Ainda
assim, a maioria dos registros para a espécie limita-se à última categoria. No Brasil, as
coletas de morcegos concentram-se principalmente na região Norte, com ênfase na
Floresta Amazônica dos estados do Pará e Amazonas (Bernard et al., 2010). Apenas
recentemente foram conduzidos inventários em estados com biodiversidade esperada
elevada, como o Amapá (Martins et al., 2006) e Rondônia (Discher et al., 2009).
Portanto, outros biomas brasileiros ficam com sua biodiversidade equivocadamente
reduzida em comparação com a Floresta Amazônica. Para V. spectrum, a ocorrência no
Pantanal brasileiro não é surpreendente, uma vez que há registros em latitudes próximas
na Bolívia, além de registros da espécie em ambientes pantanosos (Vieira, 1955;
Handley, 1976). O mesmo não pode ser dito da presença no Nordeste brasileiro, no
estado do Piauí (Gregorin et al., 2008). De fato, esforços de inventários na última
década na Caatinga brasileira resultaram na extensão de ocorrência de diversas espécies,
além da descrição de pelo menos duas novas: Chiroderma vizzotoi (Taddei & Lim,
2010) e Xeronycteris vierai (Gregorin & Ditchfield, 2005). Tais fatores evidenciam a
falta de conhecimento ecológico não somente para a espécie, mas também dos biomas
brasileiros.
Outro registro notável deste estudo é que a presença de V. spectrum no Nordeste
brasileiro (assim como o registro no Tocantins de Nunes et al., 2005) é, em termos de
adequabilidade ambiental, isolado dos demais pontos de ocorrência para a espécie,
mesmo quando analisados os modelos com threshold mínimo. No momento, algumas
hipóteses podem ser levantadas: (1) V. spectrum pode ter uma capacidade migratória
capaz de cruzar longas distâncias em condições desfavoráveis; (2) é possível que as
localidades entre as zonas favoráveis também o sejam, porém os modelos não prevejam
devido à diferença ambientais e disparidade entre as ocorrências; (3) as populações
nestes ambientes podem ser relictuais, antigos remanescentes isolados pela
fragmentação ambiental.
4.2.7 Inferências Biogeográficas
Todos os modelos apresentaram uma zona na região litorânea do Brasil,
principalmente no sul da Bahia e pouco no norte do Espírito Santo, com condições
altamente favoráveis para ocorrência de Vampyrum spectrum. É improvável que a

33
região possua populações da espécie, devido a fatores de isolamento, mas as análises
dão suporte para uma hipótese descritiva dos biomas brasileiros, a Hiléia Baiana
(Andrade-Lima, 1966). A Hiléia Baiana seria uma região disjunta da Floresta
Amazônica, mas com fauna e flora similares, devido a condições ambientais parecidas
entre ambas. Uma vez que apenas variáveis abióticas, relativas ao clima, foram
utilizadas na construção do modelo, o aparecimento dessas zonas em todos os modelos
constitui evidência para essa hipótese.
Os registros no Pantanal e Nordeste brasileiro mostram um padrão de ocorrência
no país similar à letra “C” (Figura 2), com o vão desfavorável sendo ocupado pela bacia
hidrográfica do rio Amazonas. A distribuição também sugere dois “braços”,
comunicando-se com regiões distintas da Mata Atlântica, sugerindo a existência de
corredores ecológicos, um ao norte ligando Floresta Amazônica – Caatinga – Mata
Atlântica, e um ao sul ligando Floresta Amazônica – Pantanal – Mata Atlântica. Costa
(2003) mostrou, por meio de filogeografia de pequenos mamíferos não voadores
(roedores e marsupiais), a existência de dois possíveis corredores históricos entre a
Floresta Amazônica e Mata Atlântica, similares aos sugeridos pelo presente estudo.

34
5 CONCLUSÕES
Vampyrum spectrum possui distribuição em grande parte da região Neotropical,
concentrada na Floresta Amazônica e América Central, com registros mais esparsos na
Caatinga e Pantanal brasileiros, devido a esforços de coleta e identificação serem
realizados nessas regiões de forma mais ativa na última década. Com o conhecimento
atual, ainda não é possível esclarecer precisamente a ocorrência da espécie na Caatinga,
visto que as áreas intermediárias são previstas como não adequadas para ocorrência.
Serão precisos mais dados para uma conclusão mais precisa.
Os Modelos de Distribuição Potencial de Vampyrum spectrum mostram que as
zonas de maior adequabilidade ambiental estão concentradas na região Norte do Brasil,
especialmente nos estados do Acre, Amapá e Roraima, e norte do Amazonas, Pará e
Maranhão. Tais resultados, aliados as zonas de adequabilidade ambiental alta no
Suriname, Guiana e Guiana Francesa e Venezuela, além de aproximadamente 14% das
ocorrências estarem em uma ilha (Trinidad), mostram que condições litorâneas não são
obstáculo para ocorrência de V. spectrum. A maior limitação para a espécie são
temperaturas baixas, como aquelas dos Andes.
Os Modelos indicam áreas de grande probabilidade de ocorrência para a espécie,
baseado no valor de adequabilidade ambiental elevado, nas regiões Norte e Centro-
Oeste do Brasil. Existe um número de Áreas Protegidas nessas regiões, com graus
variados de Inventariamento, sem registro de ocorrência para V. spectrum. Esforços de
coleta nas áreas levantadas podem contribuir não somente para maior compreensão da
distribuição de V. spectrum, mas também aumentar o conhecimento da quiropterofauna
brasileira.
Os thresholds de mínima presença são ineficazes para indicar áreas adequadas
para condução de inventários da quiropterofauna, pois a área prevista é quase que
totalmente a área utilizada para o estudo. O Mean Training Cell, proposto neste
trabalho, é um threshold relativo aos dados de adequabilidade ambiental que fornece
resultados mais satisfatórios para estes intentos.
Os algoritmos ENFA, Maxent, SVM e GARP possuem desempenho e variância
similares. O Random Forests, no entanto, possui comportamento muito dissimilar dos
demais. A versão disponível atualmente deve ser usada com cautela.

35
6 REFERÊNCIAS BIBLIOGRÁFICAS
Acosta, L.S., Azurduy, H. Primeras colectas del falso vampiro Vampyrum spectrum
(Phyllostomidae, Chiroptera) en el sector sur del bosque seco Chiquitano, Santa
Cruz, Bolivia. Kempffiana, 2:1, 119 – 126, 2006.
Aguirre, L.F., Mantilla, H., Miller, B., Dávalos, L. Vampyrum spectrum. In: IUCN
2012. IUCN Red List of Threatened Species. 2008.
Akkiraju, N., Edelsbrunner, H., Facello, M., Fu, P., Mücke, E.P., Varela, C. Alpha
shapes: definition and software. In: Proceedings of the International Computational
Geometry (edited by Amenta, N.). Software Workshop, 63 – 66, Geometry Center,
Minneapolis, Minnesota, United States of America, 1995.
Allouche, O., Tsoar, A., Kadmon, R. Assessing the accuracy of species distribution
models: prevalence, kappa and the true skill statistic (TSS). Journal of Applied
Ecology, 43, 1223 – 1232, 2006.
Anderson, R.P., Lew, D., Peterson, A.T. Evaluating predictive models of species’
distributions: criteria for selecting optimal models. Ecological Modelling, 162, 211
– 232, 2003.
Andrade-Lima, D. Contribuição ao estudo do paralelismo da flora amazônico-
nordestina. Boletim do Instituto de Pesquisas Agronômicas (Nova Série), 19, 3- 30,
1966.
Arita, H.T. Rarity in Neotropical bats: Correlations with phylogeny, diet, and body
mass. Ecological Applications, 3:3, 506 – 517, 1993.
Barreto, F.C.C. Modelagem de distribuição potencial de espécies como ferramenta para
a conservação: seleção e avaliação de algoritmos e aplicação com Heliconius
nattereri Felder, 1865 (Nymphalidae, Heliconiinae). Tese de Doutorado. Viçosa,
Minas Gerais, Brasil. 2008.
Bernard, E., Aguiar, L.M.S., Machado, R.B. Discovering the Brazilian bat fauna: a task
for two centuries? Mammal Review, 1 – 17, 2010.
Bonato, V., Facure, K.G., Uieda, W. Food habits of bats of subfamily Vampyrinae in
Brazil. Journal of Mammalogy, 85:4, 708 – 713, 2004.
Breiman, L. Random Forests. Machine Learning, 45, 5 – 32, 2001.
Calenge, C. Exploring habitat selection by wildlife with adehabitat.Journal of Statistical
Software, 22:6, 1 – 19, 2007.

36
Cortes, C., Vapnik, V. Support-Vector Networks. Machine Learning, 20, 273 – 297,
1995.
Costa, L.P. The historical bridge between the Amazon and the Atlantic Forest of Brazil:
a study of molecular Phylogeography with small mammals. Journal of
Biogeography, 30, 71 – 86, 2003.
Damuth, J. Population density and body size in mammals. Nature, 290, 699 – 700,
1981.
De Marco Jr., P., Siqueira, M.F. Como determinar a distribuição potencial de espécies
sob uma abordagem conservacionista? Megadiversidade, 5:1-2, 65 – 76, 2009.
Discher, D.S., Bernarde, P.S., Facure, K.G. Mammalia, Chiroptera,
Phyllostomidae,Vampyrum spectrum (Linnaeus, 1758): First record for the state of
Rondônia, Brazil, and new prey records. Check List, 5:3, 394 – 395, 2009.
Edelsbrunner, H., Kirkpatrick, D.G., Seidel, R. On the shape of a set of points in the
plane. IEEE Transactions on Information Theory, 29, 551 – 559, 1983.
Elith, J., Graham, C.H., Anderson, R.P., Dudík, M., Ferrier, S., Guisan, A., Hijmans,
R.J., Huettmann, F., Leathwick, J.R., Lehmann, A., Li, J., Lohmann, L.G., Loiselle,
B.A., Manion, G., Moritz, C., Nakamura, M., Nakazawa, Y., Overton, J.M.,
Peterson, A.T., Phillips, S.J., Richardson, K., Scachetti-Pereira, R., Schapire, R.E.,
Soberón, J., Williams, S., Wisz, M.S., Zimmermann, N.E. Novel methods improve
prediction of species’ distributions from occurrence data. Ecography, 29, 129 –
151, 2006.
Fleming, T.H., Hooper, E.T., Wilson, D.E. Three Central American bat communities:
structure, reproductive cycles, and movement patterns. Ecology, 53:4, 555 – 569,
1972.
Giannini, T.C., Siqueira, M.F., Acosta, A.L., Barreto, F.C.C., Saraiva, A.M., Alves-dos-
Santos, I. Desafios atuais da modelagem preditiva de distribuição de espécies.
Rodriguésia, 63:3, 733 – 749, 2012.
Gregorin, R., Carmignotto, A.P., Percequillo, A.R. Quirópteros do Parque Nacional da
Serra das Confusões, Piauí, nordeste do Brasil. Chiroptera Neotropical, 14:1, 366 –
383, 2008.
Gregorin, R., Ditchfield, A.D. New genus and species of nectar-feeding bat in the tribe
Lonchophyllini (Phyllostomidae: Glossophaginae) from Northeastern Brazil.
Journal of Mammalogy, 86:2, 403 – 414, 2005.

37
Grossmann, E., Ohmann, J., Kagan, J., May, H., Gregory, M. Mapping ecological
systems with a random forest model: tradeoffs between error and bias. Gap
Analysis Bulletin, 17, 16 – 22, 2010.
Guo, Q., Kelly, M., Graham, C.H. Support vector machines for predicting distribution
of Sudden Oak Death in California. Ecological Modelling, 182, 75 – 90, 2005.
Handley, C.O. Mammals of the Smithsonian Venezuelan Project.Brigham Young
University Science Bulletin.20:5, 1 – 91, 1976.
Henry, M., Cosson, J.F., Pons, J.M. Modelling multi-scale spatial variation in species
richness from abundance data in a complex neotropical bat assemblage. Ecological
Modelling, 221, 2018 – 2027, 2010.
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G., Jarvis, A. Very high resolution
interpolated climate surfaces for global land areas. International Journal of
Climatology, 25, 1965 – 1978, 2005.
Hirsch, A., Chiarello, A.G. The endangered maned sloth Bradypus torquatus of the
Brazilian Atlantic forest: a review and update of geographical distribution and
habitat preferences. Mammal Review, 1 – 20, 2011.
Hirzel, A.H., Hausser, J., Chessel, D., Perrin, N. Ecological-Niche Factor Analysis:
How to compute habitat-suitability maps without absence data? Ecology, 83:7,
2027 – 2036, 2002.
Husson, F., Josse, J., Le, S., Mazet, J. FactoMineR: Multivariate exploratory data
analysis and data mining with R, 2013. R package version 1.23. Disponível em
<http://CRAN.R-project.org/package=FactoMineR>
Hutchinson, G.E. Concluding remarks. Cold Spring Harbor Symposia on Quantitative
Biology, 22:2, 415 – 427, 1957.
Jiménez-Valverde, A. Insights into the area under the receiver operating characteristic
curve (AUC) as a discrimination measure in species distributional modelling.Global
Ecology and Biogeography, 21, 498 – 507, 2012.
Kéry, M. Ecology of small populations. Dissertation.University of Zürich, Zürich,
Switzerland. 2000 apud Hirzel, A.H., Hausser, J., Chessel, D., Perrin, N.
Ecological-Niche Factor Analysis: How to compute habitat-suitability maps
without absence data? Ecology, 83:7, 2027 – 2036, 2002.
Knudby, A., Brenning, A., LeDrew, E. New approaches to modelling fish-habitat
relationships. Ecological Modelling, 221, 503 – 511, 2010.

38
Lewin-Koh, N.J., Bivand, R. maptools: Tools for reading and handling spatial objects,
2012. R package version 0.8-16. Disponível em <http://CRAN.R-
project.org/package=maptools>
Liaw, A., Wiener, M. Classifications and Regression by randomForest. R News, 2:3, 18
– 22, 2002.
Linnaeus, C. Systema naturae.10º ed. Stockholm, L. Salvii, 1, 1 – 824, 1758.
Lobo, J.M., Jiménez-Valverde, A., Real, R. AUC: a misleading measure of the
performance of predictive distribution models. Global Ecology and Biogeography,
17, 145 – 151, 2008.
Liu, C., Berry, P.M., Dawson, T.P., Pearson, R.G. Selecting thresholds of occurrence in
the prediction of species distributions. Ecography, 28, 385 – 393, 2005.
Mantilla-Meluk, H., Jiménez-Ortega, A.M., Baker, R.J. Phyllostomid bats of Colombia:
Annotated checklist, distribution, and biogeography. Special Publications of the
Museum of Texas Tech University, 56, 1 – 37, 2009.
Martins, A.C.M., Bernard, E., Gregorin, R. Inventários biológicos rápidos de morcegos
(Mammalia, Chiroptera) em três unidades de conservação do Amapá, Brasil.
Revista Brasileira de Zoologia, 23:4, 1175 – 1184, 2006.
McNab, B.K. The structure of tropical bat faunas.Ecology, 52, 352 – 358, 1971.
Muñoz, M.E.S., Giovanni, R., Siqueira, M.F., Sutton, T., Brewer, P., Pereira, R.S.,
Canhos, D.A.L., Canhos, V.P. openModeller: a generic approach to species’
potential distribution modelling. Geoinformatica, 2009.
Navarro, D.L., Wilson, D.E. Vampyrum spectrum. Mammalian Species, 184, 1 – 4,
1982.
Nogueira, M.R., Peracchi, A.L., Moratelli, R. Subfamília Phyllostominae. In: Morcegos
do Brasil (edited by Reis, N.R, Peracchi, A.L., Pedro, W.A., Lima, I.P.).
Universidade Estadual de Londrina ,61 – 98, 2007.
Pateiro-Lopez, B., Rodriguez-Casal, A. alphahull: Generalization of the convex hull of a
sample of points in the plane, 2011. R package version 0.2-1. Disponível em
<http://CRAN.R-project.org/package=alphahull>
Pearce, J., Ferrier, S. An evaluation of alternative algorithms for fitting species
distribution models using logistic regression. Ecological Modelling, 128, 127 –
147, 2000.
Pearson, K. On lines and planes of closest fit to systems of points in space.
Philosophical Magazine 2:11, 559 – 572, 1901.

39
Peracchi, A.L., Gallo, P.H., Dias, D., Lima, I.P., Reis, N.R. Ordem Chiroptera. In:
Mamíferos do Brasil (edited by Reis, N.R., Peracchi, A.L., Fregonezi, M.N.,
Rossaneis, B.K.), Techinical Books, Rio de Janeiro, 293 – 461, 2010.
Peterson, A.T., Papeş, M., Soberón, J. Rethinking receiver operating characteristic
analysis applications in ecological niche modelling. Ecological Modelling, 213, 63
– 72, 2008.
Phillips, S.J., Anderson, R.P., Schapire, R.E. Maximum entropy modelling of species
geographic distributions. Ecological Modelling, 190, 231 – 259, 2006.
R Development Core Team. R: A language and environment for statistical computing.
R Foundation for Statistical Computing, Vienna, Austria, 2013. Disponível em
<http://www.R-project.org/>
Rafinesque, C.S. Analyse de la nature on tableau de l’Universet des corps organises.
Palermo, 1815.
Renner, I.W., Warton, D.I. Equivalence of MAXENT and Poisson Point Process
Models for species distribution modelling in ecology. Biometrics, 1 – 8, 2013.
Robinson, J.G., Redford, K.H. Body size, diet, and population density of Neotropical
forest mammals. The American Naturalist, 128, 665 – 680, 1986.
Sousa, R.F., Kreutz, C., Oliveira, S.L., Faria, K.C. Mammalia, Chiroptera,
Phyllostomidae, Vampyrum spectrum (Linnaeus, 1758): First record for the Cerrado
biome in the state of Mato Grosso, west central Brazil. Check List, 7:4, 468 – 469,
2011.
Stockwell, D.R.B., Noble, I.R. Induction of sets of rules from animal distribution data: a
robust and informative method of data analysis. Mathematics and Computers in
Simulations, 33, 385 – 390, 1992.
Taddei, V.A., Reis, N.R. Notas sobre alguns morcegos da Ilha de Maracá, Território
Federal de Roraima (Mammalia, Chiroptera). Acta Amazonica, 10:2, 363 – 368,
1980.
Taddei, V.A., Lim, B.K. A new species of Chiroderma (Chiroptera, Phyllostomidae)
from Northeastern Brazil. Brazilian Journal of Biology, 70:2, 381 – 386, 2010.
Tarca, A.L., Carey, V.J., Chen, X., Romero, R., Drӑghici, S. Machine Learning and its
applications to Biology. PLoS Computational Biology, 3:6, 953 – 962, 2007.
Terribile, L.C., Diniz-Filho, J.A.F., De Marco Jr., P. How many studies are necessary to
compare niche-based models for geographic distributions? Inductive reasoning may
fail at the end. Brazilian Journal of Biology, 70:2, 263 – 269, 2010.

40
VanDerWal, J., Falconi, L., Januchowski, S., Shoo, L., Storlie, C. SDMTools: Species
distribution modelling tools: Tools for processing data associated with species
distribution modelling exercises, 2012. R package version 1.1-12. Disponível em
<http://CRAN.R-project.org/package=SDMTools>
Vieira, C.O.C. Lista remissiva dos mamíferos do Brasil. Arquivos de Zoologia da
Universidade de São Paulo, 8, 354 – 374, 1955.
Williams, S.L., Genoways, H.H. Subfamily Phyllostominae Gray, 1825. In: Mammals
of South America. Volume 1: Marsupials, Xenarthrans, Shrews, and Bats (edited by
Gardner, A.L.). The University of Chicago Press, Chicago, 255 – 300, 2007.
Yom-Tov, Y., Kadmon, R. Analysis of the distribution of insectivorous bats in Israel.
Diversity and Distributions, 4, 63 – 70, 1998.

41
7 ANEXOS
7.1 Anexo 1: Pontos de Ocorrência de Vampyrum spectrum
ID Registro País Latitude Longitude Fonte
1 FMNH 58160 Belize 16.230 -89.109 1
2 AMNH 256825 Belize 16.530 -88.800 1
3 MSB 211451 Bolívia -14.560 -65.900 2
4 AMNH 261379 Bolívia -14.560 -65.090 3
5 AMNH 261379 Bolívia -14.560 -65.090 2
6 AMNH 261379 Bolívia -14.560 -65.090 1
7 CBFLP S/N Bolívia -14.216 -66.400 4, 5
8 - Bolívia -17.365 -65.233 5
9 MNK 3227 Bolívia -14.566 -60.900 5
10 - Bolívia -13.750 -68.350 5
11 MNK 3708 Bolívia -18.467 -58.283 6
12 MNK 3709 Bolívia -18.467 -58.283 6
13 ZUEC-MAM 2457 Brasil -11.400 -61.367 7
14 ZEE_MAM 4072 Brasil -7.557 -73.277 7
15 ZEE_MAM 116 Brasil -7.447 -73.678 7
16 MZUSP 10103 Brasil -3.400 -55.400 1
17 USNM 393007 Brasil -1.470 -48.360 1
18 FMNH 126262 Brasil -1.450 -48.483 2
19 MZUSP 12946 Brasil -1.010 -56.150 1
20 MCZ 30295 Brasil -1.000 -50.000 1
21 USNM 460693 Brasil -0.660 -58.500 1
22 MBML 1915 Brasil -0.130 -67.089 7
23 FPR-COLOMBIA 64482 Brasil 0.600 -69.200 7
24 ALP S/N Brasil -7.383 -73.650 8, 9, 10
25 INPA 2701 Brasil -2.417 -59.750 32, 9, 10
26 INPA S/N Brasil -2.500 -54.950 33, 9, 34, 10
27 - Brasil -2.650 -55.633 11, 9
28 - Brasil -15.980 -55.900 12
29 UFMT 1116 Brasil -16.350 -56.330 13
30 UFMT 1117 Brasil -16.330 -56.330 13
31 - Brasil -19.550 -55.580 35
32 M-CPAP 307 Brasil -18.980 -56.610 35
33 MZUSP 33556 Brasil -9.220 -43.498 14
34 ZUEC-MAM 2457 Brasil -11.418 -61.367 15
35 ALP S/N Brasil 0.895 -52.002 16
36 MPEG 31650 Brasil -8.617 -48.350 17
37 IEPA S/N Brasil 1.283 -51.588 18
38 IEPA S/N Brasil 2.193 -54.587 18
39 CIAH 7211 Colômbia 0.742 -72.451 3
40 ICN 8389 Colômbia 4.063 -73.515 3
41 FPR-COLOMBIA 5231 Colômbia 4.100 -73.800 7
42 LACM 38771 Costa Rica 8.697 -83.497 1

42
ID Registro País Latitude Longitude Fonte
43 ZMB 2489 Costa Rica 9.930 -85.050 1
44 LACM 14364 Costa Rica 10.100 -83.383 1
45 KU 143307 Costa Rica 10.310 -84.770 1
46 LACM 38548 Costa Rica 10.333 -85.200 2
47 USNM 562766 Costa Rica 10.350 -84.010 1
48 MSB 37722 Costa Rica 10.433 -84.000 2
49 KU 117624 Costa Rica 10.440 -85.110 1
50 KU 88190 Costa Rica 10.460 -84.010 1
51 LACM 28469 Costa Rica 10.467 -85.133 2
52 USNM 562765 Costa Rica 10.520 -85.250 1
53 USNM 568490 Costa Rica 10.600 -83.530 2
54 ROM 105870 Equador -1.040 -75.870 1
55 AMNH 66815 Equador -0.600 -79.570 1
56 USNM 528490 Equador -0.586 -79.365 2
57 FMNH 53505 Equador -0.217 -78.500 1
58 MVZ 227288 Guatemala 15.407 -88.696 2
59 USNM 564627 Guatemala 17.350 -89.900 1
60 RNH 12115 Guiana 6.130 -57.550 1
61 AMNH 42895 Guiana 6.392 -58.617 1
62 ROM S/N Guiana 4.667 -58.683 19, 20, 9
63 RNH 15906 Guiana Francesa 4.520 -52.620 1
64 AMNH 267446 Guiana Francesa 5.270 -52.920 1
65 - Guiana Francesa -3.617 -52.200 9
66 MNHN S/N Guiana Francesa 4.033 -52.700 21, 22, 9
67 AMNH 267446 Guiana Francesa 5.266 -52.917 23, 9
68 TTU 13143 Honduras 15.019 -85.533 2
69 IBUNAM 38355 México 16.100 -90.933 3
70 IBUNAM 36606 México 17.867 -89.717 3
71 USNM 78127 México 18.090 -94.250 1
72 IBUNAM 39351 México 18.487 -95.082 3
73 IBUNAM 16549 México 18.583 -95.067 3
74 MZFC 434 México 18.594 -95.096 3
75 MZFC 434 México 18.594 -95.096 3
76 IBUNAM 16548 México 18.692 -95.114 3
77 ECOAM-MAM S/N México 17.776 -95.959 24, 36
78 ECOAM-MAM 883 México 16.801 -94.718 25
79 AMNH 28993 Nicarágua 12.640 -86.900 1
80 USNM 309373 Panamá 7.800 -77.470 1
81 LSUMZ 25561 Panamá 7.855 -77.755 3
82 AMNH 18707 Panamá 8.300 -82.340 1
83 IBUNAM 6144 Panamá 8.533 -79.850 3
84 IBUNAM 6636 Panamá 8.533 -79.850 3
85 USNM 335162 Panamá 8.670 -77.460 1
86 ROM 45335 Panamá 8.850 -82.567 2
87 ROM 45335 Panamá 8.850 -82.567 26

43
ID Registro País Latitude Longitude Fonte
88 USNM 396402 Panamá 8.980 -79.590 1
89 BM 6551869 Panamá 9.000 -80.000 1
90 USNM 503833 Panamá 9.150 -79.850 1
91 USNM 335159 Panamá 9.380 -82.250 1
92 ROM 45335 Panamá 8.850 -82.567 26
93 BM 695132 Peru -12.990 -71.200 1
94 FMNH 138875 Peru -12.933 -71.250 27
95 LSUMZ 16458 Peru -12.733 -73.767 3
96 MVZ 165941 Peru -12.600 -69.073 2
97 FMNH 55402 Peru -8.300 -74.600 1
98 MVZ 154780 Peru -4.280 -78.050 1
99 MVZ 154781 Peru -4.267 -78.150 2
100 MVZ 154782 Peru -4.267 -78.150 2
101 - Peru -12.550 -69.050 28
102 MUSM S/N Peru -4.917 -77.733 29, 9
103 MUSM S/N Peru -11.900 -71.367 27, 22, 9
104 - Peru -3.748 -73.240 30
105 MUSM 22208 Peru -3.756 -80.387 31
106 FMNH 138875 Peru -12.933 -71.250 27
107 RNH 15908 Suriname 4.000 -56.000 1
108 ZMB 4619 Suriname 5.020 -54.210 1
109 RNH 25243 Suriname 5.500 -55.100 1
110 LACM 15786 Trinidad & Tobago 10.083 -61.283 2
111 AMNH 175717 Trinidad & Tobago 10.180 -61.220 1
112 AMNH 7436 Trinidad & Tobago 10.300 -61.150 1
113 FMNH 85786 Trinidad & Tobago 10.400 -61.367 1
114 FMNH 85787 Trinidad & Tobago 10.400 -61.367 2
115 FMNH 61924 Trinidad & Tobago 10.483 -61.217 1
116 FMNH 61945 Trinidad & Tobago 10.483 -61.217 2
117 FMNH 61946 Trinidad & Tobago 10.483 -61.217 2
118 TTU 5357 Trinidad & Tobago 10.500 -61.250 2
119 USNM 337312 Trinidad & Tobago 10.520 -61.060 1
120 MVZ 167391 Trinidad & Tobago 10.567 -61.133 1
121 TTU 9836 Trinidad & Tobago 10.583 -61.117 2
122 TTU 9837 Trinidad & Tobago 10.583 -61.117 2
123 TTU 11439 Trinidad & Tobago 10.583 -61.117 2
124 FMNH 80930 Trinidad & Tobago 10.583 -61.117 2
125 FMNH 80931 Trinidad & Tobago 10.583 -61.117 2
126 FMNH 51220 Trinidad & Tobago 10.650 -61.517 2
127 BM 938101 Trinidad & Tobago 10.670 -61.470 1
128 TTU 47873 Trinidad & Tobago 10.683 -61.400 2
129 TTU 47874 Trinidad & Tobago 10.683 -61.400 2
130 FPR-COLOMBIA 64479 Venezuela 1.721 -65.033 7
131 FPR-COLOMBIA 64478 Venezuela 2.500 -65.183 7
132 FPR-COLOMBIA 64476 Venezuela 3.067 -63.133 7

44
ID Registro País Latitude Longitude Fonte
133 FPR-COLOMBIA 64481 Venezuela 5.244 -67.795 7
134 FPR-COLOMBIA 64480 Venezuela 5.350 -66.100 7
135 USNM 407630 Venezuela 5.383 -66.100 1
136 USNM 385520 Venezuela 5.983 -61.433 1
137 FPR-COLOMBIA 64477 Venezuela 6.583 -67.217 7
138 FPR-COLOMBIA 64475 Venezuela 7.278 -71.919 7
139 TTU 48106 Venezuela 8.436 -67.433 2
140 PBDB 212910 Venezuela 8.567 -67.583 3
141 FMNH 18786 Venezuela 9.050 -72.230 1
142 USNM 385521 Venezuela 10.333 -65.915 1
143 - Venezuela 7.750 -61.167 9
144 - Venezuela 8.041 -61.766 9
145 - Venezuela 7.992 -70.767 9

45
7.2 Anexo2: Lista de Instituições
ALP: Coleção Peracchi, Universidade Federal Rural do Rio de Janeiro.
AMNH: American Museum of Natural History
BM: British Museum
CBFLP: Colección Boliviana de Fauna en La Paz
CIAH: Colecciones Instituto Alexander von Humboldt
ECOAM-MAM: Coléccion de Referencia de Mamíferos del Laboratorio de Ecología
Animal del Centro Interdisciplinario de Investigación para el Desarrollo Integral
Regional, Unidad Oaxaca del Instituto Politecnico Nacional de México
FMNH: Florida Museum of Natural History
FPR-COLOMBIA: Fundación Puerto Rastrojo
IBUNAM: Instituto de Biologia da Universidad Nacional Autónoma de México
ICN: Instituto de Ciencias Naturales
IEPA: Instituto de Pesquisas Científicas e Tecnológicas do Amapá
INPA: Instituto Nacional de Pesquisas da Amazônia
KU: University of Kansas
LACM: Natural History Museum of Los Angeles
LSUMZ: Louisiana State University Museum of Natural Science
M-CPAP: Embrapa Pantanal’sVertebrate Reference Collection
MBML: Museu de Biologia Professor Mello Leitão
MCZ: Museum of Comparative Zoology of Harvard
MNHN: Muséum National d’Histoire Naturelle
MNK: Museo de Historia Natural Noel Kempff Mercado
MPEG: Museu Paraense Emílio Goeldi
MSB: Museum of Southwestern Biology, Division of Mammals
MUSM: Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos
MVZ: Museum of Vertebrate Zoology of Berkeley
MZFC: Coléccion de Mamíferos del Museo de Zoología “Alfonso L. Herrera”, México
MZUSP: Museu de Zoologia da Universidade de São Paulo
PBDB: Paleobiology Database
RNH: Rijksmuseum van Natuurlijke Historie
ROM: Royal Ontario Museum
TTU: Museum of Texas Tech University
UFMT: Coleção Zoológica da Universidade Federal do Mato Grosso
USNM: National Museum of Natural History
ZEE_MAM: Zoneamento Ecológico Econômico do Acre – Mammalia
ZMB: Museum fur Naturkunde, Berlin
ZUEC-MAM: Coleção de Mamíferos do Museu de Zoologia da Unicamp

46
7.3 Anexo 3: Fontes dos Pontos de Ocorrência
1. Dados não publicados do Dr. Albert David Ditchfield.
2. Mammal Networked Information System (MaNIS). Disponível em
<http://manisnet.org>
3. Global Biodiversity Information Facility (GBIF). Disponível em <http://gbif.org>
4. Aguirre, L.F. Structure of a neotropical savanna bat community. Journal of
Mammalogy, 83:3, 775 – 784, 2002.
5. Vargas-Espinoza, A., Aguirre, L.F., Swarner, M., Emmons, L., Teran, M.
Distribución de Vampyrum spectrum en Bolivia y comentários sobre su estado de
conservación. Ecología en Bolivia, 39:2, 46 – 51, 2004.
6. Acosta, L., Azurduy, H.. Primeras colectas del falso vampiro Vampyrum spectrum
(Phyllostomidae, Chiroptera) en el sector sur del Bosque Seco Chiquitano, Santa
Cruz, Bolivia. Kempffiana, 2:1,119 – 126, 2006.
7. speciesLink. Disponível em<http://splink.cria.org.br>
8. Nogueira, M.R., Pol, A., Peracchi, A.L. New records of bats from Brazil with a list of
additional species for the chiropteran fauna of the state of Acre, western Amazon
basin. Mammalia, 63:3, 363 – 368, 1999.
9. Barnett, A.A., Sampaio, E.M., Kalko, E.K.V., Shapley, R.L., Fischer, E., Camargo,
G., Rodríguez-Herrera, B. Bats of Jaú National Park, central Amazônia, Brazil.
Acta Chiropterologica, 8:1, 103 – 128, 2006.
10. Bernard, E., Tavares, V.C., Sampaio, E.M. Compilação atualizada das espécies de
morcegos (Chiroptera) para a Amazônia Brasileira. Biota Neotropica, 11:1, 35 – 46,
2011.
11. Kalko, E.K.V., Handley, C.O. Neotropical bats in the canopy: diversity, community
structure, and implications for conservation. Plant Ecology, 153, 319 – 333, 2001.
12. Vieira, C.O.C. Lista remissiva dos mamíferos do Brasil. Arquivos de Zoologia da
Universidade de São Paulo, 8, 354 – 374, 1955.
13. Silva, A.P., Rossi, R.V. New records of Vampyrum spectrum (Chiroptera,
Phyllostomidae) for the Pantanal domain in Brazil, with notes on the species natural
history, biometry, and lower incisors arrangement. Chiroptera Neotropical, 17:1,
836 – 841, 2011.
14. Gregorin, R., Carmignotto, A.P., Percequillo, A.R. Quirópteros do Parque Nacional
da Serra das Confusões, Piauí, nordeste do Brasil. Chiroptera Neotropical, 14:1,
366 – 383, 2008.

47
15. Discher, D.S., Bernarde, P.S., Facure, K.G. Mammalia, Chiroptera, Phyllostomidae,
Vampyrum spectrum (Linnaeus, 1758): First record for the state of Rondônia,
Brazil, and new prey records. Check List, 5:3, 394 – 395, 2009.
16. Peracchi, A.L., Raimundo, S.D.L., Tannure, A.M. Quirópteros do Território Federal
do Amapá, Brasil (Mammalia, Chiroptera). Arquivos da Universidade Federal
Rural do Rio de Janeiro, 7:2, 89 – 100, 1984.
17. Nunes, A., Marques-Aguiar, S., Saldanha, N., Silva, R.S., Bezerra, A. New records
on the geographic distribution of bat species in the Brazilian Amazonia. Mammalia,
69:1, 109 – 115, 2005.
18. Martins, A.C.M., Bernard, E., Gregorin, R. Inventários biológicos rápidos de
morcegos (Mammalia, Chiroptera) em três unidades de conservação do Amapá,
Brasil. Revista Brasileira de Zoologia, 23:4, 1175 – 1184, 2006.
19. Lim, B.K., Engstrom, M.D. Bat community structure at Iwokrama Forest, Guyana.
Journal of Tropical Ecology, 17, 647 – 665, 2001.
20. Lim, B.K., Engstrom, M.D. Species diversity of bats (Mammalia: Chiroptera) in
Iwokrama Forest, Guyana, and the Guianan subregion: implications for
conservation. Biodiversity and Conservation, 10, 613 – 657, 2001.
21. Brosset, A., Charles-Dominique, P. The bats from French Guiana: a taxonomic,
faunistic and ecological approach. Mammalia, 54:4, 509 – 560, 1990.
22. Voss, R.S., Emmons, L.H. Mammalian diversity in Neotropical lowland rainforests:
A preliminary assessment. Bulletin of the American Museum of Natural History,
230, 1 – 120, 1996.
23. Simmons, N.B., Voss, R.S. The mammals of Paracou, French Guiana: a neotropical
lowland rainforest fauna: Part 1. Bats. Bulletin of the American Museum of Natural
History, 237, 1 – 219, 1998.
24. Alfaro, A.M., García-García, J.L., Santos-Moreno, A. Mamíferos de los municípios
Santiago Jocotepec y Ayotzintepec, Chinantla Baja, Oaxaca. Naturaleza y
Desarrollo, 4:1, 19 – 23, 2006.
25. Santos-Moreno, A., Orozco, S.G., Cruz, E.E.P. Records of bats from Oaxaca,
Mexico. The Southwestern Naturalist, 55:3, 454 – 456, 2010.
26. Peterson, R.L., Kirmse, P. Notes on Vampyrum spectrum, the false vampire bat, in
Panama. Canadian Journal of Zoology, 47, 140 – 142, 1969.

48
27. Pacheco, V., Patterson, B.D., Patton, J.L., Emmons, L.H., Solari, S., Ascorra, C.F.
List of mammal species known to occur in Manu Biosphere Reserve, Peru.
Publicaciones del Museo de Historia Natural UNMSM (A) 44, 1 – 12, 1993.
28. Woodman, N., Timm, R.M., Arana, C.R., Pacheco, V., Schmidt, C.A., Hooper,
E.D., Pacheco, C.A. Annotated checklist of the mammals of Cuzco Amazonico,
Peru. Occasional Papers of the Museum of Natural History of the University of
Kansas, 145, 1 – 12, 1991.
29. Ascorra, C.F., Gorchov, D.L., Cornejo, F. The bats from Jenaro Herrera, Loreto,
Peru. Mammalia, 57:4, 533 – 552, 1993.
30. Klingbeil, B.T., Willig, M.R. Seasonal differences in population-, ensemble- and
community-level response of bats to landscape structure in Amazonia. Oikos, 119,
1654 – 1664, 2010.
31. Pacheco, V., Cadenillas, R., Velazco, S., Salas, E., Fajardo, U. Noteworthy bat
records from the Pacific Tropical rainforest region and adjacent dry forest in
northwestern Peru. Acta Chiropterologica, 9:2, 409 – 422, 2007.
32. Sampaio, E.M., Kalko, E.K.V., Bernard, E., Rodríguez-Herrera, B., Handley, C.O.
A Biodiversity assessment of bats (Chiroptera) in a Tropical Lowland Rainforest of
central Amazônia, including methodological and conservation considerations.
Studies on Neotropical Fauna and Environment, 38:1, 17 – 31, 2003.
33. Bernard, E., Fenton, M.B. Species diversity of bats (Mammalia: Chiroptera) in
forest fragments, primary forests, and savannas in central Amazonia, Brazil.
Canadian Journal of Zoology, 80, 1124 – 1140, 2002.
34. Bernard, E., Fenton, M.B. Bats in a fragmented landscape: Species composition,
diversity and habitat interactions in savannas of Santarém, Central Amazonia,
Brazil. Biological Conservation, 134, 332 – 343, 2007.
35. Silveira, M., Munin, R.L., Tomás, W.M., Fischer, E., Bordignon, M.O., Silveira,
G.A. The distribution of the spectral bat, Vampyrum spectrum, reaches the Southern
Pantanal. Biota Neotropica, 11:1, 173 – 175, 2011.
36. Santos-Moreno, A., Espinosa, A.M.A. Mammalian prey of barn owl (Tyto alba) in
Southeastern Oaxaca, México. Acta Zoológica Mexicana (n.s.), 25:1, 143 – 149,
2009.

49
7.4 Anexo 4: Variáveis Ecogeográficas
BIO1: Temperatura Média Anual
BIO2: Temperatura Média Diurna (Média Mensal (Máxima – Mínima))
BIO3: Isotermalidade ((BIO2/BIO7)*100)
BIO4: Sazonalidade da Temperatura (Desvio Padrão * 100)
BIO5: Temperatura Máxima no Mês Mais Quente
BIO6: Temperatura Mínima no Mês Mais Frio
BIO7: Variação Anual da Temperatura (BIO5 – BIO6)
BIO8: Temperatura Média no Trimestre Mais Úmido
BIO9: Temperatura Média no Trimestre Mais Seco
BIO10: Temperatura Média no Trimestre Mais Quente
BIO11: Temperatura Média no Trimestre Mais Frio
BIO12: Precipitação Anual
BIO13: Precipitação no Mês Mais Úmido
BIO14: Precipitação no Mês Mais Seco
BIO15: Sazonalidade da Precipitação (Coeficiente de Variação)
BIO16: Precipitação no Trimestre Mais Úmido
BIO17: Precipitação no Trimestre Mais Seco
BIO18: Precipitação no Trimestre Mais Quente
BIO19: Precipitação no Trimestre Mais Frio
DEM: Modelo Digital de Elevação do Terreno
SLOPE: Inclinação do Terreno
Copyright © 2022 FDOKUMEN




















