
Species distinction and evolutionary relationships of the Italian hare (Lepus corsicanus) as...
13
Molecular Ecology (1999) 8 , 1805 –1817 © 1999 Blackwell Science Ltd Blackwell Science, Ltd Species distinction and evolutionary relationships of the Italian hare ( Lepus corsicanus ) as described by mitochondrial DNA sequencing M. PIERPAOLI, F. RIGA, V. TROCCHI and E. RANDI Istituto Nazionale per la Fauna Selvatica (INFS), Via Cà Fornacetta 9, 40064 Ozzano dell’Emilia (BO), Italy Abstract The taxonomic status of the Italian hare Lepus corsicanus has been uncertain since its first description by W. E. de Winton in 1898 (de Winton WE. Annual Magazine of Natural History , London , 1898, 1, 149 –158). The distribution range of this species has shrunk severely over the last few decades owing to overhunting and restocking with nonindigenous brown hares ( L. europaeus ) in central and southern Italy and Sicily. Recently, scanty populations of Italian hares were rediscovered, and samples for morphological and molecular ana- lyses were collected. Nucleotide sequences of the mitochondrial DNA (mtDNA) control region and cytochrome b indicate that L. corsicanus is a phylogenetically distinct species, which can be identified by concordant morphological and mtDNA traits. It seems to be reproductively isolated and apparently does not hybridize with sympatric brown hares. Phylogenetic analyses suggest that Italian and brown hares are not closely related sister taxa, but belong to distinct evolutionary lineages that dispersed in western Europe in dif- ferent periods during the early Pleistocene. The Italian hare probably differentiated in isolated refuges in southern Italy during the last glaciation. Comparative analyses of mis- match distributions suggest that hares have had different demographic histories during the Pleistocene, which resulted in phylogeographical structuring in Italian hares, but not in brown and mountain ( L. timidus ) hares. The Italian hare is an endangered endemism and needs urgent conservation efforts. Keywords : Lepus corsicanus , mismatch analysis, mtDNA control region, phylogenetics, population genetics, taxonomy Received 11 April 1999; revision received 15 June 1999; accepted 15 June 1999 Introduction Evolution, phylogenetics and population genetics of hares ( Lepus ; Mammalia; Lagomorpha) are poorly known and taxonomic distinctions in some species complexes are confusing (Petter 1961; Angermann 1983; Flux 1983; Chapman & Flux 1990; Hoffmann 1993), despite recent attempts to elucidate the relationships among certain problematic taxa (Yom-Tov 1967; Pérez-Suarez et al . 1994; Palacios 1996, 1998; Hoffmann 1998; Halanych et al . 1999). A typical case of confusion is represented by the uncer- tain status of hares in Italy. The mountain hare L. timidus is distributed in the Alps with relatively undisturbed populations. In contrast, many local populations of the brown hare L. europaeus , originally widespread in north- ern and central Italy with subspecies meridiei , may have been totally replaced by introduced allocthonous hares of different subspecies. Overhunting and the release of brown hares began early in 1900, reaching a peak after World War II; this also included areas in central and southern Italy, a region that is beyond this species’ natural range (Toschi 1965). The Mediterranean hare L. capensis mediterraneus , which was introduced to Sardinia in the 16th century (Vigne 1992), probably belongs to some north African or Middle Eastern populations of the Cape hare L. capensis . The Cape hare is widely distributed and includes disjunct and morphologically differentiated populations with still obscure taxonomy (Yom-Tov 1967; Hoffmann 1998; Suchentrunk et al . 1998). Consequently, it is not clear if the Sardinian hare should be considered a subspecies of the Cape hare (i.e. L. capensis mediterraneus ; Amori et al . 1996), or if it deserves full species rank (i.e. L. Correspondence: E. Randi. Fax: +39 (0) 51 79 66 28; E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Species distinction and evolutionary relationships of the Italian hare (Lepus corsicanus) as...

Molecular Ecology (1999)
8
, 1805–1817
© 1999 Blackwell Science Ltd
Blackwell Science, Ltd
Species distinction and evolutionary relationships of the Italian hare (
Lepus corsicanus
) as described by mitochondrial DNA sequencing
M. PIERPAOLI , F. RIGA, V. TROCCHI and E . RANDI
Istituto Nazionale per la Fauna Selvatica (INFS), Via Cà Fornacetta 9, 40064 Ozzano dell’Emilia (BO), Italy
Abstract
The taxonomic status of the Italian hare
Lepus corsicanus
has been uncertain since its firstdescription by W. E. de Winton in 1898 (de Winton WE.
Annual Magazine of Natural History
,
London
, 1898, 1, 149–158). The distribution range of this species has shrunk severely overthe last few decades owing to overhunting and restocking with nonindigenous brownhares (
L. europaeus
) in central and southern Italy and Sicily. Recently, scanty populationsof Italian hares were rediscovered, and samples for morphological and molecular ana-lyses were collected. Nucleotide sequences of the mitochondrial DNA (mtDNA) controlregion and cytochrome
b
indicate that
L. corsicanus
is a phylogenetically distinct species,which can be identified by concordant morphological and mtDNA traits. It seems to bereproductively isolated and apparently does not hybridize with sympatric brown hares.Phylogenetic analyses suggest that Italian and brown hares are not closely related sistertaxa, but belong to distinct evolutionary lineages that dispersed in western Europe in dif-ferent periods during the early Pleistocene. The Italian hare probably differentiated inisolated refuges in southern Italy during the last glaciation. Comparative analyses of mis-match distributions suggest that hares have had different demographic histories duringthe Pleistocene, which resulted in phylogeographical structuring in Italian hares, but notin brown and mountain (
L. timidus
) hares. The Italian hare is an endangered endemismand needs urgent conservation efforts.
Keywords
:
Lepus corsicanus
, mismatch analysis, mtDNA control region, phylogenetics, populationgenetics, taxonomy
Received 11 April 1999; revision received 15 June 1999; accepted 15 June 1999
Introduction
Evolution, phylogenetics and population genetics ofhares (
Lepus
; Mammalia; Lagomorpha) are poorly knownand taxonomic distinctions in some species complexesare confusing (Petter 1961; Angermann 1983; Flux 1983;Chapman & Flux 1990; Hoffmann 1993), despite recentattempts to elucidate the relationships among certainproblematic taxa (Yom-Tov 1967; Pérez-Suarez
et al
. 1994;Palacios 1996, 1998; Hoffmann 1998; Halanych
et al
. 1999).A typical case of confusion is represented by the uncer-
tain status of hares in Italy. The mountain hare
L. timidus
is distributed in the Alps with relatively undisturbedpopulations. In contrast, many local populations of thebrown hare
L. europaeus
, originally widespread in north-
ern and central Italy with subspecies
meridiei
, may havebeen totally replaced by introduced allocthonous haresof different subspecies. Overhunting and the release ofbrown hares began early in 1900, reaching a peak afterWorld War II; this also included areas in central andsouthern Italy, a region that is beyond this species’ naturalrange (Toschi 1965). The Mediterranean hare
L. capensismediterraneus
, which was introduced to Sardinia in the16th century (Vigne 1992), probably belongs to somenorth African or Middle Eastern populations of the Capehare
L. capensis
. The Cape hare is widely distributed andincludes disjunct and morphologically differentiatedpopulations with still obscure taxonomy (Yom-Tov 1967;Hoffmann 1998; Suchentrunk
et al
. 1998). Consequently, itis not clear if the Sardinian hare should be considered asubspecies of the Cape hare (i.e.
L. capensis mediterraneus
;Amori
et al
. 1996), or if it deserves full species rank (i.e.
L.
Correspondence: E. Randi. Fax: +39 (0) 51 79 66 28; E-mail:[email protected]
MEC766.fm Page 1805 Friday, October 29, 1999 10:55 AM

1806
M . P I E R P A O L I
E T A L
.
© 1999 Blackwell Science Ltd,
Molecular Ecology
, 8, 1805–1817
mediterraneus
; Palacios 1998). Throughout this report wewill use the provisional subspecific name
L.c. mediterraneus
,pending future genetic analyses that are in progress.
The Italian hare
L. corsicanus
, formerly distributed in cen-tral and southern Italy, Sicily and introduced to Corsicaprobably earlier than the 16th century (Vigne 1992; Palacios1996), was described in 1898 by W. E. de Winton. Its speciesrank was soon rejected by Miller (1912) and others(Ellerman & Morrison-Scott 1951; Toschi 1965), who con-sidered
corsicanus
a subspecies of
L. europaeus
. However,Palacios (1996) studied museum specimens, collected towardsthe end of the last century, and described new diagnosticskull, dental and pelage traits that provided phenetic sup-port for the species rank of the Italian hare. However, nogenetic information, defining interspecific divergence andphylogenetic relationships of the Italian hare, was available.
With the exception of two individuals collected in1974–75 in southern Italy (M. Spagnesi, personal commun-ication), and more numerous samples collected in Sicilyfrom 1994 to 1996 (Lo Valvo
et al
. 1997), only scanty andanecdoctal data were available to document persistence,distribution and population consistency of the Italianhare after the turn of the century (Palacios 1996; Fig. 1).
Extensive human disturbance (overhunting and restock-ing) could have threatened, severely restricted and even-tually eradicated the Italian hare from most of its formerhistorical range. Historical distributions suggested thatnatural populations of Italian and brown hares were allo-patric with putative contact zones running from centralTuscany to the Gargano promontory (Palacios 1996;Fig. 1). However, no data were available to document theeffective degree of reproductive isolation or eventualgene flow among populations in contact. Moreover, harescan hybridize in nature, and the integrity of local Italianhare gene pools could have been disrupted by gene intro-gression owing to released, nonindigenous brown hares(Rhymer & Simberloff 1996; Thulin
et al
. 1997a).The uncertain status of the Italian hare stimulated an
assessment of its present distribution range, and the evalu-ation of its population genetic structure and phylogen-etic relationships. Between 1997 and 1998 we collected19 living hares from southern Italy and Sicily showingmorphological traits concordant with the description of
L.corsicanus
(de Winton 1898; Palacios 1996; Riga
et al
. 1998).These findings provided new evidence to support thesurvival of the Italian hare and provided material forgenetic analyses. In this study we aimed to assess thegenetic distinction of
L. corsicanus
, and in particular: (i) toinvestigate the extent of genetic variability among Italianhares collected in peninsular Italy and Sicily; (ii) to recon-struct the phylogenetic relationships of the Italian andother European and African species of hares; and (iii) tooutline a framework for the evolution of
Lepus
in Europe,within the background of Pleistocene events. Maternalinheritance and the absence of recombination makesmitochondrial DNA (mtDNA) analysis an appropriatechoice for reconstructing the recent history of popula-tions and species (Avise
et al
. 1987, 1998). We thereforesequenced the fast-evolving domain I of the mtDNA con-trol region (CR-I) and part of the cytochrome
b
gene(CYB). mtDNA sequences provide the first evidence that
L. corsicanus
is genetically distinct and deeply divergentfrom the other European and African hares. The Italianhare is still persistent in southern Italy and Sicily withpopulations that are genetically isolated from introducedsympatric brown hares.
Materials and methods
Sample collection and species identification
A total of 108 samples, belonging to seven species of
Lepus
,were collected in Europe (Italy, Hungary, Romania, Serbia)and east Africa (Ethiopia) (Table 1). Species were identifiedaccording to the state of multiple diagnostic morpho-logical traits. In particular, individual hares collected inthe Italian lowlands and Apennines were phenotypically
Fig. 1 Approximate distributions and sampling locations ofLepus in Italy and Corsica. Vertical bars and squares: distributionrange and sampling locations of the mountain hare L. timidusin the Italian Alps. White area and dots: natural distributionof the brown hare L. europaeus. Grey area and triangles: histor-ical distribution of the Italian hare L. corsicanus; in Sardinia L.mediterraneus (*) is present. Distributions have been summarizedaccording to Palacios (1996).
MEC766.fm Page 1806 Friday, October 29, 1999 10:55 AM
G E N E T I C S O F
L E P U S C O R S I C A N U S
1807
© 1999 Blackwell Science Ltd,
Molecular Ecology
, 8, 1805–1817
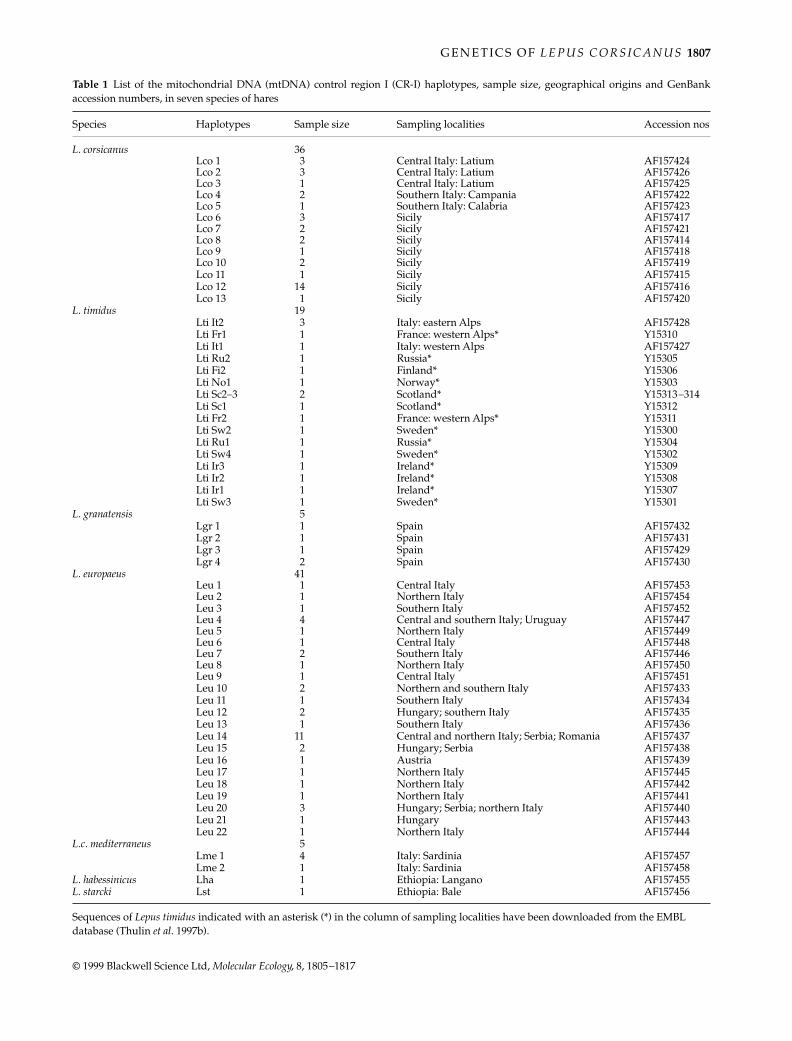
Table 1
List of the mitochondrial DNA (mtDNA) control region I (CR-I) haplotypes, sample size, geographical origins and GenBankaccession numbers, in seven species of hares
Species Haplotypes Sample size Sampling localities Accession nos
L. corsicanus
36Lco 1 3 Central Italy: Latium AF157424Lco 2 3 Central Italy: Latium AF157426Lco 3 1 Central Italy: Latium AF157425Lco 4 2 Southern Italy: Campania AF157422Lco 5 1 Southern Italy: Calabria AF157423Lco 6 3 Sicily AF157417Lco 7 2 Sicily AF157421Lco 8 2 Sicily AF157414Lco 9 1 Sicily AF157418Lco 10 2 Sicily AF157419Lco 11 1 Sicily AF157415Lco 12 14 Sicily AF157416Lco 13 1 Sicily AF157420
L. timidus
19Lti It2 3 Italy: eastern Alps AF157428Lti Fr1 1 France: western Alps* Y15310Lti It1 1 Italy: western Alps AF157427Lti Ru2 1 Russia* Y15305Lti Fi2 1 Finland* Y15306Lti No1 1 Norway* Y15303Lti Sc2–3 2 Scotland* Y15313–314Lti Sc1 1 Scotland* Y15312Lti Fr2 1 France: western Alps* Y15311Lti Sw2 1 Sweden* Y15300Lti Ru1 1 Russia* Y15304Lti Sw4 1 Sweden* Y15302Lti Ir3 1 Ireland* Y15309Lti Ir2 1 Ireland* Y15308Lti Ir1 1 Ireland* Y15307Lti Sw3 1 Sweden* Y15301
L. granatensis
5Lgr 1 1 Spain AF157432Lgr 2 1 Spain AF157431Lgr 3 1 Spain AF157429Lgr 4 2 Spain AF157430
L. europaeus
41Leu 1 1 Central Italy AF157453Leu 2 1 Northern Italy AF157454Leu 3 1 Southern Italy AF157452Leu 4 4 Central and southern Italy; Uruguay AF157447Leu 5 1 Northern Italy AF157449Leu 6 1 Central Italy AF157448Leu 7 2 Southern Italy AF157446Leu 8 1 Northern Italy AF157450Leu 9 1 Central Italy AF157451Leu 10 2 Northern and southern Italy AF157433Leu 11 1 Southern Italy AF157434Leu 12 2 Hungary; southern Italy AF157435Leu 13 1 Southern Italy AF157436Leu 14 11 Central and northern Italy; Serbia; Romania AF157437Leu 15 2 Hungary; Serbia AF157438Leu 16 1 Austria AF157439Leu 17 1 Northern Italy AF157445Leu 18 1 Northern Italy AF157442Leu 19 1 Northern Italy AF157441Leu 20 3 Hungary; Serbia; northern Italy AF157440Leu 21 1 Hungary AF157443Leu 22 1 Northern Italy AF157444
L.c. mediterraneus
5Lme 1 4 Italy: Sardinia AF157457Lme 2 1 Italy: Sardinia AF157458
L. habessinicus
Lha 1 Ethiopia: Langano AF157455
L. starcki
Lst 1 Ethiopia: Bale AF157456
Sequences of
Lepus timidus
indicated with an asterisk (*) in the column of sampling localities have been downloaded from the EMBL database (Thulin
et al
. 1997b).
MEC766.fm Page 1807 Friday, October 29, 1999 10:55 AM

1808
M . P I E R P A O L I
E T A L
.
© 1999 Blackwell Science Ltd,
Molecular Ecology
, 8, 1805–1817
classified as brown hares (
L. europaeus
) or Italian hares(
L. corsicanus
) according to external body size measure-ments, pelage colour patterns, and skull and dentaltraits, as described by Palacios (1996) and based on newunivariate and multivariate analyses (Riga
et al
. 1998).Additional samples of European brown hares introducedto South America were obtained from Uruguay. More-over, Mediterranean hares (
L. capensis mediterraneus
) werecollected in Sardinia, mountain hares (
L. timidus
) in theItalian Alps, and east African
L. starcki
and
L. habessinicus
in Ethiopia. Spanish
L. granatensis
was obtained from alocal breeder. Species distributions and sampling localitiesin Italy are shown in Fig. 1.
DNA amplification and sequencing
Total DNA was extracted from blood samples stored ina Tris–SDS buffer (Longmire
et al
. 1988), or from othertissues (liver, heart) stored in 95% ethanol, using guani-dinium thiocyanate and diatomaceous silica particles(Gerloff
et al
. 1995). mtDNA fragments, including theentire CR-I, were polymerase chain reaction (PCR)amplified in all the samples using primers LepCyb2L(5
′
-GAAACTGGCTCCAATAACCC-3
′
) and LepD2H(5
′
-ATTTAAGAGGAACGTGTGGG-3
′
), the 5
′
terminalnucleotides of which bind, respectively, to positions14 788 (towards the second half of the CYB) and 15 998 (ina central conserved region of the CR) of the rabbitmitochondrial genome (Gissi
et al
. 1998). These primersare complementary to previously published mtDNAsequences of
L. europaeus
(Casane
et al
. 1997), and weredesigned specifically to prevent the amplification of bothshort and long tandem repeats that map upstream of theorigin of the H-strand in
Lepus
and other lagomorphs(Casane
et al
. 1997; Gissi
et al
. 1998), and thus will avoidthe amplification of possible heteroplasmic regions. CYBwas amplified in two samples for each species, with theexception of
L. habessinicus
and
L. starcki
, using primersOcuCybP12 (5
′
-AACCACTCTCTAATCGACCTTCCC-3
′
)and OcuCybM1 (5
′
-TGGGCCTCTTCTCTGATGTCCC-3
′
),which map at positions 14 217 and 15 374 of the rabbitmtDNA (Gissi
et al
. 1998), respectively.The PCR products obtained using these primers were
single bands, showed no evidence of heteroplasmy andwere identical in sequence to DNAs extracted from the sameindividuals using tissues with high (i.e. liver) or low (i.e.blood) mitochondrial density. All nucleotide sequenceswere very clean, with no double or ambiguous bands, andwere easily aligned with homologous published mtDNAsequences of lagomorphs. Therefore, we were confidentthat these sequences represented true mitochondrial genesand not nuclear copies (Zhang & Hewitt 1996).
Amplifications were performed in a Perkin-Elmer9600 GeneAmp cycler using the following thermal profile:
a first denaturating step at 94
°
C for 2 min; 30 cyclesat 94
°
C for 15 s, 50–55
°
C (depending on the optimalannealing temperature of the primers) for 15 s and 72
°
Cfor 30 s; and a final extension step at 72
°
C for 10 min. Weused
≈
30–50 ng of the DNA extract and 2 m
m
MgCl
2
in a total volume of 10
µ
L for each PCR amplification.Negative controls were used in all the PCR amplificationexperiments. PCR products were purified by digestingsingle-stranded primers and excess nucleotides with exo-nuclease (0.5 units/
µ
L) and shrimp alkaline phosphatase(0.1 units/
µ
L).Double-stranded DNA cycle sequencing was performed
using the ABI Prism Dye Terminator procedure in an ABI373 automatic sequencer, using CR primers LepD2H orLepD1H (5
′
-GGACAAGAAAGGATTTGACTCGG-3
′
, whichbind at position 15 804), and CYB primers OcuCybP12,OcuCybM1 and OcuCybP2 (5
′
-CCTAGAAACCTGAAA-CATTGGC-3
′
, which bind at position 14 508). Moreover,we downloaded, from GenBank, 16 CR sequences of
L. timidus
(accession nos Y15299–Y15315; Thulin
et al
.1997b). The new mtDNA CR and CYB sequences used inthis study have been deposited in GenBank with acces-sion nos AF157414–AF157467 (see Table 1).
Sequence analysis
Sequences were automatically aligned using
clustal w
(Thompson
et al
. 1994). Pairwise genetic distances wereestimated using the model of Tamura & Nei (TN93),which is appropriate for describing the pattern of evo-lution of CR-I sequences (Tamura & Nei 1993). Phylo-genetic trees were reconstructed using distance matrixprograms, that is, the neighbour-joining procedure (NJ;Saitou & Nei 1987) as implemented in
mega
(Kumar
et al
. 1993); programs
neighbour
and
fitch
in
phylip
(Felsenstein 1993), used with randomized imput orderof pairwise distances and global branch-swappingoptimization; and quartet-puzzling maximum likelihood(ML, with the Hasegawa–Kishino–Yang model HKY85;Hasegawa
et al
. 1985) and unweighted maximum parsi-mony (MP) methods, as implemented in
puzzle
(Strimmer& von Haeseler 1996) and
paup
(Swofford 1993), respect-ively. Support for the internodes in the phylogenetic treeswas assessed by bootstrap percentages (BP) after 1000resampling steps (Felsenstein 1985) with
mega
or
paup
,or reliability percentage (RP) after 25 000 puzzling stepswith
puzzle
. Moreover, we computed the decay index(DI; Bremer 1988), i.e. the number of extra steps requiredto disrupt the monophyly of each clade, using the com-puter program
autodecay
(Eriksson 1998). Phylogenetictrees were rooted using the CR and CYB sequences ofthe rabbit
Oryctolagus cuniculus
. However,
Lepus
hasno closely related extant outgroups and
Oryctolagus
issufficiently divergent to destabilize ingroup topologies
MEC766.fm Page 1808 Friday, October 29, 1999 10:55 AM

G E N E T I C S O F
L E P U S C O R S I C A N U S
1809
© 1999 Blackwell Science Ltd,
Molecular Ecology
, 8, 1805–1817
(Halanych
et al
. 1999). Therefore, we also used the mid-point rooting procedure (Swofford 1993). Heterogeneityof rates of molecular evolution in the different lineageswere tested by comparing the log-likelihood (
lnL
) of thetrees computed with and without the assumption of amolecular clock (Felsenstein 1988).
Values of DNA polymorphism within populations,i.e. haplotype diversity (
h
), nucleotide diversity (
π
) andmean number of pairwise differences (
k
) (Nei 1987), wereestimated using
dnasp
(Rozas & Rozas 1997). Significanceof the partition of genetic diversity among conspecificpopulations sampled in different geographical areas wastested using the analysis of molecular variance (
amova
)procedure (Excoffier
et al
. 1992) as implemented in
arlequin
(Schneider
et al
. 1997). Molecular divergencewas obtained by the observed number of pairwise sub-stitutions between CR-I haplotypes, and the significanceof between-population variance (
F
ST
) was determined byrandomization analysis using 10 000 permutations.
The observed nucleotide diversity (
π
) should corre-spond to the parameter:
θ
= 2
N
ef
µ
(where
N
ef
is the effective maternal population size and
µ
is the mutation rate per site per generation) if popu-lations are in demographic/genetic equilibrium and if thestudied nucleotide sequences are not selected (Nei 1987;Kuhner
et al
. 1998). Therefore, we estimated
θ
either usingthe number of segregating sites(s) and assuming stablepopulation size and genetic equilibrium (Watterson 1975),or from the genealogical relationships among haplotypesand allowing for historically fluctuating population sizeby the coalescent Metropolis-Hastings Markov Chainmethod (MHMC) as implemented in
fluctuate
(Kuhner
et al
. 1998). Assuming that
µ
is constant for CR-I indifferent populations and species, then
θ is proportionalto the historical maternal effective population size. Theassumption of neutrality of mutations was confirmed bythe test of Tajima (Tajima 1989) using dnasp.
The distribution of pairwise genetic differencesbetween individual haplotypes within a population(mismatch distributions) was analysed using the suddenexpansion model of Rogers & Harpending (1992) asimplemented in dnasp. The shape of a mismatch dis-tribution is affected by historical demographic changes ofthe population: recent sudden expansions or bottlenecksgenerate unimodal mismatch distributions, while stableor slowly declining populations produce a variety ofmultimodal distributions. In the model of Rogers &Harpending (1992), a population at equilibrium withinitial effective size N0 suddenly changes its size to N1 atτ units of mutational time in the past, and the processgenerates a mismatch distribution which is described by:
θ0 = 2N0u; θ1 = 2N1u; and τ = 2ut
where u is the mutation rate per generation of thenucleotide sequence studied.
Results
Sequence variability and phylogenetic relationships in Lepus
We sequenced 293–295 nucleotides of the mtDNACR-I, which included 127 (43%) variable and 101 (34%)parsimony-informative sites in 108 samples of sevenspecies of hare. The alignment, which can be accessed atftp://ftp.ebi.ac.uk/pub/databases/embl/align, no. DS38811,was obtained by introducing 10 insertions/deletions(indels) of single nucleotides and identified 57 differenthaplotypes (13 in L. corsicanus, 22 in L. europaeus, 16 inL. timidus, four in L. granatensis and two in L.c. medi-terraneus), plus L. habessinicus and L. starcki, which werenot shared between the different species. In particular,Italian and brown hares had different haplotypes thatdiverged by an average TN93 genetic distance of 0.151(Table 2). CR-I genetic distances among the other speciesranged from 0.020 (in pairwise comparison of L.
Table 2 Interspecific genetic distances (Tamura & Nei 1993) computed using cytochrome b (CYB) (above the diagonal) and control regionI (CR-I) (below the diagonal) sequences in seven species of hares
Species L. corsicanus L. timidus L. granatensis L. europaeus L. c. mediterraneus L. habessinicus
L. corsicanus — 0.033 0.111 0.120 0.127 —L. timidus 0.102 — 0.108 0.118 0.110 —L. granatensis 0.145 0.140 — 0.097 0.120 —L. europaeus 0.151 0.155 0.171 — 0.111 —L.c. mediterraneus 0.160 0.172 0.168 0.119 — —L. habessinicus 0.180 0.180 0.188 0.118 0.180 —L. starcki 0.170 0.180 0.177 0.117 0.170 0.020
CYB in Lepus habessinicus and L. starcki could not be amplified using the primers listed in the Materials and methods.
MEC766.fm Page 1809 Friday, October 29, 1999 10:55 AM
1810 M . P I E R P A O L I E T A L .
© 1999 Blackwell Science Ltd, Molecular Ecology, 8, 1805–1817
habessinicus and L. starcki) to 0.188 (between L.habessinicus and L. granatensis). Interspecific CYB geneticdistances were ≈ 25% lower than the correspondingCR-I distances (Table 2). Values of intraspecific CR-Isequence divergence, estimated as π (Tajima 1983), were0.015, 0.013 and 0.057 in L. corsicanus, L. europaeus andL. timidus, respectively (Table 3), i.e. they were 50–70%lower than their interspecific genetic distances (Table 2).
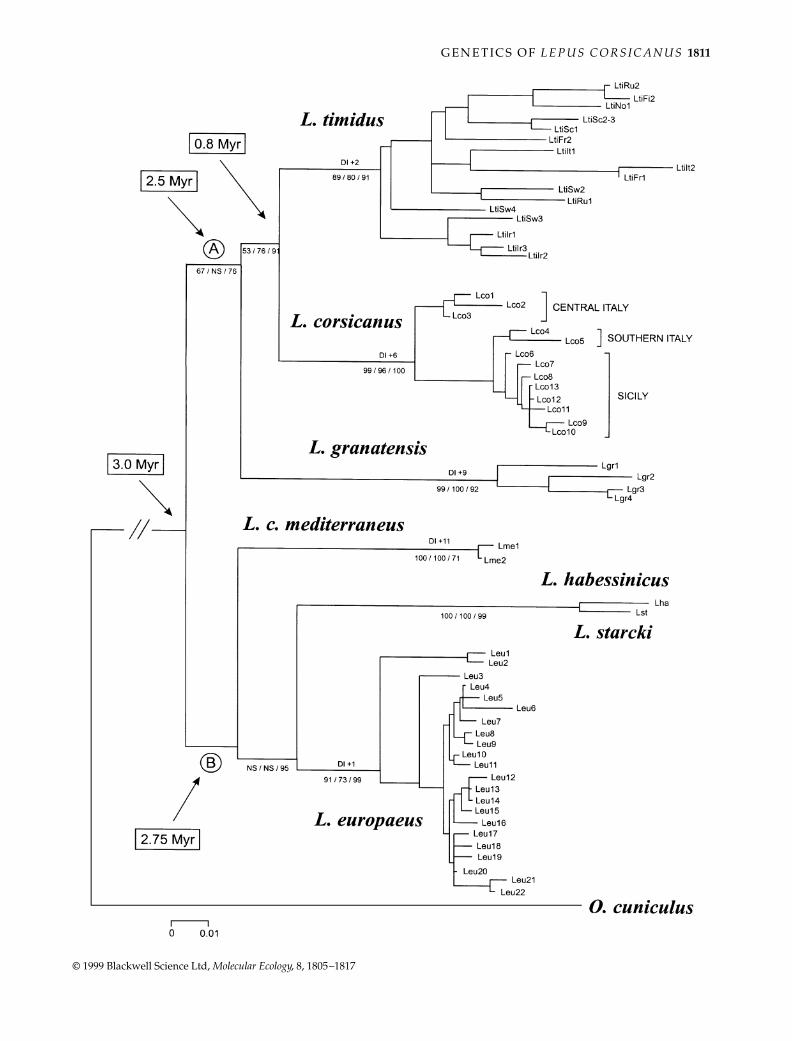
The phylogenetic trees obtained using NJ, MP and MLanalyses showed concordant topologies (a rooted NJ treeis shown in Fig. 2). Trees rooted using the rabbit CR-Isequence as an outgroup, or by midpoint rooting the long-est internal branch, concordantly showed a main geneticgap splitting the studied taxa into two groups: the first(clade A) included sister taxa L. corsicanus and L. timidus,with L. granatensis in basal position; the second (clade B)joined L. europaeus and the two closely related Africantaxa L. habessinicus and L. starcki, with L.c. mediterraneusin basal position (bootstrap values were lower than 50%because the position of L.c. mediterraneus was unstableand shifted to form a basal polytomy in the rooted trees).Randomizing the input order of pairwise distances hadno effect on the optimized topologies computed usingthe programs neighbour and fitch, which were identicalto the tree shown in Fig. 2. paup produced more than100 MP trees with identical length (L = 361), consistencyindex (CI = 0.557) and retention index (RI = 0.860), whichdepicted different, but parsimony equivalent, relationshipsamong CR-I haplotypes within species. The next multipleMP trees (> 100) had L = 362–363, that is they were one totwo steps longer than the most-parsimonious solutions.
Also, the next most-parsimonious trees produced onlyminimal rearrangements of interspecific haplotype relation-ships and preserved the monophyly of all the main clades,as represented in Fig. 2. Values of the DI indicated that sixadditional steps were necessary to disrupt the monophylyof L. corsicanus (Fig. 2).
CR-I sequences evolved at similar rates among hareswhen trees were computed excluding the rabbit sequence(lnL ratio test = 61.47, degrees of freedom (d.f.) 57,P = 31.92%). In contrast, inclusion of the rabbit sequencesintroduced significant heterogeneity (lnL ratio test = 95.78,d.f. = 58, P = 0.13%). However, including the outgroup didnot change the topological relationships among the ingroupsequences, although it reduced some of the bootstrap values.
CR-I sequences of L. corsicanus, L. timidus and L.granatensis grouped into three different and stronglysupported monophyletic clades (with bootstrap values> 80%, Fig. 2), which were concordant with morpho-logical classifications. L.c. mediterraneus, L. europaeus, L.habessinicus and L. starcki grouped into three stronglysupported monophyletic clades (with bootstrap values> 71%, Fig. 2), which were concordant with morphologicalclassifications.
Genetic variability within and among populations of Italian, brown and mountain hares
Mountain hares showed approximately one differentCR-I haplotype per individual (i.e. 16 haplotypes over20 samples), while brown and Italian hares showed onehaplotype per two and three individuals, respectively
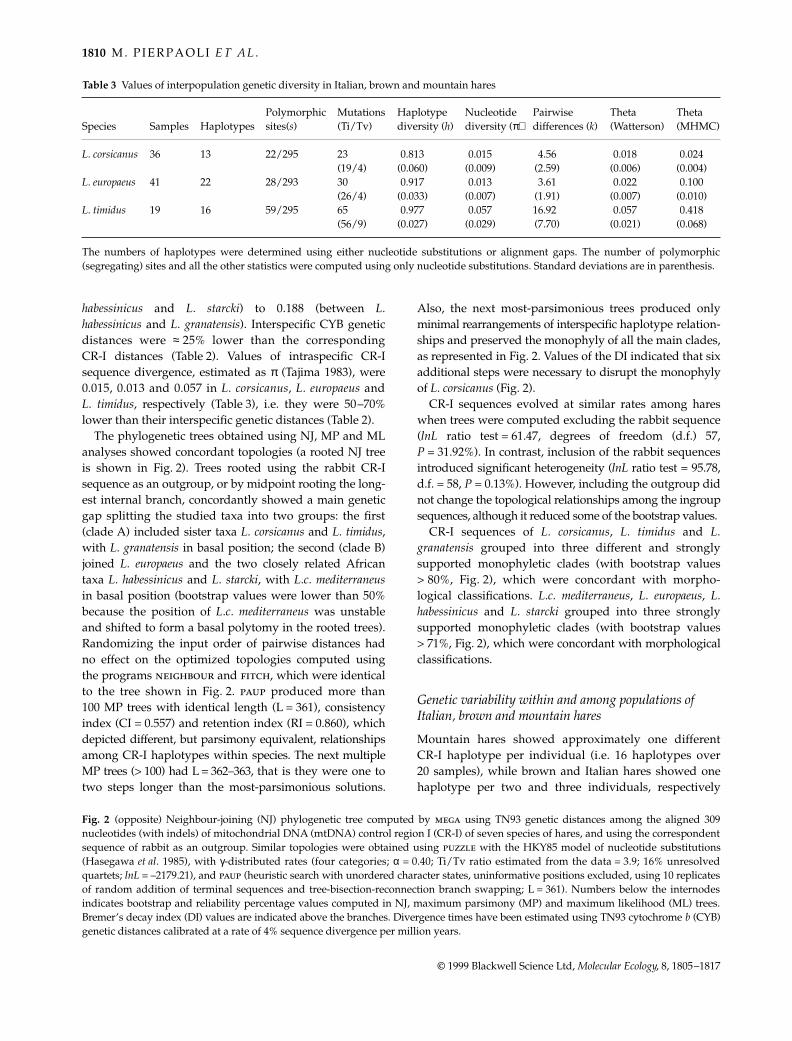
Table 3 Values of interpopulation genetic diversity in Italian, brown and mountain hares
Species Samples HaplotypesPolymorphic sites(s)
Mutations (Ti/Tv)
Haplotype diversity (h)
Nucleotide diversity (π)
Pairwise differences (k)
Theta (Watterson)
Theta (MHMC)
L. corsicanus 36 13 22/295 23 0.813 0.015 4.56 0.018 0.024(19/4) (0.060) (0.009) (2.59) (0.006) (0.004)
L. europaeus 41 22 28/293 30 0.917 0.013 3.61 0.022 0.100(26/4) (0.033) (0.007) (1.91) (0.007) (0.010)
L. timidus 19 16 59/295 65 0.977 0.057 16.92 0.057 0.418(56/9) (0.027) (0.029) (7.70) (0.021) (0.068)
The numbers of haplotypes were determined using either nucleotide substitutions or alignment gaps. The number of polymorphic(segregating) sites and all the other statistics were computed using only nucleotide substitutions. Standard deviations are in parenthesis.
Fig. 2 (opposite) Neighbour-joining (NJ) phylogenetic tree computed by mega using TN93 genetic distances among the aligned 309nucleotides (with indels) of mitochondrial DNA (mtDNA) control region I (CR-I) of seven species of hares, and using the correspondentsequence of rabbit as an outgroup. Similar topologies were obtained using puzzle with the HKY85 model of nucleotide substitutions(Hasegawa et al. 1985), with γ-distributed rates (four categories; α = 0.40; Ti/Tv ratio estimated from the data = 3.9; 16% unresolvedquartets; lnL = –2179.21), and paup (heuristic search with unordered character states, uninformative positions excluded, using 10 replicatesof random addition of terminal sequences and tree-bisection-reconnection branch swapping; L = 361). Numbers below the internodesindicates bootstrap and reliability percentage values computed in NJ, maximum parsimony (MP) and maximum likelihood (ML) trees.Bremer’s decay index (DI) values are indicated above the branches. Divergence times have been estimated using TN93 cytochrome b (CYB)genetic distances calibrated at a rate of 4% sequence divergence per million years.
MEC766.fm Page 1810 Friday, October 29, 1999 10:55 AM
G E N E T I C S O F L E P U S C O R S I C A N U S 1811
© 1999 Blackwell Science Ltd, Molecular Ecology, 8, 1805–1817
MEC766.fm Page 1811 Friday, October 29, 1999 10:55 AM

1812 M . P I E R P A O L I E T A L .
© 1999 Blackwell Science Ltd, Molecular Ecology, 8, 1805–1817
(Table 3). The number of polymorphic sites was similar inItalian and brown hares (s = 22 and 28), and it was ≈ twotimes greater in mountain hares (s = 59). Accordingly, moun-tain hares had the highest values of h, π and k (Table 3).Watterson’s and MHMC estimates of θ were three to 20times higher in mountain hares than in Italian and brownhares, suggesting either a higher mutation rate or largerhistorical population size in L. timidus (Table 3). However,CR-I sequences evolved at constant rates among hares,and Tajima’s neutrality tests were not significant. It istherefore probable that genetic variability within and amongpopulations has been affected mainly by divergencetimes and different historical demographic trajectories,and not by heterogeneous patterns and rates of substitution.
Genealogical relationships among CR-I sequences andgeographical locations of the Italian hare samples werestrictly concordant (Figs 1, 2). Hares from central Italyshowed three unique haplotypes (Lco1, -2 and -3) thatclustered apart from haplotypes of the hares sampledin southern Italy (Lco4 and -5) and Sicily (Lco6 to -13;Fig. 2). The amova test indicated significant pairwise andtotal FST values (total FST = 0.80; P < 0.001), and consequentabsence of gene flow (Nm < 0.3), among the Italian haressampled in central Italy (Latium; n = 7), southern Italy(Campania and Calabria; n = 3) and Sicily (n = 26).
Three divergent brown hare haplotypes (Leu1, Leu2and Leu3), which grouped separately from the others(Fig. 2), were found in samples collected in isolated moun-tain areas in northern (Mount Cusna), central (AbruzzoNational Park) and southern Italy (Mount Pollino)(Fig. 1). The other 38 brown hares showed 19 closely relatedhaplotypes, which congregated in a single cluster withstar-like topology and did not show any obvious correla-tion with the sampling areas. For example, the mostcommon haplotype, Leu14, was shared by 11 hares sampledfrom three localities in central and northern Italy, Romaniaand Serbia (Table 1).
Average sequence divergence between mountain harehaplotypes was ≈ four times greater than among brownhare haplotypes, suggesting an ancient origin of moun-tain hare mtDNA lineages. However, genealogical rela-tionships did not indicate any correlation of CR-I lineagesof mountain hares and particular geographical locations,perhaps with the exception of closely related haplotypesfrom Ireland and Scotland (Fig. 2). Therefore, both brownand mountain hares had shallow geographical populationstructure, either in Italy and Europe, in accordance withother published findings (Hartl et al. 1993; Thulin et al.1997b; Suckentrunk et al. 1998).
Mismatch distributions and inference on ancient population dynamics
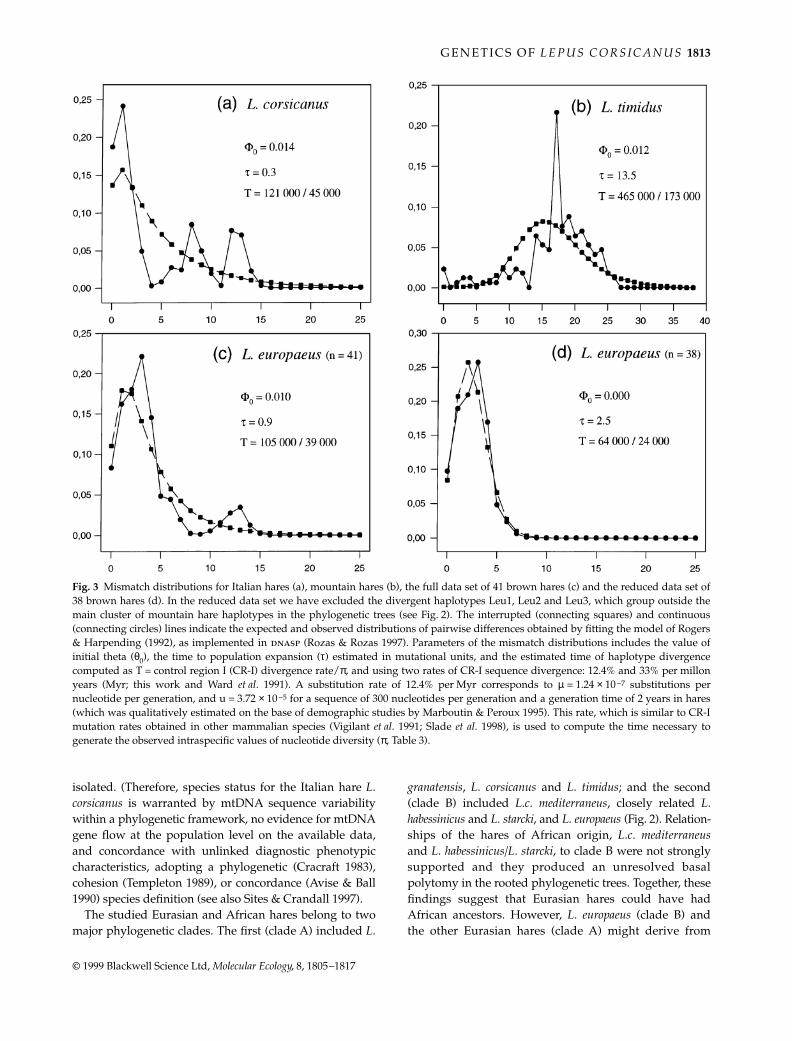
The observed mismatch distribution in Italian hares was
multimodal, with three major peaks determined by thedivergent haplotypes from central Italy, southern Italyand Sicily, and did not fit the distribution expected aftera sudden population growth (Fig. 3a). The estimated θ-value in the initial population was similar to the valuesfor the present-day populations (θ0 = 0.014; Watterson’sθ = 0.018; MHMC θ = 0.024; Table 3 and Fig. 3a) andτ = 0.3 was very small (Fig. 3a). Watterson’s and MHMCθ-values were very similar (Table 3), suggesting thatItalian hares had, historically, a stable population size.Therefore, mismatch distibution in Italian hares wasmainly affected by historical fragmentation and phylo-geographical structuring, and not by fluctuating popula-tion size.
The mismatch distribution in brown hares showed aminor peak owing to the three divergent haplotypes:Leu1, Leu2 and Leu3 (Fig. 3c). If these haplotypes wereremoved, the mismatch distribution of the reduced dataset of the other 38 haplotypes became unimodal and cor-responded to the expected distribution generated by asudden population change at τ = 2.5 (Fig. 3d). Using thedata from the complete brown hare data set, MHMC θwas ≈ five times greater than Watterson’s θ (i.e. 0.100 vs.0.022; Table. 2), and θ0 was 0.010, suggesting an ≈ 10 timesexpansion of the population size. Analysing the alignmentof 38 haplotypes, MHMC θ was 0.190 and Watterson’sθ was 0.014, i.e. ≈ 10 times smaller, but θ0 was 0, thusleaving undetermined the dimension of population sizeexpansion (Fig. 3d).
The mismatch distribution in mountain hares wasunimodal, with a major peak at τ = 13.5, as expected aftera sudden population change (Fig. 3b). Accordingly, theθ-values indicated a strong expansion of populationsize (θ0 = 0.012; Watterson’s θ = 0.057; MHMC θ = 0.418;Table 3 and Fig. 3b).
Discussion
Species distinction and phylogenetic relationships of Lepus corsicanus
Genetic analyses indicate sharp phylogenetic distinctionamong the mtDNA sequences of L. corsicanus and theother taxa included in this study. CR-I haplotypes ofItalian hares group in a strongly supported monophyleticclade (Fig. 2), and DNA distinction is concordant withboth qualitative and quantitative morphological clas-sification Riga et al. 1998). Moreover, Italian and brownhares collected from areas of sympatry and close syntopy,which were created by restocking, do not share anymtDNA haplotypes (Table 1 and Fig. 3), thus suggestingthe absence of either past and ongoing interspecific geneflow. Italian and brown hares have a long-lasting historyof independent evolution and seem to be reproductively
MEC766.fm Page 1812 Friday, October 29, 1999 10:55 AM
G E N E T I C S O F L E P U S C O R S I C A N U S 1813
© 1999 Blackwell Science Ltd, Molecular Ecology, 8, 1805–1817
isolated. (Therefore, species status for the Italian hare L.corsicanus is warranted by mtDNA sequence variabilitywithin a phylogenetic framework, no evidence for mtDNAgene flow at the population level on the available data,and concordance with unlinked diagnostic phenotypiccharacteristics, adopting a phylogenetic (Cracraft 1983),cohesion (Templeton 1989), or concordance (Avise & Ball1990) species definition (see also Sites & Crandall 1997).
The studied Eurasian and African hares belong to twomajor phylogenetic clades. The first (clade A) included L.
granatensis, L. corsicanus and L. timidus; and the second(clade B) included L.c. mediterraneus, closely related L.habessinicus and L. starcki, and L. europaeus (Fig. 2). Relation-ships of the hares of African origin, L.c. mediterraneusand L. habessinicus/L. starcki, to clade B were not stronglysupported and they produced an unresolved basalpolytomy in the rooted phylogenetic trees. Together, thesefindings suggest that Eurasian hares could have hadAfrican ancestors. However, L. europaeus (clade B) andthe other Eurasian hares (clade A) might derive from
Fig. 3 Mismatch distributions for Italian hares (a), mountain hares (b), the full data set of 41 brown hares (c) and the reduced data set of38 brown hares (d). In the reduced data set we have excluded the divergent haplotypes Leu1, Leu2 and Leu3, which group outside themain cluster of mountain hare haplotypes in the phylogenetic trees (see Fig. 2). The interrupted (connecting squares) and continuous(connecting circles) lines indicate the expected and observed distributions of pairwise differences obtained by fitting the model of Rogers& Harpending (1992), as implemented in dnasp (Rozas & Rozas 1997). Parameters of the mismatch distributions includes the value ofinitial theta (θ0), the time to population expansion (τ) estimated in mutational units, and the estimated time of haplotype divergencecomputed as T = control region I (CR-I) divergence rate/π, and using two rates of CR-I sequence divergence: 12.4% and 33% per millonyears (Myr; this work and Ward et al. 1991). A substitution rate of 12.4% per Myr corresponds to µ = 1.24 × 10−7 substitutions pernucleotide per generation, and u = 3.72 × 10−5 for a sequence of 300 nucleotides per generation and a generation time of 2 years in hares(which was qualitatively estimated on the base of demographic studies by Marboutin & Peroux 1995). This rate, which is similar to CR-Imutation rates obtained in other mammalian species (Vigilant et al. 1991; Slade et al. 1998), is used to compute the time necessary togenerate the observed intraspecific values of nucleotide diversity (π, Table 3).
MEC766.fm Page 1813 Friday, October 29, 1999 10:55 AM

1814 M . P I E R P A O L I E T A L .
© 1999 Blackwell Science Ltd, Molecular Ecology, 8, 1805–1817
different and independent evolutionary lineages. Thus, amore comprehensive taxon sampling would be necessaryto identify the ancestors of species belonging to clades Aand B, and also of the Mediterranean hare. Moreover, thefossil record and mtDNA sequence data (Halanych et al.,in press) suggest that Lepus originated in North Americaand dispersed repeatedly to Eurasia. Therefore, improvedreconstruction of phylogenetic relationships in Lepusshould include specimens of both Eurasian and NorthAmerican hares.
Our results, rather surprisingly, indicate that the closestrelative of the Italian hare is the mountain hare and notthe European hare, as previously suggested (Miller 1912;Toschi 1965; amongst others). Although further data arerequired to improve the phylogeny of hares, divergencedates (estimated through molecular data) and the inferredbiogeographical framework for the evolution of Lepus inEurope suggest that L. corsicanus and L. timidus are relictualspecies that originated before the dispersal of L. europaeusin western Europe, and which became adapted to thewidely different Mediterranean and Alpine habitats(discussed below).
Divergence dates and biogeographical framework for the evolution of European hares
Although molecular divergence is somehow correlatedwith divergence time, the calibration of evolutionaryrates is complicated by poorly known patterns of mtDNAsequence evolution (Tamura & Nei 1993) and uncertainfossil or biogeographical references. Therefore, divergencetimes inferred from molecular data are hypothetical andassociated with very large standard errors (Hillis et al. 1996).Thus, we suggest a tentative biogeographical framework forthe evolution of Lepus in Europe, which should be testedby analyses of enlarged data sets.
To obtain an estimate of interspecific divergence timein Lepus, we applied the standard calibration of CYBdivergence rate of 2–4% per million years (Myr) (Martin &Palumbi 1993; Avise et al. 1998; Santucci et al. 1998). Thebasal splitting among the phylogenetic lineages (Fig. 2) isat ≈ 12% CYB sequence divergence, corresponding to 3–6 Myr. The lower estimate, 3 Myr, corresponding to a rateof CYB divergence of 4% per Myr, is more concordantwith the available fossil record, which documents the firstappearence of Lepus in the Villafranchian (≈ 2.5 millionyears ago (Ma); Lopez Martinez 1980).
Most of the African and European species of harestudied showed pairwise CYB divergences of 10–13%.Therefore, these species could have originated duringthe last 3 Myr, i.e. in the second half of the Pliocene andat the Plio/Pleistocene boundary. The Plio/Pleistoceneboundary was marked by great climatic and palaeogeo-graphical changes, culminating with the start of major
glacial cycles ≈ 2.5 Ma (deMenocal 1995). Pleistocene climatevariations were more intense during the last 1 Myr, a periodthat was marked by four major glaciations, lasting≈ 100 000 years and interrupted by warmer and shorterinterglacials (deMenocal 1995). It is plausible that Plio/Pleistocenic climate changes fostered allopatric isolationin glacial refuge areas located in southern parts of theIberian, Italian and Balkan peninsulas (Taberlet et al. 1998),and mediated speciation through repeated cycles of popu-lation contraction and expansion in Europe and Africa(deMenocal 1995; Hewitt 1996). Many mammalian andavian species could have originated, during those periods(Avise et al. 1998), in North America, Eurasia and aroundthe Mediterranean basin (e.g. the Alectoris partridges,Randi 1996; Erinaceus hedgehogs, Santucci et al. 1998).This calibration suggests that closely related L. timidus/corsicanus, which split at 3.3% CYB sequence divergence,might have originated ≈ 820 000 years ago (other inter-specific divergence times estimated through CYB calibra-tion are indicated in Fig. 3).
In this perspective it is tempting to speculate that thethree species of hares of clade A represent relictual taxaoriginating from a common ancestor during an earlyphase of speciation and dispersal of Lepus in westernEurope. These species could have been confined to mar-ginal distributions as a consequence of either habitatchanges or comparatively recent dispersal of a strongcompetitor, L. europaeus, in western Europe. L. granatensisand L. corsicanus have localized distributions in Spain,southern Italy and Sicily (Palacios 1996), i.e. in areas thathave been repeatedly indicated as glacial refuges forplant and animal communities (Hewitt 1996; Taberletet al. 1998). The mountain hare has a circumpolar distribu-tion and occurs in isolated populations in Ireland,Scotland and in the Alps. However, the fossil recorddocuments a much more widespread distribution of themountain hare during the last glacial period throughoutmost of central Europe and the UK (Lopez Martinez1980). Upper Pleistocene fossil mountain hares have beendiscovered, associated with Oryctolagus, in sites that werepresumably characterized by a warm climate (LopezMartinez 1980). Thus, mountain hares are not necessarilyadapted to cold climates, but they might have retreated tonorthern latitudes or isolated mountain ranges to escapeinterspecific competition with other lagomorphs, andnotably the brown hare (Lopez Martinez 1980).
The mtDNA phylogeny suggests that brown haresoriginated from an African ancestor and dispersed inEurope, perhaps recently and in at least two differentwaves. Thus, the three distinct and phylogenetically basalCR-I haplotypes, which have been found in brown haresin Italy (Fig. 2), could represent the remains of an earlycolonization of the brown hare in southwestern Europe,and they probably testify to the presence of an ancestral
MEC766.fm Page 1814 Friday, October 29, 1999 10:55 AM

G E N E T I C S O F L E P U S C O R S I C A N U S 1815
© 1999 Blackwell Science Ltd, Molecular Ecology, 8, 1805–1817
population, isolated in Italy during the last glaciation(subspecies meridiei). The presence of one of these ancienthaplotypes (Leu3) in southern Italy, outside the presumednatural range of the brown hare in north-central Italy(Fig. 1), could be the consequence of recent translocations,or it could represent a relict of an ancestral and wide-spread distribution of the brown hare, running along theApennine divide at higher altitudes. On the contrary, mostof the other haplotypes are closely related to each otherand do not show any phylogeographical structure. Theycould represent the result of a very recent populationexpansion and colonization of brown hares in Europe,which occurred probably after the last glacial maximum(18 000–20 000 years ago).
Inference on ancient population dynamics of Italian, brown and mountain hares
Hares in the Northern Hemisphere speciated and divergedduring glacial/interglacial cycles, perhaps in associationwith, and as a consequence of, dramatic fluctuations ofpopulation ranges and sizes. Therefore, demographicfactors could have markedly affected the distributionof mtDNA diversity within and between populations. Infact, mismatch and coalescent analyses of sequencedifferences and genealogical relationships among CR-Ihaplotypes suggest that Italian, brown and mountain hareshad different demographic histories (Fig. 3).
Mountain hares and the reduced data set of brownhares show evidence of sudden population expansionsfrom very low to relatively large present-day popula-tions. The value of initial θ (before the expansion) is 0.012in the mountain hare, and it is 0.418 (MHMC estimate)in contemporary populations (Table 3 and Fig. 3), thussuggesting a 35-fold increase of Nef since the expansion, att = (τ/2 µ) = 135 000 years ago. The extent of populationexpansion cannot be assessed in the brown hare becauseθ0 = 0.0, but the shape of mismatch distribution (Fig. 3c)indicates a sudden expansion at t = 25 000 years. In contrast,multimodal mismatch distribution and similar valuesof initial and present-day θ, suggest that the Italian harehad a stable population size (Fig. 3a).
Mountain hares could have differentiated during thesecond half of the Pleistocene and expanded during thelast interglacial, i.e. before the last glaciation. Divergencetimes estimated by genetic data are concordant withthe widespread presence in Europe of fossil remains ofmountain hares during the last glaciation, and with thepresent-day relictual distribution of this species. Accord-ingly, high gene flow among mountain hares in continentalEurope since the recent Holocene period prevented theaccumulation of genetic differences among present-dayfragmented populations (Thulin et al. 1997b; Suckentrunket al. 1998; this work).
Haplotype divergence in L. corsicanus was between45 000 and 121 000 years old, suggesting an ancient isola-tion in central and southern Italian glacial refuge areas.Colonization of Sicily could have been possible through-out this period because the level of the Mediterranean seawas ≈ 110 m lower than present and Sicily was connectedto the mainland. Moreover, Italian hares show significantgenetic structuring in terms of geographical distributionand sequence divergence of their CR-I haplotypes (Fig. 2).The haplotypes sampled in central Italy (Latium) areunique, group in a monophyletic clade and are notpresent in the hares sampled in southern Italy (Campaniaand Calabria) and Sicily. The separation of Sicily, since theend of the last glaciation, can explain the observed diver-gence between Sicilian and central Italian hares.
Conservation biology of L. corsicanus in Italy
It is worth noting that neither L. corsicanus nor L.c.mediterraneus were mentioned as distinct taxa in theRabbit, Hares and Pikas Status Survey and ConservationAction Plan of the International Union for the Conser-vation of Nature (Chapman & Flux 1990). Taxonomicalconfusion has contributed to worsen the many conserva-tion problems generated by overhunting and extensiverelease, in peninsular Italy, of brown hares of centralEuropean origins. Present-day distribution of the Italianhare is extremely fragmented in central and southernItaly. Populations survive at low density, mainly inprotected areas and National Parks, where the species hasmanaged to escape overhunting and competition withintroduced brown hares. Although we have not detectedcases of mtDNA introgression among Italian and brownhares and, consequently, hybridization may not be athreat to the survival of L. corsicanus in central andsouthern Italy, strict prohibition of restocking with brownhares should be enforced, at least in selected priorityareas. Despite the intensive release of brown hares duringthe last 20 years, field surveys (Lo Valvo et al. 1997; Rigaet al. 1998) have shown that hares of Sicily belong only toL. corsicanus. It is therefore possible that the peculiar condi-tions of Sicily are unsuitable for the brown hare and that itcannot survive in the Mediterranean-type climate andhabitats of the island. Although Sicilian populations of L.corsicanus are rather widespread and stable, conservationmeasures should be enforced in order to prevent the releaseof brown hares in the island and to plan a controlledhunting and wise use of the Italian hare populations.
Acknowledgements
We thank officials and staff at the Italian Ministries of Agri-culture and the Environment, directors and staff at Nationaland Regional Parks and other protected areas, Regional and
MEC766.fm Page 1815 Friday, October 29, 1999 10:55 AM

1816 M . P I E R P A O L I E T A L .
© 1999 Blackwell Science Ltd, Molecular Ecology, 8, 1805–1817
Provincial Administrations, national and local Hunting Associ-ations, Natural History Museums, Universities and all the otherinstitutions, who enabled the organization of field surveys andsample collection. In particular we wish to thank A. Tinelli,C. Raja, R. De Battisti, F. Di Sisto, A. Zilio, R. Mazzoni dellaStella, F. Dalla Vecchia, I. Farronato, L. Zini, M. Ragusa, S. Poerio,S. Troisi, G. De Filippo, M. Macchio, G. Braschi and the manyother people who kindly collaborated for sample collection.M. Lo Valvo (University of Palermo) kindly shared with us hisexpertise on the biology of the Italian hare in Sicily. A. Scappiand A. De Faveri (INFS) collaborated with us for field surveys,sample collection and preservation of museum materials. ProfessorG. M. Hewitt and two anonymous referees contributed greatlyto improve on an early version of this paper. This study wassupported by the INFS.
References
Amori G, Angelici FM, Prigioni C, Vigna Taglianti A (1996) Themammal fauna of Italy: a review. Hystrix, 8, 3–7.
Angermann R (1983) The taxonomy of Old World Lepus. ActaZoologica Fennica, 174, 17–21.
Avise JC, Ball RM (1990) Principles of genealogical concordancein species concepts and biological taxonomy. Oxford Survey inEvolutionary Biology, 7, 45–65.
Avise JC, Arnold J, Ball RM Jr. et al. (1987) Intraspecific phylo-geography: the mitochondrial DNA bridge between populationgenetics and systematics. Annual Review of Ecology and System-atics, 18, 489–522.
Avise JC, Walker DE, Johns GC (1998) Speciation durations andPleistocene effects on vertebrate phylogeography. Proceedingsof the Royal Society Of London, B, 265, 1707–1712.
Bremer K (1988) The limits of amino acid sequences in angiospermphylogenetic reconstruction. Evolution, 42, 795–803.
Casane D, Dennebouy N, de Rochambeau H, Mounolou JC,Monnerot M (1997) Nonneutral evolution of tandem repeatsin the mitochondrial DNA control region of lagomorphs.Molecular Biology and Evolution, 14, 779–789.
Chapman JA, Flux JEC (1990) Rabbits, Hares and Pikas. StatusSurvey and Conservation Action Plan. IUCN/SSC LagomorphSpecialist Group, Glaud.
Cracraft J (1983) Species concept and speciation analysis. CurrentOrnithology, 1, 159–187.
Ellerman JR, Morrison-Scott TCS (1951) Checklist of Palaearcticand Indian mammals (1758–1946). British Museum (NaturalHistory), London, pp. 810.
Eriksson T (1998) autodecay, Version 4.0. Department ofBotany, Stockholm University. Program at http://www.botan.su.se/Systematik/Folk/Torsten.html.
Excoffier L, Smouse P, Quattro J (1992) Analysis of molecularvariance inferred from metric distances among DNA haplo-types: applications to human mitochondrial DNA restrictiondata. Genetics, 131, 479–491.
Felsenstein J (1985) Confidence limits on phylogenies: anapproach using the bootstrap. Evolution, 39, 783–791.
Felsenstein J (1988) Phylogenies from molecular sequences: infer-ences and reliability. Annual Review of Genetics, 22, 521–565.
Felsenstein J (1993) PHYLIP (phylogeny inference package), version3.5c. Department of Genetics, University of Washington, Seattle.
Flux JEC (1983) Introduction to taxonomic problems in hares.Acta Zoologica Fennica, 174, 7–10.
Gerloff U, Schlotterer C, Rassmann K et al. (1995) Amplificationof hypervariable simple sequence repeats (microsatellites)from excremental DNA of wild living Bonobos (Pan paniscus).Molecular Ecology, 4, 515–518.
Gissi C, Gullberg A, Arnason U (1998) The complete mito-chondrial DNA sequence of the rabbit, Oryctolagus cuniculus.Genomics, 50, 161–169.
Halanych KM, Demboski JR, Jansen Van Vuuren B, Klein DR,Cook JA (1999) Cytochrome b phylogeny of North Americanhares and jackrabbits (Lepus, Lagomorpha) and the effectsof saturation in outgroup taxa. Molecular Phylogenetics andEvolution, 11, 213–221.
Hartl GB, Suchentrunk F, Nadlinger K et al. (1993) An integ-rative analysis of genetic differentiation in the brown hare Lepuseuropaeus based on morphology, allozymes, and mitochon-drial DNA. Acta Theriologica, 38, 33–57.
Hasegawa M, Kishino H, Yano T (1985) Dating of the human–ape splitting by a molecular clock of mitochondrial DNA.Journal of Molecular Evolution, 22, 160–174.
Hewitt GM (1996) Some genetic consequences of ice ages, andtheir role in divergence and speciation. Biological Journal ofLinnean Society, 58, 247–279.
Hillis DM, Moritz C, Mable BK (1996) Applications of molecularsystematics: the state of the field and a look to the future.In: Molecular Systematics (eds Hillis DM, Moritz C, Mable BK),pp. 515–543. Sinauer Associates, Sunderland, MA.
Hoffmann RS (1993) Order Lagomorpha. In: Mammal Species ofthe World: a Taxonomic and Geographic Reference, 2nd edn. (edsWilson DE, Reeder DM), pp. 807–827. Smithsonian InstitutionPress, Washington.
Hoffmann RS (1998) The trouble with Lepus capensis. In: Euro-American Mammal Congress, Abstracts (ed. Reig S), p. 94. Uni-versidad de Santiago de Compostela, Spain, July 19–24, 1998.
Kuhner MK, Yamato J, Felsenstein J (1998) Maximum likelihoodestimation of population growth rates based on the coalescent.Genetics, 149, 429–434.
Kumar S, Tamura K, Nei M (1993) mega: molecular evolutionarygenetic analysis, version 1.0. Pennsylvania State University,University Park, PA.
Lo Valvo M, Barera A, Seminara S (1997) Biometria e status dellalepre appenninica (Lepus corsicanus, de Winton 1898) in Sicilia.Naturalista Siciliano, IV, 67–74.
Longmire JL, Lewis AK, Brown NC et al. (1988) Isolation andmolecular characterization of a highly polymorphic cen-tromeric tandem repeat in the family Falconidae. Genomics,2, 14–24.
Lopez Martinez N (1980) Les lagomorphes (Mammalia) du Pleis-tocene superieur de Jaurens. Nouvelles Archives de la Museed’Histoire Naturelle de Lyon, 18, 5–16.
Marboutin E, Peroux R (1995) Survival pattern of European harein a decreasing population. Journal of Applied Ecology, 32, 809–816.
Martin AP, Palumbi SR (1993) Body size, metabolic rate, genera-tion time, and the molecular clock. Proceedings of the NationalAcademy of Sciences of the USA, 90, 4087–4091.
de Menocal PB (1995) Plio-Pleistocene African climate. Science,270, 53–59.
Miller GS (1912) Catalogue of the Mammals of Western Europe. BritishMuseum, London.
Nei M (1987) Molecular Evolutionary Genetics. Columbia Univer-sity Press, New York, NY, USA.
Palacios F (1996) Systematics of the indigenous hares of
MEC766.fm Page 1816 Friday, October 29, 1999 10:55 AM

G E N E T I C S O F L E P U S C O R S I C A N U S 1817
© 1999 Blackwell Science Ltd, Molecular Ecology, 8, 1805–1817
Italy traditionally identified as Lepus europaeus Pallas, 1778(Mammalia: Leporidae). Bonner Zoologische Beitragen, 56, 59–91.
Palacios F (1998) Diversity of hares in Europe. In: Euro-AmericanMammal Congress, Abstracts (ed. Reig S), p. 85. Universidad deSantiago de Compostela, Spain, July 19–24, 1998.
Pérez-Suarez G, Palacios F, Boursot P (1994) Speciation andparaphyly in western Mediterranean hares (Lepus castroviejoi,L. europaeus, L. granatensis, and L. capensis) revealed by mito-chondrial DNA phylogeny. Biochemical Genetics, 32, 423–437.
Petter F (1961) Elements d’une révision des lièvres européens etasiatiques du sous-genre Lepus. Zeitschrift für Säugetierkunde,26, 30–40.
Randi E (1996) A mitochondrial cytochrome b phylogeny of theAlectoris partridges. Molecular Phylogenetics and Evolution, 6,214–227.
Rhymer JM, Simberloff D (1996) Extinction by hybridization andintrogression. Annual Review of Ecology and Systematics, 27, 83–109.
Riga F, Trocchi V, Randi E, Toso S (1998) What, if anything, is theItalian hare? In: Euro-American Mammal Congress, Abstracts (ed.Reig S), p. 97. Universidad de Santiago de Compostela, Spain,July 19–24, 1998.
Rogers AR, Harpending H (1992) Population growth makeswaves in the distribution of pairwise genetic differences.Molecular Biology and Evolution, 9, 552–569.
Rozas J, Rozas R (1997) dnasp, Version 2.0: a novel sofwarepackage for extensive molecular population genetic analysis.Computer Applications in Bioscience, 13, 307–311.
Saitou N, Nei M (1987) The neighbor-joining method: a newmethod for reconstructing phylogenetic trees. Molecular Biologyand Evolution, 4, 406–425.
Santucci F, Emerson BC, Hewitt GM (1998) Mitochondrial DNAphylogeography of European hedgehogs. Molecular Ecology, 7,1163–1172.
Schneider S, Kueffer J-M, Roessli D, Excoffier L (1997) ARLEQUIN,Version 1.1. A software for population genetic data analysis. Geneticsand Biometry Laboratory, Department of Anthropologyand Ecology, University of Geneva, Switzerland. URL: http://anthropologie.unige.ch/arlequin.
Sites JW Jr., Crandall KA (1997) Testing species boundaries inbiodiversity studies. Conservation Biology, 11, 1289–1297.
Slade RW, Moritz C, Hoelzel AR, Burton HR (1998) Molecularpopulation genetics of southern elephant seal Miroungaleonina. Genetics, 149, 1945–1957.
Strimmer K, von Haeseler A (1996) Quartet puzzling: a quartetmaximum likelihood method for reconstructing tree topolo-gies. Molecular Biology and Evolution, 13, 964–969.
Suchentrunk F, Nadlinger K, Alkon PU, Haiden A (1998) Allozymeand mtDNA RFLP data pertinent to the evolution of the brownand Cape hares (Lepus europaeus and L. capensis). In: Euro-American Mammal Congress, Abstracts (ed. Reig S), p. 98. Univer-sidad de Santiago de Compostela, Spain, July 19–24, 1998.
Swofford DL (1993) PAUP: phylogenetic analysis using parsimony,version 3.1. Illinois Natural History Survey, Champaign, Illinois.
Taberlet P, Fumagalli L, Wust-Saucy A-G, Cosson J-F (1998)
Comparative phylogeography and postglacial colonizationroutes in Europe. Molecular Ecology, 7, 453–464.
Tajima F (1983) Evolutionary relationships of DNA sequences infinite populations. Genetics, 105, 437–460.
Tajima F (1989) Statistical methods for testing the neutral muta-tion hypothesis by DNA polymorphism. Genetics, 123, 585–595.
Tamura K, Nei M (1993) Estimation of the number of nucleotidesubstitutions in the control region of mitochondrial DNA inhumans and chimpanzees. Molecular Biology and Evolution, 10,512–526.
Templeton AR (1989) The meaning of species and speciation:a genetic perspective. In: Speciation and its Consequences (edsOtte D, Endler JA), pp. 3–27. Sinauer Associates, Sunderland,Massachusetts.
Thompson JD, Higgins DG, Gibson TJ (1994) clustal w: Improv-ing the sensitivity of progressive multiple sequence alignmentthrough sequence weighting, positions-specific gap penaltiesand weight matrix choice. Nucleic Acids Research, 22, 4673–4680.
Thulin CG, Jaarola M, Tegelström H (1997a) The occurrenceof mountain hare mitochondrial DNA in wild brown hares.Molecular Ecology, 6, 463–467.
Thulin CG, Isaksson M, Tegelström H (1997b) The origin ofScandinavian mountain hares (Lepus timidus). Gibier FauneSauvage, Game Wildlife, 14, 463–475.
Toschi A (1965) Fauna d’Italia. VII. Mammalia (Lagomorpha-Rodentia-Carnivora-Artiodactyla-Cetacea). Calderini, Bologna, pp. 467.
Vigilant L, Stoneking M, Harpending H et al. (1991) Africanpopulations and the evolution of human mitochondrial DNA.Science, 253, 1503–1507.
Vigne JD (1992) Zooarchaeology and the biogeographicalhistory of the mammals of Corsica and Sardinia since the lastice age. Mammal Review, 2, 87–89.
Ward RH, Frazier BL, Dew-Jager K, Pääbo S (1991) Extensivemitochondrial diversity within a single Amerindian tribe.Proceedings of the National Academy of Sciences of the USA, 90,10663–10667.
Watterson G (1975) On the number of segregating sites in gen-etical models without recombination. Theoretical PopulationBiology, 7, 256–267.
de Winton WE (1898) On the hares of Western Europe and NorthAfrica. Annual Magazine of Natural History, London, 1, 149–158.
Yom-Tov Y (1967) On the taxonomic status of the hares (genusLepus) in Israel. Mammalia, 31, 246–259.
Zhang D-X, Hewitt GM (1996) Nuclear integrations: challengesfor mitochondrial DNA markers. Trends in Ecology and Evolu-tion, 11, 247–251.
This study is part of an ongoing project on conservation biologyof hares in Italy. Massimo Pierpaoli and Francesco Riga weresupported by research grants at the INFS; Valter Trocchi is atechnician currently working on wildlife ecology and manage-ment at the INFS. Ettore Randi is head of Conservation Biologyand Genetics at the INFS.
MEC766.fm Page 1817 Friday, October 29, 1999 10:55 AM




















